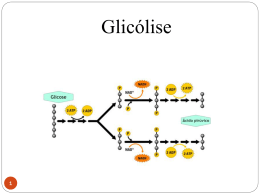
MINISTÉRIO DA EDUCAÇÃO UNIVERSIDADE FEDERAL DO PAMPA CAMPUS DOM PEDRITO BIOQUÍMICA II Unidade 2 - Metabolismo dos Carboidratos Prof ª Drª Angélica Pinho O metabolismo dos nutrientes pode ser dividido por vias metabólicas: Metabolismo dos carboidratos (glicose ou glicogênio muscular e hepático) Metabolismo das gorduras (ácidos graxos) Metabolismo das proteínas (aminoácidos). Pode ser dividido também em relação a presença de oxigênio (metabolismo aeróbio) e na ausência de oxigênio (metabolismo anaeróbio). O que vai definir qual será a via metabólica utilizada para gerar a ressintese do ATP será a velocidade e a intensidade do gasto energético. -Onde tudo inicia Digestão e absorção: Boca- Amido e glicogênio hidratados sofrem a ação da enzima alfa-amilase, presente na saliva, e são reduzidos a estruturas menores. Duodeno - A alfa-amilase presente também no suco pancreático onde os carboidratos de cadeia maior são transformados no monossacarídeo glicose, no dissacarídeo maltose, no trissacarídeo maltotriose e nas chamadas dextrinas alfa-limite. A alfa-amilase: quebra ligação α 1,4; Entretanto, algumas ligações α 1,4 correspondentes aos pontos de ramificação das moléculas glicosídicas não sofrem a ação da alfa-amilase, gerando as dextrinas alfa-limite, contendo uma média de oito unidades de glicose e uma ou mais ligações glicosídicas alfa 1,6. A hidrólise final de dia oligossacarídeos em monossacarídeos é realizada por enzimas de superfície das células epiteliais do intestino delgado (lactase, maltase, alfa-1,6 glicosidase, sacarase) liberando monossacarídeos. Di-, oligo- e polissacarídeos não hidrolisados pela alfaamilase e/ou enzimas de superfície das células epiteliais do intestino não são absorvidos na porção inferior do intestino são metabolizados por bactérias. LEMBRAR!!!! A glicose, oxidada em CO2 e H2O, é nossa fonte primária de energia. DIGESTÃO E ABSORÇÃO DOS CARBOIDRATOS Os principais carboidratos da dieta são: Amido Sacarose Lactose O glicogênio, a maltose, a glicose livre e a frutose livre constituem frações relativamente menores de carboidratos ingeridos. A absorção dos carboidratos pelas células do intestino delgado é realizada após hidrólise dos dissacarídeos, oligossacarídeos e polissacarídeos em seus componentes monossacarídeos. AS QUEBRAS OCORREM SEQÜENCIALMENTE EM DIFERENTES SEGMENTOS DO TRATO GASTROINTESTINAL POR REAÇÕES ENZIMÁTICAS α - Amilase salivar A digestão do amido inicia durante a mastigação pela ação α-amilase salivar (ptialina) que hidrolisa as ligações glicosídicas α(1→4), com a liberação de maltose e oligossacarídeos. Contudo, a α-amilase salivar não contribui significativamente para a hidrólise dos polissacarídeos, devido ao breve contato entre a enzima e o substrato. Ao atingir o estômago, a enzima é inativada pelo baixo pH gástrico. α - Amilase pancreática O amido e o glicogênio são hidrolisados no duodeno em presença da α-amilase pancreática que produz maltose como produto principal e oligossacarídeos chamados dextrinas – contendo em média oito unidades de glicose com uma ou mais ligações glicosídicas α(1→6). Certa quantidade de isomaltose (dissacarídeo) também é formada. Enzimas da superfície intestinal A hidrólise final da maltose e dextrina é realizada pela maltase e a dextrinase, presentes na superfície das células epiteliais do intestino delgado. Outras enzimas também atuam na superfície das células intestinais: Isomaltase: Hidrolisa as ligações α(1→6) da isomaltose Sacarase: Hidrolisa as ligações α,β(1→2) da sacarose em glicose e frutose Lactase: Fornece glicose e galactose pela hidrolise das ligações β(1→4) da lactose. A CAPTAÇÃO DE MONOSSACARÍDEOS DO LÚMEN PARA A CÉLULA INTESTINAL É EFETUADA POR DOIS MECANISMOS Transporte passivo (difusão facilitada) O movimento da glicose está “a favor” do gradiente de concentração (de um compartimento de maior concentração de glicose para um compartimento de menor concentração). A difusão facilitada é mediada por um sistema de transporte de monossacarídeos do tipo Na+− independente. O mecanismo tem alta especificidade para D−frutose. Transporte ativo A glicose é captada do lúmen para a célula epitelial do intestino por um co− transportador Na+−monossacarídeo (SGLT). É um processo ativo indireto cujo mecanismo é envolve a (Na+−K+)−ATPase (bomba de (Na+−K+), que remove o Na+ da célula, em troca de K+, com a hidrólise concomitante de ATP. O mecanismo tem alta especificidade por D−glicose e D−galactose. No intestino, a fosfofrutoquinase fosforila a frutose para prendê-la no interior da célula. Obs: As –quinases são importantes para prender a molécula no interior da célula através da fosforilação. Após a absorção, a glicose no sangue aumenta e as células β das ilhotas pancreáticas secretam insulina que estimula a captação de glicose principalmente pelo tecido adiposo e muscular. O fígado, o cérebro e os eritrócitos, não necessitam de insulina para captação de glicose por suas células (tecidos insulino−independentes). Outros hormônios e enzimas, além de vários mecanismos de controle, são importantes na regulação da glicemia. A frutose e a galactose somente são convertidas em glicose no fígado. Digestão se dá em duas etapas: 1. Degradação de CHO em monômeros (enzimas microbianas extracelulares associadas a membrana) 2. Absorção de monômeros e dímeros para o meio intracelular para degradação até piruvato (via de Embden-Meyerof) Digestão em Monogástricos Intestino Delgado Otimização da Absorção Pancrêas α Maltase, Lipase, α 1,6 Galactosidase Monogástricos Fígado e Rim Otimização dos processos metabólicos Gl. Salivares Ptialina Intestino Grosso Estabilização da flora Estômago Secreta Pepsina e Ác. Clorídrico Atividade enzimática em leitões Locais de absorção - Lactose: duodeno e jejuno prox - Maltose: jejuno e íleo prox - Sacarose: jejuno e íleo dist Velocidade de absorção Galactose>glicose>frutose>lactose Glicólise A glicólise é a via central do catabolismo da glicose e ocorrem no citosol de todas as células humanas. Cada molécula de glicose é convertida em duas moléculas de piruvato, cada uma com três átomos de carbonos em um processo no qual vários átomos de carbono são oxidados. Parte da energia livre liberada da glicose é conservada na forma de ATP e de NADH. A glicólise compreende dois estágios: 1º estágio (fase preparatória) → Compreende cinco reações nas quais a glicose é fosforilada por dois ATP e convertida em duas moléculas de gliceraldeído−3−fosfato. 2º estágio (fase de pagamento) → As duas moléculas de gliceraldeído−3−fosfato são oxidadas pelo NAD+ e fosforiladas em reação que emprega o fosfato inorgânico. O resultado do processo total da glicólise é a formação de 2 ATP, 2 NADH e 2 piruvato, às custas de uma molécula de glicose. Em condições de baixo suprimento de oxigênio (hipóxia) ou em células sem mitocôndrias, o produto final da glicólise é o lactato e não o piruvato, em processo denominado glicólise anaeróbica Quando o suprimento de oxigênio é adequado, o piruvato é transformado em acetil−CoA nas mitocôndrias. O grupo acetil da acetil−CoA é totalmente oxidado no ciclo do ácido cítrico com a formação de duas moléculas de CO2. ACETILCOA Glicólise Mg2+ Mg2+ * Gasto * Gasto 2x Produção Mg2+ Produção * Mg2+ Todas as células utilizam Glicose para gerar ATP Glicólise Oxidação da glicose em piruvato CARACTERÍSTICAS DA GLICÓLISE Ocorre no citosol das células; Gera ATP; Reduz o NAD para NADH; FASE AERÓBICA X FASE ANAERÓBICA Glicose 2ADP + Pi 2 ATP 2 NAD+ 2 NADH + 2H+ (2) Piruvato O2 (2) Lactato Sem ou pouco O2 A Gliconeogênese é o processo através do qual precursores como lactato, piruvato, glicerol e aminoácidos são convertidos em glicose. Durante o jejum, toda a glicose deve ser sintetizada a partir desses precursores não-glucídicos. A maioria dos precursores deve entrar no Ciclo de Krebs em algum ponto para ser convertida em oxaloacetato. O oxaloacetato é o material de partida para a gliconeogênese. Gliconeogênese Jejum Glucagon O principal destino do lactato é a conversão a glicose pela GLICONEOGÊNESE Lactato produzido pelo músculo esquelético ( e outros tecidos) é captado pelo fígado e é convertido em glicose. A Gliconeogênese ocorre principalmente no fígado e em menor extensão nos rins. Síntese da glicose a partir do piruvato - utiliza várias enzimas da GLICÓLISE. Três reações da glicólise são essencialmente IRREVERSÍVEIS: Hexoquinase Fosfofrutoquinase Piruvato quinase PRECURSORES DA NEOGLICOGENESE A gliconeogênese não é simplesmente o inverso da glicólise. Alguns passos são diferentes de tal forma que o controle de uma via não inativa a outra. Passos diferentes da glicólise. Piruvato para PEP Frutose-1,6-bisfosfato para Frutose-6-fosfato Glicose-6-FosfatoparaGlicose O piruvato é convertido a oxaloacetato antes de ser transformado em fosfoenolpiruvato. 1.Piruvato carboxilase catalisa a formação de oxaloacetato apartir de piruvatoeCO2, com gasto de ATP. 2.PEP carboxiquinase (PEPCK) converte oxaloacetato em PEP e usa GTP como agente fosforilador. PIRUVATO CARBOXILASE REQUER BIOTINA COMO COFACTOR PEP CARBOXIQUINASE ACETIL-COA REGULA A PIRUVATO CARBOXILASE Aumentos nas concentrações de oxaloacetato induzem a atividade do ciclo de Krebs e o acetilCoA é um ativador alostérico da carboxilase. Contudo, quando as concentrações de ATP e NADH estão altas e o ciclo de Krebs é inibido, o oxaloacetato vai para a gliconeogênese. VIA DAS PENTOSES A via das pentoses−fosfato é uma via metabólica alternativa à glicólise para a oxidação da glicose que não requer e não produz ATP. Seus principais produtos são: NADPH (nicotinamida adenina dinucleotído fosfato reduzido) um agente redutor empregado para os processos anabólicos. Ribose−5−fosfato um componente estrutural de nucleotídeos e de ácidos nucléicos. A via das pentoses-fosfato ocorre no citosol em duas etapas: etapa oxidativa e a etapa não−oxidativa. Na etapa oxidativa a glicose−6−fosfato é convertida à ribulose−5−fosfato acompanhada pela formação de duas moléculas de NADPH. A etapa não−oxidativa envolve a isomerização e condensação de várias moléculas diferentes de açúcar. Três intermediários do processo são utilizados em outras vias: a ribose−5−fosfato, a frutose−6−fosfato e o gliceraldeído−3−fosfato. Alternativamente, a via das pentoses− fosfato pode ser concebida como um “desvio” para a produção de frutose− 6− fosfato a partir da glicose− 6− fosfato. Tanto a glicose− 6− fosfato como o gliceraldeído−3− fosfato produzidos pela via das pentoses− fosfato podem ser metabolizados a piruvato e, finalmente, oxidado no sistema enzimático mitocondrial. CICLO DE KREBS O ciclo do ácido cítrico (ciclo de Krebs) é o estágio final da oxidação dos combustíveis metabólicos. Os átomos de carbono entram no ciclo na forma de grupos acetila derivados dos carboidratos, ácidos graxos e aminoácidos. O grupo acetila ligado a coenzima A (acetil-CoA) é oxidado em oito reações mitocondriais para formar duas moléculas de CO2 com a conservação da energia livre liberada em três moléculas de NADH, uma de FADH2 e um composto de “alta energia” (GTP ou ATP). O NADH e o FADH2 são oxidados e os elétrons são conduzidos pela cadeia mitocondrial transportadora de elétrons com a liberação de energia conservada na forma de ATP sitetizado a partir de ADP e Pi por meio de processo denominado fosforilação oxidativa. Primeiramente, o piruvato, derivado da glicose e outros açúcares através da via glicolítica, é oxidado à acetil−CoA e CO2 para entrar no ciclo do ácido cítrico. REAÇÕES DO CICLO DO ÁCIDO CÍTRICO O ciclo oxida duas unidades de carbono com a produção de duas moléculas de CO2, uma molécula de GTP, três moléculas de NADH e uma molécula de FADH2. Energia no ciclo do ácido cítrico O ciclo do ácido cítrico é a via oxidativa terminal para a maioria dos combustíveis metabólicos (piruvato, aminoácidos e ácidos graxos). Os dois carbonos do grupo acetila que participam do ciclo são oxidados completamente a CO2 e H2O. A energia liberada por essas oxidações é conservada na forma de três NADH, um FADH2 e uma molécula de GTP (ou ATP). Para cada NADH que transfere seus elétrons para a cadeia mitocondrial transportadora de elétrons, aproximadamente 2,5 ATP são produzidos a partir de ADP + Pi. Para cada FADH2, cerca de 1,5 ATP são produzidos. Assim, a completa oxidação do grupo acetila da acetil−CoA no ciclo do ácido cítrico produz 10 ATP. Obs:. As desidrogenases irão dar os H+ para o NAD+ e FAD, convertendo-os a NADH e FADH (seus cofatores respectivos), durante a fosforilação oxidativa. 2X 3 ATP * 3 ATP * * 2 ATP 3 ATP * Glicose 2 NADH 6 ATP 2 ATP Total 38 ATP Piruvato 2 NADH Acetil CoA CK 6 ATP 6 NADH 18 ATP 2 FADH2 4 ATP 2 GTP 2 ATP A GLICOSE ARMAZENADA SOB A FORMA DE GLICOGÊNIO NO FÍGADO E MÚSCULOS DESTINAM-SE A DIFERENTES FUNÇÕES: Glicogênio hepático Atua como reservatório de glicose para a corrente sangüínea com a distribuição para outros tecidos. Acumula após as refeições e, quando necessário, é degradado lentamente para manter a concentração de glicose no sangue mais ou menos constante. As reservas de glicogênio hepático no homem apresentam importante papel como fonte de glicose no período entre as refeições e, em maior extensão, durante o jejum noturno. Glicogênio muscular Serve como combustível para gerar ATP durante a atividade muscular aumentada. É formado durante o repouso após as refeições. Os níveis de glicogênio muscular apresentam menor variabilidade do que os teores hepáticos em resposta a ingestão de carboidratos. A glicogenólise degrada glicogênio para produzir glicose. Obs: O tecido adiposo também necessita glicose para a síntese de triacilglicerol (glicose Via glicolítica diidroxiacetona-P glicerol-3-P glicerol). REAÇÕES DA GLICOGÊNESE A síntese do glicogênio ocorre após as refeições, quando os teores de glicose sangüínea estão elevados. Até recentemente, presumia-se que a glicose sangüínea era a única precursora direta nesse processo. Entretanto, em condições fisiológicas, grande parte do glicogênio é produzido por um mecanismo envolvendo a seqüência: glicose da dieta → molécula C3 → glicogênio hepático. O lactato e a alanina são as principais moléculasC3 nesse processo. O lactato é formado nos eritrócitos por glicólise e é captado pelo fígado e convertido em glicose− 6− fosfato na gliconeogênese. A síntese do glicogênio se dá a partir da glicose-6fosfato derivada da glicose livre pela ação da glicocinase (no fígado) ou da hexocinase (no músculo). "O saber a gente aprende com os mestres e os livros. A sabedoria, se aprende é com a vida e com os humildes.“ Cora Coralina OBRIGADA!
Baixar