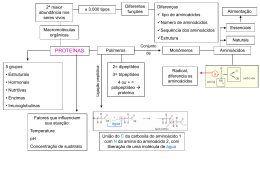
ENZIMAS As enzimas são proteínas, catalisadores (aumenta a velocidade de uma determinada reação química) biológicos (proteínas) de alta especificidade. Praticamente todas as reações que caracterizam o metabolismo celular são catalisadas por enzimas. Aceleram a velocidade de uma reação, sem, no entanto participar dela como reagente ou produto, regulando várias reações. Atuam em concentrações muito baixas e em condições suaves de temperatura e pH. As enzimas são sintetizadas através de informação genética, desta forma, caso haja erro na passagem da informação ocorrerá erro na síntese da enzima e conseqüentemente na sua ação, levando á algumas doenças. Células podem sintetizar enzimas conforme a sua necessidade. As enzimas devem: o Aumentar a velocidade das reações. o Não alterar a natureza das reações. o Pode diminuir a energia de ativação (EA). NOMENCLATURA Geralmente se utiliza o sufixo "ase" para caracterizar a enzima. Exs: Urease, Hexoquinase, Peptidase, etc., mas as enzimas primeiramente descobertas tem nomes diferenciados como pepsina e tripsina. CLASSIFICAÇÃO DAS ENZIMAS A determinação do nome das enzimas é normatizada por um comitê especializado, o Nomenclature Committee of the International Union of Biochemistry and Molecular Biology (NC-IUBMB). - Oxidorredutases: São enzimas que catalisam reações de transferência de elétrons, ou seja: reações de oxi-redução. São as Desidrogenases e as Oxidases. Se uma molécula se reduz, tem que haver outra que se oxide. - Transferases : Enzimas que catalisam reações de transferência de grupamentos funcionais como grupos amina, fosfato, acil, carboxil, etc. Como exemplo temos as Quinases e as Transaminases. - Hidrolases : Catalisam reações de hidrólise de ligação covalente. Ex: As peptidades. - Liases: Catalisam a quebra de ligações covalentes e a remoção de moléculas de água, amônia e gás carbônico. As Dehidratases e as Descarboxilases são bons exemplos. - Isomerases: Catalisam reações de interconversão entre isômeros ópticos ou geométricos. As Epimerases são exemplos. - Ligases: Catalisam reações de formação e novas moléculas a partir da ligação entre duas já existentes, sempre às custas de energia (ATP). São as Sintetases. Classes de enzimas Classe Catalisam reações de oxirredução, Oxirredutases 1 transferindo elétrons ou prótons (H+). Classe Transferases Transferem grupos químicos entre moléculas. 2 Classe Utilizam a água como receptor de grupos Hidrolases 3 funcionais de outras moléculas. Formam ou destroem ligações duplas, Classe Liases respectivamente retirando ou adicionando 4 grupos funcionais. Classe Isomerases Transformam uma molécula num isômero. 5 Classe Ligases 6 Formam ligações químicas por reações de condensação, consumindo energia sob a forma de ATP. A partir destas categorias principais, as enzimas ainda são subdivididas em outras categorias, podendo ser identificadas pelo seu número da EC; por exemplo, EC 5.4.2.2 é a fosfoglicomutase. COFATORES ENZIMÁTICOS E COENZIMAS Cofatores são pequenas moléculas orgânicas ou inorgânicas que podem ser necessárias para a função de uma enzima (apoenzima). Coenzimas A molécula orgânica que se liga às enzimas para ativar uma reação, é denominada coenzima, cada tipo apresenta uma função química particular, algumas são agentes oxi-redutoras, outras transferidoras etc. Exemplo: Coenzima: dinucleotídeo nicotinamida de adenina (NAD+). Muitas coenzimas são fortemente relacionadas com vitaminas. As vitaminas são moléculas orgânicas essenciais para o processo biológico em organismos superiores e não podem ser sintetizados por eles mesmos. Portanto estas coenzimas são essenciais aos processos metabólicos vitais dos organismos superiores. CINÉTICA ENZIMÁTICA Estuda a velocidade das reações enzimáticas, e os atores que influenciam nesta velocidade. A cinética enzimática é o estudo do mecanismo pelo qual as enzimas ligam substratos e os transformam em produtos Uma reação enzimática pode ser expressa pela seguinte equação: E + S <==> [ES] ==> E + P O complexo enzima/substrato (ES) tem uma energia de ativação ligeiramente menor que a do substrato isolado, e a sua formação leva ao aparecimento do estado de transição. A formação de "P" a partir de ES é a etapa limitante da velocidade da reação. A velocidade de uma reação enzimática depende das concentrações de enzima e de substrato. Podemos chegar à velocidade máxima de reação aumentando progressivamente a concentração de substrato, até se verificar uma velocidade constante de formação de produto (como no gráfico). A saturação acontece porque, à medida que é aumentada a concentração de substrato, aumenta também a quantidade de enzima presente sob a forma de complexo enzima-substrato (ES). À velocidade máxima, Vmax, todos os centros ativos estão ocupados (saturados) com substrato, ou seja, não existe enzima livre para ligar mais substrato e a concentração de complexo ES é igual à concentração de enzima. Em 1913 L. Michaelis e M. L. Menten postularam a teoria da ação e cinética enzimática, levando à equação que nos permite demonstrar como a velocidade de uma reação varia em função da concentração do substrato: Enzima + Substrato « [EnzimaSubstrato] -> Enzima + Produto V= Esta equação pode ser expressa graficamente, e representa o efeito da concentração de substrato sobre a velocidade de reação enzimática. O Km de um substrato para uma enzima específica é característico, e nos fornece um parâmetro de especificidade deste substrato em relação à enzima. Quanto menor o Km, maior a especificidade, e vice-versa. Curva de saturação numa reação enzimática, mostrando a relação entre a concentração de substrato ([S]) e a velocidade (V). Lineweaver-Burke linearizaram a curva de Michaelis- Menten (gráfico abaixo) Inibição Enzimática Muitos tipos de moléculas inibem as enzimas e podem agir de várias formas. A principal distinção é entre inibição reversível e inibição irreversível A forma que envolve ligações não-covalentes é reversível, através da remoção do inibidor, já a inibição irreversível, a ligação molecular é covalente. Inibição Irreversível Inibidores irreversíveis podem ser extremamente seletivos, e se ligam covalentemente às enzimas deixando-as inativas. Na maioria dos casos a substância reage com o grupo funcional no sítio ativo bloqueando o local do substrato, deixando a enzima catalíticamente inativa. Inibição Reversível Existem vários modos que estão envolvidos com ligação não covalente, eles diferenciam quanto ao mecanismo pelo qual diminuem a atividade enzimática e como eles afetam na cinética da reação. Os inibidores enzimáticos são compostos que podem diminuir a atividade de uma enzima. Existem 2 tipos de inibição enzimática reversível: Competitiva e Não competitiva Inibição Reversível Competitiva Uma molécula apresenta estrutura semelhante ao substrato da enzima se ligando ao sítio ativo, competindo com o substrato. O efeito é revertido aumentando-se a concentração de substrato Na inibição competitiva, o inibidor e o substrato competem pela enzima, isto é, não se podem ligar ao mesmo tempo à enzima. O efeito da reação modifica o Km, mas não altera a velocidade máxima , níveis mais altos de concentração do substrato são requeridos para que se atinja uma determinada velocidade, aumentando o KM aparente. Inibidor Reversível não Competitivo Ocorre quando um molécula se liga em um segundo local na superfície enzimática (não no sítio ativo), modificando a forma da enzima dificultando a catálise. Os inibidores não-competitivos podem ligar-se à enzima e ao substrato ao mesmo tempo, ou seja, nunca se ligam ao sítio ativo. O inibidor não competitivo pode ser uma molécula que não se assemelha com o substrato, mas apresenta uma grande afinidade com a enzima. É o mecanismo inverso do inibidor competitivo, porque inibe a ligação do complexo ES e não da enzima livre. O efeito da reação modifica a velocidade e o Km permanece constante. Inibidor reversível competitivo e não competitivo no gráfico de Lineweaver-Burke
Baixar