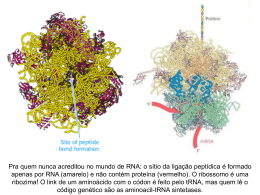
Tradução: do mRNA à Proteína Faculdades de Medicina e Medicina Dentária da Universidade do Porto Disciplina de Biologia Celular e Molecular Penas, Ana; Silva, Ana A.; Silva, Ana N.; Fontes, Andreia; Ferreira, Luís Introdução As proteínas são efectores de um grande número de processos celulares, executando uma enorme variedade de tarefas. Uma célula eucariótica típica requer centenas de proteínas diferentes num dado momento. Assim, a questão impõe-se: como é feita a síntese de proteínas? Para responder a esta questão, é oportuno debruçarmo-nos sobre a tradução, processo subjacente à síntese proteica. A tradução do mRNA é o primeiro passo na formação de uma proteína funcional. Seguidamente, a cadeia polipeptídica deve dobrar-se na conformação tridimensional apropriada e passar por diversas etapas de processamento antes de atingir a sua forma activa. Da tradução, devemos analisar as suas 3 fases - iniciação, alongamento e terminação - e, de seguida, atender ao modo como estas são reguladas. Embora a expressão da maioria dos genes seja regulada primariamente ao nível da transcrição, a expressão genética também pode ser controlada ao nível da tradução, e esse controle é um elemento fundamental da regulação genética. Finalmente, afigura-se indispensável a ligação de todo o processo à prática da Medicina. Comecemos então esta viagem do mRNA à Proteína... Alongamento Iniciação • Ligação dos Eucariotic Initiation Factors eIF-1, eIF-1A, eIF-3 e eIF-5 à subunidade 40s do ribossoma. • Ligação do eIF-2 (num complexo com GTP) ao tRNA metionil iniciador. • Reconhecimento do mRNA e transporte deste ao ribossoma pelo grupo eIF-4, sendo o 5’ cap do mRNA reconhecido pelo eIF-4E. • Ligação do eIF-4G ao eIF-4E e a uma proteína (designada PABP- poly-A binding protein) associada à cauda poli-A da extremidade 3’. Terminação • Ribossoma com três locais para a ligação do tRNA: locais P (peptidil), A (aminoacil) e E (exit). • Ligação do tRNA metionil iniciador ao local P. • Ligação do próximo tRNA aminoacil ao local A, por emparelhamento com o segundo codão do mRNA; este tRNA é acompanhado até ao ribossoma por um Eucariotic Elongation Factor – eEF-1α – que forma um complexo com o GTP. • Hidrólise do GTP a GDP (quando o tRNA aminoacil correcto é inserido no local A) e libertação do factor de alongamento ligado ao GDP. • Estabelecimento de ligação peptídica entre a metionina iniciadora e o segundo aminoácido, por acção da subunidade 60s, após quebra da ligação do eEF-1α ao ribossoma. • Transferência da metionina para o tRNA aminoacil do local A, seguida de formação de um tRNA peptidil; permanência do tRNA iniciador, sem a metionina, no local P. • Reconhecimento de ambas as extremidades dos mRNAs pelos eIFs, o que contribui para o estímulo da tradução. • Transporte do mRNA pelos eIF-4E, eIF-4G, eIF-4A e eIF4B à subunidade 40s ribossomal, ocorrendo uma interacção entre o eIF-4G e o eIF-3. • Associação do tRNA metionil e eIFs à subunidade 40s, que percorre o mRNA até identificar o codão de iniciação AUG. • Ausência de tRNAs com anticodões complementares aos codões STOP implica o reconhecimento destes pelo factor de libertação eRF-1 (Eucatiotic Release Factor). Conjuntamente com o eRF-1, actua o factor eRF-3, não envolvido no reconhecimento dos codões STOP. • Translocação, com intervenção do eEF2, acoplada à hidrólise do GTP. • Durante a translocação, deslocamento, em três nucleótidos, do ribossoma; este posiciona o próximo codão num local A vazio. • Transferência do tRNA peptidil do local A para o P e do tRNA iniciador desprovido de metionina do local P para o E. • Após identificação do codão AUG, há o desencadear, pelo eIF-5, da hidrólise do GTP ligado ao eIF-2. • Libertação dos factores de iniciação (incluindo eIF-2 ligado ao GDP) e junção da subunidade 60s à subunidade 40s para formar o complexo de iniciação. • Ligação de novo tRNA aminoacil ao local A, induzindo à libertação do tRNA iniciador do local E: ribossoma pronto para a inserção de um próximo aminoácido na cadeia polipeptídica em formação. • Reconversão, durante o alongamento, do eEF-1α libertado do ribossoma ligado ao GDP à sua forma GTP, havendo intervenção de um terceiro factor de alongamento - eEF-1βγ; este une-se ao complexo eEF-1 α/GDP e promove a troca do GDP por GTP: o eEF-1α/GTP pode acompanhar um novo tRNA aminoacil ao local A, começando um novo ciclo de alongamento. • Translocação, no local A, de codão STOP (UAA, UAG e UGA) determina fim do alongamento. Regulação da Síntese Proteica Quer nos Eucariontes quer nos Procariontes, há uma regulação do processo de tradução. Assim, enumeramos alguns mecanismos de Regulação da Tradução: 1. Ligação de proteínas repressoras: As proteínas repressoras vão ligar-se à extremidade 5’ do mRNA, inibindo deste modo a Iniciação. 2. Regulação da Actividade de Factores de Iniciação eIF-2 e eIF - 4E: No final da Iniciação, para que o eIF-2 se dissocie do GDP e fique disponível para nova ligação ao GTP, tem que intervir o factor eIF-2B. Na inibição, que se sucede é que o eIF-2 sofre fosforilação e, de seguida, liga-se ao eIF-2B, conduzindo à inactivação deste. Estando o eIF-2B inactivado, o eIF-2 não se dissocia do GDP, logo, não pode ser reutilizado. Por conseguinte, a síntese de proteínas é inibida. No que se refere ao factor eIF – 4E, existem eIF - 4E binding proteins, que quando desfosforiladas se ligam a este factor impedindo a sua ligação ao eIF – 4G. Por conseguinte, a regulação da síntese proteica resulta da fosforilação ou desfosforilação das eIF – 4E binding proteins. 3. Iniciação nos codões AUG downstream do local de início da tradução: A Iniciação começa não no primeiro codão AUG, mas no segundo ou terceiro; é um mecanismo que conduz à produção de proteínas que diferem entre si apenas no terminal amino, sendo, por conseguinte, uma “estratégia utilizada pelo gene” para produzir proteínas semelhantes, mas que vão ter, por exemplo, diferentes localizações celulares. 4. Locais de entrada internos no ribossoma: A tradução pode também iniciar-se em sequências especializadas de RNA - IRES (internal ribosome entry site). Quando células eucariotas entram na fase M do ciclo celular regista-se uma redução de 25% na taxa total de tradução, uma vez que existe uma desfosforilação do eIF-4E (diminui a afinidade para o 5’ cap). No entanto os mRNAs com IRES não sofrem o efeito do eIF-4E de modo que a sua tradução aumenta quando a célula entra na fase M. 5. Mudança de estabilidade de mRNA: A síntese de proteínas codificadas por mRNA estável persiste muito tempo para além da repressão da transcrição do gene. Nos Eucariontes, existem proteínas com um tempo de semi-vida muito longo. Outras, porém, são apenas necessárias por curtos períodos de tempo e têm que ser expressas breve e intensamente, como acontece, por exemplo, com os factores de tradução. Nestas proteínas, o mRNA correspondente tem um tempo de semi-vida curto, sendo que a sua expressão é controlada por activação e desactivação da transcrição dos seus mRNAs, que se traduzirá, naturalmente, numa activação e desactivação da tradução. Muitos destes mRNAs têm múltiplas cópias da sequência AUUUA, geralmente localizadas na UTR (untranslated region) da extremidade 3’. 6. Regulação do tamanho da cadeia poli-A dos mRNAs dos Ovócitos: Nos ovócitos, existem mRNAs cuja cadeia poli-A tem cerca de 20 nucleotídeos; estes mRNAs armazenados são utilizados para a tradução num estádio de desenvolvimento apropriado através do alongamento da cadeia poli-A até centenas de nucleotídeos. É, pois, através da regulação do tamanho da cadeia poli-A que se faz a regulação da tradução. 7. RNA interference que silencia a expressão de genes: Nas células eucarióticas existe um mecanismo de degradação de RNA, designado RNAi, que é utilizado como mecanismo de defesa, destruindo moléculas de RNA estranho. Este actua com a formação de cadeias duplas de RNA que não são traduzidas e, por isso, são facilmente degradadas. • Ligação dos factores eRF-1 e eRF-3 a um codão de terminação no seu local A que conduz à hidrólise da ligação entre o tRNA e a cadeia polipeptídica no local P, resultando na libertação do polipeptídeo completo do ribossoma. • Libertação do tRNA e dissociação ribossomais e do mRNA molde. das subunidades Ligação à Medicina Wang, C. e Pflugheber, J. (2003): Alpha Interferon Induces Distinct Translational Control Programs To Suppress Hepatitis C Virus RNA Replication • A infecção pelo vírus de hepatite C (HCV) é um grave problema de saúde pública. Dados recentes revelam que mais de 170 milhões de pessoas em todo o mundo são infectadas, sendo que estudos epidemiológicos identificaram o vírus da hepatite C como a principal causa de hepatite crónica e de doença hepática na população humana. • A infecção com o HCV é frequentemente tratada com uma terapia que recorre a um alfa interferão (α-IFN). Os IFNs são produzidos pela célula em resposta à infecção vírica. A exposição da célula aos IFNs leva, por sua vez, à expressão induzida de genes estimulados pelo IFNs (ISG), estando a maioria deles envolvidos em funções antivíricas. Assim, acredita-se que a acção dos ISGs pode limitar a replicação do vírus. • Há, pelo menos, três vias de controlo de tradução a nível celular desencadeados como resposta à presença de vírus e, por conseguinte, de INFs: a via da sintétase do 2’-5’ oligoadenilato (OAS)/ RNase L; a via da cínase da proteína R (PKR) – subunidade alfa do factor de iniciação eucariótico (eIF-2α); a via do P56-eIF-3. • A activação da enzima RNase L pelo OAS pode levar a uma supressão na tradução através da clivagem e consequente inactivação do rRNA 28s. • A activação da PKR durante a infecção vírica conduz à reacção de fosforilação na serina-51 do factor de iniciação eIF-2α. Esta fosforilação pode bloquear a iniciação na tradução, uma vez que reduz o pool de eIF-2α, bloqueando a ligação do tRNA metionil à subunidade 40s. • A P56 foi identificada como sendo uma proteína de ligação ao eIF-3, supressora da tradução. • Em suma, este estudo caracterizou os mecanismos moleculares de actuação do IFN contra a replicação do vírus HCV, caracterizando as vias da PKR e P56 enquanto reguladores dos programas de tradução do HCV através da restrição no recrutamento de RNA viral nos ribossomas. Fig.1: Mecanismos de Repressão da Tradução do RNA do HCV (Wang, C., Pflugheber, J. (2003)) Este estudo é, pois, um exemplo elucidativo de que, actuando ao nível da tradução, pode encontrar-se a chave para a cura de muitas doenças. Bibliografia • Wang, C., Pflugheber, J..(2003) Alpha Interferon Induces Distintc Translational Control Programs to Supress Hepatitis C Virus RNA Replication. Journal of Virology. pp. 3898-3912, Vol. 77, No. 7 • Nelson D.L., Cox M.M. Lehningher Principles of Biochemistry (4th Ed.). W.H. Freeman and Company. New York. 2005.(p.1034). • G.M. Cooper. "The Cell. A Molecular Approach. (3rd Ed.). AMS Press. U.S.A. 2004. (pp. 281-317). • Alberts et al. "Molecular Biology of the Cell." - (4th Ed.). Inc. New York. 2002. • http://apps.bioneq.qc.ca/twiki/bin/view/knowledgebase/Translation
Baixar