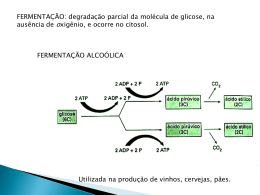
UNIVERSIDADE FEDERAL DA BAHIA FACULDADE DE FARMÁCIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA DE ALIMENTOS CARACTERIZAÇÃO DE ENZIMAS EM DOIS CULTIVARES DE CACAU Theobroma cacao L. ADRIELLE SOUZA LEÃO MACÊDO Salvador – Bahia 2014 2 ADRIELLE SOUZA LEÃO MACÊDO CARACTERIZAÇÃO DE ENZIMAS EM DOIS CULTIVARES DE CACAU Theobroma cacao L. Dissertação apresentada a Faculdade de Farmácia da Universidade Federal da Bahia, como parte das exigências do Programa de PósGraduação em Ciência de Alimentos, para obtenção do título de Mestre. Orientadora: Profª Drª. Eliete da Silva Bispo Co-orientador: Prof° Dr. Sérgio Eduardo Soares Salvador – Bahia 2014 FICHA CATALOGRÁFICA Bibliotecária: Maria Claudete Marques Barbosa Estrela CRB: 5/806 Macêdo, Adrielle Souza Leão Caracterização de enzimas em dois cultivares de cacau theobroma cacao l . / Adrielle Souza Leão Macêdo . – Salvador, 2014. 89f. Orientadora: Profª Drª. Eliete da Silva Bispo Dissertação (Mestrado) – Universidade Federal da Bahia. Programa de PósGraduação em Ciência de Alimentos. Faculdade de Farmácia. Contém referências. 1. 1. Cacau. 2. Cacau – Atividade enzimática I. Bispo, Eliete da Silva. II. Universidade Federal da Bahia, Faculdade de Farmácia. Programa de Pós-Graduação em Ciência de Alimentos . CDD: 633.74 3 Dedico Aos meus pais; Ao meu esposo; Aos meus irmãos; E, aos meus amigos; 4 “Na vida, não vale tanto o que temos, nem tanto importa o que somos, vale o que realizamos com aquilo que possuímos e, acima de tudo, importa o que fazemos de nós!” Chico Xavier 5 AGRADECIMENTOS A Deus por me dar forças e pela presença constante em minha vida; A minha família, principalmente aos meus pais Violeta e Paulo Leão, pelo incansável e verdadeiro carinho, amor, ensinamentos, torcida e apoio. Aos meus queridos irmãos, Adriano e Augusto Leão pelo incentivo e carinho; Ao meu esposo Acássio Macêdo por estar sempre ao meu lado, me inpulsionando para o progresso, por toda confiança, amor, compreensão e incentivo; À Profª. Drª. Eliete Bispo pela orientação, paciência, confiança, ensinamentos e amizade; Ao Profº. Drº. Sergio Soares pelo apoio irrestrito, auxílio e confiança; Aos colegas e agora amigos, que quero levar para sempre, e entre tantos, quero dividir em especial essa alegria com: Margareth Ribeiro, Candice Braga, Lindanor Santana, Fátima Rocha, Leonardo Maciel e Jaff Ribeiro; A Universidade Federal da Bahia pelo suporte; A CAPES pela concessão da bolsa de estudos; Ao CNPq pelo apoio fianceiro ao projeto; Aos amigos da Fazenda Lajedo do Ouro pelo acolhimento, suporte e fornecimento do material de estudo; As Professoras Janice Druzian, Mara Spínola, Alaúse Gil e Rízia de Cássia pelos conhecimentos adquiridos; Aos laboratórios LAPAAC e LAPESCA e a todos os seus colaboradores pelo suporte, acolhimento do projeto de pesquisa e pelo auxílio e parceria nas análises. Enfim, a todos que contribuíram direta ou indiretamente na realização deste trabalho. A Vocês, toda minha gratidão! 6 SUMÁRIO INTRODUÇÃO GERAL .................................................................................................. 15 OBJETIVOS ...................................................................................................................... 18 OBJETIVOS GERAIS ........................................................................................................ 18 OBJETIVOS ESPECÍFICOS .............................................................................................. 18 CAPÍTULO I REVISÃO DE LITERATURA ......................................................................................... 19 1. CACAU ........................................................................................................................... 19 1.1 Características gerais ..................................................................................................... 19 1.2 Influência genética nas características do cacau............................................................ 22 2. PRODUÇÃO DE CACAU NO MUNDO .................................................................... 24 3. PRÉ-PROCESSAMENTO DO CACAU ..................................................................... 26 3.1 Colheita e quebra dos frutos. ......................................................................................... 26 3.2 Fermentação................................................................................................................... 27 3.3 Secagem ......................................................................................................................... 30 3.4 Armazenamento ............................................................................................................. 31 4. ENZIMAS ...................................................................................................................... 32 4.1 Atividade enzimática do cacau. ..................................................................................... 33 4.1.1 Polifenoloxidases ........................................................................................................ 34 4.1.2 Invertases .................................................................................................................... 35 5. FORMAÇÃO DO FLAVOUR DO CHOCOLATE .................................................... 37 REFERÊNCIAS ................................................................................................................. 39 CAPÍTULO II CARACTERIZAÇÃO DA POLIFENOLOXIDASE EM DOIS CULTIVARES DE CACAU (Theobroma cacao L.) PRODUZIDOS NA REGIÃO SUL DA BAHIA, BRASIL.. ... 47 RESUMO ........................................................................................................................... 47 7 ABSTRACT ....................................................................................................................... 47 1. INTRODUÇÃO ............................................................................................................. 49 2. MATERIAL E MÉTODOS .......................................................................................... 51 2.1 Materiais ........................................................................................................................ 51 2.2 Reagentes e padrões....................................................................................................... 51 2.3 Fermentação................................................................................................................... 51 2.4 Coleta das amostras ....................................................................................................... 51 2.5 Caracterização da PPO .................................................................................................. 52 2.5.1 Extração da PPO da polpa .......................................................................................... 52 2.5.2. Extração da PPO das sementes .................................................................................. 52 2.5.3 Purificação da PPO dos extratos ................................................................................. 53 2.5.4 Determinação da atividade da PPO dos extratos ........................................................ 53 2.5.5 Determinação dos parâmetros cinéticos Km e Vmax da PPO ................................... 54 2.5.6 Efeito do pH e temperatura na atividade da PPO ....................................................... 54 2.5.7 Determinação do substrato preferencial da enzima .................................................... 54 2.5.8 Correlação entre os parâmetros de fermentação e a atividade enzimática dos extratos ............................................................................................................................................. 54 2.6 Determinação do teor de proteína nos extratos ............................................................. 54 2.7 Análise Estatística ......................................................................................................... 55 3. RESULTADOS E DISCUSSÃO .................................................................................. 56 3.1 Determinação da atividade da PPO dos extratos ........................................................... 56 3.2 Efeito do pH na atividade da PPO ................................................................................. 57 3.3 Efeito da temperatura na atividade da PPO ................................................................... 59 3.4 Determinação do substrato preferencial da PPO ........................................................... 60 3.5 Correlação entre os parâmetros de fermentação e a atividade enzimática dos extratos ............................................................................................................................................. 61 REFERÊNCIAS .................................................................................................................. 66 8 CAPÍTULO III EXTRAÇÃO E CARACTERIZAÇÃO DA INVERTASE EM DOIS CULTIVARES DE CACAU (Theobroma cacao L.) PRODUZIDOS NO SUL DA BAHIA, BRASIL ............................................................................................................................................. 68 RESUMO ........................................................................................................................... 68 ABSTRACT ....................................................................................................................... 68 1. INTRODUÇÃO ............................................................................................................. 70 2. MATERIAL E MÉTODOS .......................................................................................... 72 2.1 Materiais ........................................................................................................................ 72 2.2 Reagentes e padrões....................................................................................................... 72 2.3 Fermentação................................................................................................................... 72 2.4 Coleta das amostras ....................................................................................................... 72 2.5 Caracterização da Invertase ........................................................................................... 73 2.5.1 Extração da Invertase da polpa ................................................................................... 73 2.5.2. Extração da Invertase das sementes .......................................................................... 73 2.5.3 Purificação da Invertase dos extratos ......................................................................... 74 2.5.4 Determinação da atividade da Invertase dos extratos................................................. 74 2.5.5 Determinação dos parâmetros cinéticos Km e Vmax da Invertase ............................ 75 2.5.6 Efeito do pH e temperatura na atividade da Invertase ................................................ 75 2.5.7 Correlação entre os parâmetros de fermentação e a atividade enzimática dos extratos ............................................................................................................................................. 75 2.6 Determinação do teor de proteína nos extratos ............................................................. 75 2.7 Análise Estatística ......................................................................................................... 75 3. RESULTADOS E DISCUSSÃO .................................................................................. 76 3.1 Determinação da atividade da Invertase dos extratos .................................................... 76 3.2 Efeito do pH na atividade da Invertase .......................................................................... 78 3.3 Efeito da temperatura na atividade da Invertase ............................................................ 79 3.4 Correlação entre os parâmetros de fermentação e a atividade enzimática dos extratos ............................................................................................................................................. 81 9 REFERÊNCIAS .................................................................................................................. 85 CONCLUSÕES FINAIS ..................................................................................................... 87 SUGESTÕES PARA ESTUDOS FUTUROS..................................................................... 89 10 LISTA DE FIGURAS CAPÍTULO I Figura 01. (A) Corte longitudinal de um fruto de cacau - 1: Placenta; 2: Polpa Mucilaginosa; 3: Casca; (B) Corte longitudinal de uma semente de cacau - 4: Testa; 5: Cotilédone; 6: Gérmen ou embrião ................................................................................................................................ 20 Figura 02. Variedades de cacau: (A) Criollo; (B) Forastero; (C) Trinitário ..................... 21 Figura 03. (A) Frutos amontoados na plantação de cacau; (B) Quebra dos frutos ............ 26 Figura 04. A) Cocho de madeira e massa de cacau coberta com folha de bananeira; (B) Revolvimento da massa de cacau ........................................................................................ 27 Figura 05. Mudanças químicas e bioquímicas dentro da semente do cacau durante a fermentação ............................................................................................................................................. 29 Figura 06. Secagem natural ao sol em barcaça................................................................... 31 CAPÍTULO II Figura 01. Atividade da PPO para o substrato catecol em diferentes concentrações (0,05 M a 0,4 M ). (A) Polpa; (B) Semente .............................................................................................. 56 Figura 02. Parâmetros cinéticos pelo método de LINEWEVER-BURK com diferentes concentrações do substrato catecol para a enzima PPO nos dois cultivares de cacau. (A) Polpa; (B) Semente ......................................................................................................................... 57 Figura 03. Efeito do pH na atividade da PPO. (A) Cultivar PH 16; (B) Cultivar TSH 1188 ............................................................................................................................................. 58 Figura 04. Efeito da temperatura na atividade da PPO. (A) Cultivar PH 16; (B) Cultivar TSH 1188 ..................................................................................................................................... 59 Figura 05. Determinação da atividade PPO para substrato preferencial: Catequina e Epicatequina. (A) Polpa; (B) Semente ............................................................................... 61 11 Figura 06. Parâmetros de fermentação e pH ótimo de atuação da PPO. (A) PH 16 - Polpa; (B) PH 16 - Semente; (C) TSH - Polpa; (D) TSH 1188 – Semente........................................... 62 Figura 07. Parâmetros de fermentação e temperatura ótima de atuação da PPO. (A) PH 16; (B) TSH-1188 ............................................................................................................................ 64 CAPÍTULO III Figura 01. Atividade das Invertases para o substrato sacarose em diferentes concentrações (0,05 M a 0,4 M ). (A) Polpa; (B) Semente ................................................................................ 76 Figura 02. Parâmetros cinéticos pelo método de LINEWEVER-BURK com diferentes concentrações de sacarose, para as invertases nos cultivares estudados. (A) PH16 - Polpa; (B) PH16- Semente; (C) TSH 1188 - Polpa; (D) TSG 1188 – Semente.................................... 77 Figura 03. Efeito do pH na atividade das Invertases. (A) Ácida; (B) Neutra.................... 79 Figura 04. Efeito da temperatura na atividade da Invertase. (A) Ácida; (B) Neutra .......... 80 Figura 05. Acompanhamento do pH para polpa e semente durante o processo de fermentação. (A) Cultivar PH 16; (B) Cultivar TSH 1188 ....................................................................... 81 Figura 06. Parâmetros de fermentação e temperatura ótima de atuação da Invertase. (A) PH 16 – Invertase Ácida; (B) PH 16 – Invertase Neutra; (C) TSH - Invertase Ácida; (D) TSH 1188 – Invertase Neutra................................................................................................................... 83 12 LISTA DE TABELAS CAPÍTULO I Tabela 01. Composição de sementes de cacau (Forastero) do Oeste Africano ................. 21 Tabela 02. Maiores produtores mundiais de cacau............................................................. 24 Tabela 03. Principais enzimas ativas durante a fermentação de sementes de cacau .......... 33 CAPÍTULO III Tabela 01. Km e Vmax das invertases para os cultivares com o substrato sacarose. ......... 78 13 RESUMO O interesse do cultivo do cacau (Theobroma cacao L.) está relacionado no aproveitamento de suas sementes para produção de derivados, principalmente para produção de chocolate. Basicamente após a colheita do cacau, são efetuadas as etapas de separação da casca do material interno (sementes e polpa) que é conduzido à fermentação, etapa indispensável para a obtenção de amêndoas de boa qualidade devido a complexas reações bioquímicas que provocam a morte do embrião, hidrólise de açúcares e proteínas, liberação das enzimas e substratos e difusão de compostos fenólicos que entram em contato com as enzimas. Em seguida as amêndoas de cacau são conduzidas à etapa de secagem que além de auxiliar a eliminação da água, possibilita a continuidade das mudanças bioquímicas, iniciadas na fermentação. Durante estas etapas, são gerados inúmeros compostos de aroma e sabor, sendo que as reações que favorecem a formação desses precursores são levadas a cabo por enzimas endógenas do cacau. Com o objetivo de contribuir para a melhoria da qualidade do cacau na produção de chocolates monovarietais, as atividades enzimáticas de dois cultivares de cacau, PH 16 e TSH 118, produzidos na região Sul da Bahia, foram caracterizadas e correlacionadas com os parâmetros de fermentação (pH e temperatura). A atividade enzimática da PPO foi determinada através da medida do aumento da absorbância a 420nm por espectrofotometria, e para a enzima invertase (ácida e neutra), as atividades enzimáticas foram avaliadas pela dosagem de açúcares redutores segundo método de Somogyi-Nelson e determinadas por espectrofotometria. Os resultados expostos demostram que a ação da PPO será favorecida em temperaturas mais baixas, e que o aumento na frequência dos revolvimentos da massa de cacau no início da fermentação, poderá retardar a elevação da temperatura e a queda do pH. Foi observado também que as estabilidades e atividades da invertase, são fatores limitantes para a conversão total da sacarose e formação de precursores de sabor. Foi evidenciada a diferença e especificidade existente entre os cultivares de cacau, e entre polpa e semente em cada cultivar, o que sugere que durante o processo de fermentação devem ser utilizados critérios específicos para cada cultivar, tomando como base os parâmetros ideais de pH e temperatura com o intuito de prolongar o período de melhor atuação destas enzimas, contribuindo assim para a obtenção de amêndoas de maior qualidade para produção de chocolates no que diz respeito à formação e potencialização dos precursores de aroma e sabor. Palavras-chave: cacau, atividade enzimática, polifenoloxidase, invertase. 14 ABSTRACT The interest of the cultivation of cocoa (Theobroma cacao L.) is referenced in the use of its seeds for production of derivatives, mainly for the production of chocolate. Basically after harvest, cocoa steps are effected separation of the inner bark materials (pulp and seeds) which is led to the fermentation step essential for obtaining good quality beans due to the complex biochemical reactions that cause the death of the embryo, hydrolysis of sugars and protein release of enzymes and substrates and distribution of phenolic compounds that come into contact with the enzymes. Then the cocoa beans are conveyed to the drying step that also aids the elimination of water allows the continuity of biochemical changes initiated in the fermentation. During these steps, many flavor and aroma compounds are generated, and the reactions which favor the formation of these precursors are carried out by endogenous enzymes of cocoa. Aiming to contribute to improving the quality of the cocoa production monovarietais chocolates, the enzymatic activities of both cultivars cocoa, PH 16 and TSH 1188 produced in the South of Bahia, were characterized and correlated with fermentation parameters (pH and temperature). The enzymatic activity of PPO was determined by measuring the increase in absorbance at 420nm by spectrophotometry, and the enzyme invertase (acid and neutral), the enzymatic activities were evaluated by measuring reducing sugars second method of Somogyi - Nelson and determined by spectrophotometry. The above results demonstrate that the action of PPO will be favored at lower temperatures, and that the increase in the frequency of turnings of cocoa mass at the beginning of fermentation, can slow the rise in temperature and decrease in pH. It was also observed that the stabilities and activities of invertase, limit the total conversion of sucrose and formation of precursors of flavor factors. It was demonstrated and the difference between existing cultivars specificity cocoa and between each pulp and seed cultures, suggesting that during the fermentation process specific criteria for each cultivar must be used, based on optimal parameters of pH and temperature in order to prolong the activity of these enzymes better, thus contributing to the achievement of higher quality beans to chocolate production with respect to the formation and enhancement of the aroma and flavor precursors. Keywords: cocoa, enzymatic activity, polyphenol oxidase, invertase. 15 INTRODUÇÃO GERAL A partir das sementes do cacau, Theobroma cacao L, é obtido um dos alimentos mais conhecidos e apreciados: o chocolate. Seu flavour é condicionado não apenas a atributos genéticos do cacaueiro (variedade), como também a modificações que ocorrem durante seu beneficiamento (BECKETT, 1994). Basicamente após a colheita do cacau, os frutos são amontoados no chão da plantação cacaueira e abertos com facões. A casca e a placenta são separadas e o material interno (formado de sementes e polpa) é levado às etapas posteriores. A primeira delas é a fermentação que facilita a separação da polpa da semente além de proporcionar a ocorrência de uma série de reações bioquímicas (OETTERER, 2006). Esta etapa é essencial ao processamento, pois é a etapa responsável pelo desenvolvimento dos precursores e inúmeros compostos de sabor. Durante esta etapa, a polpa envoltória das sementes é degradada pela ação sucessiva de microrganismos (leveduras e bactérias ácido-lácticas e ácido-acéticas) naturais do ambiente, com a elevação da temperatura para cerca de 50ºC (CRUZ et al., 2013). Esses microrganismos atuam nos açúcares e ácidos orgânicos da polpa, que são transformados em etanol, ácido láctico e especialmente em ácido acético (SCHWAN e WHEALS, 2004). Os ácidos orgânicos gerados penetram nas sementes, e juntamente com a elevação da temperatura causada pela fermentação aeróbica, causam a morte do embrião e acidificação no tecido armazenado. Com a morte do embrião, é perdida a permeabilidade seletiva de membranas, possibilitando o contato entre enzimas e substratos (LOPEZ, 1986). Estas reações e transformações afetam significativamente a qualidade do produto final, principalmente os aspectos que envolvem a formação de sabor (SCHWAN, 1996). Depois de fermentadas, as amêndoas são secas e torradas a 7% de umidade, o que estabiliza o sabor e produz o aroma e a cor característicos do chocolate, pois muitas das reações bioquímicas iniciadas na fermentação continuam na secagem como as reações de oxidação que proporcionam redução da acidez das amêndoas, além do escurecimento dos cotilédones (ROHAN; STEWART, 1967; LOPEZ; QUESNEL, 1973; BECKETT, 1994), redução do teor de compostos fenólicos que são responsáveis pelo amargor e adstringência (FELLOWS, 2006; DOMINGUES, 2010) e remoção de compostos indesejáveis formados durante a fermentação, como por exemplo, o ácido acético. Segundo Beckett (2009) o desenvolvimento dos precursores do flavour do cacau ocorre nos cotilédones durante fermentação e secagem. Existem dois tipos 16 importantes de células dentro os cotilédones: células de armazenamento contendo gorduras e proteínas, e as células de pigmento contendo compostos polifenólicos e metilxantinas (teobromina e cafeína). Um problema recorrente na indústria de chocolate é a baixa qualidade das amêndoas de cacau, considerando que o processo de fermentação e secagem é feito ainda nas fazendas, sem qualquer controle de processo, uma porcentagem significativa das sementes não sofre as alterações necessárias (principalmente a acidificação do pH e aumento da temperatura) para que as reações enzimáticas se processem de forma satisfatória. Uma das possibilidades de solucionar este problema é o acompanhamento e intervenção, principalmente no processo de fermentação, objetivando caracterizar os compostos, enzimas e melhores condições de processo para melhor uniformizar e aumentar a qualidade das amêndoas de cacau produzidas (AQUARONE et. al., 2001). É sabido que reações que levam à formação de precursores de sabor do chocolate são levadas a cabo por enzimas endógenas da semente do cacau e que na fermentação a principal consequência é o abaixamento do pH, favorecendo a ação destas enzimas. Ao contrário de muitas outras matérias- primas fermentadas, enzimas endógenas desempenham um papel crucial no desenvolvimento do flavour do cacau (LEHRIAN; PATTERSON, 1983). A atividade enzimática, em amêndoas de cacau, durante a fermentação, é conhecida e estudada pelo menos desde a segunda metade do século XX. A confiabilidade e comparação das atividades das enzimas do cacau são complicadas, devido a variações causadas por diferentes genótipos, a origem geográfica, métodos de fermentação utilizados e tipos de cochos empregados. Embora o papel essencial de enzimas endógenas durante a fermentação do cacau tenha sido evidenciado há muitos anos, existe ainda a falta de estudos sistemáticos abordando o comportamento entre diferentes cultivares de cacau (HANSEN et al., 1998). Além disso, não está ainda elucidado como os processos enzimáticos são regulados durante a fermentação, quais substratos enzimáticos/produtos estão relacionados com o sabor de amêndoas com qualidade superior e quais os fatores limitantes para esses processos (disponibilidade de substrato e/ou enzima, genótipo, condições de cultivo ou processo de fermentação). Neste contexto, estudos regionais possibilitam o monitoramento dos parâmetros existentes nos processos de fermentação, para se estabelecer a dinâmica das principais enzimas em diferentes cultivares de cacau, com o objetivo de dimensionar as reais condições do processo e/ou 17 planejar intervenções tecnológicas que contribuam na melhoria da qualidade da matéria-prima para a produção de chocolates monovarietais. 18 OBJETIVOS OBJETIVO GERAL Caracterizar as atividades enzimáticas em dois cultivares de cacau produzidos na região Sul da Bahia, relacionando-as com as condições do processo fermentativo, fornecendo subsídios para futuras intervenções tecnológicas que possam contribuir para melhoria da qualidade da matériaprima na produção de chocolates monovarietais. OBJETIVOS ESPECÍFICOS Monitorar o processo de fermentação dos cultivares de cacau PH16 e TSH-1188, através da medida do pH e temperatura; Caracterizar as enzimas Polifenoloxidase e Invertase, presentes na polpa e sementes dos dois cultivares, antes do início da fermentação (tempo zero); Determinar as condições ótimas de atividade das enzimas caracterizadas; Correlacionar os parâmetros de fermentação, temperatura e pH, com a atividade das enzimas. 19 CAPÍTULO I REVISÃO BIBLIOGRÁFICA 1. CACAU 1.1 Características gerais O cacaueiro é uma planta da família Malvaceae, gênero Theobroma, espécie Theobroma cacao L. originada na Bacia Amazônica e cultivada nas regiões tropicais do mundo. O interesse de cultivo desta espécie está no aproveitamento de suas sementes para produção de derivados de cacau (ALVES, 2002). No início do século XVII, o cacau foi citado pela primeira vez na literatura botânica, por Charles de L’Écluse que o descreveu com o nome de Cacao fructus, porém, em 1737 foi classificado por Linneu como Theobroma fructus, para em 1753 ser modificado para Theobroma cacao L., denominação que permanece atualmente. É uma planta típica dos trópicos úmidos, e é cultivado em regiões onde o clima apresenta variações relativamente pequenas durante o ano, especialmente em termos de temperatura, radiação solar e comprimento do dia (SILVA NETO et al., 2001). Foi introduzido na região Sul da Bahia em 1746, às margens do Rio Pardo na Fazenda Cubículo, hoje município de Canavieiras, por sementes vindas do Pará. A partir daí a cultura se expandiu nessa região, onde as condições climáticas e a riqueza dos recursos naturais contribuíram bastante para torná-la a principal produtora de cacau do país (GRAMACHO et al., 1992; SANCHES, 2006). O interesse de cultivo desta espécie está no aproveitamento de suas sementes (amêndoas) para produção de cacau, de gordura e de chocolate (ALVES, 2002). O fruto tem forma oval com 15 a 20 cm de comprimento do eixo maior, e cor amarela quando maduro. O cotilédone e um pequeno gérmen de planta embrionária são recobertos por uma película denominada testa, e a semente é revestida por uma polpa branca com tons rosados, mucilaginosa e adocicada, representados na Figura 01-A (BECKETT, 1994; MARTINI, 2004; BATALHA, 2009). A polpa é constituída por um parênquima de células esponjosas mucilaginosas contendo água, frutose, glucose, sacarose, pentosanas, ácido cítrico, proteínas e vários sais inorgânicos. A testa secreta a mucilagem e atua como via de transporte entre os cotilédones e a polpa mucilaginosa. O cotilédone apresenta células contendo reservas protéicas, 20 lípides, amido e células polifenólicas. As sementes, Figura 01-B, apresentam um grande e único vacúolo preenchido por polifenóis sendo responsáveis pela cor dos cotilédones (MARTINI, 2004). Este vacúolo é lisado durante a fermentação (URBANSKI, 1992). Figura 01. (A) Corte longitudinal de um fruto de cacau - 1: Placenta; 2: Polpa Mucilaginosa; 3: Casca; (B) Corte longitudinal de uma semente de cacau - 4: Testa; 5: Cotilédone; 6: Gérmen ou embrião. A. B. 4 1 2 5 6 3 Fonte: Fazenda Lajedo do Ouro. Autoria Própria. O cacau é classificado sob três variedades: Criollo, Forastero e Trinitario (Figura 02). O Criollo, originado do lado ocidental dos Andes e o grupo Forastero do lado oriental. O terceiro grupo, Trinitario, é oriundo de um cruzamento (hibridização) entre os dois primeiros, considerado como uma primeira tentativa de melhoramento genético (GRAMACHO et al., 1992; SANCHES, 2006) e de alta qualidade (BATALHA, 2009). As variedades Trinitario e Criollo produzem um chocolate considerado de qualidade excelente pelo suave aroma e sabor (BECKETT, 2009), em contra partida o grupo Forastero responde por 80% da produção mundial, sendo este predominante nas plantações da Bahia, Amazônia, e nos países produtores da África (LEMUS et al, 2002; MARITA et al, 2001). 21 Figura 02. Variedades de cacau: (A) Criollo; (B) Forastero; (C) Trinitário. A. B. C. Fonte: FERREURA et al., (2013). Todos os tipos têm ampla variabilidade, sendo reconhecidos pelo conjunto geral de características. Há, no entanto, algumas distinções mais claras: os Criollos possuem sementes brancas ou de coloração rósea clara e frutos com casca vermelha ou verde, quando imaturos; Forasteros possuem sementes intensamente pigmentadas e frutos verdes, quando novos; e Trinitários são 43 identificados pela associação de caracteres de ambos os tipos anteriores, com coloração de frutos e sementes variáveis (PIRES, 2003). O cacau é uma matéria-prima com grande valor nutricional e sua composição (Tabela 01) varia com a época da colheita, tamanho do fruto, grau de maturação, clima, tipo de solo e manipulação pós-colheita (ZOUMAS et al., 1980). Esta matéria-prima é ingrediente base para formulação de chocolates, achocolatados, biscoitos entre outros (ISAE, 2003). Tabela 01: Composição de sementes de cacau (Forastero) do Oeste Africano. Constituintes Peso Seco (%) Cotilédones 89,60 Testa 9,63 Embrião 0,77 Lipídios 53,05 22 Umidade 3,65 Cinzas 2,63 Nitrogênio Nitrogênio Total 2,28 Nitrogênio proteico 1,50 Teobromina 1,71 Cafeína 0,085 Carboidratos Glicose 0,30 Sacarose 1,58 Amido 6,10 Pectina 2,25 Fibras 2,09 Pentosans 1,27 Polifenóis 7,54 Ácidos Acético 0,014 Oxálico 0,29 Fonte: AFOAKWA, 2010. 1.2 Influência genética nas características do cacau Um dos fatores determinantes na qualidade, sabor e intensidade do chocolate é o genótipo do cacau (AFOAKWA, 2010). Estudos relatados por American Cocoa Research Institute (ACRI) em 1991 demostraram que alguns híbridos da Estação de Pesquisas de Cacau da Costa Rica (CATIE) apresentavam características de aroma e sabor inferiores em relação a outros materiais estudados (MATILIK, 1994). Zamalloa (1994) citados por Efraim (2009) avaliaram as características químicas, físicoquímicas e sensoriais de genótipos dos tipos Forastero e Trinitário cultivados no Estado de São Paulo, em condições climáticas distintas das quais o cacaueiro vem sendo cultivado no mundo em larga escala. Tucci et al. (2002) e Efraim et al. (2006) avaliaram os mesmos genótipos, respectivamente com relação a composição em ácidos graxos, triacilgliceróis e conteúdo de 23 gordura e os teores de compostos fenólicos. Todos os três estudos encontraram diferenças entre os materiais em relação às características avaliadas. Trabalhos conduzidos por Efraim (2009) comprovaram a diferença do comportamento dos cultivares de cacau ao serem monitorados os parâmetros de temperatura e pH durante a fermentação, da mesma forma que foi encontrada diferenças nos liquors, manteiga e chocolate. Uma das principais dificuldades em avaliar comparativamente as diferenças entre variedades de cacau encontra-se na escassez de trabalhos que tenham utilizado materiais distintos submetidos aos mesmos protocolos de fermentação, secagem e torração. Ainda que se tenha dados de alguns trabalhos conduzidos com genótipos de cacaueiro e de sua influência nas características sensoriais, físicas, físico-químicas, não há uma conclusão concreta e generalizada sobre a real influência de genótipos no sabor do chocolate (BUCHELI et al., 2000). 24 2. PRODUÇÃO DE CACAU NO MUNDO O cacau tem importância econômica no contexto internacional por ser um commodity de participação relevante no comércio mundial de produtos agrícolas tanto em importações quanto exportações (GUYTON, 2003). De acordo com o International Cocoa Organization (ICCO, 2011), os maiores produtores mundiais de cacau (Tabela 02) são a Costa do Marfim seguida por Gana, Indonésia, Nigéria, Camarões, Brasil, Equador e Papua Nova Guiné. A estimativa de produção mundial para o período 2012/2013 foi de 4.003 milhões de toneladas (ICCO, 2013). No Brasil, a produção tem grande destaque na região Nordeste, principalmente na Bahia. O Sudeste da Bahia, onde se concentra a produção de cacau no estado, foi o responsável, no ano de 2004, por cerca de 80% da produção nacional. Os principais municípios produtores de cacau da região são Itabuna e Ilhéus (CEPLAC, 2005). Tabela 02: Maiores produtores mundiais de cacau. Países Produção de Cacau t/ ano Costa do Marfim 1.511.000 Gana 1.025.000 Indonésia 440.000 Nigéria 250.000 Camarões 229.000 Brasil 200.000 Papua Nova Guiné 47.000 Fonte: Annual Report 2010/2011 (ICCO, 2011). A cacauicultura foi tida por muito tempo como um dos principais fatores do desenvolvimento econômico do Estado da Bahia e de bastante importância na produção agrícola da região Amazônica (PINTO; PIRES, 1998), envolvendo investimentos em torno de R$ 2,5 bilhões, e aproximadamente 300.000 empregos diretos (FÓRUM NACIONAL DA AGRICULTURA, 1997). A partir de 1989, esse monocultivo passa a ser fortemente comprometido pela queda dos preços, descapitalização e endividamento dos produtores, já antes 25 desestimulados pelas políticas governamentais desfavoráveis, baixa competitividade do setor, exploração de plantações já antigas e em decadência, manejo inadequado da lavoura, estresse hídrico por mais de seis anos consecutivos na região (GRAMACHO et al., 1992; COUTO, 2000; SANCHES, 2006) e a ocorrência, em 1989, do fungo causador da “vassoura-de-bruxa” do cacaueiro – Moniliophthora perniciosa (AIME; PHILLIPS-MORA, 2005; SANCHES, 2006). Essa enfermidade causou danos na região, grandes o suficiente para quase dizimar a cacauicultura na região a ponto de sair de uma produção de 400.000 t/ano para menos de 150.000 t/ano (DIAS, 2001). Entre os esforços concentrados no controle da vassoura-de-bruxa no Brasil, destacam-se as pesquisas sobre novos cultivares de cacau resistentes, produtivos e que originam matériasprimas de qualidade industrial, além de estudos genéticos e de agentes químicos ou biológicos capazes de controlar a doença. Uma das formas mais eficientes encontradas para a recuperação da lavoura cacaueira baiana tem sido a seleção e utilização de clones de cacau provenientes de materiais resistentes à doença, trabalho que tem sido realizado com o apoio de instituições de pesquisa, como o Centro de Pesquisa do Cacau – CEPEC, da Comissão Executiva do Plano da lavoura cacaueira – CEPLAC, e dos próprios cacauicultores com orientação técnica do CEPEC. Com isso, a produção de cacau começou a se recuperar e o Brasil, de um pico de baixa em 1999/2000, de 196 mil toneladas, apresentou elevação da produção para 221 mil toneladas em 2006/2007, de acordo com as estimativas da FAO (FAO, 2008). 26 3. PRÉ-PROCESSAMENTO DO CACAU 3.1 Colheita e quebra dos frutos O desenvolvimento do fruto, desde a fecundação até a maturação, demanda o tempo de cerca de seis meses. Na prática, a maturidade do fruto é reconhecida geralmente pela mudança da cor do mesmo. Por ocasião da colheita, é essencial colher apenas frutos maduros, pois somente estes possuem açúcar e outros substratos em quantidade adequada para uma boa fermentação (LOPES, 2000; CRUZ, 2002). Inicialmente o fruto é colhido com podões, e a seguir os frutos são amontoados no chão da plantação de cacau (Figura 03- A) para serem abertos com o auxílio de facões. Um fator que deve ser levado em consideração é não causar nenhum corte nos frutos, pois desta forma dará inicio ao processo fermentativo antes mesmo de serem colocados nos cochos, comprometendo a qualidade das amêndoas de cacau. O intervalo de tempo para se realizar a quebra dos frutos deve considerar um intervalo de 2 a 3 dias em relação à colheita, pois a separação das sementes da casca é facilitada, caso esse tempo seja estendido acaba por comprometer a qualidade das sementes ocasionando germinação no interior dos frutos, alterando todo o mecanismo do sabor e aroma do chocolate (SERRA, 2004). A casca é então separada (Figura 03-B) e o material interno constituído de amêndoas e polpa (25%do fruto) é levado à fermentação (OETTERER, 2006). Figura 03. (A) Frutos amontoados na plantação de cacau; (B) Quebra dos frutos. A. Fonte: Fazenda Lajedo do Ouro. Autoria Própria. B. 27 3.2 Fermentação Na fermentação o cacau pode ser empilhado aos montões ou em caixas. No sistema de montões, método mais primitivo, são utilizados de 10 kg a 2 t, sendo essa massa revolvida com varas de bambu, de tempos em tempos. No sistema de caixas de madeira ou cochos (Figura 04 – A), o fundo e aas laterais das caixas possuem furos e os lotes são de 200 kg a 1,5 t, com cerca de 1 m de altura. A massa de cacau é transferida (revolvimento) de uma caixa para outra com auxílio de pás de madeira (Figura 04 – B). Durante o processo de fermentação, a massa é coberta com sacos de juta ou folhas de bananeira para reduzir as perdas de calor e evitar o ressecamento excessivo da camada superficial (OETTERER, 2006; PONTILLON, 2009). O processo de fermentação das sementes é essencial ao processamento, pois é responsável pelo desenvolvimento dos precursores e inúmeros compostos de sabor (SCHWAN, 1998). No processo de fermentação, o tipo de sistema, a temperatura do ambiente e da massa, o pH e a acidez da polpa e do cotilédone, o tempo de processo, o revolvimento da massa bem como a microflora presente são fatores de grande importância (ROHAM e CONNEL, 1964; LOPEZ; QUESNEL, 1973). Figura 04. (A) Cocho de madeira e massa de cacau coberta com folha de bananeira; (B) Revolvimento da massa de cacau. A. B. Fonte: Fazenda Lajedo do Ouro. Autoria Própria. 28 É uma etapa indispensável para a obtenção de amêndoas de boa qualidade, devido a complexas reações bioquímicas que provocam a morte do embrião, hidrólise de açúcares e proteínas, liberação de enzimas e substratos, difusão de compostos fenólicos que entram em contato com as enzimas (FORSYTH; QUESNEL, 1958; BECKETT, 1994; BRITO, 2000). A fermentação é essencial para reduzir a acidez, adstringência e amargor nas sementes, formação de açúcares redutores e aminoácidos, que são os precursores da reação de Maillard durante a torrefação (HUANG; BARRINGER, 2010). O tempo requerido para a fermentação das sementes é variável, segundo o material genético. Para a ocorrência das principais reações que levam à formação dos principais precursores de sabor do chocolate, as sementes de cacau, devem ser, geralmente, fermentadas por períodos superiores a cinco dias (BECKETT, 1994). A polpa do cacau é um excelente meio para o crescimento de microrganismos contendo 10-15% de açúcares. A atividade microbiana provoca aumento da temperatura da massa, que contribui para o término do poder germinativo da semente. Têm-se então condições propícias às reações bioquímicas, que culminam com a síntese dos precursores de sabor e aroma (EFRAIN, 2004). A polpa, em cacaus sadios, é isenta de microrganismos, contaminando-se imediatamente durante a quebra dos frutos pelas mãos dos operadores e depois pela exposição ao ambiente. Quando as sementes são removidas dos frutos, ocorre uma inoculação natural de microrganismos do ambiente para a polpa (BECKETT, 2009). Segundo Schwan e Wheals (2004) o processo de fermentação pode ser dividido em três fases: 9 Fase 1: Ação das leveduras anaeróbicas: Nas primeiras 24-36 h, as leveduras transformam o açúcar em álcool sob condições anaeróbicas em um pH abaixo de 4,0. A morte da semente geralmente ocorre no segundo dia, causada pela presença de ácido acético e álcool. É a partir deste momento que as sementes podem ser chamadas de amêndoas de cacau. Normalmente a partir do segundo dia até o final do processo, se faz o revolvimento da massa para que a temperatura não passe de 45°C e favoreça a ação das enzimas. Os revolvimentos podem acontecer de um cocho para o outro ou de um local para o outro, quando o processo for efetuado em montes, e tem por finalidade uniformizar a temperatura e oxigenar a massa (OETERRER, 2006). 9 Fase 2: Ação das bactérias láticas: Estas estão presentes desde o início do fermentação, mas só se tornam dominantes entre 48 e 96 h. Bactérias láticas convertem açúcares e alguns ácidos orgânicos em ácido lático. Por volta do terceiro dia, a massa das amêndoas tem sua temperatura 29 elevada entre 45 e 50° C. Nessa fase, há uma difusão dos conteúdos celulares, iniciando-se uma série de reações relacionadas com as alterações de sabor, aroma e cor da semente. 9 Fase 3: Ação das bactérias acéticas: responsáveis pela conversão do álcool em ácido acético com o auxílio das bactérias acéticas que tornam o tegumento permeável, fazendo com que as amêndoas sofram a ação das enzimas. Ressalta-se a oxidação dos polifenóis que formam ou não complexos com as proteínas e peptídeos levando a redução da adstringência e do amargor. Segundo Beckett (2009) o desenvolvimento dos precursores do sabor do cacau ocorre nos cotilédones durante fermentação e secagem. Existem dois tipos importantes de células dentro dos cotilédones: células de armazenamento contendo gorduras e proteínas, e as células de pigmento contendo compostos polifenólicos e metilxantinas (teobromina e cafeína). A Figura 02 ilustra as reações que ocorrem em sementes de cacau durante sua fermentação. Figura 05. Mudanças químicas e bioquímicas dentro da semente do cacau durante a fermentação. Fonte: LOPEZ, 1986, citado por BECKETT, 2009. A fermentação completa das sementes é diretamente proporcional à qualidade do chocolate. O aroma e sabor característicos são obtidos pela formação dos precursores do flavour, 30 pois as sementes não fermentadas geralmente são amargas e adstringentes, e as parcialmente fermentadas são amargas (KATTENBERG e KEMMINK, 1993; CALIGIANI et al., 2007). O escurecimento não enzimático é proveniente da acumulação de compostos insolúveis como polifenóis, 60% desses compostos são flavanóis (BRITO, 2000). Na fermentação completa do cacau a secreção de algumas enzimas por microrganismos é muito útil como a produção de enzimas pectinolíticas que, em contato com o substrato, podem melhorar a aeração da massa; enzimas amilolíticas, que produzem o aumento da concentração de açúcares redutores pela hidrólise do amido e as enzimas proteolíticas que geram a melhor concentração de aminoácidos livres. Todas estas enzimas são requeridas para a formação do flavour do cacau (AMIN et al., 1998; SCHWAN, 1998). 3.3 Secagem A secagem deve ser iniciada imediatamente após a fermentação. Não deve ser lenta ou mal conduzida para que não possibilite o desenvolvimento de fungos que, quando presentes, conferem sabor desagradável ao produto final. Por outro lado, a secagem não deve ser efetuada de forma demasiadamente rápida através do emprego de temperaturas elevadas, para evitar problemas com a gordura (manteiga de cacau) e com o desenvolvimento do sabor do chocolate (EFRAIN, 2004). O método mais utilizado no Brasil é a secagem feita em barcaças (Figura 06). Que são construções típicas constituídas por um lastro de madeira erguido sobre pilares de alvenaria, e uma cobertura que desliza sobre trilhos. A cobertura, geralmente feita de chapas de alumínio corrugado ou de zinco, é afastada para expor as amêndoas ao sol e, quando fechada, protege contra chuva, sereno e calor excessivo. As amêndoas são espalhadas sobre o lastro da barcaça em uma camada uniforme com cerca de 5 cm de espessura. O revolvimento constante é feito com um rodo de madeira, principalmente no início da secagem, a fim de evitar aglomerados. O processo mais utilizado é a secagem ao sol complementada com aquecimento artificial, via estufas aquecidas por combustão de lenha ou gás liquefeito e/ou energia solar (OETTERER, 2006). Além da eliminação da água, a secagem do cacau possibilita a continuidade das mudanças bioquímicas, iniciadas na fermentação, que vão contribuir para o sabor e aroma característico do chocolate. A secagem é também responsável pela redução da acidez das amêndoas e deve ser 31 conduzida de tal maneira a se obter um teor de umidade em torno de 7%. Secagem excessiva torna a casca quebradiça, enquanto que excesso de umidade favorece o desenvolvimento de fungos (LOPES, 2000). Figura 06. Secagem natural ao sol em barcaça. Fonte: Fazenda Lajedo do Ouro. Autoria Própria. 3.4 Armazenamento Além da importância das etapas de fermentação e secagem à qualidade dos produtos obtidos, as condições de estocagem das amêndoas devem ser observadas, evitando-se o armazenamento de grandes volumes em ambientes com elevada umidade e pouca circulação de ar, umas vez que as amêndoas de cacau são higroscópicas e seu ganho de umidade pode levar ao desenvolvimento de fungos e outros microrganismos indesejáveis (BECKETT, 1994). Esta etapa assume importância devido ao longo tempo em que o cacau pode permanecer armazenado. Começa na fazenda produtora em sacos de aniagem de 60 kg por cerca de 30 dias, fica nas cooperativas vários meses e nos armazéns dos portos por cerca de 15 dias. A amêndoa armazenada deve ter 7% de umidade e estar em equilíbrio com a umidade relativa do ar (70%) (OETERRER, 2006). Segundo Serra (2004) as instalações destinadas ao armazenamento de cacau devem possuir luminosidade e aeração adequadas. 32 4. ENZIMAS As enzimas são proteínas com ação catalítica que aceleram as reações bioquímicas, sendo de extrema importância para o controle dos processos (TORTORA et al., 2005). Fatibello-Filho e Vieira (2002) destacam que a ação enzimática pode ser alterada de acordo com o pH, temperatura, concentração do substrato e pela presença de inibidores. O local onde os substratos se ligam para que as reações se processem, é chamado de sítio ativo, constituído pela interação de alguns resíduos de aminoácidos da cadeia proteica, sendo o responsável pela atividade biológica da enzima, ou seja, no sítio ativo ocorre a reação catalítica. Essencialmente as enzimas apresentam três propriedades principais: estabilidade, atividade e especificidade (BAILEY; OLLIS, 1986; BLANCH; CLARK, 1997; GALVÃO, 2004): x Estabilidade: a capacidade de uma enzima depende de sua estrutura nativa, a qual é mantida por meio de forças de interação (pontes de hidrogênio e ligações de sulfeto, forças de Van der Waals, interações apolares e iônicas). Alterações no ambiente reacional podem debilitar essas interações, alterando a estrutura tridimensional nativa e ocasionando perda parcial ou total da sua funcionalidade biológica. Assim a estabilidade pode ser afetada por variação de temperatura, pH e presença de solventes polares. x Atividade: esta propriedade de uma enzima atua na diminuição da energia de ativação requerida para transformar um substrato em produto, aumentando a velocidade de reação. A capacidade catalítica da enzima reside no seu sítio ativo e este compreende um número pequeno de aminoácidos. O sítio ativo é uma estrutura complexa cuja configuração permite alojar a molécula de substrato na posição correta para que os grupos funcionais da enzima efetuem sua transformação química. x Especificidade: a especificidade define a afinidade de uma enzima por grupos específicos em um determinado substrato. Esta é uma propriedade imprescindível das enzimas enquanto catalisadores. Duas características estruturais são determinantes na especificidade da enzima: O substrato possui ligações químicas que podem ser atacadas pelos grupos funcionais do sítio ativo da enzima e o substrato possui grupos funcionais que se unem à enzima, permitindo seu correto alinhamento no sítio ativo para que a reação possa ocorrer. 33 4.1 Atividade enzimática no cacau A atividade enzimática, em amêndoas de cacau, durante a fermentação, é conhecida e estudada pelo menos desde a segunda metade do século XX. A confiabilidade e comparação das atividades das enzimas do cacau são complicadas, devido a variações causadas por diferentes genótipos, a origem geográfica, métodos de fermentação utilizados e tipos de cochos empregados (HANSEN et al., 1998). O desenvolvimento dos precursores do aroma do cacau envolve a ação de vários microrganismos presentes na polpa e a ação de enzimas sobre os carboidratos, proteínas e polifenóis. Ao contrário de muitas outras matérias- primas fermentadas, enzimas endógenas desempenham um papel crucial no desenvolvimento do sabor do cacau (LEHRIAN e PATTERSON, 1983). Embora a ação das enzimas endógenas durante a fermentação do cacau tenha sido evidenciado há muitos anos, existe ainda a falta de estudos sistemáticos abordando o comportamento entre diferentes cultivares de cacau (HANSEN et al., 1998). A Tabela 03 relaciona as principais enzimas que catalisam reações que ocorrem durante a fermentação das sementes de cacau, seu principal substrato e suas condições ideais de atuação. Essas enzimas possuem importância capital na formação de flavour de chocolate das amêndoas de cacau (HANSEN et al., 1998; LOPEZ; DIMICK, 1991). Tabela 03. Principais enzimas ativas durante a fermentação de sementes de cacau. Enzima Invertases Localização Substrato Produto pH T (ºC) Semente e Sacarose Glicose e 4,0 - 37 frutose 5,25 testa Glicosidases Semente e Glicosídeos, 3- β- β-galactosidase cotilédone galactosidilcianidina Cianidina e 3,8- e 3-α- açúcares 4,5 Peptídeos e 4,7 55 6,0 31,5- 45 arabinosidilcianidina Proteinases Semente e Proteínas cotilédone Polifenoloxidase Semente e aminoácidos Polifenóis (epicatequina) cotilédone Fonte: LOPEZ, 1986, citado por BECKETT, 1988. O-quinona e O-diquinona 34,5 34 As enzimas exibem diferentes estabilidades durante o processo de fermentação, principalmente no que se refere à inativação. Esta é causada pelo calor gerado durante o processo, pela formação de ácidos e presença de polifenóis, portanto o período real da acessibilidade e atuação das enzimas aos substratos, é curto. As proteases, aminopeptidases, invertases e polifenoloxidases são fortemente inativadas durante a fermentação. Já as carboxipeptidases, são parcialmente inativadas, enquanto que as endoproteases e glicosidases permanecem ativas durante todo o processo. O período real da acessibilidade das enzimas aos substratos, e o período de atuação é curto (HANSEN et al., 2000). 4.1.1 Polifenoloxidases A Polifenoloxidase (PPO) é enzima pertencente ao grupo das oxirredutases e é amplamente distribuída na natureza, sendo responsáveis pela oxidação de compostos fenólicos, as quais, na presença de oxigênio, os transformam em quinonas coloridas que participam, posteriormente, das reações de polimerização para dar origem às melonoidinas, caracterizadas pelo aparecimento da coloração marrom-escura (ROELOFSEN, 1958; VÁMOS -VIGYÁZÓ, 1981; WONG, et al., 1990). A atividade das enzimas oxidativas é desencadeada quando a integridade da célula é rompida. Nesta ocasião, os substratos fenólicos, de localização vacuolar, entram em contato com as enzimas catalisadoras das reações de oxidação de polifenóis. O processo oxidativo ocorre quando os substratos fenólicos, as enzimas, o íon metálico e o oxigênio se encontram em condições ideais de pH, temperatura e atividade de água (VITTI, 2007). Mazzafera et al. (2002), explicam que a oxidação de fenóis se dá em função da captura de elétrons por dois átomos de cobre que se encontram no sítio ativo da enzima, havendo o consumo de oxigênio durante o processo. A PPO contém cobre no centro ativo e catalisa dois tipos de reações, ambas envolvendo oxigênio. A primeira reação corresponde à hidroxilação de monofenóis formando orto-difenóis e a segunda à oxidação de orto-difenóis formando orto-quinonas. Os fenóis possuem, pelo menos, um anel benzênico, com um ou mais grupos hidroxila, livres ou substituídos. Algumas formas de fenóis podem ser convertidas em derivados com radicais de oxigênio, extremamente reativos. As polifenoloxidases atuam sobre uma grande variedade de substratos: citase p-cresol, tirosina e 35 ácido p-cumárico como substratos monofenólicos, enquanto catecol, diidroxifenilalanina e ácido clorogênico são substratos difenólicos (VAMOS-VIGYAZO, 1981; CAMPOS et al., 2004). A presença e a atividade da enzima PPO, durante a fermentação e a secagem das amêndoas de cacau, são um dos fatores responsáveis pelo desenvolvimento dos precursores do sabor, começando na fase oxidativa da fermentação e continuando na secagem (ROELOFSEN, 1958; VÁMOS-VIGYÁZÓ, 1981; WONG, et al., 1990). O papel da PPO e sua importância na produção do sabor característico de chocolate não estão bem compreendidos. Contudo, uma baixa atividade da enzima ou condições que podem levar a uma redução da atividade têm sido apontada como fatores responsáveis por uma queda na produção de precursores de sabor de chocolate. A importância desta enzima é frequentemente mencionada de forma implícita na redução do sabor adstringente do cacau (REEVES et al., 1988). 4.1.2 Invertases A enzima invertase também chamada de β-frutofuranosidase, não só catalisa a hidrólise de sacarose a glicose e frutose, mas em algumas condições também é capaz de catalisar a transfrutosilação para produzir frutooligossacarídeos (FOS) (RUBIO; RUNCO; NAVARRO, 2002). Na fermentação do cacau são responsáveis pela liberação de açúcares redutores indispensáveis à formação do sabor durante a torrefação (HANSEN et al., 1998; LOPEZ; DIMICK, 1991). As invertases são, usualmente, classificadas de acordo com seu pH e localização celular. A invertase ácida com atividade máxima em pH 5,5, é predominante em tecidos em crescimento (localizadas no vacúolo e/ou parede celular) e invertase neutra, com um máximo de atividade em pH 7,0 e com predominância em tecidos completamente expandidos (HUSSAIN et al., 2008). Ashokkumar, Kayalvizhi & Gunasekaran (2001) afirmam que a invertase catalisa a hidrólise da sacarose em quantidades equimolares de frutose e glicose, processo conhecido como inversão. Invertase hidrolisa sacarose sob concentrações de sacarose mais baixas que 10% (peso/vol.) (RUBIO; RUNCO; NAVARRO, 2002). O mecanismo de ação da invertase não é totalmente conhecido, mas estudos sugerem que há um envolvimento de um ânion carboxilato e uma histidina residual na atividade catalítica desta enzima (MARQUEZ, 2007). A sacarose resultante da fotoassimilação do CO2 é a principal 36 forma de açúcar translocada nas plantas. O metabolismo da sacarose está estreitamente ligado com a atividade das enzimas sacarose sintetase, sacarose fosfato sintetase, fosfatase e invertases. A atividade da invertase pode ser medida seguindo-se vários caminhos: polarimetricamente, seguindo-se as mudanças na rotação ótica; determinação da presença de açúcares redutores; ou estimando-se a concentração de glicose resultante da hidrólise pelo uso de glicose oxidase. 37 5. FORMAÇÃO DO FLAVOUR DO CHOCOLATE O mais notável atributo do chocolate é o seu sabor único. A combinação e o equilíbrio dos numerosos compostos que contribuem para o flavour final dependem de diversos fatores dentre os quais estão genética, condições ambientais, condições de colheita e de processamento. Essa complexidade é evidente quando se considera que, ainda hoje este flavour tão desejável não foi reproduzido pela indústria química (REINECCIUS, 2006). Durante as etapas de beneficiamento do cacau são gerados os precursores do sabor característico dos produtos de cacau, como também compostos que não mais sofrerão modificações e que contribuirão para esse sabor. No entanto, concomitantemente, nas etapas de fermentação e secagem, ocorrem as maiores perdas de compostos fenólicos presentes naturalmente e em elevadas quantidades nas sementes do cacau. Os compostos fenólicos no cacau, são componentes importantes do sabor, responsáveis principalmente pelo amargor e adstringência. Segundo alguns autores, os produtos da reação de substâncias fenólicas estão entre os componentes mais importantes do sabor do cacau, e os produtos da condensação de compostos fenólicos gerados durante a fermentação e secagem são responsáveis pela cor marrom do cacau e do chocolate (ELWERS et al, 2009). Os principais fenólicos encontrados nas sementes de cacau estão dentro das classes dos taninos e dos flavonoides. Os flavonoides presentes incluem flavanóis, flavonóis, antocianinas, flavonas e flavanonas. Entre estes, os flavanóis são os mais abundantes, sendo a (+)-catequina e a (–)-epicatequina os principais representantes. A (–)-epicatequina tem sido reportada como o principal flavanol monomérico do cacau, representando aproximadamente 35% do conteúdo total dos fenólicos (WOLLGAST; ANKLAM, 2000). Estes compostos são armazenados em células de pigmentos dos cotilédones, também chamadas células de armazenamento de polifenóis, e a quantidade de antocianinas nessas células de pigmentos dão aos grãos coloração que variam de branco a vinho. Quando os grãos de cacau passam pelos processos de fermentação e secagem, que são passos críticos no processamento do cacau, as membranas das células de pigmento se rompem e o seu conteúdo é exposto a outros componentes no grão. Com isso, os polifenóis passam por uma série de reações, como a difusão da epicatequina pela membrana da célula de armazenamento, ficando exposta a reações de oxidação e polimerização, formando taninos complexos (RUSCONI, CONTI, 2010). 38 Os polifenóis do cacau têm sido estudados há várias décadas devido à influência negativa que exercem no sabor, conferindo o amargor e a adstringência verificados em produtos com elevados teores desses compostos (ROAHN e CONNEL, 1964; CROSS et al., 1982; VILLENEUVE et al., 1985; BRITO, 2000; SOARES, 2001). Em contra partida, descobertas mais recentes sobre os efeitos benéficos desses compostos à saúde humana têm provocado interesse em mantê-los durante o processamento dos produtos obtidos do cacau, sem prejuízo do sabor (KEALEY et al., 1998; KEALEY et al., 2004; EFRAIM, 2004; RIZO, 2006). Uma das mais importantes e complexas reações que envolvem a formação do flavour é a reação de Maillard, devido presença de aminoácidos livres e açúcares redutores formados durante a etapa de fermentação do cacau. A reação de Maillard é uma reação entre carbonilas e aminas. Geralmente, as carbonilas em alimentos são açúcares redutores, enquanto aminas podem vir de aminoácidos ou proteínas (PORTE et al., 2007). Para que as amêndoas de cacau possam originar o sabor e o aroma característico de um chocolate de excelente qualidade é imprescindível que sejam fermentadas, secas e torradas num processo de complexas reações enzimáticas e químicas como a hidrólise de açúcares e proteínas, liberação de enzimas, difusão de compostos fenólicos e morte do embrião (FORSYTH; QUESNEL, 1957). Através destas reações, todos os precursores de aroma do cacau interagem para produzir componentes de sabor, como álcoois, éteres, furanos, tiazoles, piroles, ácidos, ésteres, aldeídos, iminas, aminas, oxazolas, pirazinas e pirróis (MISNAWI et al, 2003). 39 REFERÊNCIAS AFOAKWA, E, O. Chocolate Science and Technology. United Kingdom: John Wiley And Sons Ltd, New Delhi, Índia, 2010. AIME, M.C., PHILLIPS-MORA, W. The causal agents of witches’ broom and frosty pod rot of cacao (chocolate, Theobroma cacao) form a new lineage of Marasmiaceae. Mycologia, n. 97, p. 1012-22, 2005. AQUARONE, E.; BORZANI, W.; SCHMIDELL, W.; LIMA, U. A. Biotecnologia industrial. Sao Paulo: E. Blucher, 2001. vol. 4. ALVES, S. A. M. Epidemiologia da vassoura de bruxa (Crinipellis perniciosa (STAHEL) SINGER) em cacaueiros enxertados em Uruçuca, Ba. 2002..Dissertação (Mestrado em Agronomia). Universidade de São Paulo. Escola Superior de Agricultura “Luiz de Queiroz”. Piracicaba – SP, p 70, 2002. AMIN, I.; JINAP, S.; JAMILAH, B. Proteolytic Activity (Aspartic Endoproteinase and Carboxypeptidase) of Cocoa Bean during Fermentation. Journal Scienc Food Agricultural. v.76, p.123-128, 1998. ASHOKKUMAR, B., KAYALVIZHI, N. & GUNASEKARAN, P. Optimization of media for β-fructofuranosidase production by Aspergillus niger in submerged and solid state fermentation. Process Biochmistry. n. 37, p. 331-338. 2001. BAILEY, J. E. e OLLIS, D. F. Bichemical Engineering Fundamentals. Second edition. New York, 1986. BATALHA, P. G. Caracterização do cacau catongo de São Tomé e Príncipe. Lisboa. 2009. Mestrado (Mestre em Engenharia de Alimentos – Tecnologia de Produtos vegetais) Universidade Técnica de Lisboa. Instituto Superior de Agronomia. Lisboa – Portugal. BECKETT, S. T. Fabricación y utilización industrial del chocolate; trad. de Mariano Gonzaléz Alonso. 1 ed. Zaragoza: Acribia, 1988. 423 p ______. Fabricación y utlización industrial del chocolate. Zaragoza: Editorial Acribica, 1994, 432p. ______. Industrial chocolate manufacture and use. 4 ed. London: Chapman and Hall, p.20-23, 2009. BLANCH, H. W. e CLARK, D. S. Biochemical Engineering. Estados Unidos. 1997. 40 BRITO, E.S. Estudo de mudanças estruturais e químicas produzidas durante fermentação, secagem e torração do cacau (Theobroma cacao L.); e propostas de tratamento para o melhoramento de sabor. 2000. 176p. Tese (Doutorado em Tecnologia de Alimentos) Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Campinas, 2000. BUCHELI, P. et al. Strategy for assessing cocoa flavour of a large number of samples for selection and breeding. Proceedings of 13th International Cocoa Research Conference, Kata Kinobalu, Sabah, Malaysia, p. 865-870, 2000. CALIGIANI, A. et al. Detection of Chiral markers in cocoa beans of different quality and geographic origin. Chirality. v.19, p.329-334. 2007. CAMPOS, A, D et al. .Atividade de peroxidase e polifenoloxidase na resistência do feijão à antracnose. Pesq. agropec. bras., v. 39, p. 637-643, 2004. COMISSÃO EXECUTIVA DO PLANO DE RECUPERAÇÃO DA LAVOURA CACAUEIRA. Estatística do cacau na Bahia. Ilhéus: CEPLAC/CENEX, 2005. 1 p. COUTO, V. de A. O território do cacau no contexto da mundialização. Bahia Análises & Dados. Salvador-BA. SEI v.9, n.4, p.38-52, Março 2000. CROSS, E.; VILLENEUVE, F.; VINCENT, J. C. Recherche d’un índice de fermentation du cacau. Café, Cacau hé, v. 16, n. 2, p. 109-113, 1982 CRUZ, C. L. C. V. Melhoramento do sabor de amêndoas de cacau através de tratamento térmico em forno convencional e de microondas. Campinas, 2002. 101p. Dissertação (Mestre em Tecnologia de Alimentos). Faculdade de Engenharia de Alimentos. Universidade Estadual de Campinas. Campinas – SP. CRUZ, J, F, M; LEITE, P, B; SOARES, S, E; BISPO, E, Silva. Assessment of the fermentative process from different cocoa cultivars produced in Southern Bahia, Brazil. African Journal of Biotechnology, v. 12 (33), p. 5218-5225, 2013. DEUNER, S. et al. Caracterização parcial da invertase ácida solúvel em tubérculos de batata: energia de ativação e efeito de inibidores. Revista Brasileira de Agrociência, Pelotas – RS, v. 11, n. 45-50, 2005. DIAS, L. A. S.. Melhoramento genético do cacaueiro. Viçosa-MG. FUNAPE, UFG. 578 pág., 2001. DOMINGUES, 2010. E.S. Seleção de linhagens de leveduras pectinolíticas para fermentação das sementes de cacau (Theobroma cacau). 2010. 80p. Dissertação. (Mestrado) - Escola Superior de agricultura Luis de Queiroz. 2010. 41 EFRAIM, P. Estudo para minimizar as perdas de flavonóides durante a fermentação de cacau para produção de chocolate. 2004. 114 p. Dissertação (Mestrado em Tecnologia de Alimentos) – Universidade Estadual de Campinas – UNICAMP, Campinas. EFRAIM, P. et al. Teores de Compostos Fenólicos de Sementes de Cacaueiro de Diferentes Genótipos. Brazilian Journal Food Technology, v. 9, n. 4, p. 229-236, 2006.. EFRAIM, P. Contribuição à melhoria de qualidade de produtos de cacau no Brasil, através da caracterização de derivados de cultivares resistentes à vassoura de bruxa e de sementes danificadas pelo fungo. 2009. 226p. Tese (Doutor em Tecnologia de Alimentos). Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Campinas, SP.2009. EFRAIM, P. et al. Influência da fermentação e secagem de amêndoas de cacau no teor de compostos fenólicos e na aceitação sensorial. Ciência e Tecnologia de Alimentos, Campinas, v. 30, n. 1, p. 142-150, 2010. ELWERS, S. et al. Differences between the content of phenolic compounds in Criollo, Forastero and Trinitario cocoa seed (Theobroma cacao L.). European Food Research and Technology, v. 229, p. 937–948, 2009. FATIBELLO-FILHO, O; VIEIRA, I. da C. Uso analítico de tecidos e de extratos brutos vegetais como fonte enzimática. Química Nova, v. 25, n. 3, p.455-464, 2002. FELLOWS, P.J. Tecnologia do processamento de alimentos. Princípios e prática. 2° Edição. Ed Artmed, p 183-205, 602, 2006. FERREURA, A, C, R. et al. Guia de Beneficiamento de cacau de qualidade. Instituto Cabruca, 2013 FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS – FAO. Disponível em:<http://apps.fao.org/page/collections?subset=agriculture> . Acesso em 01 de fevereiro 2013. FÓRUM NACIONAL DA AGRICULTURA. Grupo Temático Cacau. Proposta básica de políticas para a cadeia produtiva do cacau. [s. l.] FNA, p. 108, 1997. FORSYTH, W.G.C; QUESNEL, V.C. Cacau glycosidase and colour changes during fermentation. Journal of the Science of Food and Agriculture, London, v. 8, p. 505-509, 1958. GALVÃO, C. M. A. Hidrólise controlada de proteínas do soro lático usando tripsina e quimotripsina imobilizadas em diferentes suportes. (Tese de doutorado em engenharia química). São Carlos, SP. 2004, 191p. 42 GOMEZ, M L P A; LAJOLO, F M; CORDENUNSI, B R. Metabolismo de carboidratos durante o amadurecimento do mamão (Carica papaya, L. cv. Solo): influência da radiação gama. Ciência e Tecnologia de Alimentos. 1999, v.19, 246-52.ISSN 0101-2061. GRAMACHO, I. DA C. P. et al. Cultivo e Beneficiamento do Cacau na Bahia. Ministério da Agricultura e Reforma Agrária. CEPLAC. Ilhéus - BA - Brasil, 1992. GUYTON, B. Commodities – Cocoa Review. 2003 Issues, trends and performance of the chocolate and confectionery industries, New York. 40p. HANSEN, C.E.; DEL OLMO, M.; BURRI, C. Enzyme activities in cocoa beans during fermentation, Journal of the Science of Food and Agriculture, v.77, 273-81, 1998. HANSEN, C.E; MAÑEZ, A; BURRI, C; BOUSBAINE, A. Comparison of enzyme activities involved in flavour precursor formation in unfermented beans of different cocoa genotypes. Journal of the Science of Food and Agriculture, London, v. 80, p. 1193-1198, 2000. HUANG, Y.; BARRINGER, S. A. Alkylpyrazines and Other Volatiles in Cocoa Liquors at pH 5 to 8, by Selected Ion Flow Tube-Mass Spectrometry (SIFT-MS). Journal of Food Science, v. 75, n. 1, 2010 HUSSAIN, A.; RASHID, M.; PERVEEN, R.; ASHRAF,M. Purification, kinetic and thermodynamic characterization of soluble acid invertase from sugarcane. Plant Physiology et Biochemistry, in press, 2008. ICCO (INTERNATIONAL COCOA ORGANIZATION). Annual Report 2010/2011. Disponível em:<http://www.icco.org/>. Acesso em: 29 de fevereiro de 2013. ICCO (INTERNATIONAL COCOA ORGANIZATION). Boletim Trimestral 2013 de Estatísticas do Cacau. Disponível em:<http://www.icco.org/>. Acesso em: 29 de fevereiro de 2013. ISAE/FGV. Cacau: Pontencialidades regionais e estudo de viabilidade econômica. Manaus: Suframa, 2003. KATTEMBERG, H.R.; KEMMINK, A. The flavor of cocoa in relation to the origin and processing of the cocoa beans. Developing food science., v. 32, p.1-22. 1993. KEALEY, K. S. et al. Mars Incorporeted. Cocoa components, edible products having enhanced polyphenol content, methods of making same medical uses. Patent Corporation Treaty (PCT) WO 98/09533, 1998. KEALEY, K, S. et al. Incorporeted. Method for producing fat and/or solids from beans and compositions containing polyphenols. US Patent Application 2004/0058022, 2004. 43 LEHRIAN , D, W; PATTERSON G. R. Cocoa fermentation, In: Biotechnology, a Comprehensive Treatise,. v. 5, p. 529–575, 1983. LEMUS, M. et al. Efecto del mezclado de cacaos tipos criollo y forastero de la localidad de cumboto sobre algunas características físicas de los granos durante la fermentación. Agronomía Tropical, v. 52, n. 1, p. 45-58, 2002. LEHRIAN, D, W; PATTERSON G. R. Cocoa fermentation, In: Biotechnology, a Comprehensive Treatise,. v. 5, p. 529–575, 1983. LOPEZ, A, S.; QUESNEL, V. C. Volatile fatty acid production in cacao fermentation and the effect on chocolate flavour. Journal of the Science of Food and Agriculture, v. 24, n. 3, p. 319324, 1973. LOPEZ, A, S. Chemical changes occurring during the processing of cacao. In: Symposium Cacao Biotechnology. Proceedings. University Park: The Pennsylvania State University, Pennsylvania, p.19-54, 1986. LOPEZ, A, P; DIMICK, P.S. Cap. 25: Enzymes involved in cocoa curing In: Food Enzymology Vol. 2. Elsevier Applied Science. p. 211-236,1991. LOPES, A. S. Estudo químico e nutricional de amêndoas de cacau (Theobroma cacao L.) e cupuaçu (Theobroma grandiflorum Schum) em função do processamento. Campinas, 2000. 130p. Dissertação (Mestre em Tecnologia de Alimentos). Faculdade de Engenharia de Alimentos. Universidade Estadual de Campinas. Campinas, SP. MALTIK, B, K. Cocoa: Post Harvest Practines, Part I. The Manufacturing Confectioner. V. 74, n. 6, p. 95-96, 1994. MARTINI, M. H. Caracterização das sementes de seis espécies de Theobroma em relação ao Theobroma cacao L. 2004. Tese (Doutorado em Alimentos e Nutrição). Faculdade de Engenharia de Alimentos. Universidade Estadual de Campinas. Campinas – SP. MARITA, J. M. et al. Analysis of genetic diversity in Theobroma cacao with emphasis on witches’ broom disease resistance. Crop science, v. 41, 2001. MARQUEZ, L. D.S. Produção de açúcar invertido pelo uso de invertase imobilizada em resinas. Uberlândia: Faculdade de Engenharia Química, UniversidadeFederal de Uberlândia, 2007. 124p. Dissertação (Mestrado). MAZZAFERA, P. et al. Extração e dosagem da atividade da polifenoloxidase do café. Sci. agric, v. 59, p. 695-700, 2002. MISNAWI, J, S; JAMILAH, B; NAZAMID, S . Sensory properties of cocoa liquor as affected by polyphenol concentration and duration of roasting. J. Food Qual. Pref.15:403-409, 2003. 44 NASCIMENTO, R. et al. Distribuição de amido, açucares solúveis e atividade de invertases em explantes de soja sob várias concentrações de sacarose e diferentes fontes de nitrogênio. Revista Brasileira de Fisiologia Vegetal, v. 10, 125-30, 1998. OETTERER, M. Tecnologias de obtenção do cacau, produtos do cacau e do chocolate. In: OETTERER, M; M., Regitano d´Arce A.; SPOTO, M.H.F. (Org.). Fundamentos de Ciência e Tecnologia de Alimentos. Barueri: Manole, 2006, v. 1, p. 1-50. PINTO, L. R. M.; PIRES, J. L.. Seleção de Plantas de Cacau Resistentes à assoura-debruxa. Boletim Técnico n. 181 – Ministério da Agricultura e do Abastecimento. CEPLAC/CEPEC. Ilhéus-BA, 1998. PIRES, J.L. Avaliação quantitativa e molecular de germoplasma para o melhoramento de cacaueiro com ênfase na produtividade, qualidade dos frutos e resistência à doenças. 226p. 2003. Tese (Doutorado em Genética e Melhoramento) – Universidade Federal de Viçosa, Viçosa, 2003. PONTILLON, J. Do cacao ao tablete. A Ciência na cozinha, São Paulo, v. 1, p. 62-71, 2009. Porte, A., Rezende, C. M., Antunes, O. A. C. Produção de voláteis via sistemas modelo de Maillard usando glicose e l-aminoácidos sob diferentes condições de pH. Rev. Univ. Rural. Sér. Ci. Exatas e da Terra, Seropédica, RJ, EDUR, vol. 26, n. 1-2, jan-dez, p. 12-32, 2007. REEVES, S.G. et al. Biochemical studies of Cocoa Bean o-diphenol O2 oxidoreductase (catechol oxidase). Food Chemistry, Kidlington, 290-219,1988. REINECCIUS, G.. Sabor chemistry and technology. Boca Raton: Taylor Francis, p. 489, 2006. RIZO, D. C. Barry Callebaut conirma el poder de los polifenoles en el chocolate. Dulcelandia, v. 65, n. 789, p. 33-37, 2006. ROELOFSEN,P.A. Fermentation dryng and storage of cocoa beans. Advances in Food Research, San Diego, p.226-229,1958. ROHAN, T. A.; CONNEL, M. he precursors of chocolate aroma: A study of the lavonoids and phenolic acids. Journal of Food Science, v. 29, n. 4, p. 460-463, 1964. ROHAN, T.H.; El Beneficio del cacao bruto destinado al Mercado. Roma, 1964. ROHAN, T.A.; STEWART, T. The precursors of chocolate aroma: production of reduction sugars during fermentation of cocoa beans. Journal of Food Science, v.32, n. 4, p. 399-402, 1967. 45 RUBIO, M.C., RUNCO, R., NAVARRO,A.R. Invertase from a strain of Rhodotorula glutinis. Phytochemstry, v.61, p.605-609, 2002. RUSCONI, M.; CONTI, A. Theobroma cacao L., the Food of the Gods: A scientific approach beyond myths and claims. Pharmacological Research, v. 61, p. 05–13. 2010. SANCHES, C, L, G. Murcha-de-ceratocystis (ceratocystis cacaofunesta) no sul da bahia: metodologia para seleção de genótipos de cacaueiro resistentes e estudos preliminares descritivos do patógeno. Dissertação apresentada à Universidade Estadual de Santa Cruz, Ilhéus, Bahia, 2006. SCHWAN, R. F. Microbiology of cocoa fermentation: a study to improve quality. In: CONFERÊNCIA INTERNACIONAL DE PESQUISA EM CACAU, 12, Salvador, BA, nov. 1996. Anais. Salvador: CEPLAC, 1996. ______. Cocoa fermentations conduced with a defined microbial cocktail inoculum. Applied and environmental microbiology. Lavras – MG. v.64, p.1477-1483. 1998. SCHWAN, R,F; WHEALS, A, E.The microbiology of cocoa fermentation and its role in chocolate quality. Critical Reviews in Food Science e Nutrição. Boca Raton, 44(4): 205-221, 2004. SERRA, W. S. Manual do Cacauicultor: com perguntas e respostas. p177-207, 2004. SILVA NETO, P. J. da; MATOS, P, G, G, de; MARTINS, A, C; SILVA, A, P. Sistema de produção de cacau para a Amazónia brasileira. 125p, Belém, CEPLAC, 2001. SERRA, W. S. Manual do Cacauicultor: com perguntas e respostas. p177-207, 2004. SILVA NETO, P. J. Sistema de Produção de cacau para a Amazônia brasileira. Belém, PA. CEPLAC. 2001. 125p SOARES, M. S. Estudo do melhoramento do sabor de cacau (Theobroma cacao L.) através de ação enzimática durante a fermentação. 2001. 107 p. Dissertação (Mestrado em Tecnologia de Alimentos) – Universidade Estadual de Campinas – UNICAMP, Campinas. SOUNIGO, O. et al. Assessment of the value of doubled haploids as progenitors in cocoa (Theobroma cacao L.) breeding. Journal of general and applied microbiology, França, v. 49, n. 3, 2003. SOUZA, A. S. L. Avaliação da estabilidade térmica e oxidativa de chocolates amargos. 2010. Dissertação (Mestrado em Ciência e Tecnologia de Alimentos), Universidade Federal da Paraíba. João Pessoa – PB, p.110, 2010. 46 TORTORA, G.J. ; FUNKE, B. R. ; CASE, C. L. Metabolismo microbiano. In:Microbiologia. 8.ed. Porto Alegre: Artmed, 2005. cap. 5, p. 111-121. TUCCI, M.L.S. Seleção de genótipos para produção de manteiga de cacau no Vale do Ribeira, São Paulo, Brasil. Dissertação (Mestrado). Escola Superior de Agricultura “Luiz de Queiroz”. Universidade de São Paulo, Piracicaba, p.120. 1997. URBANSKI, J. J. Chocolate flavor/origins and descriptions. The effects of process and bean source. The Manufacturing Confectioner, v. 72, p.69-82, 1992. WOLLGAST, J.; ANKLAM, E. Polyphenols in chocolate: is there a contribution to human health? Food Research International, Italia, v. 33, p. 449- 459, 2000. WONG,M.K.; DIMICK,P.S.; HAMMERSTEDT, R.H. Extraction and high performance liquid chromatographic enrichment of polyphenol oxidase from Theobroma cacao seeds. Journal of Food Science, Chicago, v.55,n.4,p.1108-1111,1990. VÁMOS-VIGYÁZÓ, L.Polyphenoloxidase and peroxidase in fruit and vegetables.CRCCritical Reviews in Food Science and Nutrition, n.49,1981. VILLENEUVE, F.; CROS, E.; MACHEIX, J.J. Effet de la fermentation sur lês activités peroxidasiques et polyphenoloxidasiques de la féve de cacao. Café, Cacao, Thé, Paris, v. 33, n.2, p. 113-120, 1985. VITTI, M. C. D. Respostas fisiológicas e bioquímicas de diferentes cultivares de batatas ao processamento mínimo. Tese (Doutorado), Escola Superior de Agricultura Luiz de Queiroz, 2007. ZOUMAS, B.L.; KREISER, W.R.; MARTIN, R.A. Theobromine and caffeine content of chocolate products. Journal of Food Science, v 45, p 314-316, 1980. ZAMALLOA, W. A. C. Caracterização físico-química e avaliação de metilpirazinas no desenvolvimento do sabor, em dez cultivares de cacau (Theobroma cacao L.) produzidos no Estado de São Paulo. Dissertação (Mestrado em Tecnologia de Alimentos) - Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Campinas, 1994. 47 CAPÍTULO II CARACTERIZAÇÃO DA POLIFENOLOXIDASE EM DOIS CULTIVARES DE CACAU (Theobroma cacao L.) PRODUZIDOS NA REGIÃO SUL DA BAHIA, BRASIL. RESUMO Neste estudo a Polifenoloxidase foi caracterizada na polpa e semente de duas variedades de cacau, PH 16 e TSH 1188, com o objetivo de mensurar sua atividade enzimática quanto a concentração de substrato, pH e temperatura ótimos de reação correlacionando-a com as condições do processo fermentativo. A atividade foi determinada por espectrofotometria através da medida do aumento da absorbãncia a 420 nm. Os resultados demonstram a diferença e especificidade existente entre os cultivares de cacau, e entre polpa e semente em cada cultivar. Sugere-se que sejam utilizados critérios específicos para cada cultivar, tomando como base os parâmetros ideais para prolongar o período de melhor atuação da enzima durante o processo fermentativo, contribuindo assim para a melhoria na qualidade das amêndoas de cacau e por consequência maior qualidade na produção de chocolate monovarietal. Palavras-chave: cacau, parâmetros, atividade enzimática, chocolate. ABSTRACT The present study was designed to characterize the activity of polyphenol oxidase (PPO) in the pulp and seeds of two cocoa varieties, PH 16 and TSH 1188. The PPO activity was determined as a function of optimal substrate concentration, pH and temperature and was correlated with the conditions during the fermentation process. PPO activity was determined spectrophotometrically by measuring the increase in absorbance at 420 nm. The results showed the specificity and differences between the two cocoa cultivars and between the pulp and seeds of each cultivar. It is suggested that specific criteria are adopted for each cultivar, based on the optimal PPO parameters, to prolong the period of maximum PPO activity during fermentation, contributing to 48 the improvement of the quality of cocoa nuts and consequently to higher quality in the production of monovarietal chocolate. Keywords: cocoa, parameters, enzyme activity, chocolate. 49 1. INTRODUÇÃO Atualmente sabe-se que as reações que levam à formação de precursores de sabor do chocolate são levadas a cabo por enzimas endógenas presentes na semente do cacau e que o processo de fermentação tem como principal consequência o abaixamento do pH, favorecendo a ação destas enzimas. Durante a fermentação, a polpa envoltória das sementes do cacau é degradada pela ação sucessiva de microrganismos (leveduras e bactérias ácido-lácticas e ácidoacéticas) naturais do ambiente, com elevação da temperatura para cerca de 50ºC (CRUZ et al., 2013). A atividade enzimática em amêndoas de cacau durante a fermentação, é conhecida e estudada pelo menos desde a segunda metade do século XX. De acordo com Hansen et al., (1998), atualmente acredita-se que as seguintes enzimas tenham importância capital na formação do flavour do chocolate: a Polifenoloxidase, a Invertase e a Protease. A Polifenoloxidase (PPO) é amplamente distribuída na natureza, e é responsável por catalisar reações de oxidação de compostos fenólicos, na presença de oxigênio, cujos produtos se polimerizam, formando compostos de cor escura (ROBINSON et al., 1991). Sua presença e atividade durante a fermentação e a secagem das amêndoas de cacau, são fatores responsáveis pelo desenvolvimento dos precursores do sabor, começando na fase oxidativa da fermentação e continuando na secagem (LIMA et al., 2001). A importância da atividade desta enzima é também mencionada de forma implícita na redução do sabor amargo e adstringente do cacau (REEVES et al., 1988). A confiabilidade e comparação das atividades das enzimas do cacau são complicadas, devido a variações causadas por diferentes genótipos, a origem geográfica, métodos de fermentação utilizados e tipos de cochos empregados. Embora o papel essencial de enzimas endógenas durante a fermentação tenha sido evidenciado há muitos anos, existe ainda a falta de estudos sistemáticos abordando a comparação entre diferentes cultivares de cacau (HANSEN et al., 1998). Além disso, ainda não está elucidado como os processos enzimáticos são regulados durante a fermentação, quais substratos enzimáticos/produtos estão relacionados com o sabor de amêndoas com qualidade superior e quais os fatores limitantes para esses processos (disponibilidade de substrato e/ou enzima, genótipo, condições de cultivo ou processo de fermentação). 50 Neste contexto, este estudo tem por objetivo caracterizar a atividade enzimática da PPO em dois cultivres de cacau produzidos na região Sul da Bahia e relacioná-la com as condições do processo fermentativo, fornecendo subsídios para futuras intervenções tecnológicas que possam contribuir para melhoria da qualidade da matéria-prima na produção de chocolates monovarietais. 51 2. MATERIAL E MÉTODOS 2.1 Materiais Foram estudados dois cultivares de cacau, PH 16 e TSH-1188, produzidos na Fazenda Lajedo do Ouro (S 14° 06' 15.2" WO 39° 38' 45.8"), na cidade de Ibirataia, região sul do estado da Bahia, Brasil. O material em estudo foi definido pelos produtores, segundo sua disponibilidade no período de realização do experimento. O cultivar PH 16 (Híbrido Forastero - resultado do cruzamento do Forastero do Alto Amazônico com o Trinitario) foi identificado em 1996 em uma população de cacaueiros híbridos da Fazenda Porto Híbrido, no município de São José da Vitória – BA. Já o cultivar TSH -1188 (Trinidad Selected Hybrids - híbrido trinitario), é originario de Trinidad and Tobago, localizada próximo a costa oriental da Venezuela e apresenta resistência à vassoura-de-bruxa e excelente produtividade. 2.2 Reagentes e padrões Todos os reagentes e padrões utilizados neste estudo foram adiquiridos na Sigma Aldrich Co. 2.3 Fermentação A fermentação foi realizada em cochos de madeira (70 x 70 x 75 cm), com capacidade para 400 kg de massa de cacau, apresentando cerda de 20 furos (1,27 cm cada um) na parte inferior e nas laterais a fim de permitir o escoamento do mel do cacau produzido pela polpa durante a feremntação. Este processo teve duração de sete dias, sendo que a cada 48 horas a massa de cacau era revolvida tendo por finalidade oxigenar a massa e uniformizar a temperatura. Para cobrir a massa de cacau, foram utilizadas folhas de bananeira. 2.4 Coleta das amostras As amostras foram coletadas (aproximadamente 500 g) antes do início da fermentação 52 (tempo zero) e a cada 12 horas até o final do processo. Foram congeladas a -18º C para interrupção enzimática. Durante a fermentação, foram realizadas medidas de temperatura (Digital Thermometer MINIPA, modelo MT – 450) e pH (Medidor de pH portátil PHtek Digital) da massa (AOAC, 2000). 2.5 Caracterização da PPO A atividade da enzima foi determinada na polpa e semente dos dois cultivares de cacau antes do início da fermentação (tempo zero), tendo como parâmetros a temperatura, o pH e o substrato preferencial da enzima. Os resultados obtidos para temperatura e pH da enzima foram confrontados com aqueles obtidos durante o processo de fermentação, visando indentificar a etapa do processo onde as condições são mais favoráveis para a ação da PPO. 2.5.1 Extração da PPO da polpa A extração foi realizada segundo o descrito por Lima et al., (2001), utilizando-se 100 g de cacau. As polpas foram separadas das sementes manualmente e imersas em solução tampão fosfato de potássio 0,02 M (pH 7,5), contendo 5 % de polietilenoglicol e 5 mM de ácido ascórbico, o material foi então triturado com o auxílio de um triturador, na mesma solução tampão, na proporção 1:2 (p/v) e homogeneizado durante 30 minutos a 4 °C. O homogeneizado foi centrifugado a 11.000.g durante 15 minutos a 0 °C em Centrífuga Refrigerada HITACHI, modelo CR22GIII. O sobrenadante (extrato) foi então estocado a -18 ºC para realização das análises. 2.5.2 Extração da PPO das sementes A extração também foi realizada segundo o descrito por Lima et al., (2001). As sementes foram liofilizadas (Liofilizador Liotop, modelo L108) e posteriormente, desengorduradas, utilizando-se éter de petróleo como solvente (YUSEP et al., 2002). As sementes secas e desengorduradas foram suspensas em solução tampão fosfato de potássio 0,02 M (pH 7,5), contendo 5 % de polietilenoglicol e 5 mM de ácido ascórbico, na proporção 1:10 (p/v) e 53 homogeneizadas em agitador magnético durante 30 minutos a 4 °C. A suspensão foi então centrifugada a 11.000.g durante 15 minutos a 0 °C. O sobrenadante (extrato) foi estocado a -18 ºC para realização das análises. 2.5.3 Purificação da PPO dos extratos A purificação foi realizada segundo o descrito por Erzengin (2009), onde foi adicionado aos extratos sulfato de amônio [(NH4)2SO4], em quantidade suficiente para fornecer 80 % de saturação. O sal foi adicionado lentamente com agitação branda a 4 ºC e a mistura centrifugada a 20000.g por 60 minutos a 4 °C, reservando-se o precipitado. O precipitado foi suspenso em solução tampão fosfato potássio 0,02 M (pH 6,5) e dialisado contra o mesmo tampão em membrana de acetato por 24 horas a 4 °C. Após a diálise, a fração proteica foi precipitada com acetona na proporção 2:1 (v/v), e sua separação realizada por centrifugação a 11.000.g durante 15 minutos a 0 °C. O precipitado foi novamente suspenso em tampão fosfato potássio 0,02 M (pH 6,5) e armazenado a -18 °C para realização das demais análises (PERONE et al., 2007). 2.5.4 Determinação da atividade da PPO dos extratos A atividade enzimática foi determinada por espectrofotometria (Biochrom, Modelo Libra S50), através da medida do aumento na absorbância a 420 nm para o substrato catecol, nas concentrações de 0,05 M a 0,4 M. A mistura da reação foi constituída de 2,5ml de solução tampão fosfato de sódio 0,2 M (pH 7,0), 0,3 ml de substrato em diferentes concentrações e 0,2 ml do extrato contendo a enzima, atingindo um volume final de 3,0 ml. A amostra em branco teve apenas 0,3 ml da solução de substrato e 2,7 ml de solução tampão fosfato de sódio 0,2 M (pH 7,0). A temperatura foi mantida constante a 25°C. A porção linear da curva de atividade foi utilizada para expressar a atividade da enzima. Uma unidade de atividade da enzima (UE) foi definida como a quantidade de enzima que provoca um aumento na absorbância de 0,001.ml-1.min-1 (ERZENGIN, 2009). 54 2.5.5 Determinação dos parâmetros cinéticos Km e Vmax da PPO Os valores de Km e Vmax foram determinados para diferentes concentrações de catecol (0,05 M a 0,4 M). Os dados foram representados graficamente como 1/V0 e 1/[S] de acordo com o método de LINEWEVER-BURK (1934). 2.5.6 Efeito do pH e temperatura na atividade da PPO Para as determinações do pH e temperaturas ótimas da reação, foi utilizada a mesma metodologia descrita para a medida da atividade enzimática, sendo que para o pH foram testados os pHs 2, 3, 4, 5, 6, 7 e 8 e para a temperatura foram testadas as temperaturas de 20, 25, 30, 40, 50, e 60 ºC. 2.5.7 Determinação do substrato preferencial da enzima A atividade enzimática foi determinada utilizando-se a (+)-catequina e a (–)-epicatequina como substrato, visto que estes são os principais compostos fenólicos presentes no cacau (EFRAIM et al., 2011). A mesma metodologia descrita para o substrato catecol foi utilizada, assim como os valores de pH e temperatura ótimos já fixados. 2.5.8 Correlação entre os parêmetros de fermentação e a atividade enzimática dos extrato. Os resultados da atividade enzimátia da PPO nos extratos da polpa e semente dos cultivares no tempo zero, foram confrontados com os parâmetros de fermentação (pH e temperatura), com o objetivo de avaliar o comportamento da enzima no decorrer do processo. 2.6 Determinação do teor de proteína nos extratos. Para o cálculo da atividade enzimática específica, o teor de proteínas foi determinado pelo método de Lowry et al. (1951). 55 2.7 Análise Estatística. Todas as análises foram realizadas em sextuplicata e o desvio padrão dos dados foi determinado. 56 3. RESULTADOS E DISCUSSÃO 3.1 Determinação da atividade da PPO dos extratos Na Figura 01-A, pode-se observar uma grande diferença na atividade enzimática das polpas dos dois cultivares. A atividade da PPO no cultivar TSH-1188 é aproximadamente 6 vezes maior que a do PH 16, mostrando uma maior presença da enzima nesse cultivar. Essa maior atividade enzimática no cultivar TSH-1188, muito provavelmente terá reflexo na concentração e no comportamento dos compostos fenólicos durante e após a fermentação, pois segundo Misnawi et al. (2002), a PPO atua intensamente sobre esses compostos durante a fase aeróbica da fermentação, convertendo-os em quinonas, o que pode acarretar alterações nas características sensoriais como cor e sabor (MISNAWI et al., 2003). Já na semente, onde a atividade enzimática é menor que na polpa (Figura 01-B), a PPO atua de forma semelhante nos dois cultivares, pois as enzimas possuem atividades muito próximas. Contudo, existe uma diferença maior entre as atividades da polpa e semente no cultivar TSH-1188 do que no cultivar PH 16. Figura 01. Atividade da PPO para o substrato catecol em diferentes concentrações (0,05 M a 0,4 M ). (A) Polpa; (B) Semente. A. B. 80 TSH 1188 PH 16 3,5 60 -1 Atividade (UE. mg proteína ) -1 Atividade (UE. mg proteína ) 70 4,0 TSH 1188 PH 16 50 40 30 20 10 0 0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 0,40 0,45 -1 [catecol] (mg.ml ) 3,0 2,5 2,0 1,5 1,0 0,5 0,0 0,00 0,05 0,10 0,15 0,20 0,25 0,30 -1 [catecol] (mg.ml ) 0,35 0,40 0,45 57 A diferença de atividade observada entre os cultivares, pode ser evidenciado na Figura 02 através dos parâmetros cinéticos. A PPO possui boa afinidade com o catecol, uma vez que quanto menor o valor de Km maior será a afinidade da enzima com o substrato. Na polpa (Figura 02-A) os valores de Km e Vmax encontrados no cultivar PH 16 foram 0,107 mg.ml-1 e 10,6 mg.ml min-1 respectivamente, já para o cultivar TSH 1188 o Km foi de 0,089 mg.ml-1 e Vmax de 81,3 mg.ml min-1. Na semente (Figura 02-B) foi encontrado Km 0,047 mg.ml-1 e Vmax 2,3 mg.ml min-1 para PH 16 e para o TSH 1188 foi encontrado Km de 0,02 mg.ml-1 e Vmax de 2,3 mg.ml min-1. Figura 02. Parâmetros cinéticos pelo método de LINEWEVER-BURK com diferentes concentrações do substrato catecol para a enzima PPO nos dois cultivares de cacau. (A) Polpa; (B) Semente. A. B. 0,4 0,9 y = 0,0101x + 0,0942 R² = 0,8981 0,3 0,25 TSH 1188 0,2 PH 16 1/ V0 (mg.ml min -¹) 1/ V0 (mg.ml min -¹) 0,35 y = 0,0207x + 0,4392 R² = 0,9359 0,75 0,6 0,45 0,15 y = 0,0087x + 0,4256 R² = 0,9991 0,3 0,1 TSH 1188 PH 16 y = 0,0011x + 0,0123 R² = 0,9771 0,15 0,05 0 0 0 5 10 15 20 1/[ S ] (mg.ml -¹) 25 0 5 10 15 20 25 1/[ S ] (mg.ml -¹) A afinidade da PPO pelo tipo e concentração de substrato, é dependente não apenas da origem da enzima (tipo de planta), como foi observado por Oktay et al. (1995), como também da espécie e do cultivar. 3.2 Efeito do pH na atividade da PPO Apesar de apresentar uma maior atividade em maiores concentrações de catecol 58 (Figura 01), foi escolhida a concentração de 0,1 M, tanto para polpa quanto para semente, visto que o comportamento nessa concentração foi semelhante entre os cultivares e também pelo baixo desvio encontrado na análise. Os valores de pH ótimos para a atuação da PPO na polpa e semente de cacau foram determinados no intervalo de pH 2 a 8 sob temperatura constante a 25ºC, com concentração já fixada do catecol para reação de 0,1 M. Os valores de pH ótimos para o cultivar PH 16 (Figura 03- A) na polpa foi de 6,5 e semente de 5,8, apresentando atividade enzimática de 7,1 UE. mg proteína-1 e 6,5 UE. mg proteína-1 respectivamente. No cultivar TSH 1188 (Figura 03-B), o pH ideal para polpa foi de 6,6 e para semente 6,0. Como já evidenciado anteriormente, na polpa a atividade da enzima foi superior que a atividade na semente, apresentando valores de 43,1 UE. mg proteína-1 e 4,3 UE. mg proteína-1 respectivamente. Isso pode ser explicado por que o pH ótimo para máxima atividade da PPO nas plantas varia dependendo do método de extração, natureza dos substratos utilizados para o ensaio, e a localização da enzima na célula da planta (Dogan et al., 2005). Já no cultivar PH 16, a atividade na polpa e semente apresentam comportamento semelhantes. Figura 03. Efeito do pH na atividade da PPO. (A) Cultivar PH 16; (B) Cultivar TSH 1188. A. B. 50 -1 8 Atividade (UE. mg proteína ) Polpa Semente -1 Atividade (UE. mg proteína ) 10 6 4 2 0 2 3 4 5 pH 6 7 8 40 Polpa Semente 30 20 10 0 0 1 2 3 4 5 6 7 8 pH De acordo com os resultados apresentados pode-se concluir que, para os cultivares estudados, em valores baixos de pH, a PPO apresenta baixa atividade e que em pH alcalino há 59 uma diminuição brusca da atividade, chegando a representar entre 70 a 90 % de perda no potencial de atividade. Este fato também foi observado em estudos realizados por Toralles et al. (2005) em pêssegos, onde a PPO apresentou o pH ótimo variando numa faixa de 6,0 a 7,5, dependendo do substrato e do tecido vegetal. Abaixo desse pH a enzima foi menos ativa e acima desse pH ocorreu a auto oxidação do substrato. 3.3 Efeito da temperatura na atividade da PPO A partir da concentração de catecol (0,1 M) e de pH (polpa e semente, 6,6 e 6,0 para o TSH-1188 e 6,5 e 5,8 para o PH 16,respectivamente) já fixados, a temperatura ótima de atuação da PPO foi determinada numa faixa de temperatura de 20 a 60°C. Para o PH 16 (Figura 4-A), a PPO atingiu máxima atividade a 30ºC para a semente e 27ºC para a polpa, apresentando atividade de 9,7 UE. mg proteína-1 e 7,1 UE. mg proteína-1 respectivamente. Figura 04. Efeito da temperatura na atividade da PPO. (A) Cultivar PH 16; (B) Cultivar TSH 1188. A. B. 50 12 Polpa Semente Polpa Semente Atividade (UE. mg proteína ) 40 -1 -1 Atividade (UE. mg proteína ) 10 8 6 4 2 0 30 20 10 0 20 30 40 Temperatura (°C) 50 60 20 30 40 50 60 Temperatura (°C) No cultivar TSH-1188, a temperatura ideal foi de 25ºC, apresentando atividade de 4,3 UE. mg proteína-1 para semente e 40,3 UE. mg proteína-1 para polpa, representados na Figura 4-B. A 60 queda do potencial da atividade (aproximadamente de 90% para 10 %) da PPO em altas temperaturas é, provavelmente, devido ao desdobramento da estrutura terciária e desnaturação da enzima. Este fato também foi observado por Todaro et al. (2011), que caracterizou a PPO em berinjela, e encontrou uma perda significativa na atividade em temperaturas elevadas. Pode-se observar que nos dois cultivares, tanto na polpa quanto na semente, a atividade diminui gradativamente com o aumento da temperatura, o que sugere que altas temperaturas não favorecem a atuação da PPO, e implica em afirmar que a zona de maior atuação da enzima é entre 25 e 30°C. 3.4 Determinação do substrato preferencial da PPO Na figura Figura 05, pode ser observado que tanto para polpa como para semente nos dois cultivares, há maior afinidade da enzima pela catequina, apresentando valores de atividade na polpa de 11,93 UE. mg protein-1 e 20,3 UE. mg protein-1, e na semente 4,97 UE. mg protein-1 e 11,69 UE. mg protein-1, para PH 16 e TSH-1188, respectivamente. Diante destes resultados, observa-se que a enzima não possui maior afinidade com o composto presente em maior concentração na semente do cacau, pois segundo Kwik-Uribe (2005), em sementes de cacau não fermentadas in natura, foi observado que a quantidade de (–)-epicatequina é vinte vezes maior que a de (+)-catequina. Por outro lado, durante o processo de fermentação, o teor de polifenóis totais diminui cerca de 70%, sendo que a (–)-epicatequina sofre redução de 90% de sua concentração inicial (EFRAIM, 2011). Payne et al. (2010), relatou que a redução da epicatequina, pode ser explicada devido à oxidação biológica e ou a polimerização de monómeros, além da epimerização da (-)epicatequina a (-)-catequina, devido a elevação da temperatura durante a fermentação. Durante as etapas de processamento do cacau para a produção de chocolate, Cross (1999) observou que os flavanóis sofrem uma série de reações químicas, como oxidação, complexação e lixiviação, que, associadas às demais reações em curso – como a hidrólise de proteínas a aminoácidos, e de açúcares – levam à formação de compostos aromáticos, contribuindo significativamente para a formação do sabor desejável e a redução do amargor e da adstringência. 61 Figura 05. Determinação da atividade PPO para substrato preferencial: Catequina e Epicatequina. (A) Polpa; (B) Semente. A. B. 12 -1 Atividade (UE. mg proteína ) Epicatequina Catequina -1 Atividade (UE. mg proteína ) 20 15 10 5 0 10 8 6 4 2 0 PH 16 TSH 1188 Cultivar Epicatequina Catequina PH 16 TSH 1188 Cultivar Sendo assim, é possível afirmar que a redução drástica sofrida pela (-) epicatequina durante o processamento do cacau não pode ser atribuída com exclusividade a ação da PPO, mas a outros mecanismos mais importantes descritos pelos autores já citados. 3.5 Correlação entre os parêmetros de fermentação e a atividade enzimática dos estratos Nas Figuras 06 e 07, estão representados os parâmetros de fermentação, pH e temperatura, respectivamente e o pH e temperatura ótimos para a atuação da enzima PPO nos dois cultivares de cacau estudados. Para a polpa do cultivar PH 16 (Figura 06-A), a maior atividade da PPO ocorre em pH 6,5, contudo todo o processo de fermentação dessa variedade ocorre em pH na faixa de 4,5, somente após 5 dias (120 horas) de fermentação o pH atingiu um valor próximo de 6,0 e novamente voltou a valores próximos a 4,5. Portanto, na fermentação da polpa do cultivar PH 16, temos apenas 70% da atividade máxima da enzima durante todo o processo. Já para a semente (Figura 06-B), a maior atividade (6,5 UE. mg protein-1) é obtida em pH 5,8. Durante a fermentação, o pH da semente no início do processo está em torno de 6,5 e vai reduzindo lentamente até valores próximos a 4,5. Neste caso, a ação da enzima é favorecida nessa fase 62 inicial do processo, nos primeiros 3 dias (72 horas), após esse período a atividade vai reduzindo até permanecer próxima a 70% de sua atividade máxima. Sendo assim, embora a atividade da PPO seja menor na semente do que na polpa, a ação da enzima na semente é favorecida durante o processo de fermentação, quando considerado o pH do meio. Figura 06. Parâmetros de fermentação e pH ótimo de atuação da PPO. (A) PH 16 - Polpa; (B) PH 16 Semente; (C) TSH - Polpa; (D) TSH 1188 – Semente. B. 10 80 60 4 40 2 20 2 3 4 5 6 7 8 9 pH Polpa Tempo de Fermentação 80 60 20 40 10 20 4 5 pH 60 4 40 2 20 2 3 4 5 6 7 8 9 0 6 7 8 0 160 140 Semente Tempo de Fermentação 5 -1 Atividade (UE. mg proteína ) 100 30 3 80 6 Tempo de Fermentação (horas) 120 2 6 pH 140 40 0 100 D. 160 50 120 8 0 0 140 Semente Tempo de Fermentação 120 4 100 3 80 60 2 40 1 0 20 2 3 4 5 6 7 Tempo de Fermentação (horas) 6 160 -1 120 100 C. -1 140 8 0 Atividade (UE. mg proteína ) 12 Atividade (UE. mg proteína ) Polpa Tempo de Fermentação -1 Atividade (UE. mg proteína ) 10 160 Tempo de Fermentação (horas) 12 Tempo de Fermentação (horas) A. 0 pH No cultivar TSH-1188, a PPO presente na polpa (Figura 06-C) tem maior atividade (43,1 UE. mg proteína-1) em pH 6,6. Como o valor médio de pH da fermentação desse cultivar também 63 está em torno de 4,5, conclui-se que durante a fermentação desse cultivar, a enzima atinge somente 45% da sua atividade máxima, 18 UE. mg proteína-1. Já na semente (Figura 06-D), a fermentação teve início em pH 6,6 e foi reduzindo ao longo do processo até a estabilização em pH em torno de 4,8. Portanto, o pH da fermentação permaneceu na zona de maior atividade da enzima nas primeiras 72 horas do processo, favorecendo a ação da PPO nesse período. Ainda para este cultivar, após 72 horas, o pH da fermentação permaneceu numa faixa onde a enzima possui aproximadamente 70% de sua atividade máxima. Com isso, pode-se afirmar, que a ação da PPO na semente é favorecida também pelo pH no estágio inicial da fermentação (primeiros 3 dias) e que a diferença nas atividades da polpa e semente são compensadas durante o processo. Deve-se ressaltar que o pH encontrado na polpa no início do processo de fermentação, em torno de 4,5, para as duas cultivares está de acordo com os dados encontrados na literatura para amêndoas fermentadas na mesma região da Bahia (CRUZ et al., 2013), Contudo, segundo Amin et al. (2002), para obtenção dos melhores resultados na qualidade das amêndoas, o pH da fermentação deve estar na faixa de 5,0 a 5,5. Melhores resultados podem ser obtidos com os cultivares estudados, se medidas forem adotadas, visando minimizar a fermentação precoce que ocorre devido à quebra dos frutos no campo e o transporte da massa para os cochos, já em estado de fermentação. Com relação à temperatura, para o cultivar PH 16 (Figura 07- A), a polpa apresentou maior atividade a 30°C e a semente em 27ºC. De acordo com o gráfico, a temperatura somente irá favorecer a ação da enzima na polpa nas primeiras 48 horas de fermentação. Após esse período, a medida que a temperatura da fermentação aumenta, a atividade da enzima na polpa cai inicialmente 50% até que aos 45ºC a atividade da enzima é reduzida a somente 10% da atividade máxima. Para a semente, a ação da enzima também será favorecida nas primeiras 48 horas de fermentação, contudo a redução da atividade com o aumento da temperatura será menor, aproximadamente 40%. De forma análoga ao ocorrido com o pH, embora a atividade da enzima na polpa seja maior que na semente, sua resistência a altas temperaturas é menor, apresentando uma redução drástica após 48 horas, enquanto a enzima da semente permanece com maior atividade durante todo o processo. No cultivar TSH-1188 (Figura 07-B), a máxima atividade da enzima ocorre a 25° C, tanto para polpa quanto para semente. Com isso, assim como para a cultivar PH 16 durante a fermentação, a ação da PPO é maior apenas nos 2 primeiros dias para a polpa, havendo uma 64 redução da atividade de aproximadamente 60% quando a temperatura atinge os 35ºC. Para a semente a redução é mínima, visto que a enzima, embora apresente atividade bem menor que aquela presente na polpa, mantem-se no mesmo patamar durante todo o processo. Figura 07. Parâmetros de fermentação e temperatura ótima de atuação da PPO. (A) PH 16; (B) TSH1188. A. B. 10 120 8 100 80 6 60 4 40 2 0 20 20 30 40 Temperatura (°C) 50 60 0 50 140 120 40 100 30 80 60 20 40 10 0 20 20 30 40 50 60 Tempo de Fermentação (horas) 140 160 Polpa Semente Tempo de Fermentação -1 -1 160 Atividade (UE. mg proteína ) 12 Atividade (UE. mg proteína ) 60 Polpa Sememnte Tempo de Fermentação Tempo de Fermentação (horas) 14 0 Temperatura (°C) As correlações apresentadas evidenciam a diferença e especificidade existente entre as enzimas da polpa e semente dos cultivares, assim como entre os cultivares de cacau. Esta observação também foi relatada nos trabalhos conduzidos por Efraim (2010), que comprovaram a diferença do comportamento dos cultivares de cacau ao monitorar os parâmetros de temperatura e pH durante a fermentação, da mesma forma que foi encontrada diferenças nos liquors, manteiga e chocolate. Os resultados expostos no presente estudo sugerem, que no decorrer do processo de fermentação sejam utilizados critérios específicos para cada cultivar de cacau, tomando como base suas especificidades e parâmetros ideais, para que se possa prolongar e potencializar a atividade da enzima PPO, visto que a oxidação dos compostos fenólicos presentes na semente, realizada pela enzima, é de suma importância para características sensoriais do chocolate a ser produzido, como amargor e adstringência. Logo, prolongar a atuação da PPO em temperaturas mais baixas, através do aumento na frequência e quantidade dos revolvimentos da massa de 65 cacau, no início do processo de fermentação, pode retardar a elevação da temperatura e a queda do pH, o que irá favorecer a atuação da enzima nos cultivares em estudo. O controle do pH e temperatura irá proporcionar maior eficiência da enzima, prolongando seu período de melhor atuação durante o processo fermentativo, podendo contribuir para a melhoria nas características das amêndoas de cacau, e por consequência maior qualidade na produção do chocolate. 66 REFERÊNCIAS ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS. Official Methods of Analysis of AOAC International, 2000. AMIN, I. et al. Analysis of vicilin (7S)- class globulin in cocoa cotyledons from various genetic origins. Journal of the Science of Food and Agriculture, London, v. 82, p. 28-732, 2002. CROSS, E. Cocoa flavor development. The Manufacturing Confectioner, Princeton, v. 79, n. 2, p. 70-77, 1999. ROBINSON, D.S.; ESKIN, N.A.M. Oxidative Enzymes in Foods. Elsevier Applied Science, 1991. Cap 1, p.1-47; Cap 6, p.217-273 CRUZ, J, F, M; LEITE, P, B; SOARES, S, E; BISPO, E,S. Assessment of the fermentative process from different cocoa cultivars produced in Southern Bahia, Brazil. African Journal of Biotechnology. 2013. 12, 5218-5225. DOGAN, S. et al. Characterization and Purification of Polyphenol Oxidase from Artichoke (Cynara scolymus L.) Journal of Agricultural and Food Chemistry. 2005, 53, 776−785 EFRAIM, P. et al. Influence of cocoa beans fermentation and drying on the polyphenol content and sensory acceptance. Ciência e Tecnologia Alimentos. 2010, 30(1):142-50. EFRAIM, P; ALVES, A, B; JARDIM, D, C, P. Revisão: Polifenóis em cacau e derivados: teores, fatores de variação e efeitos na saúde. Braz. J. Food Technol. 2011, 14, p. 181-201. ERZENGIN, M. Affinity Purification and Characterization of Polyphenol Oxidase from Helianthus tuberosus L. Hacettepe Journal of Biology & Chemistry. 2009, 37, 313-25. TORALLES, R. P. et al. Controle da atividade da polifenoloxidase de pêssego por interação do pH, da temperatura e da concentração de ácido ascórbico. Brazilian Journal of Food Technology, 201 0Campinas, v. 13, n. 2, p. 120-127. HANSEN, C.E.; DEL OLMO, M.; BURRI, C. Enzyme activities in cocoa beans during fermentation. Journal of the Science of Food and Agriculture, 1998, .77, 273-81. OKTAY, M. et al. Polyphenoloxidase from Amasya apple. Journal of Food Science. 1995, 60, n.3, 494-505. KWIK-URIBE, C. Potential Health Beneits of Cocoa Flavanols. The Manufacturing Confectioner, Princeton. 2005, 85, n. 10, 43-49. LIMA, E, D, P, A. et al. Obtenção e utilização da enzima polifenoloxidase extraída de polpa de pinha (annona squamosa l.) madura no melhoramento do sabor do cacau (Theobroma cacao L) Revista Brasileira de Fruticultura. Jabotivabal – SP, 2001, v. 23, n. 3, 709-713. 67 LINEWEVER H.; BURK, D. The determination of enzyme dissociation constants. Journal of American Chemical Society. 1934, 56, 658-666. LOWRY, O. H. et al. Protein measurement with the Folin fenol reagent. Journal of Biological Chemistry. 1951, 193, 265- 275. MISNAWI; JINAP, S.; NAZAMID,S.; JAMILAH, B. Activation of remaining key enzymes in dried under-fermentedcocoa beans and its effect on aroma precursor formation. Food Chemistry, v. 78, 407-17, 2002. MISNAWI, JINAP, S., JAMILAH, B; NAZAMID, S. Sensory properties of cocoa liquor as affected by polyphenol concentration and duration of roasting. Journal of Food Quality and Preference, v.15, p. 403-409, 2003. PAYNE, M, J. et al. Impact of Fermentation, Drying, Roasting, and Dutch Processing on Epicatechin and Catechin Content of Cacao Beans and Cocoa Ingredients. Journal of Agricultural and Food Chemistry. 2010, 58, 10518 – 10527. PERONE, C A S. et al. Purificação parcial e caracterização cinética da enzima Polifenoloxidase de banana nanica (Musa acuminata). Revista do Instituto de Ciências da Saúde. 2007, 25, 239-46. REEVES, S.G. et al. Biochemical studies of Cocoa Bean o-diphenol O2 oxidoreductase (catechol oxidase). Food Chemistry, Kidlington, 1988, 290-219. YORUK, R; MARSHALL, M, R. Physicochemical properties and function of plant polyphenol oxidase: a review'. Journal of Food Biochemistry. 2003, 27, 361-422. TODARO, A. et al. Study and Characterization of Polyphenol Oxidase from Eggplant (Solanum melongena L.). Journal of Agricultural and Food Chemistry. Italia, 2011, 59, 11244–11248. YUSEP, I. et al. . Influence of carboxypeptidases on free aminoacid, peptide and methylpyrazine contents of under-fermented cocoa beans. Journal Science Food Agriculture. 2002, 82, 1584-92. 68 CAPÍTULO III EXTRAÇÃO E CARACTERIZAÇÃO DA INVERTASE EM DOIS CULTIVARES DE CACAU (Theobroma cacao L.) PRODUZIDOS NO SUL DA BAHIA, BRASIL. RESUMO A invertase foi caracterizada em dois cultivares de cacau, PH 16 e TSH 1188 na polpa e semente, objetivando identificar a atividade enzimática quanto a concentração de substrato, pH e temperatura ótimos de reação correlacionando-a com as condições do processo fermentativo. As atividades enzimáticas (invertase ácida e neutra) foram avaliadas pela dosagem de açúcares redutores segundo método de Somogyi-Nelson e determinadas por espectrofotometria. Os resultados evidenciam a diferença e especificidade da enzima e que a invertase ácida é cineticamente mais eficiente do que a neutra podendo ser a principal forma de enzima vacuolar envolvida na hidrólise da sacarose. O controle da temperatura e pH irá prolongar o período de melhor atuação durante o processo fermentativo, propiciando uma melhor qualidade das amêndoas de cacau e por consequência, maior qualidade na produção de chocolates monovarietais, no que diz respeito à formação e potencialização dos precursores de aroma e sabor. Palavras-chave: cacau, enzimas, chocolate monovarientais. ABSTRACT The invertase was characterized in two cultivars of cocoa, PH 16 and TSH 1188 the pulp and seeds in order to identify the enzymatic activity and substrate concentration , pH and temperature optima of reaction correlated with the conditions of the fermentation process . The enzymatic activities ( acid and neutral invertase ) were assessed by measurement of reducing sugars second method of Somogyi - Nelson and determined by spectrophotometry . The results show a 69 difference and specificity of the enzyme and that acid invertase is kinetically more efficient than the neutral may be the main form of vacuolar enzyme involved in the hydrolysis of sucrose . The control of temperature and pH will prolong the best performance during the fermentation process , providing a better quality of cocoa beans and therefore higher quality production monovarietais chocolates , with regard to training and enhancement of flavor precursors and flavor. Keywords: cocoa, enzymes, monovarietais chocolate. 70 1. INTRODUÇÃO O processo de fermentação é uma etapa muito importante no processamento de cacau. Estudos mostraram que quando o cacau não é submetido ao processo de fermentação, não irá desenvolver o sabor desejável ao chocolate durante o processo de torração, ficando excessivamente adstringente e amargo (MISNAWI et al., 2002). Durante este processo, ocorrem importantes reações bioquímicas que, por meio da hidrólise de proteínas e de açúcares presentes na polpa e semente do cacau em conjunto com a elevação da temperatura a valores próximos a 50°C, levam à formação dos precursores de sabor, em particular aminoácidos livres e açúcares redutores (PEZOA-GARCÍA, 1989). É sabido que enzimas endógenas desempenham um importante papel na produção dos precursores de aroma e sabor e na degradação de compostos durante a fermentação do cacau (STURM, 1999). A enzima invertase atua durante este processo na hidrólise da sacarose em glicose e frutose (açúcares redutores), indispensáveis à formação do sabor que serão potencializados durante a etapa de torração (HANSEN et al., 1998; LOPEZ; DIMICK, 1991). Esta enzima possui duas isoenzimas: uma ácida com atividade máxima em pH 5,5 e predominante em tecidos em crescimento e outra neutra, com um máximo de atividade em pH 7,0 e com predominância em tecidos completamente expandidos (HUSSAIN et al., 2008). Contudo, ainda é pouco o conhecimento sobre invertases neutras/ácidas devido a dificuldades de suas purificações e atividades enzimáticas baixas e instáveis (DU, et al., 2013; ROITSCH; GONZALEZ, 2004). A identificação e caracterização das atividades enzimáticas no cacau é sensível e trabalhosa, pricipalmente com relação a inúmeras variações que ocorrem durante as etapas de processamento do cacau (STURM, 1999). A quantificação e a comparação dessas atividades são complexas, considerando-se as variações causadas pelas diferenças dos genótipos, diferenças geográficas e dos métodos de fermentação (HANSEN et al., 1998). Além disso, ainda não é totalmente compreendido como esses processos enzimáticos são regulados, quais seus parâmetros limitantes e quais substratos/produtos estão relacionados com obtenção de amêndoas de maior qualidade. Diante do exposto, este estudo tem por objetivo caracterizar a atividade enzimática da invertase do cacau nos cultivres PH 16 e TSH 1188, produzidos na região Sul da Bahia, visando 71 fornecer subsídios para possíveis intervenções tecnológicas que possam contribuir para melhoria da qualidade da matéria-prima na produção de chocolates monovarietais. 72 2. MATERIAL E MÉTODOS 2.1 Materiais Foram estudados dois cultivares de cacau, PH 16 e TSH-1188, produzidos na Fazenda Lajedo do Ouro (S 14° 06' 15.2" WO 39° 38' 45.8"), na cidade de Ibirataia, região sul do estado da Bahia, Brasil. O material em estudo foi definido pelos produtores, segundo sua disponibilidade no período de realização do experimento. O cultivar PH 16 (Híbrido Forastero - resultado do cruzamento do Forastero do Alto Amazônico com o Trinitario) foi identificado em 1996 em uma população de cacaueiros híbridos da Fazenda Porto Híbrido, no município de São José da Vitória – BA. Já o cultivar TSH -1188 (Trinidad Selected Hybrids - híbrido trinitario), é originario de Trinidad and Tobago, localizada próximo a costa oriental da Venezuela e apresenta resistência à vassoura-de-bruxa e excelente produtividade. 2.2 Reagentes e padrões Todos os reagentes e padrões utilizados neste estudo foram adiquiridos na Sigma Aldrich Co. 2.3 Fermentação A fermentação foi realizada em cochos de madeira (70 x 70 x 75 cm), com capacidade para 400 kg de massa de cacau, apresentando cerda de 20 furos (1,27 cm cada um) na parte inferior e nas laterais a fim de permitir o escoamento do mel do cacau produzido pela polpa durante a feremntação. Este processo teve duração de sete dias, sendo que a cada 48 horas a massa de cacau era revolvida tendo por finalidade oxigenar a massa e uniformizar a temperatura. Para cobrir a massa de cacau, foram utilizadas folhas de bananeira. 2.4 Coleta das amostras As amostras foram coletadas (aproximadamente 500 g) antes do início da fermentação 73 (tempo zero) e a cada 12 horas até o final do processo. Foram congeladas a -18º C para interrupção enzimática. Durante a fermentação, foram realizadas medidas de temperatura (Digital Thermometer MINIPA, modelo MT – 450) e pH (Medidor de pH portátil PHtek Digital) da massa (AOAC, 2000). 2.5 Caracterização da Invertase A atividade da enzima foi determinada na polpa e semente dos dois cultivares de cacau antes do início da fermentação (tempo zero), tendo como parâmetros, a temperatura, o pH e o substrato preferencial da enzima. Os resultados obtidos para temperatura e pH da enzima foram confrontados com aqueles obtidos durante o processo de fermentação, visando indentificar a etapa do processo onde as condições são mais favoráveis para a ação da Invertase. 2.5.1 Extração da Invertase da polpa A extração foi descrita por Gomez et al., (1999) utilizando-se 100g de cacau. As polpas foram separadas das sementes manualmente e imersas em solução tampão fosfato de sódio 50mM pH 7,5 contendo NaCl 50mM, glicerol 5%, sulfato de manganês 5mM e β-mercaptoetanol 1mM, na proporção de 1:2 (p/v) e homogeneizadas durante 30 minutos a 4°C. O homogeneizado foi centrifugado em centrífuga refrigerada HITACHI, modelo CR22GIII a 20000.g durante 10 min a 0°C. O sobrenadante (extrato) foi estocado a -18ºC para realização das análises. 2.5.2 Extração da Invertase das sementes A extração também foi realizada segundo o descrito por Gomez et al., (1999). As sementes removidas da extração das polpas foram liofilizadas (Liofilizador Liotop, modelo L108) e posteriormente, desengorduradas, utilizando-se éter de petróleo como solvente (YUSEP et al., 2002). As sementes secas e desengorduradas foram suspensas em solução tampão assim como descrito para as polpas. A suspensão foi então centrifugada a 20000.g durante 10 min a 0°C. O sobrenadante foi então estocado a -18ºC para realização das análises. 74 2.5.3 Purificação da Invertase dos extratos A purificação foi realizada segundo o descrito por Deuner et al. (2005), onde foi adicionado aos extratos sulfato de amônio [(NH4)2SO4], em quantidade suficiente para fornecer 80 % de saturação. O sal foi adicionado lentamente com agitação branda a 4 ºC e a mistura centrifugada a 20000.g por 60 minutos a 4 °C, reservando-se o precipitado. Este foi suspenso em 4ml de água deionizada. A seguir, o extrato foi dialisado por 24 horas a 4°C contra tampão fosfato potássio 100 mM e pH 7,0 obtendo-se o extrato enzimático bruto. Após a diálise, o extrato foi precipitado com acetona na proporção 2:1 (v/v), e sua separação realizada por centrifugação a 11.000.g durante 15 minutos a 0 °C. O precipitado foi novamente suspenso em tampão fosfato de potássio pH 6,5 e armazenado a -18 °C para realização das demais análises. 2.5.4 Determinação da atividade da Invertase dos extratos A atividade enzimática foi determinada segundo Nascimento et al., (1998), por espectrofotometria (Biochrom, Modelo Libra S50), para o substrato sacarose, nas concentrações de 0,05 M a 0,4 M. A mistura da reação (4,0 ml) foi constituída de 0,5 ml do extrato enzimático, 1,0 ml de sacarose nas diferentes concentrações e 2,5 ml de solução tampão no pH de reação indicado para cada isoenzima, sendo tampão acetato de potássio 0,2 M, pH 4.5, para invertase ácida e tampão fosfato de potássio 0,2 M, pH 7.5, para invertase neutra. O meio de reação foi incubado em banho maria a 37°C durante 30 minutos. A reação foi interrompida pelo aquecimento a 100°C. As atividades enzimáticas foram avaliadas pela dosagem de açúcares redutores segundo protocolo para determinação de açúcares redutores pelo método de SomogyiNelson (NELSON, 1960). Uma unidade da enzima foi definida como sendo a quantidade de enzima capaz de liberar 1 mg de açúcar redutor por mg de proteína por hora (mg glicose.mg proteína-1.h-1). 2.5.5 Determinação dos parâmetros cinéticos Km e Vmax da Invertase Os valores de Km e Vmax foram determinados para diferentes concentrações de 75 sacarose (0,05 M a 0,4 M). Os dados foram representados graficamente como 1/V0 e 1/[S] de acordo com o método de LINEWEVER-BURK (1934). 2.5.6 Efeito do pH e Temperatura na atividade da Invertase Para as determinações do pH e temperatura ótimos de reação, foi utilizada a mesma metodologia descrita para a medida da atividade enzimática, sendo que para a invertase ácida somente foi testado pH 4,5 e para a invertase neutra os pHs 6,5, 7,5 e 8,5. Para a temperatura foram testadas as temperaturas de 20, 30, 40, 50, e 60ºC para as duas isoenzimas. 2.5.7 Correlação entre os parêmetros de fermentação e a atividade enzimática dos extratos Os resultados da atividade enzimátia da Invertase (ácida e neutra) nos extratos da polpa e semente dos cultivares no tempo zero, foram confrontados com os parâmetros de fermentação (pH e temperatura), com o objetivo de avaliar o comportamento da enzima no decorrer do processo. 2.6 Determinação do teor de proteína nos extratos Para o cálculo da atividade enzimática específica, o teor de proteínas foi determinado pelo método de Lowry et al. (1951). 2.7 Análise Estatística. Todos as análises foram realizadas em sextuplicata e o desvio padrão dos dados foi analisado. 76 3. RESULTADOS E DISCUSSÃO 3.1 Determinação da atividade da Invertase dos extratos A partir da Figura 01-A, pode ser observado que a enzima invertase apresenta atividades diferentes na polpa, nos cultivares PH 16 e TSH 1188, sendo maior para invertase ácida do que para invertase neutra. Já na semene, Figura 01-B, a invertase atua de forma semelhante nos dois cultivares, pois a enzima possui atividades muito próximas. Comparando-se polpa e semente nos dois cultivares, fica evidente que a invertase possui maior atividade na polpa do que na semente, uma vez que a polpa do cacau é rica em açúcares (10-15%) e é um excelente meio para o crescimento de microrganismos. Quando as sementes são removidas dos frutos, ocorre uma inoculação natural de microrganismos do ambiente para a polpa (BECKETT, 2009). Figura 01. Atividade das Invertases para o substrato sacarose em diferentes concentrações (0,05 M a 0,4 M ). (A) Polpa; (B) Semente. A. B. -1 -1 0,8 TSH 1188 - Invertase Ácida PH 16 - Invertase Ácida TSH 1188 - Invertase Neutra PH 16 Invertase Neutra Atividade (mg glicose.mg proteína .h ) -1 -1 Atividade (mg glicose.mg proteína .h ) 1,0 0,6 0,4 0,2 0,0 0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 0,40 0,45 -1 [sacarose] (mg.ml ) 0,14 0,12 TSH 1188 - Invertase Ácida PH 16 - Invertase Ácida TSH 1188 - Invertase Neutra PH 16 - Invertase Neutra 0,10 0,08 0,06 0,04 0,02 0,00 0,00 0,05 0,10 0,15 0,20 0,25 0,30 0,35 0,40 0,45 -1 [Sacarose] (mg ml ) A atuação da enzima invertase sobre a sacarose está relacionada com a hidrólise desse substrato em glicose e frutose (açúcares redutores) durante o processo de fermentação. Esses açúcares são importantes precursores de sabor do chocolate, e suas atuações serão potencializadas nas etapas posteriores do beneficiamento do cacau, sendo de vital importância na participação da 77 reação de Maillard, que é a principal responsável pela formação do sabor desejável do cacau. No estágio inicial, essa reação envolve a condensação de grupos carbonila de açúcares redutores com grupos amina provenientes de aminoácidos livres, seguida da degradação dos produtos originados, formando diversos compostos oxigenados (PEZOA-GARCÍA, 1989; ROSLI et al., 1996). Figura 02. Parâmetros cinéticos pelo método de LINEWEVER-BURK com diferentes concentrações de sacarose, para as invertases nos cultivares estudados. (A) PH16 - Polpa; (B) PH16- Semente; (C) TSH 1188 - Polpa; (D) TSG 1188 – Semente. A. B. 6 120 y = 0,0405x + 3,5508 R² = 0,9826 100 4 3 Invertase Ácida Invertase Neutra y = 0,0446x + 1,2328 R² = 0,9473 2 1/ V0 (mg.ml min -1) 1/ V0 (mg.ml min -1 5 1 y = 5,2762x - 4,8034 R² = 0,9981 80 60 Invertase Ácida Invertase Neutra 40 20 0 y = 4,579x + 2,7951 R² = 0,9221 0 0 5 10 15 20 25 0 5 1/[ S ] (mg.ml -1) D. 14 1/ V0 (mg.ml min -1) 12 10 Invertase Ácida Invertase Neutra 8 y = 0,1353x + 2,1119 R² = 0,9207 6 15 20 25 250 y = 0,2407x + 7,674 R² = 0,9926 4 1/ V0 (mg.ml min -1) C. 10 1/[ S ] (mg.ml -1) 200 y = 10,693x - 31,968 R² = 0,9038 Invertase Ácida 150 Invertase Neutra 100 50 y = 5,4286x - 3,5714 R² = 0,9849 2 0 0 0 5 10 15 1/[ S ] (mg.ml -1) 20 25 0 5 10 15 20 25 1/[ S ] (mg.ml -1) A boa afinidade da enzima com a sacarose pode ser evidenciada na Figura 02 através dos valores de Km e Vmax (Tabela 01) encontrados para as isoenzimas, invertase ácida e neutra, nos 78 cultivares estudados. Quanto menor o valor de Km maior será a afinidade da enzima com o substrato. Tabela 01. Km e Vmax das invertases para os cultivares com o substrato sacarose. Invertase Ácida PH 16 Vmax (mg.ml Invertase Neutra TSH 1188 PH 16 TSH 1188 Polpa Semente Polpa Semente Polpa Semente Polpa Semente 0,8 0,4 0,5 0,3 0,3 0,2 0,1 0,03 0,036 1,638 0,064 1,520 0,011 1,098 0,031 0,334 min-1) Km (mg.ml-1) Segundo DU at al., (2013), ainda não há dados conclusivos quanto a estabilidade da enzima invertase. As invertases normalmente apresentam valores de Km muito baixos com o substrato sacarose, o que é positivo, porém com velocidade de hidrólise lenta, assim como encontrado no presente estudo. 3.2 Efeito do pH na atividade da Invertase Os valores de pH ótimos para a atuação da invertase na polpa e semente de cacau foram determinados no intervalo de pH 6,5, 7,5 e 8,5 para invertase neutra e pH 4,5 para invertase ácida sob temperatura de incubação constante a 37ºC e interrupção da reação a 100°C, em concentrações de sacarose fixadas para os dois cultivares, sendo para invertase ácida, polpa e semente 0,2 M e 0,4 M, respectivamente e invertase neutra 0,3 M polpa e semente. A escolha destas concentrações é justificada pela maior atividade das invertases encontradas e baixo desvio padrão. Estão representados na Figura 03-A as atividades da invertase ácida com pH fixo 4,5. Para os cultivares PH 16 e TSH 1188, apresentam maior atividade na polpa (0,66 mg glicose.mg proteína-1.h-1 e 0,31 mg mg glicose.mg proteína-1.h-1 respectivamente) do que na semente (0,14 mg glicose.mg proteína-1.h-1 e 0,14 mg glicose.mg proteína-1.h-1 respectivamente). As invertases ácidas não foram testadas em outros valores de pH (2,5 e 3,5) pois os valores das atividades 79 poderiam não ser confiáveis, uma vez que a conversão da sacarose em glicose e frutose poderia ocorrer, provavelmente, pela acidificacação do meio e não pela ação da enzima. Figura 03. Efeito do pH na atividade das Invertases. (A) Ácida; (B) Neutra. A. B. 0,40 0,5 0,4 0,3 0,2 0,1 0,0 PH 16 -1 -1 Polpa Semente Atividade (mg glicose.mg proteína .h ) 0,6 -1 -1 Atividade (mg glicose.mg proteína .h ) 0,7 TSH 1188 - Polpa PH16 - Polpa TSH 1188 - Senmente PH 16 - Semente 0,35 0,30 0,25 0,20 0,15 0,10 0,05 0,00 TSH 1188 Cultivar 6,5 7,0 7,5 8,0 8,5 pH Na Figura 03-B, podemos observar que para invertase neutra, a polpa nos dois cultivares também apresenta maior atividade que na semente. Para o cultivar PH 16, a polpa apresenta pH ótimo de reação de 6,5 com atividade de 0,34 mg glicose.mg proteína-1.h-1 e para semente de pH 7,5 com atividade de 0,07 mg glicose.mg proteína-1.h-1. Já no cultivar TSH 1188, apresenta pH ótimo de 7,5 para polpa e semente com atividades de 0,12 mg glicose.mg proteína-1.h-1 e 0,08 mg glicose.mg proteína-1.h-1, respectivamente. Os valores ótimos encontrados, assemelham-se com o relatado por Darren et al., (1998), no qual encontrou valor ótimo de pH 7,2 para cana de açúcar, encontram-se dentro da faixa de pH ótimo para invertase neutra que varia de pH 6,8, a 8,0. 3.3 Efeito da temperatura na atividade da Invertase As temperaturas ótimas de atuação das invertases foram determinadas numa faixa de temperatura de 20 a 60°C, a partir da concentração de sacarose e pH já fixados. Na invertase ácida (Figura 04-A), para o cultivar PH 16, a enzima atingiu máxima atividade na polpa a 33ºC 80 (0,78 mg glicose.mg proteína-1.h-1) e para a semente apresentou 02 picos de maior atividade, 28°C (0,82 mg glicose.mg proteína-1.h-1) e 43°C (0,89 mg glicose.mg proteína-1.h-1). Para o cultivar TSH 1188, apresentou maior atividade a 31ºC (0,39 mg glicose.mg proteína-1.h-1) para polpa e 27° (0,73 mg glicose.mg proteína-1.h-1) e 43°C (0,70 mg glicose.mg proteína-1.h-1) para semente. Pode ser observado que a atividade é maior na semente do que na polpa. Figura 04. Efeito da temperatura na atividade da Invertase. (A) Ácida; (B) Neutra. A. B. 0,5 0,8 0,6 0,4 0,2 0,0 20 30 TSH 1188 - Polpa PH 16 - Pola TSH 1188 - Semente PH 16 - Semente -1 -1 1,0 Atividade (mg glicose.mg proteína .h ) TSH 1188 - Polpa PH 16 - Polpa TSH 1188 - Semente PH 16 Semente -1 -1 Atividade (mg glicose.mg proteína .h ) 1,2 40 50 60 Temperatura (°C) 0,4 0,3 0,2 0,1 0,0 20 30 40 50 60 Temperatura (°C) Para a invertase neutra (Figura 04-B), as temperaturas ótimas de atuação para o cultivar PH 16 foi de 52ºC com atividade de 0,43 mg glicose.mg proteína-1.h-1para a polpa e 50ºC e atividade de 0,17 mg glicose.mg proteína-1.h-1 para a semente. Já para o TSH 1188 a temperatura ideal foi de 50°C, apresentando atividades de 0,26 mg glicose.mg proteína-1.h-1 para polpa e 0,18 mg glicose.mg proteína-1.h-1para semente. Com isso pode-se concluir que a atuação desta enzima, em ambos os cultivares, é maior na polpa do que na semente, e que altas temperaturas favorecem sua atuação, o que implica afirmar que a zona de maior atividade para esta enzima é entre 50 e 52°C. Comparando-se a atuação das duas isoenzimas, é possível afirmar que a atividade da invertase ácida é maior do que na invertase neutra. 81 3.4 Correlação entre os parêmetros de fermentação e a atividade enzimática dos extratos Estão representados nas Figuras 05 e 06, os parâmetros de fermentação, pH e temperatura, respectivamente e as temperaturas ótimas para a atuação das invertases nos dois cultivares de cacau estudados. A ação da invertase ácida, com pH fixo estabelecido 4,5, é maior na polpa do que na semente para ambos os cultivares, uma vez que o pH da polpa durante a maior parte do processo de fermentação ocorre nessa faixa de pH (Figura 05-A e B). Sendo assim a atuação desta enzima é favorecida mais na polpa que na semente. Na semente a ação da enzima, ainda que baixa, será favorecida no PH 16 nos últimos dias de fermentação (após 120 horas e no TSH 1188 no quinto dia de fermentação (120 horas). Figura 05. Acompanhamento do pH para polpa e semente durante o processo de fermentação. (A) Cultivar PH 16; (B) Cultivar TSH 1188. A. B. 7,0 Polpa Semente 7,0 6,0 5,5 5,5 5,0 5,0 4,5 4,5 4,0 4,0 3,5 3,5 3,0 3,0 2,5 0 20 40 60 80 100 120 140 Tempo de Fermanteção (horas) 160 180 Polpa Semente 6,5 6,0 pH pH 6,5 2,5 0 20 40 60 80 100 120 140 160 180 Tempo de Fermentação (horas) A invertase neutra, apesar de apresentar valores ótimos de pH fixos para maior atividade da enzima (PH 16, polpa pH de 6,5 e semente pH 7,5; TSH 1188, polpa e semente pH 7,5), atua fora da faixa de pH durante todo o processo de fermentação como pode ser observado na Figura 05- A e B (valores de pH entre 4,1 a 5,78 na polpa e 4,5, a 6,84 na semente para o cultivar PH 16 e valores de pH entre 4,16 a 4,97 na polpa e 4,58 a 6,89 na semente, para TSH 1188). Isso 82 implica dizer que a invertase neutra não possui atuação nestas condições de fermentação para os cultivares estudados. As variações do pH, assim como encontrada na polpa e semente dos cultivares estudados durante o processo de fermentação, afeta a ionização de resíduos de aminoácidos essenciais no sítio ativo, o qual está envolvido na ligação do substrato e o catalizador. Alguns resíduos ionizáveis podem ser localizados na periferia do sítio ativo, vulgarmente conhecido como resíduos não essenciais. Esta ionização pode provocar distorções no sítio ativo e, portanto, afetar indiretamente a atividade da enzima (BHATTI et al., 2006). Com relação à temperatura ótima de atuação, a invertase ácida para o cultivar PH 16 (Figura 06- A), apresentou na polpa maior atividade a 33°C e na semente apresentou 02 picos para ótima atividade da enzima, o primeiro a 28°C e o segundo a 43ºC. A partir do gráfico, fica evidenciado que a temperatura somente irá favorecer a ação da enzima na polpa nas primeiras 36 horas de fermentação, no restante do processo apenas 32% da atividade máxima da enzima será atingida. Para a semente, a maior atividade da enzima será favorecida do primeiro ao terceiro dia de fermentação (20 a 72 horas aproximadamente). Para a invertase neutra (Figura 06–B), como já evidenciado anteriormente, na polpa a enzima apresenta maior atuação do que na semente. Apesar de sua maior atividade (52°C e atividade de 0,43 mg glicose.mg proteína-1.h-1) encontrar-se fora da faixa de temperatura alcançada durante a fermentação (temperatura máxima de 51°C), mas ainda assim a atividade é maior no final da fermentação. Na semente, a atividade da enzima é maior a 50°C (0,17 mg glicose.mg proteína-1.h-1), e sua maior atuação ocorre no sexto dia de fermentação. No cultivar TSH-1188 (Figura 06-C), a máxima atividade da enzima ocorre a 31°C para a polpa em relação à invertase ácida. A ação desta enzima é maior apenas nos 2 primeiros dias, havendo uma redução da atividade de aproximadamente 44% quando a temperatura atinge os 43°C. Para a semente a máxima atividade é alcançando a 27ºC (0,73 mg glicose.mg proteína-1. h-1) e a 43°C (0,70 mg glicose.mg proteína-1.h-1), assim após 48 horas do início da fermentação a atividade da enzima é reduzida a 33% e novamente volta a atingir sua máxima atividade em 72 horas, permanecendo por 12 horas e então é outra vez reduzida a 64% da atividade máxima. Para a invertase neutra (Figura 06-D), tanto polpa quanto semente apresenta comportamento bastante semelhante, alcançando maior atividade da enzima a 50ºC. 83 Como pode ser observada a partir do gráfico, essa maior atividade é alcançada a partir do quarto dia do processo de fermentação. Figura 06. Parâmetros de fermentação e temperatura ótima de atuação da Invertase. (A) PH 16 – Invertase Ácida; (B) PH 16 – Invertase Neutra; (C) TSH - Invertase Ácida; (D) TSH 1188 – Invertase Neutra. B. 120 0,8 100 0,6 80 60 0,4 40 0,2 0,0 20 20 30 40 50 60 0 (Atividade (mg glicose.mg proteína-1.h-1) 140 Tempo de Fermentação (horas) 1,0 0,5 160 Polpa Semente Tempo de Fermentação -1 0,4 100 0,3 80 0,2 60 40 0,1 20 0,0 20 30 40 50 60 0 Temperatura (°C) C. D. Polpa Semente Tempo de Fermentação 0,75 120 100 0,60 80 0,45 60 0,30 40 0,15 0,00 20 20 30 40 Temperatura (°C) 50 60 0 -1 -1 140 Atividade (mg glicose.mg proteína .h ) -1 0,90 0,30 160 Tempo de Fermentação (horas) 1,05 -1 140 120 Temperatura (°C) Atividade (mg glicose.mg proteína .h ) 160 Polpa Semente Tempo de Fermentação 160 Polpa Semente Tempo de Fermentação 0,25 140 120 0,20 100 0,15 80 60 0,10 40 0,05 0,00 20 20 30 40 50 60 Tempo de Fermentação (horas) -1 Atividade (mg glicose.mg proteína .h ) 1,2 Tempo de Fermentação (horas) A. 0 Temperatura (°C) A partir das correlações apresentadas, fica evidente que em ambos os cultivares, a semente apresenta maior atividade do que a polpa em relação a invertase ácida e para invertase neutra ocorre o inverso, a polpa apresenta maior atividade que na semente. Comparando-se as isoenzimas, pode-se afirmar que a atividade da invertase ácida é maior que a atividade da 84 invertase neutra tanto na polpa como na semente (4 a 5 vezes maior) durante o processo de fermentação acompanhado nos dois cultivares. Este fato indica que esta enzima é cineticamente mais eficiente e pode ser a principal forma de enzima vacuolar envolvida na hidrólise da sacarose (SACHDEVA et al., 2003). Os resultados expostos no presente estudo comprovam a diferença e especificidade existentes entre os cultivares de cacau, e entre polpa e semente em cada cultivar. Sugere-se que no decorrer do processo de fermentação, sejam utilizados critérios específicos para cada cultivar de cacau, tomando como base suas especificidades e parâmetros ideais para prolongar e potencializar a atividade da invertase, uma vez que sua estabilidade e atividade são fatores limitantes para a conversão total da sacarose e formação de precursores de sabor durante a fermentação (HANSEN et al., 1998). As enzimas exibem diferentes estabilidades durante o processo de fermentação, principalmente no que se refere à inativação. Esta é causada pelo calor gerado durante o processo, pela formação de ácidos e presença de polifenóis, portanto o período real da acessibilidade e atuação das enzimas aos substratos, é curto (HANSEN et al., 2000). Sendo assim, o controle da temperatura e do pH irá proporcionar maior eficiência das isoenzimas, prolongando o período de melhor atuação durante o processo fermentativo, propiciando uma melhor qualidade das amêndoas de cacau e por consequência, maior qualidade na produção de chocolates monovarietais, no que diz respeito à formação e potencialização dos precursores de aroma e sabor. 85 REFERÊNCIAS ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS. Official Methods of Analysis of AOAC International, 2000. BHATTI, N, H. et al. Studies on Kinetics and Thermostability of a Novel Acid Invertase from Fusarium solani. Journal of Agricultural and Food Chemistry. 2006, 54, p. 4617−4623. BECKETT, S. T. Industrial chocolate manufacture and use. 4 ed. London: Chapman and Hall, p.20-23, 2009. DEUNER, S. et al. Caracterização parcial da invertase ácida solúvel em tubérculos de batata: energia de ativação e efeito de inibidores. Revista Brasileira de Agrociência, Pelotas-RS, v. 11, 45-50, 2005. DARREN J. VORSTER and FREDERIK C. BOTHA. Partial purification and characterisation of sugarcane neutral invertase. Phytochemistry. 1998, 49, p. 651-655. DU, L. et al. Characterization of an Invertase with pH Tolerance and Truncation of Its NTerminal to Shift Optimum Activity toward Neutral pH. PLOS ONE. 2013, 8, e62306. GOMEZ, M L P A. et al. Metabolismo de carboidratos durante o amadurecimento do mamão (Carica papaya, L. cv. Solo): influência da radiação gama. Ciência e Tecnologia de Alimentos. 1999, v.19, 246-52. HANSEN, C, E; OLMO, M; BURRI, C. Enzyme Activities in Cocoa Beans During Fermentation. Journal Science Food Agricultural. 1998, 77, 273 – 281. HANSEN, C, E; ÄEZ, A, M; BURRI, C; BOUSBAINE, A. Comparison of enzyme activities involved in flavour precursor formation in unfermented beans of different cocoa genotypes. Journal of the Science of Food and Agriculture. 2000. 80, p. 1193-119. HUSSAIN, A.; RASHID, M.; PERVEEN, R.; ASHRAF,M. Purification, kinetic and thermodynamic characterization of soluble acid invertase from sugarcane. Plant Physiology et Biochemistry, in press, 2008. LINEWEVER H.; BURK, D. The determination of enzyme dissociation constants. Journal of American Chemical Society. 1934, 56, 658-666. LOPEZ, A.P.; DIMICK, P.S. Cap. 25: Enzymes involved in cocoa curing In: Food Enzymology Vol.2. Elsevier Applied Science. p. 211-236,1991. LOWRY, O. H. et al. measurement with the Folin fenol reagent. Journal of Biological Chemistry. 1951, 193, 265- 275. 86 MISNAWI, JINAP, S; NAZAMID, S; JAMILAH, B. Activation of remaining key enzymes in dried under-fermented cocoa beans and its eơect on aroma precursor formation. Food Chemistry. 2002, 78, 407–417. NASCIMENTO, R. et al. Distribuição de amido, açucares solúveis e atividade de invertases em explantes de soja sob várias concentrações de sacarose e diferentes fontes de nitrogênio. Revista Brasileira de Fisiologia Vegetal. Viçosa- MG, v. 10, 125-30, 1998. NELSON, N. A photometric adaptation of Somogyi method for determination of glucose. Journal of Biologic Chemistry, Bethesda, Md, v. 153, n. 2, p. 375-380, Feb. 1960. PEZOA-GARCÍA, N,H. Contribuition a I’étude d’um capteur por controler em continu procede de torréfaction.170p. 1989. These (Docteur) – Université de Technologie de Compiegne, Compiegne, 1989. ROITSCH, T; GONZALEZ, M, C. Function and regulation of plant invertases: sweet sensations. Trends Plant Sci. 2004, 9, p. 606–613. ROSLI, W, I, W; JINAP, S; RUSSLY, A, R. Effect of roasting time and temperature on volatile components profiles during nib roasting. In Proceendings of the 12th Internacional Cocoa Reaserch Conference. Salvador, Brasil, Ccocoa Producers. P977-991, 1996. SACHDEVA, M; MANN, A, E, S; BATTA, S, K. Multiple Forms of Soluble Invertases in Sugarcane Juice: Kinetic and Thermodynamic Analysis. Sugar Tech. India, 2003, 5, p.31 35. STURM, A. Invertases primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiology, Rockville, v.121, p.1-7, 1999. YUSEP, I. et al. Influence of carboxypeptidases on free aminoacid, peptide and methylpyrazine contents of under-fermented cocoa beans. Journal Science Food Agriculture. 2002, 82, 1584-92. 87 CONCLUSÕES FINAIS x Os resultados expostos no presente estudo evidenciam a diferença e especificidade existente entre os cultivares de cacau (PH 16 e TSH 1188) e entre polpa e semente em cada cultivar, em relação à atividade enzimática da polifenoloxidase e invertase. x A PPO apresentou boa afinidade para o substrato catecol, assim como a invertase apresentou para o substrato sacarose, uma vez que quanto menor o valor de Km maior será a afinidade da enzima pelo o substrato. x Com relação ao efeito do pH na atividade da PPO, foi observado que para o cultivar TSH 1188 a polpa apresentou maior atividade que a semente, e para o cultivar PH 16, as atividades possuem comportamentos semelhantes. Foi evidenciado que a ação da PPO na semente em ambos os cultivares, é favorecida pelo pH no estágio inicial da fermentação (primeiros 3 dias). Em pH alcalino há uma diminuição brusca da atividade, chegando a representar entre 70 a 90 % de perda no seu potencial. Para a invertase em ambos os cultivares, a atuação da invertase ácida é favorecida mais na polpa do que na semente. A invertase neutra, apesar de apresentar valores ótimos de pH fixos para maior atividade da enzima, atua fora da faixa de pH durante todo o processo de fermentação, o que implica dizer que esta enzima não possui atuação nestas condições de fermentação para os cultivares estudados, porém apresenta maior atividade na polpa do que na semente. x Com relação à temperatura ótima de maior atuação das enzimas, foi observado que nos dois cultivares, tanto na polpa quanto na semente, a atividade da PPO diminuiu gradativamente com o aumento da temperatura, o que sugere que altas temperaturas não a favorecem, e implica em afirmar que zona de maior atuação desta enzima é entre 25 e 30°C. Para a enzima invertase para ambos os cultivares, foi evidenciado que a atividade da invertase ácida é maior na semente do que na polpa. Para invertase neutra, a polpa apresenta maior atividade que a semente e sua atuação é favorecida sob altas temperaturas, apresentando zona de maior atividade entre 50 e 52°C. Comparando-se a atuação das duas isoenzimas (ácida e neutra) nos dois cultivares, tanto na polpa quanto na semente, é possível afirmar que a atividade da invertase ácida é maior (4 a 5 vezes) do que na invertase neutra. Este fato indica que esta enzima é cineticamente mais eficiente e pode ser a principal forma de enzima vacuolar envolvida na hidrólise da sacarose durante o processo de fermentação. 88 x As correlações apresentadas neste estudo sugerem que no decorrer do processo de fermentação, sejam utilizados critérios específicos para cada cultivar de cacau, tomando como base suas especificidades e parâmetros ideais, para que se possa prolongar e potencializar a atividade destas enzimas. Para a PPO, o controle desses parâmetros é de suma importância, visto que a oxidação dos compostos fenólicos presentes no cacau, realizada pela enzima, é determinante para às características sensoriais do chocolate a ser produzido, principalmente no que diz respeito à redução do amargor e da adstringência. Logo, prolongar a atuação da PPO em temperaturas mais baixas, através do aumento na frequência e na quantidade dos revolvimentos da massa de cacau no início do processo de fermentação, pode retardar a elevação da temperatura e a queda do pH, o que poderá favorecer a atuação destas enzimas nos cultivares estudados. Com relação à invertase, o controle desses parâmetros é crucial para prolongar sua atividade, uma vez que a estabilidade e atividade desta enzima são fatores limitantes para a conversão total da sacarose e formação de precursores de sabor durante a fermentação e que serão continuados e potencializados nas etapas seguintes de secagem e principalmente na torrefação. x O controle da temperatura e do pH irá proporcionar maior eficiência das enzimas estudadas, prolongando o período de melhor atuação durante o processo fermentativo, uma vez que estas exibem diferentes estabilidades durante o processo, principalmente no que se refere à inativação. Esta é causada pelo calor gerado durante o processo, pela formação de ácidos e presença de polifenóis, portanto o período real da acessibilidade e atuação das enzimas aos substratos é curto. Logo, o controle das condições de fermentação e até mesmo a possibilidade de intervenções tecnológicas, como a aplicação de enzimas exógenas comerciais de padrão de atividade semelhantes às nativas do cacau, irão propiciar uma melhor qualidade das amêndoas de cacau e por consequência, maior qualidade na produção de chocolates monovarietais, no que diz respeito à formação e potencialização dos precursores de aroma e sabor. 89 SUGESTÕES PARA ESTUDOS FUTUROS x Sugere-se a extração e caracterização da enzima protease e suas isoenzimas, nos mesmos cultivares estudados no presente trabalho, para que a caracterização das principais enzimas presentes no cacau seja completa; x A partir da caracterização e determinação das condições ótimas de atuação das enzimas polifenoloxidade, invertases e proteases, poderão ser fornecidos subsídios necessários para promover possíveis intervenções tecnológicas durante o processo de fermentação do cacau e a aplicação do controle dos parâmetros de fermentação apontados, visando a melhoria da qualidade do cacau produzido e principalmente melhoria na qualidade do chocolate a ser processado.
Baixar