☰
Explorar
Assinar em
Inscrever-se
Envio
×
Baixar
Sem categoria
Fernando Barreto Rodrigues
Distribuição Normal
Introdução à estatística – Desvio-padrão, variância e coeficiente de
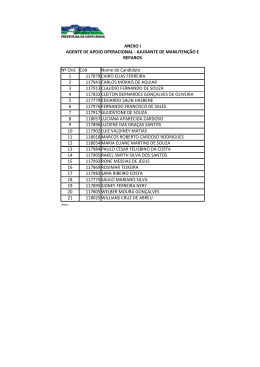
ANEXO I Nome do Candidato 1 117879 CAIRO ELIAS FERREIRA 2
CERTIFICADO DE CONCLUSÃO
291 ESTIMATIVA DE CONSUMO DE BÜFALOS E BOVINOS EM
aula11norm
Alex Martins Varela de Arruda Graduação em Zootecnia pela
Quando há inúmeras variáveis empíricas distribuídas
Lista dos Aprovados - Museu Paraense Emílio Goeldi
Aula – Introdução ao estudo da Química - Docentes