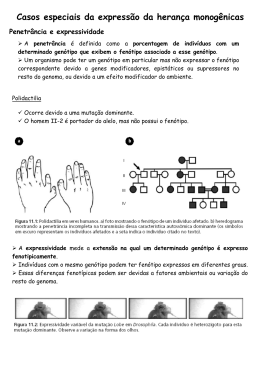
Genes Ligados Sutton (1903): “se houvesse apenas um gene em cada cromossomo, o número de características diferentes de um indivíduo não poderia exceder o número de cromossomos nos gametas, o que é indubitavelmente contrário aos fatos” Portanto, deve haver muitos genes em um mesmo cromossomo. Sutton (1903): “se os cromossomos retêm permanentemente as suas individualidades, então todos os alelomorfos presentes em um mesmo cromossomo devem ser herdados juntos” E a lei da segregação independente? Testando a 2ª lei: Segregação Independente x Ligação Gênica Os resultados observados em cada cruzamento estão de acordo com o esperado pela lei da segregação independente? Como você explicaria a ocorrência desse desvio da proporção esperada? Por que os fenótipos encontrados nos indivíduos da prole de cada cruzamento-teste são os mesmos fenótipos dos indivíduos parentais? Resultado esperado havendo segregação independente... AaBb Genótipos produzidos pela associação dos gametas masculinos e femininos... EMPARELHAMENTO TIPO I EMPARELHAMENTO TIPO II (tipo não parental) (tipo não parental) Fase de ligação: configuração cis (ou acoplamento) = AB/ab configuração trans (ou repulsão) = Ab/aB Cruzamentos-teste em que os genes apresentaram ligação completa: Por convenção, os alelos que se encontram em um cromossomo são separados por uma barra daqueles localizados no cromossomo homólogo. Ex.: DE/de ou De/dE, A forma mais comum de ligação gênica é a ligação incompleta, onde, apesar de os genes estarem localizados no mesmo cromossomo, gametas recombinantes podem ser produzidos durante a meiose de indivíduos heterozigóticos. Como isto é possível? A DESCOBERTA DA LIGAÇÃO GÊNICA 1905 - William Bateson, Edith Saunders e Reginald Punnett publicaram os resultados de experimentos em que cruzavam variedades de ervilhas-de-cheiro que diferiam quanto à cor da flor e ao tamanho proporção observada na F2 desse cruzamento não estava de acordo com a proporção fenotípica esperada pela 2ª Lei de Mendel Desvio da proporção esperada (1:1:1:1)! Parecia existir duas regras: (1) (2) As classes fenotípicas mais comuns são dos tipos parentais e apareciam na freqüência de 43,7% para cada classe. As classes fenotípicas recombinantes também apresentavam freqüências iguais (6,3% para cada classe), mas bem menores do que o esperado. Bateson e seus colaboradores concluíram que os genes que condicionam estas duas características, de alguma forma, apresentavam um acoplamento. Contudo, este acoplamento não era total, pois classes fenotípicas recombinantes também foram observadas na prole desses cruzamentos. Genes Ligados PERMUTAÇÃO E FREQÜÊNCIA DE RECOMBINAÇÃO O processo de troca entre segmentos das cromátides não-irmãs dos cromossomos homólogos acontece no início da prófase da primeira divisão meiótica e é chamado de permutação (ou do inglês, crossing-over). Se em todas as células meióticas de um indivíduo houver permutação entre dois genes ligados (100% de permutação), serão produzidos 50% de gametas recombinantes para esses genes. Portanto, a freqüência de recombinação (FR) máxima entre dois genes quaisquer será 50%! Você se lembra de qual é a freqüência de recombinação para dois genes com segregação independente? Como podemos fazer para calcular a FR entre dois genes ligados? Durante a divisão celular, quanto mais distantes, em um mesmo cromossomo, dois genes estiverem, maior a probabilidade de que ocorra uma permuta entre eles em cada uma das meioses que darão origem ao seu conjunto de gametas. Logo, maior será a freqüência com que serão produzidos os gametas recombinantes. Não é possível ver os genes, mas é possível fazer uma estimativa da distância entre dois genes que estão em um mesmo cromossomo através da freqüência de indivíduos com fenótipo recombinante que surgem na prole de um cruzamento-teste. A FR é calculada através da porcentagem de indivíduos com fenótipos recombinantes, ou seja, a porcentagem de indivíduos com fenótipos diferentes dos fenótipos das linhagens parentais. Para chegar a esse valor, precisamos somar o número de indivíduos observados nas duas classes fenotípicas recombinantes: selvagemmutante B e mutante A-selvagem, e dividir essa soma pelo total de indivíduos analisados (2.695 indivíduos). Assim, FR entre A e B = (79 + 82) / 2.695 = 0,06 ou 6,0%. OS GRUPOS DE LIGAÇÃO corresponde ao conjunto de genes presente em um cromossomo. Uma propriedade interessante da análise da FR é a previsão do número de indivíduos que esperamos encontrar, em cada classe fenotípica, nos cruzamentos envolvendo genes ligados. Por exemplo, se a FR entre dois genes é 30%, para cada 100 indivíduos esperamos encontrar 15 em cada uma das classes recombinantes (0,30/2) e 35 em cada uma das classes parentais (100% – 30% = 70%; 0,70/2 = 35). Desse modo, após calcularmos a FR entre dois genes em um experimento, podemos utilizar esse valor para prever os resultados esperados em cruzamentos futuros.
Baixar