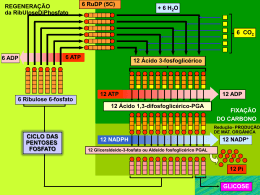
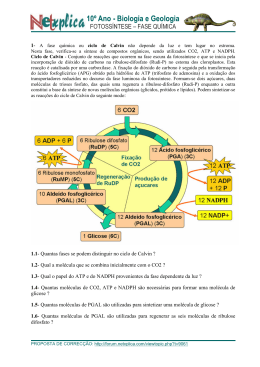
EIXO BIOLÓGICO Unidade 3 Metabolismo Autor: Professor Carlos Eduardo Anunciação I. Introdução II. Princípio evolutivo do metabolismo III. Membrana bilipídica e metabolismo celular IV. Moeda de energia, o ATP V. Fosforilação no nível do substrato VI. Atmosfera primitiva e obtenção de energia VII. Equilíbrio dinâmico VIII. Referências #M2U3 I. Introdução Nesta unidade, vamos tratar do mecanismo básico que rege o funcionamento das células e seu processo de produção de energia para se manter viva. Vamos ver que as biomoléculas que constituem nosso organismo possuem características especiais, capazes de “trocar” energia com outros compostos e até de possuir atividade mecânica como a locomoção. Veremos, também, que todas as biomoléculas são regidas por princípios básicos como as leis da termodinâmica, e que é necessário algum tipo de energia para a realização de tais processos. Vamos ver, ainda, que no curso da evolução, as células passaram a apresentar meios para a utilização de diferentes formas de energia. Esses meios permitiram a realização de seus processos, “construindo” vias alternativas para vencer as grandes barreiras energéticas, às vezes, requeridas para as suas sínteses. Todos os seres vivos sejam unicelulares ou multicelulares, teciduais ou não, precisam de “alimento” para manutenção da célula/vida. É desse alimento que são retiradas as biomoléculas básicas para a construção de seu organismo, como os aminoácidos. Essas biomoléculas são modificadas, para assim “construir” outras novas para sua estrutura e multiplicação ou para “desmontá-las” e obter energia para manter viva a célula e suas funções. A esse processamento das biomoléculas do organismo, seja ele no sentido de construir ou desmontar, chamamos de metabolismo. Nesse caso, a palavra “construir” é usada no sentido de se sintetizar uma molécula mais complexa, apresentando maior valor de energia livre a partir de compostos mais simples, com menor valor de energia livre. No mesmo contexto, a palavra “desmontar” é usada no sentido inverso. 220 Módulo II — Processos biológicos na captação e na transformação da matéria e energia P Eixo Biológico BSC Em outras palavras, a soma dos processos da biotransformação (químico ou físico) das moléculas nas células, desmontando-as, é que torna a energia disponível, pronta para a manutenção do tônus celular e para a biossíntese de novas biomoléculas, mantendo o organismo vivo. Dessa forma, os seguintes processos ocorrem nas células de forma a permitir a manutenção da sua atividade biológica: Apresentação de uma estrutura celular/molecular definida que comporte seus constituintes e a proteja. Uso de aparatos capazes de selecionar e buscar substâncias energéticas, além de transferi-las para seu interior. Controle de um sistema ágil de biotransformação, que lhe permita desmontar as substâncias energéticas ingeridas com mínima perda de energia. Armazenamento da energia em um sistema que permita utilizá-la ou transferi-la para outras reações ou biomoléculas. Respeito aos princípios da termodinâmica nas construções e transformações da matéria e da energia. #M2U3 II. Princípio evolutivo do metabolismo. Você já parou para pensar como nosso organismo ou nossas células selecionam o que irá processar? Como o alimento selecionado é percebido entre milhões de substâncias? Como após selecionar o alimento, trabalhamos sobre sua estrutura química retirando o máximo de energia útil, armazenando-a para a usar posteriormente? Bem, poderíamos formular muitas outras perguntas sobre como nosso organismo funciona bioquimicamente, mas, para entendermos o funcionamento desse mecanismo complexo e para alcançarmos essas respostas de maneira mais fácil, precisamos, antes, pensar no funcionamento de indivíduos mais simples e unicelulares como as bactérias e, também, sobre como devem ter ocorrido alguns aspectos da evolução ou o princípio evolutivo do metabolismo. O ambiente primitivo, que originou a primeira célula, era rico na variedade de biomoléculas resultantes das transformações químicas e físicas dos elementos disponíveis na atmosfera, e que eram depositados no mar. Em alguma região desse mar, onde possivelmente se concentraram grupos de biomoléculas (os coacervados), é que surgiram, do contato entre eles, as primeiras células. Destas “experiências” celulares, surgiu um tipo funcional que adquiriu propriedades replicativas, evoluindo e originando as células como as conhecemos hoje. www. Saiba mais sobre a origem da vida no site: http://www.brasilescola.com/biologia/origem-vida.htm Essas “experiências celulares”, apesar de terem sido testadas aleatoriamente em toda a superfície do planeta, foram bem-sucedidas apenas no mar. Este último fenômeno não aconteceu por acaso, uma vez que as biomoléculas das nossas células possuem propriedades físico-químicas muito interessantes quando se encontram na água. Tais propriedades possibilitam sua organização ou agrupamento, formando estruturas como as membranas e propiciando a ocorrência de reações químicas diversas de moléculas, no sentido de desagregá-las ou desmontá-las, liberando, também, energia. Consórcio Setentrional de Ensino a Distância 221 B # M2U3 Metabolismo Todas estas atividades só foram possíveis porque no mar, ou melhor, na água do mar, há presença de sais, íons e metais diversos, além do pH moderado (em torno de 7,0), o que dá origem a propriedades físicas e químicas especiais das biomoléculas e, em alguns casos, as torna mais energéticas ou reativas. Como exemplo, temos os lipídios, que podem apresentar diferentes padrões de interação com a água, como no caso dos óleos, que interagem de forma diferente que os ácidos graxos e, também, os lipídeos de membrana (fosfolipídios). A água possui uma estrutura molecular organizada e dinâmica, apesar de fluida. Ela se organiza ordenando as faces de suas moléculas de acordo com a possibilidade de interagirem com pontes de hidrogênio (H:::H). Essa interação permite que se forme uma estrutura fluida cristalina e organizada (Figura 1ª) com um baixo grau de energia, sendo que qualquer substância adicionada à água interferirá na estrutura cristalina (pontes de hidrogênio), desorganizando-a e aumentando seu grau de entropia (∆S). Assim, quanto mais pontes de hidrogênio uma substância realizar com a água, mais solúvel em água ela será. A substância será considerada insolúvel quando não puder realizar interações ou quando realizar poucas interações, comparadas com seu peso molecular. a) b) Figura 1: a) Estrutura cristalina da água; b) Dissolução de partículas na água. 222 Módulo II — Processos biológicos na captação e na transformação da matéria e energia P Eixo Biológico Saiba mais Óleos comestíveis são formados predominantemente por triacilgliceróis, que não apresentam polaridade nem caráter anfipático. Os ácidos graxos, que são parte dos triacilgliceróis, estão esterificados ao glicerol, perdendo, portanto, sua característica de polaridade. Os ácidos graxos livres, apesar de presentes, são componentes minoritários, não interferindo, significativamente, na solubilidade em água. BSC Experimente colocar duas gotas de óleo em uma lâmina de água parada e tente explicar termodinamicamente o que acontece. Em uma situação ou em que as moléculas de ácidos graxos são colocadas em contato com a água, um padrão de interação diferente pode ser observado. Essas moléculas, pouco solúveis, quando jogadas dispersas sobre a água, tendem a se agrupar aumentando sua estabilidade ou tornando sua co-existência com moléculas de água mais viável. Isso ocorre porque a sua estrutura química possui uma cauda imiscível ou apolar (hidrocarbonada (–CH2-CH2-CH2)n-CH3) ligada a um grupo ácido carboxílico, um ácido fraco, que, portanto, tem natureza polar (cabeça polar ou -COOH) com afinidade pela água. (Figura 2) Figura 2: Fórmula estrutural e molecular da cadeia de hidrocarbonetos de um ácido graxo (lipídio). Uma vez na água, este grupo ácido se dissocia ionizando-se negativamente e atraindo um cátion. Assim, isolado, este ácido graxo se posiciona de forma a expor a cabeça hidrofílica para a água, mas com a cauda hidrofóbica procurando uma condição de maior estabilidade, ou falando físico-quimicamente, procurando um ∆G0 mais favorável ou, ainda, procurando uma condição energética mínima em contato com a água. (Figura 3a). Quando existem outras moléculas de ácido graxo na água, a estrutura cristalina desfeita pela presença da cabeça polar “pressionando”, localmente, reorganiza-se para adquirir uma menor entropia ou estado energético, favorecendo o agrupamento das moléculas de ácido graxo. Cada molécula de ácido graxo ocupa um espaço razoável na água, de forma que moléculas de água, próximas à região apola,r efetuem um número de pontes menor do que poderiam, devido às restrições espaciais. Isso faz que moléculas de água mais próximas da região hidrofóbica sejam menos estáveis que moléculas de água em outras regiões, levando as moléculas mais instáveis a uma mobilidade maior. Assim, ao invés de vários pontos de conflito e quebra da estrutura cristalina em locais diversos, a água adquire apenas alguns pontos, resultando numa menor entropia possível, com menor ∆G0. Consórcio Setentrional de Ensino a Distância 223 B # M2U3 Metabolismo Para o ácido graxo, este encontro também é favorável, pois diminui o conflito da presença da cauda apolar com a água. Ele se posiciona com as caudas em bloco, agrupadas a uma distância que permite interações hidrofóbicas, o que dá maior força de coesão para as caudas apolares e reduz a entropia (∆G0 mais favorável). Este efeito aglutinador pode ser explicado, também, pela força de coesão das partículas de água. A água possui uma forte interação entre seus átomos, e a presença de moléculas apolares, como os lipídios, provoca a perturbação da estrutura cristalina do meio. As moléculas de água são agrupadas pela afinidade que elas têm de se agruparem. Agora, quando se agita a água, contendo cátions monovalentes imersos como o Na+ e K+, os ácidos graxos expõem toda a cauda apolar no meio aquoso, aumentam seu conflito e, para atingir um menor grau de entropia ou uma condição mais estável, se agrupam em pequenas micelas ou bolhas, expondo sua cabeça polar para a água e as caudas apolares para dentro, esta é a estrutura cristalina do sabão. (Figura 3 b). Figura 3: a) dissolução de lipídios na água; b) formação de micelas. 224 Módulo II — Processos biológicos na captação e na transformação da matéria e energia Saiba mais Existem vários modelos que ilustram a forma e o comportamento da micela. Um dos mais aceitos é o modelo de Stigter, no qual cada unidade de ácido graxo se organiza em forma esférica, onde todas as porções hidrofóbicas estão voltadas para o centro (núcleo micelar) e as cabeças hidrofílicas ficam na superfície, em contato com a água (camada de Stern). P Eixo Biológico BSC A propriedade de formar micelas é muito útil na formação das membranas, por um tipo especial de lipídios denominados de diacilgliceróis. Esses lipídios são os principais constituintes das membranas biológicas e possuem duas caudas hidrocarbonatadas hidrofóbicas e uma cabeça polar. Na água, esses lipídios tendem a se organizar como nas micelas descritas anteriormente, mas devido à sua forma cilíndrica (duas caudas), ao se disporem lado a lado, eles tendem a formar folhas que proporcionam a formação de grandes “micelas” em dupla camada, pois a forma cilíndrica dificulta a formação de micelas pequenas. (Figura 4) Figura 4: Bicamada lipídica com a cabeça polar dos lipídios voltadas para o lado externo e a cauda apolar, para o lado interno. Assim, a cauda apolar é escondida da água, de forma a alcançar um estado termodinâmico mais estável, sendo que os lipídios se dispõem lado a lado, formando folhas e protegendo a ponta descoberta da cauda apolar. Posteriormente, os lipídios agregamse em duas camadas unidas pela “ponta” apolar e as cabeças polares dos ácidos carboxílicos, ficam voltados para a face externa que, por sua vez, fica voltada para a água. Saiba mais O pH sanguíneo está entre 7,35 a 7,45, ou seja, é levemente alcalino. Dessa forma, o equilíbrio dependerá da proporção de bicarbonato e ácido carbônico presente. O desequilíbrio do pH pode causar acidose ou alcalose metabólica ou respiratória. Outro exemplo do que ocorre na água ocorre no ar com o CO2 ou gás carbônico. Como este gás de alta estabilidade química se dispersa e se adquire um estado de menor entropia? Embora seja um gás, ele se dissolve na água. O que torna isso possível? Lançado na água, o CO2 pode reagir com ela formando o bicarbonato, conforme a reação abaixo, e resultando num grau de energia maior (∆ G0). Esta reação pode ocorrer espontaneamente já que e a diferença do ∆G entre o CO2 livre e sua forma hidratada é pequena, e a energia desse processo pode ser fornecida pelo calor do ambiente. Na forma hidratada, este sal se dissocia gerando o ânion carbamato (HCO3-) que possui um pK (constante de dissociação) de 7,4, ou seja, age como um tampão mantendo o pH em torno de 7,4 e, por esta propriedade, constitui um dos principais tampões do nosso organismo e da água do mar. Esse tampão é tão importante que são encontradas enzimas especiais capazes de catalisar esta reação e aumentar sua formação, de forma a auxiliar no tamponamento do pH sanguíneo. Reação de hidratação do CO2 CO2 + H2O → H2CO3 → H+ + HCO3- Consórcio Setentrional de Ensino a Distância 225 B # M2U3 Metabolismo #M2U3 III. Membrana bilipídica e metabolismo celular Voltando à origem das biomoléculas, além de apresentarem novas propriedades físico-químicas na água, as biomoléculas foram produzidas agregando energia retirada da atmosfera primitiva. As primeiras células primitivas, possivelmente, foram selecionadas em função da capacidade de utilizar estas biomoléculas para sua arquitetura celular, diretamente ou indiretamente, por meio de remontagem utilizando parte delas. Para todas estas transformações, união e quebra de ligações químicas, é preciso energia. O surgimento de enzimas, moléculas capazes de atuar como catalizadores, diminuindo a energia de ativação necessária para tais reações, favoreceu as transformações dependentes de energia nos seres vivos. Em posse de biomoléculas ricas em energia, da construção em membranas e das enzimas, como é possível retirar e conservar a energia destas moléculas, mantê-las organizadas gerando um processo de fluxo contínuo gerador de energia e, ainda, outro meio utilizador dessa energia? Pelo princípio da termodinâmica, a energia da natureza se conserva e pode ser transformada. Em outras palavras, para realizar trabalho precisamos de uma fonte de energia para, a partir dela, fazer as transformações biológicas necessárias: fonte de energia (luz, química) – transformação. (Figura 5) ATP NADH2 FADH2, etc. Energia Resíduos ou produtos metabólicos liberados. Trabalho celular: Aminoácidos, DNA, lipídios, etc. Transformações químicas. Figura 5: Fonte de energia. Essas transformações só se tornaram possíveis por conta do ambiente aquático, das enzimas (excelentes catalisadores biológicos), do alto nível de energia contido nas biomoléculas, do desenvolvimento de uma forma de agregar e estocar esta energia e transferi-las para outras reações químicas, como as biomoléculas ATP, NADH, FADH2, fosfocreatina e outras. A célula trabalha segundo a primeira lei da termodinâmica, a chamada lei de conservação de energia. Exemplificando: oxidamos moléculas ricas em energia obtida do ambiente, aproveitando a energia de suas ligações químicas e transferindo-a para manter e sintetizar nossas biomoléculas também ricas em energia. Então, se somos transformadores químicos de energia, podemos nos considerar organismos químicos? 226 Módulo II — Processos biológicos na captação e na transformação da matéria e energia P Eixo Biológico BSC Não, certamente que não, pois utilizamos biomoléculas organizadas em organelas, células e tecidos que são mantidos às custas de transformações químicas e da utilização de sua energia. Pensar, agir e enxergar são ações biológicas processadas e mantidas pela energia retirada de moléculas ricas na chamada energia ingerida, como os carboidratos. E como nossas biomoléculas, então, conseguem utilizar estas moléculas energeticamente ricas e manipular sua energia? Se podemos transferir energia química, podemos, também, sintetizar qualquer tipo de composto químico? Para obtermos essa resposta, precisamos retomar nossa construção de uma célula metabólica funcional. A nossa “célula”, ao constituir a membrana e se tornar fechada, isolou seu ambiente das reações de fora (meio externo) pela barreira “impenetrável” da membrana. Por ela, podem passar apenas pequenas moléculas, principalmente de natureza apolar. Curiosidade A diferença de potencial ocasiona o deslocamento espontâneo de cargas elétricas. Este fenômeno ocorre ao ligar um aparelho elétrico e pode ser observado também entre as nuvens carregadas e a superfície da Terra, o que provoca a descida espontânea da carga até o solo e vice-versa, ou seja, o relâmpago. As reações que se processaram lá dentro resultaram na alteração do pH por meio da seleção de moléculas tampões que mantêm um pH mais apropriado para as reações químicas. Foram, também, desenvolvidas as membranas internas, pequenas micelas de bicamada, em que reações especiais compartimentalizadas geraram funções específicas (as organelas). Aprisionados desta forma, nesses compartimentos, foram acumulados e até gerados novos compostos, por meio de ações metabólicas, tornando o meio interno diferente do externo. Esse conflito tornou o meio interno mais energético do que o externo, ou seja, o ambiente interno passou a possuir um ∆G0 maior e carregado de moléculas mais concentradas em energia. Quando esse meio interno adquiriu, então, constituintes diferentes, com cargas e um pH diferente, no que isso resultou termodinamicamente falando? Você se lembra do sistema gerador de energia elétrica? Da pilha química? Pois é, a célula utiliza o princípio semelhante ao sistema gerador de energia elétrica, o qual acumula água em um âmbito mais alto para gerar trabalho e transformá-lo em energia. Acontece com a pilha química, também, em que colocamos em contato, por um fio ou ligado a um aparelho elétrico como um rádio, a dois compostos em compartimentos isolados, um de alto nível de energia (baixo Redox) e outro de baixo nível e ávido por elétrons (alto Redox), forçando a transferência dos elétrons ao acoplamento de reação de oxidação-redução. A célula fechada do ambiente externo, pelos diferentes compostos gerados, tornouse mais concentrada e em conflito com o meio externo, mais pobre e diluído. Pela segunda lei da termodinâmica, o compartimento celular mais concentrado para se tornar estável, tem de difundir e distribuir seus compostos uniformemente pelos ambientes. Mas ao se concentrar, a célula cria uma diferença de potencial, sendo este potencial o de concentração que pode ser utilizado para gerar trabalho útil e energia. Assim, um composto rico em energia pode ser obtido e agregado contra a corrente interna de concentração que possui enzimas transportadoras localizadas estrategicamente na membrana. Dessa forma, a célula pode lançar mão dessa diferença de potencial, trazer o composto energético para dentro de si, levando o que tinha de mais concentrado em seu interior para fora, acoplado à entrada desse composto energético. Além disso, os íons, liberados da quebra de biomoléculas e os retidos internamente, pela sua incapacidade de atravessar a barreira lipídica, por serem solúveis em água, geram um potencial por sua concentração e uma resultante de cargas diferente do meio externo. Consórcio Setentrional de Ensino a Distância 227 B # M2U3 Metabolismo Observa-se na célula, ou nas faces da membrana de compartimentos, uma verdadeira pilha, contendo uma apreciável diferença de potencial eletroquímico (Figura 6). Figura 6: Modelo de potencial eletroquímico entre a membrana interna e a externa da mitocôndria. Assim, usando a segunda lei da termodinâmica, esse potencial eletroquímico pode ser utilizado para permuta de compostos com mesma carga presentes no exterior da membrana. Quando o composto ou íon tem duas cargas, ele pode ser trocado por um cátion bivalente ou por dois monovalentes. De acordo com a força iônica destes íons, a troca pode não ser eqüitativa, como acontece com o sódio e o potássio. Usando a energia do ATP e a enzima ATPase, a célula traz para o seu interior dois íons K+ e envia para fora 3Na+, auxiliando a manutenção do ambiente interno, relativo à maior concentração de K. Além desse facilitador para o transporte, a diferença química das concentrações dos elementos químicos transportados (cátions ou ânions), também, é evidenciada pela membrana. Quando estes elementos químicos estão mais concentrados em um meio físico, sua concentração pode ser utilizada como energia direcional para efetuar transporte. Para esses transportes, a célula utiliza a energia contida no meio ambiente, gerada pelo seu metabolismo e pela diferença de concentração dos meios interno e externo. Ou seja, a membrana (célula) é capaz de diferenciar um elemento químico específico e usar tanto sua concentração diferencial nos meios interno/externo, quanto a sua carga para realizar trabalho. Este trabalho é, basicamente, utilizado para o transporte de substâncias, não sendo utilizado para a biossíntese molecular, exceto para a síntese do ATP, por meio de um conjunto de enzimas e estruturas especiais existentes nas membranas internas das mitocôndrias, dos cloroplastos e nas membranas dos procariotos, um conjunto de reações denominado de fosforilação oxidativa. Estudaremos este conceito detalhadamente nos próximos módulos, mas podemos, resumidamente, dizer que consiste na geração de uma diferença de potencial na membrana pela concentração de prótons (H+), sendo que a energia para o seu acúmulo vem da oxidação das biomoléculas energéticas (carboidratos, lipídeos, etc.). Os prótons são lançados para um meio externo e podem ser retornados para dentro do compartimento por uma enzima especial (F0F1 ATPase). Esta enzima é capaz de acoplar a energia deste transporte à catalise da reação ADP + Pi → ATP + H2O. 228 Módulo II — Processos biológicos na captação e na transformação da matéria e energia P Eixo Biológico BSC O ATP (adenosina tri-fosfato) é a moeda universal de energia livre em sistemas biológicos e é constantemente formado e destruído. (Figura 7) NH2 N O- O P O OO P OO O P HC O CH2 O H H H OH OH C C N O C N N CH H Figura 7: Estrutura molecular do ATP. Para a maioria das reações de biossínteses endergônicas (consumidoras de energia), executar alguns trabalhos osmóticos e mecânicos, a energia de catálise precisa ser fornecida pelas moléculas transportadoras intermediárias de energia, como o ATP e seu análogo o GTP, o NADH, FADH2 e NADPH. O NADH e o FADH2 são os principais carreadores de energia bruta na forma de elétrons, retirados das moléculas ricas durante a oxidação das moléculas alimentares. Esta transferência de energia é uma obrigatoriedade, uma lei, conhecida como Lei de Lipmann. Então, a célula trabalha contra o princípio número 1 da lei da termodinâmica? O que você acha disso? #M2U3 IV. Moeda de energia, o ATP Em virtude da necessidade de realizar reações diversas entre construção de biomoléculas e estruturas (anabolismo) e guardar energia, as células que desenvolveram um sistema que permitisse acumular substâncias ricas em energia, contra um gradiente de concentração, transformando energia química em trabalho químico e físico foram favoravelmente selecionadas. A célula vence os princípios que podem ser vencidos pelo aporte de energia, mas não consegue burlar todos eles. Para acumular substâncias, ela não pode vencer a barreira do equilíbrio químico e das reações, ou seja, a concentração máxima alcançada pela célula depende da energia livre da substância. Em outras palavras, a célula trabalha em observância das leis da química e da física. Para vencer algumas barreiras, ela precisa de energia. Para concentrar substâncias dentro de si, ela utiliza um sistema de transporte que implica o gasto de energia, mas se esse pro- Consórcio Setentrional de Ensino a Distância 229 B # M2U3 Metabolismo duto possui passagem facilitada pela barreira da membrana, sua concentração dependerá de um equilíbrio entre o sistema interno e externo. Se esse princípio difusor fosse o único sistema facilitador do transporte de nutrientes utilizados pela célula, ela sobreviveria somente por um fluxo constante pelo meio externo destas moléculas energéticas e, morreria quando houvesse longas interrupções deste fluxo. Durante o processo evolutivo, algumas células não ficaram dependentes dessa variação de concentração dos nutrientes, por terem desenvolvido um sistema transportador contra-corrente, uma enzima. (Figura 8) Figura 8: Enzima de membrana de sistema de transporte contra-corrente com gasto de energia (ATP). A enzima transportadora, posicionada sobre a membrana, possui um “canal”, uma forma de colocar em contato o meio externo com o interno e pode transportar íons ou moléculas ricas em energia contra a corrente de concentração, o que ocorre com gasto de ATP. Ao acoplar a energia do ATP à reação de transporte, a enzima consegue passar o composto adsorvido à superfície externa para o meio interno, por meio de mudanças na sua conformação. Na verdade, é um processo mais complexo que só poderemos estudar mais a frente, após conhecermos mais sobre a estrutura das proteínas. Mas explicando de maneira mais simples, o processo ocorre quando a enzima entra e contato com o composto do meio externo, ela catalisa a quebra da molécula do ATP em ADP + Pi e a energia desta reação provoca o fechamento da “porta” externa e a abertura da “porta” para o meio interno, liberando o composto para dentro da célula. Funciona como se fosse um alçapão, uma armadilha molecular. Como resultado do acúmulo de íons e da alteração do pH, a molécula energética que entrou encontra um ambiente químico diferente. Ela pode se tornar menos ou mais ávida por elétrons, pode sofrer ionização, passando a ter um nível de energia ainda mais elevado, devido à mudança da sua organização eletrônica, que pode potencializar reações. Contudo, para vencer a entropia do sistema e concentrar substâncias, a célula precisa investir energia. Tal energia tem de ser retirada de outras reações químicas e embora todas as biomoléculas possuam energia, nem todas podem ser utilizadas para este fim. 230 Módulo II — Processos biológicos na captação e na transformação da matéria e energia Saiba mais O sistema contra-corrente é amplamente utilizado pelas células, não somente na produção de energia, mas também para manter o estoque de substâncias e íons, como, por exemplo, as células nervosas que acumulam neurotransmissores e utilizam o gradiente eletroquímico do sistema de sódio e potássio. P Eixo Biológico BSC Muitas moléculas são mais valiosas e devem ser preservadas, como as estruturais das membranas e ribossomos. Outras possuem um nível de energia baixo ou são de difícil degradação como os gangliosídeos e o colesterol ou são escassas na natureza. Na evolução, as moléculas ricas em energia, como o amido e a celulose (fontes de glicose), direcionaram e agiram como seletivas no desenvolvimento metabólico dos animais. Assim, os primeiros animais hábeis em degradar as biomoléculas fonte de açúcares tiveram mais sucesso no curso da evolução, de forma que todos os seres vivos, dos microrganismos aos mamíferos superiores, possuem um conjunto de enzimas hábeis para degradar a glicose, organizadas numa cadência de eventos bioquímicos, denominada de glicólise ou via glicolítica. Antes de vermos como se organizaram estas e outras vias de metabolismo, vamos voltar à compreensão da captação de energia das moléculas e do sistema de produção de trabalho. #M2U3 V. Fosforilação no nível do substrato Afinal, por que algumas moléculas possuem uma capacidade reativa maior dentro da célula que no meio externo? Os átomos, ao se ligarem, tendem a “saturar” seus orbitais, os completando com elétrons para alcançar estabilidade, e estas ligações possuem diferentes “forças” de acordo com a natureza dos ligantes. Os compostos de carbono possuem propriedades especiais e uma delas é a ressonância dos elétrons ao redor dos ligantes, ou seja, da cadeia carbônica. Com relação às ligações, envolvendo os elementos C; O; N; P e S, geralmente o carbono é o átomo central das moléculas orgânicas e a presença de outros átomos, principalmente associados ao oxigênio, como os fosfatos (—H2PO4), sulfatos (—H3SO3), ácidos carbônicos (COOH) e nitratos (—HNO3), pela natureza eletronegativa do oxigênio, tende a atrair a nuvem eletrônica que gira ao redor da cadeia carbônica, deslocando-a para próximo de si. Quanto mais eletronegativo o grupo químico ligado à cadeia dos carbonos, maior é a atração da nuvem eletrônica, maior o conflito na ressonância dos elétrons e maior é a tensão entre os átomos para reter seus elétrons. Quanto maior essa tensão, mais energia conterá a ligação entre estes ligantes. Se o pH celular for favorável à ionização do grupo, como acontece com os fosfatos (—PO4-), os ácidos carboxílicos (COO-) e o grupo amino (—NH3+), maior ainda será essa tensão eletrônica. Estes grupos, comuns nas moléculas orgânicas, tendem a se ionizar no pH fisiológico. Assim, uma molécula, ao entrar na célula, pode se ionizar e aumentar a energia contida em suas ligações químicas e, além da ionização, liberar prótons H+. O citossol, tornando-se mais ácido, vai se diferenciando do pH externo, acirrando o gradiente de prótons e íons entre as membranas (lado interno e o meio externo), isto é, a diferença de potencial. Bem, a nossa célula pode, então, transportar moléculas utilizando as enzimas transportadoras e como energia, a translocação de átomos, e íons ou utilizando a energia do ATP. A molécula que entrou pode ou não se dissociar e elevar seu nível de energia. Mas para degradá-la (catabolizar) e retirar energia do processo de desmontagem, é preciso patrocinar simplesmente a quebra de ligações, fazendo reações de óxido-redução, o que resulta em um processo de liberação de grande quantidade de energia, ou seja, em uma verdadeira combustão, que provocará a morte da célula. Consórcio Setentrional de Ensino a Distância 231 B # M2U3 Metabolismo Dessa forma, o processo de quebra deve ser gradual e controlado. Mas a quebra pode resultar, também, na liberação de pouca energia útil aproveitada ou gerar uma molécula que não possui um conflito eletrônico (de baixa energia), o qual favoreça a continuação do processo de óxido-redução dos compostos oxidantes da célula. E mais, “queimar” diretamente a molécula com reações enzimáticas consecutivas de óxido-redução, pode gerar uma molécula intermediária de ∆ G0 baixo, parando as reações catabólicas, pelo fato do ∆ G0 ser muito inferior ao do NAD+, FAD+ ou ADP. Como conseguir obter um alto rendimento energético, contínuo destas oxidações, até a queima total em CO2? Para entender como isso ocorre, observe na tabela o ∆ G0 (Tabela 1 ) de alguns compostos químicos da célula e, em especial dos derivados da glicose. Tabela 1 - Energia Livre-padrão de Hidrólise de alguns compostos fosfatados. ∆G 0´ kcal/mol Fosfoenolpiruvato 3-fosfogliceroilfosfato ( -14,8 3-fosfoglicerato + P i ) fosfocreatina -11,8 -10,3 ADP ( AMP + P i ) -7,3 ATP ( ADP + P i ) -7,3 AMP ( adenosina + P i ) -3,4 Glicose 1-fosfato -5,0 Frutose 6-fosfato -3,8 Glicose 6-fosfato -3,3 Glicerol 1-fosfato -2,2 Quando um composto entra na célula, mesmo que se torne mais energético, as enzimas trabalham sobre ele, aumentando seu nível de energia e adicionando grupos que contenham um dos radicais citados acima, em especial o grupo fosfato (um tipo de oxidação). Este grupo irá, então, atrair os elétrons da nuvem eletrônica da molécula, aumentando o conflito pelos elétrons e o estado de energia das ligações. Se a energia contida não for alta o suficiente para gerar uma molécula de reserva de energia como o ATP (Figura 9), pode-se aumentar seu estado energético aumentando o conflito pela nuvem eletrônica, por meio da remoção de prótons e da geração de duplas ligações entre carbonos e entre carbono e oxigênio, ou adicionando novos radicais como o próprio fosfato. Quanto mais eletrofílico for o grupo adicionado, maior será a “disputa” pelos elétrons e, conseqüentemente, mais energia renderá a quebra das ligações onde houver este conflito. Alterações desse tipo são freqüentes no nosso catabolismo e ocorrem geralmente para a produção de moléculas de energia como o ATP, em reações denominadas fosforilação no nível de substrato. Explicando de outra forma, quando se insere um fosfato (H2PO4) em uma molécula como um carboidrato, fazemos uma oxidação que pode resultar no aumento da energia deste carboidrato. 232 Módulo II — Processos biológicos na captação e na transformação da matéria e energia Saiba mais Um dos importantes eventos no qual ocorre a fosforilação em nível do substrato é o Ciclo de Krebs, uma rota anfibólica (ou seja, anabólica e catabólica), realizado na mitocôndria, fazendo parte de uma das etapas da respiração celular, responsável pela degradação da molécula de piruvato e produção, com isso, produção de energia utilizada na cadeia de elétrons. P Eixo Biológico BSC Catabolismo + Figura 9: Transferência de força redutora das reações catabolicas àquelas biossintéticas, por meio do ciclo do NADP. Para gerar um ATP, pode-se transferir diretamente este grupo fosfato para a molécula do ADP, caso a ligação fosfato no carboidrato tenha um nível de energia maior que a da reação ADP+PO4 →ATP (∆ G= 31kJ mol-1; fosforização a nível de substrato). Caso tenha uma energia menor, a molécula precisa ser modificada pela adição, de duplas ligações com remoção de prótons, elevando o nível de conflito eletrônico e o ∆ G0 da ligação. Daí o grupo fosfato de alta energia pode ser transferido para o ADP. A energia excedente é perdida na forma de calor. Passando para um exemplo real, um dos passos que veremos a seguir é o da degradação da glicose, a cadeia de 6 carbonos é quebrada em duas moléculas de 3 carbonos, cada uma ligada a um grupo fosfato de baixa energia, o 3fosfoglicerato (3Pgli) (Figura 10) 2 Figura 10: Molécula de 3-fosfoglicerato, mostrando os grupos com carga fosfato e ácido. Apesar da molécula do 3Pglicerato possuir dois grupos eletrofílicos fortes (fosfato e carboxíla), cada um atrai a nuvem eletrônica para si, sem muita resistência do grupo – OH do carbono 2. Dessa forma, a energia contida na ligação do carbono e do fosfato (-CH2~P04) contém pouca energia, não permitindo a transferência do grupo fosfato para uma molécula do ADP. Então, para “enriquecer” a ligação com o fosfato, ele é translocado para o carbono 2 que passa a disputar mais acirradamente a nuvem eletrônica, portanto, com aumento do ∆ G. Saiba mais Isômeros são substâncias químicas com diferentes propriedades, mas que possuem a mesma fórmula. Entretanto, como a energia ainda não é suficiente, a enzima enolase retira dois prótons deixando uma dupla ligação entre os carbonos. Como essa dupla ligação possui uma forte atração de elétrons, provoca uma redistribuição dos elétrons da nuvem eletrônica, aumentando a tensão das ligações, com conseqüente aumento da energia da ligação C~P, tornando-a capaz para transferência para o ADP. O conteúdo energético das ligações químicas é, então, fundamental para governar as reações e indicar a importância das biomoléculas. A conservação da energia química retirada do catabolismo das moléculas na forma de ATP, GTP, etc., é uma forma ativa das células conservarem a energia química retirada das reações (Figura 11). Consórcio Setentrional de Ensino a Distância 233 B # M2U3 Metabolismo 3-Fosfoglicerato (2) P O CH 2 CH C O O OH 2-Fosfoglicerato (2) 2H 2 O CH 2 CH OH O O P P 2 Fosfoenolpiruvato (2) CH 2 2ADP C O O O O C C O O O O O O P P 2 ATP ATP O O O O Piruvato (2) CH 3 C O O C Figura 11: Transformação na molécula do 3-Fosfoglicerato para aumentar seu conteúdo energético e obter energia para síntese do ATP. O O Alguns princípios podem ser tirados quanto ao metabolismo, à capacidade metabólica das biomoléculas e às ligações ricas em energia: a) Quando as ligações químicas, ricas em energia, são de igual número e teor nas moléculas dos reagentes e produtos, a reação de transferência de energia (grupos funcionais) pode acontecer em qualquer sentido, como ocorre a transferência do grupo da creatina fosfato para o ADP, na reação Creatina~P + ADP → Creatina-OH + ATP. Quando o substrato e o produto possuem o mesmo número de ligações pobres em energia, a reação também pode acontecer livremente, como entre isômeros químicos: Gli-6P ↔ Gli-1P. b) Quando o total de energia das ligações do substrato é maior que a dos produtos, a reação tende a ocorrer livremente, liberando o saldo de energia, geralmente na forma de calor (exergônica). São reações do tipo Gli + ATP → Gli 6P + ADP. O mesmo se pode dizer quanto ao saldo energético das reações de quebra de compostos pobres em energia. (Figura 12) c) Quando, ao contrário, os produtos possuem mais ligações químicas ricas em energia que os substratos (endergônica), a reação tende a não acontecer. Para que a reação se proceda é necessário doar energia através, geralmente, do ATP, O doando diretamente o grupo fosfato ou a sua energia (Figura 13). O p O H O H C OH H2C O H C C H HO C C OH H HO H C C H OH H H C C H C H 2C H 2 CO O O OH H2C C OH O O H HO H C C H OH OH OH H H C C OH OH OH H C H 2C OH O H 2 CO O -p p O O O Glicose-6-P Glicose-6-P O - O - H2C O ATP ATP ADP ADP HO H C C H OH H H C C OH OH H C H 2C OH p O O Frutose-6-P Frutose-6-P O - O - O - O O H H 2 CO O -p - O C C HO O O p O H2C C C C HO O O O O -p p O - O - Frutose-1,6-BP Frutose-1,6-BP Figura 12: Reação endergônica da primeira fase da degradação da glicose. 234 Módulo II — Processos biológicos na captação e na transformação da matéria e energia P Eixo Biológico Gliceraldeído-3-fosfato (2) BSC O P O CH 2 2P i CH C H OH 2NAD + 2 NADH + H + 1,3-Bifosfoglicerato (2) O P O CH 2 CH C O OH 2ADP P 2 ATP 3-Fosfoglicerato (2) O P O CH 2 CH C O OH 2-Fosfoglicerato (2) O CH 2 CH OH O C O P 2H 2O O Fosfoenolpiruvato (2) CH 2 2ADP C C O O P 2 ATP Piruvato (2) O CH 3 C O C O Figura 13: Reação exergônica da segunda fase da gicólise. (reação do 1,3 Bifosfoglicerato + ADP→ 3-fosfoglicerato + ATP Consórcio Setentrional de Ensino a Distância 235 B # M2U3 Metabolismo Os tipos de reações químicas mais comuns utilizadas para as transformações metabólicas, são: óxido-redução, transferência, hidrólise, eliminação, isomerização e ligação. Você se lembra destes tipos de reações? Procure, então, fazer uma síntese de cada uma delas elaborando exemplos. #M2U3 VI. Atmosfera primitiva e obtenção de energia Falamos muito, aqui, que o catabolismo é um meio pelo qual a célula obtém energia e a transfere de nutrientes para as moléculas transportadoras intermediárias de energia como o ATP e o NADH. Falamos, também, que esta ação é catalisada pelas enzimas e que este catabolismo deve ser feito de modo controlado, em passos, para evitar uma combustão descontrolada da célula, provocando a liberação de calor em excesso e a morte celular. Mas, pensando neste ambiente primitivo, reflita: não era possível catabolizar uma molécula como um carboidrato até a produção de CO2. Isto só foi possível, certamente, depois que encontravam-se nas células todos os elementos necessários para esta conversão química, ou seja, todas as enzimas e recursos celulares compatíveis para esse catabolismo. Lembre-se de que a atmosfera rica em oxigênio foi fruto da atividade fotossintética dos vegetais (algas e plantas), surgidos milhões de anos mais tarde. Mas sem oxigênio, como as células primitivas conseguiam obter energia suficiente para os seus processos celulares metabólicos? Quais poderiam ser os produtos finais do metabolismo? Pesquise sobre a importância da biolixiviação para a mineração e a formação geológica (rochas) do globo terrestre. Provavelmente, a reunião de todas as enzimas ou seus respectivos genes, que possibilitaram a queima de moléculas de carbono ricas em energia até CO2, foi fruto da fusão de células diferentes ou da troca de material genético entre elas, como acontece com as bactérias atuais. Vamos imaginar o ambiente primitivo com água, metais reduzidos, ausência de O2 e atmosfera rica em Nitrogênio e CO2 resultante da atividade vulcânica. Para a obtenção de energia no ambiente primitivo, limitado às biomoléculas, as primeiras células desenvolveram sistemas de remoção de energia da fonte mais abundante, do sol e dos compostos químicos das rochas. A partir destas fontes, as algas e as plantas foram capazes de sintetizar todos compostos orgânicos necessários, por isso foram denominados de seres autotróficos. Como retiram energia da luz, são chamados de seres fototróficos. No primeiro sistema fototrófico, a energia da luz é utilizada para gerar energia e reduzir o CO2 como fonte de carbono para construção de suas biomoléculas, estes seres são chamados de fotolitotróficos (Tabela 2). Complete a tabela abaixo: Tabela 2 Classificação dos microrganismos em relação a fonte de energia Tipo de fonte de energia Fototróficos Fotolitotróficos Autotróficos Heterotróficos Químiolitotróficos Foto-organotróficos Quimiorganotróficos 236 Módulo II — Processos biológicos na captação e na transformação da matéria e energia P Eixo Biológico BSC Outros microrganismos apresentaram a habilidade de retirar energia de uma outra fonte abundante, os elementos reduzidos das rochas. Eles utilizavam as enzimas para retirar energia (elétrons) por meio de reações de óxido-redução de compostos inorgânicos (principalmente dos metais), obtendo, então, energia para utilizar o CO2 como fonte de carbono. Estes seres são chamados de quimiolitotróficos. Estes últimos foram muito úteis para a dissolução das rochas, liberando os mais diversos componentes químicos e liberando grande quantidade de matéria orgânica, possibilitando a vida de novos seres especialistas, que puderam utilizar as biomoléculas de carbono, construídas a partir do CO2 fixado, os foto-organotróficos, como as algas Rhodopseudomonas e os quimiorganotróficos, hábeis em oxidar matéria orgânica reduzida e dela retirar energia e carbono, como a Escherichia coli e todos invertebrados e vertebrados. A energia luminosa e química possibilitou, por milhares de anos, a existência de variados microrganismos na atmosfera primitiva e a experimentação de variadas formas de enzimas, biomoléculas, organização e funcionamento destes componentes na célula. www. Rhodopseudomonas é um gênero de cianobactéria, para saber mais sobre estes organismos, visite o site: http://www.enq.ufsc.br/labs/probio/disc_eng_bioq/ trabalhos_pos2004/microorganismos/CIANOBACTERIAS.html A fotossíntese, nestes microrganismos, ocorre em um processo pouco diferente do que observamos nos vegetais, afinal eles possuem organelas até maiores que estes microrganismos, como os cloroplastos. O princípio deste funcionamento é muito semelhante entre as bactérias e vegetais, mas essas reações serão vistas apenas no nosso próximo módulo, pois ainda precisamos conhecer alguns princípios básicos do metabolismo. Explicando de forma resumida, a luz é a fonte de energia utilizada para a quebra da molécula de água (fotólise) em H20 → ½ O2 + 2H+ , por um conjunto de enzimas especiais, que carregam um átomo de cloro central em um componente não protéico chamado de coenzima. A energia obtida a partir desta reação é utilizada pelos vegetais para reduzir o CO2 e convertê-lo em glicose. Essa energia, também, é utilizada para a produção de intermediários ricos em energia que podem ser prontamente utilizáveis pelas células para trabalho bioquímico, como as moléculas do ATP, NADH, FADH2, NADPH2, etc. Podem, também, sintetizar biomoléculas, realizar transporte, movimento muscular e variadas transformações químicas que precisam energia. À luz destes novos conhecimentos sobre bioquímica, reflita: “A descoberta de água em marte e da existência de atmosferas oxidadas em outros planetas é um indício da possibilidade de vida fora do globo terrestre”. Vamos, ainda, continuar imaginando a atmosfera primitiva, agora já mais adiantada, aonde as descargas elétricas geravam biomoléculas que despejavam o nitrogênio no mar e, como decorrência da atividade metabólica dos microrganismos, uma grande quantidade de ácidos orgânicos e inorgânicos estavam sendo constantemente despejados, liberando uma grande quantidade de metais e outros átomos, como o oxigênio (Figura 14). Essa “sopa” de substâncias solubilizadas das rochas (oxidadas) e de diversas moléculas orgânicas e sintetizadas permitiu o surgimento de outros microorganismos fotolitotróficos e quimiolitotróficos, muito importantes para os dias atuais. Com a redução da disponibilidade de fonte de carbono na água do mar, produzida pelos microrganismos fotolitotróficos e quimiolitotróficos, surgiram organismos hábeis em oxidar estas moléculas orgânicas e dela retirar energia para seus processos. Tais microrganismos são denominados fotolitotróficos e quimiorganotróficos. Consórcio Setentrional de Ensino a Distância 237 B # M2U3 Metabolismo Metal reduzido ou LUZ Energia Trabalho bioquímico (síntese de Biomoléculas; fixação CO2, fotólise H2O) Elétrons e- Aceptor final (redução) Metal oxidado Figura 14: Redução do metal e acepção dos elétrons. Os elétrons retirados no catabolismo destes compostos pelos microorganismos fotolitotróficos e quimiorganotróficos foram endereçados para compostos inorgânicos oxidados, como o óxido de cobre, oxalato de cálcio, óxido de ferro, etc. levando-os a diferentes estados de redução ou, até mesmo, ao seu estado elemental. Estes átomos ou metais dissolvidos na “sopa” química do mar funcionaram (e ainda funcionam) para esta classe de microrganismos, como aceptores finais de elétrons da oxidação dos compostos orgânicos. O resultado desta ação redutora foi a síntese de compostos inorgânicos que se depositaram no leito do mar, formando grandes sedimentos, como podemos observar hoje nas jazidas de cálcio nas rochas calcárias (ardósias, mármores) que formam as diversas grutas calcárias espalhadas pelo globo, e de várias jazidas de minérios. Você já havia relacionado alguma vez o porquê da concentração de alguns destes elementos minerais em determinadas regiões do nosso planeta? É claro que os microrganismos não foram os responsáveis pela concentração de minerais de todas as jazidas observadas atualmente, como as jazidas de calcário. Muitas jazidas foram resultantes do resfriamento gradual do planeta quando a temperatura ambiente atingiu o ponto de condensação de modo seletivo e gradual, como as jazidas de minério de ferro, mas, muitas das jazidas de minerais, podemos creditar as atividades dos microrganismos. Baseado no que acabamos de estudar, você compreende agora a importância que os cientistas dão à pesquisa da constituição e à estrutura dos meteoritos? Curiosidade Como resultado das diversas formas de vida no ambiente primitivo e explorando formas distintas de organização celular metabólica, surgiu a evolução de uma outra forma celular, as células eucarióticas, cuja organização celular é mais complexa e agregam princípios funcionais iguais aos das células procarióticas, as bactérias. Estes seres, diferentemente evoluídos, se adaptaram para controlar o processo de crescimento e diferenciação dos tecidos, mas, metabolicamente falando, os processos bioquímicos mantiveram muito do que os seres primitivos desenvolveram. Graças a esta forma de “como fazer” dos organismos primitivos, temos hoje variadas formas de enzimas e coenzimas hábeis em metabolizar diferentes compostos, em condições adversas. Reportando a nossa célula funcional, agora que vimos que a retirada de energia química pode gerar trabalho, e, que os compostos orgânicos reduzidos são fonte dessa energia, precisamos entender melhor como ocorre o controle dessa produção ou a regulação do metabolismo (Figura 15). 238 Módulo II — Processos biológicos na captação e na transformação da matéria e energia O contato direto da rocha base com a água do mar é chamado de falésia. A partir disso, a ação do intemperismo (físico e químico), com o constante e lento desgaste da água do mar criam fendas, cavernas, salões e as grutas submersas exploradas pelo turismo de mergulho. P Eixo Biológico BSC Figura 15: Esquema geral de catabolismo e anabolismo. #M2U3 VII. Equilíbrio dinâmico Se considerarmos a fonte de compostos ricos em energia uma constante (alimentação), teremos uma continua oferta de energia. Mas todas as células têm momentos de maior e de menor necessidade de energia, momentos de grande consumo como em pleno exercício físico e de baixo consumo como em repouso. Como é possível o controle dos passos metabólicos e o controle do nível de produção e consumo de energia em nível celular? Para se manter viva, a célula precisa manter um equilíbrio dinâmico e estrito entre os processos de produção e consumo de energia, isto é, um ato contínuo de transformação da energia para realizar trabalho, afinal somos seres transformadores de energia. Além disso, precisamos manter um equilíbrio químico favorável à célula entre os diferentes compostos dos meios externo e interno. Tal equilíbrio é mantido às custas da energia obtida por meio da combustão de outras moléculas, para acumular sais e variadas moléculas energéticas. Como é possível manter esse gradiente de concentração de íons e das moléculas na célula? Isso é possível pelas trocas de íons, como vimos anteriormente, pela impermeabilidade da membrana e de poros existentes na sua superfície. Estes poros são constituídos por proteínas que agem, seletivamente, no transporte das moléculas, também denominadas de enzimas transportadoras. O controle da atividade destas enzimas, permitindo um controle funcional da célula (auto-regulação), e o controle do nível de produção de altas demandas como em pleno exercício físico, e de redução da produção, como durante o repouso ou o sono, são conseqüentes ao controle da atividade destas admiráveis biomoléculas, as enzimas. Consórcio Setentrional de Ensino a Distância 239 B # M2U3 Metabolismo Uma enzima é um catalisador biológico que trabalha em condições moderadas de temperatura e pH para efetuar uma catálise química, sem produzir produtos intermediários ou produtos alterados. Ela é, na realidade, uma proteína que apresenta uma excepcional capacidade de ação funcional. Esta fantástica biomolécula é capaz de assumir as mais variadas atividades na célula que vão desde o catabolismo/anabolismo, ao transporte pelas membranas como vimos anteriormente, passando pela proteção imunológica, etc. Antes de entendermos como ela pode processar estas atividades, vamos entender como é sua estrutura molecular. Essas enzimas, para permitir que as células mantivessem um fluxo contínuo de produção de energia e mantivessem, também, seus processos vitais (catabolizar e anabolizar compostos ao mesmo tempo) precisaram estar sob um controle funcional estrito que adequasse o nível de produção de energia à necessidade de seu consumo para os processos anabólicos. Isto é, um controle metabólico, em que a produção de compostos ricos em energia fosse compatível com seu gasto. Por meio da tradução de novas enzimas, a célula pode controlar o volume global da atividade metabólica, no entanto, o nível em que elas ocorrem é controlado pelo número de unidades ativas ou moduladas destas enzimas. Muitas enzimas possuem apenas um sítio catalítico onde as reações químicas acontecem, mas outras possuem, também, formando parte de sua estrutura biológica, um ou mais sítios de interferência, denominados de sítios de regulação. Assim, quando o organismo necessita reduzir, para determinada reação ou mesmo para uma via metabólica, enzimas reguladas, estrategicamente posicionadas numa via, tem suas atividades moduladas, por meio do acoplamento ao sítio regulador de uma substância reguladora. A enzima ligada covalentemente ou adsorvida a esse composto pode ter sua atividade estimulada ou inibida. Geralmente, as interações covalentes entre modulador e enzima são modulações mais fortes e de longa duração, enquanto as adsorções acarretam efeitos fracos e de pouca duração. Para controle mais efetivo do catabolismo ou anabolismo em uma via metabólica, considerando-se o volume de produtos gerados ou consumidos, a presença de enzimas reguladas, tanto no início quanto no fim do processo metabólico, permite maior controle do gasto de substratos ou da geração do produto final. Assim, é possível controlar o funcionamento e o desempenho das vias metabólicas, respectivamente. Quando uma via metabólica gera, em um dos passos, um composto também utilizado para iniciar outra via de metabolismo (substrato intermediário), a enzima, logo abaixo desta bifurcação, também é regulada, permitindo, simultaneamente, frear uma via e estimular a outra. Os agentes interferentes (inibidores/ estimuladores) usados podem ser agentes químicos diferentes ou mesmo um composto produzido como produto final da via. Quando essa inibição é feita por produtos finais, gerados pela via metabólica, é denominada inibição por feedback ou retroalimentação, muito efetiva para o controle do nível de metabolização, pois quanto mais produto se gerar, maior será o efeito sobre o controle dos passos iniciais da via. 240 Módulo II — Processos biológicos na captação e na transformação da matéria e energia P Eixo Biológico BSC A regulação depende, portanto, da concentração do agente inibidor e pode ser reversível ou temporária. Quando se objetiva uma inibição mais ativa, ou mesmo o bloqueio por longos períodos ou até anos de uma via, os inibidores que se ligam à enzima podem não ser produtos da via metabólica e ligam-se à enzima em sítios especiais e até por ligações covalentes. Dessa forma, para o bloqueio da atividade de uma via, os inibidores não precisam atingir altas concentrações na célula e podem ser lançados em número pouco superior ao número de unidades ativas da enzima em questão (Figura 16). - Figura 16: Passos enzimáticos na via metabólica na síntese do aminoácido isoleucina exemplificando o mecanismo de regulação por feedback. Então, quando o volume de matéria orgânica rica em energia, que é absorvida por um organismo, torna-se constante para ele realizar todas as suas funções metabólicas funcionais (catabolismo, anabolismo, trabalhos químico, osmótico e mecânico), diz-se que o organismo está em homeostase metabólica (Figura 17) HOMEOSTASE: Compostos ricos em energia: Carboidratos, lipídios, etc. Metabolismo eprocessos: osmótico, químicos e mecânicos. Compostos pobres em energia: CO2, ácido úrico, O2, etc. Figura 17: Esquema geral da homeostase metabólica. Consórcio Setentrional de Ensino a Distância 241 B # M2U3 Metabolismo Em uma célula procariótica, a dinâmica de funcionamento apresentada é muito mais simples que numa célula eucariótica. Mas por outro lado, a presença de compartimentos celulares, envoltos por membranas, torna as reações químicas mais ativas e seletivas, permitindo múltiplas ações metabólicas ao mesmo tempo, sem o risco de interferência entre os processos. Entretanto, estes seres eucariotos só surgiram depois que a atmosfera foi inicialmente alterada pelos efeitos ambientais e após a atividade dos microrganismos pioneiros, pois a redução dos gases, a penetração da luz e a atmosfera rica em oxigênio permitiram um surgimento expressivo de novas formas de vida na superfície do planeta. Surgindo os vegetais unicelulares e as plantas sem sementes que dominaram inicialmente a superfície terrestre, permitindo sua colonização por outros seres vivos. #M2U3 VIII. Referências LEHNINGER , A.; NELSON, D.I.; COX, M.M. Princípios de bioquímica. 2ª ed., São Paulo: Savier, 1995 em diante. ROSKOSKI. Bioquímica. Rio de Janeiro: Guanabara Koogan, 1997 STRYER, L. Biochemistry. 4ª. ed., São Francisco: W.E. Freeman and Company, 1995. R. K. MURRAY; et al. Harper’s biochemistry. 25ª. ed., Ed. Appleton & Lange, 2000. MARZZOCO, A.; TORRES, B.B. Bioquímica básica. 2ª. ed., Rio de Janeiro: Ed. Guanabara, 1999. FUTUYMA, Douglas. J. Biologia evolutiva, Ribeirão Preto: Soc. Bras. de Genet, 1997. 242 Módulo II — Processos biológicos na captação e na transformação da matéria e energia Saiba mais As plantas sem sementes englobam algumas algas, briófitas (musgos), pteridófitas (samambaias) e as gimnospermas (eucalipto, pinheiros).
Download