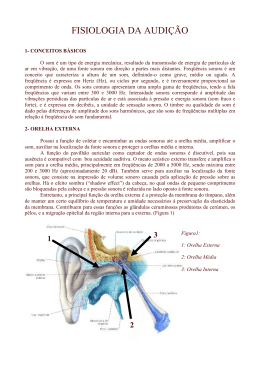
PARTE 2 Neurociência Sensorial Capítulo 8 Os Sons do Mundo Estrutura e Função do Sistema Auditivo Clique nas setas verdes para avançar/voltar ou ESC para retornar ao menu geral O som é produzido pela vibração de objetos sólidos que põem em movimento as partículas do ar circundante. Criam-se regiões de compressão e rarefação das partículas, que se deslocam para fora como superfícies esféricas de raios crescentes. Nas ondas transversais as partículas vibram em direção perpendicular à sua propagação (A), enquanto nas ondas longitudinais, vibração e propagação têm a mesma direção (B). Os tons puros são ondas senoidais. Neste experimento imaginário, medese a densidade de partículas em um ponto fixo durante algum tempo (A). Verifica-se que a densidade naquele ponto varia no tempo de acordo com uma curva senoidal. Depois (B) medese a densidade em três pontos diferentes, simultaneamente. Encontram-se as mesmas curvas em todos os pontos, mas um pouco deslocadas uma em relação à outra. Amplitude (A) é diferente de frequência. Enquanto a primeira permite determinar a quantidade de energia (E) contida na onda sonora em cada ponto do ciclo (A1 < A2, logo E1 < E2), a frequência representa a quantidade de ciclos que ocorrem em um certo período de tempo. As ondas sonoras interagem, somando-se algebricamente. A representa a soma de duas ondas em coincidência de fase, produzindo uma onda resultante de maior amplitude e mesma frequência. B representa um caso de oposição de fase, em que as duas ondas iguais que interagem se anulam. C mostra a resultante da interação de três ondas diferentes. É assim complexa a maioria dos sons que ouvimos. As curvas mostram o limiar de audibilidade para uma população de indivíduos. Os níveis de intensidade sonora que os indivíduos são capazes de ouvir ficam acima de cada curva. O grupo de indivíduos com melhor audição (1%) está representado pela curva cinza. As demais curvas representam cada uma delas uma maior proporção de pessoas na população. A curva cinza de cima mostra o limiar para dor provocada por intensidades sonoras muito fortes. Pode-se decompor matematicamente em ondas senoidais simples a onda complexa produzida pelo som de um instrumento musical como o clarinete. Neste caso, haverá uma frequência fundamental característica de um tom (dó, ré etc.), e uma composição de harmônicos característica do instrumento. A cóclea, órgão receptor do sistema auditivo, fica no labirinto (A), uma estrutura membranosa incrustada no osso temporal. O corte de uma volta da cóclea (B) mostra que ela é formada por canais ou escalas, e que as células receptoras ficam situadas entre duas membranas (tectorial e basilar). A maioria das fibras auditivas é aferente, e seus somas ficam no gânglio espiral. Visto de um outro ângulo e em maior ampliação (C), o nervo auditivo contém fibras aferentes (em verde-escuro) mas também fibras eferentes (em roxo) que inervam os receptores. Todos os níveis do SNC apresentam componentes do sistema auditivo. A é uma vista dorsal do tronco encefálico, do ângulo assinalado pela luneta no pequeno encéfalo acima. No encéfalo estão também representados os planos dos cortes (números circulados) mostrados em B. Tanto em A como em B, os neurônios auditivos estão representados em roxo e preto (os aferentes) e em vermelho (os eferentes). A posição das áreas auditivas corticais no homem pode ser visualizada na face lateral do encéfalo (A), e mais completamente se removermos a parte superior dos hemisférios (B) para revelar o assoalho do sulco lateral (C). Através de ressonância magnética funcional a área A1 aparece (D) quando se oferece estimulação sonora a um indivíduo, que provoca o aumento do fluxo sanguíneo da região, resultante da atividade neuronal. A reconstrução por computador mostra os focos de ativação bilateral (em vermelho) no giro temporal superior de ambos os hemisférios. As vistas de C e D são indicadas pela luneta em B. W = área de Wernicke. A membrana basilar vibra a cada som que entra no ouvido (A), e vibra mais forte quando o som incidente é também mais forte (B). A cóclea está aqui representada como se estivesse desenrolada (pequeno detalhe em A). A relação de proporcionalidade entre a intensidade do som e a resposta dos axônios aferentes foi medida experimentalmente (C). Constatou-se que a frequência de PAs é maior (curva verde) para sons mais fortes. Hermann von Helmholtz (à esquerda) e Georg Von Békésy (à direita). No centro, esquema da tonotopia coclear desenhado por Békésy. Experimentos de registro eletrofisiológico indicaram que as variações da frequência do potencial receptor das células estereociliadas da cóclea acompanham a frequência do som incidente (A). O mesmo ocorre com a frequência das salvas de PAs das fibras do nervo auditivo (B). Mas isso só é verdade para os tons graves e médios (entre 300 e 1.000 Hz). E os agudos?. A tonotopia representa uma especialização da membrana basilar: os sons mais graves fazem vibrar o ápice (A), e os mais agudos movimentam a base (B). Na verdade, cada frequência faz vibrar seletivamente um local da membrana basilar. Desse modo o sistema auditivo discrimina (separa) os tons, mesmo os mais agudos que não são acompanhados pelo princípio das salvas. As curvas de sintonia das células estereociliadas da cóclea (A) e das fibras do nervo auditivo (B) revelam uma frequência característica individual (linhas tracejadas). Nessa frequência ocorre o disparo de um PR ou de PAs, respectivamente, para um som incidente de intensidade mínima. Quando os sons incidentes se afastam dessa frequência para mais ou para menos, é preciso aumentar a intensidade para ativar a célula ou a fibra. A frequência característica da célula azul, por exemplo (em A) e da fibra laranja (em B) é de quase 1 kHz. As frequências vizinhas não são tão eficazes para elas, mas podem ativar outras, representadas em cinza. A organização tonotópica aplica-se a todo o sistema auditivo, da membrana basilar às áreas corticais. Em todas essas regiões se encontram mapas tonotópicos, isto é, uma distribuição ordenada de neurônios que respondem à série de frequências audíveis. O detalhe acima mostra o mapa tonotópico de A1. No exemplo, a cadeia de neurônios ativada para o som que faz vibrar a membrana basilar (abaixo) está representada em vermelho em todos os estágios do sistema auditivo. A eletromotilidade é possibilitada por proteínas como a prestina, que de algum modo detectam a despolarização da membrana causada pelo estímulo sonoro e contraem-se, provocando o encurtamento da célula ciliada. A. A fotomicrografia eletrônica mostra os estereocílios alinhados das células ciliadas externas. B mostra a presença de caderina (pontos verdes), uma proteína que contribui para a abertura dos canais de potássio na ponta dos estereocílios. C representa a motilidade dessas células receptoras em função do potencial de sua membrana. Quando ocorre uma despolarização provocada pelas fibras eferentes olivococleares (à direita), a célula se contrai, “puxando” a membrana basilar para cima e tornando mais rígido o conjunto. Quando um som complexo entra no ouvido, faz vibrar ao mesmo tempo diversas partes da membrana basilar, e assim ativa – em paralelo – as regiões tonotópicas correspondentes do sistema auditivo. O desenho mostra as regiões mais ativas em vermelho, e as menos ativas em cinza ao longo do sistema. A. Um som que incide de lado atinge primeiro uma das orelhas e forma uma “sombra” atrás da cabeça. A outra orelha será atingida por reflexão da onda incidente nos objetos do ambiente próximo. B. Cada um dos neurônios do complexo olivar superior, indicados em C, apresenta disparo de PAs em maior frequência para certas diferenças de fase que resultam da diferença do tempo de chegada do som às duas orelhas. Os neurônios do núcleo olivar superior lateral detectam diferenças de intensidade dos sons incidentes em cada orelha, com a intervenção de neurônios inibitórios do núcleo do corpo trapezóide (em vermelho). Este mecanismo é mais eficiente para a localização espacial dos sons agudos. Estudos experimentais no macaco (acima) têm permitido identificar diferentes áreas no assoalho do lobo temporal (visualizado por meio de um “corte” das regiões sobrepostas). A partir de A1, essas áreas mostraram-se fortemente interconectadas (setas vermelhas). No córtex humano (abaixo), os estudos não têm ainda precisão comparável, mas pode identificar-se a área 41 de Brodmann como a região auditiva primária (A1), 42 e 52 como o cinturão auditivo, e 22 e talvez 38 como o paracinturão.
Baixar