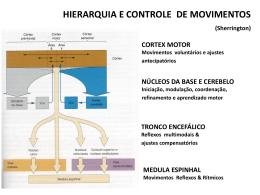
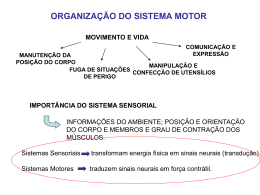
PARTE 3 NEUROCIÊNCIA DOS MOVIMENTOS Capítulo 12 O Alto Comando Motor Estrutura e Função dos Sistemas Supramedulares de Comando e Controle da Motricidade Clique nas setas verdes para avançar/voltar ou ESC para retornar ao menu geral Os ordenadores do sistema motor chegam aos motoneurônios espinhais através das vias descendentes. À esquerda estão aqueles que compõem o subsistema ventromedial, e à direita os que compõem o subsistema lateral. O pequeno encéfalo indica o plano do corte coronal à direita, e a luneta indica o ângulo de observação (dorsal) dos troncos encefálicos desenhados na parte de baixo. A figura não mostra os núcleos dos nervos cranianos e suas vias. Os axônios do feixe piramidal (em vermelho) formam as pirâmides bulbares na • superfície ventral do tronco encefálico, e cruzam na decussação piramidal, visível a olho nu. A ilustração é do histórico livro de Anatomia do inglês Henry Gray (1827-1861). Os diferentes feixes medulares, entidades anatômicas que alojam as vias descendentes dos ordenadores motores, ocupam regiões específicas da substância branca medular. No funículo lateral situam-se os feixes corticoespinhal lateral e rubroespinhal, ambos componentes do subsistema motor lateral. No funículo ventromedial ficam os demais feixes, componentes do subsistema medial (ou ventromedial). Na figura, os feixes estão representados apenas de um lado da medula para simplificar o esquema e facilitar a compreensão. As fibras descendentes que emergem dos feixes para terminar na medula o fazem topograficamente, representando as diferentes regiões do corpo (detalhe acima). Desse modo, as fibras do funículo lateral inervam neurônios laterais do corno ventral, enquanto as do funículo ventromedial inervam neurônios situados mais medialmente. As vias descendentes do sistema medial originam-se de diferentes regiões do tronco encefálico, mas todas terminam em motoneurônios mediais do corno ventral da medula. A. Os feixes reticuloespinhais originam-se de neurônios da formação reticular pontina e da formação reticular bulbar, e os axônios projetam para os motoneurônios do mesmo lado da medula. B. Os feixes vestibuloespinhais originamse dos núcleos vestibulares situados no bulbo, e projetam a ambos os lados da medula. C. O feixe tectoespinhal origina-se no colículo superior, e seus axônios cruzam para o lado oposto antes de chegar à medula. Os desenhos representam cortes transversais do tronco encefálico, numerados em correspondência com os níveis representados no pequeno encéfalo do quadro. As vias descendentes do sistema lateral originam-se no córtex cerebral e no mesencéfalo, terminando nos motoneurônios laterais do corno ventral da medula. Os feixes corticoespinhais originam-se principalmente na área motora primária, mas só o maior deles (o lateral) cruza na decussação piramidal antes de atingir a medula (o feixe corticoespinhal medial não está ilustrado na figura). O feixe rubroespinhal origina-se no núcleo rubro e cruza no tronco encefálico alto. Os desenhos representam cortes transversos numerados em correspondência com os níveis representados no pequeno encéfalo do quadro. Alguns dos circuitos posturais têm origem nos órgãos vestibulares (à direita), outros nos fusos musculares dentro dos músculos. Desses dois órgãos receptores emergem as vias aferentes (em azul). As principais estruturas que coordenam as reações posturais são os núcleos vestibulares, que comandam a musculatura do corpo, e os núcleos motores do globo ocular, que comandam a musculatura extraocular. Por simplicidade, só estão ilustradas (em vermelho) as vias eferentes do cerebelo, núcleo abducente e núcleos vestibulares. Os axônios de comando dos movimentos oculares originam-se nos núcleos dos nervos motores do globo ocular, com um padrão específico de inervação. À esquerda estão representados cortes transversos do tronco encefálico, cuja vista dorsal está representada à direita. Os movimentos de estabilização do olhar são comandados a partir de informações veiculadas pela retina aos núcleos pretectais, que por sua vez emitem projeções até os núcleos dos nervos cranianos correspondentes. Observar que apenas o núcleo troclear emite projeções cruzadas. A representação dos movimentos oculares no colículo superior pode ser revelada experimentalmente. O pesquisador registra a atividade neuronal no colículo superior (pontos numerados, em B), e procura no campo visual a posição dos campos receptores correspondentes (círculos brancos numerados, em A). Depois estimula eletricamente a região através do mesmo eletródio, e registra o movimento ocular produzido (setas em A). Observa-se assim que os olhos do animal se movem do centro do campo visual para o centro do campo receptor, cada vez que a região correspondente do colículo superior é estimulada. O meridiano horizontal do campo visual (em A) e sua representação no colículo (em B) estão representados por uma linha pontilhada, e o meridiano vertical por uma linha contínua grossa. Os demais meridianos estão representados por linhas contínuas finas. As cores representam os quadrantes do campo (em A), e correspondem às regiões do colículo (em B). Os movimentos sacádicos são comandados pelo córtex frontal e pelo colículo superior (neurônios vermelhos) através da formação reticular pontina do lado oposto. Os neurônios desta (em azul) projetam aos núcleos motores do globo ocular. As áreas motoras corticais estão representadas em tons de azul. As áreas representadas em tons de verde conectam-se com as primeiras, mas não são consideradas partes do sistema motor. O desenho de cima ilustra a face lateral do hemisfério esquerdo, e o desenho de baixo ilustra a face medial do hemisfério direito. Todas as áreas representadas, entretanto, existem em ambos os hemisférios. Abreviaturas no texto. Os números referem-se à classificação citoarquitetônica de Brodmann. Ferrier (à esquerda) realizou seus experimentos em macacos, mas logo extrapolou os dados •• para o cérebro humano. Segundo ele, os pontos motores do cérebro humano são os numerados de 2 a 12, o que corresponde ao que se conhece atualmente. Desenho à direita de D. Ferrier (1876) Functions of the Brain. Putnam, EUA. A somatotopia é um importante princípio de organização de M1. A. A estimulação elétrica de partes do giro pré-central permite idealizar um homúnculo que representaria o “mapa motor” do corpo humano na superfície cortical. B. Os experimentos feitos no cérebro de macacos indicaram que cada ponto estimulado pode provocar a ativação de vários músculos. O desenho de baixo representa uma ampliação do desenho de cima, e os campos em preto representam as partes do corpo do macaco que se movem quando cada ponto do córtex é estimulado eletricamente. Divergência e convergência dos axônios corticoespinhais. A representa um único neurônio corticoespinhal que projeta para diferentes motoneurônios (divergência), cada um deles responsável pelo comando de um músculo diferente. B. A morfologia de um único axônio corticoespinhal (em vermelho) revela ramificações em diferentes setores do corno ventral da medula. C. A estimulação elétrica de um único axônio corticoespinhal (indicadapelo asterisco sobre o traçado gráfico) ativa os músculos 1-4 (ondas assinaladas pelos pontos vermelhos), mas não os músculos 5 e 6. D representa a convergência: vários neurônios corticoespinhais projetam para um único motoneurônio medular. Imagens de ressonância magnética funcional de um indivíduo durante o movimento dos dedos da mão direita. Aparecem ativas lateralmente as áreas motora primária (M1) e somestésica primária (S1), e medialmente a área motora suplementar (MS). A representa um plano horizontal mais profundo que B, como indicado em C. S1 é ativada por que o próprio movimento causa estimulação somestésica. O experimento de Evarts. A. O macaco era treinado a estender ou fletir o punho, e o pesquisador ao mesmo tempo registrava a atividade neuronal do seu córtex motor com um microeletródio. B. O traçado azul representa o disparo de potenciais de ação (cada traço vertical) de um neurônio motor cortical, enquanto o traçado vermelho indica os movimentos do punho. Observar que o neurônio começa a disparar antes da flexão (B1), e que a frequência de PAs é maior quando a força muscular empregada é também maior (B2). Quando a flexão é “ajudada” por um peso colocado contra os músculos extensores, o punho flete sem mesmo a necessidade de uma contração muscular (B3). Na técnica de estimulação magnética transcraniana (A), estimulando-se o córtex motor primário (M1), obtém-se um potencial evocado motor no músculo oponente do polegar, graças à ativação corticoespinhal e ao registro por meio de eletrodos posicionados sobre o músculo (foto abaixo). À direita se pode ver a região de representação da mão de um paciente biamputado e bitransplantado, após 2 (B) e 10 (C) meses da realização da cirurgia. Notar que com o tempo a região cortical que comanda a mão amplia-se significativamente. Cortesia do Laboratório de Neuropsicologia da Ação, Além da força muscular, os neurônios corticais também comandam a direção do movimento. A. O macaco é treinado a realizar movimentos em determinadas direções, enquanto o pesquisador registra a atividade elétrica dos neurônios corticais. B. O comprimento de cada traço azul representa a frequência de disparo de PAs de um neurônio cortical antes de um movimento direcionado. As setas pretas indicam a resultante da atividade da população neuronal que dispara antes de cada movimento. Pode-se ver que a resultante é muito próxima da direção efetiva do movimento. Planejamento e comando motor envolvem áreas diferentes do córtex cerebral. A. O movimento simples de um dedo provoca a ativação de M1 e S1 no hemisfério esquerdo. B. Um movimento complexo envolvendo vários dedos em sequência provoca a ativação de várias áreas em ambos os hemisférios. C. Pensar no movimento anterior, sem executá-lo, ativa apenas a região de planejamento motor. O experimento de Passingham. Enquanto o indivíduo tenta descobrir a sequência correta de movimentos (A), as áreas ativadas são diferentes de quando ele a descobre (B). A. Regiões onde foram encontrados neurôniosespelho, ligadas estritamente ao sistema visuomotor do córtex (em azul), a sistemas cognitivos complexos (em verde), e ao processamento de emoções (em laranja, representando a ínsulaA e a amígdalaA). B. Buba ou quiqui, qual é um, qual é o outro? O cerebelo possui um córtex na superfície e núcleos profundos no seu interior. A. Vista dorsal do cerebelo (indicada pela luneta no pequeno encéfalo acima e à direita), com os núcleos profundos desenhados em verde “por transparência”. B. Vista ventral do cerebelo (luneta no pequeno encéfalo abaixo à direita), com os pedúnculos cerebelares cortados. A. Do ponto de vista das suas conexões, o cerebelo é subdividido em três regiões: verme, hemisférios intermédios e hemisférios laterais. Os núcleos profundos recebem aferentes seletivos de cada subdivisão, como se pode ver pela equivalência de cores. B. As subdivisões conectivas do cerebelo são também funcionais, e relacionam-se com os subsistemas motores, definindo o vestibulocerebelo, o espinocerebelo e o cerebrocerebelo. Os diagramas de blocos representam os aferentes e os eferentes de cada subdivisão funcional. Uma pequena fatia do córtex cerebelar (detalhe acima, à esquerda) é representativa de todas as regiões. A representa as fibras aferentes do cerebelo (em vermelho). A seta aponta para um glomérulo cerebelar. B representa as fibras eferentes do córtex cerebelar, que emergem das células de Purkinje (em vermelho). C ilustra os interneurônios principais (também em vermelho). O circuito básico do cerebelo pode ser representado esquematicamente. Notar que as fibras que saem do cerebelo a partir dos núcleos profundos são excitatórias, mas as fibras de Purkinje que sobre elas incidem são fortemente inibitórias. As fibras aferentes que chegam ao cerebelo do tronco encefálico e da medula são excitatórias. Os núcleos da base (em verde) ficam no interior do encéfalo, e são atravessados pela cápsula interna (em azul). A. Representação “por transparência” dos núcleos da base, atravessados por dois dos feixes da cápsula interna. B. Representação do corte indicado pela linha branca em A, mostrando também os núcleos da base em relação à cápsula interna. Um plano de corte oblíquo como esse permite visualizar ao mesmo tempo os componentes telencefálicos, diencefálicos e mesencefálicos dos núcleos da base. A substância negra pode ser vista claramente acima do pedúnculo cerebral, no mesencéfalo (desenho de baixo em B). As informações de entrada que o corpo estriado processa vêm de extensas regiões do córtex cerebral (A), enquanto as informações de saída seguem para certos núcleos do tálamo, que as transmite para as regiões motoras do córtex, e para o colículo superior do mesencéfalo (B). Os núcleos da base constituem, entre si, um complexo circuito de processamento (C) que transforma as informações de entrada em comandos para iniciar ou terminar um movimento simples, uma sequência deles, ou mesmo comportamentos complexos e elaborados. Os aferentes da célula espinhosa média do corpo estriado são conhecidos quanto à sua origem, seu local preciso de terminação, e o seu neurotransmissor. ACh = acetilcolina; Cd = caudado; DA = dopamina; GABA = ácido gamaaminobutírico; Glut = glutamato; Pu = putâmen; SN = substância negra. A. Representação dos principais circuitos dos núcleos da base em um indivíduo normal, com os neurônios inibitórios representados em vermelho e os excitatórios em azul. As conexões recíprocas não estão representadas, por simplicidade. B. Nos doentes parkinsonianos, neurônios negro-estriados degeneram. C. Nos pacientes com balismo, degeneram os neurônios subtalâmico-pálidos, e nos pacientes com doença de Huntington (D), são os neurônios espinhosos médios do corpo estriado que degeneram. Alguns dos sintomas dessas doenças podem ser explicadas analisando os circuitos (veja o texto). Cd = caudado; GPe = globo pálido externo; GPi = globo pálido interno; Pu = putâmen; SN = substância negra; ST = núcleo subtalâmico.
Baixar