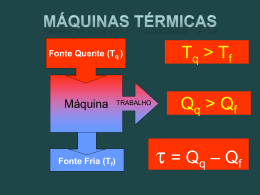
Pró-Reitoria de Graduação Curso Física CARLOS DAde SILVA DOS SANTOS Trabalho de Conclusão de Curso VISÃO TERMODINÂMICA DE ALGUNS PROCESSOS BIOLÓGICOS VISÃO TERMODINÂMICA DE ALGUNS PROCESSOS BIOLÓGICOS Monografia apresentada ao curso de graduação em Licenciatura em Física da Universidade Católica de Brasília, como requisito parcial para a obtenção do Título de Licenciado em Física. Orientador: Henrique Alves Autor: CarlosDr.daPaulo Silva dos Santos Guimarães Orientador: Dr. Paulo Henrique Alves Guimarães Brasília 2010 Brasília - DF 2010 1 CARLOS DA SILVA DOS SANTOS VISÃO TERMODINÂMICA DE ALGUNS PROCESSOS BIOLÓGICOS Monografia apresentada ao curso de graduação em Licenciatura em Física da Universidade Católica de Brasília, como requisito parcial para a obtenção do Título de Licenciado em Física. Orientador: Guimarães Brasília 2010 Dr. Paulo Henrique Alves 2 AGRADECIMENTO Ao Senhor Deus pelos dons da vida e da inteligência, pois, sem os quais e sem Ele, não haveria chegado até aqui. Aos meus pais, pelo apoio e incentivo ao longo dessa jornada. Aos meus amigos por também terem sempre me apoiado. Aos caros colegas, professores, e amigos de faculdade, que não citarei nomes para não correr o risco de esquecer alguém, por me aturarem todos esses anos e contribuírem de forma direta para minha formação. Por todas as discussões que tivemos, pois isso me instigou ainda mais a buscar o conhecimento e a verdade. Às instituições de ensino em que trabalhei, por terem acreditado no meu potencial e deixarem desenvolver habilidades e conhecimentos adquiridos na Universidade. Ao professor Dr. Paulo Henrique Alves Guimarães pela orientação e respaldo na realização deste trabalho. 3 "Como pode a Matemática, sendo produto do pensamento humano, independente da experiência, se adaptar tão admiravelmente aos objetos da realidade?" ALBERT EINSTEIN 4 VISÃO TERMODINÂMICA DE ALGUNS PROCESSOS BIOLÓGICOS CARLOS DA SILVA DOS SANTOS Resumo: Neste trabalho será realizada uma análise do não funcionamento da bomba de sódio-potássio, por meio da Segunda Lei da Termodinâmica. Serão feitas duas análises. A primeira consistirá em calcular a variação da entropia, utilizando-se os valores referentes ao meio interno e ao meio externo da célula. Esse cálculo será realizado tanto para o estado de funcionamento da bomba como para o de não funcionamento da mesma. Na segunda análise será comparado o valor da entropia nos estados inicial e final, mas será considerado somente o interior da célula. As consequências desse estado de não funcionamento para o sistema serão discutidas e uma visão física será abordada para esse processo. PALAVRAS-CHAVE: Entropia. Segunda Lei da Termodinâmica. Bomba de sódio-potássio. 1. INTRODUÇÃO 1.1 APRESENTAÇÃO Processos e sistemas biológicos são complexos e, assim como qualquer outro processo que ocorra na natureza, podem ser explicados pela física e devem obedecer as suas leis. A termodinâmica é o ramo da física que possui maior afinidade com esses sistemas e seu entendimento traz uma visão mais clara do funcionamento deles. Células apresentam mecanismos de transporte ativo que são indispensáveis para a manutenção da ordem local e da vida dela. Estão continuamente consumindo energia para fazê-los funcionar perfeitamente. O não funcionamento deles provocará a morte celular, fenômeno que pode ser comprovado pela termodinâmica, mais especificamente, por meio da Segunda Lei da Termodinâmica. A segunda lei em um dos seus enunciados trata de uma função termodinâmica chamada entropia. Naturalmente, os fenômenos que ocorrem na natureza, passam do estado de menor energia para o estado de maior energia e maior entropia. Para entender bem o comportamento de alguns mecanismos de transporte ativo será necessário o conhecimento sobre eletricidade, pois solutos, geralmente, apresentam carga elétrica e podem interagir com o meio, e com outros solutos, devido a essa propriedade. A bomba de sódio-potássio é um potente, e eficiente, mecanismo de transporte ativo. Trabalha contra os gradientes eletroquímicos dos solutos que transporta e, por esses solutos 5 apresentarem carga elétrica e não serem transportados na mesma quantidade, no mesmo ciclo, a bomba é dita eletrogênica (GUYTON, 1989). Vamos trabalhar em cima da hipótese de não funcionamento dessa bomba e analisar como a termodinâmica está envolvida nesse processo, como se comporta a entropia nesse sistema e nas vizinhanças e a importância da manutenção dela a níveis baixos. O comportamento dos solutos que apresentam carga elétrica frente as interações elétricas presentes no sistema antes e depois do impedimento do funcionamento da bomba. 1.2 JUSTIFICATIVA Tendo em vista que a entropia é um dos conceitos mais difíceis de serem compreendidos dentro da própria Física iremos mostrar uma aplicação prática dela em outros sistemas e outra área do conhecimento, nesse caso, a Biologia. Esse conceito é utilizado por outras áreas do conhecimento, na maioria das vezes, de forma errônea e a definem somente como a medida do nível de desordem de um sistema. Poucas referências existem sobre como ela atua em sistemas biológicos e qual sua importância nos processos que ocorrem nesses sistemas. Outra visão, do funcionamento de alguns sistemas, pode ser dada através da utilização dessa função. 2. SEGUNDA LEI DA TERMODINÂMICA 2.1 HISTÓRICO A Segunda Lei (ou Segundo Princípio) da Termodinâmica apresenta vários enunciados formulados por alguns cientistas, mas todos eles são equivalentes entre si. Esta lei, em várias de suas formulações, explica os fenômenos que permitem uma máquina térmica funcionar. Este princípio não se aplica somente à elas mas também a qualquer outro dispositivo que funcione de maneira semelhante, sendo utilizado também em outras ciências como a Biologia e a Química. Sadi Carnot, engenheiro francês, publicou em 1824 um trabalho que apresenta suas idéias sobre um ciclo termodinâmico que leva seu nome. Baseado nele, desenvolveu um teorema, o qual afirma que: o rendimento de uma máquina térmica funcionando de acordo com o ciclo de Carnot é maior do que o de qualquer outra máquina que opere entre as mesmas temperaturas e que esse rendimento é independente da substância utilizada nas transformações (TIPLER;MOSCA, 2006). 6 A máquina de Carnot opera entre fontes de calor, “frias” e “quentes”, em ciclos formados por quatro etapas reversíveis, podendo ser realizado com qualquer estado da matéria. “Um processo reversível é aquele que, tendo ocorrido, pode ser invertido passando pelos mesmos estados de equilíbrio” (TIPLER; MOSCA, 2006, p. 675). Em termodinâmica, para que um processo seja reversível devem ser obedecidas algumas condições. O processo deve ser quase-estático, isto permite ao sistema estar sempre num estado de equilíbrio e que não haja turbulências. As trocas de calor devem ocorrer entre corpos com temperaturas iguais ou infinitesimalmente próximas e não pode haver forças dissipativas atuando no sistema. Para que o processo possa ser invertido as mudanças na vizinhança também devem ser infinitesimais. Se essas condições não forem obedecidas o processo é dito irreversível. Este pode ser quase-estático ou não. Na natureza a maioria dos processos são irreversíveis. Um exemplo de processo irreversível é o atrito entre dois corpos, como é uma força dissipativa, a energia degradada fica indisponível ao sistema para realização de trabalho (TIPLER; MOSCA, 2006). Na primeira etapa do ciclo de Carnot o calor é absorvido de uma fonte quente em um processo isotérmico. Na segunda etapa há expansão e redução da temperatura até a temperatura da fonte fria em um processo adiabático. Na terceira etapa há liberação de calor para a fonte fria em um processo isotérmico e, por fim, na quarta etapa há uma compressão adiabática até retornar ao estado inicial (ver Figura 1). A Lei da conservação de energia pode ser utilizada para obtermos a relação entre calor e trabalho realizado por uma máquina térmica. Qq W Qf onde Q q é a quantidade de calor fornecida pela fonte quente (calor absorvido), Q f é a energia cedida à fonte fria (calor cedido) e W é o trabalho realizado pela máquina. A quantidade de energia que “entra” no sistema é, em módulo, igual a que “sai”. Fazendo-se uma análise mais detalhada do ciclo vamos obter algumas relações importantes. 7 Figura 1: Diagrama PV do Ciclo de Carnot Da primeira lei da termodinâmica, que é uma lei de conservação de energia, temos que: U (1) Q W ou seja, a variação da energia interna de um sistema é igual ao trabalho realizado por ele subtraído do calor recebido pelo mesmo. A energia interna de um sistema (gás ideal) depende apenas da temperatura em que o mesmo se encontra, dessa forma, a variação da energia interna depende apenas da variação de temperatura. Na primeira etapa do ciclo, há absorção de calor em um processo isotérmico. Como não há variação da energia interna temos que U 0 , portanto Qq W . O trabalho realizado em um processo isotérmico pode ser calculado utilizando-se a seguinte relação: Wisotérmico nRT ln Vf (2) Vi onde: n é o número de mols do gás utilizado no processo, R é a constante universal dos gases perfeitos e T é a temperatura. Para esta etapa temos que o valor do calor recebido é: Qq nRTq ln V2 V1 (2.1) Na segunda etapa temos uma expansão adiabática. Nas transformações adiabáticas não há trocas de calor entre o sistema e o ambiente e vice-versa. Das transformações adiabáticas temos a seguinte relação: TV TqV2 (2.3) 1 T f V3 1 1 C te . Para esta etapa temos: 8 Na terceira etapa, há liberação de calor por um processo isotérmico e temos, novamente, a relação entre trabalho e calor: Q f W . Como o volume final é maior do que o inicial, o calor cedido pelo sistema pode ser calculado da seguinte forma: Qf nRT f ln V3 V4 (2.4) Na quarta etapa, há compressão adiabática, novamente podemos utilizar a relação: TqV1 1 T f V4 1 (2.5) Fazendo-se uma divisão membro a membro com as equações (2.3) e (2.5) podemos obter a seguinte relação: V2 V1 V3 V4 (3) Dividindo-se membro a membro as equações (2.1) e (2.4) e utilizando a relação (3), chegamos à seguinte conclusão: Qq Tq Qf Tf , ou ainda, Qq Qf Tq Tf (4) Esta igualdade é muito útil dentro da termodinâmica, por meio dela podemos definir outras grandezas. William Thomson, conhecido com Lord Kelvin, título que lhe fora concedido, analisando o trabalho de Carnot propôs, através da relação (4), uma escala absoluta de temperaturas. Sabendo que todas as máquinas que operam segundo o ciclo de Carnot tem o mesmo rendimento se funcionarem entre as mesmas temperaturas, percebeu que poderia determinar a temperatura de um corpo Tc . Para isso, colocar-se-ia uma máquina de Carnot em funcionamento entre uma fonte de calor a uma temperatura conhecida T f e o corpo. Sabendo-se a quantidade de calor trocado entre a máquina e o corpo e entre a máquina e a fonte, pode-se calcular o valor da temperatura do corpo utilizando a relação (4). Seguindo esse raciocínio, Kelvin definiu uma escala de temperatura absoluta que leva seu nome, a escala Kelvin, que é utilizada atualmente como escala padrão no Sistema Internacional de Medidas. A obra de Carnot tambem chamou a atenção de Rudolf Clausius, um físico alemão. Clausius estudando detalhadamente o trabalho de Carnot chegou à conclusão que para cada ciclo: 9 “A soma dos calores recebidos ou cedidos pelo corpo, tomados com os respectivos sinais e divididos pelas temperaturas absolutas das fontes de calor que os cedeu ou recebeu, é negativa ou nula.” (CLAUSIUS apud FIOLHAIS C; FIOLHAIS M; GÜÉMEZ, 1998, p. 113). Em termos matemáticos a afirmação de Clausius fica assim: n Qi Ti i 0 Adotando-se uma transformação reversível que leve o sistema de um estado para outro, mas que o processo seja realizado com variações infinitesimais, podemos calcular o somatório das quantidades Q / T desse processo através de uma integral: f i d 'Q T O valor dela só depende dos estados inicial e final e não do processo que leva de um estado a outro. Essa quantidade de Q / T calculada nos somatórios Clausius a definiu como uma nova grandeza e em 1865 batizou-a de entropia, designando a letra S para representá-la. “Se se procura para S um nome característico, pode-se-lhe dar o nome de conteúdo de transformação do corpo (...) Mas eu prefiro ir buscar às línguas antigas os nomes de quantidades científicas importantes, a fim de que eles possam ser os mesmos em todas as línguas vivas; proponho chamar a quantidade S entropia do corpo, do nome grego para transformação. Formei daí a palavra entropia, de maneira que se parecesse com a palavra energia: as duas quantidades têm uma tal analogia no seu significado físico que me pareceu útil uma analogia na sua designação. (CLAUSIUS apud FIOLHAIS C; FIOLHAIS M; GÜÉMEZ, 1998, p. 429). A variação infinitesimal da entropia num processo reversível é dada por: dS d 'Q T (5) Integrando-se ambos os lados com os limites de integração adequados temos: S Q T (5.1) 2.2 ENTROPIA A entropia é uma grandeza que não varia ao longo de processos adiabáticos, portanto é variável de estado de um sistema. Geralmente, quando trabalhamos com essa grandeza tratamos de sua variação, pois assim com a energia potencial o mais importante é medir a variação da quantidade. Utilizaremos, agora, dois exemplos clássicos, uma expansão isotérmica e uma expansão livre ambos com gases ideais. Imagine um gás ideal em certa temperatura T em um 10 recipiente com uma das paredes móvel, fornecendo-lhe calor o mesmo pode expandir-se isotermicamente realizando trabalho sobre a vizinhança. Para calcular a variação da entropia utilizaremos a equação (5.1) e, como num processo isotérmico não há variação da energia interna utilizaremos a relação (2), isso resultará em: S nR ln Vf (5.3) Vi Do ponto de vista do sistema teremos: S sistema Q / T , já que o gás recebeu calor e que o volume final é maior que o inicial. O sinal positivo indica que houve aumento da entropia do sistema. Realizando o mesmo procedimento para a vizinhança encontraremos: S vizinhança Q / T , isso se deve ao fato da vizinhança ter fornecido calor ao sistema e o sinal negativo indica que houve diminuição da entropia da vizinhança. A entropia é uma grandeza extensiva, ou seja, a entropia líquida é a soma da entropia das partes. Para calcularmos a variação da entropia líquida do universo, conjunto formado pelo sistema e suas vizinhanças, devemos somar as suas variações de entropia. A variação de entropia do universo é igual a zero, deste resultado concluímos que a variação da entropia do universo em um processo reversível é nula. Fato que podemos observar da mesma forma no ciclo de Carnot. Suponhamos que este mesmo gás ideal esteja, agora, em um recipiente constituído de paredes fixas e adiabáticas e que contenha dois compartimentos. Estes são separados por uma parede impermeável que nos permite confinar o gás em um dos compartimentos (ver Figura 2). Figura 2: Recipiente de confinamento do gás. Em dado momento retiramos essa parede impermeável, isso permitirá que o gás ocupe todo o volume do recipiente. A expansão livre é um processo isotérmico, a diferença consiste que aquela é irreversível, isso porque depois de uma expansão é quase impossível que o gás retorne espontaneamente ao estado inicial. Na expansão livre de um gás, conforme as 11 condições citadas, não há variação da energia interna, portanto podemos usufruir da equação (5.3). Sendo o volume final maior que o inicial a variação da entropia é positiva e isso significa que ela aumentou. Neste processo não há mudanças nas vizinhanças, então a variação da entropia do gás é a própria variação de entropia do universo, desse procedimento concluímos que em um processo irreversível a entropia do universo sempre aumenta. Através dessas conclusões podemos afirmar que a entropia do universo nunca diminui em qualquer processo. A entropia também é tratada como a medida do nível de desordem de um sistema. Essa grandeza possui uma interpretação probabilística formulada por Ludwig Boltzmann. Está relacionada à probabilidade ou número de estados acessíveis ao sistema. Nesse caso a desordem não tem o mesmo significado que costumamos utilizar, ou seja, desordem não é caos, embora outras áreas do conhecimento assim a utilizem. Em sua forma matemática: S k B ln p onde k B é a constante de Boltzmann e p é uma função que determina a probabilidade de tal evento ocorrer. Quanto maior for o número de estados acessíveis maior será a entropia. Essa equação é interessante porque podemos perceber que um processo irreversível, na verdade, é apenas improvável de acontecer. A expansão livre pode ilustrar bem isso. As moléculas estão em movimento aleatório, nada as impedem de estarem todas do mesmo lado do recipiente. Esse fenômeno é pouco provável que aconteça porque a probabilidade de tal evento ocorrer é pequeníssima. Se isso ocorresse, a entropia do universo diminuiria, fato que tem chance ínfima de ocorrer na natureza, sendo não observado. 3. CARGA ELÉTRICA E SUAS INTERAÇÕES Carga elétrica é uma característica intrínseca de algumas partículas elementares. Ela pode ser positiva ou negativa. É uma grandeza quantizada, ou seja, quase sempre aparece em um múltiplo inteiro de um valor denominado unidade fundamental de carga elétrica. A unidade fundamental é representada por e e tem módulo igual a 1,602176 x10 19 Coulomb (C). Os quarks, que são partículas elementares que formam os bárions e os mésons, apresentam carga elétrica 1 e e 3 2 e , no entanto estas partículas não aparecem de forma 3 isolada, mas sempre de forma que a carga total seja múltiplo da carga elementar. Os átomos são formados por partículas que apresentam carga elétrica. O próton possui carga e , o nêutron tem carga nula e o elétron possui um valor de carga elétrica igual a e. 12 Em geral, os átomos são eletricamente neutros, pois possuem a mesma quantidade de prótons e elétrons. Quando um átomo tem uma quantidade maior de prótons ou elétrons, apresentará carga elétrica resultante não nula. Isso também pode ocorrer com moléculas e em corpos de escala macroscópicas, eles podem ser eletrizados tirando-se elétrons dos átomos que os constituem, deixando-os assim eletrizados positivamente. Outro corpo que receber esses elétrons ficará eletrizado negativamente. A quantidade de carga elétrica no universo é conservada em qualquer fenômeno. Esta é a lei de conservação da carga elétrica que é uma das leis fundamentais da natureza. Assim como a massa, a carga elétrica também pode alterar as características do espaço ao seu redor. A massa cria um campo gravitacional de modo que todas as partículas e/ou corpos que possuem massa estarão sujeitas à ação de uma força gravitacional. Analogamente, a carga elétrica cria ao seu redor um campo elétrico de modo que todas as partículas carga que possuem carga elétrica estarão sujeitas a uma força elétrica. Essas duas forças são forças de campo (ou forças de interação à distância), ou seja, não é necessário haver contato entre as partículas para existir força. A diferença entre elas consiste no fato de que a força gravitacional é somente atrativa e força elétrica é atrativa ou repulsiva, isso dependerá do sinal das cargas. Cargas de mesmo sinal interagem entre si através de uma força de repulsão e cargas de sinais opostos interagem entre si através de uma força de atração. O campo elétrico é representado por meio de linhas de força. Estas, por convenção, se orientam de modo a “saírem” da carga positiva e “entrarem” na carga negativa. Indicando o sentido da força exercida em uma carga de prova positiva inserida no campo. “A força eletrostática é conservativa e, portanto, existe uma energia potencial associada com as configurações (posição relativa dos objetos) de um sistema no qual forças eletrostáticas agem.” (HALLIDAY; RESNICK; KRANE, 2004, p. 75). A variação da energia potencial de um sistema pode ser descrita como sendo igual ao negativo do trabalho para movê-lo entre duas posições.. b b U Ub Ua Wab F .ds q 0 E.ds a a (6) O valor dessa integral é independente do caminho adotado, dependendo apenas dos pontos inicial e final do sistema. Diferente da força e do campo elétrico a energia potencial não é uma grandeza vetorial e sim escalar, pois ela é o resultado do produto escalar entre dois vetores (duas grandezas vetoriais). Existe outra função escalar, associada ao campo elétrico, o potencial elétrico. Ele é, por definição, a quantidade de energia ganha ou perdida por cada unidade de carga. 13 V U q0 A unidade de potencial elétrico é a de energia por unidade de carga, ou seja, J / C , chamada de volt V . O potencial elétrico é uma propriedade que depende apenas da distância que está da carga geradora de campo, ou seja, é uma propriedade do ponto. Suponhamos um campo elétrico uniforme criado por uma carga elétrica, a ele estão associado vários valores de potenciais elétricos que dependem apenas da distância da carga geradora. Podemos colocar uma carga de prova em qualquer ponto desse campo e ela terá um valor de potencial elétrico, 1V por exemplo, ela realizará, ou sofrerá realização de trabalho de 1 joule para cada unidade de carga que possuir. Retirando-se essa carga de prova o potencial ainda existirá e terá o mesmo valor de antes. O potencial elétrico, geralmente e convenientemente, é ajustado como zero em um ponto infinitamente distante do sistema de cargas geradoras de campo. Dependendo apenas do valor da carga geradora e da distância de que se está dela, podem existir vários pontos onde o potencial elétrico apresenta o mesmo valor, formando o que chamamos de superfícies equipotenciais. Elas são perpendiculares as linhas de campo elétrico. Uma carga elétrica ao se mover sobre essas superfícies não realiza ou não é realizado trabalho sobre ela. O valor do potencial elétrico pode ser positivo, negativo ou nulo. Onde o potencial é nulo, não significa que o campo elétrico ou a força elétrica sejam nulos. Em um sistema com cargas pontuais a visualização das superfícies equipotenciais é simples. Figura 3: Superfícies equipotenciais. Fonte: Propriedades das Superfícies Equipotenciais (UnB) Assim como a energia potencial o mais importante é medir a variação da grandeza, ou seja, a diferença de potencial elétrico (ddp), conhecida como tensão elétrica. 14 V U q0 b a E.ds (7) Uma bateria convencional de automóveis tem uma ddp de 12V entre seus terminais, isto não significa, necessariamente, que o pólo positivo tem um potencial de 12V e o negativo 0V. O pólo positivo está a um potencial 12V maior que o pólo negativo. A ddp possibilita o deslocamento de cargas nos condutores. Se ligarmos um fio condutor na bateria citada acima, teremos uma tensão elétrica de 12V aplicada entre as extremidades do mesmo. As cargas, pelo sentido convencional, tenderão ir em direção da diminuição do potencial, ou seja, do positivo para o negativo. Esse fenômeno do movimento das cargas elétricas chamamos de corrente elétrica. 4. MEMBRANAS E TRANSPORTE 4.1 MEMBRANA PLASMÁTICA As membranas definem o espaço físico ocupado pela célula e regulam a entrada e saída de moléculas. O trânsito é regulado por sua permeabilidade, podendo, dessa forma, reter algumas substâncias no meio intracelular e excluir outras. Ela não é miscível com o líquido intracelular (LIC) e nem com o líquido extracelular (LEC), atuando como barreira para a maioria das moléculas hidrossolúveis, no entanto, substâncias lipossolúveis, como os gases e alcoóis, podem passar facilmente através dela. Atualmente o modelo mais aceito para a estrutura da membrana celular é o mosaico fluido. A unidade estrutural básica é composta por fosfolipídios que formam uma bicamada, elástica e muito fina, onde as regiões não polares estão orientadas para o centro da bicamada e os grupos polares para o exterior. Há várias moléculas de proteínas incrustadas nessa bicamada, sendo algumas integrais, atravessando toda a bicamada, e algumas periféricas, aderidas somente em um lado da bicamada. Por conta disso, a bicamada lipídica não possui simetria, tanto estrutural quanto funcional. (GUYTON, 1989) As proteínas integrais atuam como canais estruturais entre o LIC e o LEC pelos quais podem passar substâncias hidrossolúveis e íons. Elas possuem propriedades seletivas que permitem somente a passagem de algumas substâncias. Algumas atuam como carreadoras transportando moléculas grandes demais para passarem pelos orifícios da membrana. As proteínas periféricas atuam, quase que exclusivamente, como enzimas e limitando a mobilidade de proteínas integrais fazendo a ligação delas com estruturas intracelulares. 15 4.2 TRANSPORTE ATRAVÉS DA MEMBRANA A barreira imposta pela bicamada lipídica define, de certa forma, a composição do LIC e do LEC. A quantidade de cada substância no LIC pode ser controlada e é bem diferente das concentrações do LEC. A quantidade de sódio e cloreto no LEC é muito maior que no LIC, entretanto a quantidade de potássio e fosfatos presentes no LIC é muito superior a quantidade presente no LEC. A tabela abaixo mostra isso muito bem. Tabela 1 – Composição do LIC e LEC Na K Ca 2 Mg 2 Cl HCO 3 Fosfatos SO 42 Glicose Aminoácidos pH Proteínas Fonte: GUYTON (1989) LEC 142 mEq/L 4 mEq/L 5 mEq/L 3 mEq/L 103 mEq/L 28 mEq/L 4 mEq/L 1 mEq/L 90 mg% 30 mg% 7.4 2g% (5mEq/L) LIC 10 mEq/L 140 mEq/L <1 mEq/L 58 mEq/L 4 mEq/L 10 mEq/L 75 mEq/L 2 mEq/L 0 a 20 mg% 200 mg% 7.0 16g% (40 mEq/L) O transporte através da membrana pode ser realizado através de dois mecanismos básicos: a difusão, também chamada de transporte passivo, e o transporte ativo. A difusão depende do movimento aleatório das partículas, seja pelos orifícios entre a membrana ou pelas proteínas transportadoras. Íons e moléculas estão em constante movimento aleatório devido à temperatura do sistema em que se encontram. Este movimento recebe o nome de movimento browniano e é proporcional a k BT , onde k B é a constante de Boltzmann e T é a temperatura, medida em kelvin, que o sistema se encontra. A difusão, ou transporte passivo, pode ser simples ou facilitada. A difusão simples se deve ao movimento browniano e se dá através dos orifícios da membrana ou pelos canais aquosos de algumas proteínas sem a necessidade de energia extra ou fixação em proteínas carreadoras. O que faz com que uma substância se difunda pela membrana é a diferença de concentração entre os compartimentos intracelular e extracelular. Essa diferença de concentração gera pressão, maior no lado mais concentrado, e isso faz o soluto se movimentar através da membrana na tentativa de igualar as concentrações. Esse tipo de transporte ocorre com compostos não polares. (GUYTON, 1989) A difusão facilitada, ao contrário da difusão simples, necessita de proteína carreadora específica para realizar o transporte de substâncias através da membrana, ou seja, a proteína 16 irá facilitar a difusão da substância. A substância a ser transportada fixa-se no canal, em seguida há alteração conformacional e o canal se abre do outro lado da membrana. Os canais apresentam “comportas” que podem ser ativadas por ddp (voltagem dependentes) ou por fixação de outra molécula de proteína, chamada de ligando (ligando dependente). Esses canais são bastante seletivos, permitindo a passagem somente de substâncias que apresentam maior permeabilidade ao canal. (GUYTON, 1989) As substâncias presentes no nosso organismo estão em concentrações muito diferentes no LIC e no LEC. Naturalmente, as moléculas tendem a passar do meio mais concentrado para o meio menos concentrado, mas em alguns processos é necessário ser feito exatamente o oposto. Moléculas podem se mover a favor do gradiente eletroquímico e seu retorno é praticamente impossível. Para que essas moléculas retornem ao meio mais concentrado é necessário fornecer energia. O transporte ativo necessita de energia externa e é realizado contra o gradiente eletroquímico. (GUYTON, 1989) O transporte ativo depende, além de energia, de proteínas carreadoras. A energia utilizada nesse processo advém da adenosina trifosfato (ATP). “As proteínas carreadoras do transporte ativo possuem atividade de ATPase, o que significa que podem clivar o ATP, para formar adenosina difosfato (ADP) ou adenosina monofosfato (AMP), com liberação de energia das ligações de fosfato de alta energia.” (GUYTON, 1989, p. 80). O transporte ativo pode ser classificado em transporte ativo primário e transporte ativo secundário. No transporte ativo primário a substância é acoplada à proteína carreadora pelo fornecimento de energia e através disso é “bombeada” de um meio para outro. O transporte ativo secundário ocorre quando dois solutos diferentes são transportados pela mesma proteína carreadora. Quando são transportados no mesmo sentido denomina-se cotransporte, e quando é realizado em sentidos opostos é denominado contratransporte. O transporte ativo secundário só ocorrerá se as duas substâncias estiverem fixadas na proteína carreadora. 5. DIFERENÇAS DE POTENCIAIS TRANSMEMBRANA 5.1 POTENCIAL DE REPOUSO O LIC e o LEC são soluções eletrolíticas, ou seja, são compostas por partículas que apresentam cargas elétricas, e contém a mesma quantidade de íons positivos e negativos sendo assim eletricamente neutros. (GUYTON, 1989) 17 Há dois mecanismos básicos que levam ao desenvolvimento de potenciais de membrana, a difusão de íons através da membrana e o transporte ativo de íons. Esses dois mecanismos criam desequilíbrio entre as cargas elétricas. No LIC há muitas moléculas com carga negativa. Elas são grandes para se difundirem pela membrana e acumulam-se na face interna dela. Uma quantidade de íons positivos acumula-se na face externa da membrana. Com a diferença de cargas entre a membrana, será gerada uma diferença de potencial elétrico, chamada de potencial de membrana. (GUYTON, 1989). Supondo a célula permeável somente ao sódio, ele tenderá a entrar na célula devido à diferença de concentração. Haverá um momento em que a difusão cessará devido a neutralização das forças que permitem o íon se difundir, a força elétrica e a força devida ao potencial químico. Elas terão mesmo módulo, porém sentidos opostos, neste momento podemos calcular o potencial de equilíbrio através da equação: V RT C externa ln zF C int erna (8) Em que C representa as concentrações do meio interno e externo, z é a valência do íon e F é a carga de um mol de partículas, que equivale a 96500 C. Este cálculo dará um valor por volta de +61 mV (GUYTON, 1989). Essa é a ddp de equilíbrio do sódio. Fazendo a mesma suposição para o potássio, este tenderá a sair da célula devido ao gradiente de concentração. Quando as forças que causam a difusão se neutralizarem podemos utilizar a equação (8) para calcular a ddp de equilíbrio do potássio. O valor da ddp será de aproximadamente -94 mV (GUYTON, 1989). Conhecendo-se a ddp de equilíbrio de cada íon podemos calcular o valor do potencial de membrana. Existe um mecanismo de transporte ativo de sódio e potássio, a Na K ATPase, ou bomba de sódio-potássio, que também contribui para o potencial de membrana. Ela bombeia simultaneamente três íons de sódio para fora da célula e dois íons de potássio para dentro. A Na K ATPase troca íons de mesma carga elétrica, efetivamente, a cada ciclo ela “joga” uma carga positiva para o meio externo, criando um acúmulo na parte externa da membrana e isto criará ddp na mesma. Esta bomba é eletrogênica porque pode criar diferença de potencial devido ao seu funcionamento. (GUYTON, 1989). Com todos esses fatores influenciando, a diferença de potencial “resultante” de membrana será algo em torno de -90 mV, ou seja, o lado interno está a um potencial de 90 mV menor que o lado externo. 18 5.2 POTENCIAL DE AÇÃO Algumas células são excitáveis, ou seja, são capazes de propagar variações de potencial, chamados potencial de ação. Sinais neurais são transmitidos dessa forma através de células nervosas. Quando a célula está no estado de repouso diz-se que ela está polarizada. No repouso a membrana celular é bem mais permeável ao potássio do que ao sódio, isso permite que o potássio saia da célula, “levando com ele” carga positiva. Os canais de sódio, assim como os canais de potássio, são voltagem dependentes. Em um dado momento a ddp entre a membrana diminuirá o bastante para ativar os canais de sódio, aumentando a permeabilidade ao sódio e permitindo a entrada dele na célula. Com a entrada do sódio o interior da célula fica cada vez mais positivo, podendo ultrapassar o valor de 0 mV e chegar próximo do valor do potencial de equilíbrio do sódio, essa etapa é chamada despolarização. Ao chegar a certo valor de ddp os canais de sódio são inativados e ele não pode mais entrar na célula. Quase simultaneamente os canais de potássio se abrem permitindo a entrada desse íon. Esses processos levam a célula de volta ao potencial de repouso, sendo chamado de repolarização. Esses dois canais atuam em conjunto com a Na K ATPase na geração do potencial de ação e na repolarização da membrana. (GUYTON, 1989). 6. A CÉLULA E A TERMODINÂMICA 6.1 SISTEMAS ABERTOS Sistemas abertos são aqueles que permitem trocas de energia e matéria com o meio externo. Ao tratar esse tipo de sistema algumas modificações são necessárias nas equações que descrevem as funções termodinâmicas. Novas variáveis também são necessárias pra descrevê-las. Quando se adiciona ou retira matéria do sistema a energia interna sofrerá alterações, e ela se tornará dependente do número de mols trocados. A energia interna pode ser descrita como: dU dn Em que µ é o chamado potencial químico e dn representa a variação infinitesimal do número de mols, “[...] o princípio do potencial químico fornece significado à grandeza conjugada com o número de moles: o potencial químico indica como varia a energia com o número de moles.” (FIOLHAIS, 1998, p.242). 19 Através da função geral da energia interna é possível determinar a entropia do sistema, mesmo que haja variação no número de mols. A variação da entropia pode ser encontrada a partir da seguinte equação: dS dU T PdV T dn T (9) Sistemas biológicos são exemplos de sistemas abertos, estão sempre trocando energia e matéria com o meio externo, desde o nível celular até o nível de sistemas. Tratando-se somente da célula, podemos observar que ela sempre está trocando energia e matéria com o meio exterior, seja por transporte ativo ou passivo. Esse sistema trabalha em desequilíbrio com o meio externo. A própria vida pode ser considerada um fenômeno que acontece devido a esse desequilíbrio. Isso é o que diferencia seres com vida de objetos inanimados. Estes, desde que não haja nenhuma interferência externa, estão em equilíbrio com o ambiente. O equilíbrio de um sistema biológico é dinâmico, não pode ser estático, pois se assim fosse, não haveria vida. Algumas áreas do conhecimento tratam a entropia simplesmente como a medida do nível de desordem de um sistema. Esse tipo de interpretação pode ser utilizada, mas com certa precaução. Essa visão será útil para o entendimento de algumas questões sobre os sistemas biológicos. A célula troca matéria e consome muita energia para manter a entropia local baixa, ou seja, manter o nível de organização mais alto possível. Evidente que para manter esse alto nível de organização a célula requer gasto energético, isso possibilita a organização local à custa do aumento da entropia do meio externo, estando esse processo de acordo com a segunda lei da termodinâmica: em qualquer processo a entropia do universo nunca diminui. Um exemplo básico da manutenção da “ordem” no interior da célula é o funcionamento da bomba de sódio-potássio, um mecanismo de transporte ativo. Ela, a cada ciclo, repõe o potássio que saiu da célula e retira o sódio que entrou. Este processo é possível devido à energia fornecida pelo ATP, já que o bombeamento desses íons é feito contra o gradiente eletroquímico. O seu funcionamento é imprescindível para que a célula possa manter-se viva. 6.2 RELAÇÃO ENTRE A ENTROPIA E A VIDA CELULAR A inativação da bomba de sódio e potássio pode ser feita a partir de uma droga, a dopamina. “A dopamina inibe a atividade da NA-K-ATPase no túbulo proximal (10 –30%) na alça ascendente de Henle, no túbulo distal e no ducto coletor”. (BELICK; COSTA, 1996) 20 Suponhamos que a Na / K ATPase tenha seu funcionamento impedido. Uma das funções dela é manter as concentrações de sódio de potássio em níveis normais. O potássio, devido à permeabilidade da membrana, irá sair da célula e certa quantidade de sódio irá entrar. Até que a concentração de sódio no LIC seja igual ao LEC esse soluto continuará a entrar na célula. Quando as concentrações tornarem-se iguais, a célula irá arrebentar, pois seu volume é pequeno para suportar o volume de água necessário para que o equilíbrio seja estabelecido. A entropia desse sistema irá aumentar demasiadamente visto que esse processo é irreversível mesmo existindo canais permeáveis a esses solutos. A explosão da célula tambem é um fator que justifica a irreversibilidade desse processo. Não havendo explosão, deve ser realizado trabalho pra que se possa fazer o sistema voltar ao estado inicial, porque ele não retornará espontaneamente. Podemos fazer uma analogia desse sistema constituído pela célula, com o funcionamento da bomba impedido, com um recipiente constituído de paredes fixas e adiabáticas, contendo dois compartimentos. Estes são separados por uma parede semipermeável e flexível como a própria membrana celular. Os volumes dos dois compartimentos serão considerados iguais, assim poderemos ter uma noção, em média, da quantidade de partículas que compõe cada um deles. Apresentarão valor V, igual ao volume celular. O LIC e o LEC seriam análogos aos compartimentos e a membrana juntamente com seus canais seria a parede semipermeável. Nas bordas da parede, assim como na membrana, é estabelecida uma ddp com intensidade igual ao potencial de repouso. Continuando nessa linha de raciocínio, podemos aproximar o comportamento dos íons ao de um gás ideal, já que o tamanho deles é desprezível em relação ao volume da célula e ao volume do LEC. Não há reação entre sódio e potássio, pois, quando ionizados, apresentam configurações estáveis iguais aos gases nobres. Esses solutos irão ocupar os compartimentos da caixa, cada um na sua concentração original. Utilizaremos a equação (9) para calcularmos a entropia desse sistema. A medida será feita no início do processo, com a bomba funcionando perfeitamente, e no fim, com a interrupção do funcionamento da bomba. A concentração de cada soluto é apresentada na Tabela 1 em miliequivalentes por litro de solução. Podemos encontrar o número de partículas para determinada concentração através da expressão: C zn V (10) 21 No caso do sódio e do potássio z tem valor igual a um, pois são monovalentes. A célula, em seu pleno funcionamento, apresenta um nível de organização bastante elevado. A esse nível de organização há um valor de entropia associado. Este valor, quando comparado com outras situações, como a de perfeito funcionamento da bomba, é baixo. O exterior da célula, da mesma forma, apresenta organização e valor de entropia associado. O corpo, ou qualquer outro sistema vivo, tende a manter a temperatura constante. Na maioria das vezes há um gradiente de temperatura, mas, em média, a temperatura é a mesma para locais próximos. Consideraremos constante a temperatura, o potencial químico, visto que o sódio não reagirá com outros elementos nesse processo e o volume (antes que a célula exploda). Tendo sido feito essas considerações a equação (9) pode ser reduzida a: dS dn T (11) Integrando-se ambos os lados e utilizando como limites dois estados diferentes teremos a seguinte equação: S T (n 2 (11.1) n1 ) O número de mols contidos no sistema em cada estado pode ser trocado pela concentração conhecendo-se também o volume através da equação (10). O volume dos dois compartimentos são iguais, como foi estabelecido, e sabemos que o sódio é monovalente. Então a equação acima pode ser reescrita como: S V T (C 2 (11.2) C1 ) A variação da entropia numa primeira análise, com o perfeito funcionamento da bomba de sódio-potássio, pode ser medida através dos valores da entropia do meio externo e do meio interno (os compartimentos da caixa). Fazendo S S2 S1 com S 2 representando o valor da entropia do meio interno e S 1 representando o valor da entropia do meio externo, e substituindo os respectivos valores das concentrações obteremos, em média, o seguinte valor para a entropia: S 132 V T Isso indica que entropia em um dos compartimentos é maior, e essa situação é realmente necessária para que o sistema possa trabalhar perfeitamente, mantendo a célula viva. 22 Seguindo a hipótese da interrupção do funcionamento da bomba, as concentrações dos meios interno e externo tenderão a se igualar. Pensando na entropia como a medida do nível de desordem de um sistema, nessa situação, o meio interno tende a igualar o nível de organização com o meio externo. Sendo assim, o valor da entropia, associado ao sódio, do meio interno será igual à do meio externo. Podemos comprovar isso através da equação (11.2). O resultado será: S 0J / K Esse valor indica a morte celular, pois o nível de organização dos dois meios (ou compartimentos) são iguais. No estado de não funcionamento da bomba a força motriz e a ddp, ambas criadas pela diferença de concentração, não existirão mais. A célula não poderá realizar trabalho para retornar ao estado inicial, porque os mecanismos responsáveis por essa tarefa não funcionam. Se não há força não há realização de trabalho. Vimos que a célula irá explodir devido ao volume de água necessário para igualar a concentração com o meio externo. Não faz sentido existir célula e meio externo se aquela não funciona mais, a explosão transforma os dois compartimentos em um só meio. Outra análise pode ser feita admitindo somente o meio interno à célula. Nesse caso calcularemos a variação da entropia a partir dos estados final e inicial da célula. Faremos S S final S Inicial . Lembrando-se de que o estado final é o de não funcionamento da ATPase. Na / K O valor da concentração do estado final deve ser igual ao valor da concentração no estado inicial. Seguindo o modelo e a proposta apresentada, no estado final, a célula apresentará um valor de concentração igual a 76 mEq/L, este valor pode ser obtido somandose os valores das concentrações do LIC e do LEC e dividindo-o para os dois compartimentos. Calculando a partir da equação (11.2) e com base em tudo que já foi dito a respeito da concentração dos dois estados teremos, em média, para o modelo apresentado, o seguinte valor para a variação da entropia na célula: S S final S Inicial 66 V T Isso valida o modelo apresentado, pois foi defendido que a entropia, no estado de não funcionamento da bomba de sódio e potássio, aumentaria no interior da célula. Segundo o modelo, a entropia do meio externo diminui e a do meio interno aumenta. A verificação deste aumento é o mais importante nessa análise. Podemos perceber nitidamente que o valor da variação da entropia é alto, mostrando que o valor dela no estado final é maior que no estado 23 inicial. Portanto, ao atingir esse nível de entropia, ou o valor acima para a variação da mesma, a célula morrerá. Em células excitáveis, além dela não poder realizar trabalho para se manter viva, também não poderá gerar e propagar potenciais de ação. Com a interrupção da propagação dos potenciais de ação, informações necessárias para o perfeito funcionamento dos sistemas, e consequentemente do corpo, ficarão comprometidas. Nesta situação também poderá ser evidenciado aumento na entropia. 7. CONCLUSÃO O funcionamento de mecanismos de transporte ativo é importantíssimo em sistemas que apresentam vida. Eles mantém o sistema funcionando perfeitamente dentro padrões estabelecidos, pelo próprio sistema, como normais. O consumo de energia é indispensável, pois, como vimos, os processos na natureza tendem a ocupar o estado de menor energia e maior entropia. Esses mecanismos trabalham contra gradientes eletroquímicos e quando os “transportados” apresentam carga elétrica, o transporte deve ser realizado contra o sentido do campo elétrico, no sentido do maior potencial elétrico. Os sistemas biológicos são sistemas considerados complexos, mas ainda obedecem às leis físicas da termodinâmica e do eletromagnetismo. Observarmos que o não funcionamento de um mecanismo de transporte ativo como a Na / K ATPase certamente levará a célula ao estado de morte devido ao aumento da entropia do sistema. O aumento dessa grandeza, quando comparado os estados inicial e final, é inevitável nessa situação. Foi mostrado ainda que, quando a bomba tem seu funcionamento impedido, a tendência da célula é igualar a sua entropia como o meio externo (vizinhança) que nesse caso é o LEC. O nível de organização dos dois sistemas torna-se o mesmo e a célula explode, tornando o processo irreversível. Sendo assim, todo o processo está de acordo com a Segunda Lei da Termodinâmica. Esses sistemas estão sempre trabalhando para manter a entropia baixa. O aumento dessa função termodinâmica, que é um processo natural, causará a morte. Talvez essa seja uma possível explicação do porquê de sistemas que apresentam vida irão, em algum momento, morrer. REFERÊNCIAS 24 BELIK, J.; COSTA, Karina N.; Suporte circulatório neonatal: Agentes farmacológicos. Trabalho apresentado no XI Congresso Brasileiro de Perinatologia. 1996, Rio Grande do Sul. Disponível em: <http://www.paulomargotto.com.br/documentos/farm_card.doc>. Acesso em: 25 jun. 2010 GÜÉMEZ, Julio; FIOLHAIS, Carlos; FIOLHAIS, Manuel. Fundamentos Termodinâmica do equilíbrio. 1ª edição, Ed. Fundação Calouste Gulbenkian, 1998. de GUYTON, Arthur C.; Tratado de fisiologia médica. 7ª edição, Editora Guanabara Koogan S.A.; Rio de Janeiro, 1989. HALLIDAY, David; RESNICK, Robert; KRANE, Kenneth S.; Física 3. 5ª edição, LTC, Rio de Janeiro, 2004. MUNDIM, Kleber C.; Propriedades das Superfícies equipotenciais. UnB, Agosto de 1999. Disponível em: <http://vsites.unb.br/iq/kleber/EaD/Eletromagnetismo/EquipotenciaisProp/EquipotenciaisP rop.html>. Acesso em: 14 mar. 2010. TIPLER, Paul; MOSCA, Gene; Física para cientistas e engenheiros – Volume 1 – Mecânica, oscilações e ondas e termodinâmica. 5ª edição, LTC, Rio de Janeiro, 2005. BIBLIOGRAFIA FEYMMAN, Richard Phillips; Lições de Física – Volume 1, 2ª edição, Ed. Bookman Companhia Editora ltda, 2008. GÜÉMEZ, Julio; FIOLHAIS, Carlos; FIOLHAIS, Manuel. Fundamentos Termodinâmica do equilíbrio. 1ª edição, Ed. Fundação Calouste Gulbenkian, 1998. de GUYTON, Arthur C.; Tratado de fisiologia médica. 7ª edição, Editora Guanabara Koogan S.A.; Rio de Janeiro, 1989. HALLIDAY, David; RESNICK, Robert; KRANE, Kenneth S.; Física 3. 5ª edição, LTC, Rio de Janeiro, 2004. MUNDIM, Kleber C.; Propriedades das Superfícies equipotenciais. UnB, Agosto de 1999. Disponível em: <http://vsites.unb.br/iq/kleber/EaD/Eletromagnetismo/EquipotenciaisProp/EquipotenciaisP rop.html>. Acesso em: 14 mar. 2010. OLIVEIRA, Mário José de; Termodinâmica. Editora Livraria da Física, São Paulo, 2005. SERWAY, Raymond A.; JEWETT, John W; Princípios de Física: Movimento ondulatório e Termodinâmica – Volume 2. 3ª edição, Thomson Learning Edições, São Paulo, 2006. TIPLER, Paul; MOSCA, Gene; Física para cientistas e engenheiros – Volume 1 – Mecânica, oscilações e ondas e termodinâmica. 5ª edição, LTC, Rio de Janeiro, 2005. 25
Baixar