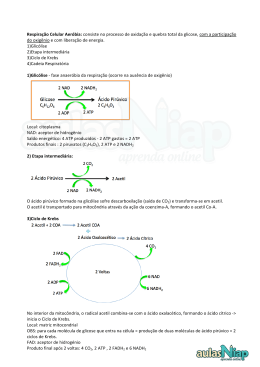
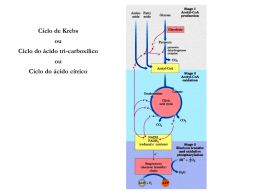
Aula: 13 Temática: Metabolismo aeróbio – parte I Hoje iremos conhecer o ciclo de Krebs e qual a sua importância no metabolismo aeróbio. Acompanhe! O Ciclo de Krebs ou Ciclo do Ácido Cítrico A molécula de glicose pode ser metabolizada a piruvato, de modo anaeróbio, sintetizando ATP através da via glicolítica. Contudo, a glicólise rende apenas uma fração do ATP disponível da glicose. O processamento aeróbio da glicose é a fonte da maior parte do ATP gerado no metabolismo. O processamento aeróbio da glicose começa com a oxidação completa de derivados da glicose até dióxido de carbono. Esta oxidação ocorre no chamado ciclo de Krebs ou ciclo do ácido cítrico, com uma série de reações também conhecida como ciclo dos ácidos tricarboxílicos (GAT ou TGA, de tricarboxylic acid cycle). Este ciclo é a via final comum para a oxidação de moléculas dos alimentos, aminoácidos, ácidos graxos e carboidratos. A maioria delas adentra o ciclo como acetil CoA. O ciclo do ácido cítrico é a função metabólica central da célula. É a porta para o metabolismo aeróbio de qualquer molécula que possa ser transformada em um grupamento acetila ou em ácido dicarboxílico. O ciclo é também uma fonte importante de precursores, não somente para as formas de armazenamento de energia, mas também para os blocos de construção de quaisquer outras moléculas como aminoácidos, bases de nucleotídeos, colesterol e porfirinas. O ciclo de Krebs (ciclo dos ácidos tricarboxílicos ou ciclo do ácido cítrico) é uma continuação da via glicolítica e requer oxigênio, portanto, ocorre somente em organismos aeróbios. Consiste em converter os ácidos láticos e pirúvicos (produtos finais da glicólise), em CO2 e água. Além disto, o ciclo de Krebs constitui um mecanismo comum não só para a oxidação dos produtos da glicólise, mas também para a oxidação final de produtos resultantes do metabolismo de ácidos graxos e de aminoácidos. Assim sendo o ciclo de Krebs BIOQUÍMICA é via comum final do metabolismo dos membros de várias classes de substâncias. O ácido pirúvico ocupa posição central em várias seqüências metabólicas, mas é importante notar que ele pode ter diferentes destinos metabólicos. Consideremos por exemplo a transformação do piruvato em gás carbônico e água: C3H4O3 + 5/2 O2 + 15 ADP + 15 Pi 3CO2 + 2H2O + 15 ATP O piruvato entra no ciclo de Krebs perdendo um CO2 e sendo transformado num radical acetil ligado à coenzima-A (CoA). A unidade acetil entra então no ciclo, condensando-se com ácido oxaloacético, um ácido dicarboxílico com quatro carbonos, dando um ácido tricarboxílico de seis carbonos, o ácido cítrico. Graças à perda e ganho sucessivos de água, este composto é transformado em ácido isocítrico, que se oxida, dando origem ao ácido oxalossuccínico, que por sua vez perde CO2 formando o ácido α-cetoglutárico. O ácido succínico é formado a partir deste composto graças a uma oxidação e perda de CO2. A partir do ácido succínico obtém-se novamente ácido oxaloacético graças a uma oxidação, uma hidratação e uma segunda oxidação (figuras 1 e 2). BIOQUÍMICA Fig. 1 – Esquematização primeira etapa do Ciclo de Krebs. As enzimas implicadas são: (1) oxidase-pirúvica, (2) enzima de condensação (modificado de BENNET & FRIEDEN, 1987). O ciclo de Krebs envolve três passos, nos quais há produção de NADH2 ou NADPH2. Estes nucleotídeos reduzidos naturalmente podem dar origem à formação de ATP pelo processo de fosforilação oxidativa. Fosforilação oxidativa refere-se à fosforilação do ADP em ATP, utilizando para isso a energia liberada nas reações de óxido-redução. É o processo metabólico de síntese de ATP a partir da energia liberada pelo transporte de elétrons na cadeia respiratória, que veremos mais tarde. Em condições aeróbias, o piruvato gerado da glicose sofre descarboxilação oxidativa, formando acetil CoA. As reações do ciclo do ácido cítrico ocorrem dentro da mitocôndria, ao contrário das da glicólise, que ocorrem no citosol (parte fluida do citoplasma). Muitos metabólitos, alguns com função bem conhecida, outros não, surgem como intermediários no ciclo de Krebs. Por exemplo, o ácido cítrico que ocorre em concentração elevada nos tecidos de plantas superiores é um produto do ciclo de Krebs, também o inicio da síntese de muitos aminoácidos. Outros ácidos formados neste ciclo têm papel funcional em fungos e tecidos de plantas. Em cada fase do ciclo há o consumo de 1 mol de acetato sob a forma de acetilcoenzima A, com a produção de dois moles de CO2. O oxaloacetato consumido inicialmente é novamente regenerado, o que permite que o processo funcione continuamente desde que haja entrada de acetil e remoção de água e gás carbônico. Em cinco etapas do ciclo a energia utilizável torna-se disponível, para ser capturada sob a forma de fosfatos de alta energia (ATP). BIOQUÍMICA As reações são estereoespecíficas somente no ácido succínico, onde observamos que os átomos de carbono provenientes da acetil-CoA confundemse com átomos do oxaloacetato. Fig. 2 – Continuação do esquema do Ciclo de Krebs da figura 1. As enzimas implicadas são: (3) aconitase, (4) e (5) desidrogenase isocítrica, (6) oxidase α-cetoglutâmica, (7) sucinil tiocinase, (8) desidrogenase succínica, (9) fumarase, (10) desidrogenase málica. (modificado de BENNET & FRIEDEN, 1987). Um composto de quatro carbonos (oxaloacetato) se condensa com urna acetila, com dois carbonos, resultando um ácido tricarboxílico de seis carbonos BIOQUÍMICA (citrato). Um isômero do citrato é a seguir descarboxilado. O composto resultante, com cinco carbonos (α-cetoglutarato), também sofre descarboxilação oxidativa, originando um composto de quatro carbonos (succinato). O oxaloacetato é então regenerado a partir do succinato. Dois átomos de carbono entram no ciclo corno uma acetila, e dois átomos de carbono deixam o ciclo, na forma de duas moléculas de dióxido de carbono. Três iontes hidreto (portanto, seis elétrons) são transferidos para três moléculas de nicotinamida adenina dinucleotícleo (NAD+), enquanto um par de átomos de hidrogênio (portanto, dois elétrons) é transferido para uma molécula de flavina adenina dinucleotídeo (FAD). A função elo ciclo de Krebs é captar elétrons de alta energia de moléculas dos alimentos. Observe que este ciclo não gera uma grande quantidade de ATP nem inclui o oxigênio como reagente. Ao contrário, o ciclo remove elétrons da acetil-CoA e os utiliza para formar NADH e FADH. No esquema abaixo e nas figuras 1 e 2, observa-se os passos que produzem NADH2- ou NADPH2-. Depois da oxidação de NADH2 ou NADPH2 (via NADH2), vão se originar 3 moles de ATP por molécula de NADH2 ou NADPH2. A conversão de sucinil- BIOQUÍMICA CoA em ácido sucínico, na qual intervém a guanosina-difosfato (GDP), também resulta na produção de um mol de ATP. Nesta reação toma parte o nucleotídeo flavina, FAD. A forma reduzida de FAD é FADH2, pode ser oxidada no sistema dos citocromos, dando origem a duas moléculas de ATP. A oxidação de acetil-CoA para CO2 e H2O no ciclo de Krebs gera um total de 12 moles de ATP por mol de acetil consumido. Visto que 12 moles de fosfato inorgânico (P~) e de ADP são utilizados, e como o acetil entra na forma de acetil-CoA, podemos deduzir a soma geral da oxidação do seguinte modo: A acetil CoA é a fonte de energia para o ciclo de Krebs. Esta molécula importante é formada pela degradação de glicogênio (a forma de armazenamento da glicose), lipídeos e muitos aminoácidos. De fato, os lipídeos contém séries de unidades de dois carbonos que são primeiramente oxidadas a acetil CoA e dai completamente oxidadas a CO2 pelo ciclo de Krebs. BIOQUÍMICA Envie suas dúvidas para nossa tutoria, não deixe que acumulem. Um forte abraço e até a próxima aula! BIOQUÍMICA
Baixar