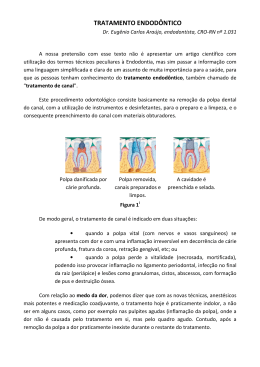
Sipert, Carla Renata Si74p Produção de MIP-1alfa e SDF-1 por fibroblastos de polpa dental humana em cultura frente ao desafio com Enterococcus faecalis inativado por calor/ Carla Renata Sipert -- Bauru, 2007. xx, 65 p. : il. ; 30 cm. Dissertação (Mestrado) -- Faculdade de Odontologia de Bauru. Universidade de São Paulo. Orientador: Prof. Dr. Carlos Ferreira dos Santos Autorizo, exclusivamente pra fins acadêmicos e científicos, a reprodução total ou parcial desta tese, por processos fotocopiadores e/ou meios eletrônicos. Assinatura do autor: Data: Comitê de Ética em Pesquisa em Seres Humanos da FOBUSP: projeto de pesquisa aprovado em 30 de novembro de 2005. Número do protocolo: 131/2005 Dados Curriculares Dados Curriculares Carla Renata Sipert Nascimento Santo André – São Paulo 14 de setembro de 1979 Graduação em Odontologia na Faculdade de Odontologia de Bauru / Universidade de São Paulo 1998 - 2001 Especialização em Endodontia pelo Hospital de Reabilitação de Anomalias Craniofaciais / Universidade de São Paulo 2002 - 2004 Mestrado em Endodontia na Faculdade de Odontologia de Bauru / Universidade de São Paulo 2005 - Atual iv “O conhecimento começa (...) quando, olhando para um objeto, sentimos que ele está a nos dizer: Decifra-me ou te devoro! Mas essa ordem do objeto, somente os curiosos a ouvem. É a curiosidade que nos faz fazer perguntas. É na curiosidade que o pensamento se inicia.” “ Quem está possuído pela curiosidade não descansa.” Rubem Alves, 2005 v Dedicatória e Agradecimentos Dedicatória e Agradecimentos ou Sobre limas e fibroblastos... A história que vou contar agora pode não ser muito original, mas de qualquer forma foi única para a minha vida: o meu Mestrado! Uma fase com a qual eu já sonhava desde a graduação e que em 19 de abril de 2004, o dia da prova de seleção, começava a ser real. Esta é a típica circunstância que deixa qualquer pessoa fora de seu estado emocional natural. Teria sido assim comigo se não fosse a providencial presença de meus queridos amigos de graduação dias antes da tão esperada prova. Graças a André Marchiori (Tony), Ednir Matheus Rossi, Daniel Silva (Mona), Gustavo Souza (Talento), Danilo Souza, Fernando Ueno (Tanakama), Luis Fernando Lopes (Slot) e Marina Vaz de Lima, não houve espaço para tensão em meio a tanto bom humor e critatividade. Os demais amigos que não estavam em Bauru, como Camila Rodini, Andréia Dias, Ana Carolina Jaña (Calanga), Marcelo Montanha (Montanha), Pedro Coesta e Deise Yamashiba devem ter feito uma bela corrente de pensamentos positivos de forma que algumas semanas depois da prova, meu nome constava dentre os alunos selecionados para o mestrado em endodontia – 2005/2007. Tive o privilégio de dividir esta comemoração com Celso Kenji Nishiyama, Renata Hussne e Edninha, pessoas que me incentivaram a fazer o mestrado mesmo ainda na especialização. Meus divertidos colegas: Victor Hugo Lopes, Fábio Aznar, Arnaldo Santana, Carolina Novaretti, Micheli Izeppe, Bianca Zambon, Carina Martini, Tatiana Bella, Marina Vaz de Lima, Daniele Fakhoury e Juliana Baldani, já deixavam saudades logo que o curso de especialização terminou. Pareceram muito curtos os 6 meses que precederam as aulas do mestrado. Um longo ciclo de disciplinas básicas teve início e, no decorrer do aprendizado que tivemos acerca de bases de dados, bioestatística, normas bibliográficas, revisões sistemáticas, softwares milagrosos, docência na Universidade e fotografia ao alcance de todos; tivemos a oportunidade de fazer amigos dentre todas as áreas, alguns dos quais levei por todo o curso: Luciana Rezende, Nicolas, Dafna, Érica Bariri, Simone Faustino, Ana Carolina Fracalossi, Janaína Ghizoni, Marcela Claudino, Ana Paula, Renata, Marta, Dani Fa, Flora, Gustavo Lauttenschläger, dentre muitos outros. Os meses que se seguiram foram de intensa convivência com os colegas da endodontia. Alguns dias depois das aulas já iniciadas, um deles ainda faltava: Ramiro Oropeza, o boliviano. Foi preciso pouco tempo para que todos percebessem a calma e tranqüilidade com que ele lidava com os trabalhos. Aliás, calma essa que contagiou a turma de tal forma que foi possível que formássemos um grupo tão unido que em todos estes meses de trabalho, compromissos e tensões, nunca houve uma única desavença entre nós. Pelo contrário, aos poucos conheceríamos a história e a personalidade de cada um. A Roberta, que por tantas vezes deixava seu filho Alexandre com os avós para executar os trabalhos em grupo, nunca faltou com sua simpatia e disposição durante estes momentos. A Márcia, sempre elegante e delicada mesmo após horas de trabalho seguidas. O Bruno, com tanta praticidade e agilidade, vi Dedicatória e Agradecimentos que me fazia muitas vezes pedir para o Fernando traduzir o que ele nos dizia! Este, por sua vez, sempre disposto a ajudar a todos e a rir de nossas piadas. A Clarice, com seu carisma, criatividade e muita animação, conquistou logo o carinho de toda turma. Essa convivência agradável era melhorada ainda mais pela presença da Patrícia com sua alegria constante e bom humor aliados sempre à prudência e maturidade em suas colocações. Mas sem dúvida, não houve maior parceiro nestes dois anos que pudesse superar o Ronan: não bastasse sermos colegas da mesma área, nos tornamos parceiros de dança, executamos nossos trabalhos no mesmo laboratório, compartilhamos nossas dificuldades e sucessos sempre da mesma maneira: rindo muito! Tanto convivemos que eu nem pensei em outra pessoa para me atender quando eu precisei de um endodontista. Evidentemente não fiquei livre do seu humor pontiagudo nem mesmo nesta situação. Em meados de abril de 2005, a minha orientação pelo professor dr. Carlos Ferreira dos Santos finalmente havia sido formalizada e poderíamos então pensar no projeto que seria executado durante o mestrado. Após um contato feito com a profa Dra Sandra Helena Penha de Oliveira, da Faculdade de Odontologia da UNESP de Araçatuba, o prof. Carlos e a dra Sandra me propuseram juntos um trabalho no qual eu faria cultura de fibroblastos de polpa para estudar a resposta destas células frente a um estímulo bacteriano. Imediatamente me apaixonei pela idéia e em julho deste mesmo ano passei 15 dias no laboratório da profª Sandra onde tive a oportunidade de entrar em contato pela primeira vez com parte daquilo que eu executaria nos próximos meses. A drª Sandra mostrou‐se completamente solícita tanto no ambiente de trabalho como em sua própria casa onde me recebeu. A sua ajuda acabou se estendendo por todo o período de execução do projeto: mesmo longe, ela se mostrava pronta a sanar minhas dúvidas a qualquer hora do dia. Ainda em Araçatuba, tive a oportunidade de trabalhar novamente com uma contemporânea minha de especialização, na época mestranda e orientada da drª Sandra, Alessandra Gomes, cuja ajuda foi fundamental para o máximo aproveitamento desta minha fase de estágio. Fiz mais alguns novos amigos, incluindo o prof. Dr. João Eduardo Gomes a quem devo muitas horas de conversas extremamente agradáveis, inteligentes e bem humoradas. Foi também nesta ocasião que conheci a amiga Melaine Lawal. Ao ser aprovada no doutorado em Bauru, ela se mudou para meu apartamento e então pudemos conviver por mais de um ano num clima de muita descontração e também de aprendizado mútuo. Ao voltar de Araçatuba, então, providenciei a redação do projeto que, por decisão do prof. Carlos, foi submetido à FAPESP para a solicitação de um auxílio. Em outubro do ano seguinte, este financiamento foi concedido ao meu orientador. Ainda no segundo semestre de 2005, cumprindo disciplinas e atividades, tive a imensa satisfação de acompanhar, pela primeira vez, a atividade dos alunos da graduação. Foram meses de trabalho vendo colegas aprendendo endodontia exatamente no mesmo local e da mesma forma como eu aprendi. Eram inevitáveis os momentos de saudosismo e menos ainda os momentos cômicos no decorrer desta convivência com alunos sempre tão alegres e tão divertidos. Momentos estes que eram reforçados pela nossa querida d. Neide, que sempre complementava nossa rotina com muita alegria e sabedoria. Alegria, por sinal, era algo que dificilmente faltava no ambiente de trabalho quando o Edimauro se fazia presente com uma velocidade de raciocínio incrível: tanto para suas ciladas como para seu trabalho. O convívio com os professores do departamento deixou para nós, alunos de mestrado, exemplos a serem vii Dedicatória e Agradecimentos levados para o resto de nossas vidas: a experiência do prof. Dr.Norberti Bernardinelli, a gentileza e minuciosidade do prof. Dr.Roberto Brandão Garcia, a competência e a seriedade do prof. Dr. Clóvis Bramante e o constante carinho e sabedoria do prof. Dr. Ivaldo Gomes de Moraes. Grandes mestres que me motivaram durante a graduação e que me ensinaram a motivar durante este mestrado. Em todos os momentos que precisamos de alguma assistência, tivemos a orientação da secretária Suely Bettio e ainda a imensa ajuda da Patrícia. A ajuda de todos os funcionários foi fundamental para nós, alunos, durante estes dois anos. Iniciado o ano de 2006, tornava‐se urgente o início da execução do meu projeto e para que o mesmo pudesse ser executado em Bauru, precisei contar com a ajuda da Prof ª Drª Ana Paula Campanelli, que cederia, por mais de um ano, um espaço no seu laboratório e muito tempo de sua agenda para supervisionar minhas atividades. O convívio com novos colegas se iniciou e aos poucos se tornou uma amizade bem mais intensa do que eu poderia imaginar. Trabalhar com muitas pessoas competentes já seria por si só estimulante, mas ainda melhor com pessoas que fazem o possível para que o ambiente de trabalho se mantenha agradável. Foi assim que fui inserida no grupo formado por Rodrigo, Victor, Aninha, Vanessa, Michele ,Tatiana Sales, Valéria Gelani, Thaís Gasparoto e Eduardo Figueira. A estes últimos quatro devo imensa gratidão por tamanha ajuda, atenção e orientação em tantos momentos em que me senti perdida perante coisas novas a aprender. O convívio diário no laboratório acabou deixando alguns momentos marcantes entre nós. A cada momento de dificuldade e desânimo, pude contar com o olhar de solidariedade da Tati! A experiência e sensatez da Val sempre me inspiraram uma imensa confiança de maneira a sempre recorrer a suas palavras em momentos difíceis. A ajuda e paciência do Eduardo salvaram as culturas por incontáveis vezes. Finalmente, o exemplo de alto astral e dedicação da Thaís serviram constantemente como injeção de ânimo e inspiração para que eu executasse meu trabalho. O convívio entre os colegas ia muito além do laboratório e dos corredores da faculdade. Intermináveis horas de conversas culturais e divertidas ocorreram em nossas casas, bares e festas. Ronan, Clarisse, Fernando, Thaís, Minke Gasparoto, Wagner Baseggio e Marcela Claudino, Ericson Janolio e Melina Bortolo são os responsáveis por tantos momentos animados e descontraídos durante esses dois anos. Em meio a estes encontros informais, uma amizade fantástica se estabeleceu entre mim e Minke Gasparoto. Sua presença rende, na mesma proporção, gargalhadas incontroláveis alternadas com riquíssimas conversas sobre filmes, músicas e, obviamente, direito internacional. Outros amigos apareceram em momentos cruciais: Rosário Zedebski me ajudou muito no início do trabalho com a realização da fotografia das culturas. Dentre nossos encontros em corredores, nunca houve ocasião em que seu alto astral não me contagiasse. Outros amigos reapareceram depois de muito tempo longe, como Milton Santa Maria Jr., que eu conheci ainda na minha graduação e que ao retornar a Bauru revelou‐se um grande amigo. Juntamente com sua noiva Aline, muitos momentos de descontração se passaram tanto em Bauru como em trânsito para Ribeirão Preto. Outro amigo de longa data a quem ajudei foi Heitor Honório: somente aceitei ser voluntária de sua pesquisa pela estima e consideração que tenho por ele desde a época da graduação. No entanto, o aparelho que nós, voluntários, usávamos dia e noite (pingando açúcar, colocando de molho na Coca‐Cola, etc.) para seu estudo de desmineralização de esmalte, serviria perfeitamente como um modelo de indução viii Dedicatória e Agradecimentos de estresse em seres humanos. Isso sem contar todas as explicações que eram fundamentais quando se almoçava fora de casa, quando toda a parafernália da pesquisa tinha que ser levada a tiracolo! Na rotina do laboratório, também pude contar com a imensa ajuda da secretária Dalva Ribeiro de Oliveira, que se mostrou uma grande amiga por diversas horas de conversa descontraída nos intervalos do trabalho. A companhia do técnico André Luis da Silva também contribuiu diversas vezes para trabalhos bem feitos num clima bastante agradável. Cláudia também se fez fundamental durante toda a organização de material para a execução dos protocolos. Para que o meu projeto tivesse início, algumas parcerias tiveram que ser feitas com colegas cirurgiões‐dentistas que me ajudariam a providenciar os terceiros molares para a padronização das culturas primárias. Foi fundamental nesta fase a ajuda que recebi de Érica Kuriki, Fernando Giglio, Karin Modena e Bella Luna Colombini. Durante a padronização do protocolo, pude contar com a ajuda da colega Vivien Sakai, o que foi de imensa valia naquele momento. Outros colegas do laboratório de farmacologia deram seu apoio e sua solidariedade durante diversas fases do trabalho como Adriana Calvo, Thaís Marchini, Ana Elisa Akashi e Thiago Cruvinel. Aos poucos eu também comecei a participar dos protocolos de cirurgia do LAFFIC e tive a oportunidade de conhecer melhor os alunos de iniciação científica Thiago, Leonardo, Paulo e Anderson. Foi muito gratificante poder contribuir e aprender com o trabalho destes queridos alunos! Durante a execução dos experimentos, eu precisei me deparar com muitas dificuldades, como já era previsto. Não raro, o trabalho era perdido em razão de culturas contaminadas, PCR´s borrados e citometrias frustradas. No entanto, nunca me faltou a ajuda de profissionais experientes e extremamente generosos como o dr. Fernando Cunha e o dr. João Santana (FMRP/USP) que nos cederam os anticorpos para a execução de ELISA. Juliana Bertozzi, funcionária da referida escola, gentilmente realizou a análise das amostras dos meus experimentos. Para a citometria de fluxo, contamos com a colaboração do Hemonúcleo do Hospital Amaral Carvalho através da profissional Fátima Barrach. A todos, o meu muito obrigado pela colaboração e paciência! Muitos profissionais do campus tiveram participação fundamental durante toda esta fase. A um grande funcionário desta escola dedico um imenso carinho e admiração: Thiago José Dionísio. Alguém que só há pouco concluiu sua graduação, mas já se mostra um perfeito mestre em paciência, bom humor, simpatia e principalmente, competência. Características estas também coroadas, de maneira unânime, pela profissional Vera Rufino Rosa. Muitos outros funcionários fizeram parte do convívio destes anos sempre de maneira solícita e agradável como Marilza, Eva, Ovídio, Gilmar, Thelma, Solange, seu Luís, Zilei, Mônica, Sueli. Cada qual essencial à escola em suas atividades. Sobre o significado para mim da Faculdade de Odontologia de Bauru haveria páginas do que falar, mas me contentarei em declarar a paixão que trago comigo desde o dia em que aqui pisei pela primeira vez na ocasião da matrícula para o curso de graduação. Neste local, muito mais sobre a vida aprendi do que ser cirurgiã‐dentista ou especialista ou mestre. Dedico a gratidão que carrego comigo ao atual diretor da escola prof. Dr. Luis Fernando Pegoraro e aos ex‐diretores durante todo o período em que estive aqui: Profª Drª Maria Fidela de Lima Navarro, atual secretária geral da USP, ix Dedicatória e Agradecimentos e ao prof. Dr. Aymar Pavarini, que me concedeu o grau de cirurgiã‐dentista no ano de 2001. À presidente da pós‐graduação, Profª Drª Maria Aparecida de Andrade Moreira Machado, meus agradecimentos pelo empenho e dedicação pela pós‐graduação desta escola. Agradeço também às secretárias da pós‐graduação Giane Tenório Quintela, Ana Letícia Momesso e Maria Margareth Mokarzel por tamanha paciência durante a solução dos trâmites burocráticos. Essencial agradecer ainda ao governo federal pela disponibilização da bolsa CNPq que proveu meu sustento ao longo destes dois anos. Ao governo do Estado de São Paulo deixo minha admiração por ainda manter uma universidade pública do nível da Universidade de São Paulo, respeitada dentre todas as grandes universidades do mundo. Agradeço mais uma vez ao governo estadual pelo auxílio concedido pela agência FAPESP, o que tornou este projeto e alguns futuros, passíveis de realização. Muito conhecimento é necessário para que se idealize um projeto com propensões a ser mundialmente aceito e respeitado. A realização deste grande sonho devo à profª Drª Sandra Oliveira, que me ofereceu algo para que eu pudesse trabalhar com paixão. Muito mais do que apenas atenção e paciência recebi da profª Drª Ana Paula Campanelli. Inimaginável foi o crescimento que desenvolvi durante estes meses de convivência em seu laboratório. Por aquele que me recebeu como sua orientada sem nem sequer ter sido sua aluna, e ainda assim me dedicar tamanho voto de confiança que eu jamais esperaria receber; o meu imenso respeito e admiração! Declaro a minha gratidão ao meu orientador Prof. Dr. Carlos Ferreira dos Santos, um homem que carrega consigo uma verdadeira lista de exemplos a ser seguida por qualquer pessoa que deseja ser mais humana, competente, respeitada e acima de tudo, humilde. Muitos familiares torciam e se preocupavam comigo durante estes 2 anos de curso. Muito carinho recebi à distância dos primos Mônica, Paulo, Rosmeri e Silvani Prediger, família Sipert (Luís, tio Arno, Marcos, Nico, Hugo, Manfred, tia Voni, Hari), família Reimann (Liki, Mário, Wagner, Vinícius), família Reffatti (Márcio e Tatiana, tio Chico, Elizete e Eliane), Vera e Gilberto Agibert. Em especial, agradeço a atenção que recebo sempre de minha tia Betti Sipert, uma pessoa que se fez presente em minha infância e assim continua sendo até hoje! A Eunice Kusuda, minha gratidão por abrir as portas de sua casa para mim de forma tão amável! Por sempre me acolher com carinho e, especialmente, alegria; o meu muito obrigada! Por mais que o programa de mestrado e a convivência com tantos profissionais competentes fossem responsáveis por tanto aprendizado e crescimento durante esta fase, levo comigo a certeza de que grande parte deste amadurecimento não teria sido tão evidente sem a parceria e o companheirismo daquele que comigo dividiu cada entusiasmo, emoção, decepção, dúvida e desabafo. A presença de Ricardo Kusuda nesta fase de minha vida fez com que todos os momentos tivessem um brilho especial! Poucas coisas na vida são tão acalentadoras como o abraço de uma avó. A Melida Alma Reffatti meu muito obrigado por tanto carinho e por tantas coisas lindas que eu aprendo a cada dia com essa pessoa tão especial. Também agradeço ao meu avô Anicleto Reffatti por toda força e determinação que passam a cada geração. Aos meus saudosos avós Erna Ziel Sipert e Evaldo Sipert, de quem herdo meu sobrenome, a promessa de honrá‐lo sempre com honestidade e humildade. x Dedicatória e Agradecimentos Ao amigo de todas as horas, ao que trouxe consigo ao nascer a alegria sem limites, meu irmão Tiago Alexandre Sipert, minha imensa gratidão por tanto carinho e amor, por tanta saudade compartilhada, por tantas frases só por você interpretadas. Tenho certeza de que a sua existência faz a minha muito mais feliz. Aos meus amados pais Vili Sipert e Marli Reffatti Sipert eu dedico minha dissertação com todas as honras que vocês merecem. Somente pais sonhadores como vocês compreenderiam os anseios de um filho também sonhador. Agradeço pelo alento e força em tantos momentos de dificuldade, saudade e desânimo. Serei eternamente grata pela compreensão quanto à minha ausência e falta de tempo para com a família. E que Deus permita que vocês vivam por muitas décadas para que eu ainda possa honrá‐los por muito mais tempo como ordena seu Santo Mandamento. E ao Criador dos Céus e da Terra, eu declaro minha gratidão e alegria pelo privilégio da vida. Agradeço a Deus, meu Pai maior, por possibilitar tantas oportunidades maravilhosas como todas estas que vivi durante o meu curso de Mestrado. Ao dividir minha rotina entre Clínica e Bancada, vocês todos fizeram com que eu pudesse me realizar ao aprender cada vez mais sobre limas e fibroblastos!... Muito obrigada!!! Carla Sipert xi Sumário RESUMO 1 Introdução e Síntese Bibliográfica xx 3 2 Proposição 19 3 Material e métodos 23 3.1 Coleta do tecido pulpar..............................................................................................23 3.2 Cultura dos fibroblastos.............................................................................................23 3.3 Preparo do Enterococcus faecalis morto por calor....................................................24 3.4 Estimulação dos Fibroblastos Pulpares por Enterococcus faecalis morto por calor.24 3.5 ELISA........................................................................................................................24 3.6 Análise Estatística .....................................................................................................25 4 Resultados 29 4.1 Cultura dos fibroblastos.............................................................................................29 4.2 Produção de MIP-1α/CCL3 por ELISA....................................................................33 4.3 Produção de SDF-1/CXCL12 por ELISA .................................................................37 5 Discussão 43 6 Conclusão 51 Referências bibliográficas 55 ABSTRACT 65 Lista de Figuras FIGURA 1 Cultura de fibroblastos de polpa dental humana FIGURA 2 Produção de MIP-1α por fibroblastos de polpa dental humana 35 estimulados por Enterococcus faecalis inativado por calor FIGURA 3 Valores individuais da produção de MIP-1α/CCL3 por 35 fibroblastos de polpa dental humena estimulados por Enterococcus faecalis inativado por calor FIGURA 4 Produção de SDF-1 por fibroblastos de polpa dental humana 43 estimulados por Enterococcus faecalis inativado por calor FIGURA 5 Valores individuais da produção de SDF-1/CXCL12 por 53 fibroblastos de polpa dental humana estimulados por Enterococcus faecalis inativado por calor 31 Lista de Tabelas TABELA 1 Médias e erros-padrão da média (média ± EPM) referentes aos 33 valores de produção de MIP-1α/CCL3 por fibroblastos de polpa dental humana. TABELA 2 Médias e erros-padrão da média (média ± EPM) referentes aos 37 valores de produção de SDF-1/CXCL12 por fibroblastos de polpa dental humana. Lista de Abreviaturas e Símbolos ° = graus % = por cento et al = e colaboradores LTA = Lipoteichoic acid - Ácido lipoteicóico LPS = Lipopolysaccharide – Lipopolissacarídeo VEGF = Vascular endothelial growth factor – Fator de crescimento vascular endotelial TGF = Transforming Growth Factor – Fator de crescimento transformador VBNC = Viable but non-culturable – Viável mas não cultivável IL = Interleucin – Interleucina GRO = Growth-related Oncogene – Oncogene relacionado ao crescimento SDF = Stromal Cell-Derived Factor – Fator celular derivado de estroma MCP = Monocyte Chemotactic Protein – Proteína quimiotática para monócito MIP = Macrophage Inflammatory Protein – Proteína inflamatória para macrófago TNF = Tumor Necrosis Factor – Fator de necrose tumoral IP = Interferon-gama-inducible Protein – Proteína indutora de Interferon-gama NK = Natural Killer – Células do sistema imune denominadas de “Exterminadoras Naturais” IFN = Interferon CD = Cluster of differentiation – Agrupamento de diferenciação TLR = Toll-like Receptor – Receptor do tipo Toll MyD = Myeloid Differentiation – Diferenciação mielóide NOD = Nucleotide-binding Oligomerization Domain – Domínio de oligomerização ligado a nucleotídeo A.a = Aggregatibacter Actinomycetemcomitans HGF = Hepatocyte Growth Factor – Fator de crescimento para hepatócito ATM = Articulação Têmporo-mandibular IDO = Indoleamine-pyrrole 2,3-dioxygenase RANTES = Regulated on Activation Normal T-cell Expressed and Secreted CpG DNA = Notação utilizada para distinguir uma Citosina seguida de uma Guanina RT-PCR = Reverse Transcriptase – Polymerase chain reaction – Transcriptase reversa seguida de reação em cadeia da polimerase DMEM = Dulbecco´s Modified Eagle´s Medium – Meio de Eagle modificado por Dulbecco BHI = Brain Heart Infusion – Infusão de cérebro e coração PBS = Phosphate Buffered Saline – Tampão fosfato salina ELISA = Enzyme Linked Immuno Sorbent Assay = Ensaio imuno-enzimático FOB/USP = Faculdade de Odontologia de Bauru / Universidade de São Paulo Resumo Resumo A polpa dental é formada de tecido conjuntivo frouxo sendo constituída por diversas células, dentre as quais os fibroblastos são as mais numerosas. Ao serem submetidas a agressões diversas, estas células respondem com a liberação de substâncias, tais como citocinas e quimiocinas, que participam de maneira ativa no processo inflamatório. Assim sendo, este trabalho teve como proposição: 1. avaliar a capacidade de fibroblastos de polpa dental humana em cultura em produzirem as quimiocinas MIP-1α /CCL3 e SDF-1/CXCL12; 2. avaliar a produção destas quimiocinas pelos fibroblastos quando estimulados por Enterococcus faecalis morto por calor com relação à quantidade de bactérias por célula e 3. avaliar a liberação destas quimiocinas com relação ao tempo de estímulo. Para o estabelecimento das culturas, foi coletada a polpa de terceiro molar hígido de um paciente saudável. O tecido foi extraído, armazenado e picotado em meio de cultura para fibroblastos (DMEM), os quais foram utilizados a partir da quarta passagem. Após adesão das células a placas de 24 poços, o meio de cultura contendo Enterococcus faecalis morto por calor numa concentração correspondente a 1, 10 e 100 bactérias por fibroblasto foi adicionado aos poços. Após 1, 6 e 24 horas, o sobrenadante das células foi coletado para a análise por ELISA. A análise estatística foi realizada aplicando-se o teste Kruskal-Wallis com nível de significância de 5%. A produção de MIP-1α /CCL3 e SDF-1/CXCL12 pelas células pôde ser detectada por ELISA. Os fibroblastos pulpares se mostraram capazes de produzir SDF-1 constitutivamente sendo que o estímulo bacteriano levou a uma diminuição estatisticamente significativa desta produção. A produção de MIP-1α também foi detectada tanto de maneira constitutiva como em resposta ao desafio microbiano. Enquanto a concentração intermediária de bactéria por fibroblasto (10:1) mostrou uma produção semelhante ao grupo controle, as concentrações de 1 e 100 bactérias por fibroblasto induziram aumento maior na primeira hora de estímulo. Essas diferenças, entretanto, não foram estatisticamente significativas. A capacidade dos fibroblastos secretarem quimiocinas, como MIP-1α e SDF-1, reforça a importância dessas células dentro do contexto de imunidade e inflamação pulpar, principalmente por serem as células mais numerosas deste microambiente. Palavras-chave: Fibroblastos. Quimiocinas. Inflamação. xx Introdução e Síntese Bibliográfica 3 1 Introdução e Síntese Bibliográfica A estrutura do tecido pulpar A polpa dental é constituída de tecido conjuntivo frouxo derivado das células da crista neural ou do ectomesênquima e encontra-se confinada no interior da câmara pulpar e dos canais radiculares (CHIEGO JR1, 2002). Este tecido interage com outros como o periodonto e o sistema nervoso central e pela sua integração com a dentina, anatômica e funcionalmente, constitui juntamente com ela o complexo dentinopulpar (GOODIS2, 2002). A polpa contém células que desempenham de funções divididas em odontogênica, nutritiva, sensorial e defensiva, as quais permitem a preservação da sua vitalidade durante a manutenção da homeostasia e o reparo após a agressão (CHIEGO JR1, 2002). Apesar da polpa apresentar muitas propriedades em comum com outros tecidos conjuntivos, sua localização peculiar lhe impõe uma série de características ímpares a começar pela distribuição celular. Uma camada de odontoblastos, células especializadas que produzem dentina, circunscreve a periferia do tecido pulpar com seu corpo celular na polpa e seus prolongamentos no interior dos túbulos dentinários. Subjacente aos odontoblastos, encontra-se uma área livre de células – zona de Weil – constituída em sua maioria por fibras neurais não mielinizadas, capilares e processos de fibroblastos. Mais profundamente situa-se a zona rica em células constituída de fibroblastos em grande quantidade, células mesenquimais indiferenciadas, células de defesa (macrófagos e linfócitos), capilares e fibras nervosas. Internamente a esta área, localiza-se uma massa central de tecido conjuntivo conhecida por centro da polpa. Esta região é composta por fibroblastos, vasos sangüíneos, nervos, células mesenquimais indiferenciadas, macrófagos, linfócitos e fibras colágenas (OKIJI3, 2002). Dentre todos os grupos celulares que povoam a polpa, os fibroblastos são os mais numerosos, podendo ser encontrados em densidade ainda maior na zona rica em células (OKIJI3, 2002). Possuem um formato de estrela com longas extensões citoplasmáticas que permanecem em contato com as demais células (odontoblastos e outros fibroblastos) por meio de junções do tipo “gap” (TAKASHIBA; NARUISHI; MURAYAMA4, 2003). 4 Introdução e Síntese Bibliográfica Os estímulos agressores que desafiam a polpa dental e os tecidos periapicais O tecido pulpar é freqüentemente agredido por fatores ambientais como calor, trauma mecânico, bactérias e suas toxinas, sendo estas consideradas o fator etiológico principal da inflamação pulpar. Bactérias e seus derivados invadem a polpa dental em decorrência da cárie (a maior fonte de bactérias em infecções da polpa e da área periapical), de fraturas, de formações dentárias anômalas ou como conseqüência de procedimentos restauradores (TROWBRIDGE5, 2002). Os microrganismos sob lesões cariosas têm sido isolados e observa-se, de maneira geral, a presença de Streptococcus sp., Actinomyces sp., Eubacterium sp. e Lactobacillus sp. em alta freqüência em lesões de cárie superficial, enquanto que Eubacterium sp., Propionibacterium sp. e Bifidobacterium sp. são isolados de lesões de cárie profunda (LOVE; JENKINSON6, 2002). A presença de Enterococcus sp. também é relatada em dentina cariada (MARTIN et al.7, 2002). A importância de microrganismos na etiopatogenia da doença periapical já tem sido amplamente 8 elucidada pela literatura (KAKEHASHI; STANLEY; 9 FITZGERALD , 1965, MOLLER et al , 2004). Especial atenção tem sido destinada ao perfil microbiológico dos casos de insucesso do tratamento endodôntico. Neste contexto, vários estudos têm relacionado a prevalência de Enterococcus faecalis em lesões periapicais refratárias aos protocolos de tratamento atualmente empregados (KAUFMAN et al.10, 2005, SEDGLEY et al.11, 2005, WILLIAMS et al.12, 2006, ZOLETTI; SIQUEIRA; SANTOS 13, 2006). Enterococus habitam, de forma comensal, o trato gastrintestinal de humanos, mas podem desencadear uma grande variedade de doenças como infecções urinárias, endocardite bacteriana, dentre outras (JETT; HUYCKE; GILMORE14, 1994). A presença de Enterococcus faecalis também se dá na cavidade oral em locais como saliva, sulco gengival, dorso da língua e canais radiculares infectados (SEDGLEY; LENNAN; CLEWELL15, 2004, SEDGLEY; BUCK; APPELBE16, 2006). Entretanto, é notável a alta prevalência em casos de insucesso do tratamento endodôntico, ou seja, quando da persistência da inflamação periapical ou seu aparecimento após a intervenção profissional. Em estudos que utilizam métodos de identificação por meio de biologia molecular, encontram-se prevalências deste microrganismo de 12,1% (KAUFMAN et al.10, 2005), 43% (WILLIAMS et al.12, 2006), 80% (ZOLETTI; SIQUEIRA; SANTOS13, 2006) e 89,6% (SEDGLEY et al.11, 2006). WILLIAMS et al.12, 2006, relatam que E. faecalis foi de 2 a 3 vezes mais 5 Introdução e Síntese Bibliográfica freqüente em infecções refratárias do que em primárias. As possíveis razões para esses achados podem estar relacionadas à capacidade de E. faecalis em se adaptar a condições adversas (FLAHAUT et al.17, 1996), como ocorre durante as diferentes fases do tratamento endodôntico (WILLIAMS et al.12, 2006). Estes mecanismos de defesa estão diretamente relacionados aos seus fatores de virulência, como por exemplo, adesinas que se ligam ao colágeno tipo I (ROZDZINSKI et al.18, 2001), principal componente orgânico da dentina (LINDE; GOLDBERG19, 1993), conferindo à bactéria a capacidade de se alojar profundamente nos túbulos dentinários, permanecendo inatingíveis pela ação do preparo biomecânico dos canais radiculares (WILLIAMS et al.12, 2006). Por ser o trato gastrintestinal seu nicho natural, o ambiente alcalino proporcionado pelo uso da medicação intracanal de hidróxido de cálcio não é capaz de inviabilizá-lo como a outros microrganismos (BYSTROM; CLAESSON; SUNDQVIST20, 1985, DISTEL; HATTON; GILLESPIE21, 2002). Nestas circunstâncias, E. faecalis é capaz de sobreviver no interior dos canais radiculares mesmo na ausência de outros microrganismos (MOLLER et al.9, 2004). Além disso, em ambientes desfavoráveis, E. faecalis é capaz de assumir um estado denominado VBNC (“viable but nonculturable” – viável mas não cultivável) (SIGNORETO et al.22, 2000), uma estratégia de sobrevivência que o mantém metabolicamente ativo até que no meio no qual ele se encontra, as condições favoráveis sejam restabelecidas (OLIVER23, 2005). Outro fator de virulência importante é o ácido lipoteicóico (LTA), um componente importante presente na membrana celular externa de microrganismos Gram positivos, como é o caso de E. faecalis. O LTA é uma adesina anfifílica associada à membrana com um fator de virulência potente que se assemelha ao LPS (lipopolissacarídeo) de membrana. O LTA age como uma molécula de adesão que facilita a colonização e a invasão de bactérias para o interior dos tecidos (GINSBURG24, 2002). Os fibroblastos do tecido pulpar respondem de diferentes formas a estímulos agressores Em resposta aos estímulos agressores que acometem o tecido pulpar, como calor e trauma mecânico e principalmente toxinas bacterianas, as células da polpa possuem a capacidade de se reparar, se diferenciar em odontoblastos e produzir uma série de proteínas da matriz durante o processo de reparo (CHAN et al.25, 2005). Nem toda 6 Introdução e Síntese Bibliográfica reação inflamatória da polpa, portanto, resulta em dano permanente. Quando a lesão é eliminada antes de comprometer em definitivo a polpa, a inflamação caminha para a resolução e começa então o processo de reparo (TROWBRIDGE5, 2002) e nesta fase, a síntese de colágeno é acelerada (OKIJI3, 2002). Mediante a ação de citocinas como o fator de crescimento tumoral (TGF)-β1 e TGF-β2 e interleucina (IL)-1β, a deposição de colágeno no tecido pulpar mostra-se aumentada (BARKHORDAR et al.26, 2002, CHAN et al.25, 2005), provável razão pela qual fibroblastos inflamados depositam colágeno em maior quantidade do que as mesmas células em condições normais (BARKHORDAR et al.26, 2002). Esta síntese aumentada de colágeno pelos fibroblastos durante o processo inflamatório é um evento fundamental para o reparo da polpa dental humana (CHAN et al.25, 2005). Recentemente, tem sido discutido que em resposta a estímulos variados, os fibroblastos, além de produzirem colágeno, são capazes de secretar também vários tipos de citocinas (SISMEY-DURRANT; HOPPS27, 1991, TAMURA et al.28, 1992, YAMAJI et al.29, 1995, WANG et al.30, 1999, SUGIYAMA et al.31, 2000, WANG et al.32, 2000, HOSOKAWA et al.33, 2005, UEHARA; TAKADA34, 2007) e da mesma forma os fibroblastos pulpares (TOKUDA; NAGAOKA; TORII35, 2002, YANG et al.36, 2003, YANG et al.37, 2003, COIL; TAM; WATERFIELD38, 2004, YANG et al.39, 2004). Citocinas são proteínas secretadas por células de imunidade tanto inata como adaptativa que atuam como mediadores destas mesmas células. São consideradas reguladores fundamentais dos mecanismos de homeostasia como resposta imune, inflamação e reparo (OLLIER40, 2004). As citocinas podem ser produzidas em resposta a microrganismos e seus produtos, sendo que diferentes citocinas estimulam diversas respostas em células constituintes da imunidade e inflamação. Dentre as citocinas, destaca-se uma grande família de homólogos estruturais que estimulam o deslocamento de leucócitos e regulam sua migração do sangue aos tecidos. A estas citocinas, deu-se o nome de quimiocinas (citocina quimiotática). Elas são produzidas por várias células em resposta a estímulos inflamatórios recrutando, assim, leucócitos ao local da inflamação (ABBAS; LICHTMAN41, 2003). As quimiocinas são todas polipeptídeos entre 8 e 14 kDa sendo classificadas em subfamílias com base no número e posição dos resíduos de cisteína N-terminal. As duas maiores subfamílias são a CC (resíduos de cisteína estão adjacentes) e a CXC (os resíduos são separados por um aminoácido). Um número menor de quimiocinas é classificado como C (apenas um resíduo de cisteína) ou CX3C (três aminoácidos separando os resíduos) (ZLOTNIK; YOSHIE42, 2000). As Introdução e Síntese Bibliográfica 7 quimiocinas dos grupos CC e CXC são produzidas por leucócitos, células endoteliais, células epiteliais e também por fibroblastos. Como exemplos de quimiocinas da subfamília CXC, podemos citar interleucina-8 (IL-8), oncongene relacionado com o crescimento (GRO) e fator estroma derivado de célula (SDF). Já dentre os componentes da subfamília CC, destacam-se a proteína quimiotática para monócito (MCP) e a proteína inflamatória para macrófago (MIP) (SCHALL et al.43, 1993, OGURA et al.44, 2005). Em algumas destas células, a liberação de quimiocinas é induzida por microrganismos e por outras citocinas como o fator de necrose tumoral (TNF) e IL-1. Essa liberação constitui um importante mecanismo de defesa, pois recruta leucócitos para a região onde ocorre a infecção (ABBAS; LICHTMAN41, 2003). A quimiotaxia ocorre em resposta a um gradiente local de quimiocinas no espaço extracelular (DIEUNOSJEAN et al.45, 1999). Os fibroblastos pulpares têm-se mostrado capazes de produzir a quimiocina IL-8/CXCL8 como constatado por alguns estudos (NAGAOKA et al.46, 1996, YANG et al.36, 2003). As quimiocinas MIP-1α/CCL3 e SDF-1/CXCL12 As primeiras quimiocinas foram descobertas no fim da década de 80 devido a suas ações angiostática (por exemplo IP-10/CXCL10) e quimiotática para neutrófilo (por exemplo IL-8/CXCL8) (DIEU-NOSJEAN et al.45, 1999). O interesse pelo estudo de quimiocinas tem crescido cada vez mais, principalmente após evidências demonstradas a respeito da ação protetora que essas citocinas exercem sobre o HIV (CHOE et al.47, 1996). A partir de então, estudos têm mostrado a participação de quimiocinas em diversos processos biológicos como embriogênese, desenvolvimento, angiogênese, hematopoiese, resposta imune e inflamação (BAGGIOLINI; DEWALD; MOSER48, 1997). Embora inicialmente as quimiocinas dos grupos CC e CXC tenham sido caracterizadas como atrativos para monócitos/macrófagos e neutrófilos respectivamente (ROLLINS49, 1997), muitos estudos têm mostrado que não há uma correlação simples entre a estrutura primária de uma quimiocina e uma célula alvo específica (DIEU-NOSJEAN et al.45, 1999). O membro CCL3 (humano MIP-1α/LD78α) da família da proteína inflamatória para macrófago-1 (MIP-1) é decodificado por genes localizados no cromossomo 11 (MENTEN; WUYTS; VAN DAMME50, 2002). Células diretamente envolvidas na resposta imune como macrófagos, linfócitos T e B, neutrófilos, células dendríticas, 8 Introdução e Síntese Bibliográfica mastócitos e células NK (“Natural killer”) são capazes de produzir grandes quantidades de MIP-1. Plaquetas, osteoblastos, células epiteliais e fibroblastos, dentre outras, também o fazem, mas em quantidades mais discretas. A produção de MIP-1 pode ser induzida por diversos agentes pró-inflamatórios, LPS, substância P, infecções virais, TNF-α, IFN-γ, IL-1α/β. Outras citocinas como IL-4 ou IL-10 podem levar a uma diminuição da produção de MIP-1 (MAURER; STEBUT51, 2004). Esta quimiocina participa tanto da fase aguda como crônica da inflamação por meio do recrutamento de células pro-inflamatórias ao local de dano ou infecção. MIP-1α/CCL3 exerce quimiotaxia para monócitos (migração transendotelial), linfócitos B, neutrófilos ativados (BONECCHI et al.52, 1999) e eosinófilos (MAURER; VON STEBUT51, 2004). É capaz de atrair ainda linfócitos T CD4+ e CD8+ tanto sob fenótipo de memória como naive. (SCHALL et al.43, 1993). As evidências mostradas pela literatura constatam que MIP-1α/CCL3 desempenha uma função muito importante na indução e modulação da resposta imune e inflamatória (MAURER; VON STEBUT51, 2004). O fator derivado de estroma ou SDF-1/CXCL12 é uma quimiocina produzida constitutivamente por muitos tecidos distintos (BLEUL et al.53, 1996). O gene que a codifica, sdf1, está localizado no cromossomo 10 (SHIROZU et al.54, 1995). Este gene é traduzido como isoformas α e β que diferem em seu Cterminal. No entanto, não há indícios de quaisquer diferenças no que tange a expressão ou função. SDF-1/CXCL12 é quimiotática para leucócitos incluindo neutrófilos, monócitos, linfócitos B e T e células progenitoras hematopoiéticas (BLEUL et al.53, 1996, AIUTI et al.55, 1997, CHAN; HYDUK; CYBULSKY56, 2001). SDF-1/CXCL12 parece contribuir para o retorno de neutrófilos senescentes para a medula uma vez que estas células se mostram ainda mais responsivas a esta quimiocina quando idosas (MARTIN et al.57, 2003). Células dendríticas respondem a um amplo espectro de quimiocinas incluindo MIP-1α/CCL3 e SDF-1/CXCL12. No entanto, com a maturação, estas células perdem responsividade para muitas quimiocinas, mas não para SDF-1/CXCL12 (DIEU et al.58, 1998). Existem alguns indícios na literatura de que pode haver diminuição da produção de SDF1/CXCL12 durante o processo inflamatório (FEDYK et al.59, 2001, HOSOKAWA et al.33, 2005). A produção constitutiva de SDF-1/CXCL12 foi primeiramente constatada em células do estroma de medula óssea (AIUTI et al.55, 1997). No entanto, a produção desta quimiocina já pode ser demonstrada por fibroblastos sinoviais da articulação têmporo- Introdução e Síntese Bibliográfica 9 mandibular humana (OGURA et al.44, 2005) e por fibroblastos de gengiva humana (FEDYK et al.59, 2001, HOSOKAWA et al.33, 2005). Fibroblastos, imunidade inata e inflamação A participação dos fibroblastos na resposta imune inata tem sido demonstrada com base em achados acerca de receptores vinculados ao reconhecimento de antígenos bacterianos. A expressão de CD14 (agrupamento de diferenciação - proteína expressa na superfície de algumas células do sistema imune, envolvida no reconhecimento de LPS segundo WRIGHT et al.60, 1990) por fibroblastos de gengiva humana foi demonstrada por WANG et al. 61 , 1998. Estas células são capazes de se ligarem ao LPS de Escherichia coli e Porphyromonas gingivalis. A produção de IL-6 se mostrou dependente de CD14. Em fibroblasto de gengiva de camundongos, o CD14 só se mostrou expresso após estímulo por LPS (BOTERO et al.62, 2006). HATAKEYAMA et al.63, 2003, mostraram presença expressiva de CD14 em fibroblastos de gengiva humana contrastando com a presença discreta deste receptor em fibroblastos de ligamento periodontal de um mesmo indivíduo. Fibroblastos também parecem expressar uma classe de receptores denominados tipo “Toll-like receptors”(TLR) – receptores do tipo Toll (MAHANONDA et al.64, 2007, UEHARA; TAKADA34, 2007). Estes receptores são mediadores do reconhecimento inicial de patógenos microbianos, o que dá início à resposta imune inata. Até o presente, são conhecidos 12 membros da família TLR (AKIRA; UEMATSU; TAKEUSHI65, 2006). Estes receptores são expressos de maneira diferente pelas células imunes e parecem responder a estímulos diferentes (AKIRA; TAKEDA; KAISHO66, 2001). Entretanto, a expressão destes receptores vem sendo observada em outros grupos de células incluindo fibroblastos gengivais (MAHANONDA et al.64, 2007), fibroblastos de ligamento periodontal (HATAKEYAMA et al.63, 2003) e odontoblastos (DURAND et al.67, 2006). A expressão de TLR4, receptor para LPS, foi relatada em fibroblastos de gengiva humana por WANG et al.32, 2000. Este receptor pareceu influenciar a produção de IL-1 induzida por LPS. Novamente, porém em fibroblastos de gengiva de camundongos, BOTERO et al. 62, 2006, mostram o aumento da expressão de TLR4, assim como a de CD14, como já mencionado acima. Em estudos mais amplos com fibroblastos de gengiva humana, MAHANONDA et al.64, 2007 e 10 Introdução e Síntese Bibliográfica UEHARA; TAKADA34, 2007, observaram uma grande variedade de receptores relacionados ao reconhecimento de microrganismos. MAHANONDA et al.64, 2007, demonstraram a expressão de TLR1, TLR2, TLR3, TLR4, TLR5, TLR6 e TLR9 além da relevância funcional de TLR2, 3, 4 e 5 que tiveram envolvimento na produção de IL8 pelos fibroblastos. UEHARA; TAKADA34, 2007, observaram a expressão de TLR1, 2, 3, 4, MD-2, TLR5, 6, 7, 8 e 9, MyD88, NOD1 e NOD2. O desafio com agonistas de TLR2/6, TLR3, TLR4, TLR7/8, TLR9, NOD1 e NOD1 levou as células a produzirem IL-6, IL-8 e MCP-1. Estes achados suportam o papel dos fibroblastos na resposta imune inata contra bactérias. As diversas respostas dos fibroblastos frente a estímulos de origem microbiana vêm sendo elucidadas de maneira bastante variada pela literatura. Dentre os desafios escolhidos para o estudo destas células no contexto de imunologia e inflamação, encontram-se: sobrenadante de bactérias (LETZELTER et al.68, 1998, YANG et al.36, 2003, YANG et al.37, 2003, YANG et al.39, 2004), bactéria íntegra inativada (SUGIYAMA et al.31, 2000, HOSOKAWA et al.33, 2005), extrato sonicado de bactérias (YAMASAKI et al.69, 1998), estruturas bacterianas isoladas como vesícula extracelular e proteína externa de membrana (SUTHIN et al.70, 2003), fímbria (SUGIYAMA et al.31, 2000) e peptidoglicana (HATAKEYAMA et al.63, 2003). Têm sido amplamente estudados também os componentes de parece celular como o ácido lipoteicóico (LTA) (SUGIYAMA et al.71, 1996, WANG et al.72, 2001, TELLES et al.73, 2003), proveniente de bactérias gram-positivas; e o lipopolissacarídeo (LPS) (SISMEY-DURRANT; HOPPS27, 1991, TAMURA et al.28, 1992, YAMAJI et al.29, 1995, WANG et al.61, 1998, WANG et al.30, 1999, WANG et al.32, 2000, TOKUDA; NAGAOKA; TORII35, 2002, BOTERO et al.74, 2003, SUTHIN et al.70, 2003, COIL; TAM; WATERFIELD38, 2004, TARDIF; ROSS; ROUABHIA75, 2004, GUTIERREZ-VENEGAS et al.76, 2005, HOSOKAWA et al.77, 2005, BOTERO et al.62, 2006, MAHANONDA et al.64, 2007, UEHARA; TAKADA34, 2007) proveniente de bactérias gram-negativas. Além de agentes microbianos, diversas citocinas têm sido utilizadas como estímulo a fibroblastos: IL-10 (WANG et al.30, 1999, TOKUDA; NAGAOKA; TORII35, 2002), IL1β (TAKASHIBA et al.78, 1992, SUGIYAMA et al.71, 1996), IL-1α (SUGIYAMA et al.31, 2000, FEDYK et al.59, 2001), TNF-α (TAKASHIBA et al.78, 1992, FEDYK et al.59, 2000, TOKUDA, NAGAOKA, TORII35, 2002, HOSOKAWA et al.33, 2005, OGURA et al.44, 2005, MAHANONDA et al.64, 2007,) e IFN-γ (TAMURA et al.28, 11 Introdução e Síntese Bibliográfica 1992, HOSOKAWA et al.33, 2005, MAHANONDA et al.64, 2007). Com todos os estímulos acima citados, fibroblastos oriundos de diferentes tecidos demonstraram alguma resposta relevante para o processo inflamatório dos mesmos. A toxicidade bacteriana a fibroblastos no que concerne à proliferação destas células em cultura, foi demonstrada por LETZELTER et al.68, 1998, com fibroblastos de gengiva humana que tiveram seu crescimento diminuído com o sobrenadante de Prevotella nigrescens, Capnocytophaga ochracea , Aggregatibacter actinomycetemcomitans e Peptostreptococus micros. A mesma inibição pôde ser observada por YAMASAKI et al.69, 1998, em fibroblastos periapicais quando estimulados pelo extrato sonicado de Porphyromonas endodontalis, Porphyromonas gingivalis e Fusobacterium nucleatum. De forma contrária, o extrato sonicado de Prevotella intermedia levou ao estímulo do crescimento celular. Dentre os microrganismos estudados, como mais tóxicos se mostraram P. gingivalis e F. nucleatum. A capacidade de um componente de parede celular como o LTA de Streptococcus mutans em induzir apoptose foi demonstrado em células da polpa de dentes decíduos por WANG et al.72, 2001. No grupo tratado com LTA, os autores observaram degradação do DNA das células, típica do processo de apoptose. A literatura tem mostrado que os fibroblastos constituem um grupo de células capazes de produzir mediadores inflamatórios frente a estímulos variados. A produção do mRNA de VEGF foi mostrada com aumento por YANG et al.39, 2004 em fibroblastos de polpa humana estimulados pelo sobrenadante de P. endodontalis, P. gingivalis e P. intermedia. A produção desta mesma proteína por fibroblastos de tecido gengival murino, como elucidado por TELLES et al.73, 2003, foi encontrada de maneira constitutiva por estas células mas que não tiveram produção aumentada pelo LTA de Streptococcus sanguinis e S. mutans. Por outro lado, em fibroblastos de gengiva humana, SUTHIN et al.70, 2003, encontraram níveis significativamente mais altos de VEGF quando as células foram estimuladas por vesícula extracelular e proteína externa de membrana tanto de P. gingivalis como A.a. No entanto, o LPS de P.gingivalis não foi capaz de induzir aumento dos níveis basais de VEGF. A IL-6 é uma citocina amplamente estudada como resultado a estímulos variados. Sua produção por fibroblastos de ligamento periodontal humano foi demonstrada por YAMAJI et al.29, 1995, frente a desafio por LPS. Com este mesmo antígeno, WANG et al.30, 1999, e WANG et al.61, 1998, mostraram a produção da mesma citocina em fibroblastos de gengiva humana. No entanto, os autores enfatizam a 12 Introdução e Síntese Bibliográfica inibição desta produção pelo tratamento conjunto com IL-10 (WANG et al.30, 1999) e com anticorpo anti-CD14 (WANG et al.61, 1998). Os fibroblastos de polpa dental humana também foram capazes de expressar mRNA para IL-6 frente ao desafio por sobrenadante de bactérias gram-negativas conforme YANG et al.37, 2003, e a produção desta mesma citocina foi confirmada por COIL; TAM; WATERFIELD38, 2004, especialmente se estimuladas por LPS, quando a produção foi significativamente aumentada. A produção de IL-1β também tem sido demonstrada por diversos estudos. A produção desta citocina por fibroblastos oriundos de periósteo de rato foi estudada por SISMEY-DURRANT; HOPPS27, 1991. Os níveis basais de IL-1β eram baixos, mas o estímulo com LPS de P. gingivalis levou a um aumento de até 6 vezes. TARDIF; ROSS; ROUABHIA75, 2004, compararam a produção de IL-1β por fibroblastos de derme e gengiva humanos. Frente ao desafio com LPS de Escherichia coli, ambos os grupos produziram a citocina, porém as células da derme em níveis mais elevados. Também em fibroblastos de gengiva humana, TAMURA et al.28, 1992, já haviam mostrado a produção de IL-1β porém de maneira discreta. Fibroblastos de polpa dental humana em cultura originados de tecido inflamado produziram níveis maiores de IL-1β do que as células provenientes do tecido saudável segundo observado por BARKHORDAR et al.26, 2002. Outras citocinas, cuja produção já foi relatada como resposta de fibroblastos a diferentes estímulos, podem ser citadas. O fator de crescimento para hepatócito (HGF) teve sua produção basal aumentada em fibroblastos de gengiva humana estimulados por LTA de S. sanguinis e E. faecalis. O estímulo concomitante com IL-1β levou ao aumento na produção deste fator de maneira sinergística com LTA de S. sanguinis (SUGIYAMA et al.71, 1996). Em estudo mais recente, SUGIYAMA et al.31, 2000, demonstraram novamente nestas células o aumento de HGF induzido por P. gingivalis (bactéria íntegra e inativada) e por fímbria deste mesmo microrganismo. O aumento de HGF também foi observado quando do tratamento destes fibroblastos com IL-1α. A produção de TGF-β por fibroblastos de ligamento periodontal humano foi demonstrada por YAMAJI et al.29, 1995. A produção constitutiva desta citocina não foi afetada por estímulo com LPS de P. gingivalis e de E. coli. TAMURA et al.28, 1992, mostraram produção de IL-1α por fibroblastos de gengiva humana estimulados por LPS de P. intermedia e P. gingivalis. Introdução e Síntese Bibliográfica 13 A capacidade de fibroblastos em produzirem quimiocinas também vem sendo estudada de forma variada. TAMURA et al.28, 1992, mostraram aumento da expressão de mRNA para IL-8 (CXCL8) por fibroblastos de gengiva humana após desafio com LPS de P. intermedia. Quando as células eram tratadas previamente com IFN-γ, a expressão desta quimiocina foi ainda maior. Em fibroblastos de mesma origem, TAKASHIBA et al.78, 1992 observaram o aumento da expressão de mRNA para IL8/CXCL8 após tratamento das células com IL-1β e TNF-α. O estudo também demonstra a capacidade destes fibroblastos em induzir a migração de neutrófilos. Ao estudar fibroblastos de ligamento periodontal humano perante estímulo por LPS de P. gingivalis e E. coli, YAMAJI et al.29, 1995, também observaram aumento de IL8/CXCL8 como resposta ao desafio. O aumento da expressão do mRNA para IL8/CXCL8 por fibroblastos de polpa dental humana foi observado por YANG et al.36, 2003, ao estimularem as células com sobrenadante de P. endodontalis, P. gingivalis e P. intermedia. A produção de IL-8/CXCL8 por fibroblastos pulpares humanos já havia sido observada por TOKUDA et al.35, 2002. Quando tratadas previamente com IL-10, as células estimuladas por LPS de P. intermedia, IL-1β e TNF-α tiveram inibidas a expressão de mRNA para IL-8/CXCL8 induzida por estes estímulos. Em fibroblastos sinoviais da articulação temporo-mandibular (ATM) humana, OGURA et al.44, 2005 observaram a produção de IL-8/CXCL8 e de outras quimiocinas como GRO-α/CXCL1, MCP-1/CCL2 e RANTES/CCL5 significativamente aumentadas pelo tratamento com TNF-α. MAHANONDA et al.64, 2007, e UEHARA; TAKADA34, 2007, observaram também, em fibroblastos de gengiva humana, o aumento da produção de IL-8/CXCL8 e IDO (molécula imunossupressora) e de IL-8/CXCL8, MCP-1/CCL2 e IL-6, respectivamente, como resposta ao desafio com diferentes agonistas de TLRs. A produção de CXCL12 (SDF-1α/β) foi demonstrada em fibroblastos de gengiva e derme por FEDYK et al.59, 2001. A produção constitutiva se deu em níveis altos, mas foi completamente inibida pelo tratamento das células com IL-1α e TNF-α. Por outro lado, nas mesmas condições, os autores observaram aumento do mRNA para IL-8/CXCL8 e MCP-1/CCL2. Também em fibroblastos de gengiva humana, HOSOKAWA et al.33, 2005, observaram a produção constitutiva de CXCL12, que por sua vez foi aumentada com estímulo por TNF-α, IFN-γ, TGF-β, RANTES/CCL5, MIP3α/CCL20 e fractalquina/CX3CL1, além de LPS de E. coli e CpGDNA. De maneira contrastante, o LPS de P. gingivalis diminuiu esta produção. O fenômeno se confirma 14 Introdução e Síntese Bibliográfica quando P. gingivalis na forma íntegra morta por calor foi utilizada como estímulo: enquanto a produção de IL-8/CXCL8 aumentou, a de CXCL12 diminuiu. Com relação à produção de MIP-1α/CCL3, OGURA et al.44, 2005, relatam presença tão discreta desta quimiocina no sobrenadante das células em cultura, que chega a estar próxima ao nível limite de detecção em fibroblastos de ATM humana. Os trabalhos buscados na literatura mostram ser cada vez mais evidente o fato de que fibroblastos constituem uma família heterogênea de células. Diversos relatos têm documentado diferenças tanto em características fenotípicas como funcionais em fibroblastos originados em diferentes sítios anatômicos (BROUTY-BOYE et al.79, 2000, HATAKEYAMA et al.63, 2003). A produção de quimiocinas por fibroblastos de tecido hematopoiético da medula óssea, baço mielometaplástico, pulmão fetal e adulto e tecido brônquico distal a tumor, estroma metastático de melanoma de pulmão, tecido de mama e carcinoma de mama foi estudada por BROUTY-BOYÉ et al.79, 2000, comparando as células de acordo com o tecido de origem. Uma grande, porém variável quantidade de IL-8/CXCL8, SDF-1/CXCL12, MCP-1/CCL2 e eotaxina/CCL11 foram analisadas por RT-PCR. Ao analisar as respostas de fibroblastos provenientes da gengiva e do ligamento periodontal, HATAKEYAMA et al.63, 2003, mostraram que num mesmo indivíduo, os fibroblastos gengivais expressaram mais CD14 do que os fibroblastos de ligamento periodontal. Os fibroblastos de gengiva também produziram quantidades mais elevadas de IL-8/CXCL8 quando estimulados por LPS de Salmonella enterica do que as células do ligamento periodontal, que de uma maneira geral mostraram-se mais responsivas ao estímulo por peptidoglicana e muramilpeptídeo (componentes da parede celular de bactérias gram-positivas). A expressão de TLR2 se mostrou mais intensa nos fibroblastos de ligamento periodontal enquanto a de TLR4 foi muito semelhante para ambas células. Estes achados conjuntos contribuem para o crescente interesse do estudo de fibroblastos oriundos de diferentes regiões anatômicas incluindo os microambientes existentes na cavidade bucal. Neste contexto, o comportamento dos fibroblastos pulpares dentro da resposta imune inata e inflamatória se torna necessário e justificável. A relevância dos fibroblastos no contexto de inflamação pulpar A polpa dental vem sendo objeto de estudo não apenas pelas implicações clínicas dos seus processos patológicos, mas também tem-se observado notável interesse por este tecido devido a fatores específicos como sua localização interna a um Introdução e Síntese Bibliográfica 15 tecido duro com paredes inflexíveis, seu suprimento vascular particular e dinâmica celular, que juntos levam a um interesse crescente pela resposta que a polpa exerce a agressões, especialmente infecções bacterianas (GOODIS2, 2002). A literatura tem elucidado o comportamento dos odontoblastos na resposta imune inata da polpa como primeira linha de defesa no processo de cárie tanto com relação à capacidade de reconhecimento de antígeno como de recrutamento de células de defesa por meio da produção de quimiocinas (BOTERO et al.74, 2003, BOTERO et al.62, 2006, DURAND et al.67, 2006). No entanto, pelo fato dos fibroblastos serem as células mais numerosas a povoarem o tecido pulpar, a sua participação na resposta imune e inflamação deste tecido podem exercer um papel de fundamental importância (CHIEGO JR1, 2002). Além disso, a literatura apresenta evidências inequívocas de que fibroblastos não constituem um grupo único de células, mas sim um grupo com uma vasta variedade em relação à forma, proliferação, expressão de receptores de membrana, função, além de outras características (BROUTY-BOYE et al.79, 2000, HATAKEYAMA et al.63, 2003. Tendo em vista que em processos inflamatórios, de maneira geral, as quimiocinas exercem papel fundamental no recrutamento de células de defesa, torna-se também de interesse significativo o estudo da liberação delas por fibroblastos da polpa mediante agressão por microrganismos freqüentemente vinculados a infecções endodônticas como no caso do Enterococcus faecalis. O estudo da possibilidade de recrutamento de células de defesa (produção de quimiocinas) por fibroblastos dentro do contexto de inflamação pulpar torna-se verdadeiramente necessário, uma vez que os fenômenos da inflamação não ocorrem da mesma maneira em todos os sítios preenchidos por tecido conjuntivo (RAVANTI et al.80, 1999, BARKHORDAR et al.26, 2002, MARTINEZ; ARAUJO81, 2004). 16 Introdução e Síntese Bibliográfica Proposição 19 2 Proposição Justificativa: devido à importância dos fenômenos presentes na inflamação do tecido pulpar, existe a necessidade de conhecê-los em detalhes uma vez que a inflamação não ocorre por igual em todos os tecidos conjuntivos do organismo. Pela predominância numérica dos fibroblastos no tecido pulpar e pelos recentes relatos acerca da capacidade destas células em responderem a estímulos externos por meio da liberação de diversas citocinas, dentre elas quimiocinas, que por sua vez desempenham importante função na formação do infiltrado inflamatório, o presente trabalho teve como objetivos: Avaliar a capacidade de fibroblastos de polpa dental humana em cultura quanto à produção das quimiocinas MIP-1α/CCL3 e SDF-1/CXCL12. Avaliar a liberação das quimiocinas MIP-1α/CCL3 e SDF-1/CXCL12 por fibroblastos de polpa dental humana em cultura quando estimulados por Enterococcus faecalis inativado por calor, em diferentes quantidades de bactérias por fibroblasto e em diferentes períodos de estimulação. Material e Métodos 23 3 Material e Métodos 3.1 Coleta do Tecido Pulpar O dente recentemente extraído (terceiro molar hígido) de um paciente livre de doenças sistêmicas foi obtido no Laboratório de Fisiologia e Farmacologia Clínica da FOB/USP. O dente foi doado pelo paciente em formulário específico devidamente aprovado pelo Comitê de Ética em Pesquisa da FOB/USP (Processo no 131/2005). Após remoção do ligamento periodontal e desinfecção do dente com solução de gluconato de clorexidina 0,12% (Colgate – Palmolive Ind. e Com. LTDA, São Paulo, Brasil), o armazenamento foi feito em soro fisiológico até sua utilização para a extração do tecido pulpar. Um sulco transversal com ponta diamantada (#2200, KG Sorensen, São Paulo, Brasil) foi realizado na região da junção amelocementária em toda a circunferência do dente. Com a ajuda de dois alveolótomos, o dente foi fraturado e aberto em condições assépticas, removendo-se assim sua polpa, a qual foi imediatamente imersa em meio de cultura Dulbecco´s modified Eagle´s medium (DMEM) (Gibco, Invitrogen Corporation, Califórnia, EUA) e acondicionada em placa de Petri de 100x10 mm (diâmetro x altura). 3.2 Cultura dos Fibroblastos Os tecidos provenientes da polpa foram picotados com lâmina de bisturi no 15 (BD, São Paulo, Brasil) permanecendo imersos em meio de cultura DMEM suplementado com soro bovino fetal 10% (Cultilab, Campinas, Brasil), penicilina 100 µg/mL, estreptomicina 100µg/mL e anfotericina B 0,5 mg/mL (Invitrogen, Califórnia, EUA). Os fragmentos de polpa foram centrifugados e armazenados em novo meio de cultura e mantidos em estufa a 37ºC e a 5% CO2. As culturas foram mantidas até os fibroblastos alcançarem confluência com troca de meio a cada 2-3 dias. Após confluência, os fibroblastos foram tripsinizados e a utilização das células ocorreu entre a 4a e a 8a passagens. O treinamento para a obtenção de cultura primária de fibroblastos a partir de tecidos bucais foi realizado no Laboratório de Farmacologia da Faculdade de Odontologia de Araçatuba (UNESP) sob orientação da Profa Dra Sandra Helena Penha de Oliveira. 24 Material e Métodos 3.3 Preparo do Enterococcus faecalis morto por calor A cepa de Enterococcus faecalis ATCC 29212 foi obtida do Laboratório de Microbiologia e Imunologia, Departamento de Ciências Biológicas, da FOB/USP. Os microrganismos foram reativados em meio BHI (brain heart infusion) (Difco, Detroit, MI, USA) até atingirem crescimento exponencial. Os tubos de ensaio contendo as bactérias foram aquecidos a 80ºC por 30 minutos conforme protocolo descrito por Noel et al.82, 1995. Uma alíquota de 100µL foi semeada em BHI estéril para comprovar a eficácia do processo. Estas culturas foram centrifugadas a 4.000 rpm por 20min em temperatura ambiente e lavadas por 2 vezes em PBS (fosfato tamponado em salina), ressuspensas em meio DMEM e diluídas para o uso. Ainda em PBS, foi realizada a contagem das bactérias com o auxílio de um espectrofotômetro (Ultrospec 1000, Pharmacia Biotech, Westerville, EUA) no comprimento de onda de 530nm tomando como curva padrão a escala de Mc Farland (BIER83, 1985). 3.4 Estimulação dos Fibroblastos Pulpares por Enterococcus faecalis morto por calor Os fibroblastos foram distribuídos em placas de 24 poços na concentração de 5 x 104 células por poço e deixados para aderirem até atingirem confluência. As células foram mantidas em incubadora a 37ºC com 5% de CO2. O meio de cultura contendo E. faecalis morto na proporção de 1:1, 10:1 e 100:1 bactérias:fibroblasto foi adicionado às placas em duplicata. Após 1, 6 e 24 h, o sobrenadante das células foi coletado e armazenado para a dosagem de quimiocinas por ensaio imunoenzimático (ELISA). 3.5 ELISA (Enzyme Linked Immuno Sorbant Assay) A produção de MIP-1α/CCL3e SDF-1/CXCL12 extracelular foram quantificadas usando-se o método de ensaio imunoenzimático (ELISA). Placas de 96 poços foram recobertas e incubadas durante 15-18h a 4ºC, com anticorpos anti-MIP-1α (Anti-human CCL3/MIP-1α Antibody – AF-270-NA – R&D Systems, Minneapolis, EUA) ou anti-SDF-1 (Monoclonal Anti-human/mouse CXCL12/SDF-1 Antibody – MAB 350 – R&D Systems, Minneapolis, EUA) diluídos em tampão de fosfato de cálcio. Após o tempo de incubação, as placas foram lavadas com PBS, contendo Tween 20 0,5% e bloqueadas durante 1 hora, em temperatura ambiente, com PBS contendo soroalbumina bovina 1%. As placas, então, foram lavadas com PBS e incubadas com os sobrenadantes das células ou com quantidades conhecidas de MIP-1α/CCL3 ou SDF- Material e Métodos 25 1/CXCL12 quimiocinas recombinantes (R&D Systems: MIP-1α - 270-LD-010; SDF1α - 350-NS, Minneapolis, EUA) (curvas-padrão) durante 1 h a 40C. Após este período, as placas foram lavadas com PBS e incubadas com anticorpo biotinilado (Biotinylated Anti-human CCL3/MIP-1α Antibody – BAF 270 - R&D Systems; Biotinylated Antihuman CXCL12/SDF-1 Antibody – BAF 310 – R&D Systems, Minneapolis) por 30 minutos a 370C. As placas foram lavadas com PBS e o substrato (peróxido de hidrogênio e tetrametilbenzidina na proporção de 1:1) adicionado conforme as instruções do fabricante. Após 30 minutos, a solução de paralisação da reação (ácido sulfúrico 2N) foi adicionada e a leitura foi realizada em espectrofotômetro ajustado para o comprimento de onda de 490 nm. A obtenção da cultura primária de fibroblastos e sua manutenção, bem como a manipulação do microrganismo e ainda a execução dos experimentos de estímulo dos fibroblastos e ELISA foram realizados no Laboratório de Microbiologia e Imunologia, Departamento de Ciências Biológicas, da FOB/USP sob orientação da Profa Dra Ana Paula Campanelli. 3.6 Análise Estatística A análise estatística foi realizada por meio do teste Kruskal-Wallis seguido de pós-teste de Miller. Foi adotado nível de significância de 5% para que as diferenças fossem consideradas estatisticamente significativas. Para esta análise, contamos com a colaboração do Prof. Dr. José Roberto Pereira Lauris, docente da disciplina de Bioestatística da FOB/USP. Este trabalho contou com financiamento da Fundação de Amparo à Pesquisa do Estado de São Paulo por meio de auxílio a pesquisa (Processo FAPESP 05/60167-0) em nome do orientador Prof. Dr. Carlos Ferreira dos Santos. 26 Material e Métodos Resultados 29 4 Resultados 4.1 Cultura dos fibroblastos A técnica de cultura primária de fibroblastos a partir da extração do tecido proveniente de polpa dental humana se mostrou passível de reprodução durante a execução deste trabalho. Para tanto, os protocolos experimentais foram adaptados para que fosse viável a estabilização da cultura de fibroblastos a partir de tecido humano, ao invés do uso de tecido proveniente de rato ou camundongo, que representam espécies tastante utilizadas para experimentos desta natureza. Adaptações também foram necessárias para que a extração da polpa fosse realizada de maneira asséptica e a tempo das células não sofrerem necrose após a extração dos dentes. Seguem, a seguir, fotografias dos fibroblastos pulpares de acordo com o tempo de cultura (visualização em microscópio óptico invertido). 30 Resultados Resultados 31 Figura 1: Cultura de fibroblastos de polpa dental humana. A polpa dental de um terceiro molar foi utilizada para a obtenção de cultura primária de fibroblastos. Em a, b e c, observam-se células com 15 dias de cultura, antes de formarem confluência. Em d, e e f, fibroblastos com 30 dias de cultura. Em f, pode ser observado aspecto de confluência. Em g, h e i, fibroblastos de polpa dental humana com 60 dias de cultura após a 4a passagem. Em i, pode ser observada a ocorrência de mitose (seta). Visualização em microscópio óptico invertido com aumento de 100x em a, b, d, e e f, e de 320x em c, g, h e i. 32 Resultados 33 Resultados 4.2 Produção de MIP-1α/CCL3 por ELISA A produção de MIP-1α/CCL3 pelas células em cultura foi detectada por ELISA a partir da análise do sobrenadante das células em cultura. Os fibroblastos de polpa dental humana foram estimulados por Enterococcus faecalis morto por calor nas proporções de 1 bactéria por fibroblasto (EF1:1), 10 bactérias por fibroblasto (EF10:1) e 100 bactérias por fibroblasto (EF100:1) nos tempos experimentais de 1, 6 e 24 h. Foram utilizadas como controle células não submetidas a estímulo (CLE). A produção de MIP1α/CCL3 pelos fibroblastos pulpares pôde ser constatada por ELISA e a resposta dos fibroblastos frente a este estímulo variou de acordo com sua concentração (Figuras 2 e 3). Enquanto no grupo controle a produção de quimiocina parece se acumular no meio de cultura ao longo das 24 h, nas concentrações de 1:1 e 100:1 bactérias:fibroblasto, a produção maior ocorreu logo na primeira hora de estímulo, diminuindo nos períodos seguintes, e chegando a zero no caso da concentração mais elevada de bactérias. Com o estímulo de 10:1 bactérias:fibroblasto, a produção desta quimiocina aumentou gradativamente ao longo do período experimental. A análise estatística por meio do teste Kruskal-Wallis, entretanto, não mostrou diferença estatisticamente significativa entre os grupos. As médias e erros-padrão seguem na tabela abaixo: Tabela 1: Médias e erros-padrão das médias (média ± EPM) referentes aos valores de produção de MIP-1α/CCL3 por fibroblastos de polpa dental humana estimulados por Enterococcus faecalis inativado por calor nas proporções de 1:1, 10:1 e 100:1 bactérias:fibroblasto nos tempos experimentais de 1, 6 e 24 h. Valores de controle (CLE) são relativos ao grupo sem estímulo (n=3). 1h 6h 24 h CLE 2,55 ± 1,84 1,49 ± 1,23 7,19 ± 0,84 EF1:1 8,48 ± 0,93 1,43 ± 0,08 2,48 ± 2,31 EF10:1 0,94 ± 0,32 2,04 ± 2,04 6,28 ± 3,16 EF100:1 7,25 ± 3,62 4,23 ± 4,23 0±0 As diferenças não foram consideradas estatisticamente significativas pelo teste Kruskal-Wallis (p>0,05). 34 Resultados 35 Produção de MIP-1alfa (pg/mL) Resultados 10.0 1h 6h 24 h 7.5 5.0 2.5 0.0 CLE EF 1:1 EF 10:1 EF 100:1 Produção de MIP-1alfa (pg/mL) Figura 2: Produção de MIP-1α/CCL3 por fibroblastos de polpa dental humana estimulados por Enterococcus faecalis inativado por calor. Cultura primária de fibroblastos foi obtida utilizando-se a polpa de um terceiro molar hígido. Após a 4a passagem, estas células foram estimuladas por E. faecalis morto por calor em concentrações de 1:1, 10:1 e 100:1 bactérias:fibroblasto. Como controle, foi colocado sobre as células o meio de cultura sem estímulo (CLE). A análise do sobrenadante por ELISA mostrou que fibroblastos de polpa dental humana produzem MIP-1α. Os resultados apresentados mostram aumento da produção desta quimiocina ao longo das 24h nos grupos controle (CLE) e 10:1 bactérias:fibroblasto. Com a adição de E. faecalis nas proporções 1:1 e 100:1 bactérias:fibroblasto, observa-se tendência de aumento de produção de MIP-1α logo na 1a hora de estímulo. O teste Kruskal-Wallis não revelou diferenças significativas entre os grupos experimentais (n=3). 15 10 5 0 CLE CLE CLE EF1:1 EF1:1 EF1:1 1h 6h 24h 1h 6h 24h EF10:1 EF10:1 EF10:1 EF100:1 EF100:1 EF100:1 1h 6h 24h 1h 6h 24h Concentração de bactéria e tempo Figura 3: Valores individuais da produção de MIP-1α/CCL3 por fibroblastos de polpa dental humana estimulados por Enterococcus faecalis inativado por calor. A detecção de MIP-1α produzida pelos fibroblastos ocorreu de forma bastante heterogênea mesmo dentro do mesmo grupo. Grupos controle (CLE) relativos ao grupo experimental sem estímulo. EF1:1, EF10:1 e EF100:1: grupos experimentais com adição de estímulo nas proporções de 1:1, 10:1 e 100:1 bactérias:fibroblasto, respectivamente. Tempos experimentais de 1, 6 e 24h de contato dos fibroblastos com o meio de cultura com ou sem estímulo (n=3). 36 Resultados 37 Resultados 4.3 Produção de SDF-1/CXCL12 por ELISA A produção de SDF-1/CXCL12 também foi avaliada por ELISA. Os resultados mostram que fibroblastos pulpares em cultura são capazes de produzir esta quimiocina. Constitutivamente, a produção se mostrou elevada no período experimental de 24 horas, mas não pôde ser detectada nos períodos anteriores. O estímulo com Enterococcus faecalis inativado por calor causou aumento da produção de SDF1/CXCL12 na concentração de 1:1 bactéria:fibroblasto no período de 6 h. Ao analisar a relação número de bactérias com a resposta no período experimental de 24 h, observouse diminuição estatisticamente significativa da produção desta quimiocina com o aumento da concentração de estímulo bacteriano (p = 0,000655) (Figuras 4 e 5) Tabela 1: Médias e erros-padrão das médias (média ± EPM) referentes aos valores de produção de SDF-1/CXCL12 por fibroblastos de polpa dental humana estimulados por Enterococcus faecalis inativado por calor nas proporções de 1:1, 10:1 e 100:1 bactérias:fibroblasto nos tempos experimentais de 1, 6 e 24 h. Valores de controle (CLE) são relativos ao grupo sem estímulo (n=4). 1h 6h 24 h CLE 0±0 0±0 173,00 ± 6,35* EF1:1 0±0 40,50 ± 23,38 159,00 ± 17,03# EF10:1 0±0 0±0 22,00 ± 12,70 EF100:1 0±0 0±0 14,00 ± 0& Diferenças consideradas estatisticamente significativas pelo teste Kruskal-Wallis: * com relação aos tempos correspondentes à ausência de estímulo (CLE) e ainda com relação às concentrações 10:1 e 100:1 no mesmo período experimental; # com relação ao período de 1h da mesma concentração de microrganismo(EF1:1) e ainda com relação às concentrações EF10:1 e EF100:1 no mesmo período experimental; e & com relação aos demais períodos avaliados na mesma concentração de E. faecalis. 38 Resultados 39 Produção de SDF-1 (pg/mL) Resultados 200 * 1h 6h 24 h # 100 & 0 CLE EF 1:1 EF 10:1 EF 100:1 Produção de SDF-1 (pg/mL) Figura 6: Produção de SDF-1/CXCL12 por fibroblastos de polpa dental humana estimulados por Enterococcus faecalis inativado por calor. Cultura primária de fibroblastos foi obtida utilizando-se a polpa de um terceiro molar hígido. Após a 4a passagem, estas células foram estimuladas por E. faecalis morto por calor em concentrações de 1:1, 10:1 e 100:1 bactérias:fibroblasto. Como controle, foi colocado sobre as células o meio de cultura sem estímulo (CLE). A análise do sobrenadante por ELISA mostrou que fibroblastos pulpares humanos produzem SDF-1 constitutivamente. O estímulo com E. faecalis levou à diminuição da produção desta quimiocina conforme o aumento da concentração de bactérias no período experimental de 24h. Diferenças significativas foram encontradas em * com relação aos tempos correspondentes à ausência de estímulo e ainda com relação às concentrações 10:1 e 100:1 no mesmo período experimental; # com relação ao período de 1h da mesma concentração de microrganismo e ainda com relação às concentrações EF10:1 e EF100:1 no mesmo período experimental; e & com relação aos demais períodos avaliados na mesma concentração de E. faecalis (n=4). 200 100 0 CLE CLE CLE EF1:1 EF1:1 EF1:1 EF10:1 EF10:1 1h 6h 24h 1h 6h 24h 1h 6h EF10:1 EF100:1 EF100:1 EF100:1 24h 1h 6h 24h Concentração de bactéria e tempo Figura 7: Valores individuais da produção de SDF-1/CXCL12 por fibroblastos de polpa dental humana estimulados por Enterococcus faecalis inativado por calor. A detecção de SDF1 produzida pelos fibroblastos ocorreu de forma marcante no período de 24 horas. Com a colocação e aumento da quantidade de bactérias como estímulo, esta produção diminuiu de forma significativa segundo teste Kruskal-Wallis. Grupos controle (CLE) relativos ao grupo experimental sem estímulo. EF1:1, EF10:1 e EF100:1: grupos experimentais com adição de estímulo nas proporções de 1:1, 10:1 e 100:1 bactérias:fibroblasto, respectivamente. Tempos experimentais de 1, 6 e 24h de contato dos fibroblastos com o meio de cultura com ou sem estímulo (n=4). 40 Resultados Discussão 43 5 Discussão Os fibroblastos são células que povoam o centro do tecido pulpar e embora não sejam a primeira linha de defesa deste tecido, como os odontoblastos (DURAND et al.67, 2006), são as células mais numerosas deste microambiente (CHIEGO JR1, 2002). Sendo inegável a importância do conhecimento acerca dos fenômenos imunológicos e inflamatórios que afetam o tecido pulpar, estudar o papel dos fibroblastos neste contexto torna-se fundamental para o crescimento da ciência na endodontia. Embora haja evidências inequívocas da participação dos fibroblastos de outros tecidos como derme (FEDYK et al.59, 2001, TARDIF; ROSS; ROUABHIA75, 2004), articulação têmporomandibular (OGURA et al.44, 2005), ligamento periodontal (YAMAJI et al.29, 1995, HATAKEYAMA et al.63, 2003) e gengiva (SISMEY-DURRANT; HOPPS27, 1991, TAKASHIBA et al.78, 1992, TAMURA et al.28, 1992, SUGIYAMA et al.71, 1996, WANG et al.61, 1998, WANG et al.30, 1999, SUGIYAMA et al.31, 2000, WANG et al.32, 2000, FEDYK et al.59, 2001, SUTHIN et al.70, 2003, TARDIF; ROSS; ROUABHIA75, 2004, GUTIERREZ-VENEGAS et al.76, 2005, HOSOKAWA et al.33, 2005, MAHANONDA et al.64, 2007, UEHARA; TAKADA34, 2007) nos fenômenos inflamatórios e imunológicos através da capacidade destas células em produzirem citocinas, dentre elas quimiocinas, além de outros agentes pró-inflamatórios, a capacidade destas células oriundas da polpa dental ainda não foi completamente elucidada (WANG et al.72, 2001, TOKUDA; NAGAOKA; TORII35, 2002, YANG et al.36, 2003, YANG et al.37, 2003, COIL; TAM; WATERFIELD38, YANG et al.39, 2004). Estudos com esse objetivo se justificam, uma vez que se considera consagrado o fato de que fibroblastos não constituem um grupo celular único entre diferentes regiões anatômicas (BROUTY-BOYE et al.79, 2000). Mais do que isso, fibroblastos oriundos de regiões diferentes parecem apresentar características fenotípicas também diferentes (PHIPPS et al.84, 1989) o que faz com que fibroblastos possam ser considerados em subgrupos de células (PHIPPS et al.84, 1989, BROUTY-BOYE et al.79, 2000), mesmo quando originados de diferentes tecidos da cavidade bucal (HATAKEYAMA et al.63, 2003). A escolha da utilização de Enterococcus faecalis como estímulo neste trabalho deveu-se principalmente à relevância que este microrganismo assume no contexto de 44 Discussão infecção em endodontia. O destaque que este microrganismo recebe se deve especialmente à freqüência com que ele é encontrado em lesões refratárias ao tratamento endodôntico (KAUFMAN et al.10, 2005, SEDGLEY et al.11, 2006, WILLIAMS et al.12, 2006, ZOLETTI; SIQUEIRA; SANTOS13, 2006). Isso se deve principalmente à capacidade deste microrganismo em resistir a situações de ambiente desfavorável ao seu crescimento (FLAHAUT et al.17, 1996). Dentre os fatores de virulência conhecidos destacam-se a substância de agregação ROZDZINSKI et al.18, 2001, e o ácido lipoteicóico (SIGNORETTO et al.22, 2000. A escolha por estruturas isoladas da parede celular de bactérias como estímulo para células é amplamente utilizada na literatura por simplificar a análise e atividade destes componentes como no caso de ácido lipoteicóico, peptidoglicanas, dentre outros. No entanto, a atividade destas estruturas pode ser modificada de maneira importante pela arquitetura e composição da parede celular em sua forma completa (NOEL et al.82, 1995). Em bactérias gram-positivas, como o Enterococcus faecalis, alguns componentes, como o ácido lipoteicóico, estão localizados no interior da parede celular, ao contrário do LPS em bactérias gram-negativas, o qual se localiza externamente à parede celular (SHOCKMAN; BARRET85, 1983). O modelo de estímulo com o microrganismo intacto inativado por calor tem sido utilizado por diversos autores e tem como vantagem principal considerar a virulência do microrganismo em seu estado íntegro (NOEL et al.82, 1995, SUGIYAMA et al.31, 2000, HOSOKAWA et al.33, 2005). Dada a importância de se conhecer mais sobre o papel dos fibroblastos no contexto de inflamação do microambiente pulpar, o presente estudo procurou investigar a capacidade dos fibroblastos pulpares produzirem in vitro duas quimiocinas: MIP1α/CCL3 e SDF-1/CXCL12. Nossos dados mostram que constitutivamente estas células foram capazes de produzir pequenas quantidades de MIP-1α, as quais aparentemente se acumularam ao longo dos períodos experimentais. Com o desafio antigênico proporcionado por Enterococcus faecalis morto por calor, a produção desta quimiocina pelas células foi alterada. Embora estatisticamente as diferenças observadas não tenham sido significativas, podemos sugerir uma tendência de aumento da produção de MIP1α/CCL3 na primeira hora de contato dos fibroblastos com o estímulo na proporção 1:1 e 100:1 bactérias:fibroblasto, sendo que nos períodos experimentais seguintes essa produção tende a diminuir chegando a 0 na concentração mais elevada de bactérias. A produção de MIP-1α/CCL3 induzida pelo estímulo na razão de 10 bactérias:fibroblasto Discussão 45 apresentou perfil de aumento gradativo ao longo do período experimental de 24 h, de certa forma semelhante ao ocorrido sem estímulo. Pelo nosso conhecimento, não existem relatos na literatura acerca da produção de MIP-1α/CCL3 por fibroblastos de polpa dental humana. O relato da produção de uma quimiocina pertencente ao grupo de proteína inflamatória para macrófago em polpas inflamadas foi realizado por NAKANISHI et al.86, 2005, por meio de imunohistoquímica. A marcação para MIP3α/CCL20 pôde ser localizada em monócitos/macrófagos CD68-positivos acumulados de forma ajdacente à lesão cariosa; em contrapartida, esta quimiocina foi marcada de maneira escassa no tecido pulpar normal. Fibroblastos sinoviais da ATM humana foram capazes de produzir MIP-1α/CCL3 mas em níveis muito próximos ao limite inferior de detecção por ELISA (OGURA et al.44, 2005). Sendo assim, nossos achados podem contribuir para o conhecimento da participação dos fibroblastos na imunidade e inflamação da polpa dental por meio da síntese de MIP-1α/CCL3, uma quimiocina com participação tanto na resposta imune inata como adaptativa. Conforme observado, a intensidade do estímulo agressor pode levar a diferentes perfis de produção de quimiocina por estas células. A produção aumentada de citocinas e quimiocinas por fibroblastos em decorrência de desafio microbiano tem sido documentada na literatura. HOSOKAWA et al.33, 2005, mostraram um aumento na produção de IL-8/CXCL8 por fibroblastos gengivais estimulados por Porphyromonas gingivalis morta por calor. Utilizando como estímulo o sobrenadante de bactérias, três trabalhos distintos de um mesmo grupo demonstraram que em fibroblastos pulpares há indução da expressão de mRNA para IL8/CXCL8 (YANG et al.36, 2003), IL-6 (YANG et al.37, 2003) e VEGF (YANG et al.39, 2004), sendo que o mRNA para estes três mediadores não havia sido detectado constitutivamente. Nossos resultados acerca da produção de MIP-1α corroboram o fenômeno do aumento da produção de citocinas conforme observado por YAMAJI et al.29, 1995, com o aumento de mRNA de IL-6 por LPS de E. coli e P. gingivalis em fibroblastos de ligamento periodontal; MAHANONDA et al.64, 2007, com o aumento de IL-8/CXCL8 por estímulos antigênicos variados em fibroblastos de gengiva; e UEHARA; TAKADA34, 2007 com o aumento da produção de IL-6, IL-8 e MCP1/CCL2 também por fibroblastos de gengiva. Com relação à produção de SDF-1/CXCL12, nossos resultados mostraram produção constitutiva desta quimiocina numa quantidade que pode ser importante para 46 Discussão manter a homeostasia do tecido pulpar, como já sugerido acerca do tecido periodontal por HOSOKAWA et al.33, 2005. Com o estímulo bacteriano e seu aumento em concentração, pudemos observar diminuição gradual, e estatisticamente significativa, da produção desta quimiocina. Estes dados podem ser considerados relevantes uma vez que corroboram o fenômeno relatado por HOSOKAWA et al.33, 2005, no qual Porphyromonas gingivalis morta por calor levou à diminuição na produção de CXCL12 conforme o aumento da concentração de bactérias indicada pela densidade óptica. Por outro lado, os autores relatam que a produção de IL-8/CXCL8 comportou-se de maneira contrária, aumentando juntamente com a concentração bacteriana. Utilizando-se de cultura primária de fibroblastos oriundos da derme e da gengiva, FEDYK et al.59, 2001, demonstraram haver também produção constitutiva de SDF-1/CXCL12 por fibroblastos de tecidos diferentes assim como observamos produção constitutiva desta quimiocina pelos fibroblastos pulpares. No entanto, no estudo daqueles autores, quando as células foram estimuladas por IL-1α e TNF-α, a produção de CXCL12 foi completamente inibida, assim como a migração de linfócitos B Nalm-6. Mais uma vez, mediante o estímulo pelas mesmas citocinas, os fibroblastos foram capazes de secretar quantidades mais elevadas de IL-8/CXCL8 conforme o aumento da concentração das citocinas IL1α e TNF-1 (FEDYK et al.59, 2001). Os nossos achados, juntamente com os de HOSOKAWA et al.33, 2005, e FEDYK et al.59, 2001, reforçam a possibilidade de diminuição da produção de SDF-1/CXCL12 em resposta à infecção. O fato da produção constitutiva das duas quimiocinas estudadas neste trabalho ter sido alterada em função da colocação de um microrganismo em contato com os fibroblastos sugere a possível ocorrência do reconhecimento de antígenos pelas células em cultura (MAHANONDA et al.64, 2007, UEHARA; TAKADA34, 2007). A capacidade de reconhecimento de antígenos por fibroblastos, em especial os pulpares, ainda não foi completamente elucidada. WANG et al.61, 1998, mostraram a presença de receptores CD14 em fibroblastos gengivais comprovando, ainda, a capacidade destes receptores em se ligar a LPS de P. gingivalis. O bloqueio com anticorpo anti-CD14 diminuiu, mas não inibiu completamente, a produção de IL-6, o que sugere a presença deste receptor no reconhecimento de antígenos por fibroblastos gengivais, mas também de outros receptores não elucidados no estudo. Ao comparar fibroblastos originários da gengiva e ligamento periodontal de um mesmo indivíduo, HATAKEYAMA et al.63, 2003, mostraram expressão de CD14 significativamente mais elevada por fibroblastos Discussão 47 de gengiva, expressão de TLR2 discretamente mais elevada por fibroblastos de ligamento periodontal e expressão semelhante de TLR4 por ambos grupos de célula. MAHANONDA et al.64, 2007, demonstraram a expressão de receptores TLR em fibroblastos de gengiva humana. Os receptores expressos foram: TLR1, TLR2, TLR3, TLR4, TLR5, TLR6 e TLR9. Ligantes específicos para TLRs 2, 3, 4 e 5 levaram ao aumento significativo da produção de IL-8 por estas células. Em estudo semelhante, UEHARA; TAKADA34, 2007, demonstraram a expressão de TLR1-9, MD-2, MyD88, NOD1 e NOD2, (todos moléculas reconhecedoras de antígenos). O estímulo com ligantes específicos para TLRs 2/6, 3, 4, 7/8, 9 e para NOD1 e NOD2, causou aumento significativo da produção de IL-6, IL-8 e MCP-1. Estes achados juntos sugerem não somente a presença destes receptores em fibroblastos, mas também sua relevância funcional (UEHARA; TAKADA34, 2007). Nas células da polpa dental, a existência de um destes receptores, TLR4, já foi descrita na camada de odontoblastos de polpas humanas por JIANG et al.87, 2006. Em células odontoblasto-like e células indiferenciadas de linhagem murina, BOTERO et al.62, 2006, observaram a expressão de CD14 e TLR4. Finalmente, vários receptores TLR (TLR1, TLR2, TL3, TLR4, TLR5, TLR6 e TLR9) foram expressos em odontoblastos humanos em cultura conforme relatado por DURAND et al.67, 2006. Até o presente, não há relatos da expressão destes receptores sabidamente capazes de reconhecer antígenos em cultura primária de fibroblastos de polpa dental humana, mas nossos achados contribuem para sua futura investigação, o que abre perspectivas para trabalhos futuros. A produção de SDF-1/CXCL12 presente no período experimental de 24h e ausente nos períodos anteriores sugere resposta celular mais demorada do que aquela que desencadeou a produção de MIP-1α/CCL3 ainda na primeira hora de estímulo. A produção de SDF-1/CXCL12, ao contrário da síntese de MIP-1α/CCL3, pode ter ocorrido não diretamente em resposta ao reconhecimento do antígeno, mas indiretamente, se considerarmos a provável produção de outras citocinas inflamatórias pelos fibroblastos em cultura. A produção de quimiocinas por diversas células e inclusive por fibroblastos pode ser influenciada por citocinas, resultando tanto no aumento como na diminuição de sua produção (GOUWY et al.88, 2005). Fibroblastos de gengiva humana aumentaram a produção de IL-8 com estímulo por IL-1β e TNF-α conforme estudo de TAKASHIBA et al.78, 1992. Em fibroblastos sinoviais da ATM, TNF-α mostrou ser capaz de aumentar a produção de IL-8/CXCL8, GRO-α/CXCL1, 48 Discussão MCP-1/CCL2 e RANTES/CCL5. Entretanto, a produção de MIP-1α/CCL3, MIP1β/CCL4 e SDF-1α/CXCL12 permaneceram muito baixas mesmo mediante incubação com TNF-1α (OGURA et al.44, 2005). De forma contrária, FEDYK et al.59, 2001, demonstraram que TNF-α e IL-1β diminuíram a produção de SDF-1/CXCL12 por fibroblastos de gengiva e derme humanas. No mesmo estudo, estas citocinas desencadearam aumento da produção de IL-8/CXCL8 e MCP-1/CCL2. Embora nosso estudo não tenha investigado a possível interferência de citocinas na produção de SDF1/CXCL12 pelos fibroblastos pulpares, não podemos descartar a possibilidade de ocorrência de produção e consumo de citocinas pelos fibroblastos, o que pode ter levado, por meio delas, à estimulação das células culminando na síntese de SDF1/CXCL12. Pelo fato de haver relatos de diminuição na produção de SDF-1/CXL12 tanto por estímulo com bactéria morta por calor (HOSOKAWA et al.33, 2005) como pela incubação com citocinas (FEDYK et al.59, 2001), estudos futuros devem ser delineados para possibilitar melhor compreensão dos mecanismos de sinalização autócrina, tais como aqueles envolvidos no reconhecimento de antígenos por fibroblastos de polpa dental humana. A inflamação do tecido pulpar é um fenômeno que, não raro, leva à perda de suas estruturas devido ao caráter de irreversibilidade que a pulpite assume conforme se instalam os fenômenos do processo inflamatório (KOKKAS et al89, 2007). Conhecer as bases da inflamação pulpar poderá contribuir, a longo prazo, para o avanço e crescimento da ciência em endodontia. O estudo dos diferentes grupos celulares que compõem a polpa dental permite conhecer o papel que os mesmos desempenham, isoladamente, dentro da resposta imune. O presente estudo relata, de maneira inédita, a capacidade de fibroblastos de polpa dental humana produzirem em cultura as quimiocinas MIP-1α e SDF-1. Além disso, mostramos que frente ao desafio microbiano com Enterococcus faecalis, estas células alteraram a produção destas quimiocinas. Nossos achados demonstram que os fibroblastos, mesmo não compondo a primeira linha de defesa do tecido pulpar contra os agentes bacterianos da cárie dental, provavelmente desempenham papel importante dentro da resposta inflamatória da polpa. Podemos sugerir que os fibroblastos são capazes de recrutar células de defesa, não se limitando, portanto, a funções estruturais, mas agindo também como verdadeiras sentinelas do tecido pulpar. Conclusão 51 6 Conclusão Nas condições em que este trabalho foi realizado, pode-se concluir que: Fibroblastos oriundos da polpa dental humana são capazes de produzir MIP-1α e SDF-1 in vitro constitutivamente. Esta produção se alterou em resposta ao desafio por Enterococcus faecalis inativado por calor. A concentração de microrganismos parece induzir respostas diferentes quanto à produção destas quimiocinas; pois os fibroblastos demonstraram uma tendência para o aumento da produção de MIP-1α enquanto a produção de SDF-1 diminuiu com o aumento da concentração bacteriana. O tempo de estímulo determinou diminuição da produção de MIP-1α nas concentrações mais baixa e mais elevada, enquanto no grupo controle e na concentração intermediária de bactérias, a quimiocina parece se acumular no sobrenadante. A produção de SDF-1 só pôde ser detectada no período experimental de 24h, o que sugere a participação de vias autócrinas envolvidas na síntese desta quimiocina. Referências Bibliográficas 55 Referências bibliográficas 1. Chiego Jr. D. Histology of the pulp. In: Avery J, editor. Oral development and histology. 3rd ed. ed. Stuttgart: Georg Thieme Verlag; 2002. p. 190-212. 2. Goodis H. History of pulp biology. In: Hargreaves K, Goodis H, editors. Seltzer and Bender´s Dental Pulp. 1 ed. Carol Stream: Quintessence publishing co.; 2002. p. 111. 3. Okiji T. Pulp as a connective tissue. In: Hargreaves K, Goodis H, editors. Seltzer and Bender´s Dental Pulp. Carol Stream: Quintessence Publishing Co.; 2002. p. 95-122. 4. Takashiba S, Naruishi K, Murayama Y. Perspective of cytokine regulation for periodontal treatment: fibroblast biology. J Periodontol 2003;74(1):103-10. 5. Trowbridge H. Histology of pulpal inflamation. In: Hargreaves K, Goodis H, editors. Seltzer and Bender´s Dental Pulp. Carol Stream: Quintessence publishing Co.; 2002. p. 227-245. 6. Love RM, Jenkinson HF. Invasion of dentinal tubules by oral bacteria. Crit Rev Oral Biol Med 2002;13(2):171-83. 7. Martin FE, Nadkarni MA, Jacques NA, Hunter N. Quantitative microbiological study of human carious dentine by culture and real-time PCR: association of anaerobes with histopathological changes in chronic pulpitis. J Clin Microbiol 2002;40(5):1698704. 8. Kakehashi S, Stanley HR, Fitzgerald RJ. The Effects of Surgical Exposures of Dental Pulps in Germ-Free and Conventional Laboratory Rats. Oral Surg Oral Med Oral Pathol 1965;20:340-9. 9. Moller AJ, Fabricius L, Dahlen G, Sundqvist G, Happonen RP. Apical periodontitis development and bacterial response to endodontic treatment. Experimental root canal infections in monkeys with selected bacterial strains. Eur J Oral Sci 2004;112(3):207-15. 10. Kaufman B, Spangberg L, Barry J, Fouad AF. Enterococcus spp. in endodontically treated teeth with and without periradicular lesions. J Endod 2005;31(12):851-6. 11. Sedgley C, Nagel A, Dahlen G, Reit C, Molander A. Real-time quantitative polymerase chain reaction and culture analyses of enterococcus faecalis in root canals. J Endod 2006;32(3):173-7. 12. Williams JM, Trope M, Caplan DJ, Shugars DC. Detection and Quantitation of E. faecalis by Real-time PCR (qPCR), Reverse Transcription-PCR (RT-PCR), and Cultivation During Endodontic Treatment. J Endod 2006;32(8):715-21. 56 Referências Bibliográficas 13. Zoletti GO, Siqueira JF, Jr., Santos KR. Identification of Enterococcus faecalis in Root-filled Teeth With or Without Periradicular Lesions by Culture-dependent andIndependent Approaches. J Endod 2006;32(8):722-6. 14. Jett BD, Huycke MM, Gilmore MS. Virulence of enterococci. Clin Microbiol Rev 1994;7(4):462-78. 15. Sedgley CM, Lennan SL, Clewell DB. Prevalence, phenotype and genotype of oral enterococci. Oral Microbiol Immunol 2004;19(2):95-101. 16. Sedgley C, Buck G, Appelbe O. Prevalence of Enterococcus faecalis at multiple oral sites in endodontic patients using culture and PCR. J Endod 2006;32(2):104-9. 17. Flahaut S, Hartke A, Giard JC, Benachour A, Boutibonnes P, Auffray Y. Relationship between stress response toward bile salts, acid and heat treatment in Enterococcus faecalis. FEMS Microbiol Lett 1996;138(1):49-54. 18. Rozdzinski E, Marre R, Susa M, Wirth R, Muscholl-Silberhorn A. Aggregation substance-mediated adherence of Enterococcus faecalis to immobilized extracellular matrix proteins. Microb Pathog 2001;30(4):211-20. 19. 728. Linde A, Goldberg M. Dentinogenesis. Crit Rev Oral Biol Med 1993;4(5):679- 20. Bystrom A, Claesson R, Sundqvist G. The antibacterial effect of camphorated paramonochlorophenol, camphorated phenol and calcium hydroxide in the treatment of infected root canals. Endod Dent Traumatol 1985;1(5):170-5. 21. Distel JW, Hatton JF, Gillespie MJ. Biofilm formation in medicated root canals. J Endod 2002;28(10):689-93. 22. Signoretto C, Lleo MM, Tafi MC, Canepari P. Cell wall chemical composition of Enterococcus faecalis in the viable but nonculturable state. Appl Environ Microbiol 2000;66(5):1953-9. 23. Oliver JD. The viable but nonculturable state in bacteria. J Microbiol 2005;43 Spec No:93-100. 24. Ginsburg I. Role of lipoteichoic acid in infection and inflammation. Lancet Infect Dis 2002;2(3):171-9. 25. Chan CP, Lan WH, Chang MC, Chen YJ, Lan WC, Chang HH, et al. Effects of TGF-beta s on the growth, collagen synthesis and collagen lattice contraction of human dental pulp fibroblasts in vitro. Arch Oral Biol 2005;50(5):469-79. 26. Barkhordar RA, Ghani QP, Russell TR, Hussain MZ. Interleukin-1beta activity and collagen synthesis in human dental pulp fibroblasts. J Endod 2002;28(3):157-9. Referências Bibliográficas 57 27. Sismey-Durrant HJ, Hopps RM. Effect of lipopolysaccharide from Porphyromonas gingivalis on prostaglandin E2 and interleukin-1-beta release from rat periosteal and human gingival fibroblasts in vitro. Oral Microbiol Immunol 1991;6(6):378-80. 28. Tamura M, Tokuda M, Nagaoka S, Takada H. Lipopolysaccharides of Bacteroides intermedius (Prevotella intermedia) and Bacteroides (Porphyromonas) gingivalis induce interleukin-8 gene expression in human gingival fibroblast cultures. Infect Immun 1992;60(11):4932-7. 29. Yamaji Y, Kubota T, Sasaguri K, Sato S, Suzuki Y, Kumada H, et al. Inflammatory cytokine gene expression in human periodontal ligament fibroblasts stimulated with bacterial lipopolysaccharides. Infect Immun 1995;63(9):3576-81. 30. Wang PL, Shirasu S, Shinohar M, Azuma Y, Daito M, Yasuda H, et al. IL-10 inhibits Porphyromonas gingivalis LPS-stimulated human gingival fibroblasts production of IL-6. Biochem Biophys Res Commun 1999;263(2):372-7. 31. Sugiyama A, Ogawa T, Daikuhara Y, Takada H. Enhancement of hepatocyte growth factor (scatter factor) production by human gingival fibroblasts in culture stimulated with Porphyromonas gingivalis fimbriae. J Med Microbiol 2000;49(4):31925. 32. Wang PL, Azuma Y, Shinohara M, Ohura K. Toll-like receptor 4-mediated signal pathway induced by Porphyromonas gingivalis lipopolysaccharide in human gingival fibroblasts. Biochem Biophys Res Commun 2000;273(3):1161-7. 33. Hosokawa Y, Hosokawa I, Ozaki K, Nakae H, Murakami K, Miyake Y, et al. CXCL12 and CXCR4 expression by human gingival fibroblasts in periodontal disease. Clin Exp Immunol 2005;141(3):467-74. 34. Uehara A, Takada H. Functional TLRs and NODs in Human Gingival Fibroblasts. J Dent Res 2007;86(3):249-54. 35. Tokuda M, Nagaoka S, Torii M. Interleukin-10 inhibits expression of interleukin-6 and -8 mRNA in human dental pulp cell cultures via nuclear factorkappaB deactivation. J Endod 2002;28(3):177-80. 36. Yang LC, Huang FM, Lin CS, Liu CM, Lai CC, Chang YC. Induction of interleukin-8 gene expression by black-pigmented Bacteroides in human pulp fibroblasts and osteoblasts. Int Endod J 2003;36(11):774-9. 37. Yang LC, Tsai CH, Huang FM, Liu CM, Lai CC, Chang YC. Induction of interleukin-6 gene expression by pro-inflammatory cytokines and black-pigmented Bacteroides in human pulp cell cultures. Int Endod J 2003;36(5):352-7. 38. Coil J, Tam E, Waterfield JD. Proinflammatory cytokine profiles in pulp fibroblasts stimulated with lipopolysaccharide and methyl mercaptan. J Endod 2004;30(2):88-91. 58 Referências Bibliográficas 39. Yang LC, Tsai CH, Huang FM, Su YF, Lai CC, Liu CM, et al. Induction of vascular endothelial growth factor expression in human pulp fibroblasts stimulated with black-pigmented Bacteroides. Int Endod J 2004;37(9):588-92. 40. Ollier WE. Cytokine genes and disease susceptibility. Cytokine 2004;28(45):174-8. 41. Abbas A, Lichtman A. Cytokines. In: Abbas A, Lichtman A, editors. Cellular and molecular immunology. 5th ed. ed. Philadelphia: Saunders; 2003. p. 243-274. 42. Zlotnik A, Yoshie O. Chemokines: a new classification system and their role in immunity. Immunity 2000;12(2):121-7. 43. Schall TJ, Bacon K, Camp RD, Kaspari JW, Goeddel DV. Human macrophage inflammatory protein alpha (MIP-1 alpha) and MIP-1 beta chemokines attract distinct populations of lymphocytes. J Exp Med 1993;177(6):1821-6. 44. Ogura N, Tobe M, Sakamaki H, Nagura H, Abiko Y, Kondoh T. Tumor necrosis factor-alpha increases chemokine gene expression and production in synovial fibroblasts from human temporomandibular joint. J Oral Pathol Med 2005;34(6):35763. 45. Dieu-Nosjean MC, Vicari A, Lebecque S, Caux C. Regulation of dendritic cell trafficking: a process that involves the participation of selective chemokines. J Leukoc Biol 1999;66(2):252-62. 46. Nagaoka S, Tokuda M, Sakuta T, Taketoshi Y, Tamura M, Takada H, et al. Interleukin-8 gene expression by human dental pulp fibroblast in cultures stimulated with Prevotella intermedia lipopolysaccharide. J Endod 1996;22(1):9-12. 47. Choe H, Farzan M, Sun Y, Sullivan N, Rollins B, Ponath PD, et al. The betachemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell 1996;85(7):1135-48. 48. Baggiolini M, Dewald B, Moser B. Human chemokines: an update. Annu Rev Immunol 1997;15:675-705. 49. Rollins BJ. Chemokines. Blood 1997;90(3):909-28. 50. Menten P, Wuyts A, Van Damme J. Macrophage inflammatory protein-1. Cytokine Growth Factor Rev 2002;13(6):455-81. 51. Maurer M, von Stebut E. Macrophage inflammatory protein-1. Int J Biochem Cell Biol 2004;36(10):1882-6. 52. Bonecchi R, Polentarutti N, Luini W, Borsatti A, Bernasconi S, Locati M, et al. Up-regulation of CCR1 and CCR3 and induction of chemotaxis to CC chemokines by IFN-gamma in human neutrophils. J Immunol 1999;162(1):474-9. Referências Bibliográficas 59 53. Bleul CC, Fuhlbrigge RC, Casasnovas JM, Aiuti A, Springer TA. A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1 (SDF-1). J Exp Med 1996;184(3):1101-9. 54. Shirozu M, Nakano T, Inazawa J, Tashiro K, Tada H, Shinohara T, et al. Structure and chromosomal localization of the human stromal cell-derived factor 1 (SDF1) gene. Genomics 1995;28(3):495-500. 55. Aiuti A, Webb IJ, Bleul C, Springer T, Gutierrez-Ramos JC. The chemokine SDF-1 is a chemoattractant for human CD34+ hematopoietic progenitor cells and provides a new mechanism to explain the mobilization of CD34+ progenitors to peripheral blood. J Exp Med 1997;185(1):111-20. 56. Chan JR, Hyduk SJ, Cybulsky MI. Chemoattractants induce a rapid and transient upregulation of monocyte alpha4 integrin affinity for vascular cell adhesion molecule 1 which mediates arrest: an early step in the process of emigration. J Exp Med 2001;193(10):1149-58. 57. Martin C, Burdon PC, Bridger G, Gutierrez-Ramos JC, Williams TJ, Rankin SM. Chemokines acting via CXCR2 and CXCR4 control the release of neutrophils from the bone marrow and their return following senescence. Immunity 2003;19(4):583-93. 58. Dieu MC, Vanbervliet B, Vicari A, Bridon JM, Oldham E, Ait-Yahia S, et al. Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites. J Exp Med 1998;188(2):373-86. 59. Fedyk ER, Jones D, Critchley HO, Phipps RP, Blieden TM, Springer TA. Expression of stromal-derived factor-1 is decreased by IL-1 and TNF and in dermal wound healing. J Immunol 2001;166(9):5749-54. 60. Wright SD, Ramos RA, Tobias PS, Ulevitch RJ, Mathison JC. CD14, a receptor for complexes of lipopolysaccharide (LPS) and LPS binding protein. Science 1990;249(4975):1431-3. 61. Wang PL, Sato K, Oido M, Fujii T, Kowashi Y, Shinohara M, et al. Involvement of CD14 on human gingival fibroblasts in Porphyromonas gingivalis lipopolysaccharide-mediated interleukin-6 secretion. Arch Oral Biol 1998;43(9):687-94. 62. Botero TM, Shelburne CE, Holland GR, Hanks CT, Nor JE. TLR4 mediates LPS-induced VEGF expression in odontoblasts. J Endod 2006;32(10):951-5. 63. Hatakeyama J, Tamai R, Sugiyama A, Akashi S, Sugawara S, Takada H. Contrasting responses of human gingival and periodontal ligament fibroblasts to bacterial cell-surface components through the CD14/Toll-like receptor system. Oral Microbiol Immunol 2003;18(1):14-23. 64. Mahanonda R, Sa-Ard-Iam N, Montreekachon P, Pimkhaokham A, Yongvanichit K, Fukuda MM, et al. IL-8 and IDO Expression by Human Gingival Fibroblasts via TLRs. J Immunol 2007;178(2):1151-7. 60 Referências Bibliográficas 65. Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell 2006;124(4):783-801. 66. Akira S, Takeda K, Kaisho T. Toll-like receptors: critical proteins linking innate and acquired immunity. Nat Immunol 2001;2(8):675-80. 67. Durand SH, Flacher V, Romeas A, Carrouel F, Colomb E, Vincent C, et al. Lipoteichoic acid increases TLR and functional chemokine expression while reducing dentin formation in in vitro differentiated human odontoblasts. J Immunol 2006;176(5):2880-7. 68. Letzelter C, Croute F, Pianezzi B, Roques C, Soleilhavoup JP. Supernatant cytotoxicity and proteolytic activity of selected oral bacteria against human gingival fibroblasts in vitro. Arch Oral Biol 1998;43(1):15-23. 69. Yamasaki M, Nakata K, Imaizumi I, Iwama A, Nakane A, Nakamura H. Cytotoxic effect of endodontic bacteria on periapical fibroblasts. J Endod 1998;24(8):534-9. 70. Suthin K, Matsushita K, Machigashira M, Tatsuyama S, Imamura T, Torii M, et al. Enhanced expression of vascular endothelial growth factor by periodontal pathogens in gingival fibroblasts. J Periodontal Res 2003;38(1):90-6. 71. Sugiyama A, Arakaki R, Ohnishi T, Arakaki N, Daikuhara Y, Takada H. Lipoteichoic acid and interleukin 1 stimulate synergistically production of hepatocyte growth factor (scatter factor) in human gingival fibroblasts in culture. Infect Immun 1996;64(4):1426-31. 72. Wang PL, Shirasu S, Daito M, Ohura K. Streptococcus mutans lipoteichoic acidinduced apoptosis in cultured dental pulp cells from human deciduous teeth. Biochem Biophys Res Commun 2001;281(4):957-61. 73. Telles PD, Hanks CT, Machado MA, Nor JE. Lipoteichoic acid up-regulates VEGF expression in macrophages and pulp cells. J Dent Res 2003;82(6):466-70. 74. Botero TM, Mantellini MG, Song W, Hanks CT, Nor JE. Effect of lipopolysaccharides on vascular endothelial growth factor expression in mouse pulp cells and macrophages. Eur J Oral Sci 2003;111(3):228-34. 75. Tardif F, Ross G, Rouabhia M. Gingival and dermal fibroblasts produce interleukin-1 beta converting enzyme and interleukin-1 beta but not interleukin-18 even after stimulation with lipopolysaccharide. J Cell Physiol 2004;198(1):125-32. 76. Gutierrez-Venegas G, Maldonado-Frias S, Ontiveros-Granados A, KawasakiCardenas P. Role of p38 in nitric oxide synthase and cyclooxygenase expression, and nitric oxide and PGE2 synthesis in human gingival fibroblasts stimulated with lipopolysaccharides. Life Sci 2005;77(1):60-73. 77. Hosokawa Y, Nakanishi T, Yamaguchi D, Nakae H, Matsuo T. Expression of fractalkine (CX3CL1) and its receptor, CX3CR1, in periodontal diseased tissue. Clin Exp Immunol 2005;139(3):506-12. Referências Bibliográficas 61 78. Takashiba S, Takigawa M, Takahashi K, Myokai F, Nishimura F, Chihara T, et al. Interleukin-8 is a major neutrophil chemotactic factor derived from cultured human gingival fibroblasts stimulated with interleukin-1 beta or tumor necrosis factor alpha. Infect Immun 1992;60(12):5253-8. 79. Brouty-Boye D, Pottin-Clemenceau C, Doucet C, Jasmin C, Azzarone B. Chemokines and CD40 expression in human fibroblasts. Eur J Immunol 2000;30(3):914-9. 80. Ravanti L, Hakkinen L, Larjava H, Saarialho-Kere U, Foschi M, Han J, et al. Transforming growth factor-beta induces collagenase-3 expression by human gingival fibroblasts via p38 mitogen-activated protein kinase. J Biol Chem 1999;274(52):37292300. 81. Martinez EF, Araujo VC. In vitro immunoexpression of extracellular matrix proteins in dental pulpal and gingival human fibroblasts. Int Endod J 2004;37(11):74955. 82. Noel RF, Jr., Sato TT, Mendez C, Johnson MC, Pohlman TH. Activation of human endothelial cells by viable or heat-killed gram-negative bacteria requires soluble CD14. Infect Immun 1995;63(10):4046-53. 83. Bier O. Técnicas bacteriológicas. In: Bier O, editor. Microbiologia e Imunologia. 24ª ed. São Paulo: Melhoramentos; 1985. p. 919-998. 84. Phipps RP, Penney DP, Keng P, Quill H, Paxhia A, Derdak S, et al. Characterization of two major populations of lung fibroblasts: distinguishing morphology and discordant display of Thy 1 and class II MHC. Am J Respir Cell Mol Biol 1989;1(1):65-74. 85. Shockman GD, Barrett JF. Structure, function, and assembly of cell walls of gram-positive bacteria. Annu Rev Microbiol 1983;37:501-27. 86. Nakanishi T, Takahashi K, Hosokawa Y, Adachi T, Nakae H, Matsuo T. Expression of macrophage inflammatory protein 3alpha in human inflamed dental pulp tissue. J Endod 2005;31(2):84-7. 87. Jiang HW, Zhang W, Ren BP, Zeng JF, Ling JQ. Expression of toll like receptor 4 in normal human odontoblasts and dental pulp tissue. J Endod 2006;32(8):747-51. 88. Gouwy M, Struyf S, Proost P, Van Damme J. Synergy in cytokine and chemokine networks amplifies the inflammatory response. Cytokine Growth Factor Rev 2005;16(6):561-80. 89. Kokkas AB, Goulas A, Varsamidis K, Mirtsou V, Tziafas D. Irreversible but not reversible pulpitis is associated with up-regulation of tumour necrosis factor-alpha gene expression in human pulp. Int Endod J 2007;40(3):198-203. 62 Referências Bibliográficas Abstract 65 Abstract Production of MIP-1alfa and SDF-1 by cultured human dental pulp fibroblasts challenged by heat killed Enterococcus faecalis Dental pulp is a connective tissue structure constituted by many different cell types. Among them, the fibroblasts are the most frequent ones. When challenged by different aggressive agents, these cells are able to release some substances like cytokines and chemokines, which are essential to trigger the inflammatory process. The aims of this study were: 1. to evaluate the ability of fibroblasts to produce the chemokines MIP-1α/CCL3) and SDF-1/CXCL12; 2. to evaluate the expression of these chemokines by fibroblasts when challenged by heat killed Enterococcus faecalis in gradual concentrations and 3. to evaluate the production of these chemokines in a time course manner. The dental pulp from non-carious third molar was collected from a healthy patient. Explants were made and stocked in culture medium (DMEM) for fibroblasts growth. The cells were used since passage four. In a 24-well plate and after reaching confluence, culture medium alone or containing heat killed E. faecalis at proportion 1:1, 10:1 and 100:1 bacteria:fibroblast, were added to the fibroblasts. After 1, 6 and 24 hours, the supernatants were collected for analysis. The protein detection of MIP-1α/CCL3 and SDF-1/CXCL12 was performed by ELISA. For statistical analysis, data were assessed by Kruskal-Wallis followed by Miller post-test. Significance levels of 5% were adopted. Production of both chemokines was detected by ELISA. Pulp fibroblasts were able to produce SDF-1 constitutively. This production decreased with the increase in the number of heat killed E. faecalis increased (p<0.05). Production of MIP-1α was detected in unchallenged and challenged cells. The median bacterial concentration (10:1) presented a profile production similar to that of unstimulated cells. Bacterial concentrations of 1 and 100 microrganisms/cell showed a highly enhanced production of MIP-1α at the first hour of stimulum; however, these data were not statistically significant (p>0.05). Fibroblasts ability to produce chemokines, like MIP1α and SDF-1, confirms their importance at immune and inflammatory events in dental pulp, specially being fibroblasts the most abundant cells at this microenvironment. Keywords: Fibroblasts. Chemokines. Inflammation.
Baixar