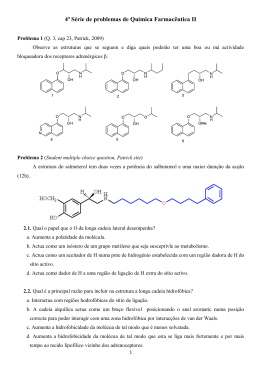
ANDERSON RICARDO SOARES LIGNIFICAÇÃO DE RAÍZES DE SOJA SOB AÇÃO DE L-3,4-DIIDROXIFENILALANINA (L-DOPA) Dissertação apresentada ao Programa de Pósgraduação em Ciências Biológicas (área de concentração Biologia Celular) da Universidade Estadual de Maringá, para obtenção do grau de Mestre em Ciências Biológicas. Maringá Abril – 2006 Dados Internacionais de Catalogação-na-Publicação (CIP) (Biblioteca Central - UEM, Maringá – PR., Brasil) S676L Soares, Anderson Ricardo Lignificação de raízes de soja sob a ação de L-3,4diidroxifenilalanina (L-Dopa) / Anderson Ricardo Soares. – Maringá, PR : [s.n.], 2006. 45 f. : il. color. Orientadora : Prof. Dr. Maria de Lourdes Lucio Ferrarese. Dissertação (mestrado) - Universidade Estadual de Maringá. Programa de Pós-graduação em Ciências Biológicas, 2006. 1. Soja - Alelopatia (L-DOPA). 2. Soja - Lignina Determinação. 3. Soja - Raízes - Crescimento. 4. Soja Compostos fenólicos - Determinação. 5. Soja - Peroxidases Determinação das atividades. 6. Soja - Fenilalanina amônia liase (PAL) - Determinação da atividade. I. Universidade Estadual de Maringá. Programa de Pós-graduação em Ciências Biológicas. II. Título. CDD 21.ed.635.655 ANDERSON RICARDO SOARES LIGNIFICAÇÃO DE RAÍZES DE SOJA SOB AÇÃO DE L-3,4-DIIDROXIFENILALANINA (L-DOPA) Profª Drª Maria de Lourdes Lucio Ferrarese Orientadora Maringá Abril – 2006 2 APRESENTAÇÃO Esta dissertação é composta de uma Revisão e de um Artigo científico que trata dos efeitos do L-DOPA na lignificação de raízes de soja. O artigo foi redigido de acordo com as normas da revista Journal of Chemical Ecology. Anderson Ricardo Soares; Maria de Lourdes Lucio Ferrarese; Rita de Cássia Siqueira; Franciele Mara Lucca Zanardo Böhm and Osvaldo Ferrarese-Filho. L-DOPA increases lignification associated to Glycine max root growth-inhibition (submetido). 3 RESUMO GERAL As plantas superiores regularmente liberam compostos orgânicos no ambiente os quais são adicionados ao solo. Alguns destes compostos têm sido citados como agentes de interações entre plantas, fenômeno conhecido como alelopatia. Estes compostos são liberados por vários mecanismos incluindo a lixiviação pela água pluvial, exsudação pelas raízes, decomposição natural de partes das plantas na superfície ou no próprio solo. A ação primária dos compostos alelopáticos ainda não foi estabelecida, mas alguns efeitos fisiológicos são conhecidos. Tipicamente os aleloquímicos interferem nas plantas superiores suprimindo a germinação, causam injúrias durante o crescimento da raiz e meristemas e, então, inibem o crescimento das plantas. Embora as plantas de cobertura do solo forneçam valor à colheita quando adicionadas em sistemas agronômicos, muitas espécies utilizadas como coberturas são tóxicas devido às altas concentrações de fitoquímicos, próprios da alelopatia. Mucuna pruriens (L.) DC. var. utilis é um exemplo de planta de cobertura do solo bem sucedida e com vários produtos naturais altamente ativos. Mucuna tem sido amplamente cultivada em áreas tropicais como enriquecedora do solo, cobertura para controle de plantas daninhas e silagem. Tem sido relatado que vários agentes químicos secundários são produzidos pelas sementes, folhas e raízes de Mucuna. O principal composto fitotóxico encontrado é o aminoácido não-protéico L-3,4-dihidroxifenilalanina (L-DOPA), que é utilizado no tratamento sintomático da doença de Parkinson. Além disso, este composto é liberado das raízes de Mucuna para o solo e inibe o crescimento de espécies de plantas vizinhas. Esta ação alelopática tem sido relatada em um grande número de espécies de plantas, mas o conhecimento sobre o mecanismo de ação do L-DOPA é escasso. Nas plantas, L-DOPA é precursor de muitos alcalóides, catecolaminas, flavonóides, melanina e fenilpropenóides. A via dos fenilpropenóides é uma das mais importantes devido ao seu papel na síntese de compostos fenólicos e de uma ampla série de produtos secundários em plantas, incluindo lignina. Fenilalanina amônia liase (PAL) e peroxidase (POD) têm sido associadas com a polimerização de monolignóis e consequentemente a lignificação. Não há relatos sobre os efeitos de L-DOPA exógeno em raízes de soja. Devido ao importante papel da lignificação no crescimento da raiz, esta pesquisa tem por objetivo investigar a absorção e os efeitos de L-DOPA nas atividades da PAL e peroxidases, nos conteúdos de compostos fenólicos e de lignina e no crescimento das raízes de soja. 4 Plântulas obtidas após três dias de germinação, foram cultivadas em solução nutriente, pH 6.0, contendo ou não L-DOPA (0.1 a 1.0 mM). Os experimentos foram acondicionados em uma câmara de crescimento (25°C, 12-h luz/12-h escuro, irradiação de 280 µmol m-2 s-1) durante 24 horas. Depois da incubação, a absorção de L-DOPA pelas raízes e os efeitos sobre as atividades enzimáticas, compostos fenólicos, conteúdos de lignina e o crescimento da raiz foram avaliados. Os dados são expressos como médias de três a seis experimentos independentes ± erro padrão da média. Para testar a significância das diferenças observadas foi realizada a análise de variância. A diferença entre os parâmetros foi avaliada pelo teste de múltiplas comparações de Dunnett e valores de P<0,05 foram considerados estatisticamente significantes. Os resultados mostraram que 1) a média total do comprimento das raízes foram 20.9 a 76,7% menores do que os controles para tratamentos com 0.25 a 1.0 mM, respectivamente. As biomassas frescas das raízes diminuíram 11,2 e 14,3% nos tratamentos com 0.5 e 1.0 mM, respectivamente. Nas mesmas concentrações, as biomassas secas diminuíram 16,1 e 14,9% em relação aos controles. 2) Os teores de L-DOPA aumentaram progressivamente nas raízes. Após tratamento com 0.1 mM, os conteúdos foram 0,099 mg g-1 de matéria seca enquanto, a 1.0 mM, este contedúdo foi 0,306 mg g-1 de matéria seca. 3) L-DOPA aumentou a atividade da PAL de 82% a 106%, após os tratamentos com 0.25 a 1.0 mM. As atividades das peroxidases solúveis aumentaram cerca de 39% enquanto as atividades das peroxidases ligadas à parede celular aumentaram em torno de 31% independentemente das concentrações de L-DOPA. 4) Os conteúdos de compostos fenólicos aumentaram significativamente de 84% (a 0.1 mM) a 145% (a 1.0 mM). 5) Os conteúdos de lignina aumentaram 56%, de 0.25 a 1.0 mM de L-DOPA. Os resultados revelam que o crescimento da raiz (comprimento e biomassa) diminuiu após a absorção de L-DOPA, enquanto as atividades da PAL e POD, os compostos fenólicos e os conteúdos de lignina aumentaram. Com exceção da soja, a redução do crescimento da raiz por LDOPA foi relatada em várias espécies de plantas. Algumas plantas são tolerantes, outras não. Em termos gerais, a redução do crescimento de raízes de plântulas tem sido associada com a lignificação precoce das paredes celulares induzida pelo aleloquímico, acompanhada por aumento nos teores de compostos fenólicos e atividades da PAL e POD. Sabe-se que as paredes celulares se tornam lignificadas quando a expansão celular cessa, quando se diferenciam em xilema ou quando a célula está sob estresse abiótico ou biótico. A POD e polifenoloxidase mediam a oxidação de L-DOPA para formar dopaquinona e, posteriormente, melanina. Durante esta via, espécies reativas ao oxigênio (EROs) são liberadas e as plântulas podem sofrer estresse. 5 Como resposta, a via de fenilpropenóides é acelerada aumentando as atividades da PAL e das peroxidases, produzindo compostos fenólicos e lignina. Este parece ser o caso relatado no presente trabalho. Concluindo, os resultados indicam a suscetibilidade da soja ao L-DOPA e reforçam o papel deste aminoácido não-protéico como forte aleloquímico. Além disso, os resultados sugerem que a inibição induzida por L-DOPA no crescimento das raízes de plântulas de soja, pode ser devido a um processo de enrijecimento da parede celular relacionado à produção de lignina. Se a parede celular é o sítio de ação do L-DOPA, estas descobertas fortalecem as informações a respeito de como este composto afeta o crescimento das plantas. 6 GENERAL ABSTRACT Since higher plants regularly release organic compounds in the environment, their decay products are often added to the soil matrix and a few have been reported as agents of plant-plant interactions, a characteristic of allelopathy. These compounds are released by various mechanisms, which include release in soil by rainwater, excretion or exudation from roots, and by natural decay of parts of plants lying above or below the ground. The primary mode of action has not been established for any allelopathic compound, albeit some physiological actions are known. Allelochemicals, active against higher plants, typically suppress seed germination, cause injury to root growth and other meristems, and inhibit seedling growth. Although cover crops provide an added crop value in the agronomic system, many cover-crop species are too toxic due to high concentrations of phytochemicals, proper to allelopathy. Mucuna pruriens (L.) DC. var. utilis is an example of a successful cover crop with several highly active natural products. Mucuna has long been cultivated in tropical areas as soilimproving crop, as cover crop to control weeds, and as forage plants. It has been reported that several secondary chemicals agents are produced by seeds, leaves and roots of Mucuna. The main phytotoxic compound encountered is held to be the non-protein amino acid L-3,4dihydroxyphenylalanine (L-DOPA), which is chiefly used in treating the symptoms of Parkinson's disease. Moreover, this compound is released from the Mucuna roots into soils and inhibits the growth of nearby plant species. Allelopathic action has been reported in many plant species but knowledge on L-DOPA’s mechanism is scanty. In plants, L-DOPA is a precursor of many alkaloids, catecholamines, flavonoids, melanin and phenylpropenoids. The phenylpropenoid pathway is one of the most important metabolic pathways since it synthesizes phenolic compounds and a wide range of secondary products in plants, including lignin. Phenylalanine ammonia-lyase (PAL) and peroxidase (POD) have been associated with monolignol polymerization and, consequently, with lignification. No reports on the effects of exogenous L-DOPA on soybean roots are available so far. Due to the important role of lignification in root growth, current research investigates the absorption of L-DOPA by soybean roots and their effects on PAL and POD activities, phenolic compounds and lignin contents. Three-day-old seedlings were cultivated in nutrient solution, pH 6.0, containing or not 0.1 to 1.0 mM L-DOPA. Experiments were carried in a growth chamber (25°C, 12-h light/12-h dark 7 cycle, irradiance of 280 µmol m-2 s-1) during 24 hours. After incubation, the absorption of LDOPA by roots and their effects on enzymatic activities, phenolic compounds and lignin contents and root growth were evaluated. Data are expressed as means of three to six independent experiments ± S.E. Whereas variance tested the significance of the observed differences, the difference between parameters was evaluated by Dunnett´s multiple comparison test and P values <0.05 were considered to be statistically significant. Results showed that 1) mean total root lengths were 20.9 to 76.7% less than controls for 0.25 to 1.0 mM treatments, respectively. Fresh root weights decreased 11.2 and 14.3% at 0.5 and 1.0 mM treatments, respectively. In similar concentrations, dry root weights decreased 16.1 and 14.9% less than those of controls. 2) L-DOPA was progressively absorbed by the roots. After exogenous 0.1 mM treatment, the L-DOPA content in the roots reached 0.099 mg g-1 dry weight while, at 1.0 mM, this content amounted to 0.306 mg g-1 dry weight. 3) L-DOPA increased PAL activities from 82% to 106% after 0.25 to 1.0 mM treatments. Soluble POD activities increased around 39%, while cell wall-bound POD activities approximately increased 31% regardless of LDOPA concentrations. 4) Phenolic compound content was significantly increased from 84% (at 0.1 mM) to 145% (at 1.0 mM). 5) Lignin content increased around 56% from 0.25 to 1.0 mM LDOPA. Results show that root growth (length and weight) decreased, while PAL and POD activities and phenolic compounds and lignin contents increased under L-DOPA action. Except in soybean, reduction of root growth by L-DOPA has been related in several plants. Some species are tolerant but others are not. In general terms, reduction of seedling roots has been associated with premature allelochemical-induced lignification of the cell walls accompanied by increase in phenolic compounds and PAL and POD activities. Cell walls are know to become lignified when cell expansion ceases, when it differentiates to particular specialization, notably the xylem, or when the cell is under abiotic or biotic stresses. It is known that POD and polyphenoloxidase (PPO) mediate the oxidation of L-DOPA to form dopaquinone and, further, melanin. During this pathway, reactive oxygen species (ROS) are released and seedlings may be stressed. As response, the phenylpropenoid pathway is accelerated, increasing PAL and POD activities, producing phenolics compounds, and consequently lignin. This seems to be the case reported in current research. Current results indicate the susceptibility of soybean to L-DOPA and reinforce the role of this non-proteic amino acid as a strong allelochemical. Results further suggest that L-DOPA- 8 induced inhibition of soybean root growth may be due to cell wall stiffening process associated with lignin production. If the cell wall is a target site of L-DOPA, findings corroborate the manner the compound affects the growth of plant species. 9 REVISÃO BIBLIOGRÁFICA Aspectos gerais O termo alelopatia foi originalmente sugerido por Hans Molish, em 1937, para definir o fenômeno da interinfluência entre plantas (incluindo microorganismos), sem a necessidade de contato físico (Tang et al., 1995). Tal interação à distância é promovida pela liberação de biossintatos para o ambiente por um produtor e sua absorção pelo receptor (Rice, 1984). O estabelecimento de relações causais entre cada um deles e os efeitos alelopáticos é complexo: um mesmo organismo produz e segrega inúmeros aleloquímicos; estes uma vez liberados interagem entre si e com outros agentes bióticos e abióticos do meio. Além disso, cada espécie vegetal desenvolveu um modo próprio de responder ao estresse alelopático. Assim, de um modo geral, infere-se que a interação alelopática depende da concentração e estabilidade dos aleloquímicos, da atividade microbiana e da resistência da planta (Blum et al., 1984). Os compostos com ação alelopática são primordialmente oriundos do metabolismo secundário. Dentre os milhares de aleloquímicos destacam-se os compostos fenólicos, os terpenos e os compostos nitrogenados entre os quais os aminoácidos não-protéicos, motivo do presente estudo. Os aleloquímicos interferem em muitos processos vitais das plantas e sua atuação não é específica. As funções prejudicadas, com maior freqüência, são a utilização de água e assimilação de nutrientes (Lyu e Blum, 1990; Bergmark et al., 1992; Booker et al., 1992), o crescimento de raízes e expansão de folhas (Blum e Dalton, 1985), a fotossíntese e a síntese de proteínas (Mersie e Singh, 1993), a respiração celular e a permeabilidade da membrana celular e atividades enzimáticas (Baziramakenga et al., 1995; Politycka, 1996, 1997, 1998). Grande parte dos estudos no campo da alelopatia está voltada para o isolamento e identificação de estruturas dos compostos que causam efeito alelopático. Há abundantes informações sobre as estruturas químicas e aspectos pertinentes de sua interação com o ambiente. Entretanto, investigações mais consistentes voltadas à absorção desses compostos (Lehman e Blum, 1999; Ferrarese et al., 2000; Silva et al., 2000, Herrig et al., 2000), bem como seus efeitos metabólicos nas plantas (Baleroni, 1999; Baleroni et al., 2000; Herrig et al. 2002; Ng et al., 2003; Doblinski et al., 2003; Santos et al., 2004; Böhm et al., 2006) apontam para a necessidade de maior atenção dos pesquisadores das áreas de fisiologia e bioquímica das plantas. 10 O maior desafio desses cientistas é determinar o mecanismo de ação dos aleloquímicos. Não existe uma simples equação que explique o modo de ação dos aleloquímicos. Outra questão em aberto, é que não tem sido possível separar, com clareza, os efeitos primários dos secundários. Diferentes componentes estruturais da célula podem sofrer os efeitos dos aleloquímicos, muito embora as perturbações a nível de membrana indicam ser este o ponto inicial para as múltiplas ações destes compostos. Os dados obtidos até o momento levam a crer que a atividade destes compostos, sobre as membranas, desencadeia subseqüentes alterações fisiológicas. É possível que a fitotoxicidade de muitos destes compostos não seja devida a um sítio localizado de interferência, mas talvez decorrente da ação sobre múltiplos sítios (Einhellig, 1995). Além disso, um dos grandes déficits de conhecimento acerca das atividades dos aleloquímicos é uma clara explicação para as diferenças de sensibilidade entre as espécies. Não há um aleloquímico genérico! Diferentes modos de ação dentre os inúmeros compostos são possíveis, e têm sido freqüentemente relatados (Macias, 1995). Como se nota, estudos sobre a ação de aleloquímicos, sua absorção pelas raízes, seus efeitos sobre o metabolismo celular constituem, de um modo geral, assunto indiscutivelmente fascinante. A variabilidade de respostas, os controversos efeitos sobre as espécies estudadas são pontos que desafiam os pesquisadores na busca do entendimento do modo de ação desses compostos. O aleloquímico L-DOPA As leguminosas vêm sendo utilizadas na agricultura desde os tempos remotos, há mais de dois mil anos. O gênero Mucuna (Leguminosae) compreende aproximadamente 160 espécies. As espécies mais comumente encontradas são deeringiana, utilis, pruriens, cochichinensis, nivea, capitata, hassjoo, diabolica, aterrima e cinérea. Estas espécies são muito utilizadas como silagem e planta de cobertura do solo devido à grande quantidade de matéria orgânica de alta digestibilidade que produz, além da sua capacidade de controlar fitonematóides (Vargas-Ayala et al., 2000). Mucuna spp (Fig. 1), de ocorrência tropical e amplamente usada como cobertura do solo, contém em suas folhas, sementes e raízes um importante agente alelopático, L-DOPA (L3,4-dihidroxifenilalanina) (Prakash et al., 2001; Nishihara et al., 2002). 11 Fig. 1. A planta e seu composto alelopático L-3,4-diidroxifenilalalnina (L-DOPA). L-DOPA, cuja formação é resultado da hidroxilação da tirosina, é um aminoácido nãoprotéico, precursor de alcalóides, fenilpropenóides, flavonóides, lignina e melanina. Além desta característica, recente estudo envolvendo sua aplicação em diferentes espécies vegetais revelou que ele possui atributos típicos de um aleloquímico (Nishihara et al., 2004). Estes autores observaram variabilidade das respostas de diferentes plantas ao L-DOPA. Por exemplo, os tratamentos de espécies de Gramíneae e Leguminosae mostraram-se menos afetados, no que tange ao crescimento das raízes, que os tratamentos de espécies de Brassicaceae, Compositae, Hydrophyllaceae e Cucurbitaceae. Estes resultados permitiram concluir que L-DOPA constitui um poderoso aleloquímico haja vista que as concentrações requeridas para ocasionar 50% de inibição (EC50) variaram de 5 a 50 µg/ml. Além da possibilidade de afetar o crescimento de várias plantas, em especial o crescimento das raízes, outro aspecto a ser destacado é a capacidade destas plantas detoxificarem L-DOPA. Estas respostas protetoras ao L-DOPA são consideradas como resultantes da sua oxidação. Durante este processo, L-DOPA pode ser descarboxilado formando dopamina, pela ação da L-DOPA descarboxilase, ou formando 3-O-metildopa pela ação da catecol-O-metiltransferase (Often et al., 2001). L-DOPA tem despertado muita atenção devido sua ação preventiva contra a doença de Parkinson, caracterizado pela deficiência na síntese do neurotransmissor dopamina, nas células nervosas. L-DOPA é descarboxilado a dopamina quando entra nas células nervosas. Vered et al (1994) relataram que o conteúdo de L-DOPA nas plântulas de Vicia faba é alto. A injeção do extrato destas plântulas em ratos aumentou o conteúdo de dopamina no cérebro e a excreção pela 12 urina de L-DOPA, dopamina e seus metabólitos de maneira similar àquela observada após a administração de L-DOPA na sua forma sintética. O estresse ambiental é fator limitante à produtividade vegetal e ao rendimento das colheitas. Muitos dos processos deletérios sofridos por plantas submetidas às condições adversas são mediados por espécies reativas de oxigênio (EROs) como peróxido de hidrogênio (H2O2), ânion superóxido (•O2) e radicais hidroxil (•OH) e perhidroxil (•O2H) geradas em diferentes compartimentos celulares como consequência do funcionamento inadequado das vias metabólicas e em processos fisiológicos normais (Allen, 1995; Apel e Hirt, 2004). Estas moléculas reativas, especialmente •OH, são altamente destrutivas para lipídios, ácidos nucléicos e proteínas podendo ocasionar a morte celular. De um modo geral tem sido aceito que EROs, ao lado de outros mediadores como o Ca2+, atuam como mensageiros secundários nas respostas aos estresses bióticos e abióticos e na sinalização hormonal. Neste cenário, H2O2, que é relativamente estável e difusível através das membranas, é reconhecido atualmente como uma molécula sinalizadora, atuando em diversas respostas fisiológicas, bioquímicas e moleculares, tanto em nível celular como da planta inteira. Assim, a adição de H2O2 ou a sua produção experimental em tecidos vegetais tem demonstrado que esta espécie química atua como sinal indutor da expressão gênica de diversas proteínas, entre elas, enzimas antioxidativas. Outro aspecto não menos importante, é que EROs como •O2 e H2O2 são também requeridas para lignificação (Buchanan et al., 2000). Existem vários processos biológicos capazes de provocar reduções eletrônicas gerando EROs. Todas estas espécies são mais reativas que o O2 e, em conseqüência, mais tóxicos (Boveris, 1984; Fridovich, 1998). EROs podem se originar nas mitocôndrias, via cadeia respiratória. A NADH desidrogenase e a ubiquinona, nas formas reduzidas, são capazes de transferir elétrons ao O2 gerando •O2 (Gardner e Boveris, 1990). Além disso, peroxissomos vegetais contém várias oxidases capazes de produzir •O2 e H2O2 (Del Rio et al., 1998). Entretanto, a fonte mais importante de EROs nas plantas é o cloroplasto. Muitos intermediários da cadeia transportadora de elétrons são encontrados na forma reduzida, o que aumenta o risco de transferência de elétrons ocasionando a formação de •O2 (Palatnik et al., 2002). Após sucessivas reações esta espécie química pode gerar significativo número de espécies reativas. Normalmente, os mecanismos protetores das células (superóxido dismutase, catalase, peroxidases, por exemplo) podem eliminar estes compostos sem maiores dificuldades. Todavia, quando estes sistemas de defesa falham, ou quando aumenta anormalmente a produção de EROs, 13 o balanço entre oxidantes e antioxidantes favorece os primeiros e se estabelece a condição conhecida como estresse oxidativo. Os complexos ferro/enxofre das metaloproteínas, em particular os centros Fe-S presentes em enzimas como a aconitase e a fumarase, são rapidamente destruídos por •O2 com inativação das enzimas (Asada, 1999). O H2O2 inibe reversivelmente várias enzimas do ciclo de fixação do carbono e de outras vias metabólicas pela oxidação de tióis funcionais, sendo também capaz de ocasionar a peroxidação de lipídios e pigmentos (Dietz, 2003). Albrecht e Kohlenbach (1990) constataram que L-DOPA é encontrado em quantidades significativas nos vacúolos de folhas de Vicia faba e o H2O2 formado nos cloroplastos se difunde para os vacúolos. Peroxidase vacuolar oxida L-DOPA a dopacromo e outros compostos como a melanina. A oxidação do L-DOPA também tem sido relatada por Takahama (1992) em protoplastos de células do mesófilo de Vicia faba e em vacúolos isolados dos protoplastos, quando tratados com H2O2. Peroxidase vacuolar, isolada de folhas de Vicia faba, oxidam DOPA lentamente (Takahama e Oniki, 1998). Bem recentemente, Hachinohe e Matsumoto (2005) estudando o metabolismo de L-DOPA em Echinochloa crus-galli e Lactuca sativa relataram que pode ocorrer formação de EROs durante a biossíntese de melanina, gerada por uma cascata de autoxidações do L-DOPA. Estas reações ocorrem sem catálise enzimática, mas a taxa é aumentada na presença de concentrações traços de íons metálicos (Fe3+ e Cu2+). A perda de um elétron de L-DOPA produz o radical semiquinona DOPA-SQ-. Este pode ser oxidado para dopaquinona (DOPA-Q) que é um intermediário da via de oxidação de L-DOPA (O’Brien, 1991; Bolton et al., 2000). Este processo pode ocorrer pela transferência de elétrons a íon metálico com propriedade redox adequada, O2 molecular ou outros aceptores de elétrons como o peróxido de hidrogênio (Beyer, 1992). DOPA-Q também pode ser gerado pela perda direta de dois elétrons de L-DOPA através de reação enzimática. Nesse caminho tem sido proposto que a oxidação do L-DOPA pode resultar em danos a outras moléculas através de reações diretas ou indiretas (O’Brien, 1991; Bolton et al., 2000). DOPA-SQ• pode transferir elétrons para outras moléculas, ou retirar átomos de hidrogênio. Danos indiretos podem ocorrer pela produção de EROs, via redução direta de peróxidos ou via redução de O2 molecular para radicais superóxidos (•O2) e subsequente dismutação destas espécies para H2O2; na presença de alguns íons metais de transição, H2O2 pode formar radicais HO•. DOPA-Q pode ser oxidado e os produtos deste processo são compostos indólicos, que podem sofrer reações posteriores formando melanina. Em adição, a taxa de oxidação de L- 14 DOPA é marcadamente dependente do pH. Em pH ácido L-DOPA é relativamente estável; em pH fisiológico a oxidação ocorre em poucas horas enquanto que em solução básica a oxidação é muito rápida (Pattison et al., 2002). Outros estudos têm mostrado que a incubação de L-DOPA, ou dos seus produtos de oxidação, com proteínas (por exemplo, creatina quinase) na presença de Fe3+ pode resultar na geração de ambos DOPA-SQ• e radicais sobre a proteína alvo. A formação de radicais pode ocorrer via múltiplos mecanismos incluindo átomos de hidrogênio retirados direto da proteína por DOPA SQ-. Como resultado há formação de •OH e subsequente ataque desta espécie sobre a proteína, podendo levar à inibição enzimática (Pattison et al., 2002). A inativação mediada por oxidação direta de grupos SH no sítio ativo da enzima por DOPA-SQ• foi proposta por Takasaki e Kawakishi (1997). Estes autores revelaram que os produtos de oxidação do L-DOPA, como dopaquinona, se ligam aos grupos SH de proteínas (cisteína) formando 5-S-cisteinildopa. Dopaquinona é um intermediário altamente reativo e na ausência de compostos SH sofre ciclização eventualmente direcionada para a formação de melanina. Pattison et al. (2002) relataram que a incubação de Fe3+ (ou Cu2+) unido a ADP com LDOPA na presença de DNA plasmídio, resulta na formação de uma forma circular relaxada como resultado da quebra da fita simples, com grandes danos resultantes do aumento das concentrações de ADP-Fe3+ e L-DOPA. Os problemas observados foram amenizados pela adição de catalase e manitol, que diminuem os níveis de H2O2 e de •OH nas células. Interações de DOPA com Mn2+, Ni2+ e Co2+ na presença de DNA foram investigadas, porém, sem a constatação de qualquer dano, o que tem sugerido que isto possa ser decorrente da ausência de radicais •OH, eventualmente produzidos. Vários estudos têm constatado que a citotoxicidade do L-DOPA, nas células animais, é atribuída às EROs geradas durante sua oxidação para melanina. Em células de neuroblastoma, Lai e Yu (1997) demonstraram correlação entre a citotoxicidade de L-DOPA e a formação de melanina. As células sofreram danos oxidativos causados por EROs que romperam a integridade da membrana plasmática resultando na redução do crescimento ou morte celular. Mena et al. (1997) relataram que a incubação de L-DOPA e neurônios pós-natais de camundongos, contendo superóxido dismutase mutada, causou morte celular. Estes autores sugerem que a enzima mutada aumenta a formação de radicais livres, sujeitando a célula ao estresse oxidativo, o qual pode eventualmente desencadear a morte celular. 15 Os danos oxidativos do L-DOPA em plantas, avaliados pela quantificação da peroxidação lipídica nas membranas celulares de Echinochloa crus-galli, Lactuca sativa e Daucus carota foram estimados por Hachinohe e Matsumoto (2005). Os níveis de peróxidos lipídicos foram maiores em comparação com experimentos controles, sugerindo o envolvimento de EROs num mecanismo similar às células animais. Os efeitos citotóxicos descritos foram completamente reduzidos por antioxidantes (ácido ascórbico e α-tocoferol) ou enzimas (catalase e superóxido dismutase). A via dos fenilpropenóides A presença da parede celular, acima de todas as outras características, distingue as células vegetais das células animais. Ela contém uma variedade de enzimas, além de desempenhar papel importante na absorção, transporte e secreção de substâncias e de atuar na defesa do vegetal contra agentes patogênicos como bactérias ou fungos (Buchanan et al., 2000). Depois da celulose, o polímero mais abundante das paredes celulares é a lignina, sendo que sua incorporação na parede celular ocasiona rigidez estrutural e resistência dos tecidos das plantas (Polle et al., 1994). Isto se dá devido ao mecanismo de espessamento que diminui a extensibilidade da parede celular em decorrência da formação de pontes difenil entre polímeros da parede, por ação da peroxidases (Sánchez et al., 1996). Muito do que se sabe a respeito da lignificação nas plantas se deve aos estudos de enzimas envolvidas neste processo, principalmente peroxidases, solúveis e ligadas à parede celular, catalase, fenilalanina amônia liase (PAL), cinamil álcool desidrogenase (CAD), além de outras (Abreu, 1994). Lignina é um polímero formado a partir de monômeros de álcool cinamílico sendo que ocorrem variações nos seus tipos, dependendo de onde esta foi sintetizada. Nas gimnospermas, a lignina é composta principalmente de unidades de álcool coniferílico, e nas angiospermas, apresenta polímeros onde há igual proporção de unidades de álcoois coniferílico e sinapílico. Existe ainda, uma pequena porção de álcool p-cumarílico em ambos os tipos de lignina. As ligações intermoleculares que formam estes polímeros também são bastante complexas, sendo que se atribui às ligações denominadas de pontes difenil a rigidez característica das paredes celulares durante sua maturação (Whethen et al., 1998). Existem várias enzimas envolvidas na síntese de lignina, sendo a sequência metabólica que conduz a esse processo conhecida como via de fenilpropenóides. A primeira reação da via é 16 catalisada pela PAL, que converte L-fenilalanina em ácido trans-cinâmico. Este, apesar de não ser ainda um composto fenólico, é precursor imediato dos mesmos (Jones, 1984). Outra etapa na via de síntese de fenilpropenóides, de grande importância no desenvolvimento de plantas é a que gera fenólicos precursores para síntese e deposição de lignina na parede celular. Os precursores são relatados por Hahlbrock e Schell (1989), como sendo monômeros de álcoois cinamílicos hidroxilados e metoxilados que são sintetizados em dois passos pela cinamil álcool-CoA redutase (CCR) e cinamil álcool desidrogenase (CAD). Esta última atua na síntese de álcoois cinamílicos, dos seus correspondentes cinamaldeídos, e é considerada, por sua alta especificidade, como marcadora do processo de lignificação. Ao final da via de fenilpropenóides, precursores monoméricos de lignina são enzimaticamente desidrogenados a radicais fenóxidos por peroxidases. Estes radicais se polimerizam espontaneamente, gerando complexas ligações rígidas entre proteínas, ligninas e polissacarídeos (Polle et al., 1994). Peroxidases têm sido implicadas nessas ligações cruzadas pelo fato de terem sido localizadas nas paredes celulares de tecidos lignificantes. A presença da peroxidase (aniônica e catiônica) ligada à parede celular pode estar relacionada às isoenzimas com altas afinidades com essa parte da célula. Deste modo, acredita-se que peroxidase ligada à parede celular, em especial a catiônica, têm grande afinidade por precursores de lignina. Os citados autores verificaram que, em hipocótilos de abeto (Picea abies L.), em processo de lignificação, peroxidase catiônica estava localizada no xilema durante o espessamento secundário. Ao mesmo tempo, enzimas envolvidas na síntese de fenilpropenóides, como PAL e cinamato 4-hidrolase, foram localizadas no citosol e vesículas de Golgi, respectivamente, pressupondo-se co-regulação durante a síntese de lignina. A lignificação por peroxidases requer H2O2 como co-substrato. Aumento na produção de H2O2 leva ao aumento da atividade da peroxidase ligada à parede celular e aumento na síntese de dímeros de ácidos fenólicos. A formação desses dímeros e a ativação da síntese de lignina na parede celular aumentam a rigidez desta ocasionando a inibição do crescimento das raízes (Ishii, 1998; Santos et al., 2004). L-DOPA e fenilpropenóides: os objetivos Como se notou a via de fenilpropenóides é de fundamental importância para o desenvolvimento das plantas, em especial nos estádios iniciais do crescimento das raízes. Por ela são produzidos compostos fenólicos com ampla variedade de funções, com destaque ao processo 17 de lignificação. Como visto anteriormente, os efeitos do L-DOPA no processo de lignificação das raízes não têm sido descritos na literatura especializada. Levando em conta as informações descritas é que o presente trabalho está sendo apresentado. As questões fundamentais que se pretendeu responder foram: 1. as enzimas da via de fenilpropenóides, especialmente aquelas relacionadas à lignificação, seriam afetadas, nas raízes de soja, por níveis crescentes de L-DOPA? e 2. as possíveis alterações enzimáticas causadas pelo L-DOPA comprometeriam o crescimento das raízes de soja? L-DOPA L-DOPA Fig. 2 Visão geral do suposto modo de ação do L-DOPA nas raízes de soja. 18 REFERÊNCIAS BIBLIOGRÁFICAS ABREU, H.S. Biossíntese da lignificação. Rio de Janeiro: EDUR, p.63.1994. ALBRECHT, C.; KOHLENBACH, H. W. L-DOPA content, peroxidase activity, and response to H2O2 of Vicia faba L. and V. narbonensis L. in situ and in vitro. Springer Verlag Wien, v. 154, p.144 -150, 1990. ALLEN, R.D. Dissection of oxidative stress tolerance using transgenic plants. Plant Physiology, v.107, p. 1049-1054, 1995 APEL, K.; HIRT, H. Reactive oxygen species: metabolism, oxidative stress and signal transduction. Annual Review of Plant Biology and Plant Molecular Biology, v.55, p. 373-399, 2004. ASADA, K. The water-water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annual Review of Plant Physiology and Plant Molecular Biology, v.50, p.601-639, 1999. BALERONI, C.R.S. Germinação de sementes e composição de ácidos graxos em cotilédones de canola (Brassica napus L.) sob influência de ácidos fenólicos. Maringá: Universidade Estadual de Maringá, 72p. Dissertação de Mestrado - Produção Vegetal, 1999. BALERONI, C. R. S.; FERRARESE, M.L.L.; BRACCINI, A.L.; SCAPIM, C.A.; FERRARESE-FILHO, O. Effects of ferulic and p-coumaric acids on canola (Brassica napus L. cv. Hyola 401) seed germination. Seed Science and Technology, v.28, p.333-340, 2000. BAZIRAMAKENGA, R.; LEROUX, G.D.; SIMARD, R.R. Effects of benzoic and cinnamic acids on membrane permeability of soybean roots. Journal of Chemical Ecology, v.21, p.12711285, 1995. 19 BERGMARK, C. L.; JACKSON, W. A.; VOLK, R. J.; BLUM, U. Differential inhibition by ferulic acid of nitrate and ammonium uptake in Zea mays L. Plant Physiology, v.98, p.639-645, 1992. BEYER, R.E. An analysis of the role of coenzyme Q in free radical generation and as an antioxidante. Biochemistry and Cell Biology, v.70, p.390-403, 1992 BLUM, U.; DALTON, B. R. Effects of ferulic acid, an allelopathic compound, on leaf expansion of cucumber seedlings grown in nutrient culture. Journal of Chemical Ecology, v.11, p.279-301, 1985. BLUM, U.; DALTON, B. R.; RAWLINGS, J. O. Effects of ferulic acid and some of its microbial metabolic products on radicle growth of cucumber. Journal of Chemical Ecology, v.10, p.1169-1191, 1984. BÖHM, P.A.F.; ZANARDO, F.M.L.; FERRARESE, M.L.L.; FERRARESE-FILHO, O. Peroxidase activity and lignification in soybean root growth-inhibition. Biologia Plantarum, v.50, p.315-317, 2006. BOLTON, J.L.; TRUSH, M.A.; PENNING, T.M.; DRYHURST, G.; MONKS, T.J. Role of quinones in toxicology. Chemical Research and Technology, v.13, p.135-160, 2000. BOOKER, F. L.; BLUM, U.; FISCUS, E.L. Short-term effects of ferulic acid on ion uptake and water relations in cucumber seedlings. Journal of Experimental Botany, v.43, p.649-655, 1992. BOVERIS, A. Determination of the production of superoxide radicals and hydrogen peroxide in mitochondria. Methods Enzymology, v.105, p.429-435, 1984. BUCHANAN, B.B.; GRUISSEM, W.; RUSSELL, J.L. Biochemistry and Molecular Biology of Plants, p.1158-1203, 2000. 20 DEL RIO, L.A.; PASTORI, G.M.; PALMA, J.M.; SANDALIO, L.M.; SEVILLA, F.; CORPAS, F.J.; JIMÉNEZ, A.; LÓPEZ-HUERTAS, E.; HERNÁNDEZ, J.A. The activated oxygen role of peroxisomes in senescente. Plant Physiology, v.116, p.1195-1200, 1998. DIETZ, K.J. Plant peroxiredoxins. Annual Review of Plant Biology and Plant Molecular Biology, v.54, p.93-107, 2003. DOBLINSKI, P. M. F; FERRARESE, M. L. L.; HUBER, D. A.; SCAPIM, C. A.; FERRARESE FILHO, O. Peroxidase and lipid peroxidation of soybean roots in response to p-coumaric and phydroxybenzoic acids. Brazilian Archives of Biology and Technology, v.46, p.193-198, 2003. EINHELLIG, F.A. Allelopathy. Current status and future goals. In: Inderjit, Dakshini K.M.M., Einhellig F.A. (eds) Allelopathy. Organisms, Processes and Applications, New York: American Chemical Societies, Series 582. pp 1-24, 1995. FERRARESE, M.L.L.; FERRARESE-FILHO, O.; RODRIGUES, J. D. Ferulic acid uptake by soybean root in nutrient culture. Acta Physiologiae Plantarum, v.22, p.121-124, 2000. FRIDOVICH, I. Oxygen toxicity: a radical explanation. Journal of Experimental Biology, v.201, p.1203-1209, 1998. GARDNER, J.F.; BOVERIS, A. Generation of superoxide anion by the NADH dehydrogenase of bovine heart mitochondria. Biochemical Journal, v.191: p.421-427, 1990 HACHINOHE, M.; MATSUMOTO, H. Involvement of reactive oxygen species generated from melanin synthesis pathway in phytotoxicty of L-DOPA. Journal of Chemical Ecology, v.31, p.237-246, 2005. HAHLBROCK, K.; SCHEEL, D. Physiology and molecular biology of phenylpropanoid metabolism. Annual Review of Plant Physiology and Plant Molecular Biology, v.40, p.347-369, 1989. 21 HERRIG, V.; SILVA. H.C; FERRARESE, M.L.; FERRARESE-FILHO, O. Ferulic acid depletion by cultured soybean seedlings under action of glucose and methionine. Brazilian Archives of Biology and Technology, v.43, p.515-518, 2000. HERRIG, V.; FERRARESE, M.L.L., SUZUKI, L.S., RODRIGUES, J.D., FERRARESEFILHO, O. Peroxidase and phenylalanine ammonia-lyase activities, phenolic acid contents, and allelochemicals-inhibited root growth of soybean. Biological Research, v.35, p.51-58, 2002 ISHII, T. Structure and function of feruloylated polysaccharides. Plant Science, v.127, p.111127, 1998. JONES, D.H. Phenylalanine ammonia-lyase: regulation of its induction and its role in plant development. Phytochemistry, v.23, p.1349-1359, 1984. LAI, C. T.; YU, P. H. Dopamine and L-ß-Dihydroxyphenylalanine hydrochloride (L-DOPA)induced cytotoxicity towards catecholaminergic neuroblastoma SH-SY5Y cells. Biochemical Pharmacology, v.53. p.353-372, 1997. LEHMAN, M.E.; BLUM, U. Evaluation of ferulic acid uptake as a measurement of allelochemical dose: effective concentration. Journal of Chemical Ecology, v.25, p.2585-2600, 1999. LYU, S-W; BLUM, U. Effects of ferulic acid, an allelopathic compound, on net P, K, and water uptake by cucumber seedlings in a split-root system. Journal of Chemical Ecology, v.16, p.24292439, 1990. MACIAS, F. Allelopathy in the search for natural herbicides model. Allelopathy. Current status and future goals. In: Allelopathy. Organisms, Processes and Applications (Inderjit, Kakshini, K. M. M., and Einhellig, F. A. eds.), New York: American Chemical Societies. Series 582, pp. 311329, 1995. 22 MENA, M.A.; KHAN, U.; TOGASAKI, D.M.; SULZER, D.; EPSTEIN, C.J.; PRZEDBORSKI, S. Effects of wild-type and mutated copper/zinc superoxide dismutase on neuronal survival and L-DOPA-induced toxicity in postnatal midbrain culture. Journal of Neurochemistry, v.69, p.2133, 1997. MERSIE, W.; SINGH, M. Phenolic acids affect photosynthesis and protein synthesis by isolated leaf cells of velvet-leaf. Journal of Chemical Ecology, v.19, p.1293-1301, 1993. NG, P.L.L.; FERRARESE, M.L.L.; HUBNER, D.A.; RAVAGNANI, A.L.S.; FERRARESEFILHO, O. Canola (Brassica napus L.) seed germination influenced by cinnamic and benzoic acids and derivatives: effects on peroxidase. Seed Science and Technology, v.31, p.39-46, 2003. NISHIHARA, E.; ARAYA, H.; HIRADATE, S.; FUJII, Y. The inhibition of lettuce growth by diffused L-3,4,-dihydroxyphelalanine (L-DOPA) in Mucuna accessions. Third World Congress in Allelopathy, p.246. 2002. NISHIHARA, E.; PARVEZ, M.M.; ARAYA, H.; FUJII, Y. Germination growth response of different species to the allelochemical L-3,4-dihydroxyphelalanine (L-DOPA). Plant Growth Regulation, v.42, p.181-189. 2004. O’BRIEN, P.J. Molecular mechanisms of quinone cytotoxicity. Chemical and Biological Interactions, v.80. p.1-41, 1991. OFTEN, D.; PANES, H.; GALILI-MOSBERG, R.; MELAMED, E. Catechol-O- methyltransferase decreases levodopa toxicity in vitro. Clinical Neuropharmacology, v.24, p.2730, 2001. PALATNIK, J.F.; VALLE, E.M.; CARRILLO, N. Oxidative stress and damage in chloroplasts from dawn to dusk. Advances in Plant Physiology, v.4, p.75-88, 2002. PATTISON, D.I.; DEAN, R.T.; DAVIES, M.J. Oxidation of DNA, proteins and lipids by DOPA, protein-bound DOPA, and related catechol(amine)s. Toxicology. v.177, p.23-37, 2002. 23 POLITYCKA, B. Peroxydase activity and peroxidation lipidic in roots of cucumber seedlings influenced by derivatives of cinnamic and benzoic acids. Acta Physiologiae Plantarum, v.18, p.365-370, 1996. POLITYCKA, B. Free and glucosylated phenolics, phenol-β-glucosyltransferase activity and membrane permeability in cucumber roots as affected by derivatives of cinnamic and benzoic acids. Acta Physiologiae Plantarum, v.19, p.311-317, 1997. POLITYCKA, B. Phenolics and the activities of phenylalanine ammonia-lyase, phenol-βglucosyltransferase and β-glucosidase in cucumber roots as affected by phenolic allelochemicals. Acta Physiologiae Plantarum, v.20, p.405-410, 1998. POLLE, A.; OTTER, T.; SEIFERT, F. Apoplastic peroxidases and lignification in needles of Norway spruce (Picea abies L.). Plant Physiology, v.106, p.53-60, 1994. PRAKASH, D.; NIRANJAN, A.; TEWARI, S.K. Some nutritional properties of the seeds of three Mucuna species. International Journal of Food Science and Nutrition, v.52, p.79-82, 2001. RICE, E. L. Allelopathy. Academic Press, Orlando, 1984. SÁNCHEZ, M.; PENA, M.J.; REVILLA, G.; ZARRA, I. Changes in dehydrodiferulic acids and peroxidase activity against ferulic acid associated with cell walls during growth of Pinus pinaster hypocotyl. Plant Physiology, v.111, p.941-946. 1996. SANTOS, W.D.; FERRARESE, M.L.L.; FINGER, A.; TEIXEIRA, A.C.N.; FERRARESEFILHO, O. Lignification and related enzymes in soybean root growth-inhibition by ferulic acid. Journal of Chemical Ecology, v.30, p.1199-1208, 2004 SILVA, H. C.; HERRIG, V.; FERRARESE, M.L.L.; FERRARESE-FILHO, O. Consumption of ferulic acid and p-hydroxybenzoic acids by soybean root tips. Brazilian Archives of Biology and Technology, v.43, p.281-284, 2000. 24 TANG, C.S.; CAI, W.F.; KOHL, K.; NISHIMOTO, R.K. Plant Stress and Allelopathy. ACS symposium Series 558; American Chemical Society. Washington, DC, pp. 142-157, 1995. TAKASAKI, S.; KAWAKISHI, S. Formation of protein-bound 3,4 dihydroxyphenilalanine and 5-S-Cysteinyl-3,4-dihydroxyphenylalanine as new cross-linkers in gluten. Journal of Agricultural and Food Chemistry, v.45, p.3472-3475, 1997. TAKAHAMA, U. Hydrogen peroxide scavenging systems in vacuoles of mesophyll cells of Vicia faba. Phytochemistry, v.31, p.1127-1133, 1992. TAKAHAMA, U.; ONIKI T. 3,4-Dihydroxyphenylalanine is oxidized by phenoxyl radicals of hydroxycinnamic acid esters in leaves of Vicia faba. Journal of Plant Research, v.111. p.487494, 1998. VARGAS-AYALA, R.; RODRIGUES-KABANA, R.; MORGAN-JONES, G.; McINROY, J.A.; KLOEPPER, J.W. Shifts in soil microflora induced by velvetbean (Mucuna deeringiana) in cropping systems to control root-knot nematodes. Biological Control, v.17, p.11-22, 2000. VERED, Y.; RABEY, JM.; PALEVEITCH, D.; GROSSKOPF, I.; HARSAT, A.; YANOWSKI, A. Bioavailability of levodopa after consumption of Vicia faba seedlings by Parkinsonian patients and control subjects. Clinical Neuropharmacology, v.17. p.46–138, 1994. WHETHEN, R.W.; MacCAY, J.J.; SEDEROFF, R.R. Recent advances in understanding lignin biosynthesis. Plant Molecular Biology, v.49, p.585-609, 1998. 25 L-DOPA INCREASES LIGNIFICATION ASSOCIATED TO Glycine max ROOT GROWTH-INHIBITION ANDERSON RICARDO SOARES; MARIA DE LOURDES LUCIO FERRARESE; RITA DE CÁSSIA SIQUEIRA; FRANCIELE MARA LUCCA ZANARDO BÖHM AND OSVALDO FERRARESE-FILHO Correspondence to: Maria de Lourdes Lucio Ferrarese Laboratory of Plant Biochemistry Department of Biochemistry State University of Maringá Av. Colombo, 5790 87020-900, Maringá, PR BRAZIL E-mail: [email protected] Fax: +55 44 2633655 26 L-DOPA INCREASES LIGNIFICATION ASSOCIATED TO Glycine max ROOT GROWTH-INHIBITION ANDERSON RICARDO SOARES; MARIA DE LOURDES LUCIO FERRARESE∗; RITA DE CÁSSIA SIQUEIRA; FRANCIELE MARA LUCCA ZANARDO BÖHM AND OSVALDO FERRARESE-FILHO Laboratory of Plant Biochemistry, Department of Biochemistry, State University of Maringá, Av. Colombo, 5790, 87020-900, Maringá, PR, Brazil Abstract – L-DOPA (L-3,4-dihydroxyphenylalanine), an allelochemical exuded from the roots of velvet bean (Mucuna pruriens (L.) DC. var. utilis) displays a high inhibitory action to plant growth. L-DOPA´s effects on phenylalanine ammonia-lyase (PAL, EC 4.3.1.5) and peroxidase (POD, EC 1.11.1.7) activities, phenolic compounds and lignin contents in soybean (Glycine max (L.) Merr.) roots were investigated to determine its possible phytotoxic mechanism. Three-dayold seedlings were cultivated in half-strength Hoagland nutrient solution (pH 6.0), without or with 0.1 to 1.0 mM L-DOPA, in a growth chamber (25°C, 12L:12D photoperiod, irradiance of 280 µmol m-2 s-1) for 24 hr. In general, length, fresh weight and dry weight of roots decreased, while PAL and POD activities, phenolic compounds and lignin contents increased after L-DOPA treatments. Data indicate the susceptibility of soybean to L-DOPA and reinforce the role of this non-proteic amino acid as a strong allelochemical. Results also suggest that L-DOPA-induced inhibition in soybean roots may be due to a cell wall stiffening process related to the formation of cross-linking between cell wall polymers linked to lignin production. Key Words – Allelopathy, L-DOPA, lignin, peroxidase, phenolic compounds, phenylalanine ammonia-lyase, root growth, soybean. ∗ To whom correspondence should be addressed. E-mail: [email protected] 27 INTRODUCTION For years it has been known that plants release organic compounds into the environment from their aerial or sub-aerial parts as exudates, volatiles, and/or decomposition residues. These compounds may accumulate in the soil environment and affect the growth and development of neighboring plants, an interaction called allelopathy (Einhellig, 1995). Velvet bean (Mucuna pruriens (L.) DC. var. utilis) is one of these plant species which has been long cultivated tropical regions as soil-improving crop, as cover crop to control weeds, and as green manure and forage plant (Anaya et al., 1999; Tarawali et al., 1999; Vargas-Ayala et al., 2000). Using in vitro bioassays and field studies, Fujii et al. (1991) identified secondary chemical agents produced and released by leaves and roots of Mucuna. The main phytotoxic compound encountered was the non-protein amino acid L-3,4-dihydroxyphenylalanine (L-DOPA), which represented about 1% of the fresh tissues. L-DOPA exudes from the Mucuna roots to soils (Furubayashi et al., 2005) and its concentrations may reach 1 ppm in water culture solution and 50 ppm in the vicinity of roots (Anaya et al., 1999). This concentration is enough to affect the growth of neighboring plants. It reduces seed germination and suppresses root growth more than shoots of different plant species (Fujii et al., 1991; Fujii, 1999; Nakajima et al., 1999; Nishihara et al., 2004; Hachinohe et al., 2004; Nishihara et al., 2005). L-DOPA´s physiological mechanism in plants, however, is poorly understood. In animals L-DOPA is a well-known precursor of the neurotransmitter dopamine and the most effective therapeutic agent for the symptomatic relief of Parkinson’s disease (Mercuri and Bernardi, 2005). In plants L-DOPA is a precursor of many alkaloids, catecholamines, flavonoids, melanin and phenylpropenoids (Hahlbrock and Scheel, 1989). The phenylpropenoid pathway is one of the most important metabolic pathways since it synthesizes phenolic compounds and a wide range of secondary products in plants, including lignin. Phenylalanine ammonia-lyase (PAL) is regarded as the primary enzyme of the phenylpropenoid biosynthetic pathway, wheareas peroxidase (POD) within the cell wall, in either the free or bound state, has been shown to be associated with monolignol polymerization and, therefore, in lignin synthesis (Passardi et al., 2005). To date, no reports on the effects of exogenous L-DOPA on soybean roots are available. Due to the important role of lignification in root growth, the question in current research was whether L-DOPA affects PAL and POD activities, phenolic compounds and lignin contents of soybean roots. 28 METHODS AND MATERIALS General Procedures. Soybean (Glycine max (L.) Merr. cv. BRS-133) seeds, surfacesterilized with 2% sodium hypochlorite for 5 min and rinsed extensively with deionized water, were dark-germinated (at 25°C) on three sheets of moistened filter paper. Twenty-five 3-day-old seedlings of uniform size were supported on an adjustable acrylic plate and transferred into a glass container (10 × 16 cm) filled with 200 ml of half-strength Hoagland’s solution (pH 6.0), without or with 0.1 to 1.0 mM L-DOPA. The container was kept in a growth chamber (25°C, 12L:12D photoperiod, irradiance of 280 µmol m-2 s-1). Roots were measured at the start and at the end of experiments (24 hr). Fresh root weight was determined immediately after incubation and dry weight estimated after oven-drying at 80°C, for 24 hr. L-DOPA was purchased from Sigma Chemical Co. (St Louis, USA) and all other reagents used were of the purest grade available or chromatographic grade. Enzymatic Assays. After incubation, all treated or untreated seedling roots were detached and enzymes were extracted. Phenylalanine ammonia-lyase (PAL) was extracted, as described by Ferrarese at al. (2000). Fresh roots (2 g) were ground at 4ºC in 0.1 M sodium borate buffer (pH 8.8). Homogenates were centrifuged (2,200g, 15 min) and the supernatant was used as the enzyme preparation. The reaction mixture (100 µmoles sodium borate buffer, pH 8.7, and a suitable amount of enzyme extract in a final volume of 1.5 ml) was incubated at 40°C, for 5 min, for PAL activity assay. Fifteen µmoles of L-phenylalanine were added to start the reaction which was stopped after 1 hr of incubation by the addition of 50 µl 5 N HCl. Samples were filtered through a 0.45 µm disposable syringe filter and analyzed (20 µl) with a Shimadzu® Liquid Chromatograph (Tokyo, Japan) equipped with a LC-10AD pump, a Rheodine® injector, a SPD10A UV detector, a CBM-101 Communications Bus Module, and a Class-CR10 workstation system. A reversed-phase Shimpack® GLC-ODS (M) column (150 × 4.6 mm, 5 µm) was used at room temperature, with an equivalent pre-column (10 × 4.6 mm). The mobile phase was methanol:water (70%:30%) with a flow rate of 0.5 ml min-1. Absorption was measured at 275 nm. Data collection and integration were performed with Class-CR10 software (Shimadzu®, Tokyo, Japan). t-Cinnamate, the product of PAL, was identified by comparing its retention time with that of standard’s. Parallel controls without L-phenylalanine or with t-cinnamate (added as internal standard in the reaction mixture) were made as described elsewhere (Ferrarese et al., 2000). PAL activity was expressed as µmol t-cinnamate hr-1 g-1 of fresh weight. 29 Peroxidase (POD) was extracted from fresh roots (0.5 g) with 67 mM phosphate buffer (5 ml, pH 7.0). Extract was centrifuged (2,200g, 5 min, 4°C), and the supernatant was used to determine the activity of soluble POD. For cell wall-bound POD isolation, the pellet was washed with deionized water until no soluble POD activity was detected in the supernatant. Pellet was then incubated in 1 M NaCl (2 ml, 1 hr, 4°C), and the homogenate was centrifuged (2,200g, 5 min). The supernatant contained the cell wall-(ionically)-bound POD. Guaiacol-dependent activities of the soluble and cell wall-bound POD were determined according to Cakmak and Horst (1991), with slight modifications. The reaction mixture (3 ml) contained 25 mM sodium phosphate buffer, pH 6.8, 2.58 mM guaiacol and 10 mM H2O2. Reaction started by adding the enzyme extract in phosphate buffer. Guaiacol oxidation was followed for 5 min, at 470 nm, and enzyme activity was calculated from the extinction coefficient (25.5 mM-1 cm-1) for tetraguaiacol. Blank consisted of a reaction mixture without enzyme extract whose absorbance was subtracted from the mixture with enzyme extract. POD activities were expressed as µmol tetraguaiacol min-1 g-1 fresh weight. Extraction and quantification of L-DOPA in roots. Exogenous L-DOPA-treated roots (0.1 g dry weight) were put into 5 ml of deionized water in 30 ml polypropylene tubes. Extraction was performed by immersing the tubes into a sonication bath for 5 min (St-Laurent et al., 2002). Extracts were then centrifuged at 2,200g for 2 min and diluted (1:1) in the chromatographic mobile phase containing water:methanol:phosphoric acid (97.5:2:0.1). The solution was filtered through 0.45 µm disposable syringe filter and analyzed (20 µl) with a Shimadzu® Liquid Chromatograph (Tokyo, Japan), as described earlier. A reversed-phase Shimpack® GLC-ODS (M) column (150 × 4.6 mm, 5 µm) was used, at room temperature, with an equivalent pre- column (10 × 4.6 mm). The mobile phase was applied with a flow rate of 0.5 ml min-1. Absorption was measured at 280 nm. Data collection and integration were performed with ClassCR10 software (Shimadzu®, Tokyo, Japan). L-DOPA was identified by comparing its retention time with that of standard’s. L-DOPA content was expressed as mg g-1 of dry weight. Phenolic Compounds Quantification. Dry root (0.25 g) was boiled for 30 min in 5 ml of 2 N HCl. After cooling, the homogenate was filtered through a Whatman® paper and the filtrate used to determine total phenolic compounds. Samples (5 ml) were mixed with 0.75 ml of 1.9 M Na2CO3 plus 0.25 ml of Folin-Ciocalteau phenol reagent. This mixture was kept in the dark, at room temperature (23 ºC - 25 ºC), for 1 hr, before its absorption was read at 750 nm (Blum et al., 1991). Ferulic acid was used as standard. Since L-DOPA absorbed by the roots might to 30 overestimate results, data obtained by HPLC (described earlier) were subtracted from total phenolics. Results were expressed as mg total phenolics g-1 dry weight. Lignin Quantification. After the incubation period, dry roots (0.3 g) were homogenized in 50 mM potassium phosphate buffer (7 ml, pH 7.0) with mortar and pestle, and transferred into a centrifuge tube (Ferrarese et al., 2002). The pellet was centrifuged (1,400g, 4 min) and washed by successive stirring and centrifugation, as follows: twice with phosphate buffer pH 7.0 (7 ml); × 3 with 1% (v/v) Triton® X-100 in pH 7.0 buffer (7 ml); × 2 with 1 M NaCl in pH 7.0 buffer (7 ml); × 2 with distilled water (7 ml); and × 2 with acetone (5 ml). Pellet was dried in an oven (60°C, 24 hr) and cooled down in a vacuum desiccator. The dry matter obtained was defined as protein-free cell wall fraction. Further, all dry protein-free tissue was placed into a screw-cap centrifuge tube containing the reaction mixture (1.2 ml of thioglycolic acid plus 6 ml of 2 M HCl) and heated (95ºC, 4 hr). After cooling at room temperature, the sample was centrifuged (1,400g, 5 min) and the supernatant decanted. The pellet was washed × 3 with distilled water (7 ml) and the product extracted by shaking (30ºC, 18 hr, 115 oscillations min-1) in 0.5 M NaOH (6 ml). After centrifugation (1,400g, 5 min), the supernatant was stored and mixed with supernatant obtained from a second pellet washed with 0.5 M NaOH (3 ml). The combined alkali extracts were acidified with concentrated HCl (1.8 ml). The lignothioglycolic acid (LTGA), formed after 4 hr at 0ºC was recovered by centrifugation (1,400g, 5 min) and washed × 2 with distilled water (7 ml). The pellet was dried at 60°C, dissolved in 0.5 M NaOH, and diluted to yield an appropriate absorbance for spectrophotometric determination at 280 nm. Lignin was expressed as mg LTGA g-1 dry weight. Statistical Design. The experimental design was completely randomized and each plot was represented by one glass container with twenty-five seedlings. Data are expressed as mean of three to six independent experiments ± S.E. Significance of differences was undertaken by oneway variance analysis with GraphPad Prism package (Version 2.0, GraphPad Software Inc., USA, 1995). Difference between parameters was evaluated by Dunnett´s multiple comparison test and P values <0.05 were considered statistically significant. 31 RESULTS To detect absorption of L-DOPA, soybean roots were incubated with this compound (0.1 to 1.0 mM) for 24 hr. Figure 1 shows that L-DOPA has been progressively absorbed by the roots. After exogenous 0.1 mM treatment, L-DOPA content determined in the roots reached 0.099 mg g-1 dry weight while, at 1.0 mM, this content amounted to 0.306 mg g-1 dry weight. Root lengths and root fresh and dry weights decreased with increasing concentrations in soybean seedlings grown during short-term exposure (24 hr) in nutrient solution containing LDOPA (Table 1). Mean total root lengths were 20.9, 72.5 and 76.7% less than controls for 0.25, 0.5 and 1.0 mM treatments, respectively. Low concentration (0.1 mM) did not impair root length. Root fresh weight decreased 11.2 and 14.3% at 0.5 and 1.0 mM treatments, respectively, when compared to control. A similar behavior was also evident in root dry weights determined as 16.1 and 14.9% less than those of controls with 0.5 and 1.0 mM L-DOPA. No appreciable changes in fresh and dry weights of roots exposed to L-DOPA at low concentrations (≤ 0.25 mM) were recorded. L-DOPA-affected PAL activities were significantly different from controls (Figure 2). The allelochemical increased the enzymatic activities from 82% to 106% after 0.25 to 1.0 mM treatments. No significant change was observed in PAL activity after 0.1 mM L-DOPA exposure. Soluble POD activities (Figure 3A) increased around 39% and cell wall-bound POD activities (Figure 3B) increased around 31% regardless of L-DOPA concentrations, when compared to control conditions. Phenolic compound content was significantly increased from 86% (at 0.1 mM) to 150% (at 1.0 mM) when compared to untreated roots (Figure 4). Finally, data revealed that the lignin content increased around 56% from 0.25 to 1.0 mM L-DOPA, while no change was recorded at 0.1 mM treatment when compared to untreated roots (Figure 5). DISCUSSION The main fact revealed in current research is that root growth (length and weight) decreased (Table 1) after L-DOPA absorption (Fig. 1), while PAL and POD activities and phenolic compounds and lignin contents increased (Fig. 2 to 5). 32 Reduction of root growth by L-DOPA has been reported in several plant species. Nishihara et al. (2004) used a treatment of 250 µg/ml (about 1.27 mM) and showed that L-DOPA strongly inhibited (<25% of control) root growth of cabbage (Brassica oleraceae), komatsuna (Brassica campestris), qing gin cai (Brassica chinensis), baby blue eyes (Nemophila menziesii), watercress (Nasturtium officinale), pumpkin (Cucurbita moschata), lettuce (Lactuca sativa) and cucumber (Cucumis sativus). However, tall fescue (Festuca arundinacea), perennial ryegrass (Lolium perenne), creeping bentgrass (Agrostis palustris), red clover (Trifolium pratense), milk vetch (Astragalus sinicus), hairy vetch (Vicia villosa) and corn (Zea mays) were less inhibited (>75% of control) and suggested a possible resistance to L-DOPA. The effect of L-DOPA (0.01 or 0.1 mM) on root growth of 32 species was examined at the germination stage (Hachinohe et al., 2004). Root growth was <50% at the control value for 7 or 19 of the species treated with 0.01 or 0.1 mM L-DOPA, respectively. Barnyard grass (Echinochloa crus-galli) was more tolerant than lettuce, one of the most susceptible (about 80% of inhibition by 0.1 mM L-DOPA). Since LDOPA reduced the soybean root growth and fresh and dry weights (Table 1), its susceptibility was indicated and the amino acid’s role as a strong allelochemical was reinforced. Investigations about absorption of L-DOPA in plant species are scanty. Using 14C-L-DOPA, Hachinohe et al. (2004) reported that this compound was largely absorbed and metabolized by roots of barnyard grass and lettuce. The authors verified that metabolites (dopamine, phenylalanine and tyrosine) derived from 14 C-L-DOPA are not directly involved in L-DOPA phytotoxicity. Since results reported in the present work indicate that L-DOPA has been effectively absorbed by soybean roots (Fig. 1), reduction of root growth by L-DOPA, verified after 24 hr treatments (Table 1), may be, at least partly, due to this active form in the roots. The discovery that L-DOPA stimulated the lignin production (Fig. 5) is of particular interest. Reduction of seedling roots has been associated with premature allelochemical-induced lignification of the cell walls accompanied by increases in phenolic compounds and PAL and POD activities (Devi and Prasad, 1996; Politycka, 1999: Herrig et al., 2002; Santos et al., 2004). PAL is regarded as the entry enzyme into the phenylpropenoid pathway responsible for the synthesis of a diverse array of phenolic metabolites, whereas structural lignin has been reported to be a response of plants against allelopathic stress. PAL participates in maize (Devi and Prasad, 1996), cucumber (Politycka, 1999) and soybean (Herrig et al., 2002; Santos et al., 2004) root growth reductions, under stress of cinnamic and benzoic acids derivatives, in association with cell wall stiffening related to the formation of cross-linking among cell wall polymers and lignin 33 production. Likewise, higher activities of PAL (Fig. 2) in soybean root tissues were accompanied by increase in phenolic compounds contents (Fig. 4) after L-DOPA treatments. Phenolic compounds play a vital role in plant growth, regulation and are mainly produced to protect plants from abiotic and biotic stresses. They are known for their anti-oxidative properties scavenging free radicals (Appel, 1993). Phenolic compounds are derived originally by enzymatic deamination of phenylalanine and tyrosine via nitrogen-free skeletons of cinnamate or pcoumarate. Once formed, the latter compound is converted, whitin the phenylpropanoid metabolism, into a broad range of secondary metabolites, such as caffeic, ferulic and sinapic acids and, further, lignin (Boerjan et al., 2003). PAL and POD are involved in this pathway and the production of phenolic compounds has been often observed when levels of phenylpropenoid pathway intermediates or end products are artificially increased. Stress caused by addition of exogenous allelochemicals (ferulic and vanillic acids) in soybean stimulated the accumulation of phenolic compounds and lignin (Herrig et al., 2002; Santos et al., 2004). As mentioned earlier, L-DOPA is a precursor of phenylpropenoids. It has been shown that L-DOPA-treated roots of cucumber accumulated phenylalanine and tyrosine supposedly due to its metabolic detoxification (Nakajima et al., 1999). Dopamine (product of enzymatic decarboxylation of L-DOPA), tyrosine (substrate of tyrosine decarboxylase to form dopamine) and phenylalanine (substrate of PAL in the first reaction of phenylpropenoid pathway) failed to increase in lettuce roots after exogenous L-DOPA treatments (Hachinohe et al., 2004). In the authors’ opinion, these metabolites are not directly involved in the phytotoxicity of L-DOPA since they may be further metabolized into other compounds. Like lettuce, soybean is susceptible to L-DOPA (Table 1) and an accumulation of phenylalanine and tyrosine may not occur because they are precursors of phenylpropenoids and lignin. It seems that these facts may explain results obtained for PAL (Fig. 2) and phenolic compounds (Fig. 4) reported in this study. Increased activities of soluble POD induced by L-DOPA treatment (Fig. 2A) are consistent with the role of this enzyme in plants. It has been frequently emphasized that one of the most accepted functions for soluble POD is to catalyze the breakdown of H2O2, with the concomitant dependent oxidation of a wide variety of substrates (Bolwell and Wojtaszek, 1997; Passardi et al., 2005). Therefore, it is often regarded as an antioxidant enzyme, protecting cells from the destructive influence of reactive oxygen species (ROS). Under normal circumstances, plants scavenge ROS by invoking this enzymatic defensive system. However, if the capacity of the cells to scavenger ROS is exceeded, the phenolic substrates oxidation by soluble POD produces 34 quinones which cause oxidative stress (Appel, 1993). The final consequence may be the depolarization of the root cell membrane enhancing its permeability with electrolyte leakage, blocking the plant nutrient uptake and hindering the root growth (Einhellig, 1995). Another relevant fact is that POD mediates the oxidation of L-DOPA to form dopaquinone and, further, melanin (Albrecht and Kohlenbach, 1990; Takahama and Egashira, 1991). Melanin biosynthesis is linked to production of ROS (1O2, O2, HO● and H2O2) or other L-DOPA oxidation products, such as semiquinone radicals, quinones and metal ion-L-DOPA complexes (Kruk et al., 1999; Pattison et al., 2002). The toxicity of L-DOPA in animals has been attributed to its auto-oxidative breakdown in the presence of molecular oxygen. Highly unstable electrophilic L-DOPA-quinones are formed which may induce oxidative damage (Bindoli et al., 1992). Hachinohe and Matsumoto (2005) have recently reported that L-DOPA increased the levels of melanin and thiobarbituric acid reactive substances (TBARS) in lettuce, which suggests that this action was due to ROS released during melanin synthesis. Although low temperature at flowering leads to the formation of a melanin-like compound in soybean seed coats (Takahashi and Akiyama, 1993), it is not possible to state with surety that melanin is synthesized in soybean roots. Since soybean radicle becomes light brown or black after 0.1 and 1.0 mM L-DOPA treatments, respectively, it is plausible that increase in soluble POD activities (Fig. 3A) may be due to oxidation of L-DOPA and, presumably, to melanin formation. L-DOPA also increased the cell wall-bound POD activities (Fig. 3B). There are longstanding evidences that this enzymatic form causes oxidative polymerization of monolignols from phenolic compounds, such as ferulic, p-coumaric and caffeic acids. This process appears to need oxidative coupling, dependent on H2O2, which causes a rapid cross-linking of cell wall polymers. H2O2 may act as a signaling molecule triggering secondary defenses which may cause an untimely cell wall lignification, thereby, decreasing cellular viability and resulting in growth reduction (Passardi et al., 2005). If cell wall-bound POD regulates cell wall stiffening and lignin polymerization, there must be a sufficient supply of H2O2. As mentioned earlier, this relatively stable form of ROS may be formed during oxidative transformation of L-DOPA into melanin (Kruk et al., 1999). Other emerging evidences suggest that H2O2 may be also produced by pH dependent cell wall POD and by NADPH oxidase complex (Bolwell and Wojtaszek, 1997) or its levels may be regulated by the POD´s peroxidative cycle (Passardi et al., 2005). It has become clear from this study that L-DOPA treatment of soybean seedlings resulted in an increase of PAL activity producing phenolic compounds. Further, H2O2-dependent POD 35 reactions convert phenolic substances into phenoxyl radicals which may produce lignin monomers. These monomers form a complex network that solidifies the plant cell and subsequently restrict the root growth. According to the results of current research, it is probable that L-DOPA-induced inhibition in soybean roots may be due to a cell wall stiffening process related to the formation of cross-linking among cell wall polymers and lignin production. If this is the L-DOPA´s working mode, the cell wall may be a site for allelopathic action and indicate how the compound affects the growth of plant species. One may ask whether these effects have been exclusively due to L-DOPA itself or to products of its metabolism. Notwithstanding the fact that L-DOPA has been absorbed by roots (Fig. 1), its metabolism has not explored in soybean seedlings. However, Hachinohe and Matsumoto (2005) evaluated the L-DOPA metabolism in other plant species and suggested that its phytoxicity is due to metabolism and not to its absorption. Therefore, the possibility cannot be discarded that data reported here are results of LDOPA-induced oxidative stress due to ROS released during its transformation into melanin and/or due to its auto-oxidative breakdown. Further studies are required to elucidate L-DOPA metabolism in soybean roots and to identify ROS generated during this pathway. This is the challenge of a new study in progress. Acknowledgements – Research was financially supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). O. Ferrarese-Filho and M.L.L. Ferrarese are research fellow of CNPq. A.R. Soares is the recipient of a CNPq fellowship. The authors kindly thank Aparecida M. D. Ramos and Gisele A. Bubna for their technical assistance. REFERENCES ALBRECHT. C. and KOHLENBACH, H.W. 1990. L-DOPA content, peroxidase activity, and response to H2O2 of Vicia fava L. and V. narbonensis L. in situ and in vitro. Protoplasma 154:144-150. ANAYA, A.L. 1999. Allelopathy as a tool in the management of biotic resources in agroecosystems. Crit. Rev. Plant Sci. 18:697-739. APPEL, H.M. 1993. Phenolics in ecological interactions. The importance of oxidation. J. Chem. Ecol. 19:1521-1552. 36 BINDOLI, A., RIGOBELO, M.P., and DEEBLE, D.J. 1992. Biochemical and toxicological properties of the oxidation products of catecholamines. Free Rad. Biol. Med, 13:391–405. BLUM, U., WENTWORTH, T.R., KLEIN, K., WORSHAM, A.D. KIMG, L.D., GERIG, T.M., and Lyu, S.W. 1991. Phenolic acid content of soils from wheat-no till, wheat-conventional till, and fallow-conventional till soybean cropping systems. J. Chem. Ecol., 17:1045-1068. BOERJAN, W., RALPH, J., and BAUCHER, M. 2003. Lignin biosynthesis. Annu. Rev. Plant Biol, 54:519-546. BOLWELL, G.P., and WOJTASZEK, P. 1997. Mechanisms for the generation of reactive oxygen species in plant defense - a broad perspective. Physiol. Mol. Plant Pathol. 51:347-366. CAKMAK, I., and HORST, W.J. 1991. Effect of aluminum on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 83: 463-468. DEVI, S.R. and PRASAD, M.N.V. 1996. Ferulic acid mediates changes in oxidative enzymes of maize seedlings: Implications in growth. Biol. Plant. 38: 387-395. EINHELLIG. F.A. 1995 Allelopathy. Mechanism of action of allelochemicals in Allelopathy. pp. 97-116, in Inderjit, K.M.M. Dakshini, and F.A. Einhellig (eds.). Allelopathy. Organisms, Processes and Applications, ACS Symposium Series 582, American Chemical Societies. Series 582, Washington, D.C. FERRARESE, M.L.L., RODRIGUES, J.D., and FERRARESE-FILHO, O. 2000. Phenylalanine ammonia-lyase activity in soybean roots extract measured by reversed-phase high performance liquid chromatography. Plant Biol. 2:152-153. FERRARESE, M.L.L., ZOTTIS, A., and FERRARESE-FILHO, O. 2002. Protein-free lignin quantification in soybean (Glycine max) roots. Biologia. 57:541-543. FUJII, Y., SHIBUYA, T., and YASUDA, T. 1991. L-3,4-dihydroxyphenylalanine as an allelochemical candidate from Mucuna pruriens (L.) DC. var. utilis. Agric. Biol. Chem. 55:617-618. FUJII, Y. 1999. Allelopathy of hairy vetch and Mucuna; their application for sustainable agriculture, pp. 289-300, in C.H. Chou (ed). Biochemistry and Allelopathy from Organisms to Ecosystems in the Pacific. Academic Sinica. Taipei. FURUBAYASHI, A., HIRADATE, S., and FUJII, Y. 2005. Adsorption and transformation reactions of L-DOPA in soils. Soil Sci. Plant Nutr. 51:819-825. 37 HACHINOHE, M., SUNOHARA, Y., and MATSUMOTO, H. 2004. Absorption, translocation and metabolism of L-DOPA in barnyard grass and lettuce: their involvement in species-selective phytotoxic action. Plant Growth Reg. 43:237-243. HACHINOHE, M., and MATSUMOTO, H. 2005. Involvement of reactive oxygen species generated from melanin synthesis pathway in phytotoxicity of L-DOPA. J. Chem. Ecol. 31:237-246. HAHLBROCK, K., and SCHEEL, D. 1989. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol. 40: 347-369. HERRIG, V., FERRARESE, M.L.L., SUZUKI, L.S., RODRIGUES, J.D., and FERRARESE-FILHO, O. 2002. Peroxidase and phenylalanine ammonia-lyase activities, phenolic acid contents, and allelochemicals-inhibited root growth of soybean. Biol. Res. 35:59-66. KRUK, I., LICHSZTELD, K., BOUNIAS, M., KLADNA, A., and KUBERA-NOWAKOWSKA, L. 1999. Chemosphere 39:443-453. MERCURI, N.B., and BERNARDI, G. 2005. The ´magic´ of L-dopa: why is it the gold standard Parkinsosn´s disease therapy? Trends Pharm. Sci. 26:341-344. NAKAJIMA, N., HIRADATE, S., and FUJII, Y. 1999. Characteristics of growth inhibitory effect of L3,4-dihydroxyphenylalanine (L-DOPA) on cucumber seedlings. J. Weed Sci. Tech. 44:132138. NISHIHARA, E., PARVEZ, M.M., ARAYA, H., and FUJII, Y. 2004. Germination growth response of different plant species to the allelochemical L-3,4-dihydroxyphenylalanine (L-DOPA). Plant Growth Reg. 42:181-189. NISHIHARA, E., PARVEZ, M.M., ARAYA, H., KAWASHIMA, S., and FUJII, Y. 2005. L-3-(3,4dihydroxyphenyl)alanine (L-DOPA), an allelochemical exuded from velvetbean (Mucuna pruriens) roots. Plant Growth Reg. 45:113-120. PASSARDI, F., COSIO, C., PENEL, C., and DUNAND, C. 2005. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 24:255-265. PATTISON, D.I., DEAN, R.T., and DAVIES, M.J. 2002. Oxidation of DNA, proteins and lipids by DOPA, protein-bound DOPA, and related catechol(amine)s. Toxicol. 177:23-37. POLITYCKA, B. 1999. Ethylene-dependent activity of phenylalanine ammonia-lyase and lignin formation in cucumber roots exposed to phenolic allelochemicals. Acta Soc. Bot. Pol. 68:123-127. SANTOS, W.D., FERRARESE, M.L.L., FINGER, A., TEIXEIRA, A.C.N., and FERRARESE-FILHO, O. 2004. Lignification and related enzymes in Glycine max root growth-inhibition by ferulic 38 acid. J. Chem. Ecol. 30:1199-1208. ST-LAURENT, L., LIVESEY, J., ARNASON, J.T., and BRUNEAU, A. 2002 Variation in L-dopa concentration in accessions of Mucuna pruriens (L.) DC. and in Mucuna brachycarpa Rech. pp. 352-374, in M.B. Flores, M. Eilittä, R. Myhrman, L.B. Carew, and R.J. Carsky (eds.). Proceedings of the International Workshop "Food and Feed from Mucuna: Current Uses and the Way Forward", CIDICCO, Tegucigalpa, Honduras. TAKAHAMA, U., and EGASHIRA, T. 1991. Peroxidases in vacuoles of Vicia faba leaves. Phytochem. 30:73-77. TAKAHASHI, R., and AKIYAMA, T. 1993. Characterization of a melanin associated with low temperature-induced browning in soybean seed coats. Phytochem. 34:587-588. TARAWALI, G., MANYONG, V.M., CARSKY, R.J., VISSOH, P.V., OSEI-BONSU, P., and GALIBA, M. 1999. Adoption of improved fallows in West Africa: lessons from mucuna and stylo case studies. Agroforest. Syst. 47:93-122. VARGAS-AYALA, R., RODRIGUES-KABANA, R., MORGAN-JONES, G., MCINROY, J.A.. and KLOEPPER, J.A. 2000. Shifts in soil microflora induced by velvetbean (Mucuna deeringiana) in cropping systems to control root-knot nematodes. Biol. Control 17:11-22. 39 L-DOPA in root (mg g-1 dry weight) 0.4 0.3 0.2 0.1 0.0 0.1 0.25 0.5 1.0 L-DOPA (mM) in nutrient solution FIG. 1. L-DOPA contents in roots treated exogenously with L-DOPA (0.1 to 1.0 mM) after 24 hr (N = 3 ± SE). 40 TABLE 1. CHANGES IN ROOT LENGTH, ROOT FRESH AND DRY WEIGHTS OF SOYBEAN SEEDLINGS TREATED WITH L-DOPA FOR 24 HR. L-DOPA (mM) 0 0.1 0.25 0.5 1.0 ROOT LENGTH (cm) 2.15 ± 0.101 2.37 ± 0.222ns 1.70 ± 0.066* 0.59 ± 0.056* 0.50 ± 0.066* % 20.9 72.5 76.7 FRESH WEIGHT (g) 2.58 ± 0.012 2.77 ± 0.024 ns 2.44 ± 0.052 ns 2.29 ± 0.099* 2.21 ± 0.053* % 11.2 14.3 DRY WEIGHT (g) 0.174 ± 0.001 0.192 ± 0.001ns 0.162 ± 0.005ns 0.146 ± 0.009* 0.148 ± 0.003* % 16.1 14.9 Note. Means (N = 5 ± SE) significantly smaller than the experiment control (Dunnett´s multiple comparison test) are marked *. ns = not significant at 0.05 level. The symbol % represents inhibition of statistically significant means when compared to control (0 mM). 41 PAL (µmol hr-1 g-1 fresh weight) 0.5 * 0.4 * * 0.3 0.2 0.1 0.0 0 0.1 0.25 0.5 1.0 L-DOPA (mM) FIG. 2. Effects of L-DOPA on phenylalanine ammonia-lyase (PAL). *Values (N = 4 ± SE) differ statistically (Dunnett´s multiple comparison test) from control (P < 0.05). 42 2.0 A * * * * B * 6 * * * 1.5 4 1.0 2 0.5 0 0.0 0 0.1 0.25 0.5 1.0 L-DOPA (mM) 0 Bound POD (µmol min-1 g-1 fresh weight) Soluble POD (µmol min-1 g-1 fresh weight) 8 0.1 0.25 0.5 1.0 L-DOPA (mM) FIG. 3. Effects of L-DOPA on soluble (A) and cell wall-bound (B) peroxidases (POD). *Values (N = 5 ± SE) differ statistically (Dunnett´s multiple comparison test) from control (P < 0.05). 43 Phenolics (mg g-1 dry weight) 35 * 28 * * 0.25 0.5 * 21 14 7 0 0 0.1 1.0 L-DOPA (mM) FIG. 4. Effects of L-DOPA on total phenolics contents. *Values (N = 4 ± SE) differ statistically (Dunnett´s multiple comparison test) from control (P < 0.05). 44 Lignin (mg g-1 dry weight) 25 * * * 20 15 * 10 * 5 0 0 0.1 0.25 0.5 1.0 L-DOPA (mM) FIG. 5. Effects of L-DOPA on lignin contents. *Values (N = 6 ± SE) differ statistically (Dunnett´s multiple comparison test) from control (P < 0.05). 45
Baixar