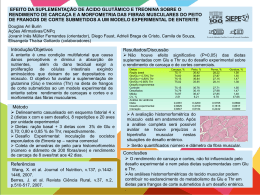
Universidade do Minho Escola de Ciências Ricardo Filipe Duarte da Cruz Indução e Recuperação do Stresse Stresse Hídrico em Variedades Portuguesas de Milho Setembro de 2006 Universidade do Minho Escola de Ciências Ricardo Filipe Duarte da Cruz Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho Tese de Mestrado Biologia do Stresse em Plantas Trabalho efectuado sob a orientação de Professor Doutor Jorge Marques da Silva Professora Professora Doutora Ana Cunha Setembro de 2006 É AUTORIZADA A REPRODUÇÃO INTEGRAL DESTA TESE APENAS PARA EFEITOS DE INVESTIGAÇÃO, MEDIANTE DECLARAÇÃO ESCRITA DO INTERESSADO, QUE A TAL SE COMPROMETE. “Have courage for the great sorrows of life and patience for the small ones; and when you have laboriously accomplished your daily task, go to sleep in peace. God is awake.” Victor Hugo (1802 - 1885) Agradecimentos Agradecimentos Ao Professor Jorge Marques Silva por me ter sugerido o tema deste trabalho, e me ter acolhido prontamente na Faculdade de Ciências da Universidade de Lisboa. Agradeço-lhe também a liberdade que me permitiu ao longo do trabalho e a discussão dos problemas que foram surgindo. Agradeço-lhe ainda a leitura cuidadosa e com espírito crítico do presente manuscrito. À Professora Ana Cunha pela disponibilidade e pelo acompanhamento ao longo de todo o trabalho, em especial o realizado na Universidade do Minho. Agradeço-lhe também pela leitura cuidadosa e crítica do presente manuscrito. Ao Professor Pedro Fevereiro e ao Engenheiro Silas Pego pelo apoio ao projecto e o auxílio na obtenção das sementes. À Professora Anabela Bernardes da Silva pelo auxílio prestado na determinação das actividades da PEPC e EM-NADP. À Professora Celeste Arrabaça pelo auxílio prestado na determinação das actividades da RuBisCO. Ao Banco Português de Germoplasma Vegetal, Braga, pelas sementes das diferentes variedades portuguesas de milho. À Ana Elisabete, à Ana Sofia, à Elsa Elias e ao André Costa pela ajuda imprescindível que me ofereceram ao longo de todo o trabalho, quer na utilização dos aparelhos, quer na metodologia aplicada. Também quero agradecer-lhes a amizade e o apoio ao longo do trabalho. À Alexandra Marques pela discussão dos problemas que foram surgindo, pela amizade e apoio que demonstrou. À Cátia Nunes pela ajuda preciosa na quantificação dos aminoácidos livres e prolina, na Universidade do Minho. À funcionária Manuela Lucas pela ajuda preciosa, pela amizade e apoio que sempre demonstrou. A todos os docentes da Secção de Fisiologia e Bioquímica Vegetais pelo ambiente e apoio que demonstraram ao longo do trabalho. Agradeço à minha família, à Leonor e a todos os meus amigos pelo apoio nos momentos de maior dificuldade. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho I Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho II Resumo Resumo Este trabalho teve como objectivo geral, na primeira fase, caracterizar as estratégias de resposta ao stresse hídrico, com ênfase no metabolismo fotossintético, em seis variedades portuguesas de milho (Zea mays L.) e, na segunda fase, aprofundar o conhecimento sobre o comportamento fisiológico das duas variedades com respostas mais contrastantes. Utilizaramse dois sistemas de indução de stress, um rápido e um lento, e recorreu-se, na primeira fase, à medição das trocas gasosas através da análise de gases por radiação infra-vermelha e de fluorescência modulada da clorofila a. Nas duas variedades seleccionadas para a segunda fase, estudou-se adicionalmente a resposta do aparelho fotossintético ao stress hídrico sob diferentes regimes de PPFD e diferentes concentrações de CO2. Analisaram-se também as actividades das principais enzimas do metabolismo fotossintético (RuBisCO, PEPC, EMNADP) e a variação do teor em prolina. Nos ensaios de indução lenta do stresse ocorreu uma diminuição de todos os parâmetros de trocas gasosas (A, E, gs, WUE) nas seis variedades. Destacaram-se as variedades PB260 e PB269, como aquelas onde os parâmetros sofreram uma diminuição progressiva e mais lenta, e a variedade PB369, onde os efeitos negativos do stresse hídrico se fizeram sentir de forma mais abrupta, com uma diminuição rápida dos valores. As variedades recuperaram os valores iniciais da maioria dos parâmetros após irrigação, exceptuando-se a variedade PB369 cuja recuperação se deu de forma mais lenta. Nos ensaios de fluorescência da clorofila a, verificou-se uma diminuição de vários parâmetros (Fv/Fm, ΦPSII, qP, ETR) ao longo do stresse lento e do stresse rápido. Por outro lado, qN aumentou em todas as variedades, à excepção de PB269, na qual não variou. Comparando os ensaios dos dois tipos de stresse, verificou-se que os parâmetros fisiológicos que diminuíram o fizeram de forma mais marcante no stresse rápido, em todas as variedades. As variedades PB269 e PB369 foram seleccionadas como sendo a mais e a menos tolerante ao stresse hídrico, respectivamente. A partir das determinações dos potenciais hídricos constatou-se que a variedade PB269 conseguiu manter um potencial hídrico mais elevado do que a variedade PB369 em condições de stresse hídrico semelhantes e que possui uma parede celular mais elástica, permitindo a manutenção da turgescência até THR mais baixos apresentando-se esta como uma boa estratégia de tolerância ao stresse hídrico. Através da decomposição do parâmetro de fluorescência qN, verificou-se que houve um aumento de qE com a intensificação do stresse hídrico, na variedade PB369. Por outro lado, na variedade PB269 houve a manutenção deste parâmetro até THR de cerca de 50%, indicando uma maior capacidade de manter activos os processos fotossintéticos sem recorrer a processos de amortecimento não-fotoquímicos. Nos ensaios enzimáticos, observou-se que em ambas as variedades houve diminuição das actividades enzimáticas da RUBISCO, PEPC e EM-NADP com a diminuição do THR, nos ensaios de stresse lento. Contudo, nos ensaios de stresse rápido tal diminuição só aconteceu na RUBISCO, mantendo-se as actividades das outras enzimas constantes. Os dados parecem indicar que ambas as variedades possuem uma reserva de prolina que poderá ter sido rapidamente mobilizada no período de recuperação do stresse, principalmente na variedade PB269. No seu conjunto, os resultados obtidos parecem mostrar que existe um espectro alargado de respostas fisiológicas face ao stresse hídrico que, a ter uma base genética de elevada hereditabilidade e estabilidade, poderão constituir uma base de selecção em futuros programas de melhoramento. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho III Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho IV Abstract Induction and Recovery of Water Stress in Portuguese Maize Cultivars Abstract This work aims for the identification of several aspects of water stress physiological response strategies in different Portuguese maize cultivars (Zea mays L.), in a first stage, and expand the knowledge of the most tolerant and less tolerant cultivars physiological responses to water stress. The responses were studied under two water stresse regimes – slow stress and rapid stress – relevant in field conditions, and in the recovery process after irrigation. In the first part of the work, a preliminary characterization of six cultivars was made measuring gas exchange with an infrared gas analyser and chlorophyll a fluorescence. In the second part of the work, a deeper characterization of those two cultivars was made through the study of water relations and photosynthetic metabolism under different PPFD and external CO2 conditions, the enzymatic activities (RuBisCO, PEPC, NADP-ME) and proline content variation. In slow stress, all the parameters of gas exchange (A, E, gs, WUE) suffered a reduction in all six cultivars. In PB260 and PB269, the reduction was made in a slower progressive way, and, in contrast, in PB369 the parameters declined abruptly. In general, the parameters recovered upon irrigation, with the exception of PB369, in which the recovery was slower. In rapid stress, the parameters responded in a similar but very fast way. PB369 was the only cultivar with a slower response. Chlorophyll fluorescence parameters (Fv/Fm, ΦPSII, qP, ETR) suffered a decrease, in slow and rapid stress. On the other hand, qN increased in all cultivars, with the exception of PB269, in which it maintained its values. Comparing slow and rapid stress, the parameters decreased faster in rapid stress, in all cultivars. PB269 and PB369 were selected as the cultivars more and less tolerant to water stress. PB269 is able to maintain higher leaf water potential under water stress and as a more elastic cell wall that allows it to maintain turgescence at lower RWC, resulting in a good tolerance strategy to water stress. The decomposition of the qN fluorescence parameter, showed a rise of the qE component, in PB369, with increased water stress. PB269 maintained this parameter to a RWC of 50%, indicating a suitable photosynthesis without activating major nonphotochemical quenching processes. On the enzymatic assays, there was a decrease in the activities of RUBISCO, PEPC and NADP-ME, with RWC decrease, in slow stress. However, in rapid stress this decrease only occurred in RUBISCO. Both cultivars appear to have a proline reserve that could be easily mobilised during recovery, especially in PB269. The data presented in this work appears to indicate the existence of a wide spectrum of physiological responses to water stress that, having a highly hereditable and stable genetic background, can establish the basis of future plant breeding programs. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho V Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho VI Índice Índice Agradecimentos ………………………………………………………………………………………………….. I Resumo ………………………………………………………………………………………………….……... III Abstract ………………………………………………………………………………………………….…….... V Índice ………………………………………………………………………...………………………………... VII Símbolos e Abreviaturas …………………………………………………………………………………….... IX p 1. Objectivos e Abordagem do Problema ……………………………………………………………………… 1 p 2. Introdução Geral ............................................................................................................................................... 3 2.1. Descrição Botânica do Milho .……………..………………...….……………………………………….…. 3 2.2. O Milho na Economia Mundial e em Portugal ……………....…..……………….…………………….…… 4 2.3. Relações Hídricas em Plantas ……………………………………………………………………………….. 4 2.4. Stresse Hídrico nas Plantas ………………………………………………………………………………….. 6 2.4.1. Aspectos Gerais ……………...…………………………………………………………………………….. 6 2.4.2. Respostas Estomáticas ao Stresse Hídrico ……………………………………………………………….... 8 2.4.3. Metabolismo do Carbono e Efeitos do Stresse Hídrico na Fotossíntese ………………………………...… 9 2.5. Estratégias de Sobrevivência à Seca ……………………………………………………………………….. 14 2.5.1. Aspectos Gerais ……...………………………………………………………………………………….... 14 2.5.2. Modificações Anatómicas, Morfológicas e Metabólicas na Resposta ao Stresse Hídrico …………….…. 15 2.5.3. Solutos Compatíveis e Ajustamento Osmótico …………………………………………………………... 19 2.6. Técnicas de Melhoramento e Selecção para Tolerância ao Stresse Hídrico ……………...……...………… 24 p 3. Metodologia Geral ……………………………………………………………………………………..……. 29 3.1. Material Vegetal ……………………………………………………………………………………………. 29 3.2. Indução do Stresse Hídrico ………………………………………………………………………..……….. 30 3.3. Relações Hídricas ………………………………………………………………………………………...… 30 3.3.1. Determinação do Teor Hídrico Relativo da Folha …………………………………………….………… 30 3.3.2. Determinação do Teor Hídrico do Solo ………………………………………………………………….. 31 3.4. Medição das Trocas Gasosas por Análise de Gases por Radiação Infra-Vermelha (IRGA) …...………….. 31 3.5. Medição da Fluorescência Modulada da Clorofila a …………...……………………...………..…………. 33 3.6. Medição da Libertação de O2 ………………………………………………………………………………. 36 3.7. Medição de Actividades Enzimáticas …………………………………………………………………….... 37 3.8. Análise Estatística ………………………………………………………………………………………….. 37 p 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho …………………………………. 39 4.1. Introdução …………………………………………………………………………………………………... 39 4.2. Metodologia ………………………………………………………………………………………………... 39 4.2.1. Material Vegetal …………………………………………………………………………………………. 39 4.2.2. Relações Hídricas ………………………………………………………………………………………… 39 4.2.3. Medição das Trocas Gasosas …………………………………………………………………………….. 39 4.2.4. Medição da Fluorescência Modulada da Clorofila a …………………………………………………..… 41 4.3. Resultados e Discussão …………………………………………………………………………………..… 42 4.3.1. Relações Hídricas ……………………………………………………………………………………….... 42 4.3.2. Trocas Gasosas ………………………………………………………………………………………...…. 47 4.3.2.1. Stresse Lento ………………………………………………………………………………………….... 48 4.3.2.2. Stresse Rápido ………………………………………………………………………………………….. 59 4.3.3. Fluorescência Modulada da Clorofila a ………………………………………………………………….. 68 4.3.3.1. Stresse Lento ………………………………………………………………………………………….... 68 4.3.3.2. Stresse Rápido ……………………………………………………………………………………….…. 75 4.4. Conclusões ……………………………………...………………………………………………………….. 83 4.4.1. Conclusões das Trocas Gasosas ………...………………………………………………………………... 83 4.4.2. Conclusões da Fluorescência Modulada da Clorofila a ………………………...………………………... 83 4.4.3. Conclusões da Fase I ……………………………………………………………….…………………….. 84 p Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho VII Índice 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico …..... 85 5.1. Introdução ………………………………………………………………………………………………..…. 85 5.2. Metodologia ………………………………………………………………………………………………... 85 5.2.1. Material Vegetal ………………………………………………………………………………………….. 85 5.2.2. Relações Hídricas ……………………………………………………………………………………….... 85 5.2.2.1. Determinação do Teor Hídrico Relativo da Folha ……………………………………….…………….. 85 5.2.2.2. Determinação do Potencial Hídrico das Folhas ………………………………………………………... 86 5.2.2.3. Estabelecimento de Curvas Pressão/Volume …………………………………………………………... 86 5.2.3. Medição das Trocas Gasosas …………………………………………………………………………...… 87 5.2.3.1. Curvas A/PPFD ………………………………………………………………………………………… 87 5.2.3.2. Curvas A/CO2a ………………………………………………………………………………………….. 87 5.2.3.3. Determinação da Libertação de O2 …………………………………………………………...………… 88 5.2.4. Medição da Fluorescência Modulada da Clorofila a …………………………………………………..… 88 5.2.5. Determinação de Clorofilas e Carotenóides ……………………………………………………………… 89 5.2.6. Determinação de Actividades Enzimáticas ………………………………………………………………. 89 5.2.6.1. Extracção ……………………………………………………………………………………………….. 90 5.2.6.2. Ribulose-1,5-Bisfosfato Carboxilase/Oxigenase (RuBisCO) ………………………………………..… 90 5.2.6.3. Fosfoenolpiruvato Carboxilase (PEPC) ……………………………………………………………...… 91 5.2.6.4. Enzima Málica Dependente de NADP (EM-NADP) …...…………………………………………….... 91 5.2.7. Quantificação de Proteína Solúvel …………………………………………………………………….…. 91 5.2.8. Quantificação de Solutos Compatíveis …………………………………………………………………... 92 5.2.8.1. Extracção ……………………………………………………………………………………………….. 92 5.2.8.2. Quantificação de Aminoácidos Livres e Prolina ……………………………………………………….. 92 5.3. Resultados e Discussão …………………………………………………………………………………….. 93 5.3.1. Relações Hídricas ……………………………………………………………………………………….... 93 5.3.2. Trocas Gasosas ………………………………………………………………………………………...…. 96 5.3.2.1. Curvas A/PPFD ………………………………………………………………………………………… 96 5.3.2.2. Curvas A/CO2a ………………………………………………………………………………………. 105 5.3.2.3. Libertação de O2 ………………………………………………………………………………………. 112 5.3.3. Fluorescência Modulada da Clorofila a ……………………………………………………………….... 114 5.3.4. Clorofilas Totais e Carotenóides ………………………………………………………………………... 125 5.3.5. Metabolismo do Carbono ……………………………………………………………………………… 128 5.3.5.1. Ribulose-1,5-Bisfosfato Carboxilase/Oxigenase (RuBisCO) ………………………………………… 129 5.3.5.2. Fosfoenolpiruvato Carboxilase (PEPC) ………………………………………………………………. 133 5.3.5.3. Enzima Málica Dependente de NADP (EM-NADP) ……...………………………………………….. 136 5.3.6. Solutos Compatíveis …………………………………………………………………………………..… 137 5.3.6.1. Aminoácidos Livres ………………………………………………………………………………… 138 5.3.6.2. Prolina ………………………………………………………………………………………………. 140 5.3.7. Conclusões da Fase II ………………………………………………………………………………….... 142 p 6. Conclusões Gerais …………………………………………………………………………………………. 145 p 7. Referências Bibliográficas ……………………………………………………………………………….... 147 p Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho VIII Símbolos e Abreviaturas Símbolos e Abreviaturas A taxa fotossintética A/CO2a taxa fotossintética em função da concentração externa de CO2 A/PPFD taxa fotossintética em função da densidade de fluxo fotónico fotossintético ABA ácido abcísico Apot potencial fotossintético ATP adenosina trisfosfato ATPase ATP sintase C3 metabolismo fotossintético cujo primeiro composto estável formado possui 3 átomos de carbono C4 metabolismo fotossintético cujo primeiro composto estável formado possui 4 átomos de carbono Ca clorofila a 2+ Ca ião cálcio Cb clorofila b Ca+b clorofila total CA1P 2-carboxiarabinitol-1-fosfato Ci concentração interna de CO2 CO2 dióxido de carbono CO2a concentração externa de CO2 Cx+c carotenóides totais DMSO dimetilsulfóxido DTT ditiotritol E taxa transpiratória EDTA ácido etilenodiamina tetracético EM-NADP enzima málica dependente de NADP Eq. equação ETR taxa de transporte de electrões no PSII Fm fluorescência máxima Fm’ fluorescência máxima medida à luz Fo fluorescência basal Fo’ fluorescência basal medida após iluminação Fv fluorescência variável Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho IX Símbolos e Abreviaturas Fig. figura gs condutância estomática H2O água H2O2 peróxido de hidrogénio HCl ácido clorídrico HEPES ácido N-(2-hidroxietil) piperazina N’-2-etanosulfónico IRGA infra-red gas analyser (analisador de gases por radiação infra-vermelha) K+ ião potássio Kcat constante de catálise KCl cloreto de potássio KOH hidróxido de potássio LHCII complexo antena do PSII M molar MDH malato desidrogenase MPa mega Pascal Mg2+ ião magnésio MgCl2 cloreto de magnésio mV milivolt N2 azoto molecular NADH nicotinamida adenina dinucleótido reduzida NADP fosfato de nicotinamida adenina dinucleótido NADP+ fosfato de nicotinamida adenina dinucleótido oxidado NADPH fosfato de nicotinamida adenina dinucleótido reduzido NaHCO3 hidrogenocarbonato de sódio NaH14CO3 hidrogenocarbonato de sódio marcado radioactivamente NPQ amortecimento não-fotoquímico O2 oxigénio molecular p probabilidade P5CS ∆1-pirrolina-5-carboxilato sintetase PAM pulse amplitude modulated (amplitude de pulso modulada) PDH prolina desidrogenase PEG polietilenoglicol PEP fosfoenolpiruvato PEPC fosfoenolpiruvato carboxilase Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho X Símbolos e Abreviaturas Pf peso fresco pH logaritmo do inverso da concentração hidrogeniónica PPFD densidade de fluxo fotónico fotossintético ppm partes por milhão PRK fosforibulocinase Ps peso seco PSII fotossistema II Pt peso túrgido PVP polivinilpirrolidona QA quinona A QB quinona B QAC composto quaternário de amónio qE coeficiente de amortecimento do estado de alta energia qI coeficiente de amortecimento fotoinibitório qN coeficiente de amortecimento não-fotoquímico qP coeficiente de amortecimento fotoquímico qT coeficiente de amortecimento de transição de estado QTL quantitative trait loci (loci de características quantitativas) r 2 coeficiente de determinação rbcS subunidade menor da RUBISCO ROS reactive oxygen species (espécies reactivas de oxigénio) RuBisCO ribulose-1,5-bisfosfato carboxilase/oxigenase RuBP ribulose-1,5-bisfosfato SLA área foliar específica SnCl2.H2O cloreto de estanho hidratado THR teor hídrico relativo THS teor hídrico do solo V volt WUE eficiência do uso da água ΦPSII rendimento quântico efectivo do PSII Ψg potencial gravítico Ψp potencial de pressão Ψs potencial osmótico Ψw potencial hídrico Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho XI Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho XII 1. Objectivos e Abordagem do Problema 1. Objectivos e Abordagem do Problema O milho (Zea mays L.) é originário da América Central, tendo sido rapidamente distribuído pelos quatro cantos do Mundo através das viagens marítimas do século XVI. Ao longo de gerações, a agricultura produziu numerosas variedades locais de milho. Actualmente, estas variedades voltam a adquirir elevada importância, mas num contexto de valorização dos produtos regionais. A diversidade genética das variedades portuguesas de milho constitui um recurso valioso para o melhoramento da espécie e, além disso, a comparação de variedades com diferentes capacidades de resistência ao stresse pode fornecer informações importantes sobre os mecanismos fisiológicos que a conferem. O stresse hídrico é a mais importante limitação à produtividade vegetal. O desenvolvimento de variedades resistentes à seca assume particular importância no panorama de alterações climáticas, em que o aumento da temperatura prolonga o período de restrições hídricas das plantas (Bruce et al., 2002; Taiz e Zeiger, 2002; Chaves et al., 2002, 2003). Para contrariar a influência destas alterações na produtividade, deverá ser desenvolvido um programa de selecção para uma maior tolerância ao stresse hídrico, mantendo uma boa eficiência do uso da água e da capacidade fotossintética, de modo a garantir um aumento de produção nestas condições. Para melhor compreender as limitações fisiológicas e bioquímicas da fotossíntese, torna-se necessário elucidar aspectos fundamentais do metabolismo fotossintético e da gestão da água ao nível celular e da planta inteira e, assim, avançar no conhecimento dos mecanismos de resposta ao stresse. A tolerância é o resultado duma resposta integrada, determinada por múltiplos genes. Deste modo, a identificação de um elevado número de alterações de valor adaptativo revestese de enorme importância para o planeamento de estratégias de melhoramento da resistência à seca de espécies de interesse agrossilvícola. Existem ainda lacunas de conhecimento sobre os mecanismos que afectam a fisiologia e o metabolismo em condições de stresse hídrico, nomeadamente sobre a contribuição relativa dos processos estomáticos e não-estomáticos (ou metabólicos) na redução da fotossíntese (Lawlor e Cornic, 2002). Sob stresse moderado parecem predominar as limitações estomáticas, mas, em certas circunstâncias, à medida que o stresse se intensifica, as limitações não-estomáticas (como a alteração da actividade e estrutura das enzimas do metabolismo fotossintético do carbono) ganham peso (Marques da Silva e Arrabaça, 2004a). A extensão de cada variação, contudo, é desconhecida. Foi sugerido ser possível incluir diferentes espécies Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 1 1. Objectivos e Abordagem do Problema em dois grupos com respostas ao stresse distintas: nas plantas do tipo I, as limitações ao nível do mesófilo aumentam linearmente com a intensidade do stresse, enquanto que, nas plantas do tipo II, não há limitações do mesófilo até um determinado nível de stresse hídrico, mas, uma vez atingido o limiar, observa-se um rápido aumento dessas limitações (Lawlor e Cornic, 2002). Não se conhecem as razões porque uma espécie responde da forma I ou II e se comporta de forma distinta, em diferentes circunstâncias, admitindo-se, no entanto, que a velocidade de imposição do stresse poderá ser relevante. Deste modo, também a forma como cada variedade responde ao stresse é determinante para seleccionar a estratégia de melhoramento. Uma resposta quase universal ao défice hídrico é a acumulação de solutos osmoticamente compatíveis com o metabolismo celular (Bohnert e Shen, 1999). Esta resposta pode conduzir a um ajustamento osmótico e à manutenção de um estado hídrico favorável para o metabolismo e, neste caso, os solutos denominam-se osmorreguladores. Noutros casos, a síntese de certos solutos compatíveis pode ter um papel fundamental na protecção de proteínas (nomeadamente enzimas), na estabilização de membranas celulares e na sequestração de espécies reactivas de oxigénio (ROS, do inglês reactive oxygen species) (Bohnert e Shen, 1999), cuja produção aumenta em condições de défice hídrico (IturbeOrmaetxe et al., 1998). Neste caso, designam-se os solutos de osmoprotectores. Vários estudos revelam que a produção de solutos compatíveis permitiu uma maior actividade de enzimas do ciclo de Calvin em condições de défice hídrico (Shen et al., 1997b; Heuer, 2003). Dada a importância de conhecer a influência do stresse hídrico no desenvolvimento, crescimento e produtividade das plantas agrícolas, tornou-se necessário o desenvolvimento e aplicação de técnicas que forneçam parâmetros com os quais seja possível avaliar os efeitos do stresse ambiental. Algumas dessas metodologias serão aqui abordadas, nomeadamente a medição da fluorescência da clorofila a e das trocas gasosas por análise de gases na banda do infra-vermelho (IRGA). Neste trabalho, proceder-se-á à análise de seis variedades portuguesas de milho através destas técnicas, isolando as duas que mais se distinguem, positiva e negativamente, em termos de tolerância ao stresse hídrico. Ulteriormente, a investigação seguirá novas abordagens dessas duas variedades, de modo a identificar características que permitam explicar algumas diferenças entre a variedade mais tolerante e a variedade mais susceptível ao stresse hídrico, nomeadamente actividades de enzimas-chave do metabolismo do carbono e quantificação de solutos compatíveis, como a prolina. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 2 2. Introdução Geral 2. Introdução Geral 2.1. 2.1. Descrição Botânica do Milho Durante séculos os investigadores interrogaram-se sobre a origem geográfica do milho. Actualmente, todos estão de acordo que a região de origem desta espécie é o Novo Mundo, seja no México, na América Central ou na América do Sul. A espécie de milho que hoje utilizamos é incapaz de sobreviver sem a intervenção do Homem, uma vez que não possui mecanismos de disseminação, nem órgãos de resistência. É uma espécie relativamente recente, com alguns milhares de anos, tendo surgido provavelmente a partir do período em que o Homem iniciou o desenvolvimento da agricultura. O milho pertence à Divisão Magnoliophyta, Classe Liliopsida, Ordem Poales, e Família das Poaceae (também conhecida como Gramineae). É uma espécie muito polimórfica, apresentando populações muito precoces, em que a maturação é atingida em 60-70 dias, e populações muito tardias, que exigem 10 a 11 meses. Também o número de folhas por planta pode variar entre 8 e 48, a altura do caule entre 60 centímetros e 6 metros e a dimensão da espiga entre 2,5 e 30 centímetros (Floyd et al., 2002). Embora existam grandes diferenças morfológicas entre as populações de milho, todas as variedades cultivadas têm características morfológicas comuns, nomeadamente os órgãos vegetativos (raízes, caule, folhas) e os órgãos reprodutores (panícula e espigas). No milho, existem dois sistemas radiculares: um seminal ou provisório e um coronário ou definitivo. As raízes seminais são as primeiras a formar-se permitindo à planta desenvolver-se até formar as raízes coronárias, as quais permitem também fixar a planta ao solo (Floyd et al., 2002). O caule do milho é formado por nós e entrenós, variando o número de nós entre 8 e 48. Ao nível de cada nó estão inseridos uma folha e um gomo axilar. Quando os gomos axilares da base do caule se desenvolvem dão origem a rebentos que, tal como o pé principal, também são formados de caule, folhas e inflorescências (Floyd et al., 2002). Num pé de milho as folhas apresentam dimensões diferentes. As folhas da base são pequenas. O engrossamento do caule provoca a ruptura dos vasos que as alimentam fazendo com que elas deixem de ser funcionais. As folhas inseridas na zona mediana do caule (ao nível da inserção das espigas) são as mais longas e mais largas. No terço superior do caule, as Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 3 2. Introdução Geral folhas vão diminuindo de dimensão à medida que a sua inserção se aproxima da panícula (Floyd et al., 2002). 2.2. O Milho na Economia Mundial Mundial e em Portugal Actualmente, o milho é cultivado em diversos pontos do planeta, desde países como Canadá e Rússia até ao Brasil e países africanos; desde locais situados abaixo do nível do mar junto do Mar Cáspio até aos 3600 metros de altitude nos Andes peruanos; desde zonas semiáridas onde a precipitação anual não ultrapassa 250 milímetros até às regiões onde ela ultrapassa os 1000 milímetros; em regiões com Verão muito curto (Canadá) e nas de Verão permanente (Equador). Nenhuma outra cultura tem uma distribuição geográfica tão vasta (CIMMYT, 2001). Depois do trigo e do arroz, o milho é o cereal mais cultivado, com uma produção de mais de 400 milhões de toneladas por ano, sendo os EUA o maior produtor mundial, com cerca de 40% da produção (CIMMYT, 2001). Em Portugal, a área cultivada é de cerca de 180.000 hectares. Os milhos mais cultivados são os híbridos, calculando-se que ocupem cerca de 71,4 % da área global da cultura. A cultura do milho decorre no período da Primavera-Verão, quando as temperaturas são mais elevadas, sendo a temperatura óptima para o crescimento do milho de 24 a 30ºC (Syngenta, 2004). Este facto levanta sérios problemas ao cultivo de milho, dado que estas temperaturas, associadas ao aumento da evapotranspiração e à diminuição dos recursos hídricos no solo, limitam a água disponível para a planta durante o seu desenvolvimento o que é determinante para garantir uma boa produtividade. 2.3. 2.3. Relações Hídricas em Plantas Plantas A água é o líquido mais abundante à face da Terra, tendo uma função muito importante nas plantas como responsável pela dissolução e transporte de iões e moléculas e influenciando também a estrutura de proteínas, ácidos nucleicos, polissacáridos e outros constituintes da membrana celular. A água forma o ambiente no qual ocorre a maioria das reacções celulares, e participa directamente em muitas reacções bioquímicas (Taiz e Zeiger, 2002). As células vegetais, quando completamente desenvolvidas, possuem um enorme vacúolo constituído de uma solução aquosa de natureza diversa, restando apenas 5 a 10% do Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 4 2. Introdução Geral volume celular para o citoplasma. A água constitui cerca de 80 a 95% da massa dos tecidos vegetais em crescimento. Por outro lado, estruturas como a madeira, constituída essencialmente por células mortas, contém quantidades baixas de água. As sementes possuem 5 a 15% de água, estando entre os tecidos vegetais com menor quantidade de água, embora na germinação necessitem de absorver enormes quantidades (Taiz e Zeiger, 2002). As plantas estão continuamente a absorver e a perder água, grande parte dela perdida por transpiração foliar, através dos estomas, à medida que o CO2, necessário para a fotossíntese, é absorvido da atmosfera. O fluxo de água também é um meio importante de adquirir os minerais existentes no solo através da absorção nas raízes. O movimento de água é conduzido por difusão simples ou fluxo de massa, ou através de uma combinação destes dois mecanismos de transporte, em resposta a uma diferença de potencial hídrico (Steudle e Peterson, 1998; Steudle, 2000). A difusão acontece devido ao constante movimento térmico das moléculas, que ocorre na presença de um gradiente de concentrações. Por outro lado, existe o movimento em fluxo de massa, ao longo do contínuo solo-planta-atmosfera (Wei et al., 1999; Lawlor, 2002). Actualmente, a manutenção deste fluxo é explicada por dois modelos complementares, o modelo da pressão radicular, segundo o qual a água ascende na planta devido a uma pressão provocada pela entrada de água nas raízes, e o modelo de tensão-coesão-adesão, segundo o qual a coluna de água ascende devido às propriedades específicas das moléculas de água, referidas no nome do modelo (Kramer e Boyer, 1995). A água é absorvida pelas raízes e o seu movimento pode ocorrer através de duas vias possíveis: apoplástica (ao longo dos espaços intercelulares) e simplástica (atravessando paredes celulares e membranas plasmáticas e depois difundindo-se pelos plasmodesmos) (Steudle e Peterson, 1998). No caso das células, designa-se por osmose a difusão de água através de membranas, dependendo de um gradiente da energia livre da água através da membrana, normalmente medido como diferença de potencial hídrico (Taiz e Zeiger, 2002). A concentração de solutos no citosol e vacúolo e a pressão hidrostática exercida pela membrana plasmática na parede celular são os dois factores principais que afectam o potencial hídrico, embora, à medida que a distância vertical aumenta, a gravidade também possa desempenhar um papel importante. Os componentes do potencial hídrico (Ψw) podem ser resumidos na expressão Ψw = Ψs + Ψp + Ψg, onde Ψs corresponde ao potencial osmótico (efeito dos solutos na energia livre da água), Ψp corresponde à pressão hidrostática ou de turgescência, e Ψg corresponde ao efeito da gravidade no movimento da água (nas plantas de Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 5 2. Introdução Geral maiores dimensões aumenta cerca de 0,1 MPa / 10 m) (Taiz e Zeiger, 2002). O potencial hídrico é normalmente um bom indicador do estado hídrico da planta. Ao contrário das células animais, as células vegetais podem suportar uma elevada pressão intracelular, denominada pressão de turgescência, resultante da presença da parede celular. Esta pressão é necessária para diversos processos fisiológicos, nomeadamente a elongação celular, as trocas gasosas nas folhas, o transporte floémico, e diversos processos de transporte membranar. A pressão de turgescência também contribui para a manutenção da rigidez e da estabilidade mecânica em tecidos vegetais não-lenhificados. 2.4. 2.4. Stresse Hídrico nas nas Plantas Plantas 2.4. 2.4.1. Aspectos Gerais As plantas estão frequentemente expostas a stresses ambientais que podem resultar em défice hídrico, toxicidade por cálcio e outros metais, deficiência em iões e fotoinibição, entre outras. A reduzida disponibilidade de água é um dos factores ambientais mais importantes na regulação do crescimento e desenvolvimento das plantas e uma das principais causas para a redução do rendimento das colheitas que afectam a maioria das zonas cultivadas por todo o mundo. À medida que os recursos hídricos para utilização agronómica se vão tornando cada vez mais limitantes, o desenvolvimento de linhas de plantas tolerantes à seca tem surgido como um objectivo importante a atingir (Stoop et al., 1996; Hare et al., 1999; Jiang e Zhang, 2002; Bruce et al., 2002). Durante o ciclo de vida de uma planta ocorrem frequentemente períodos de défice hídrico nos solos e/ou na atmosfera, mesmo fora de regiões áridas/semi-áridas, como foi documentado nas florestas decíduas temperadas (Burghardt e Riederer, 2003) ou nas florestas tropicais (Asner et al., 2004). Na natureza, as plantas podem estar sujeitas a dois tipos de stresse hídrico de acordo com a sua duração, podendo ser de longo prazo (de dias a semanas ou meses) ou curto prazo (de horas a dias) (Chaves et al., 2003). As respostas das plantas à escassez de água são complexas, envolvendo alterações adaptativas e/ou efeitos deletérios. Em condições naturais, estas respostas podem ser sinergistica ou antagonisticamente modificadas por sobreposição de outros stresses, como é o caso do stresse salino (Xiong et al., 2002). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 6 2. Introdução Geral As plantas terrestres também podem estar sujeitas à desidratação durante a exposição a temperaturas baixas (Hare et al., 1999), uma vez que a água pode não estar disponível na forma líquida para ser absorvida pelas raízes e, também, devido à diminuição da absorção radicular, nestas condições, mesmo quando está disponível. Algumas das diferenças entre espécies, no que respeita ao crescimento e sobrevivência, podem ser atribuídas a diferentes capacidades para a aquisição de água, transporte e acumulação, e não a diferenças drásticas do metabolismo num determinado estado hídrico. No entanto, a regulação do metabolismo fotossintético também está dependente de factores como as limitações da difusão de CO2 na folha, repartição na alocação de carbono para órgãos não-fotossintéticos e moléculas de defesa, ou mudanças na bioquímica da folha, que podem ser afectados pelo stresse hídrico (Chaves et al., 2002). Em condições de défice hídrico, também ocorre o aumento da produção de radicais livres de oxigénio nas plantas, e a acumulação de solutos compatíveis poderá ter algum efeito protector contra danos oxidativos nas proteínas, interagindo com os grupos que as constituem (Iturbe-Ormaetxe et al., 1998). As respostas iniciais ao stresse hídrico permitem a sobrevivência imediata. No entanto, para que a planta seja capaz de suportar o stresse imposto é necessária uma aclimatação, a qual produz novas capacidades metabólicas e/ou estruturais mediadas pela alteração da expressão génica (Sharp et al., 2004). As respostas fisiológicas ao stresse ambiental têm que ser activadas antes de ocorrerem danos significativos nos tecidos afectados (Hare et al., 1999). Contudo, é a resposta integrada ao nível da planta inteira, incluindo a assimilação de carbono e a alocação de fotoassimilados para diferentes partes da planta e a capacidade reprodutora, que vai determinar a sobrevivência e persistência em condições de stresse ambiental (Chaves e Oliveira, 2004). Para providenciar uma base sólida às estratégias de melhoramento da produção vegetal é necessário uma melhor caracterização dos receptores moleculares envolvidos na percepção do stresse e dos eventos moleculares que especificam a expressão da tolerância ao stresse (Hare et al., 1999). Os grandes avanços na área da genómica permitiram um enorme progresso na caracterização de genes individuais e das suas respostas específicas ao stresse, de modo a determinar a cascata de eventos que estarão associados a vias de sinais inter- e intracelulares do défice hídrico, como é o caso do ácido abcísico (ABA) (Griffiths e Parry, 2002). Contudo, como a maioria dos esforços são dirigidos para a transformação genética das plantas (Casas et Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 7 2. Introdução Geral al., 1993), as abordagens fisiológicas tradicionais são negligenciadas. Até agora, foram desenvolvidas poucas variedades com maior tolerância ao stresse ambiental. Contudo, no mesmo período de tempo, o conhecimento das respostas fisiológicas aumentou substancialmente (Heuer, 2003). 2.4.2 .4.2. .2. Respostas Estomáticas ao Stresse Hídrico Hídrico Os estomas ocupam um papel central nas trocas gasosas, funcionando como o controlo a curto prazo da perda de água por transpiração e da assimilação do CO2 pelas plantas (Jones, 1998). Foram identificadas duas respostas gerais (Tipo I e Tipo II) da fotossíntese ao stresse hídrico, que se distinguem pelo conteúdo relativo de água foliar abaixo do qual surgem os efeitos não-estomáticos (metabólicos) (Lawlor, 2002). Contudo, durante a imposição gradual de défices hídricos moderados (normalmente até –1,8 MPa durante um período de 15 dias ou mais), as respostas baseiam-se quase exclusivamente na limitação da condutividade estomática (gs) (Cornic e Fresneau, 2002; Yordanov et al., 2003). Quando os défices hídricos se desenvolvem, um dos primeiros acontecimentos a tomar lugar nas plantas é o fecho estomático, em resposta a um declínio da turgescência da folha e/ou do seu potencial hídrico (Yordanov et al., 2003), ou ainda em resposta ao aumento da concentração de ABA no fluxo transpiratório (Sharp e LeNoble, 2002; Chaves et al., 2003). Este encerramento permite evitar a desidratação da planta e fenómenos de cavitação, os quais poderiam comprometer a sobrevivência das plantas. A diminuição do CO2 intracelular, após o fecho estomático, e a consequente baixa eficiência do uso da luz pode induzir, a longo prazo, uma regulação da maquinaria fotossintética, no sentido de se adequar ao carbono disponível (Chaves et al., 2002). À medida que a seca progride, nas plantas cultivadas no campo, o fecho estomático ocorre por períodos cada vez mais longos, durante o dia, tendo início a meio da manhã (Tallman, 2004). Esta diminuição da condutância estomática tem como efeito simultâneo a redução da assimilação de carbono e das perdas de água no período de maior défice de pressão de vapor entre a folha e a atmosfera, e leva a uma optimização da assimilação de carbono, em relação à disponibilidade de água (Jones, 1998). Foi encontrada claramente uma dependência do ciclo diurno no que respeita à resposta estomática, tendo em conta a humidade do ar e ao estado hídrico da folha numa dada hora Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 8 2. Introdução Geral (Tallman, 2004), sugerindo que algumas das alterações diurnas na função estomática podem resultar de processos metabólicos com um ritmo circadiano. 2.4. 2.4.3. Metabolismo do Carbono e Efeitos do Stresse Hídrico na Fotossíntese As plantas terrestres apresentam uma enorme diversidade de formas e estratégias adaptativas aos desafios impostos pelo meio ambiente. Também ao nível dos processos fotossintéticos demonstram esta variabilidade. As principais vias fotossintéticas denominamse C3 e C4, devido ao facto de se ter observado que o primeiro produto estável da fixação de CO2 era uma molécula com três e quatro átomos de carbono, respectivamente (Ehleringer e Cerling, 2002). Existe uma terceira via fotossintética denominada CAM (de crassulacean acid metabolism) em que há uma fixação do carbono durante o período nocturno, altura em que os estomas estão abertos, ao contrário do que acontece com as outras duas vias fotossintéticas (Lüttge, 2004). A fotossíntese C3 é um processo com várias etapas nas quais o CO2 atmosférico é fixado em moléculas orgânicas estáveis, ocorrendo praticamente em todas as células do mesófilo das folhas. No primeiro passo, a ribulose-1,5-bisfosfato carboxilase/oxigenase (RuBisCO) cataliza a reacção de carboxilação de uma molécula de ribulose-1,5-bisfosfato (RuBP) com uma molécula de CO2 para formar duas moléculas de fosfoglicerato. Contudo, a RuBisCO também apresenta afinidade para o O2, iniciando uma via metabólica – conhecida por fotorrespiração – levando à perda de carbono e energia (Taiz e Zeiger, 2002; Ehleringer e Cerling, 2002). A fotossíntese C4 representa uma modificação bioquímica da fotossíntese C3, de modo a reduzir a actividade oxigenásica da RuBisCO, aumentando a concentração de CO2 junto desta enzima. Nas plantas C4, a fotossíntese C3 encontra-se restrita às células da bainha dos feixes vasculares (Fig. 2.1). É nas células do mesófilo nas quais se encontra a enzima fosfoenolpiruvato carboxilase (PEPC), a qual fixa o HCO3- (a partir do CO2 numa reacção levada a cabo pela anidrase carbónica) e PEP em oxaloacetato, um ácido C4 que se difunde para as células da bainha do feixe vascular, onde é descarboxilado (Ehleringer e Cerling, 2002). A descarboxilação deste composto e a sua introdução na via C3 normal é realizado de três formas, consoante o tipo de planta C4: através da enzima málica dependente de NADP (EM-NADP) (como é o caso do milho) (Fig. 2.1); através da enzima málica dependente de Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 9 2. Introdução Geral NAD (EM-NAD) (como é o caso de Flaveria trinervia); e através da enzima fosfoenolpiruvato carboxicinase (PEPCK) (como é o caso de Panicum maximum) (Edwards et al., 2001; Walker et al., 2002). No entanto, esta divisão tem sido vista como uma simplificação do que realmente acontece, dado que o milho também possui uma actividade descarboxilativa considerável pela enzima PEPCK (Wingler et al., 1999) e na espécie Panicum maxicum a descarboxilação também é realizada pela enzima EM-NAD (Walker et al., 2002). Independentemente da forma como é descarboxilado o composto C4, a actividade elevada da PEPC conduz a uma elevada concentração de CO2 junto da RuBisCO, limitando a actividade fotorrespiratória (Ehleringer e Cerling, 2002). Fig. 2.1 – Esquema da via fotossintética nas plantas C4, do subtipo EM-NADP, como é o caso do milho (adaptado de Taiz e Zeiger, 2002). As plantas C4, como o milho, têm sido documentadas como melhor adaptadas a condições de défice hídrico devido a uma resistência estomática mais elevada, apresentando, uma melhor eficiência do uso de água e a capacidade de manter a actividade fotossintética em condições de baixo potencial hídrico (Bruce et al., 2002). O milho é capaz de tolerar um nível mais elevado de stresse e manter o crescimento das suas raízes e partes aéreas a níveis de stresse que são inibitórios para plantas C3, como o trigo (Triticum aestivum) (Siddique et al., 1999; Nayyar, 2003). Quando o stresse hídrico é imposto lentamente, como acontece normalmente em condições naturais, pode ocorrer uma redução na capacidade bioquímica de assimilação e Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 10 2. Introdução Geral utilização de carbono, juntamente com restrições na difusão gasosa (Chaves et al., 2002). Diferenças na resposta da fotossíntese à concentração interna de CO2 (Ci) indicam que a necessidade bioquímica de CO2 é regulada em função da diminuição da disponibilidade de CO2, associada ao fecho dos estomas induzido pela seca (Tallman, 2004). Existem dados experimentais que demonstram que a diminuição das taxas de assimilação de CO2 encontradas em folhas sujeitas a stresse hídrico não pode ser revertida simplesmente aumentando o fornecimento externo de CO2, indicando que o stresse hídrico também deve afectar o metabolismo do mesófilo (Lawlor, 2002; Parry et al., 2002; Tang et al., 2002; Cornic e Fresneau, 2002). Esta resposta do mesófilo torna-se progressivamente mais importante com o aumento da deficiência em água (Parry et al., 2002). Numa experiência de campo com videiras, a assimilação de CO2 foi significativamente limitada devido ao fecho dos estomas, à medida que a seca se instalava durante o período de Verão, mas também houve uma redução na actividade de diversas enzimas do ciclo de Calvin (Maroco et al., 2002). Foi demonstrado que os efeitos do défice hídrico na capacidade fotossintética do tremoceiro dependiam da temperatura da folha e da luz incidente (Chaves et al., 1992 in Chaves et al., 2002). Foram obtidos dados semelhantes em diversas solanáceas, mostrando que a fotossíntese era significativamente menos inibida por temperaturas acima dos 38-40ºC em plantas desidratadas comparadas com plantas bem regadas (Havaux, 1992). Numa experiência com duas espécies de Quercus não se encontraram diferenças significativas nas taxas de assimilação de carbono quando estas se comparavam entre plantas em situação de humidade suficiente no solo e plantas sujeitas a um stresse hídrico moderado no início de Julho. Contudo, no fim do Verão quente e seco (Setembro), as trocas gasosas, a meio do dia, na azinheira (Quercus ilex L.) eram menos reduzidas que as observadas no sobreiro (Quercus suber L.) (Faria et al., 1998 in Chaves et al., 2002). Isto sugere um efeito menos severo do stresse hídrico na primeira relativamente ao último. De facto, foram observados valores significativamente mais elevados de potencial hídrico nas folhas no fim do Verão de 1999 na azinheira (-1,52 MPa) quando comparado com os -2,38 MPa medidos no sobreiro. Uma das hipóteses é a de que as raízes da azinheira eram capazes de retirar água de camadas de solo mais profundas, permitindo a esta espécie manter o influxo de água e taxas de assimilação de carbono mais elevadas nas folhas, por um período de tempo mais longo, do que as do sobreiro (Chaves et al., 2002). No final da época seca (Setembro), foi observada uma diminuição da fotossíntese na azinheira e no sobreiro, comparativamente com o início do Verão (Julho). Durante o período Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 11 2. Introdução Geral mais quente do dia, ocorreu uma diminuição da fotossíntese, associada a uma diminuição do rendimento quântico do fotossistema II. Neste período do dia, a assimilação de carbono estava limitada pela diminuição da condutância estomática, podendo aquela ser vista como um importante mecanismo protector contra a foto-oxidação e, portanto, contra a seca nestas árvores. Esta regulação da fotossíntese resulta de uma dissipação térmica do excesso de energia de excitação nos cloroplastos, como foi mostrado pelo aumento do amortecimento não-fotoquímico (NPQ) em Setembro quando comparado com Julho (Chaves et al., 2002). Noutras espécies, sabe-se que a seca rigorosa diminui a quantidade da proteína RuBisCO (Vu et al., 1987). A quantidade de RuBisCO nas folhas é controlada pela sua taxa de síntese e degradação. Mesmo sob stresse hídrico, a holoenzima da RuBisCO é relativamente estável, com um tempo de semi-vida de vários dias (Webber et al., 1994 in Parry et al., 2002). Contudo, o stresse hídrico no tomate (Bartholomew et al., 1991), em Arabidopsis (Williams et al., 1994 in Parry et al., 2002) e no arroz (Vu et al., 1999 in Parry et al., 2002) leva a uma rápida diminuição na abundância de transcritos da subunidade menor da RuBisCO (rbcS), o que pode indicar uma diminuição na sua síntese (Marques da Silva e Arrabaça, 2004b). Em alguns estudos, o potencial fotossintético (Apot), medido em condições saturantes de CO2, não é afectado por uma pequena diminuição do teor hídrico relativo (THR) mas torna-se progressivamente mais inibido, e menos estimulado por elevadas concentrações de CO2, abaixo de um determinado valor de THR (resposta Tipo I). Noutros estudos, Apot e a estimulação da taxa fotossintética por elevadas concentrações de CO2 diminui progressivamente à medida que THR diminui (resposta Tipo II) (Lawlor, 2002). Diversos estudos sugeriram que a diminuição da capacidade fotossintética resulta da insuficiente regeneração de ribulose-1,5-bisfosfato (RuBP) (Giménez et al., 1992). Se isto é provocado ou não pela diminuição da síntese de ATP na fase fotoquímica continua a ser discutido (Gunasekera e Berkowitz, 1993; Tezara et al., 1999; Lawlor, 2002). Embora a actividade da RuBisCO possa não ser a principal limitação do metabolismo cloroplastidial, os efeitos do stresse hídrico na quantidade e actividade da RuBisCO não podem ser ignorados. No estudo de Tezara et al. (1999), a disponibilidade de ATP diminuiu, tendo sido identificada como um factor chave na tolerância ao stresse hídrico. No entanto, as alterações nos níveis de ATP foram inferiores às verificadas para a actividade da RuBisCO, em potenciais hídricos baixos, quando a assimilação de CO2 estava muito reduzida. A actividade da RuBisCO é regulada para ajustar à capacidade da folha em regenerar RuBP, sendo modulada in vivo quer pela reacção com CO2 e Mg2+ para carbamilar o resíduo Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 12 2. Introdução Geral de lisina no local catalítico, quer pela ligação de inibidores no local catalítico (Parry et al., 2002). A ligação dos inibidores à RuBisCO carbamilada previne a catálise, enquanto que a ligação do substrato RuBP na enzima não-carbamilada previne a carbamilação do resíduo de lisina que é essencial para a actividade. A libertação de tais compostos tão fortemente ligados necessita da participação da RuBisCO activase e da hidrólise de ATP (Zhang e Portis, 1999). Enquanto que a medição da actividade da RuBisCO imediatamente após a extracção (actividade inicial) reflecte a actividade in vivo, a carbamilação potencial (actividade total) pode ser determinada incubando os extractos com elevadas concentrações de CO2 e Mg2+ antes do ensaio. Contudo, a carboxilação máxima potencial (actividade máxima) apenas é revelada se forem dados passos para, em primeiro lugar, remover quaisquer inibidores que estejam ligados aos locais activos (Parry et al., 2002). A maioria dos inibidores pode ser removido in vitro através de altas concentrações de sulfato (Parry et al., 1997). No tabaco (Nicotiana tabacum L.), a seca diminuiu a actividade inicial e total da RuBisCO. A diminuição da actividade aparenta ser o resultado de uma diminuição do Kcat aparente e não de uma alteração do estado de activação. Também têm sido observadas diminuições semelhantes na Kcat da RuBisCO em resposta ao stresse hídrico de longa duração no trevo (Medrano et al., 1997 in Parry et al., 2002). Foi demonstrado que a ligação do inibidor CA1P à RuBisCO protege a enzima da degradação proteolítica (Khan et al., 1999). A RuBisCO que se encontra activa na carboxilação da RuBP é relativamente resistente à degradação por proteases; por outro lado, a RuBisCO não-carbamilada com locais activos disponíveis é muito susceptível à proteólise. As interacções com inibidores podem apresentar vantagens in vivo, de modo a prevenir a degradação por proteases da RuBisCO não activa. Como já foi mencionado, a libertação dos inibidores de ligação forte necessita da participação da RuBisCO activase e da hidrólise de ATP (Zhang e Portis, 1999). A remoção dos inibidores pela RuBisCO activase pode ser impedida porque, como já foi sugerido por alguns trabalhos, as concentrações de ATP são reduzidas por acção da seca (Lawlor, 2002). Além disso, a RuBisCO activase é susceptível a temperaturas elevadas (Crafts-Brandner e Salvucci, 2000) que podem estar associadas ao stresse hídrico. No trigo, a diminuição do THR não conduziu ao aumento dos inibidores de ligação forte. Este resultado contradiz estimativas indirectas da percentagem de locais catalíticos bloqueados por inibidores (calculados a partir das actividades máxima e total), o qual sugeria que um número elevado de locais deveriam estar bloqueados por inibidores. Esta discrepância Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 13 2. Introdução Geral pode ser explicada por outros factores, dado que o trigo não contém grandes quantidades de CA1P (Servaites et al., 1986). Existem dois mecanismos a ser considerados em relação à diminuição da actividade da RuBisCO. Primeiro, a RuBisCO reverte a um estado de baixa actividade in vitro através de uma mudança conformacional (Schmidt et al., 1984). No entanto, a hipótese deste mecanismo ter um papel regulador in vivo tem recebido pouca atenção. A segunda hipótese é a inibição da RuBisCO por ligação da RuBP aos locais não-carbamilados (Parry et al., 2002). Em resumo, em condições de campo, quando o stresse hídrico moderado se desenvolve lentamente, a diminuição do CO2 intracelular após o fecho estomático aparentemente induz, a longo prazo, a regulação da maquinaria fotossintética, nomeadamente da RuBisCO, de modo a ajustar ao fluxo de carbono disponível (Chaves et al., 2002). 2.5. 2.5. Estratégias de Sobrevivência à Seca Seca 2.5. 2.5.1. Aspectos Gerais A adaptação das plantas à seca tem sido classificada em três estratégias: escapar, evitar e tolerar (Chaves et al., 2003). No entanto, estas estratégias não são mutuamente exclusivas e, na prática, as plantas combinam uma variedade de tipos de resposta (Ludlow, 1989 in Chaves et al., 2003). As plantas que escapam à seca exibem um elevado grau de plasticidade no seu desenvolvimento, sendo capazes de completar o seu ciclo de vida antes da ocorrência do défice hídrico. Estas estratégias têm como base o sucesso reprodutivo antes do stresse se agravar, sendo de particular importância nas plantas de locais áridos, em que há combinação de ciclos de vida curtos com elevadas taxas de crescimento e de trocas gasosas, maximizando o aproveitamento dos recursos hídricos, florescendo e produzindo sementes antes que as reservas de água se esgotem (Chaves et al., 2002, 2003). Outro mecanismo para um sucesso reprodutivo consiste em mobilizar parte dos recursos para o desenvolvimento de frutos, mecanismo associado à capacidade da planta em armazenar reservas em alguns órgãos (caules e raízes) e mobilizá-los para a produção de frutos, resposta bem documentada em algumas plantas cultivadas, como os cereais (Gebbing e Schnyder, 1999; Bruce et al., 2002) e alguns legumes (Chaves et al., 2002). Esta capacidade aumenta em plantas sujeitas à seca (Rodrigues et al., 1995). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 14 2. Introdução Geral As plantas também podem superar as condições de seca evitando a desidratação dos tecidos, enquanto mantém o potencial hídrico nos tecidos o mais elevado possível, ou tolerando um potencial hídrico baixo no tecido. A desidratação é evitada em plantas anuais ou perenes e está associada com uma variedade de características adaptativas. Estas envolvem diminuição das perdas de água e maximização da tomada de água. As perdas de água podem ser minimizadas pelo fecho dos estomas, por redução da superfície de contacto com a atmosfera através do enrolamento das folhas (Price et al., 2002), através de uma camada densa de tricomas aumentando a reflexão (Werker, 2000), através de ângulos fechados das folhas, ou diminuindo a área da copa através da redução do crescimento e da queda das folhas mais velhas. A tomada de água é maximizada ajustando o padrão de alocação de carbono, nomeadamente aumentando o investimento na formação de raízes (Jackson, 2002). Foram obtidos aumentos significativos na produção de espécies agrícolas, cruzando-as com plantas de regiões semi-áridas, resultando num melhoramento da profundidade de enraizamento (Blum, 1984 in Chaves et al., 2003). A dormência parcial das plantas como sobrevivência à estação seca é outra estratégia encontrada. A tolerância extrema à desidratação encontra-se nas denominadas plantas da ressurreição, bem como em alguns fetos, plantas não-vasculares (musgos), algas e líquenes. As folhas das plantas da ressurreição podem equilibrar com o ar perto de 0-2% (v/v) de humidade relativa e ainda conseguirem recuperar totalmente a sua actividade fisiológica após re-hidratação (Chaves et al., 2003). 2.5. 2.5.2. Modificações Anatómicas, Morfológicas e Metabólicas na Resposta esposta ao Stresse Hídrico Hídrico Para além da diminuição dos recursos hídricos, a seca do solo também induz uma diminuição de nutrientes, em particular o azoto (mas também o cálcio), com fortes efeitos interactivos no crescimento e funcionamento da planta (Chaves et al., 2003). Esta é uma área de investigação de grande importância, necessitando de métodos inovadores que permitem distinguir os efeitos causados pelo défice hídrico dos efeitos causados pela deficiente nutrição mineral, ambos causados pela seca. Os ajustamentos da planta aos baixos recursos dos ambientes áridos incluem alteração da estrutura foliar, aumento da proporção de assimilados mobilizados para as raízes (Poorter e Nagel, 2000), reduzidas taxas de crescimento e baixas taxas de turnover dos órgãos (Enquist e Niklas, 2002). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 15 2. Introdução Geral Enquanto que órgãos com curtos períodos de vida podem ser eliminados em resposta ao stresse, órgãos com longos períodos de vida têm que optimizar o seu ganho em recursos. Este é o caso da mudança de demografia foliar observado na espécie desértica perenial Cryptantha flava, cujo período de vida foliar duplicou em condições de seca (Casper et al., 2001). Esta resposta resulta em taxas de turnover foliar reduzidas e assim elevada eficiência do uso de nutrientes. Isto também significa que as plantas podem compensar parcialmente taxas de ganho de carbono mais reduzidas investindo menos na construção de novas folhas. As folhas com maior longevidade necessitam sobreviver a períodos em que as condições são mais hostis. Para tal, necessitam de diversas protecções que variam desde características anatómicas/morfológicas, como a esclerofilia para resistir a climas extremos e à herbivoria (Lucas et al., 2000; Burghardt e Riederer, 2003), uma densa camada de tricomas como na oliveira (Olea europaea L.) para aumentar a reflectância da luz e manter a humidade (Werker, 2000), ou ângulos foliares fechados (Lizana et al., 2006), até mecanismos bioquímicos que permitem a dissipação do excesso de energia radiante (Young et al., 1997). As folhas esclerófilas resistem a condições climáticas extremas e à herbivoria (Lucas et al., 2000; Burghardt e Riederer, 2003), e são normalmente pequenas e grossas, possuindo uma reduzida área foliar específica (SLA) e baixa concentração de azoto por unidade de massa foliar. Uma diminuição na SLA também pode ocorrer nas plantas herbáceas como resposta à seca, resultando num aumento no investimento de tecidos estruturais, o que permite uma maior resistência a condições ambientais desfavoráveis (Chaves et al., 2003). Outras protecções incluem mecanismos bioquímicos de dissipação de energia, como no caso do ciclo das xantofilas (Young et al., 1997). Folhas pequenas estão bem adaptadas à elevada luminosidade e à elevada temperatura que existem na maioria das regiões áridas, porque o seu tamanho permite uma maior dissipação do calor e um controlo eficaz das perdas de água através do fecho dos estomas (Jarvis e McNaughton, 1986 in Chaves et al., 2003). A senescência, que leva a uma diminuição da reserva de açúcares na planta, é um factor importante na interrupção da formação do fruto, em condições de stresse hídrico. Assim, os fenótipos com senescência retardada podem ser importantes em culturas onde o rendimento seja limitado pelos recursos e a acumulação e utilização das reservas dos caules seja insuficiente para suportar o crescimento do fruto em condições de seca. Contudo, nos cereais, observa-se uma interacção interessante entre a disponibilidade hídrica e a disponibilidade de azoto. Em condições de disponibilidade de água não-limitante, azoto abundante atrasou a senescência mas reduziu o rendimento da produção de grão, enquanto Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 16 2. Introdução Geral que em condições controladas ou suaves de seca no solo, elevada disponibilidade de azoto aumentou a utilização dos hidratos de carbono armazenados e o rendimento da produção de grão (Yang et al., 2000). As hormonas vegetais e açúcares também estão envolvidos na regulação da senescência da parte aérea. Outros estudos revelaram que elevadas concentrações de açúcares solúveis reprimem a expressão de genes associados à fotossíntese, num processo em que as hexocinases estão envolvidas (Smeekens e Rook, 1997). Um efeito importante da sobre-expressão de hexocinases é uma senescência acelerada (Dai et al., 1999). Para além disso, a queda das folhas mais velhas contribui para a diminuição do gasto de água, podendo ser visto como um programa de reciclagem no interior da planta, permitindo a remobilização dos nutrientes armazenados nas folhas mais velhas para os caules ou folhas mais jovens. Durante a indução da senescência pela seca, algumas proteases específicas da seca são activadas, as quais não ocorrem no processo normal de envelhecimento, como é o caso de algumas formas de proteases de cisteína (Solomon et al., 1999). Também existem diferenças entre as espécies tolerantes e as susceptíveis nomeadamente no grau de estimulação da actividade endoproteolítica (protease aspártica), sendo superior nas espécies susceptíveis comparativamente com as tolerantes (Cruz de Carvalho et al., 2001 in Chaves et al., 2003). As folhas mais jovens que sobrevivem à seca apresentam, por vezes, taxas fotossintéticas superiores (Ludlow, 1991) e maior conteúdo de RuBisCO por unidade de área foliar (David et al., 1998 in Chaves et al., 2003) do que folhas de idade semelhante em plantas bem regadas, no que pode ser chamado de suspensão do envelhecimento foliar. As folhas sujeitas a stresse hídrico normalmente atingem a maturidade mais cedo, tornando-se rapidamente numa fonte de carbono, ao contrário do que acontece nas folhas de plantas bem regadas (Schurr et al., 2000). A capacidade da planta captar água de um solo seco através do desenvolvimento de outras características morfológicas, como raízes mais profundas e de uma maior exploração do solo, ou através de um ajustamento osmótico são estratégias capazes de estabilizar e aumentar o fluxo de assimilados encaminhados para as sementes em desenvolvimento (Bruce et al., 2002). Algumas das alterações metabólicas que ocorrem como resultado da seca são, elas próprias, uma consequência da resistência do aparelho fotossintético à desidratação (Cornic, 2000). Enquanto as árvores e os arbustos desenvolveram estratégias de tolerância e de evitar o stresse, as plantas herbáceas e as anuais dependem maioritariamente do crescimento rápido Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 17 2. Introdução Geral para escapar aos “stresses de Verão”, bem como das respostas rápidas da maquinaria fotossintética e do metabolismo do carbono aos sinais iniciais do stresse, incluindo armazenamento de reservas no caule ou nas raízes (Chaves et al., 2002). A actividade da invertase aumentou sob stresse moderado e diminuiu drasticamente com a imposição de um défice hídrico severo. A resposta particular da invertase pode estar relacionada com o papel central desempenhado por esta enzima na modelação das respostas da planta aos stresses biótico e abiótico (Roitsch et al., 2003). O tremoceiro (Lupinus albus L.) sujeito a stresse hídrico mantém a sua produção de sementes devido à sua capacidade de acumular temporariamente assimilados nas partes aéreas que mais tarde são encaminhadas para as vagens, durante o estádio de formação das sementes (Chaves et al., 2002). Essa capacidade do tremoceiro, confirma os dados anteriores de Withers e Forde (1979 in Chaves et al., 2002) demonstrando que o stresse hídrico estimula a capacidade de depósito das sementes e vagens. No caso da azinheira e do sobreiro, a regulação da fotossíntese foi associada com o aumento da actividade do ciclo das xantofilas e com a diminuição simultânea na concentração de clorofila da folha, as quais foram observadas durante o período de Verão. Em videiras sujeitas a precipitação natural após um período prolongado de seca, observou-se uma diminuição do conteúdo em xantofilas e da eficiência dos centros de reacção abertos do PSII, resultantes da diminuição da quantidade de proteínas e clorofila (Maroco et al., 2002). O tremoceiro também é capaz de remover o excesso de energia luminosa através da dissipação térmica, associada com um aumento da concentração de xantofilas – zeaxantina e anteraxantina – à custa de violaxantina, como foi observado a meio do dia, especialmente em plantas sujeitas a stresse hídrico (Chaves et al., 2002). As temperaturas às quais a planta foi sujeita durante o crescimento também influenciam a quantidade total de xantofilas relativamente à de clorofilas presentes, ocorrendo os valores mais elevados em plantas cultivadas num regime de temperaturas mais baixas (15ºC dia/10ºC noite) comparativamente com o que sucede em plantas cultivadas a temperaturas mais elevadas (25ºC dia/20ºC noite). A presença de uma maior quantidade de xantofilas nas plantas no regime de temperaturas mais baixas pode servir como um mecanismo de protecção das folhas, comparativamente com as plantas cultivadas a temperaturas mais elevadas, onde a concentração de clorofila é quase o dobro (Chaves et al., 2002). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 18 2. Introdução Geral Apesar da enorme quantidade de informação acumulada, a compreensão das alterações metabólicas e da forma como as diferentes vias comunicam entre si necessita de um maior aprofundamento. 2.5. 2.5.3. Solutos Compatíveis e Ajustamento Osmótico O ajustamento osmótico faz parte dos mecanismos que evitam a seca mantendo a turgescência, através do aumento e manutenção da quantidade de solutos compatíveis intracelulares no citosol e vacúolo. Tem sido demonstrado que este mecanismo é de particular importância entre todos os mecanismos adaptativos ao stresse hídrico (Bohnert et al., 1995; Bohnert e Shen, 1999; Heuer, 2003; Nayyar, 2003; Marques da Silva e Arrabaça, 2004a). Estudos realizados indicam que ajustamento osmótico mais elevado pode ser um dos factores decisivos na tolerância ao stresse, oferecendo vantagens, no caso do milho. No caso do trigo, planta que possui um ajustamento osmótico mais baixo, o desempenho é inferior em condições de défice hídrico (Nayyar, 2003). A importância da acumulação de solutos compatíveis já tinha sido reconhecida há muito tempo (Jefferies et al., 1979). O tipo de osmólitos e a capacidade de os acumular durante o stresse pode variar entre diferentes tipos de plantas (Nayyar, 2003; Larher et al., 2003) e especificar diferentes mecanismos adaptativos. Muitos dos solutos compatíveis são osmorreguladores, isto é, metabolitos cuja alta concentração celular reduz significativamente o potencial osmótico, e, portanto, são considerados responsáveis pelo ajustamento osmótico. Também podem ter outras funções, nomeadamente na protecção de enzimas e na estrutura das membranas e na eliminação de ROS, designando-se neste caso de osmoprotectores (Bohnert e Shen, 1999). As características dos bons osmoprotectores incluem ser hidrofílicos, não-tóxicos e normalmente sem carga. Estas propriedades são semelhantes aos compostos usados como crioprotectores para a preservação de células procariotas durante o congelamento (Cleland et al., 2004). Existe um largo número de osmoprotectores orgânicos, incluindo o manitol, a trealose, a prolina e a glicina betaína (Stoop et al., 1996; Cleland et al., 2004). A maioria dos organismos aumentam a concentração celular de solutos compatíveis, quando estão em perigo de desidratar, quer por efeito de seca quer pelo aumento da pressão osmótica externa que acompanha, por exemplo, o aumento da salinidade dos solos (Delauney e Verma, 1993). Os compostos que se acumulam são compatíveis com o metabolismo celular Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 19 2. Introdução Geral normal em elevadas concentrações (Brown e Simpson, 1972 in Bohnert e Shen, 1999). A adição de solutos compatíveis aumentou a estabilidade térmica de enzimas, e evitou a dissociação do complexo gerador de oxigénio do PSII (Papageorgiou e Murata, 1995). Um dos aspectos importantes no estudo dos solutos compatíveis é compreender a mecânica da sua acção enquanto integrados numa resposta de toda a planta ao stresse, que inclui a manutenção da homeostase dos iões e das relações hídricas, distribuição carbono/azoto, alocação das reservas, ou armazenamento (e possivelmente difusão) de poder redutor (Bohnert et al., 1995). O potássio, se disponível, pode funcionar como soluto compatível em muitos organismos unicelulares (Bohnert e Shen, 1999). Se existir uma concentração suficiente de potássio no solo, esta pode levar a uma exclusão do sódio mais eficiente nas plantas superiores (Niu et al., 1995). Também aminoácidos e derivados de aminoácidos, açúcares, polióis acíclicos e cíclicos, frutanos, e compostos quaternários de azoto e enxofre agem frequentemente como solutos compatíveis (Delauney e Verma, 1993; Vijn e Smeekens, 1999; Bohnert e Shen, 1999). Foram propostos dois modelos para explicar os efeitos protectores ou estabilizadores dos solutos compatíveis na estrutura e função das proteínas. O primeiro é denominado “modelo de exclusão preferencial” (Arakawa e Timasheff, 1985), segundo o qual os solutos compatíveis são largamente excluídos da película de hidratação das proteínas, que estabiliza a estrutura da proteína ou promove as interacções proteína-proteína. Neste modelo, os solutos compatíveis não disturbariam a água de hidratação das proteínas, mas iriam interagir com a água presente no citosol. O segundo modelo, denominado “modelo da interacção preferencial”, dá mais relevância às interacções entre os solutos compatíveis e as proteínas (Schobert, 1977 in Bohnert e Shen, 1999). A película de hidratação das proteínas é crucial para a sua estabilidade estrutural. Durante o défice hídrico, os solutos compatíveis podem interagir directamente com os domínios hidrofílicos das proteínas e prevenir a sua destabilização, ou podem substituir as moléculas de água na proximidade dessas regiões. Embora os dois modelos possam parecer contraditórios, a verdadeira função dos solutos compatíveis poderá ser explicada por ambos os modelos. As estruturas dos diferentes solutos compatíveis poderiam suportar interacções hidrofóbicas, interacções de van-der-Waals, e interacções de cargas, mas é necessário mais dados para comprovar estas hipóteses (Bohnert e Shen, 1999). Os solutos compatíveis também podem funcionar na remoção de ROS. As plantas e os animais produzem radicais de oxigénio em resposta a uma variedade de stresses, entre os quais os derivados de ataques patogénicos. Dados experimentais demonstraram que agentes Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 20 2. Introdução Geral patogénicos fúngicos são protegidos pela síntese e excreção de manitol, provando-se que este é capaz de remover ROS (Halliwell et al., 1988 in Bohnert e Shen, 1999). Através de experiências que comparavam as capacidades de remoção de radicais por diferentes solutos compatíveis, verificou-se que o manitol, o sorbitol, o glicerol, a prolina, o ononitol e o pinitol eram capazes de o fazer, embora a diferentes concentrações in vitro, enquanto que à glicina betaína não lhe foi atribuída esta função. A eficiência relativa destes compostos para eliminar radicais parece dependente das suas constantes de reacção com radicais hidroxilo (Bohnert e Shen, 1999). Utilizando condições diferentes, como iluminação com luz forte, tratamento com metil viologénio, geração de H2O2, e infiltração de DMSO, foi demonstrado que as plantas contendo manitol nos seus cloroplastos apresentavam uma capacidade superior de manter elevadas taxas de fixação de carbono e um menor branqueamento clorofilino (Shen et al., 1997b). Experiências posteriores demonstraram que o manitol eliminava especificamente os radicais hidroxilo mas não eliminava peróxido de hidrogénio ou radical de oxigénio (Shen et al., 1997a) Demonstrou-se que algumas enzimas do ciclo de Calvin eram predominantemente afectadas por radicais hidroxilo. A fosforibulocinase (PRK), e provavelmente outras enzimasSH do ciclo de Calvin, demonstraram sensibilidade aos radicais hidroxilo, e a actividade da PRK foi protegida pela presença de manitol (Shen et al., 1997a). Assim, a síntese e transporte de solutos compatíveis permitem a protecção contra os efeitos directos (por hidratação) e indirectos (por eliminação de radicais livres) do stresse hídrico (Griffiths e Parry, 2002). As vias que levam à sua síntese estão ligadas a vias do metabolismo geral com elevadas taxas de fluxo. Exemplos disso são a via biossintética do manitol (Stoop et al., 1996), a via biossintética da prolina (Delauney e Verma, 1993), a síntese de glicina betaína (Hanson et al., 1985), e a via que conduz à formação de inositol metilado, D-pinitol (Ishitani et al., 1996). O manitol é um açúcar-álcool com grupos hidroxilo composto por seis carbonos, que se encontra distribuído por todo o reino vegetal. Investigações têm demonstrado que as espécies capazes de o metabolizar têm vantagens na tolerância ao stresse, nomeadamente um aumento da tolerância ao stresse hídrico e salino (funcionando como um soluto compatível) e também no papel que desempenha na resposta aos ataques de agentes patogénicos. Desta forma, o manitol desempenha, aparentemente, um papel importante quer nas respostas ao stresse biótico, quer nas respostas ao stresse abiótico. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 21 2. Introdução Geral Os açúcares osmoprotectores são a forma reduzida de açúcares da classe das aldoses e cetoses. Devido à estrutura dos grupos hidroxilo reproduzir a estrutura da água, pensa-se que estes compostos mantenham uma esfera de hidratação artificial em redor das macromoléculas (Schobert, 1977 in Bohnert e Shen, 1999). Alguns solutos, não respondem ao stresse por acumulação a níveis elevados mas são protectores mesmo a baixas concentrações (Holmström et al., 1996 in Bohnert e Shen, 1999), podendo estar envolvidos em mecanismos não-osmóticos, como por exemplo eliminação de ROS. É exemplo disto a trealose. A prolina e os compostos quaternários de amónio (QCA) (por exemplo, a glicina betaína, colina, prolina betaína) são osmólitos chave que contribuem para o ajustamento osmótico em muitas espécies cultivadas e acumulam-se sob condições de stresse ambiental (Hare et al., 1999; Heuer, 2003; Nayyar, 2003; Larher et al., 2003). A prolina tem sido implicada no alívio da acidose citoplasmática, na manutenção da relação NADP+/NADPH em valores compatíveis com o metabolismo, como antioxidante compatível com proteínas, para além do seu papel importante no desenvolvimento da planta em condições sem stresse (Hare et al., 1999; Raymond e Smirnoff, 2002). Foi demonstrado, no milho, que a prolina desidrogenase (PDH), a primeira enzima na via oxidativa da prolina, é regulada durante o stresse hídrico mas não é afectada por tratamentos com ABA, sugerindo uma regulação mediada pela seca e independente do ABA (Dallmier e Stewart, 1992). Uma rápida indução da actividade da PDH, durante a recuperação do stresse, é consistente com a importância da acumulação da prolina durante o stresse como uma reserva de carbono, azoto e energia rapidamente mobilizável (Hare et al., 1999). Foi sugerido que as alterações induzidas pelo stresse na síntese e degradação da prolina podem afectar a expressão dos genes da planta, para além da visão mais clássica do seu papel no ajustamento osmótico e nos efeitos metabólicos da sua acumulação (Hare et al., 1999; Raymond e Smirnoff, 2002). Estes dados sugerem que a PDH pode ser incluída nos alvos a seleccionar na resistência à seca nas culturas pretendidas (Bruce et al., 2002). Verificou-se que a aplicação exógena de prolina estimula o crescimento das células e das plantas e melhora o metabolismo, em condições de stresse (Heuer, 2003). Contudo, também foram descritos efeitos negativos (Manetas et al., 1986; Rodriguez e Heyser, 1988, Lin e Kao, 2001). A aplicação exógena de prolina (100 mM) diminui a síntese de prolina em células de milho em suspensão (Xin e Li, 1993). Tem sido demonstrada uma relação entre a síntese de prolina e a tolerância à seca e ao stresse salino em plantas de tabaco que expressam Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 22 2. Introdução Geral a enzima da via sintética da prolina, ∆1-pirrolina-5-carboxilato sintetase (P5CS) (Kishor et al., 1995). A glicina betaína é um osmoprotector muito comum que é utilizado por muitas bactérias, bem como algumas Archaea halofíticas (Galinski e Trüper, 1994). Também é um osmoprotector comum nos animais, nas plantas e outros eucariotas (Rhodes e Hanson, 1993 in Cleland et al., 2004). A glicina betaína é um crioprotector simples e útil que funciona para uma vasta gama de organismos procariotas sob diferentes regimes de criopreservação (Cleland et al., 2004). Os QCA, como a glicina betaína, em baixas concentrações, podem melhorar a tolerância ao stresse salino e ao frio, possivelmente por protecção dos complexos proteicos fotossintéticos (Holmström et al., 2000), reduzindo a peroxidação lipídica das membranas, estabilizando as enzimas e as membranas durante as condições de stresse (Bohnert e Shen, 1999; Sakamoto e Murata, 2000). A aplicação exógena de glicina betaína aliviou os efeitos dos stresses hídrico, salino e térmico (Harinasut et al., 1996, Agboma et al., 1997, Rajashekar et al., 1999 in Heuer, 2003). Tal como na prolina, também existem dados sobre a resposta negativa à aplicação exógena de glicina betaína (Kishitani et al., 2000 in Heuer, 2003). Estudos realizados com aplicação de cálcio (Ca2+), revelaram que o milho apresentava uma acumulação de prolina significativamente mais elevada enquanto que o trigo mostrou relativamente uma maior acumulação de QAC’s (Nayyar, 2003). As partes aéreas do milho sujeitas a stresse possuíam um maior Ψw, ajustamento osmótico, taxa de elongação e menor perda de electrólitos, do que as de trigo. O Ca2+ elevou a acumulação de osmólitos em ambas as plantas mas o ajustamento osmótico foi menos sensível a ele. Na presença de Ca2+, o trigo mostrou significativamente maior acumulação de osmólitos, maior Ψw, e maior taxa de elongação do que no milho. Os inibidores de Ca2+ reverteram parcialmente os efeitos do cálcio, indicando o seu envolvimento no controlo da acumulação de solutos (Nayyar, 2003). Tem sido atribuído ao Ca2+ o papel de mensageiro secundário e transdutor de sinal de vários estímulos de stresse (Sanders et al., 1999). Também tem sido demonstrado o seu envolvimento na ligação da percepção do stresse com as várias respostas celulares adaptativas (Torrecilla et al., 2000). O Ca2+ tem sido implicado na protecção contra o stresse por estabilização das membranas e reduzindo os danos oxidativos (Larkindale e Knight, 2002). Tem sido sugerido que o aumento no Ca2+ citosólico induzido pelo stresse funciona como gestor da osmorregulação, uma vez que um aumento no Ca2+ citosólico actua como regulador Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 23 2. Introdução Geral da síntese de prolina. A indução do transcrito para a enzima P5CS, em Arabidopsis, foi inibida por bloqueadores de canais de Ca2+ (Knight et al., 1997). Resumindo, a sobre-produção de prolina e QAC’s devido à aplicação de Ca2+ pode aumentar a protecção contra o stresse como já foi defendido em estudos anteriores (Hare et al., 1999; Raymond e Smirnoff, 2002). Num futuro próximo, poderá recorrer-se à transformação genética para direccionar diferentes solutos compatíveis para diferentes locais subcelulares para protecção da ultraestrutura dos organitos e das enzimas, e para diferentes órgãos, levando a um aumento na tolerância da planta ao stresse (Bohnert e Shen, 1999). 2.6. 2.6. Técnic Técnicas de Melhoramento e Selecção para Tolerância ao Stresse Hídrico Hídrico Ao longo dos anos, os produtores de milho têm apostado na criação de híbridos com maior potencial de rendimento na produção de grão, melhor estabilidade deste rendimento e melhor qualidade de grão para os consumidores (Bruce et al., 2002). A compreensão da natureza destas características, especialmente em ambientes predispostos ao stresse, permitirá melhorar o processo de melhoramento das plantas. Cerca de metade do melhoramento do rendimento de produção das últimas décadas foi devida à mecanização da agricultura, melhor gestão dos terrenos agrícolas e aumento das densidades de cultivo. Os melhoramentos genéticos contribuíram para a outra metade do aumento de produção (Duvick, 2001). Para o milho, o período compreendido entre a polinização e o início do desenvolvimento do grão é o mais sensível ao stresse hídrico, quando comparado com os estádios antes da floração e no final do desenvolvimento do grão. Como resultado da selecção para melhores desempenhos em ambientes com recursos hídricos limitados, o germoplasma tropical demonstrou um número final reduzido de espiguetas e uma formação de sedas precoce e vigorosa. Nas características secundárias, incluem-se o aspecto sempre verde, e reduções na esterilidade, biomassa radicular e enrolamento foliar (Bruce et al., 2002). Os processos de selecção baseiam-se no elevado rendimento de produção de grão, janela de reprodução alargada e um baixo nível de senescência foliar em condições de stresse crítico ou stresse intermédio, e rendimento de produção adequado, pequenas bandeiras e folhas verticais em condições ideais de irrigação (Bruce et al., 2002). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 24 2. Introdução Geral O rendimento da produção de grão é uma característica complexa e está dependente de muitos factores incluindo crescimento vigoroso, fornecimento adequado de água e nutrientes, melhor captação e conversão da luz solar em energia química, e superioridade genética (Russel, 1991 in Bruce et al., 2002). Com os híbridos mais recentes obtêm-se maiores rendimentos de produção comparativamente com linhas mais antigas. Os aumentos resultam de um melhor desempenho em densidades de cultivo mais elevadas. Impondo densidades de plantas mais elevadas geram-se sintomas de resposta ao stresse no milho e reduz-se o rendimento de produção por planta (Modarres et al., 1998; Tollenaar e Wu, 1999; Andrade et al., 1999). Contudo, esta redução é compensada pelo aumento do número de plantas por unidade de área, o qual aumenta o rendimento geral de produção. É provável que linhas seleccionadas para um melhor desempenho em elevadas densidades de cultivo apresentem uma maior tolerância a condições de stresse hídrico e viceversa, especialmente se os processos que afectam o desenvolvimento da maçaroca quando a fotossíntese diminui são afectados por ambos os stresses (Bruce et al., 2002). Elevadas densidades de cultivo criam, obviamente, maior competição para os recursos disponíveis, como captação de radiação solar e aquisição de água e nutrientes pelos sistemas radiculares de plantas sujeitas a essa compactação (Scheiner et al., 2000 in Bruce et al., 2002). O aumento da densidade de cultivo leva a uma menor quantidade de água disponível por planta e a um acelerar na ocorrência dos sintomas de stresse hídrico à medida que a seca progride (Tollenaar e Wu, 1999). Geralmente utiliza-se apenas o rendimento de produção como um parâmetro de selecção. Contudo, será necessário uma melhor compreensão dos componentes que para ele contribuem. O rendimento em termos de produção de grão é a principal característica na selecção. A utilização de uma característica secundária permite um aumento de 20% de êxito na selecção para o rendimento de variedades de milho, em condições de baixa concentração de azoto, em comparação com uma selecção baseada apenas no rendimento da produção de grão (Bänziger et al., 1997; Araus et al., 2002). Uma característica secundária apropriada é (1) geneticamente associada com o rendimento da produção de grão sob seca; (2) altamente hereditária; (3) de medição estável e fiável; (4) não-associada a perdas de rendimento sob condições ideais de crescimento (Edmeades et al., 2001 in Bruce et al., 2002). As características secundárias chave, sob seca, são esterilidade reduzida e reduzida senescência foliar, e, com menor importância, epinastia ou enrolamento da folha em Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 25 2. Introdução Geral condições de seca (Edmeades et al., 1999; Chapman e Edmeades, 1999). A inclusão de outras características, como alterações nos sistemas radiculares, importa apenas incluir quando forem testadas no campo e cumprirem estes parâmetros. No milho, demonstrou-se a importância de alguns atributos das raízes que contribuem para rendimentos mais elevados em ambientes propensos a stresse hídrico, tendo sido identificado o QTL correspondente (Tuberosa et al., 2002). O desvio da biomassa radicular das zonas superiores do solo para as regiões mais profundas poderia melhorar as probabilidades de encontrar novas fontes de água, revelando-se desejável desde que ocorresse como resposta à seca dos solos (Bruce et al., 2002). O aumento aparente da tolerância ao stresse, no germoplasma de milho de clima temperado (Tollenaar e Wu, 1999), tem sido atribuído a um número de factores: (1) a ocorrência de calor e seca em estufas sem irrigação disponível; (2) a utilização de elevadas densidades de plantas durante o desenvolvimento de linhas híbridas; (3) testes em larga escala de vastas áreas que incluem a utilização de estufas de Inverno; e (4) a utilização de progenitores com rendimentos altos estáveis para formar a próxima colheita de cruzamentos parentais. A eficiência na identificação de genótipos com tolerância a stresse abiótico melhorada necessita de um controlo cuidadoso do stresse hídrico imposto às variações genéticas dos fenótipos, como por exemplo a fecundidade. Estes progressos devem-se a uma selecção cuidadosa dos locais de teste. Assim, os ambientes teste necessitam ser estabelecidos onde a probabilidade de stresse hídrico seja elevada, onde a duração e intensidade do stresse possam ser modelados por irrigação, e onde haja semelhanças com a área de adaptação do germoplasma analisado (Bruce et al., 2002). Existem três elementos importantes da caracterização da seca, relevantes para que haja uma correcta selecção da tolerância: timing, duração e intensidade do stresse. Os períodos de stresse encontrados nas áreas de produção a nível mundial são transitórios, imprevisíveis e medidos sem precisão, levando a diversas dificuldades no melhoramento para a tolerância à seca (Bruce et al., 2002). Existem três parâmetros de desempenho centrais para o desenvolvimento de híbridos melhor adaptados ao stresse: (1) a produtividade média numa gama variada de ambientes; (2) a estabilidade no rendimento; (3) a performance relativa entre os híbridos experimentais e as variedades comerciais chave cultivadas nos mesmos ensaios (Bruce et al., 2002). No milho, é importante considerar o número e tamanho das maçarocas geradas por planta, uma vez que estes parâmetros são determinados em diferentes estádios do ciclo de Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 26 2. Introdução Geral vida da cultura (Andrade et al., 1999; Edmeades et al., 1999). Desta forma, os factores que afectam o desenvolvimento da maçaroca são alvos de investigação, com o intuito de obter novo germoplasma, com maior capacidade de resposta ao stresse hídrico (Bruce et al., 2002). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 27 Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 28 3. Metodologia Geral 3. Metodologia Geral O trabalho experimental foi dividido em duas fases. Na fase I, mediram-se as trocas gasosas e a emissão de fluorescência modulada da clorofila a de seis variedades portuguesas de milho, sujeitas a stresse rápido e a stresse lento, com o intuito de seleccionar a variedade mais sensível e a mais tolerante ao stresse hídrico. Na fase II, realizaram-se experiências que permitiram aprofundar o conhecimento do comportamento estomático, do metabolismo do carbono, do aparelho fotoquímico e da acumulação de solutos compatíveis, nas duas variedades seleccionadas e nos dois tipos de indução do stresse hídrico. Neste capítulo são apresentados os métodos que foram utilizados ao longo de todo o trabalho experimental e que são comuns às duas fases. 3.1. 3.1. Material Vegetal As sementes de milho (Zea mays L., das variedades AD3R, PB64, PB260, PB269, PB304, PB369) provenientes do Banco Português de Germoplasma Vegetal (BPGV, Braga) foram desinfectadas em lixívia comercial a 10%, durante 5 minutos, e colocadas em água corrente durante 24 horas. Após este período foram colocadas 3 a 4 sementes em caixas de Petri previamente esterilizadas, contendo papel de filtro embebido em água destilada, durante uma semana, às escuras, numa câmara de crescimento climatizada à temperatura de 25ºC/18ºC (dia/noite). As sementes germinadas foram colocadas em vasos (500 mL) com terra, contendo um composto fertilizante na superfície do solo (10 N : 1 P : 1 K) (Neorgan, Shacham Givatada Ltd., Israel), de modo a obter 0,3 Kg de composto.m-2. Os vasos foram colocados numa câmara de crescimento climatizada, com condições controladas de fotoperíodo (16 horas), temperatura (25ºC dia/18ºC noite), de densidade de fluxo fotónico fotossintético (PPFD) (entre 400 e 500 µmol fotões.m-2.s-1) e de humidade relativa (aproximadamente 40%). As plantas foram regadas sempre que necessário. As plantas foram utilizadas nas experiências 45 dias após o início da germinação, tendo as amostragens sido realizadas na porção média da última folha totalmente expandida (correspondente à 7ª folha). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 29 3. Metodologia Geral 3.2. 3.2. Indução do Stresse Hídrico Neste trabalho foram utilizados duas formas de indução do stresse. A indução lenta (denominada neste trabalho de stresse lento) efectuou-se por supressão da rega durante uma semana, após a qual se procedeu à irrigação das plantas, também durante uma semana, para avaliação da recuperação do stresse hídrico. A indução rápida do stresse hídrico (denominada neste trabalho de stresse rápido) foi obtida por excisão das folhas e sua exposição à atmosfera do laboratório. Com o stresse lento procuram-se criar condições semelhantes às que as plantas encontram no ambiente natural ou agrícola, durante o período de Verão, em que os recursos hídricos são mais escassos (Marques da Silva e Arrabaça, 2004a). As experiências de stresse rápido procuraram simular o meio-dia, período do dia em que a diferença de pressão de vapor entre a folha e a atmosfera é máxima (Marques da Silva e Arrabaça, 2004a). 3.3. 3.3. Relações Hídricas 3.3 3.3.1. .1. Determinação Determinação do Teor Hídrico Relativo da Folha O estado hídrico das folhas foi avaliado através do teor hídrico relativo (THR), que permite determinar a quantidade de água num determinado tecido em relação à quantidade máxima que esse tecido pode conter. Este parâmetro foi calculado segundo a equação seguinte (Catsky, 1960): THR (%) = Pf – Ps Pt – Ps × 100 (Eq. 1) em que Pf é o peso do material fresco, Pt é o peso do material túrgido e Ps é o peso do material seco. O peso fresco das amostras nas experiências de stresse lento foi determinado por excisão de uma pequena porção da folha e pesagem imediata numa balança de precisão (BP 210D, Sartorius, Germany). As amostras foram depois colocadas em caixas de Petri com água desmineralizada durante aproximadamente 24 horas, até atingirem o ponto máximo de hidratação. As amostras foram pesadas, tendo-se tido o cuidado de, previamente, retirar o excesso de água por ligeira pressão em papel absorvente. Deste modo, obteve-se o peso túrgido. Por fim, colocaram-se as amostras numa estufa a 80ºC (TV30, W. Memmert, Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 30 3. Metodologia Geral Schwabach, Germany), durante pelo menos 48 horas, obtendo desta forma o peso seco das amostras. Nas experiências de stresse rápido o procedimento foi semelhante, tendo, contudo, sido utilizados os próprios segmentos da folha analisada para determinação do THR. Os fragmentos foram pesados ao longo da experiência para ulteriormente se determinar os correspondentes THR ao longo da experiência. Foi determinada, sempre que necessário, a área das amostras com um medidor de áreas (CI-202 Portable Leaf Area Meter, CID, Inc., Camas, Washington, USA). 3.3.2. Determinação do Teor Hídrico do Solo O teor hídrico do solo (THS) foi calculado como a percentagem de massa de água em relação à massa do solo (Costa, 1985). O solo foi sendo pesado ao longo de vários dias e a quantidade de água medida com um higrómetro comercial (Ubbink, modelo 2125050, Holanda), correlacionando a escala do aparelho com o THS para uma mais prática inferência sobre o estado hídrico do solo ao longo das experiências. Para se obter a percentagem de água no solo, procedeu-se à sua pesagem após um período de 72 horas numa estufa a 104ºC. O THS máximo corresponde à sua capacidade de campo, em que o solo se encontra saturado, após eliminação do excesso de água. 3.4 3.4. Medição das Trocas Gasosas por Análise de Gases por Radiação Infra Infranfra-Vermelha (IRGA) A fotossíntese é um dos processos fisiológicos mais estudado nas plantas, sendo necessário técnicas que permitam quantificar as trocas gasosas que ocorrem nos tecidos vegetais. A importância de tais métodos no estudo das alterações fisiológicas em plantas sujeitas a stresses ambientais é notória, permitindo uma avaliação do estado em que se encontra a planta. O desenvolvimento recente ao nível da electrónica permitiu a criação de sistemas altamente complexos e sofisticados que fazem medições muito precisas. Simultaneamente, desenvolveram-se as técnicas de processamento digital que permitem, com rapidez e precisão, adquirir dados de uma forma fácil. Os critérios para decidir qual é o método mais adequado para uma determinada experiência incluem o gás a ser medido (O2 ou CO2), o meio em que as medidas vão ser Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 31 3. Metodologia Geral realizadas (aquoso ou gasoso), o nível de actividade metabólica da amostra e a resolução do aparelho, a gama de concentrações do gás a que a amostra vai ser sujeita (capacidade do analisador), se as respostas são ou não rápidas (tempo de resposta do aparelho), a necessidade de medições contínuas ou pontuais (sistemas abertos ou fechados), e se a amostra pode ou não ser sujeita a um método de análise destrutivo (Hunt, 2003). Os resultados deveriam ser avaliados utilizando uma combinação das tecnologias disponíveis, combinando a fluorescência com as técnicas de trocas gasosas, preferencialmente em associação com medidas grosseiras de O2 feitas utilizando um espectrómetro de massa (Haupt-Herting e Fock, 2002). O analisador de gases por radiação infra-vermelha (IRGA, do inglês Infra Red Gas Analyser) consiste num sistema que permite realizar medições das trocas gasosas num sistema aberto. Esta técnica segue o princípio de medição de gases por infra-vermelhos nãodispersivos (NDIR, em inglês), o qual permite medições de moléculas heteroatómicas (como o CO2), através da absorção da radiação infra-vermelha em bandas específicas do espectro das moléculas de gás. Desta forma, quanto maior a concentração de CO2, maior a quantidade de radiação infravermelha absorvida pelas moléculas, logo menor a quantidade de radiação que chega ao detector de Luft. A principal banda de absorção do CO2 é nos 4,25 µm com picos secundários nos 2,66 µm, nos 2,77 µm e nos 14,99 µm (Hill e Powell, 1968 in Hunt, 2003). Na calibração deve ser determinado o zero e também a quantidade de CO2 presente na atmosfera. Actualmente, assume-se o valor de 370 ppm de CO2, como o valor atmosférico (Hunt, 2003). É necessário ter em conta a temperatura e a pressão a que se realiza a experiência porque afectam a quantidade molar de CO2 por unidade de volume de ar (mol.m-3). O vapor de água também poderá interferir com a medição, um vez que possui um pico nos 2,7 µm, perto de um dos picos do CO2. Deve ser utilizado perclorato de magnésio para retirar a água, mas não gel de sílica uma vez que este interfere com a quantidade de CO2 presente (Hunt, 2003). Para uma medição correcta, é preciso ter em conta a taxa de fluxo que atravessa a câmara, dado que as taxas de troca gasosa de CO2 dependem dela. Isto pode ser facilmente obtido através da utilização de fluxómetros que permitam a medição e regulação do fluxo. As taxas fotossintéticas são normalmente expressas em termos de µmoles de CO2 por unidade de tempo e por um outro parâmetro da amostra (normalmente biomassa ou área). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 32 3. Metodologia Geral Esta técnica da medição da troca gasosa do CO2 é utilizada em quase todos os estudos fotossintéticos, uma vez que as alterações deste gás são proporcionalmente superiores ao do O2 e assim mais fáceis de medir com precisão. Neste trabalho foram utilizados dois modelos de IRGA (LCPro+ e LCA-2, ADC – The Analytical Development Co., Ltd., Hoddesdon, England) no stresse lento da fase I, tendo sido utilizado apenas o modelo LCPro+, nas restantes experiências. As taxas das trocas gasosas e das condutâncias estomáticas foram calculadas automaticamente no modelo LCPro+, e recorrendo às equações de von Caemmerer e Farquhar (1981) e Long e Hallgren (1985, 1993) para o modelo LCA-2, de acordo com o descrito em Marques da Silva (1999), e a eficiência do uso de água (WUE) foi calculada pela razão entre a taxa fotossintética e a taxa transpiratória. 3.5 3.5. Medição da Fluorescência Modulada da Clorofila Clorofila a A fluorescência da clorofila a tornou-se numa das técnicas mais úteis nas áreas de investigação da Fisiologia Vegetal e Ecofisiologia. Actualmente, muita da investigação em fotossíntese, sobretudo em condições de stresse, inclui dados da fluorescência (Fracheboud et al., 1999; Hymus et al., 1999; Maxwell e Johnson, 2000). A energia luminosa absorvida pelas moléculas de clorofila in vivo pode ter três destinos possíveis: pode ser utilizada para conduzir a fotossíntese (processo fotoquímico); pode ser dissipada sob a forma de calor (processos não-fotoquímicos); ou pode ser reemitido como luz (fosforescência ou fluorescência da clorofila). Existe competição entre estes três processos (Maxwell e Johnson, 2000). Através da medição da fluorescência da clorofila a, podem recolher-se informações acerca da eficiência dos processos fotoquímicos e não-fotoquímicos. Actualmente, muitos sistemas de medição utilizam luz de excitação modulada, permitindo ler apenas a fluorescência resultante da excitação pela luz utilizada, podendo as medições ocorrer na presença de luz ambiente. Estes aparelhos denominam-se por fluorómetros PAM (Pulse Amplitude Modulated). Embora a quantidade total de fluorescência da clorofila a seja muito pequena em condições in vivo, representando apenas 3 ou 4% do total de luz absorvida (Björkman e Demmig-Adams, 1995), a sua medição é muito simples. O pico de emissão de fluorescência na zona do vermelho longínquo possui um comprimento de onda maior que o da luz absorvida. Assim, o rendimento pode ser determinado expondo a folha a uma radiação Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 33 3. Metodologia Geral fotossinteticamente activa de comprimento de onda definido e medindo a quantidade de luz (fluorescência) emitida. Estas medições são apenas relativas, e qualquer análise necessita de uma forma de normalização (Maxwell e Johnson, 2000). Kautsky e os seus colaboradores descobriram que após a transferência de material fotossintético do escuro para a luz, havia um aumento no rendimento da fluorescência da clorofila a durante o período de cerca de 1 segundo. Este aumento tem sido descrito como uma consequência da redução de aceitadores de electrões na via fotossintética, a jusante do PSII, nomeadamente a plastoquinona, em particular a QA. Quando o PSII recebe luz e QA recebe um electrão, não pode aceitar outro até ser passado para o transportador de electrões posterior (QB). Durante este período de tempo, diz-se que o centro de reacção está fechado. Em qualquer altura, a presença de uma proporção de centros de reacção fechados leva a uma redução global da eficiência fotoquímica e ao correspondente aumento no rendimento da fluorescência (Maxwell e Johnson, 2000; Hunt, 2003). Em plantas adaptadas às escuras (todos os centros de reacção abertos) pode medir-se a fluorescência basal (Fo) do sistema fazendo incidir na amostra uma luz de medição de PPFD e frequência suficientemente baixa para não produzir actos fotoquímicos ou processos não fotoquímicos. Se esta amostra for sujeita a um pulso saturante de luz actínica (entre 5000 e 10000 µmol fotões.m-2.s-1) durante 0,8 segundos, os centros de reacção ficam momentaneamente fechados e, uma vez que os processos de dissipação de energia são competitivos, tem-se um rendimento máximo da fluorescência (Fm). A expressão (Fm-Fo)/Fm, normalmente referida como Fv/Fm, representa a eficiência máxima da conversão de energia pelos centros de reacção abertos dos PSII. Este valor é altamente conservado, sendo cerca de 0,8 em plantas não sujeitas a stresse (Maxwell e Johnson, 2000; Hunt, 2003) e indica que é necessária a energia de 9-10 fotões absorvidos para a produção de uma molécula de O2 na fotossíntese (Walker e Osmond, 1986 in Hunt, 2003). Uma redução neste valor indica que o PSII foi danificado devido a factores ambientais ou à aplicação de inibidores (Maxwell e Johnson, 2000). Normalmente, demora alguns minutos até a fluorescência atingir um estado de equilíbrio (F’), quando o sistema é iluminado por luz actínica. Se a amostra for então sujeita a um novo pulso de luz saturante, o rendimento da fluorescência aumenta até um valor máximo (Fm’) inferior ao valor máximo (Fm) obtido durante o período em que a amostra esteve no escuro. A expressão (Fm’-F’)/Fm’ representa o rendimento quântico efectivo do fotossistema II (ΦPSII) (Genty et al., 1989). As medições da fluorescência permitem também a medição da taxa de transporte de electrões (ETR) dada pela expressão ETR = ΦPSII × PPFD × 0,5 × 0,84, Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 34 3. Metodologia Geral em que o factor 0,5 corresponde à fracção de fotões que chega ao PSII (relativamente ao PSI) e o factor 0,84 corresponde à fracção de luz incidente que é absorvida pela folha. Durante o primeiro segundo de iluminação, há um aumento do rendimento da fluorescência da clorofila a. Contudo, a partir daí o nível de fluorescência volta a diminuir até um valor constante. Este fenómeno denomina-se amortecimento da fluorescência e pode ser explicado por dois processos. O primeiro refere-se ao aumento na taxa de transporte de electrões no PSII, devido, sobretudo, à activação induzida pela luz de enzimas envolvidas no metabolismo do carbono e à abertura dos estomas. Este amortecimento é denominado amortecimento fotoquímico, do qual se pode calcular o coeficiente de amortecimento fotoquímico (qP) que é dado pela expressão (Fm’–F’)/(Fm’–Fo’) onde Fo’ corresponde à fluorescência basal mínima obtida quando a luz actínica é desligada. Simultaneamente, existe um aumento na eficiência da conversão de energia em calor. Este processo é denominado amortecimento não-fotoquímico, calculando-se o coeficiente de amortecimento nãofotoquímico (qN) que é dado pela expressão (Fm–Fm’)/(Fm–Fo’) (Müller et al., 2001). Por sua vez, o amortecimento não-fotoquímico pode ser dividido em três componentes (Quick e Stitt, 1989): qE, que corresponde ao amortecimento de alta energia, resultante da energização dos tilacóides devido ao aumento do gradiente de pH transmembranar; qT, que corresponde ao amortecimento de transição do estado energético I para o estado energético II e que resulta da fosforilação das proteínas dos complexos antena do PSII (LHCII); e qI, que corresponde ao amortecimento fotoinibitório. De entre os três, qE é o maior componente de qN (Quick e Stitt, 1989). Numa planta típica, alterações nestes dois processos, qP e qN, estarão completas em cerca de 15-20 minutos e um estado estacionário é atingido, embora o tempo necessário para alcançar este estado possa variar significativamente entre espécies de plantas (Maxwell e Johnson, 2000; Hunt, 2003). Este é um método bastante vantajoso para avaliar o estado de uma planta uma vez que é uma técnica não-invasiva, não havendo necessidade de destruir a amostra durante as medições. Neste trabalho foram utilizados dois fluorómetros PAM, um modelo mais simplificado na fase I (PAM 210, Heinz Walz GmbH, Effeltrich, Germany) e um modelo mais complexo na fase II (PAM 101 Chlorophyll Fluorometer, Heinz Walz GmbH, Effeltrich, Germany). Os detalhes da sua utilização são descritos na metodologia dessas fases. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 35 3. Metodologia Geral 3.6. Medição da Libertação de O2 O eléctrodo de oxigénio de fase gasosa consiste num aparelho com um cátodo de platina e um ânodo de prata ligados por um electrólito contendo KCl. Este conjunto está separado da amostra por uma membrana permeável ao oxigénio. Aplica-se uma voltagem de cerca de 0,7 V de modo a que o O2 seja reduzido a H2O2 (sendo depois reduzido a iões hidroxilo) no cátodo de platina. Os quatro electrões necessários para a ocorrência desta redução são fornecidos pela oxidação do ânodo de prata, formando-se cloreto de prata. O fluxo entre o ânodo e o cátodo é directamente proporcional à concentração de O2 que se forma nos cloroplastos da amostra (Hunt, 2003). A câmara é fechada de modo a que o O2 se acumule na amostra durante a fotossíntese. Esta técnica também pode ser utilizada para estudar a respiração, na qual se observa, neste caso, uma depleção de O2. A inclinação da curva fornece uma medida da taxa de troca gasosa para o O2, a partir da qual se pode calcular a taxa fotossintética através da área da amostra. A calibração do instrumento é realizada acertando o zero através da utilização de N2 e um valor máximo saturando a câmara com O2. Outro factor a ter em atenção é a temperatura a que se conduz a experiência, uma vez que a solubilidade do O2 e a permeabilidade da membrana variam com este factor. Também é importante ter especial cuidado com a membrana do eléctrodo uma vez que é muito sensível e poderá comprometer as medições se estiver em mau estado (Hunt, 2003). Pode-se medir as trocas de O2 num disco foliar que é selado na câmara. O eléctrodo situa-se na parte inferior do disco, enquanto se fornece luz actínica na parte superior. Nas paredes da janela da câmara circula água de modo a controlar a temperatura. Uma vez que o volume da câmara é muito pequeno, fornece-se CO2 à câmara através de uma solução de bicarbonato de potássio. É necessário determinar o volume da câmara, injectando um volume de ar com uma seringa. A alteração do sinal do eléctrodo de O2 pode ser utilizado para calcular o volume da câmara, uma vez que depende na quantidade de O2 na seringa e na pressão do O2 (valores conhecidos) (Hunt, 2003). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 36 3. Metodologia Geral 3.7. Medição de Actividades Enzimáticas Num estudo envolvendo enzimas é necessário que a sua actividade seja preservada. Desta forma é necessário haver uma paragem rápida da actividade metabólica, para evitar que enzimas proteolíticas actuem sobre a enzima cuja actividade se pretende determinar (Marques da Silva e Arrabaça, 2004b). Uma das formas de assegurar que tal acontece é congelar a amostra em azoto líquido. Também o processo de extracção das enzimas é um dos passos cruciais para a avaliação da sua actividade, uma vez que é necessário manter as condições fundamentais de funcionamento, de modo a minimizar perda de actividade. No entanto, a extracção de enzimas solúveis, como é o caso da RuBisCO (EC 4.1.1.39), da PEPC (EC 4.1.1.31) e da EM-NADP (EC 1.1.1.82), envolve sempre a ruptura da parede celular, normalmente através de esmagamento manual em almofariz. No caso da RuBisCO de plantas C4, o problema agravase uma vez que está localizada nas células da bainha do feixe vascular, as quais apresentam uma parede celular mais espessa (Edwards e Walker, 1983 in Marques da Silva, 1999). Assim, é necessário que o meio de extracção possua condições que permitam manter a conformação estrutural das enzimas. Factores como o pH, temperatura, presença de agentes quelantes, como o EDTA, e concentração salina necessitam ser controlados no meio de extracção. Neste trabalho experimental, as enzimas RuBisCO, PEPC e EM-NADP foram isoladas a partir de um extracto comum. A actividade da RuBisCO foi determinada por incorporação de carbono radioactivo (14C), procedendo-se à sua medição num contador de cintilações, permitindo calcular o carbono fixado num determinado período de tempo. Por outro lado, as actividades da PEPC e da EM-NADP foram determinadas por espectrofotometria (ver 5.2.6). 3.8 3.8. Análise Estatística Estatística O tratamento estatístico dos resultados foi realizado recorrendo a análise e comparação de modelos de regressão e análise de variâncias (ANOVA), recorrendo ao software GraphPad Prism 4.0 (GraphPad Software Inc., USA). Para cada par de variáveis, foram efectuadas uma regressão linear e uma regressão quadrática, escolhendo o mais indicado através da realização de um teste F, admitindo sempre um α de 0,05. Nas curvas de resposta da taxa fotossintética líquida à PPFD foi utilizada a equação de Platt et al. (1980). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 37 3. Metodologia Geral As curvas ajustadas foram apresentadas nos gráficos, com a respectiva equação, o coeficiente de determinação e o nível de significância (p). Nos casos de ajuste da função quadrática, é indicado o nível de significância em relação à função linear. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 38 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho 4.1. Introdução Nesta fase procedeu-se à análise de seis variedades portuguesas de milho (AD3R, PB64, PB260, PB269, PB304, PB369) provenientes do Banco Português de Germoplasma Vegetal (BPGV, Braga). De modo a seleccionar a variedade mais tolerante e a mais susceptível ao stresse hídrico, foram efectuadas medições de trocas gasosas e de fluorescência modulada da clorofila a, nos dois sistemas de indução de stresse hídrico propostos. 4.2. Metodologia 4.2.1. Material Vegetal Foram utilizadas as variedades de milho AD3R, PB64, PB260, PB269, PB304 e PB369. As plantas foram obtidas segundo o descrito em 3.1.. 4.2.2. Relações Hídricas O THR e o THS foram determinados segundo o descrito em 3.3.1. e 3.3.2.. 4.2. 4.2.3 2.3. Medição das Trocas Gasosas Através da análise de gases no IRGA foi possível determinar vários parâmetros que permitem caracterizar as trocas gasosas, como a taxa fotossintética (A), taxa transpiratória (E), condutância estomática (gs) e a pressão parcial de CO2 no espaço intercelular (Ci), nas folhas das seis variedades portuguesas de milho. As medições nesta fase foram efectuadas em dois modelos de IRGA (LCPro+ e LCA2), ambos do mesmo fabricante (ADC – The Analytical Development Co., Ltd., Hoddesdon, England). Inicialmente seria apenas utilizado o modelo LCPro+, mas devido a avaria deste Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 39 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho iniciou-se ulteriormente a utilização do modelo LCA-2. Deste modo, nas experiências de stresse lento foram utilizados os dois modelos, enquanto que nas experiências de stresse rápido foi apenas utilizado o modelo LCPro+. Embora o modelo LCPro+ possua registo automático e controlo dos parâmetros integrados, o modo de funcionamento dos dois IRGA é semelhante, à excepção de alguns detalhes: no modelo LCPro+, a fonte luminosa era um LED que se colocava no topo da câmara, enquanto que, no modelo LCA-2, a fonte de luz era a de um projector de slides (Sawyer’s 302 Automatic, Belgium); a humidade e a temperatura no interior da câmara eram controladas no modelo LCPro+, algo que não acontecia com o modelo LCA-2, embora fosse possível a sua monitorização. Os ensaios foram efectuados sob condições atmosféricas de CO2 (actualmente situa-se próximo de 370 ppm). Para determinar a PPFD adequada a utilizar nesta fase, foi efectuado um ensaio preliminar em várias plantas com o modelo LCPro+. A taxa fotossintética máxima, sem fotoinibição visível na fotossíntese aparente, foi obtida com 870 µmol fotões.m-2.s-1 (ver 4.3.2.). Também foi determinado o período mínimo de iluminação das folhas para se atingir a fotossíntese máxima, sendo este de 20 minutos. Contudo, foi utilizado nas experiências um período de 30 minutos de iluminação para garantir a activação das enzimas do metabolismo do carbono. Quando foi utilizado o modelo LCPro+, foi também seleccionada uma humidade de 10 mbar e uma temperatura de 21ºC (correspondia a cerca de 25ºC ao nível da folha). Nas experiências de stresse lento, foram utilizadas folhas não destacadas, procedendose à sua medição de 2 em 2 dias, ao longo de duas semanas. Apenas era destacada uma pequena porção dessa folha para determinação do THR (segundo o descrito em 3.3.1.). Nas experiências de stresse rápido, a primeira medição era efectuada na folha ainda intacta, tendo sido realizadas ao longo de um dia. As medições posteriores foram efectuadas na mesma área da folha mas já destacada da planta (fragmento com cerca de 8 cm de comprimento), de modo a obter tecidos com diferentes níveis hídricos. No final da experiência, o fragmento foi tratado segundo o descrito em 3.3.1.. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 40 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho 4.2.4 4.2.4. 2.4. Medição da Fluorescência Modulada Modulada da Clorofila a As medições da fluorescência modulada da clorofila a foram efectuadas utilizando o fluorómetro PAM 210 (Heinz Walz GmbH, Effeltrich, Germany) ligado a um computador possuindo o software de registo de dados DA-TEACH v1.11 (Heinz Walz GmbH, Effeltrich, Germany). O método utilizado foi idêntico nas experiências de stresse lento e de stresse rápido, tendo sido utilizada a mesma folha analisada no IRGA. Para medir a variação da área complementar (normalizada) à fase rápida da curva de Kautsky, adaptou-se a folha ao escuro durante 10 minutos. Utilizou-se o Triggered Mode do software mencionado, com as seguintes definições: o pulso de luz saturante com nível 10 (correspondendo à PPFD máxima obtida neste fluorómetro de 3500 µmol fotões.m-2.s-1, considerando-se os pulsos de luz saturante suficientemente fortes para ocorrer o fecho dos centros de reacção), luz de medição com nível 10, e os parâmetros de gain e damp seleccionados para 1. Dado que o fabricante do PAM não indica os valores absolutos das intensidades da luz de medição foi assumido que a luz de medição utilizada é suficientemente fraca para manter os centros de reacção abertos. Para determinar os vários parâmetros de fluorescência, utilizou-se o Saturated Pulse Mode do software mencionado. Novamente, colocou-se a folha na escuridão, durante 5 minutos. Após este período de tempo, foi fornecido um pulso de luz saturante, de modo a determinar o rendimento quântico máximo do PSII (Fv/Fm). Para determinar o rendimento quântico efectivo do PSII (ΦPSII), a taxa de transporte electrónico (ETR) e os coeficientes de amortecimento fotoquímico (qP) e não-fotoquímico (qN) no software foi fornecida à folha uma luz actínica (nível 9, que corresponde a uma PPFD de 840 µmol fotões.m-2.s-1, próxima à utilizada no IRGA), durante 30 minutos, período após o qual se deu um pulso de luz saturante. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 41 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho 4.3. Resultados e Discussão Os ensaios realizados durante a primeira fase do trabalho permitiram a selecção de duas variedades de milho distintas na sua resposta ao stresse hídrico, sendo uma tolerante e a outra susceptível. 4.3.1. Relações Hídricas De modo a monitorizar a quantidade de água disponível no solo, foi utilizado um higrómetro comercial (Ubbink, modelo 2125050, Holanda), ao longo das experiências de stresse lento, tendo sido feita uma calibração da escala do aparelho (estabelecida entre 1 e 10) com o teor hídrico do solo (THS) (Fig. 4.1). THS (%) 30 20 10 1 2 3 4 5 6 7 8 9 10 Escala do Higrómetro y = 13,96 + 0,3642x + 0,1065x2 (r2 = 0,94 ; p < 0,05) Fig. 4.1 – Relação entre o teor hídrico do solo (THS) e a escala do higrómetro comercial, nas experiências de stresse lento. Através da análise do gráfico verifica-se que, quando se procedia à irrigação do solo, o solo apresentava um THS próximo de 30% (valor 10 da escala do higrómetro), diminuindo para cerca de 15% no final do período de stresse (valor 1 da escala do higrómetro). A partir desta calibração e assumindo o THS de 30% como o máximo da capacidade de retenção de água no solo, em que o solo está saturado e o higrómetro comercial apresentava o valor 10, foi possível observar a diminuição da capacidade de campo relativa do solo ao longo das experiências de stresse lento (Fig. 4.2), permitindo deste modo monitorizar os recursos hídricos disponíveis às plantas das diversas variedades. Após uma semana sem irrigação, o solo apresentava uma capacidade de campo relativa de 50%. Contudo, a velocidade com que a mesma era atingida não foi semelhante nas Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 42 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho diferentes variedades devido essencialmente às diferentes taxas transpiratórias. A variedade PB260 foi a que apresentou uma maior taxa de redução da capacidade de campo relativa ao fim de 2 a 4 dias, enquanto a variedade PB64 foi a que apresentou a menor taxa de redução. AD3R PB64 100 Capacidade de campo relativa (%) Capacidade de campo relativa (%) 100 80 60 40 0 2 4 6 8 10 80 60 40 12 0 Tempo (dias) 2 4 PB260 10 12 10 12 10 12 100 Capacidade de campo relativa (%) Capacidade de campo relativa (%) 8 PB269 100 80 60 40 0 2 4 6 8 10 80 60 40 12 0 Tempo (dias) 2 4 6 8 Tempo (dias) PB304 PB369 100 100 Capacidade de campo relativa (%) Capacidade de campo relativa (%) 6 Tempo (dias) 80 60 40 0 2 4 6 8 10 80 60 40 12 0 Tempo (dias) 2 4 6 8 Tempo (dias) Fig. 4.2 – Variação da capacidade de campo relativa do solo em função do tempo do ensaio, nas experiências de stresse lento. Consequentemente, as folhas de plantas de diferentes variedades também não se encontravam no mesmo estado hídrico em cada momento. A variedade PB260 foi a que apresentou a maior redução de THR ao fim de 4 e 6 dias (40%), enquanto que nas restantes Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 43 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho variedades uma diminuição significativa só se verificou ao fim de 6 dias, atingindo 60%, embora a variedade PB269 já apresentasse uma ligeira redução no dia 4 (Fig. 4.3). PB64 100 80 80 60 60 THR (%) THR (%) AD3R 100 40 40 20 20 0 0 0 2 4 6 8 10 0 12 2 4 8 10 12 10 12 10 12 PB269 100 100 80 80 60 60 THR (%) THR (%) PB260 40 20 40 20 0 0 2 4 6 8 10 0 12 0 2 Tempo (dias) 4 6 8 Tempo (dias) PB304 PB369 100 100 80 80 60 THR (%) THR (%) 6 Tempo (dias) Tempo (dias) 40 20 60 40 20 0 0 2 4 6 8 10 0 12 0 Tempo (dias) 2 4 6 8 Tempo (dias) Fig. 4.3 – Variação do teor hídrico relativo (THR) das folhas em função do tempo do ensaio, nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Após irrigação, os THR das variedades AD3R, PB64 e PB304 recuperaram para valores próximos dos iniciais. Contudo, ao contrário das outras, a variedade AD3R apresentou valores de 90% para 95%. As variedades PB260 e PB269 não passaram dos 80% e a variedade PB369 não conseguiu recuperar para além dos 70%, demonstrando, as primeiras, uma dificuldade moderada e, a última, uma dificuldade grande em recuperar os valores de Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 44 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho THR. Comparativamente com outros trabalhos, verifica-se que factores que influenciam a imposição do stresse hídrico e a recuperação após irrigação incluem o tipo de solo, dimensões do recipiente que contém a planta e a própria espécie vegetal analisada (Beardsell e Cohen, 1975; Collinson et al., 1997; Abraham et al., 2004), dificultando a comparação entre diferentes estudos de stresse hídrico. PB64 100 90 90 80 80 THR (%) THR (%) AD3R 100 70 60 70 60 50 50 40 40 30 0 100 200 30 300 0 Tempo (minutos) y = 91,18 – 0,1858x (r2 = 0,96 ; p < 0,01) 90 90 80 80 THR (%) THR (%) 100 70 60 70 60 50 50 40 40 30 200 30 300 0 Tempo (minutos) PB304 300 PB369 100 90 90 80 80 THR (%) THR (%) 200 y = 95,55 – 0,1783x (r2 = 0,98 ; p < 0,01) 100 70 60 70 60 50 50 40 40 30 100 100 Tempo (minutos) y = 94,18 – 0,1528x (r2 = 0,98 ; p < 0,01) 0 300 PB269 100 100 200 y = 96,03 – 0,1259x (r2 = 0,99 ; p < 0,01) PB260 0 100 Tempo (minutos) 200 30 300 0 Tempo (minutos) 100 200 300 Tempo (minutos) y = 93,33 – 0,2106x (r2 = 0,97 ; p < 0,01) y = 96,49 – 0,1922x (r2 = 0,98 ; p < 0,01) Fig. 4.4 – Variação do teor hídrico relativo (THR) das folhas em função do tempo do ensaio, nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 45 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Actualmente, existem abordagens experimentais em que se faz variar a duração e intensidade da seca imposta a curto (5 dias) e longo prazo (2-3 semanas), uma vez que parecem ter resultados significativamente diferentes (Griffiths e Parry, 2002; Marques da Silva e Arrabaça, 2004a). Nas experiências de stresse rápido, as diferentes variedades de milho também apresentaram diferentes THR das folhas ao longo do ensaio para os mesmos intervalos de tempo e para valores de humidade relativa semelhantes (Fig. 4.4). No final destas experiências, as folhas da variedade PB64 possuíam um THR médio de 60%, enquanto que as variedades PB260 e PB269 apresentavam um THR de 50% e as restantes apresentavam THR de 40%. No entanto, realizando uma ANOVA do último ponto registado em cada variedade, a diferença, em relação à variedade PB64, apenas foi significativa nas variedades AD3R, PB304 e PB369, indicando uma maior perda de água, nestas variedades, em stresse rápido. O THR das folhas de todas as variedades diminuiu ao longo da imposição do stresse hídrico nas experiências de stresse lento, embora com taxas diferentes. As variedades PB260 e PB269 atingiram valores mais baixos de THR foliar, tendo, no entanto, recuperado de forma significativa após o início da irrigação, mas não atingindo os valores iniciais (Fig. 4.3). Tal facto poderá estar relacionado com danos celulares ou perda de regulação estomática (Giles et al., 1974; Franks e Farquhar, 2001) ou alterações estruturais que conduzam a um aumento do tempo de recuperação dos níveis hídricos iniciais, devido a alterações do módulo elástico da parede celular (Marshall e Dumbroff, 1999). Por outro lado, a variedade PB369 demonstrou uma taxa mais lenta de recuperação do THR das folhas, relativamente às outras variedades. Kim et al. (2004) obtiveram valores de THR semelhantes no milho mas num período de stresse de cerca de 12 dias. Uma vez que o aspecto das folhas no final do ensaio era semelhante ao observado neste trabalho, em que as folhas se encontravam enroladas segundo o sulco central, Ivandic et al. (2000) sugerem que as diferenças poderão estar essencialmente ligadas à diferença de capacidade de campo dos solos utilizados. Outro aspecto muito importante nos estudos de stresse hídrico e recuperação é a velocidade de imposição stresse hídrico e a duração do mesmo, os quais podem implicar diferentes respostas em variedades da mesma espécie (Kim et al., 2004). O solo utilizado revelou ser adequado a experiências de stresse lento, permitindo a obtenção de stresse moderado e intenso, no período de uma semana. Após irrigação, o solo voltou à sua capacidade máxima de retenção de água, permitindo o estudo do comportamento nas variedades portuguesas de milho. Embora muitos investigadores tenham procedido ao Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 46 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho estudo dos efeitos do stresse hídrico sobre as plantas (Raymond e Smirnoff, 2002; Nayyar, 2003; Çakir, 2004; Marques da Silva e Arrabaça, 2004a, 2004b), o estudo da recuperação dos diversos parâmetros fisiológicos, após irrigação do solo, tem sido, em parte, negligenciado, não permitindo uma clarificação do seu efeito nesses parâmetros (Miyashita et al., 2005). Nas experiências de stresse rápido, as variedades PB64, PB260 e PB269 apresentaram as menores taxas de diminuição do THR ao longo do tempo, revelando um maior controlo estomático comparativamente às variedades AD3R, PB304 e PB369. Uma vez que se pretendia simular as condições do período de dia em que as plantas estão sujeitas a uma maior intensidade luminosa, e em que existe um maior défice hídrico (Marques da Silva e Arrabaça, 2004a), as variedades PB64, PB260 e PB269 parecem conseguir manter o seu THR durante um maior período de tempo, neste momento do dia. Devido à variação do THR das folhas ao longo do tempo não ser igual para todas as variedades, optou-se por apresentar os parâmetros analisados nos ensaios de trocas gasosas e nos ensaios de fluorescência modulada da clorofila a em função do THR e não em função do tempo. 4.3.2. Trocas Gasosas A realização de um ensaio preliminar nas variedades PB260 e PB369 permitiu averiguar qual a melhor PPFD a utilizar durante os ensaios (Fig. 4.5), em que a taxa fotossintética fosse máxima e os efeitos da fotoinibição na fotossíntese aparente não fossem visíveis. A (µ mol CO2.m-2.s-1) 15 10 5 0 0 500 1000 1500 2000 PPFD (µ µ mol fotões.m-2.s-1) y = 20,34 * ((1 – exp (– 0,03805x / 20,34)) * exp (– 0,005627x / 20,34) (r2 = 0,98 ; p < 0,01) Fig. 4.5 – Variação da taxa fotossintética líquida (A) em função da densidade de fluxo fotónico fotossintético (PPFD). Os pontos foram obtidos de 4 folhas de diferentes plantas das variedades PB260 e PB369. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 47 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Chegou-se desta forma ao valor 870 µmol.m-2.s-1, quer por estar no intervalo com as características pretendidas, quer por facilidade de programação no IRGA LCPro+ (ver 4.2.3.). Este valor foi utilizado para as restantes variedades de milho, admitindo-se que apresentam uma resposta semelhante a estas duas variedades, uma vez que estiveram sujeitas às mesmas condições de crescimento. Nas variedades utilizadas no ensaio preliminar, pode observar-se que, para PPFD superiores a 1000 µmol.m-2.s-1, a taxa fotossintética líquida apresenta uma tendência para diminuir, tornando-se visível o fenómeno da fotoinibição (Ying et al., 2002). Também as variações, dadas pelos desvios padrão para os diferentes pontos de PPFD, são relativamente pequenas, o que proporciona alguma segurança sobre o comportamento das restantes variedades. 4.3.2.1. 4.3.2.1. Stresse Lento Na Fig. 4.6 está representada a variação da taxa fotossintética líquida (A) em função do THR, nas diferentes variedades de milho, durante as experiências de stresse lento. É importante referir que, neste trabalho, todos os parâmetros medidos em função do THR apresentam pontos que são simultaneamente os últimos do período de stresse hídrico e os primeiros para o início do período de recuperação, ocorrendo sobreposição dos mesmos. Verifica-se que os valores máximos de A nas diferentes variedades variam entre os 15 e os 30 µmol CO2.m-2.s-1. Estes valores estão de acordo com os observados para o milho e outras plantas C4 por outros autores em condições de crescimento idênticas (Maroco et al., 1998; Marques da Silva, 1999). Os valores de THR a que A se anula também são diferentes consoante as variedades, rondando os 60 a 65% nas variedades AD3R, PB64, PB304 e PB369. Distinguem-se as variedades PB260 e PB269, em que A decresce suavemente num gradiente até se anular a THR baixos, de 40 e 50% respectivamente, valores próximos dos observados por Lal e Edwards (1996) para o milho. No entanto, durante a recuperação, embora estas duas variedades recuperem a função fotossintética quase totalmente, não recuperam o THR inicial (caixas a tracejado na Fig. 4.6). As variedades AD3R, PB64, PB304 e PB369 destacam-se por manifestarem um comportamento diferente. Possuem alguma variação de A até THR de 70-80%, momento em que há uma quebra dessa função, delimitando assim duas regiões de A em função do THR atingido (caixas fechadas na Fig. 4.6). Durante a recuperação, manifestam um comportamento idêntico mas no sentido inverso. Contudo, estas quatro variedades recuperam o seu THR para valores iniciais mas, no geral, não atingem a A inicial. A variedade PB369 destaca-se, ainda, por ser a única que apresenta Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 48 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho valores negativos de A, ou seja, em que a respiração é superior à taxa fotossintética, e que, de um modo geral, apresentou maiores dificuldades em recuperar. PB64 AD3R 30 A (µ µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 30 20 10 0 30 40 50 60 70 80 90 20 10 0 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () PB260 80 90 100 80 90 100 PB269 30 A (µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 70 Recuperação () 30 20 10 0 30 60 THR (%) 40 50 60 70 80 90 20 10 0 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆; - -): y = – 5,731 + 0,1902x (r2 = 0,81 ; p < 0,01) Stresse (∆; - -): y = 15,13 – 0,6625x + 0,007447x2 Recuperação () (r2 = 0,91 ; p < 0,01) Recuperação (; –): y = 16,04 – 0,7276x + 0,008393x2 (r2 = 0,86 ; p < 0,05) PB304 PB369 30 A (µ µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 30 20 10 0 30 20 10 0 40 50 60 70 80 90 40 100 50 60 70 80 90 100 THR (%) THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.6 – Variação da taxa fotossintética líquida (A) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Resumindo, as variedades PB260 e PB269 vão ajustando a sua A à medida que o THR Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 49 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho diminui, conseguem ir até THR mais baixos e após irrigação recuperam significativamente a A inicial. No entanto, PB260 apresenta valores inferiores de A na hidratação máxima. Por outro lado, as variedades AD3R, PB64, PB304 e PB369 sofrem um anulamento abrupto da sua A num valor limite de THR, com um comportamento ao qual se pode chamar “tudo-ounada”, e, embora recuperem o seu THR inicial, apresentam valores inferiores aos de A inicial, após irrigação. PB64 AD3R 0.3 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 0.3 0.2 0.1 0.0 30 40 50 60 70 80 90 0.2 0.1 0.0 30 100 40 50 80 90 100 Recuperação () Recuperação () PB269 PB260 0.3 gs (mol.m-2.s-1) 0.3 gs (mol.m-2.s-1) 70 Stresse (∆) Stresse (∆) 0.2 0.1 0.0 30 60 THR (%) THR (%) 40 50 60 70 80 90 0.2 0.1 0.0 30 100 40 50 60 70 80 90 100 THR (%) THR (%) Stresse (∆; - -): y = 0,2576 – 0,009618x + 9,24e-5x2 Stresse (∆; - -): y = – 0,08262 + 0,002449x (r2 = 0,81 ; p < 0,01) (r2 = 0,81 ; p < 0,05) Recuperação (; –): y = – 0,07998 + 0,002487x Recuperação (; –): y = 0,1093 – 0,00447x + 5,204e-5x2 2 (r = 0,76 ; p < 0,01) (r2 = 0,83 ; p < 0,05) PB304 PB369 0.3 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 0.3 0.2 0.1 0.0 30 40 50 60 70 80 90 0.2 0.1 0.0 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.7 – Variação da condutância estomática (gs) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 50 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Na Fig. 4.7 está representada a condutâmcia estomática (gs) em função do THR. Nas folhas perto da hidratação total, as variedades AD3R, PB64, PB260 e PB304 apresentam gs máximas entre os 0,10 e os 0,20 mol.m-2.s-1, valores de acordo com os observados no milho (Maroco et al., 1999; Bruce et al., 2002; Goodger et al., 2005), destacando-se a variedade PB269 com valores mais elevados (entre os 0,20 e os 0,30 mol.m-2.s-1) e a variedade PB369 com valores mais baixos a rondar os 0,10 mol.m-2.s-1. O momento em que se dá o fecho total dos estomas também diverge nas variedades, ocorrendo na PB260 a THR perto dos 40% e, por oposição, na PB64 ocorre logo aos 70%. A variedade PB260 parece ser também a mais versátil, adaptando a sua gs de uma forma mais ou menos linear com a diminuição do THR. Na variedade PB269 também ocorre alguma adaptação com a diminuição do THR, mas nas restantes ocorre o já referido mecanismo de “tudo-ou-nada”: a maior parte das observações numa janela estreita de THR elevados e amostras finais com valores muito baixos (quase nulos) para níveis de THR inferiores. Após irrigação, a gs das variedades AD3R, PB304 e PB369 recuperou para valores próximos dos iniciais. No caso de AD3R é de notar que A não havia recuperado para os valores iniciais embora gs sim. No caso da variedade PB64, embora THR tenha recuperado, os valores de gs são inferiores aos iniciais. Na variedade PB260, existe a recuperação de gs mas THR nunca recupera totalmente. Na variedade PB269, gs não recupera tal como acontece com o THR. No caso da variedade PB369, uma vez que a A apresenta valores negativos para THR em que gs ainda não se encontra perto dos valores nulos, poderá existir um efeito negativo do stresse hídrico sobre o metabolismo fotossintético do carbono, nomeadamente o efeito do stresse oxidativo na maquinaria fotossintética (Shen et al., 1997a; Kingston-Smith e Foyer, 2000; Chaves e Oliveira, 2004). Esta variedade possui um controlo estomático mais apertado, em oposição à variedade PB269. PB369 necessitará de gs mais baixas de modo reduzir as perdas de água, enquanto, na variedade PB269, o controlo possui uma regulação com uma maior janela de acção, apresentando maiores perdas de água mas mantendo A até THR mais baixos. Estes dados são cruciais na medida que fornecem informações importantes sobre diferentes graus de tolerância destas variedades ao stresse hídrico. A taxa transpiratória (E) acompanha a redução de A devido à diminuição da gs perto da hidratação máxima (Fig. 4.8). A variedade PB269 apresenta um valor de 3 mmol H2O.m2 -1 .s , semelhante ao encontrado na espécie herbácea Poa pratensis, a qual apresenta uma resistência moderada ao stresse hídrico (DaCosta et al., 2004), e a variedade PB369 o valor de 1,5 mmol H2O.m-2.s-1, este mais próximo do encontrado por Tang e Boyer (2002) para o milho (1,6 mmol H2O.m-2.s-1). Contudo, a variedade AD3R apresenta uma E praticamente Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 51 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho nula a THR de 70%, enquanto que o mesmo acontece apenas para THR inferiores a 40% na variedade PB260. PB64 AD3R 3 E (mmol H2O.m-2.s-1) E (mmol H2O.m-2.s-1) 3 2 1 0 30 40 50 60 70 80 90 2 1 0 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () 80 90 100 80 90 100 PB269 3 E (mmol H2O.m-2.s-1) 3 E (mmol H2O.m-2.s-1) 70 Recuperação () PB260 2 1 0 30 60 THR (%) 40 50 60 70 80 90 2 1 0 30 100 40 50 60 70 THR (%) THR (%) Stresse (∆; - -): y = 2,707 – 0,0981x + 0,0009918x2 Stresse (∆; - -): y = – 0,5858 + 0,02183x (r2 = 0,92 ; p < 0,01) (r2 = 0,82 ; p < 0,05) Recuperação (; –): y = – 1,209 + 0,03828x (r2 = 0,82 ; p < 0,01) Recuperação (; –): y = – 1,434 – 0,05697x + 0,0007075x2 (r2 = 0,81 ; p < 0,05) PB369 PB304 3 E (mmol H2O.m-2.s-1) E (mmol H2O.m-2.s-1) 3 2 1 0 30 40 50 60 70 80 90 2 1 0 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.8 – Variação da taxa transpiratória (E) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 52 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Após irrigação, todas as variedades recuperaram a sua E para valores próximos dos observados inicialmente, à excepção da variedade PB64, que apenas recuperou parcialmente, e da variedade PB269, o que está de acordo com os valores de gs registados. Nas variedades AD3R e PB64 verifica-se um ligeiro aumento do Ci mas apenas nas amostras finais (THR baixos) (Fig. 4.6). PB64 500 400 400 Ci (ppm) Ci (ppm) AD3R 500 300 200 100 0 30 300 200 100 40 50 60 70 80 90 0 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () 400 400 300 200 100 90 100 80 90 100 80 90 100 300 200 100 40 50 60 70 80 90 0 30 100 40 50 THR (%) 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB304 PB369 500 500 400 400 Ci (ppm) Ci (ppm) 80 PB269 500 Ci (ppm) Ci (ppm) PB260 300 200 100 0 30 70 Recuperação () 500 0 30 60 THR (%) 300 200 100 40 50 60 70 80 90 0 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆) Stresse (∆; - -): y = 522,7 – 3,937x (r2 = 0,76 ; p < 0,01) Recuperação () Recuperação () Fig. 4.9 – Variação da concentração interna de CO2 (Ci) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 53 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho As variedades PB260, PB269 e PB304 não apresentam variações significativas de Ci com a diminuição do THR, embora a dispersão dos pontos aumente um pouco. Destaca-se a variedade PB369 com um aumento de Ci significativo durante o período de stresse. Verificase, no entanto, uma manutenção de Ci para THR até 90%. O facto de ocorrer um aumento de Ci para THR inferiores a 90%, poderá estar relacionado com a modificação física da folha (distorção das células do mesófilo) (Giles et al., 1974; Lal e Edwards, 1996), ou com a existência de barreiras internas como o pH, dificultando a difusão do CO2 nesta variedade de milho (Marshall e Dumbroff; 1999; Marques da Silva, 1999), ou ainda com um aumento da respiração (Lawlor e Cornic, 2002). Nas variedades em que Ci variou, os valores regressaram aos iniciais, após irrigação, retomando as variedades a assimilação fotossintética do carbono. No entanto, a variedade PB369 apresentou valores de Ci bastante elevados durante a recuperação, o que, comparando com a Fig. 4.6, revela a dificuldade que esta variedade apresentou em retomar a sua função fotossintética. Em relação à eficiência do uso da água (WUE), verifica-se uma maior susceptibilidade ao stresse nas variedades AD3R e PB369, nas quais WUE se anula a THR de 50% (Fig. 4.10). Embora AD3R possua a maior WUE das seis variedades, após irrigação recuperou para valores inferiores aos iniciais. Por outro lado, durante a recuperação da variedade PB369, os valores de WUE continuavam praticamente nulos a THR de cerca de 80%, embora para THR superiores tenha recuperado para valores iniciais. A WUE da variedade PB64 não sofreu variações significativas durante o período de stresse, verificando-se, num entanto, um ligeiro aumento após o período de recuperação. De notar também que a variedade PB260, embora não tenha sofrido variação de WUE até THR de 40%, não recuperou a sua WUE inicial, no final do período de recuperação. A variedade PB304 também diminuiu a sua WUE para metade da inicial a um THR de 40%, recuperando parcialmente após irrigação. A WUE da variedade PB269 sofreu uma redução com a diminuição do THR mas após irrigação recuperou para valores ligeiramente superiores aos iniciais (à excepção de dois pontos) mesmo com THR na ordem dos 80-90%. Esta variedade parece ser a que possui um melhor equilíbrio entre a capacidade fotossintética e a disponibilidade de água no solo. A maior ou menor taxa de redução da WUE poderá estar relacionada com a velocidade a que o stresse hídrico se desenvolve (Richardson e McCree, 1985), reflectindo uma das características de tolerância à seca, como a adaptação deste parâmetro à água disponível. Marques da Silva (1999) registou em Setaria sphacelata (planta C4) um THR de cerca de 75% para a anulação de WUE. No entanto, nestas variedades de milho os valores de THR Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 54 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho registados em que tal sucedia eram inferiores. Tal facto poderá estar relacionado com o fenómeno de enrolamento das folhas, que permite uma melhor WUE, uma vez que diminui as perdas de água por transpiração e permite continuar a assimilação de CO2 (Richards et al., 2002). Este fenómeno foi observado em todas as variedades analisadas. As diferenças entre cada variedade poderão estar relacionadas com outros factores, nomeadamente nas diferenças entre os módulos elásticos da parede celular que afectam a difusão do CO2, uma vez que a rigidez da parede permite ou impede a passagem dos gases (Marshall e Dumbroff, 1999). PB64 WUE (µ µ mol CO2 / mmol H2O) WUE (µ mol CO2 / mmol H2O) AD3R 20 10 0 30 40 50 60 70 80 90 20 10 0 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () WUE (µ µ mol CO2 / mmol H2O) WUE (µ mol CO2 / mmol H2O) 10 50 60 70 80 90 40 50 70 Stresse (∆) Stresse (∆) Recuperação () PB304 10 60 70 80 90 100 80 90 100 PB369 WUE (µ mol CO2 / mmol H2O) WUE (µ µmol CO2 / mmol H2O) 60 THR (%) Recuperação () 50 100 10 0 30 100 20 40 90 20 THR (%) 0 30 80 PB269 PB260 40 70 Recuperação () 20 0 30 60 THR (%) 80 90 20 10 0 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.10 – Variação da eficiência do uso da água (WUE) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 55 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Observou-se uma relação linear entre A e gs (Kim e Lieth, 2003) em todas as variedades, à excepção do ocorrido durante o período de stresse na variedade PB269 (Fig. 4.11). PB64 AD3R 30 A (µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 30 20 10 0 0.0 0.1 0.2 0.3 20 10 0 0.0 0.4 0.1 0.2 0.3 0.4 gs (mol.m-2.s-1) gs (mol.m-2.s-1) Stresse (∆; - -): y = – 0,5661 + 141,6x (r2 = 0,91 ; p < 0,01) Stresse (∆; - -): y = 1,295 + 75,44x (r2 = 0,65 ; p < 0,01) Recuperação (; –): y = – 0,6958 + 93,45x (r2 = 0,91 ; p < 0,01) Recuperação (; –): y = – 0,8057 + 139,9x (r2 = 0,66 ; p < 0,01) PB260 PB269 30 A (µ µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 30 20 10 0 0.0 0.1 0.2 0.3 20 10 0 0.0 0.4 0.1 gs (mol.m-2.s-1) 0.4 -1 gs (mol.m .s ) Stresse (∆; - -): y = 1,8 + 58,84x (r = 0,50 ; p < 0,01) Stresse (∆; - -): y = 0,6314 + 132x – 176,7x2 (r2 = 0,90 ; p < 0,05) Recuperação (; –): y = 1,935 + 40,97x (r2 = 0,49 ; p < 0,01) Recuperação (; –): y = – 1,084 + 150,1x (r2 = 0,91 ; p < 0,01) PB369 PB304 30 30 A (µ µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 0.3 -2 2 20 10 0 0.0 0.2 0.1 0.2 0.3 20 10 0 0.0 0.4 gs (mol.m-2.s-1) 0.1 0.2 0.3 0.4 gs (mol.m-2.s-1) Stresse (∆; - -): y = – 0,1462 + 111,2x (r2 = 0,91 ; p < 0,01) Stresse (∆; - -): y = – 0,7673 + 139,4x (r2 = 0,90 ; p < 0,01) Recuperação (; –): y = – 1,244 + 119,8x (r2 = 0,83 ; p < 0,01) Recuperação (; –): y = – 2,22 + 127,4x (r2 = 0,96 ; p < 0,01) Fig. 4.11 – Relação entre a taxa fotossintética líquida (A) e a condutância estomática (gs), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 56 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Verificou-se que estes dados estão de acordo com os da WUE (Fig. 4.10). AD3R possui a maior A para uma gs menor, mas durante o período de recuperação ocorre uma diminuição desta relação, apresentado a recta um declive mais reduzido. Algo semelhante acontece na variedade PB260. Por outro lado, a variedade PB64 possui uma relação superior após a irrigação, apresentando uma A maior para uma gs mais baixa, comparativamente ao período de stresse. Também em PB269 se observa uma A igual aos valores iniciais mas com uma gs menor, o que justifica uma WUE superior durante a recuperação. Nesta variedade, observa-se uma tendência para um estado de equilíbrio de A com o aumento de gs o que revela alguma limitação não-estomática. Nas variedades PB304 e PB369 não houve diferenças significativas entre o período de stresse e o período de recuperação, embora na variedade PB304 não se tenham observado valores de A superiores a 15 µmol CO2.m-2.s-1, durante a recuperação. Estes dados estão, em parte, de acordo com os valores de Ci observados (Fig. 4.9). Se A varia paralelamente a gs, então Ci permanece constante. Tal facto verifica-se em todas as variedades, ocorrendo pouca variação desse parâmetro, à excepção do que acontece em PB369, em que Ci aumenta, mas possivelmente devido a mecanismos que já foram discutidos anteriormente. Observa-se que A se anula quando os estomas não estão completamente fechados, o que poderá indicar algumas limitações a nível do mesófilo. A nível ecofisiológico WUE poderá ser importante para determinar a tolerância das variedades ao stresse hídrico, dado que indica a capacidade de compromisso entre a perda de água e a assimilação de CO2, à medida que ocorre o fecho estomático. Desta forma, as variedades AD3R, PB260 e PB269 são aquelas que conseguem manter algum nível de A com gs muito reduzidas, mesmo com a intensificação do stresse hídrico, enquanto que nas restantes variedades, principalmente na PB304 e PB369, A anula-se quando os estomas ainda estão abertos. Em todas as variedades, Ci encontra-se entre os 130 e os 230 ppm perto da hidratação total, verificando-se uma relativa manutenção deste parâmetro com a variação de A (Fig. 4.12). Quando A se anula, Ci sofre um aumento para perto dos 300-350 ppm, valores semelhantes aos encontrados noutras C4 (Marques da Silva, 1999). Em PB369, observam-se valores superiores de Ci (cerca de 500 ppm) após a anulação da A, que poderão resultar, como já foi referido, da distorção das células do mesófilo e entrada de CO2 através da cutícula, associados a um módulo elástico mais elevado que impede a difusão de CO2 até aos locais de carboxilação (Giles et al., 1974; Lal e Edwards, 1996; Marshall e Dumbroff, 1999), e ainda a um aumento da respiração (Lawlor e Cornic, 2002). Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 57 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho PB64 AD3R 30 A (µ µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 30 20 10 20 10 0 0 100 200 300 400 0 500 0 100 Ci (ppm) Stresse (∆) Stresse (∆) Recuperação () Recuperação () 400 500 400 500 PB269 PB260 30 A (µ µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 300 Ci (ppm) 30 20 10 20 10 0 0 100 200 300 400 0 500 0 100 Ci (ppm) 200 300 Ci (ppm) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB304 PB369 30 30 A (µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 200 20 10 0 0 100 200 300 400 20 10 0 100 500 200 300 400 500 Ci (ppm) Ci (ppm) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.12 – Relação entre a taxa fotossintética líquida (A) e a concentração interna de CO2 (Ci), nas experiências de stresse lento. Os pontos foram obtidos de 4 a 5 folhas de diferentes plantas. Na recuperação, os valores de Ci retomaram os iniciais para todas as variedades, indicando uma retoma da assimilação fotossintética do carbono, embora as variedades AD3R e PB304 apresentassem uma recuperação mais lenta dos valores iniciais. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 58 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Em suma, nos ensaios de stresse lento, observou-se nas seis variedades, com o decréscimo do THR, uma diminuição geral dos parâmetros estudados (A, E, gs, WUE), característica deste tipo de stresse (Turner, 1974; Lawlor e Cornic, 2002), com o decréscimo do THR nas seis variedades. No entanto, Ci permaneceu relativamente constante num determinado intervalo de THR, sofrendo um aumento à medida que se intensificava o stresse hídrico. O facto dos valores de Ci se manterem com a diminuição do THR (Stuhlfauth et al., 1988) e/ou aumentarem à medida que o THR atinge valores mais baixos (Marques da Silva, 1999) poderá dever-se à existência de barreiras internas à difusão de CO2 que dificultem o seu acesso aos locais de carboxilação (Lawlor e Cornic, 2002), continuando, no entanto, a ocorrer alguma difusão cuticular após o fecho estomático (Meyer e Genty, 1998; Tezara et al., 2003), e um aumento da respiração (Lawlor e Cornic, 2002). Uma vez que existem diferenças entre gs e A relativamente ao THR a que se anulam, as condicionantes de A poderão estar relacionadas com factores não-estomáticos, nomeadamente uma alteração do metabolismo. gs e E anulam-se a um THR inferior ao verificado para A, indicando que existem trocas gasosas mas a fixação de CO2 é inferior à taxa respiratória (Lawlor e Cornic, 2002). 4.3.2.2. 4.3.2.2. Stresse Rápido Os valores de A nas experiências de stresse rápido são mais baixos em todas as variedades quando comparados com os valores das experiências de stresse lento (Fig. 4.6). Tal facto poderá estar relacionado com um curto período em que as plantas estiveram sujeitas a temperaturas mais elevadas devido a avaria da câmara de crescimento. Alterando desta forma as condições de crescimento, as plantas procederam a um ligeiro fecho dos estomas (valores inferiores aos do stresse lento observados na Fig. 4.14), e, como tal, partindo de A mais baixas também poderão ter-se anulado a THR mais altos. Com esta reserva, procedeu-se à análise dos dados obtidos. Nas experiências de stresse rápido verificou-se que A aparente se anulava a THR entre 60 e 70% nas diferentes variedades, à excepção de PB64 em que isso sucedeu a THR de cerca de 85% (ver caixas da Fig. 4.13). Observa-se que, neste caso, a fotossíntese diminuiu mais rapidamente que nas experiências de stresse lento (Fig. 4.6), sobretudo devido a um rápido fecho dos estomas nestas condições (Fig. 4.14). Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 59 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho AD3R PB64 20 A (µ µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 20 15 10 5 0 50 60 70 80 90 100 THR (%) -5 15 10 5 0 50 60 70 80 90 100 80 90 100 THR (%) -5 y = 4,684 – 0,305x+ 0,003702x2 (r2 = 0,97 ; p < 0,05) PB260 PB269 20 A (µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 20 15 10 5 0 50 60 70 80 90 100 THR (%) -5 15 10 5 0 50 60 y = – 17,2 + 0,2763x (r2 = 0,84 ; p < 0,01) y = 2,923 – 0,2474x+ 0,003161x2 (r2 = 0,97 ; p < 0,05) PB304 PB369 20 A (µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 20 15 10 5 0 50 -5 70 THR (%) -5 60 70 80 90 100 THR (%) 15 10 5 0 50 -5 y = 12,86 – 0,6169x+ 0,006596x2 (r2 = 0,99 ; p < 0,01) 60 70 80 90 100 THR (%) y = – 15,38 + 0,2578x (r2 = 0,70 ; p < 0,01) Fig. 4.13 – Variação da taxa fotossintética líquida (A) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Relativamente à gs, e também como acontece em relação a A (Fig. 4.13), a mesma anula-se a THR de cerca de 60-70% para todas as variedades, à excepção de PB64, que diminui abruptamente até um THR na ordem dos 85% e depois mantém-se sem se anular (Fig. 4.14). Observa-se um rápido fecho dos estomas com a diminuição do THR, a partir do qual os valores permanecem praticamente nulos, o que evita a perda de água através das folhas, nos períodos de maior défice hídrico (Raschke, 1970). Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 60 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho AD3R PB64 0.2 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 0.2 0.1 0.0 40 50 60 70 80 90 0.1 0.0 40 100 50 60 THR (%) y = 0,1487 – 0,005555x+ 5,4142e-5x2 (r2 = 0,88 ; p < 0,05) PB260 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 100 PB269 50 60 70 80 90 0.1 0.0 40 100 50 60 THR (%) 70 80 90 100 THR (%) y = 0,2519 – 0,009277x + 8,3765e-5x2 (r2 = 0,88 ; p < 0,05) y = 0,165 – 0,005727x + 5,084e-5x2 (r2 = 0,94 ; p < 0,05) PB304 PB369 0.2 0.2 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 90 0.2 0.1 0.1 0.0 40 80 y = 0,975 – 0,02559x+ 0,0001691x2 (r2 = 0,69 ; p < 0,05) 0.2 0.0 40 70 THR (%) 50 60 70 80 90 0.1 0.0 40 100 THR (%) 50 60 70 80 90 100 THR (%) y = 0,1702 – 0,006466x + 6,1748e-5x2 (r2 = 0,99 ; p < 0,05) y = 0,1132 – 0,003909x + 3,5903e-5x2 (r2 = 0,72 ; p < 0,05) Fig. 4.14 – Variação da condutância estomática (gs) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Pode observar-se na Fig. 4.15, a diminuição de E com a diminuição do stresse hídrico, muito semelhante a gs. De salientar as relativamente elevadas E da variedade PB64 mantidas a baixos THR devido ao não anulamento da gs. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 61 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho AD3R PB64 3 E (mmol.m-2.s-1) E (mmol.m-2.s-1) 3 2 1 0 40 50 60 70 80 90 2 1 0 40 100 50 60 THR (%) y = 2,023 – 0,07554x + 0,0007849x2 (r2 = 0,96 ; p < 0,05) 90 100 PB269 3 3 E (mmol.m-2.s-1) E (mmol.m-2.s-1) 80 y = 14,23 – 0,3733x + 0,002504x2 (r2 = 0,70 ; p < 0,05) PB260 2 1 0 40 50 60 70 80 90 2 1 0 40 100 50 60 THR (%) 70 80 90 100 THR (%) y = 2,716 – 0,1089x + 0,001091x2 (r2 = 0,91 ; p < 0,05) y = 2,688 – 0,09357x + 0,0008661x2 (r2 = 0,96 ; p < 0,05) PB304 PB369 3 3 E (mmol m-2 s-1) E (mmol m-2 s-1) 70 THR (%) 2 1 0 40 50 60 70 80 90 2 1 0 40 100 THR (%) 50 60 70 80 90 100 THR (%) y = 2,456 – 0,09103x + 0,0009155x2 (r2 = 0,99 ; p < 0,01) y = 0,7149 – 0,03273x + 0,0004125x2 (r2 = 0,75 ; p < 0,05) Fig. 4.15 – Variação da taxa transpiratória (E) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Na Fig. 4.16 está representada a variação de Ci em função do THR, onde se observa que até valores de THR próximos de 70-80%, Ci pouco varia (valores assinalados com caixas nos gráficos). Este facto poderá estar relacionado com o facto de A e gs covariarem até este valor de THR. Abaixo desse valor, à medida que o THR diminui observa-se um aumento de Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 62 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Ci, relacionado com os dados já referidos anteriormente (ver Fig. 4.9) Apenas na variedade PB64, tal facto não se observa, em parte por não se terem obtido THR mais reduzidos, mas também poderá resultar do facto de gs não se anular e o CO2 respirado ser libertado para o exterior, não ocorrendo acumulação deste. PB64 1000 750 750 Ci (ppm) Ci (ppm) AD3R 1000 500 250 0 40 500 250 50 60 70 80 90 0 40 100 50 60 THR (%) y = 2639 – 55,16x + 0,3079x2 (r2 = 0,82 ; p < 0,05) 750 750 500 250 100 500 250 50 60 70 80 90 0 40 100 50 60 THR (%) 70 80 90 100 THR (%) y = 3553 – 75,86x + 0,4261x2 (r2 = 0,67 ; p < 0,05) y = 3531 – 76,96x + 0,4351x2 (r2 = 0,93 ; p < 0,01) PB304 PB369 1000 1000 750 750 Ci (ppm) Ci (ppm) 90 PB269 1000 Ci (ppm) Ci (ppm) PB260 500 250 0 40 80 y = 762,8 – 5,731x (r2 = 0,74 ; p < 0,01) 1000 0 40 70 THR (%) 500 250 50 60 70 80 90 0 40 100 THR (%) 50 60 70 80 90 100 THR (%) y = 3082 – 67,03x + 0,3751x2 (r2 = 0,84 ; p < 0,05) y = 3305 – 69,44x + 0,9091x2 (r2 = 0,91 ; p < 0,05) Fig. 4.16 – Variação da concentração interna de CO2 (Ci) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 63 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Relativamente à WUE, verificou-se sempre uma anulação da mesma para THR de cerca de 70-75%, à excepção da variedade PB64 em que tal sucedeu a THR de 80% (Fig. 4.17). Esta variedade parece ter alguma sensibilidade ao stresse hídrico por indução rápida. Todas as variedades apresentaram uma WUE de 5-6 µmol CO2 / mmol H2O perto da hidratação total, semelhante às registadas por Kang e Zhang (2004) no milho, à excepção das variedades PB304 e PB369 que possuem uma WUE ligeiramente mais elevada (8 µmol CO2 / mmol H2O). PB64 WUE (µ mol CO2 / mmol H2O) WUE (µ µ mol CO2 / mmol H2O) AD3R 10 5 0 50 60 70 80 90 100 THR (%) -5 -10 -15 10 5 0 50 60 100 80 90 100 80 90 100 PB269 WUE (µ µ mol CO2 / mmol H2O) WUE (µ µ mol CO2 / mmol H2O) 90 -10 PB260 5 0 50 60 70 80 90 100 THR (%) -5 -10 -15 10 5 0 50 60 70 THR (%) -5 -10 -15 PB304 PB369 10 WUE (µ µ mol CO2 / mmol H2O) WUE (µ mol CO2 / mmol H2O) 80 -15 10 5 0 50 -5 70 THR (%) -5 60 70 80 90 100 THR (%) -10 -15 10 5 0 50 -5 60 70 THR (%) -10 -15 Fig. 4.17 – Variação da eficiência do uso da água (WUE) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 64 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Os valores negativos de WUE observados no gráfico resultam de existirem valores negativos de A, provavelmente resultantes de respiração celular a stresse hídrico mais elevado (THR foliar perto de 50%), associados a uma reduzida mas presente transpiração (Fig. 4.15). AD3R PB64 20 A (µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 20 15 10 5 0 0.1 -5 0.2 gs (mol.m-2.s-1) 15 10 5 0 0.1 -5 y = – 2,554 + 179,8x – 595,8x2 (r2 = 0,98 ; p < 0,05) PB260 PB269 A (µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 10 5 0 0.1 0.2 gs (mol.m-2.s-1) 15 10 5 0 0.1 -5 y = – 0,5647 + 149,8x – 512,8x2 (r2 = 0,95 ; p < 0,05) 0.2 gs (mol.m-2.s-1) y = – 1,645 + 180,2x – 659,8x2 (r2 = 0,99 ; p < 0,05) PB304 PB369 20 20 A (µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) gs (mol.m .s ) 20 15 15 10 5 0 0.1 -5 -1 y = – 1,889 + 118,7x (r2 = 0,98 ; p < 0,01) 20 -5 0.2 -2 0.2 gs (mol.m-2.s-1) 15 10 5 0 0.1 -5 y = – 1,383 + 132,3x (r2 = 0,99 ; p < 0,01) 0.2 -2 -1 gs (mol.m .s ) y = – 1,617 + 159x (r2 = 0,96 ; p < 0,01) Fig. 4.18 – Relação entre a taxa fotossintética líquida (A) e a condutância estomática (gs), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Comparando A com gs verifica-se que as variedades AD3R, PB260 e PB269 apresentam uma relação polinomial de segundo grau, enquanto que as variedades PB64, PB304 e PB369 apresentam uma relação linear (Fig. 4.18). As primeiras conseguem manter Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 65 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho níveis de A mais ou menos constantes até uma determinada gs, o que indica uma maior tolerância ao stresse hídrico, enquanto que as últimas diminuem a sua A consoante a gs, perdendo em parte produtividade e demonstrando uma maior sensibilidade ao stresse hídrico. De notar também que, tal como no stresse lento, as funções aplicadas não cruzam a origem, ou seja, quando A se anula, os estomas não estão totalmente fechados, podendo indicar algumas limitações a nível do mesófilo ou não-estomáticas (ver Fig. 4.11) (Lawlor, 2002), embora possa resultar da uma A real tão baixa que é “mascarada” pelo CO2 respirado. AD3R PB64 20 A (µ µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 20 15 10 5 0 250 500 750 1000 Ci (ppm) -5 15 10 5 0 250 PB260 A (µ µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 1000 750 1000 750 1000 20 15 10 5 0 250 500 750 1000 Ci (ppm) -5 15 10 5 0 250 500 Ci (ppm) -5 PB304 PB369 20 20 A (µ mol CO2.m-2.s-1) A (µ µ mol CO2.m-2.s-1) 750 PB269 20 15 10 5 0 250 -5 500 Ci (ppm) -5 500 750 1000 Ci (ppm) 15 10 5 0 250 -5 500 Ci (ppm) Fig. 4.19 – Relação entre a taxa fotossintética líquida (A) e a concentração interna de CO2 (Ci), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 66 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Na Fig. 4.19 verifica-se que, quando A varia, os valores de Ci se mantém constantes num determinado intervalo (cerca de 130-230 ppm), tal como sucedia no stresse lento (Fig. 4.12). No entanto, quando A se anula, os valores de Ci aumentam para valores que chegam a ser o triplo dos verificados em condições de hidratação total ou stresse moderado, provavelmente devido a um aumento da respiração (Lawlor e Cornic, 2002). A análise dos parâmetros das trocas gasosas nos ensaios de stresse rápido permitem observar uma rápida resposta das variedades portuguesas de milho ao período do dia correspondente a um maior défice hídrico e défice de pressão de vapor (Marques da Silva e Arrabaça, 2004b). Uma resposta mais rápida a uma diminuição brusca do THR foliar é fundamental para que as perdas de água sejam minimizadas e as plantas mantenham o seu estado hídrico constante. Para tal é necessário uma optimização da regulação estomática por parte das plantas de modo a que também as limitações não-estomáticas sejam minimizadas com a perda de água (Raschke, 1970). Especificamente, na variedade PB64, gs e E não se anulam e, portanto, pode desidratar mais facilmente a meio do dia. Por outro lado, nas variedades PB260 e PB269, gs diminui num determinado intervalo sem ocorrer diminuição de A e, desta forma, da produtividade. Esta resposta rápida verificou-se em todas as variedades de milho. Os parâmetros analisados (A, E, gs, WUE) anularam-se de uma forma mais rápida do que nos ensaios de stresse lento. No caso do Ci, houve uma manutenção dos valores até um determinado THR seguido de um aumento, tal como aconteceu no stresse lento. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 67 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho 4.3.3. Fluorescência Fluorescência Modulada Modulada da Clorofila a 4.3.3.1 4.3.3.1. .1. Stresse Lento Na Fig. 4.20 está representada a variação da área complementar (normalizada pela fluorescência basal) à fase rápida da curva de Kautsky, em função do THR, que nos permite inferir sobre a dimensão do pool de quinonas (Lazár, 1999). AD3R PB64 1250 (mV*ms)/(F0*1000) (mV*ms)/(F0*1000) 1250 1000 750 500 250 0 30 40 50 60 70 80 90 1000 750 500 250 0 30 100 40 50 Stresse (∆) Stresse (∆) Recuperação () PB260 (mV*ms)/(F0*1000) (mV*ms)/(F0*1000) 90 100 80 90 100 80 90 100 PB269 750 500 250 40 50 60 70 80 90 1000 750 500 250 0 30 100 40 50 THR (%) 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB369 PB304 1250 (mV*ms)/(F0*1000) 1250 (mV*ms)/(F0*1000) 80 1250 1000 1000 750 500 250 0 30 70 Recuperação () 1250 0 30 60 THR (%) THR (%) 40 50 60 70 80 90 1000 750 500 250 0 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.20 – Variação da área complementar (normalizada) à fase rápida da curva de Kautsky em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 68 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Verifica-se que, à medida que o THR diminui, o pool de quinonas também diminui. Pode observar-se que nas variedades AD3R, PB64 e PB304 a variação é maior, enquanto que nas variedades PB260, PB269 e PB369 essa variação é menor. Na variedade PB260, só em stresse intenso (THR entre 30 e 50%) é que a diminuição é visível. Após a irrigação, a variedade AD3R demonstrou maior dificuldade na recuperação do parâmetro, enquanto nas restantes variedades recuperou para valores próximos dos iniciais. PB64 AD3R 0.8 Fv/Fm (unidades relativas) Fv/Fm (unidades relativas) 0.8 0.6 0.4 0.2 30 40 50 60 70 80 90 0.6 0.4 0.2 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () Fv/Fm (unidades relativas) Fv/Fm (unidades relativas) 90 100 80 90 100 80 90 100 0.8 0.6 0.4 40 50 60 70 80 90 0.6 0.4 0.2 30 100 40 50 THR (%) 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB304 PB369 0.8 0.8 Fv/Fm (unidades relativas) Fv/Fm (unidades relativas) 80 PB269 PB260 0.6 0.4 0.2 30 70 Recuperação () 0.8 0.2 30 60 THR (%) 40 50 60 70 80 90 0.6 0.4 0.2 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.21 – Variação do rendimento quântico máximo do PSII (Fv/Fm) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 69 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Relativamente ao Fv/Fm, verifica-se um valor perto de 0,8 unidades relativas perto da hidratação total, descrito para a maioria das espécies vegetais em condições de ausência de stresse (Fracheboud et al., 1999; Nogués e Baker, 2000; Maxwell e Johnson, 2000) (Fig. 4.21). AD3R PB64 0.5 ΦPSII (unidades relativas) ΦPSII (unidades relativas) 0.5 0.4 0.3 0.2 0.1 0.0 30 40 50 60 70 80 90 0.4 0.3 0.2 0.1 0.0 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () PB260 Φ PSII (unidades relativas) Φ PSII (unidades relativas) 90 100 PB269 0.3 0.2 0.1 40 50 60 70 80 90 0.4 0.3 0.2 0.1 0.0 30 100 40 50 THR (%) 60 70 80 90 100 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB369 PB304 0.5 0.5 Φ PSII (unidades relativas) Φ PSII (unidades relativas) 80 0.5 0.4 0.4 0.3 0.2 0.1 0.0 30 70 Recuperação () 0.5 0.0 30 60 THR (%) 40 50 60 70 80 90 0.4 0.3 0.2 0.1 0.0 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.22 – Variação do rendimento quântico efectivo do PSII (ΦPSII) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 70 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Até a um THR de 80% (stresse baixo), este parâmetro mantém-se constante entre as 0,7 e 0,8 unidades relativas. Com a diminuição do THR (agravamento do stresse), há um decréscimo deste parâmetro até às 0,6 unidades relativas. As variedades AD3R e PB304 foram as únicas que apresentaram valores inferiores a este. No caso de AD3R, apresentou valores inferiores a este no intervalo de THR de 50-70% (stresse moderado), e no caso de PB304 apresentou um único ponto inferior a este para um THR de cerca de 40% (stresse intenso). Tal facto indica um efeito negativo nos centros de reacção do PSII, provavelmente devido a degradação dos componentes (van Wijk et al., 1994; Mattos et al., 1999; Maxwell e Johnson, 2000). De qualquer modo, este parece ser um parâmetro bastante robusto e pouco sensível a estes níveis de stresse testados. Após a iluminação das folhas, o ΦPSII apresentou, perto da hidratação total, um valor próximo de 0,35 nas variedades AD3R, PB260, PB304 e PB369 e próximo de 0,25 nas variedades PB64 e PB269, decrescendo à medida que o THR diminuía (Fig. 4.22). As variedades AD3R, PB64, PB304 e PB369 são aquelas em que este decréscimo foi mais acentuado, mesmo em stresse moderado (THR entre 50% e 80%). PB64 foi a variedade que apresentou um valor de ΦPSII superior ao inicialmente observado, após irrigação. Por outro lado, em PB260 e PB269 o decréscimo não foi tão marcado durante o stresse moderado, e só PB260 apresentou variações significativas em stresse intenso (THR inferior a 50%). Este parâmetro é assim um bom indicador do estado geral da fotossíntese quando comparado com a evolução de A (ver Fig. 4.6). O parâmetro qP indica-nos que, perto da hidratação total, existe um maior amortecimento fotoquímico nas variedades AD3R, PB260, PB304 e PB369 (perto de 0,7) e menor nas variedades PB64 e PB269 (perto de 0,6) (Fig. 4.23). À medida que o THR diminui, qP também diminui fornecendo uma indicação da energia utilizada para as reacções fotoquímicas (Maxwell e Johnson, 2000). Esta diminuição, também observada por Tezara et al. (2003) em Lycium nodosum e por Ögren (1990) em Salix sp., é mais visível nas variedades AD3R e PB304, enquanto que nas restantes variedades a variação é muito menor. Apenas na variedade PB260 ocorre uma diminuição mais acentuada para THR inferiores a 50% (stresse intenso), e nas variedades PB269 e PB369 o parâmetro praticamente não varia nos intervalos de THR considerados na Fig. 4.23. Na maioria das variedades analisadas, estes dados indicam que não há alterações na dissipação da energia fotoquímica, indicando uma integridade da cadeia transportadora de electrões até THR de cerca de 50%, sendo apenas afectada para stresse hídrico intenso. Após irrigação, o parâmetro recuperou para valores perto dos iniciais Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 71 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho em todas as variedades, chegando em alguns casos a apresentar valores superiores (variedades PB64 e PB260). PB64 AD3R 1.0 qP (unidades relativas) qP (unidades relativas) 1.0 0.8 0.6 0.4 30 40 50 60 70 80 90 0.8 0.6 0.4 30 100 40 50 Stresse (∆) Stresse (∆) Recuperação () PB260 qP (unidades relativas) qP (unidades relativas) 90 100 1.0 0.6 40 50 60 70 80 90 0.8 0.6 0.4 30 100 THR (%) 40 50 60 70 80 90 100 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB304 PB369 1.0 1.0 qP (unidades relativas) qP (unidades relativas) 80 PB269 0.8 0.8 0.6 0.4 30 70 Recuperação () 1.0 0.4 30 60 THR (%) THR (%) 40 50 60 70 80 90 0.8 0.6 0.4 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.23 – Variação do coeficiente de amortecimento fotoquímico (qP) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. O parâmetro qN possuía, perto da hidratação total, um valor entre 0,75 e 0,8 para todas as variedades (Fig. 4.24). Em todas as variedades este parâmetro sofreu uma evolução semelhante à do parâmetro qP mas em sentido oposto, ou seja, naquelas variedades em que Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 72 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho ocorreu variação (AD3R e PB304), esta ocorre no sentido de aumentar ligeiramente o seu valor, tal como em Lycium nodosum (Tezara et al., 2003). Assim, também a variedade PB260 só demonstra um ligeiro aumento do parâmetro para THR superiores a 50% e as variedades PB269 e PB369 praticamente não sofrem alterações. AD3R PB64 1.0 qN (unidades relativas) qN (unidades relativas) 1.0 0.9 0.8 0.7 0.6 30 40 50 60 70 80 90 0.9 0.8 0.7 0.6 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () qN (unidades relativas) qN (unidades relativas) 90 100 80 90 100 80 90 1.0 0.9 0.8 0.7 40 50 60 70 80 90 0.9 0.8 0.7 0.6 30 100 40 50 60 70 THR (%) THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB304 PB369 1.0 1.0 qN (unidades relativas) qN (unidades relativas) 80 PB269 1.0 0.9 0.8 0.7 0.6 30 70 Recuperação () PB260 0.6 30 60 THR (%) 40 50 60 70 80 90 0.9 0.8 0.7 0.6 30 100 THR (%) 40 50 60 70 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.24 – Variação do coeficiente de amortecimento não-fotoquímico (qN) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 73 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Estes dados são coerentes com os anteriores na medida em que quando a cadeia de electrões é afectada pelo stresse hídrico, o excesso de energia é dissipada por processos nãofotoquímicos, de modo a proteger a integridade da folha (Horton et al., 1994). Após irrigação o parâmetro regressou aos valores iniciais em todas as variedades. AD3R PB64 150 ETR (µ mol.m-2.s-1) ETR (µ mol.m-2.s-1) 150 120 90 60 30 0 30 40 50 60 70 80 90 120 90 60 30 0 30 100 40 50 THR (%) Stresse (∆) Stresse (∆) Recuperação () PB260 ETR (µ µ mol.m-2.s-1) ETR (µ mol.m-2.s-1) 90 100 80 90 100 80 90 100 PB269 90 60 30 40 50 60 70 80 90 120 90 60 30 0 30 100 40 50 THR (%) 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () PB304 PB369 150 150 ETR (µ µ mol.m-2.s-1) ETR (µ mol.m-2.s-1) 80 150 120 120 90 60 30 0 30 70 Recuperação () 150 0 30 60 THR (%) 40 50 60 70 80 90 120 90 60 30 0 30 100 THR (%) 40 50 60 70 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 4.25 – Variação da taxa de transporte electrónico (ETR) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 4 ou 5 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 74 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Na ETR verifica-se uma diminuição acentuada nas variedades AD3R e PB304 e nas restantes variedades uma diminuição menos acentuada, implicando uma redução do transporte de electrões com a diminuição do THR, que poderá resultar de uma diminuição do metabolismo fotossintético do carbono (Fig.4.25) (Maxwell e Johnson, 2000). Apenas na variedade PB260 se verifica uma diminuição mais acentuada para stresse hídrico intenso (THR inferior a 50%). Na variedade PB269 a variação é muito pequena, apresentando valores superiores de ETR após irrigação. Estes dados estão de acordo com os dados de A obtidos nos ensaios de trocas gasosas (Fig. 4.6). No entanto, na variedade PB369 observam-se valores elevados de ETR, mesmo durante a imposição do stresse e que parecem contradizer os valores de A observados. Tal facto poderá estar relacionado com um aumento do transporte de electrões cíclico em torno do PSII devido à reduzida actividade do ciclo de Calvin, aliviando o excesso de energia em redor do PSII, já observado em girassol (Laisk et al., 2006). Após irrigação, o parâmetro recuperou para valores perto dos iniciais, tendo as variedades PB64 e PB269 apresentado valores superiores aos observados inicialmente. 4.3.3.2. Stresse Rápido Em todas as variedades, observou-se um decréscimo da área (normalizada) complementar à fase rápida da curva de Kautsky, sendo maior na variedade PB369 e menor na variedade PB269 (Fig. 4.26), em semelhança ao já observado nos ensaios de stresse lento (Fig. 4.20). No parâmetro Fv/Fm constatou-se pouca variação do parâmetro, embora demonstrasse uma ligeira descida em todas as variedades à medida que se aproximavam do THR de 40% (Fig. 4.27). Nas variedades AD3R, PB260 e PB269 a diminuição foi bastante reduzida, sendo AD3R e PB269 aquelas que apresentavam variação mais pequena. Comparativamente ao stresse lento (Fig. 4.21), notam-se algumas variações principalmente na variedade AD3R. Esta apresentava um decréscimo acentuado no stresse lento que não se observa no stresse rápido, o que indicará que a velocidade de imposição do stresse desempenha um papel importante na evolução deste parâmetro associado com uma diminuição da eficiência fotoquímica (Lizana et al., 2006). Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 75 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho AD3R PB64 1000 (mV*ms)/(F0*1000) (mV*ms)/(F0*1000) 1000 750 500 250 0 40 50 60 70 80 90 750 500 250 0 40 100 50 60 THR (%) y = 312,5 + 4,044x (r2 = 0,50 ; p < 0,05) PB260 90 100 PB269 1000 (mV*ms)/(F0*1000) (mV*ms)/(F0*1000) 80 y = – 116,7 + 6,987x (r2 = 0,43 ; p < 0,05) 1000 750 500 250 0 40 50 60 70 80 90 750 500 250 0 40 100 50 60 THR (%) 70 80 90 100 THR (%) y = 135,6 + 4,947x (r2 = 0,64 ; p < 0,01) y = 237,9 + 3,552x (r2 = 0,75 ; p < 0,01) PB304 PB369 1000 1000 (mV*ms)/(F0*1000) (mV*ms)/(F0*1000) 70 THR (%) 750 500 250 0 40 50 60 70 80 90 750 500 250 0 40 100 THR (%) 50 60 70 80 90 100 THR (%) y = 106,3 + 6,379x (r2 = 0,81 p < 0,01) y = – 25,97 + 8,349x (r2 = 0,85 ; p < 0,01) Fig. 4.26 – Variação da área complementar (normalizada pela fluorescência basal) à fase rápida da curva de Kautsky, em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 76 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho PB64 AD3R 0.8 Fv/Fm (unidades relativas) Fv/Fm (unidades relativas) 0.8 0.7 0.6 0.5 0.4 30 40 50 60 70 80 90 0.7 0.6 0.5 0.4 30 100 40 50 THR (%) y = 0,5901 + 0,001441x (r2 = 0,64 ; p = 0,01) Fv/Fm (unidades relativas) Fv/Fm (unidades relativas) 90 100 0.8 0.7 0.6 0.5 40 50 60 70 80 90 0.7 0.6 0.5 0.4 30 100 40 50 THR (%) 60 70 80 90 100 THR (%) y = 0,5446 + 0,001746x (r2 = 0,61 ; p < 0,05) y = 0,5578 + 0,001419x (r2 = 0,76 ; p < 0,01) PB304 PB369 0.8 0.8 Fv/Fm (unidades relativas) Fv/Fm (unidades relativas) 80 PB269 PB260 0.7 0.6 0.5 0.4 30 70 y = 0,4142 + 0,002773x (r2 = 0,53 ; p < 0,05) 0.8 0.4 30 60 THR (%) 40 50 60 70 80 90 0.7 0.6 0.5 0.4 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) y = 0,479 + 0,002638x (r2 = 0,73 ; p < 0,01) y = 0,5382 + 0,002111x (r2 = 0,71 ; p < 0,01) Fig. 4.27 – Variação do rendimento quântico máximo do PSII (Fv/Fm) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Após a iluminação das folhas, o ΦPSII apresentou um valor entre 0,3 e 0,4 perto da hidratação total, decrescendo à medida que o THR diminui (Fig. 4.28). Todas as variedades apresentam um valor nulo de ΦPSII a THR de cerca de 40%, à excepção da variedade PB64, em que tal acontece a THR de cerca de 60%, e da variedade PB260 que a 40% ainda apresenta um Φ PSII de 0,1 unidades relativas. O fecho progressivo dos centros de reacção do Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 77 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho PSII explica a diminuição de A de forma semelhante (Fig. 4.13), já observado em cultivares de trigo (Loggini et al., 1999). AD3R PB64 0.4 Φ PSII (unidades relativas) Φ PSII (unidades relativas) 0.4 0.3 0.2 0.1 0.0 30 40 50 60 70 80 90 0.3 0.2 0.1 0.0 30 100 THR (%) 40 50 y = – 0,008431 – 0,001983x + 5,778e-5x2 (r2 = 0,97 ; p < 0,05) Φ PSII (unidades relativas) Φ PSII (unidades relativas) 90 100 0.4 0.3 0.2 0.1 40 50 60 70 80 90 0.3 0.2 0.1 0.0 30 100 40 50 THR (%) 60 70 80 90 100 THR (%) y = 0,1772 – 0,004758x + 6,393e-5x2 (r2 = 0,86 ; p < 0,05) y = – 0,1428 + 0,003977x (r2 = 0,94 ; p < 0,01) PB369 PB304 0.4 0.4 Φ PSII (unidades relativas) Φ PSII (unidades relativas) 80 PB269 PB260 0.3 0.2 0.1 0.0 30 70 y = – 0,3505 + 0,006734x (r2 = 0,78 ; p < 0,01) 0.4 0.0 30 60 THR (%) 40 50 60 70 80 90 0.3 0.2 0.1 0.0 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) y = 0,05687 – 0,002733x + 5,92e-5x2 (r2 = 0,98 ; p < 0,05) y = – 0,2539 + 0,006313x (r2 = 0,87 ; p < 0,01) Fig. 4.28 – Variação do rendimento quântico efectivo do PSII (ΦPSII) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Relativamente a qP, as variedades apresentam um valor de cerca de 0,7 a 0,8 unidades relativas perto da hidratação total, à excepção da variedade PB269 que apresenta um valor mais baixo (0,6 unidades relativas) (Fig. 4.29). Comparativamente ao stresse lento, existe um Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 78 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho decréscimo mais acentuado deste parâmetro ao longo da imposição do stresse, apresentando um declive mais marcado na variedade PB64 e um declive mais suave nas variedades PB260 e PB269. Estes dados indicam que o fluxo electrónico é rapidamente desviado dos processos fotoquímicos da fotossíntese para mecanismos de dissipação de energia não-fotoquímicos (Ögren, 1990; Tezara et al., 2003). PB64 AD3R 0.8 qP (unidades relativas) qP (unidades relativas) 0.8 0.6 0.4 0.2 0.0 30 40 50 60 70 80 90 0.6 0.4 0.2 0.0 30 100 40 50 THR (%) y = – 0,4104 + 0,01177x (r2 = 0,95 ; p < 0,01) qP (unidades relativas) qP (unidades relativas) 90 100 0.8 0.6 0.4 0.2 40 50 60 70 80 90 0.6 0.4 0.2 0.0 30 100 40 50 THR (%) 60 70 80 90 100 THR (%) y = – 0,01262 + 0,007054x (r2 = 0,82 ; p < 0,01) y = – 0,1592 + 0,007981x (r2 = 0,92 ; p < 0,01) PB369 PB304 0.8 0.8 qP (unidades relativas) qP (unidades relativas) 80 PB269 0.8 0.6 0.4 0.2 0.0 30 70 y = – 0,6247 + 0,01392x (r2 = 0,74 ; p < 0,01) PB260 0.0 30 60 THR (%) 40 50 60 70 80 90 0.6 0.4 0.2 0.0 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) y = – 0,07762 + 0,008503x (r2 = 0,80 ; p < 0,01) y = – 0,3384 + 0,01122x (r2 = 0,84 ; p < 0,01) Fig. 4.29 – Variação do coeficiente de amortecimento fotoquímico (qP) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 79 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho O parâmetro qN (Fig. 4.30) demonstrou um aumento maior do que aquele verificado no stresse lento. No final do período de stresse rápido, todas as variedades apresentavam um qN de 0,9 unidades relativas. A variedade PB64 foi aquela em que apresentou um maior declive (em módulo), apresentando o referido valor de qN num THR mais elevado que as restantes variedades (cerca de 60%). Por outro lado, as variedades PB260 e PB269 foram aquelas que apresentaram o menor declive (em módulo), de um modo semelhante ao que sucedeu no stresse lento (Fig. 4.24). AD3R PB64 1.0 qN (unidades relativas) qN (unidades relativas) 1.0 0.9 0.8 0.7 0.6 30 40 50 60 70 80 90 0.9 0.8 0.7 0.6 30 100 40 50 THR (%) y = 1,059 – 0,003503x (r2 = 0,92 ; p < 0,01) PB260 qN (unidades relativas) qN (unidades relativas) 90 100 PB269 0.8 0.7 40 50 60 70 80 90 0.9 0.8 0.7 0.6 30 100 40 50 THR (%) 60 70 80 90 100 THR (%) y = 0,933 – 0,002277x (r2 = 0,51 ; p < 0,05) y = 0,9666 – 0,002105x (r2 = 0,90 ; p < 0,01) PB304 PB369 1.0 1.0 qN (unidades relativas) qN (unidades relativas) 80 1.0 0.9 0.9 0.8 0.7 0.6 30 70 y = 1,184 – 0,005371x (r2 = 0,67 ; p < 0,01) 1.0 0.6 30 60 THR (%) 40 50 60 70 80 90 0.9 0.8 0.7 0.6 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) y = 1,006 – 0,003030x (r2 = 0,96 ; p < 0,01) y = 1,041 – 0,003429x (r2 = 0,82 ; p < 0,01) Fig. 4.30 – Variação do coeficiente de amortecimento não-fotoquímico (qN) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 80 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Em todas as variedades, ETR diminui de forma mais acentuada que nos ensaios de stresse lento. Mais uma vez, PB64 apresentou a variação mais acentuada, em oposição ao que acontece às variedades PB260 e PB269 (Fig. 4.31). Estes dados indicam um cessar do transporte da cadeia de electrões que poderá resultar de danos ao nível dos aceitadores de electrões, ou ao nível do metabolismo fotossintético do carbono (Chaves e Oliveira, 2004). AD3R PB64 150 ETR (µ mol.m-2.s-1) ETR (µ µ mol.m-2.s-1) 150 120 90 60 30 0 30 40 50 60 70 80 90 120 90 60 30 0 30 100 40 50 THR (%) y = – 2,886 – 0,7005x + 0,02038x2 (r2 = 0,97 ; p < 0,05) ETR (µ µ mol.m-2.s-1) ETR (µ mol.m-2.s-1) 90 100 150 120 90 60 30 40 50 60 70 80 90 120 90 60 30 0 30 100 40 50 THR (%) 60 70 80 90 100 THR (%) y = 61,64 – 1,652x + 0,02236x2 (r2 = 0,86 ; p < 0,05) y = – 50,24 + 1,403x (r2 = 0,94 ; p < 0,01) PB304 PB369 150 150 ETR (µ mol.m-2.s-1) ETR (µ µ mol.m-2.s-1) 80 PB269 PB260 120 90 60 30 0 30 70 y = – 123,8 + 2,378x (r2 = 0,78 ; p < 0,01) 150 0 30 60 THR (%) 40 50 60 70 80 90 120 90 60 30 0 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) y = 19,96 – 0,9623x + 0,02088x2 (r2 = 0,98 ; p < 0,05) y = – 89,68 + 2,229x (r2 = 0,87 ; p < 0,01) Fig. 4.31 – Variação da taxa de transporte electrónico (ETR) em função do teor hídrico relativo (THR), nas experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 81 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho Em resumo, nos ensaios de fluorescência modulada da clorofila a verificou-se uma diminuição de vários parâmetros (ΦPSII, qP, ETR), aumento de uns (qN) e manutenção de outros (Fv/Fm), alterações já referidas por outros autores (Mattos et al., 1999; Marques da Silva, 1999; Maxwell e Johnson, 2000; Tezara et al., 2003). Comparando os ensaios de stresse lento e de stresse rápido, verifica-se que os parâmetros diminuem de forma mais marcante no stresse rápido. Tal facto, revela a rapidez da resposta da planta, resultante da sensibilidade e/ou dano dos seus constituintes ao aumento do stresse hídrico. A aclimatação da planta, durante o stresse lento, permite o estabelecimento de formas de tolerância ao stresse hídrico, quer por reorientações nas vias de vários metabolitos, quer pela expressão de chaperonas (Rizhsky et al., 2002; Sharp et al., 2004), o qual não é possível no stresse rápido. Também a área complemetar (normalizada) à fase rápida da curva de Kautsky, que permite inferir sobre a dimensão do pool de quinonas (Lazár, 1999), sofreu um decréscimo em ambos os tipos de stresse (Mattos et al., 1999). Desta forma, observa-se uma sobrerredução das quinonas com a diminuição do THR que vai diminuir o transporte de electrões (Chaves e Oliveira, 2004). Além disso, observou-se um aumento de qN em quase todas as variedades, o que, em conjunto com a diminuição dos outros parâmetros, permite inferir sobre a possibilidade de existir dano em algum passo do metabolismo fotossintético do carbono, uma vez que o fluxo de electrões foi desviado, quase na sua totalidade, para mecanismos de dissipação de energia não-fotoquímicos. No caso da variedade PB269, verificou-se que qN não sofreu alterações ao longo do stresse lento, ao contrário do que se observou nos ensaios de stresse rápido. Esta variedade poderá possuir outros mecanismos que lhe permitam proteger os fotossistemas de danos provocados pela radiação luminosa, que não envolvam alterações de qN. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 82 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho 4.4 4.4. Conclusões 4.4.1. 4.4.1. Conclusões das Trocas Gasosas Relativamente às variedades analisadas, destacam-se as variedades AD3R, PB260 e PB269 como aquelas que apresentam características de uma maior tolerância ao stresse hídrico e a variedade PB369 como a que possui maiores dificuldades neste tipo de stresse. A variedade AD3R apresenta uma WUE elevada, com A elevada perto da hidratação total. No entanto, após a irrigação, embora tenha recuperado o THR, não conseguiu recuperar A para os valores iniciais, antes de se ter iniciado o período de stresse. A variedade PB260 apresenta um bom desempenho durante a imposição do stresse lento moderado e intenso, tolerando THR baixos. No entanto, após irrigação, apresenta dificuldades em recuperar, quer os valores de THR quer os valores de A, gs e E. A variedade PB269 apresenta uma elevada WUE, A e gs durante o stresse lento e em condições de ausência de stresse, demonstrando uma elevada produtividade, que se traduziu em plantas de elevado porte. Após irrigação, apresenta uma boa recuperação dos parâmetros, mesmo com THR inferiores aos registados na ausência de stresse. Por outro lado, a variedade PB369 apresenta um mau desempenho durante a imposição do stresse hídrico, com baixos valores de A e gs, e, após a irrigação, demonstra uma má recuperação dos parâmetros avaliados. 4.4.2 4.4.2. 4.2. Conclusões da Fluorescência Modulada da Clorofila a A forma como as plantas responderam ao stresse rápido foi diferente da forma como responderam ao stresse lento. Na primeira situação, a planta reage rapidamente e de forma brusca às novas condições que lhe são impostas. Todas as variedades parecem reagir de uma forma semelhante. Na segunda situação, a planta sofrendo um processo de adaptação, passando por uma fase de aclimatação, variando os parâmetros de forma quase sempre mais suave. Notam-se também maiores diferenças entre as variedades. No primeiro caso é necessário uma reacção rápida e que é certamente resultante de mecanismos constitutivos ou mesmo de danos, enquanto que, no segundo caso, a reacção ao stresse lento vai resultar mais de mecanismos induzidos e de um efeito cumulativo. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 83 4. Fase I – Análise Preliminar de Seis Variedades Portuguesas de Milho 4.4.3. 4.4.3. Conclusões Conclusões da Fase I A avaliação de todos os parâmetros analisados permitiu a escolha das variedades que mais se destacaram pelas suas respostas contrastantes ao stresse hídrico. Por um lado, as variedades PB260 e PB269 mostraram ser as que melhor toleravam o stresse hídrico, adaptando as trocas gasosas com a diminuição do THR e apresentando uma redução progressiva dos parâmetros, nomeadamente a taxa fotossintética líquida e a condutividade estomática, semelhante ao grupo de resposta ao stresse de tipo I (Lawlor e Cornic, 2002). No entanto, uma vez que PB260 não recuperou no parâmetro da WUE optouse pela variedade PB269 como a mais tolerante ao stresse hídrico. Por outro lado, a variedade PB369, demonstrou, desde cedo, ser a mais susceptível ao stresse hídrico nas experiências de stresse lento, apresentando sintomas de dano a THR mais elevados do que as restantes variedades. O comportamento desta variedade enquadra-se no modelo referido neste trabalho como “tudo-ou-nada”, que consiste na manutenção dos parâmetros até um valor crítico de THR, a partir do qual ocorria uma diminuição drástica desses mesmos parâmetros, passando de um patamar de valores para outro, semelhante ao grupo de resposta ao stresse de tipo II (Lawlor e Cornic, 2002). Também durante o período de recuperação apresentou maiores dificuldades em regressar aos valores iniciais dos parâmetros analisados. Por estas razões, foi seleccionada como a variedade mais susceptível ao stresse hídrico. Stresse Hídrico e Recuperação do Stresse em Variedades Portuguesas de Milho 84 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5. Fase II – Estudo Comparativo Comparativo Aprofundado Aprofundado das das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.1. Introdução Nesta fase, pretendeu-se aprofundar o conhecimento das duas variedades seleccionadas na fase I, PB269 e PB369. Procedeu-se a uma série de experiências em que as trocas gasosas foram medidas simultaneamente com a fluorescência modulada da clorofila a, nomeadamente as respostas a diferentes intensidades luminosas e a diferentes concentrações de CO2 das duas variedades ao longo do stresse hídrico. Foi ainda avaliada a libertação de O2 ao longo do stresse. As actividades experimentais envolvendo estes factores foram apenas realizadas em condições de stresse lento. Este tipo de ensaios tem uma duração demasiado longa para permitir a sua utilização nos ensaios de stresse rápido, que levariam a uma perda de água demasiado rápida para ser possível retirar qualquer tipo de conclusão. Foram também avaliadas as actividades enzimáticas de enzimas chaves do metabolismo do carbono e a quantificação de solutos compatíveis, como a prolina. Estes parâmetros foram analisados nos dois tipos de stresse, de modo a avaliar possíveis diferenças. 5.2. Metodologia 5.2.1. Material Vegetal Foram utilizadas as variedades de milho PB269 e PB369. As plantas foram obtidas segundo o descrito em 3.1.. 5.2.2. Relações Hídricas 5.2.2.1. Determinação do Teor Hídrico Relativo da Folha O THR foi determinado segundo o descrito em 3.3.1.. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 85 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.2.2.2. Determinação do Potencial Hídrico das Folhas O potencial hídrico das folhas foi determinado psicrometricamente, ao longo da experiência de stresse lento, através do psicrómetro HR-33T (Wescor, Logan, USA), equipado com câmaras C-52 (Wescor, Logan, USA) e ligado a um registador LKB REC101 (Pharmacia LKB Biotechnology AB, Uppsala, Sweden). Retirou-se da região analisada pelo IRGA e pelo fluorómetro, um disco de folha com 5 mm de diâmetro, tendo sido colocado no interior da câmara do psicrómetro, onde permaneceu durante 30 minutos (tempo de equilíbrio). Durante este período de tempo, o potencial hídrico da amostra vai entrar em equilíbrio com o da atmosfera da câmara, assumindo-se que a água perdida pela amostra não é suficiente para afectar o potencial hídrico da folha (Marques da Silva, 1999). Nesta experiência foram utilizadas as mesmas plantas da experiência de determinação das curvas A/PPFD (descritas em 5.2.3.1.). 5.2.2.3. Estabelecimento de Curvas Pressão/Volume As curvas pressão/volume permitiram averiguar as relações hídricas em folhas das variedades portuguesas PB269 e PB369 de milho não sujeitas a stresse hídrico, tendo sido utilizado para tal o psicrómetro HR-33T (Wescor, Logan, USA), equipado com câmaras C-52 (Wescor, Logan, USA) e ligado a um registador LKB REC101 (Pharmacia LKB Biotechnology AB, Uppsala, Sweden). Cortaram-se folhas totalmente expandidas, dividindo-se em cinco fragmentos, cada um pesado numa balança de precisão. Os fragmentos foram hidratados até à saturação, flutuando-os em água destilada durante 12 horas, à temperatura ambiente e sob condições de luminosidade reduzida. Retirou-se de um dos fragmentos um disco com 5 mm de diâmetro (com a ajuda de um furador) e colocou-se na câmara do psicrómetro, durante 30 minutos (tempo de equilíbrio). Os restantes fragmentos foram colocados em contacto com a atmosfera do laboratório, em processo de desidratação. De um dos fragmentos não se retiraram discos, tendo sido utilizado como controlo para cálculo do THR (ver 3.3.1.), sendo pesado antes e após cada medição do psicrómetro, assumindo-se como idêntico aos restantes fragmentos. Os parâmetros hídricos foram calculados segundo Matos (1990). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 86 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.2.3. Medição das Trocas Gasosas 5.2.3.1. Curvas A/PPFD Para observar a resposta das duas variedades a diferentes PPFD ao longo do stresse lento, utilizou-se o IRGA de modelo LCPro+ (ADC – The Analytical Development Co., Ltd., Hoddesdon, England), em conjunto com o fluorómetro PAM 101 (Heinz Walz GmbH, Effeltrich, Germany) (ver 5.2.4.). Os ensaios com o IRGA LCPro+ foram realizados em condições semelhantes às descritas anteriormente (ver 4.2.3.), à excepção da PPFD, fornecida por uma fonte de luz externa KL1500 LCD (Schott AG, Mainz, Germany). As plantas foram ensaiadas a sete PPFD diferentes (65, 100, 140, 200, 430, 720 e 1320 µmol fotões.m-2.s-1), sendo a primeira aplicada por um período de 15 minutos e as seguintes por períodos de 9 minutos. As plantas foram ensaiadas a cada 3 dias durante duas semanas, correspondendo ao início, meio e fim do stresse, e ao meio e fim da recuperação. Durante este ensaio foram recolhidas amostras das folhas analisadas para posterior análise das actividades enzimáticas e solutos compatíveis, tendo sido congeladas em azoto líquido e armazenadas numa arca congeladora a –70ºC (DLT-13V-85V12, Harris Manufacturing, Co., Asheville, USA). 5.2.3.2. 5.2.3.2. Curvas A/CO2a Para observar a resposta das duas variedades a diferentes concentrações de CO2 ao longo do stresse lento, utilizou-se o IRGA de modelo LCPro+ (ADC – The Analytical Development Co., Ltd., Hoddesdon, England), em conjunto com o fluorómetro PAM 101 (Heinz Walz GmbH, Effeltrich, Germany) (ver 5.2.4.). Os ensaios com o IRGA LCPro+ foram realizados em condições semelhantes às descritas anteriormente (ver 4.2.3.), à excepção da fonte de luz externa (KL1500 LCD, Schott AG, Mainz, Germany) e das diferentes concentrações de CO2, controladas pelo próprio IRGA. As plantas foram ensaiadas a seis concentrações de CO2 diferentes (100, 200, 370, 700, 1000 e 1400 ppm), iniciando-se nos 370 ppm, por um período de 15 minutos, e as seguintes por períodos de 9 minutos. Inicialmente, a concentração ia aumentando até aos 1400 ppm, com posterior diminuição até aos 100 ppm. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 87 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico As plantas foram ensaiadas a cada 3 dias durante duas semanas, correspondendo ao início, meio e fim do stresse, e ao meio e fim da recuperação. Durante este ensaio, foram recolhidos fragmentos de material vegetal para determinação da libertação de O2 (ver 5.2.3.3.). As limitações estomáticas (Ls) e não-estomáticas (Lns) foram determinadas utilizando a equação Ls = 100 × (A0 – A) / A0, em que A0 é a taxa fotossintética medida a Ci de 370 ppm e A é a taxa fotossintética medida a CO2a de 370 ppm, e a equação Lns = 100 × (AC – AS) / AC, em que AC é a taxa fotossintética de folhas controlo para Ci de 800 ppm e AS é a taxa fotossintética de folhas sujeitas a stresse hídrico para Ci de 800 ppm (Jones, 1985; Geber e Dawson, 1997; Tezara et al., 2003). 5.2.3.3. Determinação da Libertação de O2 A libertação de O2 foi efectuada sob condições saturantes de CO2 (aproximadamente 7%) recorrendo a um eléctrodo de fase gasosa (LD2, Hansatech Instruments Ltd., Norfolk, UK) à temperatura controlada de 25ºC e sob uma PPFD de 850 µmol fotões.m-2.s-1 fornecida por uma fonte luminosa (LS2, Hansatech Instruments Ltd., Norfolk, UK). Foram recolhidas porções da folha analisada, ao longo do período de stresse lento, de modo a preencher a câmara do eléctrodo. O peso fresco foi determinado numa balança de precisão (BP 210D, Sartorius, Goettingen, Germany) e a área num medidor de áreas (CI-202 Portable Leaf Area Meter, CID, Inc., Camas, Washington, USA). Nesta experiência foram utilizadas as mesmas plantas do ensaio da determinação das curvas A/CO2a (descritas em 5.2.3.2.). 5.2.4. Medição da Fluorescência Modulada da Clorofila a A medição da fluorescência modulada da clorofila a foi efectuada recorrendo ao fluorómetro PAM 101 (Heinz Walz GmbH, Effeltrich, Germany). A medição da razão Fv/Fm foi determinada aplicando um pulso de luz saturante (KL2500 LCD, Schott AG, Mainz, Germany), após as folhas estarem 10 minutos na escuridão. No ensaio das curvas A/PPFD, após a última PPFD, a luz era desligada e iniciava-se uma série de pulsos saturantes no escuro. Durante 4 minutos aplicou-se os pulsos com intervalos de tempo de 40 segundos, durante os 10 minutos seguintes com intervalos de 100 segundos e durante os últimos 5 minutos com intervalos de 200 segundos. O amortecimento Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 88 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico de alta energia (qE), o amortecimento da transição de estado (qT) e o amortecimento fotoinibitório (qI) foram calculados segundo Quick e Stitt (1989). 5.2.5. 5.2.5. Determinação de Clorofilas e Carotenóides Na determinação de clorofilas e carotenóides foram utilizados os fragmentos de folha provenientes dos ensaios de determinação da libertação de O2 (ver 5.2.3.3.). Os fragmentos de folha foram colocados em tubos de ensaio contendo 10 ml de metanol absoluto, durante 72 horas, período necessário à extracção dos pigmentos das folhas. As amostras foram lidas nos comprimentos de onda 470 nm, 652 nm e 665 nm num espectrofotómetro UV500 UV-Visible Spectro (Unicam, Waltham, USA), contra um branco de metanol absoluto. Para a determinação da quantidade de clorofilas (Ca, Cb, Ca+b) e carotenóides (Cx+c) foram utilizadas as seguintes equações (Lichtenthaler, 1987): Ca = 16,72 × A665 – 9,16 × A652 Cb = 34,09 × A652 – 15,28 × A665 Ca+b = 1,44 × A665 + 24,93 × A652 Cx+c = (1000 × A470 – 1,63 × Ca – 104,96 × Cb) / 221 5.2.6. Determinação de Actividades Enzimáticas Nesta fase do trabalho experimental foram avaliadas as actividades de três enzimas fundamentais no metabolismo fotossintético do carbono das duas variedades de milho: as duas enzimas carboxilativas fosfoenolpiruvato carboxilase (PEPC) e a ribulose-1,5-bisfosfato carboxilase/oxigenase (RuBisCO) e uma descarboxilativa, a enzima málica dependente de NADP (EM-NADP). Para além do ensaio de stresse lento, foi também realizado um ensaio de stresse rápido (ver 3.2.), durante o qual foram recolhidas amostras das folhas analisadas para posterior análise das actividades enzimáticas. Todas as amostras foram congeladas em azoto líquido e armazenadas numa arca congeladora a –70ºC (DLT-13V-85V12, Harris Manufacturing, Co., Asheville, USA). As actividades enzimáticas foram determinadas à temperatura de 25ºC ± 1ºC, devido ao facto de ter sido esta a temperatura de crescimento das plantas. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 89 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.2.6.1. Extracção As enzimas estudadas foram extraídas do material vegetal num extracto comum. O material vegetal foi recolhido durante a experiência de stresse lento para determinação das curvas A/PPFD e durante as experiências de stresse rápido da fase II. O meio de extracção foi adaptado de Edwards et al. (1988), sendo composto por HEPES-KOH 50 mM pH 7,3, DTT 10 mM, EDTA 1 mM, PVP25 5% (m/v), Polyclar AT 1% (m/v), PEG4000 6% (m/v), Triton X-100 0,5 % (v/v) e cocktail de inibidores de proteases 1% (v/v) (Sigma). Foi utilizado uma proporção de 20 ml de meio de extracção para cada 1 g de material vegetal para as amostras com THR superior a 70% e uma proporção de 30 ml de meio de extracção para cada 1 g de material vegetal para as amostras com THR inferior a 70%. As folhas foram esmagadas muito rapidamente num almofariz gelado com metade do meio de extracção a adicionar e uma pequena porção de areia de quartzo. Adicionou-se a outra metade do meio de extracção e transferiu-se o extracto para tubos Eppendorf gelados. Centrifugou-se durante 5 minutos a uma temperatura de 4ºC à velocidade de 13.500 g na centrífuga Sigma 2-16K (Sigma Laborzentrifugen GmbH, Osterode am Harz, Germany). Recuperou-se o sobrenadante e conservou-se em gelo até à determinação das actividades enzimáticas. 5.2.6.2. RibuloseRibulose-1,51,5-Bisfosfato Carboxilase/Oxigenase (RuBisCO (RuBisCO) RuBisCO) Na RuBisCO, foram determinadas as actividades inicial e total através da incorporação de carbono radioactivo (14C), segundo Perchorowicz et al. (1982). O meio para realizar o ensaio enzimático era composto por HEPES-KOH 50 mM pH 8,0 contendo MgCl2 30 mM e NaH14CO3 10 mM. Na determinação da actividade inicial, colocaram-se 500 µl de meio de ensaio juntamente com 50 µl de RuBP 6m M. Adicionaram-se 50 µl de extracto e após 1 minuto a reacção foi parada com 100 µl de HCl 2N. Para a determinação da actividade total, colocaram-se 500 µl de meio de ensaio com 50 µl de extracto. Após 3 minutos de activação, adicionaram-se 50 µl de RuBP 6 mM. A reacção foi parada de modo idêntico, ao fim de 1 minuto. As amostras foram transferidas para uma estufa a 80ºC. Após a evaporação de todo o meio, o resíduo foi dissolvido em 1 ml de água ultra-pura. Adicionaram-se 8 ml de líquido de cintilação (Ready Safe, Beckman, USA) aos frascos, tendo sido lidos num espectrómetro de cintilação LS 7800 (Beckman Instruments Inc., Fullerton, USA). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 90 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.2.6.3. Fosfoenolpiruvato Carboxilase (PEPC) Na enzima PEPC foi determinada a actividade em condições fisiológicas e a actividade máxima, adaptada a partir da metodologia de Donkin e Martin (1980). A actividade em condições fisiológicas foi determinada num meio composto por HEPES-KOH 50 mM pH 7,2, MgCl2 10 mM, NaHCO3 10 mM, PEP 2,5 mM e 12 unidades de malato desidrogenase (MDH), utilizando-se 20 µl de extracto. A reacção foi iniciada com 20 µl de NADH 10 mM. A actividade foi determinada a um comprimento de onda de 340 nm no espectrofotómetro UV500 UV-Visible Spectro (Unicam, Waltham, USA), com agitação. A actividade máxima foi determinada de modo semelhante, utilizando um meio composto por HEPES-HOH 50 mM pH 8,0 e PEP 10 mM, mantendo-se as restantes condições. 5.2.6.4. Enzima Málica Dependente Dependente de NADP (EM(EM-NADP) Na EM-NADP apenas foi determinada a actividade máxima, segundo o descrito em Simon (1987). A actividade máxima foi determinada num meio composto por HEPES-KOH 50 mM pH 8,0, MgCl2 30 mM, DTT 2,5 mM e L-malato 10 mM, utilizando-se 50 µl de extracto. A reacção foi iniciada com 20 µl de NADP+ 25 mM. A actividade foi determinada a um comprimento de onda de 340 nm no espectrofotómetro UV500 UV-Visible Spectro (Unicam, Waltham, USA), com agitação. 5.2.7. Quantificação de Proteína Solúvel A quantificação da proteína solúvel foi determinada pelo método de Bradford (1976), utilizando placas de ELISA, tendo sido lidas no espectrofotómetro MPRA4 Microplate reader (DuPont, Newton, USA). A recta de calibração foi efectuada utilizando BSA. Esta quantificação permitiu expressar as actividades enzimáticas por unidade de proteína solúvel. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 91 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.2.8. Quantificação Quantificação de Solutos Compatíveis A quantificação de solutos compatíveis, em particular a prolina (Živković et al., 2005), foi efectuada nas amostras recolhidas nos ensaios de stresse lento para determinação das curvas A/PPFD e num ensaio independente de stresse lento. Foi também realizado um ensaio de stresse rápido (ver 3.2.), durante o qual foram recolhidas amostras para posterior análise. As amostras foram congeladas em azoto líquido e armazenadas a –70ºC numa arca congeladora (DLT-13V-85V12, Harris Manufacturing, Co., Asheville, USA). 5.2.8.1. Extracção As amostras recolhidas durante os ensaios foram maceradas num almofariz arrefecido em azoto líquido. Adicionou-se 5 ml de etanol 85% (v/v) e verteu-se o extracto para um gobelé, onde permaneceu durante 30 minutos. Filtrou-se o extracto e recolheu-se num tubo de ensaio. 5.2.8.2. Quantificação de Aminoácidos Livres e Prolina Antes do início dos ensaios de quantificação de aminoácidos livres e de prolina, foram realizadas várias curvas de calibração utilizando soluções de L-leucina (Fluka, SigmaAldrich, Germany) e L-prolina (Fluka, Sigma-Aldrich, Japan) em etanol 85% (v/v), com concentrações entre 0 e 200 µg.ml-1. Para a quantificação dos aminoácidos livres e de prolina nas amostras adicionou-se em tubos Eppendorf 200 µl de extracto, 500 µl de tampão citrato 0,2 M (pH 5,0) contendo SnCl2.H2O 0,75 mM e 500 µl de solução 4% ninidrina (Fluka, Sigma-Aldrich, Chec Republic) em etilenoglicol. Foram realizadas três réplicas para cada amostra. Agitaram-se os tubos no vórtex e colocaram-se num banho de água a ferver durante 20 minutos, tendo o cuidado de fechar apenas parcialmente as tampas (de modo, a não aumentar demasiado a pressão dos tubos). Arrefeceram-se os tubos em gelo e diluíram-se as amostras em 5 ml de água-propanol 1:1 (v/v). Após 15 minutos, leu-se a absorvância das amostras no comprimento de onda 570 nm para os aminoácidos livres e 440 nm para a prolina, num espectrofotómetro UV-Visible 1-E Cary (Cary System, Varian Inc., Palo Alto, USA) utilizando o sistema Cary System (Software Issue v3.01, Instrument Software Issue v4.00, Varian Inc., Palo Alto, USA). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 92 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.3. Resultados e Discussão 5.3.1. Relações Hídricas O potencial hídrico (Ψw) das folhas foi seguido ao longo da experiência de stresse lento (Fig. 5.1). A variedade PB269 apresentou valores de cerca de −0,7 MPa perto da hidratação total, e a variedade PB369 valores de cerca −1,5 MPa. Nesta última, o valor reduzido de Ψw perto da hidratação total estará relacionado com as propriedades da parede celular, nomeadamente paredes mais rígidas (maior módulo de elasticidade) com menor capacidade de manter água ligada nas folhas (Marshall e Dumbroff, 1999). Na variedade PB269, observa-se uma diminuição mais suave do Ψw até −2 MPa (para THR de cerca de 55%), à medida que o THR diminui, do que na variedade PB369. Nesta última, há uma manutenção do Ψw em torno dos −2 MPa até THR de cerca de 70%, ocorrendo depois uma diminuição brusca deste parâmetro. As plantas da variedade PB269 apresentam valores de cerca de −5 MPa no final do período de stresse, correspondente a THR de 35%. A variedade PB369 atingiu valores de Ψw de −6,5 MPa a THR muito baixos (cerca de 20%). Estas plantas não sobreviveram e a recuperação foi realizada nas restantes plantas que apresentavam THR de cerca de 30% no final do período de stresse. 80 PB369 THR (%) 60 40 20 100 0 -1 -1 -2 -2 -3 Ψw (MPa) Ψw (MPa) 100 0 PB269 THR (%) -4 60 40 20 -3 -4 -5 -5 -6 -6 -7 80 -7 Stresse (∆; - -): y = – 9,851 + 0,1808x – 0,0009349x2 Stresse (∆; - -): y = – 8,954 + 0,1293x – 0,0005534x2 (r2 = 0,89 ; p < 0,05) (r2 = 0,97 ; p < 0,05) Recuperação () Recuperação (; –): y = – 10,33 + 0,197x – 0,001066x2 (r2 = 0,95 ; p < 0,05) Fig. 5.1 – Variação do potencial hídrico das folhas (Ψw) em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 93 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico O período de recuperação do stresse foi efectuado com a câmara climatizada a cerca de 38ºC(1), o que se traduziu em THR mais baixos e que não excederam os 75-80%, o que, no caso da variedade PB269 se traduziu numa incapacidade de recuperar o Ψw para valores próximos dos inicias, apresentando um Ψw de −2 MPa para um THR de 75%. Na Fig. 5.2 está representada a variação do Ψw em função do THR, para folhas destacadas de plantas bem irrigadas (ver 5.2.2.3). 100 0 90 80 PB269 PB369 THR (%) THR (%) 70 60 50 40 100 0 80 70 60 50 40 -1 Ψw (MPa) Ψw (MPa) -1 90 -2 -2 -3 -3 -4 -4 y = – 3,633 + 0,02748x (r2 = 0,59 ; p < 0,01) y = – 5,951 + 0,05354x (r2 = 0,77 ; p < 0,01) Fig. 5.2 – Variação do potencial hídrico das folhas (Ψw) em função do teor hídrico relativo (THR), para folhas destacadas de plantas bem irrigadas. Os pontos foram obtidos de 3 folhas de diferentes plantas. Perto da hidratação total, as duas variedades apresentam valores de Ψw de aproximadamente – 0,8 MPa. No caso da variedade PB369, este valor é superior ao observado no ensaio de stresse lento, dado que as folhas estiveram, durante a noite, a flutuar em água destilada e em condições de baixa luminosidade, garantindo um maior Ψw perto da hidratação total. A variedade PB269 não apresentou quaisquer diferenças significativas em relação ao método anterior. A partir deste ponto, verifica-se, em ambas as variedades, uma diminuição do Ψw com o THR, embora a variedade PB269 consiga manter o seu Ψw em cerca de – 2,5 MPa para um THR de 45%, enquanto que na variedade PB369, e para o mesmo THR, o Ψw diminui até cerca de – 4 MPa. A partir da Fig. 5.2, construíram-se as curvas pressão-volume da Fig. 5.3. (1) Resultado de uma avaria da câmara climatizada. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 94 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB369 1.5 1.5 1.0 1.0 1/Ψ Ψw (-MPa) 1/Ψw (-MPa) PB269 0.5 0.0 100 0.5 90 80 70 60 50 0.0 100 40 THR (%) 90 80 70 60 50 40 THR (%) y = 1,491 – 0,04241x + 0,0004196x2 (r2 = 0,74 ; p < 0,05) y = 1,423 – 0,04319x + 0,0004190x2 (r2 = 0,81 ; p < 0,05) Fig. 5.3 – Curvas pressão-volume de folhas destacadas de plantas bem irrigadas, construídas a partir da Fig. 5.2. A partir da Fig. 5.3 foi possível obter os valores de diversos parâmetros hídricos apresentados na Tabela 5.1. Tabela 5.1 – Parâmetros hídricos de folhas destacadas de plantas bem irrigadas, nas duas variedades portuguesas de milho. Parâmetro hídrico PB269 PB369 Potencial osmótico à turgescência máxima (Ψπo) (MPa) - 0,91 - 1,11 Potencial osmótico no ponto de perda de turgescência (Ψπtlp) (MPa) - 1,67 - 2,00 Teor hídrico relativo no ponto de perda de turgescência (THRtlp) (%) 72 79 0,76 0,89 Módulo de elasticidade à turgescência máxima (εo) (MPa) Embora não tenha sido possível, por insuficiência de material vegetal disponível, realizar curvas independentes, inviabilizando, portanto, uma validação estatística, a criação de uma curva única com os dados das folhas utilizadas permite fazer uma avaliação dos parâmetros hídricos nas duas variedades portuguesas de milho. A variedade PB369 atinge a perda de turgescência a THR superiores (79%) quando comparada com a variedade PB269 (72%). O valor de THR para a perda de turgescência observado na variedade PB369 é próximo do observado para o feto Ceratopteris richardii (81%), uma planta característica de locais húmidos (Augé et al., 1989). Por outro lado, a variedade PB269 apresenta um valor próximo do observado para a gramínea Seratia sphacelata (68%) (Marques da Silva, 1999), indicada como apresentando um bom comportamento em stresse hídrico moderado. Deste modo, a variedade PB269 apresenta, na turgescência máxima, um potencial osmótico ligeiramente mais elevado que na variedade PB369, o mesmo acontecendo no ponto de perda Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 95 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico de turgescência, o que poderá indicar uma melhor adaptação da variedade PB269, em caso de ocorrência de stresse hídrico, uma vez que possui uma maior capacidade “tampão” do seu sistema. Por outro lado, verifica-se um valor de módulo de elasticidade inferior na variedade PB269 (0,76 MPa), indicando uma parede mais elástica. Estes dados indicam que a variedade PB269 apresenta uma melhor capacidade de manter a turgescência das suas células, facto observado em espécies de locais mais secos (Lamberts et al., 1998). Estes dados estão de acordo com os valores observados de fotossíntese durante os ensaios. De facto, uma vez que PB269 mantém a turgescência a THR mais reduzidos, apesar de um maior potencial osmótico e uma maior elasticidade da parede, o metabolismo fotossintético do carbono poderá manter-se num nível superior ao de PB369, uma vez que a ultraestrutura das células da folha e a das próprias enzimas sofrerão menor alteração para THR mais reduzidos. 5.3.2. Trocas Gasosas Nestes ensaios procedeu-se à avaliação do efeito do stresse hídrico nas trocas gasosas em diferentes condições de PPFD e CO2 ambiente. 5.3.2.1. 5.3.2.1. Curvas A/PPFD Na Fig. 5.4 está representada a variação da taxa fotossintética líquida (A) em função da PPFD, para diferentes THR. Os valores baixos de A em condições controlo de hidratação não limitante, comparativamente aos observados por Maroco et al. (1999), poderão estar relacionados com as condições de crescimento, uma vez que o milho é uma planta que está adaptada a elevada radiação, apresentando normalmente valores superiores de A. No entanto, as câmaras climatizadas não permitiram a obtenção de valores de PPFD superiores a 500 µmol fotões.m2 -1 .s , condições semelhantes às utilizadas por Fracheboud et al. (1999), onde o milho apresenta valores de A idênticos. Também Maroco et al. (1998) obteve taxas fotossintéticas de cerca de 15 µmol CO2.m-2.s-1, em plantas de Amaranthus edulis (planta C4) crescidas em PPFD de 600 µmol fotões.m-2.s-1, tal como observado na gramínea Setaria sphacelata (Marques da Silva, 1999). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 96 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB269 Stresse PB369 Stresse 15 A (µ µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 15 10 5 0 250 -5 500 750 1000 1250 PPFD (µ µ mol fotões.m-2.s-1) 1500 10 5 0 250 -5 500 750 1000 1250 PPFD (µ µmol fotões.m-2.s-1) 1500 THR = 97,8±1,0 (;–): y = 8464 * ((1 – exp (– 0,024x / 8464)) * THR = 94,6±2,0 (;–): y = 17245 * ((1 – exp (– 0,02035x / exp (– 8,606x / 8464) (r2 = 0,94 ; p < 0,05) 17245)) * exp (– 16,53x / 17245) (r2 = 0,94 ; p < 0,05) THR = 68,8±14,9 (;- -): y = 927,7 * ((1 – exp (– 0,006057x / THR = 83,5±11,4 () 927,7)) * exp (– 0,4687x / 927,7) (r2 = 0,76 ; p < 0,05) THR = 26,7±11,2 (∆) THR = 37,0±3,2 (∆) PB269 Recuperação PB369 Recuperação 15 A (µ µ mol CO2.m-2.s-1) A (µ mol CO2.m-2.s-1) 15 10 5 0 250 -5 500 750 1000 1250 PPFD (µ µmol fotões.m-2.s-1) 1500 10 5 0 250 -5 500 750 1000 1250 PPFD (µ µmol fotões.m-2.s-1) 1500 THR = 76,8±3,2 (;–): y = 577,9 * ((1 – exp (– 0,005362x / 577,9)) THR = 73,9±1,4 (;–): y = 633,9 * ((1 – exp (– 0,006780x / 633,9)) * exp (– 0,5673x / 577,9) (r2 = 0,80 ; p < 0,05) * exp (– 0,4205x / 633,9) (r2 = 0,77 ; p < 0,05) THR = 76,1±5,7 (;- -): y = 391,6 * ((1 – exp (– 0,002228x / THR = 74,0±16,0 (;- -): y = 465,1 * ((1 – exp (– 0,003429x / 391,6)) * exp (– 0,1935x / 391,6) (r2 = 0,62 ; p < 0,05) 465,1)) * exp (– 0,2533x / 465,1) (r2 = 0,70 ; p < 0,05) THR = 37,0±3,2 (∆) THR = 26,7±11,2 (∆) Fig. 5.4 – Variação da taxa fotossintética líquida (A) em função da densidade de fluxo fotónico fotossintético (PPFD), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Perto da hidratação total, ambas as variedades apresentam um perfil semelhante da variação da A com a PPFD. Verifica-se uma certa diminuição na taxa fotossintética líquida a partir de PPFD de cerca de 1000 µmol fotões.m-2.s-1, valor em que se atinge a A máxima. Este valor também é inferior ao registado por outros autores no milho (Maroco et al., 1999; Leakey et al., 2006), e provavelmente deve-se, como já foi referido, às relativamente baixas condições de PPFD (para plantas C4) permitidas pela câmara climatizada, durante o crescimento das plantas. No entanto, com a diminuição do THR começam a surgir diferenças. Com um THR de cerca de 70%, PB269 sofreu apenas uma redução de cerca de 50% da A líquida, enquanto que na variedade PB369, em THR de cerca de 80%, a A praticamente se anulou. Tal facto poderá Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 97 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico estar relacionado com problemas a diversos níveis, como no fecho estomático, na diminuição da eficiência do transporte de electrões ou nas enzimas do metabolismo do carbono (Flexas e Medrano, 2002). A utilização do modelo de Platt et al. (1980) (y = Ps × (1 – e (-αx/Ps)) × e (-βx/Ps)), permite adaptar uma curva de saturação luminosa que apresenta fotoinibição. Através deste modelo é possível determinar o parâmetro Ps (que representa o valor de fotossíntese potencial máximo caso não houvesse fotoinibição [β = 0]), o parâmetro α (eficiência fotossintética, que caracteriza as reacções fotoquímicas da fotossíntese e se mede a baixas PPFD), e o parâmetro β (que caracteriza o processo de fotoinibição). Para determinar a capacidade fotossintética (Amáx) utilizou-se a equação Amáx = Ps × (α/(α+β) × (β/(α+β)β/α). Apresentam-se na Tabela 5.2 os parâmetros estimados pela aplicação deste modelo na Fig. 5.4. Tabela 5.2 – Parâmetros da equação de Platt et al. (1980), resultantes da aplicação do modelo à Fig. 5.4, nas duas variedades portuguesas de milho. PB269 PB369 Stresse Stresse THR (%) THR (%) Parâmetros 97,8 ± 1,0 68,8 ± 14,9 37,0 ± 3,2 94,6 ± 2,1 83,5 ± 11,4 26,7 ± 11,2 Amáx (µmol CO2.m-2.s-1) 8,88 ± 2,12 4,38 ± 1,11 - 7,18 ± 2,85 - - α 0,024 ± 0,006 0,006 ± 0,000 - 0,019 ± 0,011 - - β 8,606 ± 2,202 0,469 ± 0,282 - 16,53 ± 1,34 - - (µmol CO2 / µmol fotões) (µmol CO2 / µmol fotões) Recuperação Recuperação THR (%) THR (%) Parâmetros 37,0 ± 3,2 76,1 ± 5,7 76,8 ± 3,2 26,7 ± 11,2 74,0 ± 16,0 73,9 ± 1,4 Amáx (µmol CO2.m-2.s-1) - 1,65 ± 0,71 2,00 ± 0,99 - 2,30 ± 0,14 3,73 ± 0,87 α - 0,002 ± 0,002 0,005 ± 0,004 - 0,003 ± 0,001 0,007 ± 0,003 β - 0,194 ± 0,594 0,567 ± 0,001 - 0,253 ± 0,245 0,421 ± 0,325 (µmol CO2 / µmol fotões) (µmol CO2 / µmol fotões) Observam-se valores de Amáx de cerca de 8,9 µmol CO2.m-2.s-1, para PB269, e de cerca de 7,2 µmol CO2.m-2.s-1, para PB369, perto da hidratação total. Perto do THR de 70%, a variedade PB269 ainda apresenta uma Amáx de cerca de 4,4 µmol CO2.m-2.s-1, enquanto a variedade PB369, embora não tenha sido possível aplicar o modelo de Platt et al. (1980) no ponto de THR de cerca de 85%, não apresenta uma Amáx superior a 2 µmol CO2.m-2.s-1, Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 98 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico estimado por análise directa dos pontos obtidos. Relativamente ao valor de β, observa-se que ocorre uma maior fotoinibição na variedade PB369, quando comparada com a variedade PB269, perto da hidratação total. Com o stresse hídrico, aparentemente o efeito da fotoinibição diminui na variedade PB269, mas tal deve-se ao facto de ocorrerem outros danos e/ou alterações metabólicas que provocam uma diminuição de Amáx. Relativamente aos valores de α, não são estatisticamente diferentes entre as duas variedades, quer durante o stresse quer durante a recuperação, embora em PB269, ao longo do stresse, se observe uma diminuição deste parâmetro, que se mantém inalterado durante a recuperação. Na variedade PB369, também existe uma redução de α, na recuperação, relativamente ao valor inicial. Estes dados indicam uma perda da eficiência fotossintética com a imposição do stresse hídrico. No caso da variedade PB369, na segunda medição (THR de cerca de 85%) durante o processo de indução do stresse e também nas últimas medições do período de stresse das duas variedades, o modelo de Platt et al. (1980) não pôde ser aplicado, por não ser estatisticamente significativo. Após a irrigação, devido à alteração das condições de crescimento decorrente da avaria da câmara de crescimento, ambas as variedades apresentaram um baixo nível de recuperação da A, verificando-se um aumento progressivo da fotoinibição nas duas variedades. Na Fig. 5.5 está representada a variação da condutância estomática (gs) com a PPFD, para diferentes THR. Na hidratação total, ambas as variedades apresentam um perfil semelhante, aumentando gs de 0,03 mol.m-2.s-1, em PPFD mais baixos, até cerca de 0,1 mol.m-2.s-1, para PPFD de 750 µmol fotões.m-2.s-1, a partir do qual se mantém relativamente constante na variedade PB369, mas continuando a aumentar até 0,13 mol.m-2.s-1, na variedade PB269. À medida que o THR diminui, verifica-se uma diminuição de gs para valores de 5060% dos iniciais. PB369, com THR de cerca de 85%, apresenta valores semelhantes aos de PB269 com THR de 70%. Desta forma, a diminuição de A verificada para PB369 na Fig. 5.4 não será (principalmente) consequência de limitações estomáticas, as quais estão descritas como as principais responsáveis pela diminuição de A para stresse hídrico baixo a moderado (Flexas e Medrano, 2002), uma vez que para o mesmo valor de THR, as duas variedades apresentam valores semelhantes de gs, mas valores diferentes de A, o que implica uma alteração ou da componente fotoquímica ou das enzimas do metabolismo fotossintético do carbono. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 99 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Para valores de THR de 25-35% (stresse intenso), os estomas estão praticamente fechados, apresentando as duas variedades uma gs praticamente nula. PB269 Stresse PB369 Stresse 0.2 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 0.2 0.1 0.0 0 250 500 750 1000 1250 0.1 0.0 1500 0 PPFD (µ µmol fotões.m-2.s-1) 250 500 750 1000 1250 1500 PPFD (µ µmol fotões.m-2.s-1) THR = 97,8±1,0 (;–): y = 0,02784 + 0,0001863x – 8,3951-8x2 THR = 94,6±2,0 (;–): y = 0,00526 + 2,4681e-5x + 1,7723e-8x2 (r2 = 0,99 ; p < 0,01) (r2 = 0,96 ; p < 0,01) THR = 68,8±14,9 (;- -): y = 0,008359 + 0,000119x – 5,6e-8x2 THR = 83,5±11,4 (;- -): y = 0,01576 + 5,4562e-5x – 2,07e-8x2 (r2 = 0,99 ; p < 0,01) (r2 = 0,99 ; p < 0,01) THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 0,01291 + 0,00001405x THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 0,01574 – 0,00001067x (r2 = 0,94 ; p < 0,01) (r2 = 0,92 ; p < 0,01) PB269 Recuperação PB369 Recuperação 0.2 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 0.2 0.1 0.0 0 250 500 750 1000 1250 0.0 1500 0 PPFD (µ µmol fotões.m-2.s-1) -5 0.1 250 500 750 1000 -2 1250 1500 -1 PPFD (µ µmol fotões.m .s ) -8 2 THR = 76,8±3,2 (;–): y = 0,06259 + 8,3068e x – 3,1858e x THR = 73,9±1,4 (;–): y = 0,07203 + 9,0429e-5x – 3,3782e-8x2 (r2 = 0,99 ; p < 0,01) (r2 = 0,98 ; p < 0,05) THR = 76,1±5,7 (;- -): y = 0,03836 + 7,08e-5x – 2,7426e-8x2 THR = 74,0±16,0 (;- -): y = 0,04177 + 7,085e-5x – 2,751e-8x2 (r2 = 0,99 ; p < 0,01) (r2 = 0,99 ; p < 0.01) THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 0,01291 + 0,00001405x THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 0,01574 – 0,00001067x (r2 = 0,94 ; p < 0,01) (r2 = 0,92 ; p < 0,01) Fig. 5.5 – Variação da condutância estomática (gs) em função da densidade de fluxo fotónico fotossintético (PPFD), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Após a irrigação, ambas as variedades vão apresentando valores progressivamente superiores aos observados no final do período de stresse, e quando atingem THR de cerca de 75%, a variedade PB269 apresenta valores ligeiramente inferiores aos iniciais e a variedade PB369 apresenta valores superiores aos iniciais, antes do início do stresse hídrico. Tal facto Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 100 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico vem reforçar a ideia de limitação não-estomática da fotossíntese visível no período de recuperação (Fig. 5.4). Verifica-se também que para PPFD mais baixos, durante a recuperação, as duas variedades apresentam valores de gs superiores aos observados quer na ausência de stresse, quer durante a imposição do stresse, o que poderá ser produto de um aumento da sensibilidade à luz, no controlo estomático (Aphalo e Sánchez, 1986). Na Fig. 5.6 está representada a variação da taxa transpiratória (E) em função da PPFD, para diferentes THR. Nas duas variedades observa-se uma baixa E para valores de PPFD reduzidos, aumentando até cerca de 1,5 mmol.m-2.s-1, em PB269, e 1,3 mmol.m-2.s-1, em PB369, para PPFD de cerca de 500 µmol fotões.m-2.s-1, a partir do qual E se mantém relativamente constante para a variedade PB369, mas continua a aumentar para PB269 até 1350 µmol fotões.m-2.s-1. Esta evolução de E vai acompanhando a evolução de gs (Fig. 5.5). Perto da hidratação total, verifica-se que, em PB269, a E é sempre superior à de PB369, para qualquer valor de PPFD. À medida que o THR diminui, também E vai sendo afectado nas duas variedades, apresentando uma variação relativa semelhante, mas é em PB369 que ocorre a maior diminuição em valores absolutos, uma vez que, a THR de cerca de 70%, a variedade PB269 apresenta uma E de 1 mmol.m-2.s-1, enquanto que, para um THR de cerca de 85%, a variedade PB369 apresenta uma E de 0,76 mmol.m-2.s-1. Para THR de 25-35%, ambas as variedades apresentaram valores de E bastante reduzidos, na ordem das 0,4-0,5 mmol.m-2.s-1, verificando-se que, no final do período de stresse, a maior redução, em termos absolutos, acabou por se verificar na variedade PB269. Durante a recuperação, observou-se em PB269 um regresso quase aos valores iniciais, embora para PPFD baixas os valores fossem superiores aos registados perto da hidratação total. Em PB369, após a irrigação no final do período de stresse, os valores de E foram sempre superiores aos iniciais, situando-se em cerca de 1,5 mmol.m-2.s-1. Tal aumento poderá estar relacionado com o aumento de gs verificado. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 101 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB269 Stresse PB369 Stresse 2 E (mmol.m-2.s-1) E (mmol.m-2.s-1) 2 1 0 0 250 500 750 1000 1250 1 0 1500 0 PPFD (µ µmol fotões.m-2.s-1) 250 500 750 1000 1250 -2 1500 -1 PPFD (µ µ mol fotões.m .s ) THR = 97,8±1,0 (;–): y = 0,6563 + 0,002164x – 1,0747e-6x2 (r2 = 0,91 ; p < 0,05) THR = 94,6±2,0 (;–): y = 0,4559 + 0,002095x – 1,1367e-6x2 (r2 = 0,86 ; p < 0,05) THR = 68,8±14,9 (;- -): y = 0,2384 + 0,001723x – 8,9623e-7x2 THR = 83,5±11,4 (;- -): y = 0,3369 + 0,000775x – 3,453e-7x2 (r2 = 0,96 ; p < 0,01) (r2 = 0,99 ; p < 0,01) THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 0,2795 + 0,000166x THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 0,3248 – 0,0001199x (r2 = 0,94 ; p < 0,01) (r2 = 0,85 ; p < 0,01) PB269 Recuperação PB369 Recuperação 2 E (mmol.m-2.s-1) E (mmol.m-2.s-1) 2 1 0 0 250 500 750 1000 1250 1 0 1500 0 PPFD (µ µmol fotões.m-2.s-1) 250 500 750 1000 1250 -2 1500 -1 PPFD (µ µ mol fotões.m .s ) -7 2 THR = 76,8±3,2 (;–): y = 1,151 + 0,0006509x – 3,3446e x THR = 73,9±1,4 (;–): y = 1,346 + 0,0007121x – 3,5882e-7x2 (r2 = 0,76 ; p < 0,05) (r2 = 0,79 ; p < 0,05) THR = 76,1±5,7 (;- -): y = 0,0393 + 0,0001844x + 1,3241e-7x2 THR = 74,0±16,0 (;- -): y = 0,8313 + 0,000713x – 3,453e-7x2 (r2 = 0,91 ; p < 0,05) (r2 = 0,95 ; p < 0,05) THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 0,2795 + 0,000166x THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 0,3248 – 0,0001199x (r2 = 0,94 ; p < 0,01) (r2 = 0,85 ; p < 0,01) Fig. 5.6 – Variação da taxa transpiratória (E) em função da densidade de fluxo fotónico fotossintético (PPFD), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Na Fig. 5.7 está representada a variação da concentração interna de CO2 (Ci) em função da PPFD, para diferentes THR. Perto da hidratação total, verifica-se que para PPFD baixas, Ci é superior à observada para PPFD mais elevadas, variando desde os 350 ppm até aos 200 ppm nas duas variedades. O mesmo sucede durante o período de stresse mas os valores de Ci são sempre superiores aos observados antes da sua imposição. Perto da hidratação total, as plantas apresentam valores Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 102 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico mais elevados de Ci, em PPFD baixas, uma vez que as enzimas do metabolismo fotossintético não estão activas, principalmente a RuBisCO activase, responsável pela activação da RuBisCO, e que é activada pela luz (Furbank et al., 1996). PB369 Stresse Ci (ppm) Ci (ppm) PB269 Stresse 900 800 700 600 500 400 300 200 100 0 0 250 500 750 1000 -2 1250 1500 900 800 700 600 500 400 300 200 100 0 0 250 -1 PPFD (µ µmol fotões.m .s ) 500 750 1000 -2 1250 1500 -1 PPFD (µ µmol fotões.m .s ) THR = 97,8±1,0 () THR = 94,6±2,0 () THR = 68,8±14,9 () THR = 83,5±11,4 (;- -): y = 490,1 – 0,3665x – 0,0001857x2 THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 630 – 0,1607x (r2 = 0,88 ; p < 0,01) (r2 = 0,96 ; p < 0,01) THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 615,8 – 0,09492x (r2 = 0,79 ; p < 0,01) PB369 Recuperação 900 800 700 600 500 400 300 200 100 0 Ci (ppm) Ci (ppm) PB269 Recuperação 0 250 500 750 1000 1250 1500 900 800 700 600 500 400 300 200 100 0 0 250 PPFD (µ µmol fotões.m-2.s-1) 500 750 1000 1250 1500 PPFD (µ µmol fotões.m-2.s-1) THR = 76,8±3,2 () THR = 73,9±1,4 () THR = 76,1±5,7 (;- -): y = 398,8 – 0,1918x + 0,8339x2 THR = 74,0±16,0 () (r2 = 0,83 ; p < 0,05) THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 615,8 – 0,09492x (r2 = 0,79 ; p < 0,01) THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 630 – 0,1607x (r2 = 0,88 ; p < 0,01) Fig. 5.7 – Variação da concentração interna de CO2 (Ci) em função da densidade de fluxo fotónico fotossintético (PPFD), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Para THR da ordem dos 25-35%, as folhas apresentam um aumento de Ci situado entre os 500-600 ppm. Este aumento de Ci poderá estar relacionado com a alteração da morfologia das células do mesófilo (Giles et al., 1974), provocando uma distorção que dificulta a difusão do CO2 inorgânico até aos locais em que é fixado pela PEPC (Marques da Silva e Arrabaça, 2004b) que, conjuntamente com um aumento da respiração (Lawlor e Cornic, 2002) Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 103 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico permitiria um aumento de CO2 junto da enzima, e manter alguma taxa fotossintética, como já foi observado em folhas de tomate, durante stresse hídrico (Haupt-Herting et al., 2001). Durante a recuperação, ambas as variedades apresentaram valores de Ci entre os 300400 ppm, sendo menores que os registados no final do período de stresse, embora superiores aos registados inicialmente, perto da hidratação total. Observa-se sempre uma diminuição de Ci com o aumento de PPFD, que reflecte alguma actividade da maquinaria enzimática fotossintética. Na Fig. 5.8 está representada a variação da eficiência do uso da água (WUE) em função da PPFD, para diferentes THR. PB369 Stresse 7 WUE (µ mol CO2 / mmol H2O) WUE (µ µ mol CO2 / mmol H2O) PB269 Stresse 6 5 4 3 2 1 0 0 250 500 750 1000 1250 7 6 5 4 3 2 1 0 1500 0 PPFD (µ µmol fotões.m-2.s-1) 250 500 750 1000 1250 1500 PPFD (µ µmol fotões.m-2.s-1) THR = 97,8±1,0 (;–): y = 0,8502 + 0,01171x – 6,6337e-6x2 THR = 94,6±2,0 (;–): y = 0,8502 + 0,01171x – 6,6337e-6x2 (r2 = 0,85 ; p < 0,05) (r2 = 0,85 ; p < 0,05) THR = 68,8±14,9 (;- -): y = 0,4463 + 0,007749x – 3,9046e-6x2 THR = 83,5±11,4 (;- -): y = – 2,399 + 0,006999x – 3,0880e-6x2 (r2 = 0,92 ; p < 0,05) (r2 = 0,99 ; p < 0,01) PB369 Recuperação WUE (µ mol CO2 / mmol H2O) WUE (µ µ mol CO2 / mmol H2O) PB269 Recuperação 7 6 5 4 3 2 1 7 6 5 4 3 2 1 0 0 0 250 500 750 1000 -2 1250 0 1500 PPFD (µ µmol fotões.m .s ) THR = 76,8±3,2 (;–): y = 0,1429 + 0,002956x – 1,5741e-6x2 (r2 = 0,78 ; p < 0,05) THR = 76,1±5,7 (;- -): y = – 0,07077 + 0,003030x – 1,5799e-6x2 (r2 = 0,92 ; p < 0,05) 250 500 750 1000 1250 1500 PPFD (µ µmol fotões.m-2.s-1) -1 THR = 73,9±1,4 (;–): y = – 0,1150 + 0,004914x – 2,4606e-6x2 (r2 = 0,86 ; p < 0,05) THR = 74,0±16,0 (;- -): y = 0,2034 + 0,002898x – 1,3803e-6x2 (r2 = 0,82 ; p < 0,05) Fig. 5.8 – Variação da eficiência do uso da água (WUE) em função da densidade de fluxo fotónico fotossintético (PPFD), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 104 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Perto da hidratação total, ambas as variedades apresentaram variação da WUE, sendo esta menor para PPFD mais baixas e aumentando até cerca de 6 µmol CO2 / mmol H2O para PPFD mais elevadas. Em PB269, a WUE foi afectada com a diminuição do THR (cerca de 70%), apresentando valores negativos para PPFD mais baixos mas recuperando para valores positivos a partir de PPFD de cerca de 200 µmol fotões.m-2.s-1. Na variedade PB369, WUE foi mais afectada, como seria esperado pela análise dos dados anteriores (Fig. 5.4 e Fig. 5.6). Para THR na ordem dos 85%, apresenta valores positivos de WUE apenas a partir de PPFD de 430 µmol fotões.m-2.s-1. Por vezes, são observados aumentos na WUE, com a imposição do stresse hídrico, dado que gs diminui mais rapidamente que o metabolismo fotossintético do carbono, ocorrendo uma maior fixação de CO2 por cada mole de água transpirada (Chaves e Oliveira, 2004). No entanto, não foi o que aqui sucedeu. Os valores mais baixos de THR (stresse intenso) não estão representados na Fig. 5.8 porque apresentam valores de WUE negativos, devido a A apresentar também valores negativos (Fig. 5.4). Durante a recuperação, a WUE recuperou apenas muito parcialmente nas duas variedades, apresentando valores reduzidos para PPFD baixas e não ultrapassando os 2 µmol CO2 / mmol H2O para PPFD mais elevados, devido à conjunção de uma redução de A e de um aumento de E, provocados principalmente devido ao aumento da temperatura e diminuição da humidade relativa, resultante da avaria da câmara de crescimento. 5.3.2.2. Curvas A/CO2a Na Fig. 5.9 está representada a variação de A em função da concentração externa de CO2 (CO2a), para diferentes THR. Na variedade PB269, observaram-se inicialmente valores de trocas gasosas inferiores aos registados posteriormente no ensaio. Contudo, na segunda medição, os valores de THR ainda se encontravam perto da hidratação total (cerca de 90%), e poderão ser os mais próximos dos esperados. Com uma CO2a entre os 100 ppm e os 370 ppm verifica-se um grande aumento de A e, para valores superiores de CO2a, o aumento de A é menor, como acontece com as plantas C4 que saturam com o aumento de CO2a. Tal como já se havia verificado no ensaio das curvas A/PPFD, observa-se um menor valor de A para a variedade PB369. Esta apresenta um valor semelhante de A registado para PB269 mas para THR diferentes (cerca de 70% em PB269 e cerca de 85% em PB369). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 105 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB369 Stresse 20 15 15 A (µ mol.m-2.s-1) A (µ µ mol.m-2.s-1) PB269 Stresse 20 10 5 0 250 500 750 1000 1250 1500 5 0 250 CO2a (ppm) -5 10 500 750 1000 1250 1500 CO2a (ppm) -5 THR = 94,8±2,1 (;–): y = 2,211 + 0,002536x (r2 = 0,84 ; p <,0,01) THR = 92,3±3,2 (;–): y = 2,287 + 0,01191x – 4,2717e-6x2 THR = 89,7±8,4 (;- -): y = 4,628 + 0,01587x – 6,7091e-6x2 (r2 = 0,99 ; p <,0,05) (r2 = 0,96 ; p <,0,05) THR = 94,2±0,6 (;- -): y = 3,071 + 0,01003x – 3,5170e-6x2 THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 2,29 + 0,002004x (r2 = 0,94 ; p <,0,01) (r2 = 0,96 ; p <,0,05) THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 1,666 + 0,002033x (r2 = 0,92 ; p <,0,01) PB369 Recuperação 20 20 15 15 A (µ mol.m-2.s-1) A (µ µ mol.m-2.s-1) PB269 Recuperação 10 5 0 250 500 750 1000 1250 5 0 1500 250 CO2a (ppm) -5 10 -5 THR = 85,5±3,4 (;–): y = 3,338 + 0,004317x – 4,4179e-7x2 2 500 750 1000 1250 1500 CO2a (ppm) THR = 94,6±2,6 (;–): y = 0,0399 + 0,00229x (r2 = 0,77 ; p <,0,01) THR = 92,0±4,8 (;- -): y = 0,772 + 0,00308x (r2 = 0,89 ; p <,0,01) (r = 0,95 ; p <,0,05) -6 2 THR = 85,1±4,6 (;- -): y = 3,775 + 0,008619x – 2,4388e x THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 1,666 + 0,002033x (r2 = 0,92 ; p <,0,01) 2 (r = 0,97 ; p <,0,05) THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 2,29 + 0,002004x (r2 = 0,94 ; p <,0,01) Fig. 5.9 – Variação da taxa fotossintética líquida (A) em função da concentração externa de CO2 (CO2a), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Verifica-se que a variedade PB269 recupera melhor aqui que no ensaio da PPFD (indicando alguma susceptibilidade à temperatura), embora, tal como na fase I, não consiga recuperar o THR até aos valores iniciais. Por outro lado, a variedade PB369 não recupera após irrigação. Dado que o aumento de CO2 não restaura A para Amáx, pode afirmar-se que existe um predomínio da limitação não-estomática (Kanechi et al., 1996; Pankovic et al., 1999; Lawlor, 2002). Associando os dados de A aos valores de Ci, foi possível calcular as limitações estomáticas e não-estomáticas (Tab. 5.3) (Jones, 1985; Geber e Dawson, 1997; Tezara et al., 2003). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 106 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Tab. 5.3 – Limitações não-estomáticas e estomáticas da fotossíntese, a partir das curvas A/Ci. THR (%) Dia PB269 PB369 Limitação não- Limitação estomática estomática relativa (%) relativa (%) Limitação total Limitação não- Limitação (%) estomática (%) estomática (%) PB269 PB369 PB269 PB369 PB269 PB369 PB269 PB369 0 8 19 89 0 11 100 PB269 PB369 (*) 0 95 92 74 19 66 3 90 94 13 18 0 4 13 14 0 22 100 78 6 69 85 74 63 68 57 6 6 92 91 8 9 9 85 92 40 76 26 62 14 14 66 81 34 19 12 86 95 55 94 48 94 7 0 87 100 13 0 (*) Foi considerado o dia 3 como os valores controlo, ou seja, ainda perto da hidratação total. A variedade PB269 apresentou valores mais baixos de fotossíntese no primeiro dia de ensaio, mas no terceiro dia apresentou valores próximos dos esperados para a hidratação total, que se consideraram mais indicados para funcionarem como controlo. Verifica-se que as limitações não-estomáticas se manifestam de forma mais intensa na variedade PB369, uma vez que possuem uma limitação não-estomática relativa de cerca de 90% a um THR de 85%, enquanto que o mesmo valor foi observado na variedade PB269 num THR de cerca de 70%. A análise da Tab. 5.3 permite constatar que as limitações estomáticas da fotossíntese têm uma maior importância perto da hidratação total (dia 3). À medida que o stresse hídrico vai aumentando e o THR diminuindo, as limitações não-estomáticas começam a aumentar, e as limitações estomáticas deixam de ter a mesma relevância (dia 6) (Marques da Silva, 1999; Flexas e Medrano, 2002; Tezara et al., 2003). Durante a fase de recuperação também se registam diferenças, com a variedade PB269 a recuperar de forma mais marcante que PB369, a qual continua a sofrer uma diminuição de A, mesmo após irrigação. Relativamente às limitações da fotossíntese, verifica-se que em ambas as variedades as limitações nãoestomáticas são as responsáveis pela baixa ou praticamente inexistente recuperação da fotossíntese. Na Fig. 5.10 está representada a variação de gs em função da CO2a, para diferentes THR. Verifica-se que para CO2a mais baixos, ambas as variedades apresentam valores de gs superiores aos registados para CO2a mais elevados, o que é de esperar como resposta dos estomas ao aumento de CO2, de modo a permitir manter a actividade fotossintética (Lopez et al., 1987; Maroco et al., 1999). No entanto, na variedade PB269, há uma diminuição de gs de cerca de 0,2 mol.m-2.s-1 para cerca de 0,05 mol.m-2.s-1, enquanto que em PB369 a mesma Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 107 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico diminuição acontece de 0,1 mol.m-2.s-1 para 0,05 mol.m-2.s-1, indicando que esta última variedade apresenta estomas com uma sensibilidade inferior ao CO2 ambiente. Por outro lado, a maior sensibilidade estomática da variedade PB269 poderia explicar a razão do ajustamento da gs e da A com o Ψw e o THR (Fig. 4.6 e 4.7), como foi verificado numa variedade tolerante de Coffea canephora (Pinheiro et al., 2005). PB269 Stresse PB369 Stresse 0.3 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 0.3 0.2 0.1 0.0 0 250 500 750 1000 1250 0.2 0.1 0.0 1500 0 250 CO2a (ppm) 500 750 1000 1250 1500 CO2a (ppm) THR = 94,8±2,1 (;–): y = 0,1459 – 0,002485x + 1,2862e-7x2 THR = 92,.3±3,.2 (;–): y = 0,1299 – 0,0001555x + 6,6621e-8x2 (r2 = 0,90 ; p < 0,05) (r2 = 0,97 ; p < 0,05) THR = 89,7±8,4 (;- -): y = 0,2359 – 0,000345x + 1,5397e-7x2 THR = 94,2±0,6 (;- -): y = 0,1445 – 0,000194x + 8,6688e-8x2 (r2 = 0,96 ; p < 0,05) (r2 = 0,97 ; p < 0,05) THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 0,09507 – 0,00127x + 5,6102e-8x2 THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 0,06748 – 7,8896e-5x + 3,4431e-8x2 (r2 = 0,96 ; p < 0,05) (r2 = 0,96 ; p < 0,05) PB269 Recuperação PB369 Recuperação 0.3 gs (mol.m-2.s-1) gs (mol.m-2.s-1) 0.3 0.2 0.1 0.0 0 250 500 750 1000 1250 0.2 0.1 0.0 1500 0 250 CO2a (ppm) 500 750 1000 1250 1500 CO2a (ppm) THR = 85,5±3,4 (;–): y = 0,1785 – 0,000259x + 1,1931e-7x2 THR = 94,6±2,6 (;–): y = 0,08945 – 5,7028e-5x + 2,6638e-8x2 (r2 = 0,96 ; p < 0,05) (r2 = 0,93 ; p < 0,05) THR = 85,1±4,6 (;- -): y = 0,1699 – 0,00024x + 1,0271e-7x2 THR = 92,0±4,8 (;- -): y = 0,07465 – 7,749e-5x + 3,4997e-8x2 (r2 = 0,96 ; p < 0,05) (r2 = 0,95 ; p < 0,05) THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 0,09507 – 0,00127x + 5,6102e-8x2 THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 0,06748 – 7,8896e-5x + 3,4431e-8x2 (r2 = 0,96 ; p < 0,05) (r2 = 0,96 ; p < 0,05) Fig. 5.10 – Variação da condutância estomática (gs) em função da concentração externa de CO2 (CO2a), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. À medida que o THR diminui, em PB269, os valores de gs para CO2a baixos vão Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 108 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico diminuindo, embora para CO2a mais elevados se mantenham constantes. Em PB369, não existem alterações significativas. Comparando com a Fig. 5.9, verifica-se que para valores idênticos de gs, a A em PB369 apresenta uma maior redução. Durante a recuperação, PB269 recupera parcialmente os valores iniciais de gs, não ocorrendo a recuperação total provavelmente devido a não recuperar facilmente o THR, facto já observado na fase I. Por outro lado, na variedade PB369 há uma certa tendência para manter os valores semelhantes aos observados no final do período de stresse. PB269 Stresse PB369 Stresse 3 E (mmol.m-2.s-1) E (mmol.m-2.s-1) 3 2 1 0 0 250 500 750 1000 1250 2 1 0 1500 0 250 CO2a (ppm) 500 750 1000 1250 1500 CO2a (ppm) THR = 94,8±2,1 (;–): y = 1,929 – 0,002897x + 1,4784e-6x2 THR = 92,3±3,2 (;–): y = 1,81 – 0,0018x + 7,3107e-7x2 (r2 = 0,93 ; p < 0,05) (r2 = 0,99 ; p < 0,05) THR = 89,7±8,4 (;- -): y = 2,513 – 0,002885x + 1,1967e-6x2 THR = 94,2±0,6 (;- -): y = 1,903 – 0,00212x + 9,0264e-7x2 (r2 = 0,98 ; p < 0,05) (r2 = 0,98 ; p < 0,05) THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 1,318 – 0,001463x + 6,1415e-7x2 THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 1,134 – 0,001193x + 5,0385e-7x2 (r2 = 0,97 ; p < 0,05) (r2 = 0,97 ; p < 0,05) PB269 Recuperação PB369 Recuperação 3 E (mmol.m-2.s-1) E (mmol.m-2.s-1) 3 2 1 0 0 250 500 750 1000 1250 2 1 0 1500 0 250 CO2a (ppm) 500 750 1000 1250 1500 CO2a (ppm) THR = 85,5±3,4 (;–): y = 2,269 – 0,00241x + 1,0674e-6x2 THR = 94,6±2,6 (;–): y = 1,393 – 0,0008393x + 3,9304e-7x2 (r2 = 0,98 ; p < 0,05) (r2 = 0,95 ; p < 0,05) THR = 85,1±4,6 (;- -): y = 2,426 – 0,002936x + 1,2902e-6x2 THR = 92,0±4,8 (;- -): y = 1,337 – 0,001268x + 5,5299e-7x2 (r2 = 0,98 ; p < 0,05) (r2 = 0,96 ; p < 0,05) THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 1,318 – 0,001463x + 6,1415e-7x2 THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 1,134 – 0,001193x + 5,0385e-7x2 (r2 = 0,96 ; p < 0,05) (r2 = 0,97 ; p < 0,05) Fig. 5.11 – Variação da taxa transpiratória (E) em função da concentração externa de CO2 (CO2a), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 109 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Na Fig. 5.11 está representada a variação de E em função da CO2a, para diferentes THR. Perto da hidratação total, ambas as variedades apresentam valores de E superiores para CO2a inferiores, diminuindo à medida que CO2a aumenta. À medida que o THR diminui verifica-se uma diminuição do valor máximo de E. Na recuperação, o padrão observado é semelhante, acompanhando a evolução de gs (Fig. 5.10). Na Fig. 5.12 está representada a variação de Ci em função da CO2a, para diferentes THR. PB369 Stresse 1250 1250 1000 1000 Ci (ppm) Ci (ppm) PB269 Stresse 750 500 250 750 500 250 0 0 250 500 750 1000 1250 0 1500 0 250 CO2a (ppm) 500 750 1000 1250 1500 CO2a (ppm) THR = 94,8±2,1 (;–): y = – 24,48 + 0,8156x (r2 = 0,99 ; p < 0,01) THR = 92,3±3,2 (;–): y = – 32 + 0,6621x (r2 = 0,99 ; p < 0,01) THR = 89,7±8,4 (;- -): y = – 37,08 + 0,6373x (r2 = 0,99 ; p < 0,01) THR = 94,2±0,6 (;- -): y = – 36,44 + 0,6654x (r2 = 0,99 ; p < 0,01) THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = – 15,13 + 0,7686x (r2 = 0,99 ; p < 0,01) THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = – 17,42 + 0,7444x (r2 = 0,99 ; p < 0,01) PB369 Recuperação 1250 1000 1000 Ci (ppm) Ci (ppm) PB269 Recuperação 1250 750 500 250 750 500 250 0 0 250 500 750 1000 1250 0 1500 0 CO2a (ppm) 250 500 750 1000 1250 1500 CO2a (ppm) THR = 85,5±3,4 (;–): y = – 31,21 + 0,7868x (r2 = 0,99 ; p < 0,01) THR = 94,6±2,6 (;–): y = 16,14 + 0,8884x (r2 = 0,99 ; p < 0,01) THR = 85,1±4,6 (;- -): y = – 21,04 + 0,6306x (r2 = 0,99 ; p < 0,01) THR = 92,0±4,8 (;- -): y = 4,821 + 0,7799x (r2 = 0,99 ; p < 0,01) THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = – 15,13 + 0,7686x (r2 = 0,99 ; p < 0,01) THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = – 17,42 + 0,7444x (r2 = 0,99 ; p < 0,01) Fig. 5.12 – Variação da concentração interna de CO2 (Ci) em função da concentração externa de CO2 (CO2a), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Tal como seria esperado, observa-se um aumento linear de Ci com o aumento de CO2a. Com o aumento de A (Fig. 5.9), Ci apresenta uma ligeira diminuição com o aumento de CO2a, Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 110 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico na hidratação total. Com a diminuição do THR, Ci vai apresentando valores superiores aos iniciais, com o aumento de CO2a, indicando uma diminuição da capacidade de fixação do carbono pelo processo fotossintético (Medrano et al., 2002) ou um aumento da respiração (Lawlor e Cornic, 2002). Durante a recuperação, PB269 sofre uma evolução para valores perto dos iniciais, enquanto que em PB369 os valores de Ci continuam a aumentar. Na Fig. 5.13 está representada a variação de WUE em função da CO2a, para diferentes THR. PB369 Stresse 20 WUE (µ µ mol CO2 / mmol H2O) WUE (µ µ mol CO2 / mmol H2O) PB269 Stresse 15 10 5 0 0 250 500 750 1000 1250 20 15 10 5 0 1500 0 250 CO2a (ppm) 500 750 1000 1250 1500 CO2a (ppm) THR = 94,8±2,1 (;–): y = 0,957 + 0,005482x (r2 = 0,92 ; p < 0,01) THR = 92,3±3,2 (;–): y = 0,34 + 0,01477x – 2,9241e-6x2 THR = 89,7±8,4 (;- -): y = 0,5908 + 0,01752x – 3,9491e-6x2 (r2 = 0,99 ; p < 0,05) (r2 = 0,99 ; p < 0,01) THR = 94,2±0,6 (;- -): y = 0,6951 + 0,01456x – 2,9014e-6x2 THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 0,7389 + 0,006567x (r2 = 0,99 ; p < 0,05) (r2 = 0,99 ; p < 0,01) THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 0,6329 + 0,006695x (r2 = 0,99 ; p < 0,01) PB369 Recuperação 20 WUE (µ µ mol CO2 / mmol H2O) WUE (µ µ mol CO2 / mmol H2O) PB269 Recuperação 15 10 5 0 0 250 500 750 1000 1250 20 15 10 5 0 1500 0 250 CO2a (ppm) 500 750 1000 1250 1500 CO2a (ppm) THR = 85,5±3,4 (;–): y = 1,291 + 0,00573x (r2 = 0,99 ; p < 0,01) THR = 94,6±2,6 (;–): y = 0,8434 – 0,001895x + 3,2128e-6x2 THR = 85,1±4,6 (;- -): y = 1,081 + 0,009645x (r2 = 0,97 ; p < 0,05) (r2 = 0,99 ; p < 0,01) THR = 92,0±4,8 (;- -): y = – 0,1916 + 0,006058x THR = 68,5±8,5 (∆;⋅⋅⋅⋅): y = 0,7389 + 0,006567x (r2 = 0,93 ; p < 0,01) (r2 = 0,99 ; p < 0,01) THR = 85,2±7,5 (∆;⋅⋅⋅⋅): y = 0,6329 + 0,006695x (r2 = 0,99 ; p < 0,01) Fig. 5.13 – Variação da eficiência do uso da água (WUE) em função da concentração externa de CO2 (CO2a), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 111 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Perto da hidratação total verifica-se um aumento de WUE com o aumento de CO2a em ambas as variedades. A evolução crescente deste parâmetro é semelhante em todos os THR registados. À medida que o THR decresce, os valores de WUE para valores de CO2a superiores vão sendo menores. Durante a recuperação, com o consequente aumento de THR, PB269 apresenta uma recuperação de WUE, enquanto em PB369 continua a sofrer uma diminuição. Os dados de trocas gasosas, no seu conjunto, permitem verificar uma maior tolerância ao stresse hídrico na variedade PB269, relativamente a PB369, como já havia sido verificado na fase I deste trabalho. O padrão de “tudo-ou-nada”, na diminuição geral dos parâmetros (exceptuando Ci), parece verificar-se novamente para a variedade PB369, enquanto em PB269 vai ocorrendo uma diminuição gradual dos mesmos. 5.3.2.3. Libertação de O2 No que respeita à libertação de O2, PB369 apresenta uma diminuição da taxa, em condições de stresse, mais acentuada que a da variedade PB269 (Fig. 5.14), embora isso se possa dever ao facto de, na primeira variedade, se terem obtido poucas medições a THR baixos. Os dados estão de acordo com os obtidos nas curvas de A/CO2a (Fig. 5.9). PB269 PB369 25 Libertação de O2 20 15 10 5 0 60 70 80 90 (µ mol O2.m-2.s-1) Libertação de O2 (µ mol O2.m-2.s-1) 25 20 15 10 5 0 100 60 THR (%) 70 80 90 100 THR (%) Stresse (∆; - -): y = – 12,95 + 0,3044x (r2 = 0,55 ; p < 0,01) Stresse (∆) Recuperação (; –): y = – 7,564 + 0,2185x (r2 = 0,56 ; p < 0,01) Recuperação () Fig. 5.14 – Variação da libertação de O2 em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Pela análise da recta da variedade PB269, observou-se uma anulação da libertação de O2 a THR de 42%. Em PB369, não foi possível obter valores mais baixos de THR mas a 70% já apresenta um ponto com forte redução de libertação de O2. Também se observa que, mais uma vez, PB369 recupera o THR para valores próximos da hidratação total, após irrigação, Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 112 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico embora PB269 não (caixas da Fig. 5.14). No entanto, há uma retoma significativa da taxa de libertação de O2 nesta última, no período de recuperação. Em PB369, durante a recuperação, o complexo de formação de O2 não parece ter sido muito afectado, até THR de 85%. Através da normalização dos valores obtidos na libertação de O2 (Fig. 5.14) e dos dados obtidos nos ensaios da curva de A/CO2a, procedeu-se à construção da Fig. 5.15, admitindo os valores iniciais obtidos na ausência de stresse (exceptuando os de PB269, em que os valores utilizados foram os do segundo dia de medições, mas no qual as plantas ainda se encontravam perto da hidratação total [ver Fig. 5.9]), como o máximo de actividade fotossintética. Este gráfico permite observar a diferença entre a limitação não-estomática (a CO2 saturante) e a limitação estomática (a CO2 atmosférico) (Ghannoum et al., 2003; Marques da Silva e Arrabaça, 2004a). PB269 PB369 100 Trocas gasosas (%) Trocas gasosas (%) 100 75 50 25 0 60 70 80 90 75 50 25 0 60 100 70 THR (%) 80 90 100 THR (%) Fig. 5.15 – Normalização da variação da taxa fotossintética em condições atmosféricas normais de CO2 (370 ppm) (- -) e medida sob 7% de CO2 (libertação de O2) (–) em função do teor hídrico relativo (THR). Os parâmetros exprimem-se em % do valor máximo. A partir da Fig. 5.15, foi possível determinar as limitações estomáticas e nãoestomáticas da fotossíntese, apresentadas na Tab. 5.4. Tab. 5.4 – Limitações não-estomáticas e estomáticas da fotossíntese. THR (%) Limitação total Limitação não- Limitação Limitação relativa Limitação relativa (%) estomática (%) estomática (%) não-estomática (%) estomática (%) PB269 PB369 PB269 PB369 PB269 PB369 PB269 PB369 PB269 PB369 100 0 0 0 0 0 0 0 0 0 0 95 13 14 9 12 4 2 69 86 31 14 90 26 28 17 24 8 4 65 86 35 14 85 38 42 26 35 12 6 68 83 32 17 70 77 83 52 70 25 13 68 84 32 16 Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 113 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Verifica-se que na variedade PB269 as limitações não-estomáticas apresentam um valor relativo inferior ao registado em PB369, com a diminuição do THR. No entanto, as limitações estomáticas apresentam uma maior componente relativa na variedade PB269, relativamente a PB369. Porém, as limitações não-estomáticas apresentam um valor relativo superior ao observado para as limitações estomáticas, em ambas as variedades, para stresse baixo a moderado. Tal facto, poderá estar relacionado com a concentração de CO2 utilizada (7%) não ser saturante ou inibir a fotossíntese destas plantas C4, o que poderá provocar uma sobrevalorização da componente do mesófilo (Marques da Silva, 1999). No entanto, novos estudos têm revelado que a componente não-estomática pode ser uma componente mais importante do que aquela que tem sido descrita em stresse moderado, principalmente devido à diminuição da síntese de ATP, observada para o girassol (Tezara et al., 1999; Medrano et al., 2002). Este facto permite verificar que, embora em CO2 ambientais as duas variedades não apresentem diferenças significativas de A (Fig. 5.9), na variedade PB269 estas limitações estão relacionadas mais directamente com o fecho estomático (Fig. 5.10) do que com modificações do metabolismo fotossintético do carbono (Flexas e Medrano, 2002). Em PB369, as limitações de A estarão mais directamente ligadas a alterações do ciclo do metabolismo do carbono com a variação do THR, observações concordantes com os valores da Tab. 5.3. 5.3.3. Fluorescência Modulada Modulada da Clorofila a Os dados da fluorescência da clorofila a fornecem informações importantes sobre a competência fotossintética das espécies vegetais. A razão entre a fluorescência variável máxima (Fv) e a fluorescência máxima (Fm) indica a eficiência da conversão de energia pelos centros de reacção do PSII abertos, na ausência de fenómenos de amortecimento não-fotoquímico. Na Fig. 5.16 observa-se a variação deste parâmetro com o THR, nas duas variedades seleccionadas. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 114 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB369 PB269 0.8 Fv/Fm (unidades relativas) Fv/Fm (unidades relativas) 0.8 0.7 0.6 0.5 0.4 0.3 0.7 0.6 0.5 0.4 0.3 0.2 0.2 25 50 75 25 100 50 75 100 THR (%) THR (%) Stresse (∆) Stresse (∆; - -): y = 0,06583 + 0,01108x – 4,8227e-5x2 Recuperação () (r2 = 0,96 ; p < 0,05) Recuperação (; –): y = – 0,0276 + 0,0163x – 9,203e-5x2 (r2 = 0,95 ; p < 0,05) Fig. 5.16 – Variação do rendimento quântico máximo do PSII (Fv/Fm) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Pode observar-se que o valor se encontra próximo das 0,7 unidades relativas na hidratação máxima em ambas as variedades, valor inferior ao de 0,8 encontrado por outros autores (Maxwell e Johnson, 2000; Marques da Silva e Arrabaça, 2004b) e ao encontrado na primeira fase deste trabalho (Fig. 4.21). Tal facto, poderá reflectir algumas diferenças observadas nesta fase, nomeadamente na diminuição de A, em relação à primeira fase do trabalho. Provavelmente, essas diferenças estarão relacionadas com um maior crescimento das plantas que poderá ter ocorrido, em relação à fase I, atingindo a maturidade mais cedo que a inicialmente determinada. No entanto, podem verificar-se diferenças nas duas variedades. Na variedade PB269, não se verificam variações significativas deste parâmetro em THR entre os 50 e os 100%, sofrendo uma queda abrupta para 0,4-0,5 unidades relativas para um THR de cerca de 35% (caixas da Fig. 5.16), também observado por Marques da Silva (1999) em Setaria sphacelata. Na variedade PB369, o parâmetro analisado diminui continuamente até atingir um valor de 0,2-0,3 unidades relativas para um THR de cerca de 25%. Tal facto é um dado importante para distinguir as duas variedades, no que respeita à susceptibilidade do PSII ao stresse hídrico. Durante a recuperação, embora realizada em condições de temperatura mais elevadas (38ºC) que durante o período de stresse, ambas as variedades regressaram aos valores iniciais. Na Fig. 5.17 observa-se a variação do rendimento quântico efectivo do PSII (ΦPSII) em função da PPFD. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 115 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB369 Stresse PB269 Stresse 0.7 ΦPSII (unidades relativas) ΦPSII (unidades relativas) 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0.6 0.5 0.4 0.3 0.2 0.1 0.0 0.0 0 250 500 750 1000 -2 1250 0 1500 250 750 1000 1250 1500 -1 PPFD (µ µmol fotões.m .s ) PPFD (µ µmol fotões.m .s ) THR = 95,6±1,5 (;–): y = 0,727 – 0,00114x + 4,4544e-7x2 THR = 93,9±2,9 (;–): y = 0,7104 – 0,001137x + 4,6742e-7x2 (r2 = 0,99 ; p < 0,01) 2 (r = 0,99 ; p < 0,01) -7 2 THR = 68,8±14,9 (;- -): y = 0,4813 – 0,0007776x + 3,1195e x THR = 83,5±11,4 (;- -): y = 0,4332 – 0,00085x + 3,985e-7x2 (r = 0,95 ; p < 0,01) (r2 = 0,96 ; p < 0,05) THR = 37,0±3,2 (∆) THR = 26,7±11,2 (∆) 2 PB269 Recuperação PB369 Recuperação 0.7 0.7 0.6 ΦPSII (unidades relativas) Φ PSII (unidades relativas) 500 -2 -1 0.5 0.4 0.3 0.2 0.1 0.0 0 250 500 750 1000 1250 0.6 0.5 0.4 0.3 0.2 0.1 0.0 1500 0 250 PPFD (µ µmol fotões.m-2.s-1) 500 750 1000 -2 1250 1500 -1 PPFD (µ µ mol fotões.m .s ) -7 2 THR = 76,8±3,2 (;–): y = 0,4794 – 0,001082x + 5,4715e x THR = 73,9±1,4 (;–): y = 0,4997 – 0,0005759x – 1,4409e-7x2 (r2 = 0,99 ; p < 0,01) (r2 = 0,90 ; p < 0,01) THR = 76,1±5,7 (;- -): y = 0,5653 – 0,001119x + 5,2405e-7x2 THR = 74,0±16,0 (;- -): y = 0,426 – 0,000692x – 2,7933e-7x2 (r2 = 0,99 ; p < 0,01) (r2 = 0,99 ; p < 0,01) THR = 37,0±3,2 (∆) THR = 26,7±11,2 (∆) Fig. 5.17 – Variação do rendimento quântico efectivo do PSII (ΦPSII) em função da densidade de fluxo fotónico fotossintético (PPFD), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Assim perto da hidratação total e no início do stresse, ambas as variedades apresentam valores perto das 0,65 unidades relativas, para PPFD mais reduzidas, valor observado também noutras espécies como Cucumis sativus, Ternstroemia gymnantheria e Acacia auriculiformis (Tsuyama et al., 2003). Este valor diminui com o aumento da PPFD, o que indica o fecho progressivo dos centros de reacção do PSII, com o aumento da PPFD e também a existência de processos de fotoinibição da fotossíntese. Com a diminuição do THR, a proporção de centros de reacção do PSII abertos vai diminuindo, para as mesmas PPFD, em ambas as variedades. Esta diminuição é, contudo, Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 116 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico mais marcante em PB369, onde, para qualquer dos níveis de stresse considerados, ΦPSII diminui mais acentuadamente e se anula para PPFD mais baixas. Estes dados indicam uma progressiva perda de capacidade fotossintética com a diminuição do THR, possivelmente por dano dos centros de reacção do PSII (Matsubara e Chow, 2004) e associada a efeitos negativos nas enzimas do metabolismo fotossintético (Parry et al., 2002), com PB369 a revelar, mais uma vez, uma maior susceptibilidade. Durante a recuperação, as variedades demonstraram um progressivo regresso a valores perto dos iniciais. Contudo, o perfil inicial não foi atingido provavelmente porque os valores de THR não foram recuperados. Na Fig. 5.18 está representada a variação do amortecimento fotoquímico (qP) em função da PPFD, em diferentes THR. Verifica-se também uma diferença de comportamentos nas duas variedades à medida que o stresse hídrico se intensifica. Perto da hidratação total, até PPFD de cerca de 500 µmol fotões.m-2.s-1, qP praticamente não sofre variações (ver caixas da Fig. 5.18). Em PB369, ocorre uma diminuição de qP com o aumento da PPFD a THR de 85%. No entanto, em PB269, para THR de cerca de 70%, qP apresenta valores semelhantes aos das plantas perto da hidratação total, indicando que a cadeia fotoquímica apresenta um maior grau de tolerância ao stresse do que a variedade PB369. Com a intensificação do stresse hídrico, ambas as variedades apresentam um anulamento de qP, embora para valores de PPFD diferentes. Este anulamento deve-se ao facto de a planta sofrer danos nos centros de reacção do PSII (Matsubara e Chow, 2004) e também porque, à medida que o stresse hídrico progride, as actividades enzimáticas vão sendo afectadas negativamente, factores que contribuem para a diminuição de qP (Medrano et al., 2002; Parry et al., 2002). Na recuperação (efectuada a 38ºC), o parâmetro qP recupera totalmente nas duas variedades, embora o THR seja mais reduzido que o observado inicialmente, que poderá resultar de alguma aclimatação das plantas (Sharp et al., 2004). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 117 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB369 Stresse PB269 Stresse 1.0 qP (unidades relativas) qP (unidades relativas) 1.0 0.8 0.6 0.4 0.2 0.0 250 500 750 1000 -2 1250 0.8 0.6 0.4 0.2 0.0 250 1500 PPFD (µ µmol fotões.m .s ) THR = 93,9±2,9 () THR = 95,6±1,5 () THR = 68,8±14,9 () THR = 83,5±11,4 () THR = 37,0±3,2 (∆) THR = 26,7±11,2 (∆) PB269 Recuperação 750 1000 1250 1500 PB369 Recuperação 1.0 1.00 qP (unidades relativas) qP (unidades relativas) 500 PPFD (µ µmol fotões.m-2.s-1) -1 0.8 0.6 0.4 0.2 0.0 250 500 750 1000 1250 0.75 0.50 0.25 0.00 250 1500 PPFD (µ µmol fotões.m-2.s-1) 500 750 1000 1250 1500 PPFD (µ µmol fotões.m-2.s-1) THR = 76,8±3,2 () THR = 73,9±1,4 () THR = 76,1±5,7 () THR = 74,0±16,0 () THR = 37,0±3,2 (∆) THR = 26,7±11,2 (∆) Fig. 5.18 – Variação do coeficiente de amortecimento fotoquímico (qP) em função da densidade de fluxo fotónico fotossintético (PPFD), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Na Fig. 5.19 está representada a variação do amortecimento não-fotoquímico (qN) em função de PPFD, para diferentes THR. Verifica-se que para valores de THR perto da hidratação total, qN, que traduz a fracção de energia dissipada não-fotoquimicamente, se encontra perto da capacidade máxima para PPFD de 1300 µmol fotões.m-2.s-1, valor que não é considerado saturante para plantas C4 (Marques da Silva, 1999; Maroco et al., 1999). Tal facto poderá dever-se às condições em que as plantas foram cultivadas (PPFD de aproximadamente 500 µmol fotões.m-2.s-1). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 118 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB369 Stresse 1.00 qN (unidades relativas) qN (unidades relativas) PB269 Stresse 0.75 0.50 0.25 0.00 0 250 500 750 1000 1250 1.00 0.75 0.50 0.25 0.00 1500 0 250 PPFD (µ µmol fotões.m-2.s-1) 500 750 1000 -2 1250 1500 -1 PPFD (µ µmol fotões.m .s ) -7 2 THR = 93,9±2,9 (;–): y = 0,09664 + 0,001756x – 8,2417e x THR = 95,6±1,5 (;–): y = 0,03413 + 0,001866x – 8,6279e-7x2 (r2 = 0,99 ; p < 0,01) (r2 = 0,98 ; p < 0,01) THR = 68,8±14,9 (;- -): y = 0,5419 + 0,0008629x – 3,941e-7x2 THR = 83,5±11,4 (;- -): y = 0,586 + 0,0009194x – 4,6538e-7x2 (r2 = 0,92 ; p < 0,05) (r2 = 0,90 ; p < 0,05) THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 0,6748 + 0,0009621x – 5,5758e-7x2 THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 0,7066 + 0,0008811x – 5,1853e-7x2 (r2 = 0,59 ; p < 0,05) (r2 = 0,53 ; p < 0,05) PB369 Recuperação 1.00 qN (unidades relativas) qN (unidades relativas) PB269 Recuperação 0.75 0.50 0.25 0.00 0 250 500 750 1000 1250 1.00 0.75 0.50 0.25 0.00 1500 0 250 PPFD (µ µmol fotões.m-2.s-1) 500 750 1000 1250 1500 PPFD (µ µ mol fotões.m-2.s-1) THR = 76,8±3,2 (;–): y = 0,564 + 0,001019x – 5,2704e-7x2 THR = 73,9±1,4 (;–): y = 0,6037 + 0,0003279x (r2 = 0,97 ; p < 0,01) (r2 = 0,74 ; p < 0,05) THR = 76,1±5,7 (;- -): y = 0,4476 + 0,001202x – 5,9714e-7x2 THR = 74,0±16,0 (;- -): y = 0,603 + 0,0007018 – 3,0625e-7x2 2 (r = 0,99 ; p < 0,01) (r2 = 0,96 ; p < 0,05) -7 2 THR = 37,0±3,2 (∆;⋅⋅⋅⋅): y = 0,6748 + 0,0009621x – 5,5758e x 2 (r = 0,59 ; p < 0,05) THR = 26,7±11,2 (∆;⋅⋅⋅⋅): y = 0,7066 + 0,0008811x – 5,1853e-7x2 (r2 = 0,53 ; p < 0,05) Fig. 5.19 – Variação do coeficiente de amortecimento não-fotoquímico (qN) em função da densidade de fluxo fotónico fotossintético (PPFD), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. À medida que o stresse hídrico aumenta, as variedades vão perdendo a capacidade de dissipação da energia fotoquimicamente, mesmo para valores reduzidos de PPFD, aumentando a dissipação não-fotoquímica. No entanto, a diferença entre as duas variedades é notória. Para valores de THR de cerca de 85%, em PB369, a evolução de qN com PPFD já se destaca relativamente à observada em PB269, para valores de THR de 70%. Para os valores de THR considerados, PB369 já apresenta um valor de qN superior ao observado em PB269, implicando um desvio da energia captada para processos de dissipação não-fotoquímica, Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 119 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico relacionados com um efeito negativo na maquinaria enzimática fotossintética (dissipação fotoquímica) (Tezara et al., 2003), verificado na Fig. 5.18. Desta forma, a mesma evolução de qN é observada nas duas variedades em THR diferentes, surgindo, mais uma vez, a variedade PB269 como a mais tolerante ao stresse hídrico, dado que os seus processos fotossintéticos se mantém activos a THR mais baixos, comparativamente com a variedade PB369. Ao aprofundar o estudo de qN através da caracterização dos seus componentes (qE, qT e qI), é possível observar as diferenças de comportamento nas duas variedades. Na Fig. 5.20 está representada a variação do coeficiente de amortecimento de alta energia (qE) em função do THR. PB269 PB369 0.8 qE (unidades relativas) qE (unidades relativas) 0.8 0.7 0.6 0.5 0.4 0 25 50 75 0.7 0.6 0.5 0.4 100 0 THR (%) 25 50 75 100 THR (%) Stresse (∆) Stresse (∆; - -): y = 0,7394 – 0,002877x (r2 = 0,76 ; p < 0,01) Recuperação () Recuperação (; –): y = 0,7402 – 0,003023x (r2 = 0,76 ; p < 0,01) Fig. 5.20 – Variação do coeficiente de amortecimento de alta energia (qE) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Podem verificar-se comportamentos distintos nas duas variedades. Em PB269, existe uma manutenção de qE entre 0,5 para 0,6 unidades relativas, para THR até cerca de 50% (ver caixa na Fig. 5.20), embora para THR mais baixos aumente até valores de 0,7 unidades relativas. qE é considerado o mecanismo de dissipação de energia não-fotoquímica mais importante, reflectindo um processo de dissipação de energia desencadeado pelo aumento da concentração de protões no lúmen dos tilacóides, na presença de luz. Esta manutenção de qE, revela que a variedade PB269 não necessita activar este mecanismo até valores moderados de stresse hídrico, mantendo uma actividade fotossintética razoável (ver Fig. 5.4), activando-os apenas em stresse hídrico intenso. Este facto poderá implicar um mecanismo de adaptação para stresse hídrico moderado da variedade PB269. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 120 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Na variedade PB369, qE apresenta um valor de 0,5 unidades relativas perto da hidratação total e sofre um progressivo aumento até 0,7 unidades relativas, para THR de cerca de 20%. Este aumento do qE poderá dever-se ao facto de haver uma diminuição da actividade do ciclo de Calvin, ocorrendo uma baixa regeneração de ADP, com consequente aumento da concentração protónica no lúmen dos tilacóides, verificando-se uma energização dos mesmos (Marques da Silva, 1999; Tezara et al., 1999). Embora as condições de recuperação tenham sido alteradas, qE recupera para os valores iniciais em ambas as variedades. Na Fig. 5.21 está representada a variação do coeficiente de amortecimento de transição de estado (qT) com o THR. PB369 PB269 0.4 qT (unidades relativas) qT (unidades relativas) 0.4 0.3 0.2 0.1 0.3 0.2 0.1 0.0 0.0 0 25 50 75 0 100 25 50 75 100 THR (%) THR (%) Stresse (∆) Stresse (∆; - -): y = 0,3145 – 0,00289x (r2 = 0,91 ; p < 0,01) Recuperação () Recuperação (; –): y = 0,3879 – 0,00755x + 5,274e-5x2 (r2 = 0,94 ; p < 0,01) Fig. 5.21 – Variação do coeficiente de amortecimento de transição de estado (qT) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. qT está relacionado com a transição do estado energético I para o estado energético II, que resulta da fosforilação de polipéptidos do LHCII, permitindo-lhe o movimento lateral das zonas ricas em PSII dos grana para as zonas ricas em PSI do estroma, aumentando a eficiência da fotossíntese através de uma redistribuição de energia entre os dois fotossistemas (Quick e Stitt, 1989). Em ambas as variedades há um aumento de qT com a intensificação do stresse hídrico, embora ligeiramente maior na variedade PB369. Além disso, em PB269, qT parece não variar significativamente até THR perto de 50%. Esta variação parece indicar que o stresse hídrico tem um maior impacto na fosforilação do LHCII, no caso da variedade PB369. No entanto, na recuperação notam-se diferenças entre as duas variedades, uma vez que em PB269, qT Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 121 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico permaneceu constante no valor de 0,2 unidades relativas, enquanto que PB369 recuperou ligeiramente. Na Fig. 5.22 está representada a variação do coeficiente de amortecimento fotoinibitório (qI) em função do THR. O parâmetro qI está relacionado com o mecanismo de dissipação não-fotoquímica pelos próprios centros de reacção do PSII, quando existe uma sobre-redução do pool de quinonas, ocorrendo fotoinibição (Quick e Stitt, 1989). Na variedade PB269 há um ligeiro aumento de 0,2 para 0,4 unidades relativas quando foi atingido um THR de 30%, valor claramente inferior ao obtido na variedade PB369, de 0,6 unidades relativas para THR semelhantes. Um aumento de qI pode estar relacionado com a degradação da proteína D1 do centro de reacção (tipo lesivo) mas também poderá estar relacionado com um mecanismo adaptativo, uma vez que esse aumento pode preceder a degradação dessa proteína (Pieters et al., 2003). Na recuperação, PB269 manteve-se nas 0,4 unidades relativas, enquanto que em PB369 houve uma diminuição de 0,8 para 0,4 unidades relativas. PB269 PB369 1.0 qI (unidades relativas) qI (unidades relativas) 1.0 0.8 0.6 0.4 0.2 0.0 0 25 50 75 0.8 0.6 0.4 0.2 0.0 100 0 THR (%) 25 50 75 100 THR (%) Stresse (∆; - -): y = 0,5118 – 0,002972x (r2 = 0,66 ; p < 0,01) Stresse (∆; - -): y = 1,067 – 0,01818x + 9,4599e-5x2 Recuperação () (r2 = 0,98 ; p < 0,05) Recuperação (; –): y = 1,123 – 0,0221x + 0,0001524x2 (r2 = 0,92 ; p < 0,01) Fig. 5.22 – Variação do coeficiente de amortecimento fotoinibitório (qI) em função do teor hídrico relativo (THR), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Globalmente, a análise dos componentes de qN mostra que qE tem um peso maior na variedade PB269, enquanto qI e qT têm mais relevo na variedade PB369, durante a imposição do stresse. Isto parece sugerir que a variedade PB269 induz mecanismos mais rápidos (qE) de Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 122 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico ajustamento a situações de variação da disponibilidade hídrica e possui uma menor proporção de danos no PSII (qI). Na Fig. 5.23 está representada a variação da taxa de transporte electrónico (ETR) em função de PPFD, para diferentes THR, apresentando um perfil semelhante ao observado na variação de A com PPFD (Fig. 5.4). PB269 Stresse PB369 Stresse 80 ETR (µ mol.m-2.s-1) ETR (µ µ mol m-2 s-1) 80 60 40 20 0 0 250 500 -2 60 40 20 0 750 0 -1 500 750 -2 PPFD (µ mol fotões.m .s ) THR = 93,9±2,9 (;–): y = 241144 * ((1 – 250 -1 PPFD (µ µmol fotões.m .s ) exp (– 0,3215x / THR = 95,6±1,5 (;–): y = 221588 * ((1 – exp (– 0,3611x / 241144)) * exp (– 558,3x / 241144) (r2 = 0,98 ; p < 0,01) 221588)) * exp (– 592,4x / 221588) (r2 = 0,92 ; p < 0,01) THR = 68,8±14,9 (;- -): y = 180433 * ((1 – THR = 83,5±11,4 (;- -): y = 85740 – 8.478 * ((1 – exp (– 0,1917x exp (– 0,2428x / 180433)) * exp (– 483,1x / 180433) (r2 = 0,87 ; p < 0,01) / 85740)) * exp (– 267,1x / 85740) (r2 = 0,92 ; p < 0,01) THR = 37,0±3,2 (∆) THR = 26,7±11,2 (∆) PB269 Recuperação PB369 Recuperação 80 ETR (µ mol m-2 s-1) ETR (µ µ mol.m-2.s-1) 80 60 40 20 0 0 250 500 exp (– 0,2841x / THR = 76,1±5,7 (;- -): y = 178010 * ((1 – exp (– 0,2946x / 20 0 PPFD (µ µ mol fotões.m-2.s-1) THR = 76,8±3,2 (;–): y = 121101 * ((1 – 40 0 750 121101)) * exp (– 617,8x / 121101) (r2 = 0,90 ; p < 0,05) 60 250 500 -2 750 -1 PPFD (µ µmol fotões.m .s ) THR = 73,9±1,4 (;–): y = 162754 * ((1 – exp (– 0,2847x / 162754)) * exp (– 387,9x / 162754) (r2 = 0,83 ; p < 0,05) THR = 74,0±16,0 (;- -): y =170994 * ((1 – exp (– 0,2212x / 178010)) * exp (– 713,5e7x / 178010) (r2 = 0,92 ; p < 0,01) 170994)) * exp (– 459,6x / 170994) (r2 = 0,89 ; p < 0,01) THR = 37,0±3,2 (∆) THR = 26,7±11,2 (∆) Fig. 5.23 – Variação da taxa de transporte electrónico (ETR) em função da densidade de fluxo fotónico fotossintético (PPFD), nas experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Observa-se na Tabela 5.5, os parâmetros retirados da aplicação do modelo de Platt et al. (1980) na Fig. 5.23. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 123 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Tabela 5.5 – Parâmetros da equação de Platt et al. (1980), resultantes da aplicação do modelo à Fig. 5.23., nas duas variedades portuguesas de milho. PB269 PB369 Stresse Stresse THR (%) THR (%) Parâmetros 93,9 ± 2,9 68,8 ± 14,9 37,0 ± 3,2 95,6 ± 1,5 83,5 ± 11,4 26,7 ± 11,2 ETRmáx (mmol.m-2.s-1) 51,07 ± 22,43 33,35 ± 8,32 - 49,67 ±13,40 22,63 ± 8,57 - α 0,322 ± 0,040 0,243 ± 0,089 - 0,361 ± 0,028 0,192 ± 0,013 - (µmol CO2 / µmol fotões) Recuperação Recuperação THR (%) THR (%) Parâmetros 37,0 ± 3,2 76,1 ± 5,7 76,8 ± 3,2 26,7 ± 11,2 74,0 ± 16,0 73,9 ± 1,4 ETRmáx (mmol.m-2.s-1) - 27,03 ± 12,43 20,48 ± 6,92 - 30,27 ±12,82 43,93 ± 6,04 α - 0,295 ± 0,067 0,284 ± 0,037 - 0,221 ± 0,040 0,285 ± 0,048 (µmol CO2 / µmol fotões) Pode observar-se uma ETR máxima de cerca de 50 µmol.m-2.s-1, em ambas as variedades, para uma PPFD entre os 400 e os 500 µmol fotões.m-2.s-1, valores em que as plantas foram cultivadas na câmara climatizada. Ambas as variedades demonstram uma diminuição deste parâmetro para PPFD mais elevadas, demonstrando um efeito de fotoinibição, e um valor de eficiência fotossintética semelhante (valor de α). No que respeita ao parâmetro β, o modelo não conseguiu estimar este parâmetro com precisão, pelo que não são apresentados na Tabela 5.5. À medida que o stresse hídrico aumenta, com a consequente diminuição do THR, verifica-se uma diminuição de ETR, por possível afectação da cadeia transportadora de electrões e/ou danos no PSII e também devido a uma diminuição da taxa de actividade do ciclo de Calvin. A variedade PB269 demonstra uma maior ETR para THR mais baixos, comparativamente com a variedade PB369, bem como uma eficiência fotossintética superior (valor de α superior em PB269). No final do período de stresse, em que ambas as variedades apresentavam um THR muito reduzido, o ETR era reduzido para PPFD mais baixos e nulos para PPFD mais elevados. O transporte de electrões a THR muito baixos, quando A é pequeno ou nulo, é mantido principalmente devido ao funcionamento da fotorrespiração (Lawlor e Cornic, 2002). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 124 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Durante a recuperação verificou-se que PB269 apresentou algumas dificuldades em recuperar para os valores iniciais, embora PB369 tenha recuperado quase totalmente o seu perfil de ETR em resposta a um gradiente de luz. Na Fig. 5.23, não foram representados os pontos obtidos a PPFD de 1300 µmol fotões.m-2.s-1, uma vez que eram nulos e não estavam concordantes com as taxas fotossintéticas registadas durante este ensaio para aquela gama de PPFD, pelo que terá ocorrido uma interferência entre o fluorómetro e o PPFD elevado que foi utilizado (Maxwell et al., 1999). Desta forma, verifica-se que o stresse hídrico afecta grande parte da componente química e fotoquímica da fotossíntese nas duas variedades, embora PB269 tenha claramente surgido como a variedade mais tolerante, apresentando uma estratégia de redução progressiva da fotossíntese com a diminuição do THR, por dissipação de energia não-fotoquímica desencadeado pelo aumento da concentração de protões no lúmen dos tilacóides na presença de luz (qE), principalmente em stresse moderado. Por outro lado, a variedade PB369 apresentou sérias dificuldades durante o stresse hídrico, com danos no PSII a THR mais elevados (redução de Fv/Fm e aumento de qI) do que em PB269. 5.3.4 5.3.4. Clorofilas Totais e Carotenóides A quantificação das clorofilas e carotenóides totais foi realizada nas amostras recolhidas durante o ensaio das curvas A/CO2a, e, desta forma, o período de stresse e a recuperação na câmara climatizada foram efectuadas em condições idênticas. Na Fig. 5.24 está representada a variação da clorofila a em função do THR. Observase, em ambas as variedades, um valor de cerca de 3 mg.g-1 peso seco perto da hidratação total. Na variedade PB269, ocorreu uma diminuição para cerca de 2 mg.g-1 peso seco para um THR de cerca de 60%, e durante a recuperação permanece entre 1-2 mg.g-1 peso seco. Estes níveis reduzidos de clorofila a durante a recuperação poderão ser, em parte, responsáveis pela fraca recuperação desta variedade (ver Fig. 5.9). Por outro lado, na variedade PB369, só se obteve uma medição abaixo de THR de 80%, não ocorrendo qualquer variação significativa entre o período de stresse e o de recuperação. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 125 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB269 PB369 5 Clorofila a (mg.g-1 peso seco) Clorofila a (mg.g-1 peso seco) 5 4 3 2 1 0 60 70 80 90 4 3 2 1 0 100 60 THR (%) 70 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 5.24 – Variação da clorofila a em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Na Fig. 5.25 está representada a variação da clorofila b em função do THR. PB269 PB369 1.00 Clorofila b (mg.g-1 peso seco) Clorofila b (mg.g-1 peso seco) 1.00 0.75 0.50 0.25 0.00 0.75 0.50 0.25 0.00 60 70 80 90 100 60 THR (%) 70 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 5.25 – Variação da clorofila b em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Na variedade PB269, ocorre uma ligeira descida da clorofila b de 0,50 mg.g-1 peso seco para 0,25 mg.g-1 peso seco, com a diminuição do THR. Após irrigação, permanece em torno deste último valor. Na variedade PB369, tal como na clorofila a, não se verificam alterações, dado que só existe um ponto abaixo do THR de 80%. Na Fig. 5.26 está representada a variação da clorofila total com o THR. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 126 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB269 PB369 5 Clorofila total (mg.g-1 peso seco) Clorofila total (mg.g-1 peso seco) 5 4 3 2 1 0 60 70 80 90 4 3 2 1 0 100 60 THR (%) 70 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 5.26 – Variação da clorofila total em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. Embora estes dados não sejam estatisticamente significativos, observa-se uma variação semelhante à observada para a clorofila a, dado que a clorofila b apresenta valores muito mais baixos e não apresenta variações significativas. Durante períodos de stresse hídrico, a diminuição do conteúdo em clorofilas é comum, tendo sido observada quer em milho (Alberte et al., 1977), quer em trigo (Loggini et al., 1999). Tal facto deve-se à exposição das clorofilas a agentes de degradação, como ROS, com a diminuição da quantidade de água presente nas células e do próprio desequilíbrio entre a fase fotoquímica e a fase química (Loggini et al., 1999). Dependendo do nível de stresse a que as plantas estão sujeitas, poderá haver maior ou menor degradação de clorofila. No entanto, a variedade PB269 consegue suportar stresses hídricos elevados mantendo alguma actividade fotossintética, mas não conseguindo impedir a degradação da clorofila, o que vai implicar alguns problemas na recuperação. Por outro lado, na azinheira e no sobreiro, espécies tipicamente mediterrânicas, ocorre uma diminuição da clorofila, durante o Verão, de forma a regular a fotossíntese em condições de baixos recursos hídricos e elevada radiação solar (Maroco et al., 2002). Na Fig. 5.27 está representada a variação dos carotenóides totais com o THR. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 127 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB269 PB369 3 Carotenóides totais (mg.g-1 peso seco) Carotenóides totais (mg.g-1 peso seco) 3 2 1 0 60 70 80 90 2 1 0 100 60 THR (%) 70 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 5.27 – Variação dos carotenóides totais em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 5 folhas de diferentes plantas. A variedade PB269 apresenta um valor de carotenóides totais de cerca de 1,7 mg.g-1 peso seco em folhas totalmente hidratadas, enquanto PB369 apresenta um valor de 1,4 mg.g-1 peso seco. Com a diminuição do THR, em PB269, os carotenóides diminuem até cerca de 1 mg.g-1 peso seco. Em PB369, não se observam quaisquer diferenças significativas até THR de 85%. Durante o período de recuperação, PB369 não sofre quaisquer alterações significativas. Por outro lado, em PB269, os carotenóides totais diminuem para cerca de 0,5 mg.g-1 peso seco, durante o período de recuperação. Tal facto pode revelar mecanismos diferentes de resposta ao stresse hídrico, nomeadamente algumas alterações nas vias metabólicas dos carotenóides, que podem levar, em parte, à produção de ABA, com consequente aumento da tolerância ao stresse hídrico (Schwartz et al., 1997; Borel et al., 2001). 5.3.5. 5.3.5. Metabolismo do Carbono As actividades das principais enzimas do metabolismo fotossintético das duas variedades de milho seleccionadas foram determinadas a partir de amostras recolhidas durante o ensaio de determinação das curvas A/PPFD. Durante este ensaio, as câmaras climatizadas sofreram uma avaria durante o período de recuperação, aumentando a temperatura dos 25ºC para os 38ºC, pelo que é necessário ter este facto em conta na análise e discussão dos dados obtidos. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 128 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.3.5.1. Ribuloseibulose-1,51,5-Bisfosfato Carboxilase/O arboxilase/Oxigenase (RuBisCO) RuBisCO) Na Fig. 5.28 está representada a variação da actividade inicial da RuBisCO em função do THR, no ensaio de stresse lento. PB369 5 Actividade inicial da RuBisCO µ mol RuBP.hr-1.mg -1 proteína) (µ Actividade inicial da RuBisCO µ mol RuBP.hr-1.mg-1 proteína) (µ PB269 4 3 2 1 0 25 50 75 100 THR (%) 5 4 3 2 1 0 25 50 75 100 THR (%) Stresse (∆; - -): y = – 0,8214 + 0,03390x (r2 = 0,71 ; p < 0,01) Stresse (∆) Recuperação (; –) Recuperação (; –) Fig. 5.28 – Variação da actividade inicial da RuBisCO em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Perto da hidratação total, a variedade PB369 apresenta uma actividade inicial da RuBisCO de 3,2 µmol RuBP.hr-1.mg-1 proteína solúvel, a qual é superior à da variedade PB269 (2,6 µmol RuBP.hr-1.mg-1 proteína solúvel). Em ambas as variedades há uma diminuição da actividade inicial da RuBisCO com a diminuição do THR. Para THR de cerca de 30%, as duas variedades apresentam actividades enzimáticas iniciais da RuBisCO muito reduzidas que mantêm durante a recuperação. No entanto, no final do stresse, a A apresentava valores negativos (Fig. 5.4), que recuperou muito pouco durante o período de recuperação. O facto de A ser negativo, embora exista alguma actividade inicial da RuBisCO para o mesmo THR poderá estar relacionado com uma reduzida A bruta que corresponderá a uma A efectiva negativa. A diminuição da actividade inicial da RuBisCO também poderá estar relacionada com a inibição ou não-activação dos locais activos da enzima (Parry et al., 2002). Inibidores como o CA1P ligam-se fortemente aos locais de carbamilação em condições de luz reduzida e à noite (Holbrook et al., 1994) e a sua remoção é facilitada pela RuBisCO activase (Hammond et al., 1998), pelo que pode a sua actividade ser afectada à medida que o stresse hídrico progride. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 129 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Na Fig. 5.29 está representada a variação da actividade total da RuBisCO em função do THR, no ensaio de stresse lento. PB369 Actividade total da RuBisCO µ mol RuBP.hr-1.mg -1 proteína) (µ Actividade total da RuBisCO µ mol RuBP.hr-1.mg -1 proteína) (µ PB269 15 10 5 0 25 50 75 100 15 10 5 0 25 THR (%) 50 75 100 THR (%) Stresse (∆; - -): y = – 2,145 + 0,1083x (r2 = 0,76 ; p < 0,01) Stresse (∆) Recuperação () Recuperação () Fig. 5.29 – Variação da actividade total da RuBisCO em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Ambas as variedades apresentam um valor de actividade total da RuBisCO de cerca de 9 µmol RuBP.hr-1.mg-1 proteína solúvel, perto da hidratação total. Tal como na actividade inicial, também a actividade total diminui de forma semelhante com a diminuição do THR. Durante a recuperação verifica-se, tal como na actividade inicial, uma tendência para haver uma diminuição da actividade total, com o aumento do THR (ver caixas Fig. 5.28 e 5.29). É preciso ter em conta as condições de temperatura durante a recuperação que parecem ter afectado quer a actividade inicial, quer a actividade total da RuBisCO, que poderão resultar de alterações na conformação estrutural da enzima e, assim, a sua actividade (Parry et al., 2002). Também poderia dever-se a alterações da concentração das proteínas totais, mas tal não se observou após a quantificação das mesmas. Observa-se, em ambas as actividades, um padrão semelhante de diminuição com o progresso do stresse hídrico. No entanto, a variedade PB269 apresenta uma diminuição gradual com o THR, enquanto a variedade PB369 apresenta uma diminuição em patamar, ocorrendo uma diminuição brusca das actividades num determinado THR (cerca de 80%), como já havia sido verificado durante a fase I deste trabalho. Comparando com os dados da Fig. 5.4, em que ambas as variedades apresentam valores negativos da A para THR perto dos 30%, poderá constatar-se que para estes valores reduzidos de THR, a respiração apresenta uma contribuição relativa mais importante que a da fotossíntese (Lawlor e Cornic, 2002). No entanto, A recupera parcialmente, embora as Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 130 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico actividades da RuBisCO permaneçam inalteradas, o que poderá em parte estar relacionado com a recuperação da estrutura das folhas (que se encontravam enroladas, possivelmente com a estrutura histológica distorcida), que facilita a difusão do CO2 até aos locais de fixação, ou ainda com a diminuição da respiração, com o aumento da gs (correspondendo a uma diminuição da limitação estomática) ou com a recuperação dos centros activos do PSII. Na Fig. 5.30 está representada a activação da RuBisCO em função do THR, no ensaio de stresse lento. Perto da hidratação total, pode observar-se que a activação da RuBisCO na variedade PB369 (cerca de 40%) é ligeiramente superior à da variedade PB269 (cerca de 30%). Embora em ambas haja uma certa tendência para diminuir com a diminuição do THR, esta não é estatisticamente significativa, ou seja, as diminuições das actividades da RuBisCO não se devem a grandes alterações na activação da enzima e, provavelmente, dever-se-á a uma diminuição de Kcat (Parry et al., 2002). PB269 PB369 60 Activação da RuBisCO (%) Activação da RuBisCO (%) 60 40 20 0 25 50 75 40 20 0 25 100 THR (%) 50 75 100 THR (%) Stresse (∆) Stresse (∆) Recuperação () Recuperação () Fig. 5.30 – Variação da activação da RuBisCO em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Na redução de A, a RuBisCO tem certamente um papel importante na medida que ocorre uma diminuição das suas actividades, a qual poderá ter como origem uma diminuição da quantidade de RuBisCO presente nas folhas (Gallais e Hirel, 2004), embora tal facto só pudesse ser comprovado por uma quantificação efectiva da enzima, dado que poderá ser resultado apenas de uma inibição ou não-activação da enzima (Parry et al., 2002). No entanto, como tem sido observado ao longo deste trabalho, a RuBisCO não é o único factor a contribuir para a diminuição de A, como o demonstram os resultados de fluorescência da clorofila. O conjunto de todos os processos, desde a captação da luz até à fixação do carbono, formam uma rede complexa de interacções, as quais serão afectadas pelo stresse hídrico em Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 131 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico diferentes graus, contribuindo de formas diferentes para a redução de A, com a diminuição do THR. Na Fig. 5.31 está representada a variação das actividades inicial e total da RuBisCO em função do THR, no ensaio de stresse rápido. As actividades observadas perto da hidratação total são inferiores às observadas nas plantas do stresse lento devido ao facto de terem passado por dois dias de temperaturas elevadas (38ºC) devido a avaria da câmara climatizada. Este dado é preciso ter em conta na análise dos dados do stresse rápido. Em ambas as variedades, não há uma variação estatisticamente significativa das actividades inicial e total da RuBisCO com a diminuição do THR. Contudo, em PB269 observa-se uma descida da actividade para THR inferiores a 50%. PB369 5 Actividade da RuBisCO µ mol RuBP.hr-1.mg -1 proteína) (µ Actividade da RuBisCO µ mol RuBP.hr-1.mg-1 proteína) (µ PB269 4 3 2 1 0 25 50 75 100 5 4 3 2 1 0 25 50 THR (%) 75 100 THR (%) Actividade inicial (∆) Actividade inicial (∆) Actividade total () Actividade total () Fig. 5.31 – Variação das actividades inicial (∆) e total () da RuBisCO em função do teor hídrico relativo (THR), ao longo das experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Também na activação da RuBisCO não há uma variação estatisticamente significativa, situando-se entre 30% e 40%, ao longo do THR (Fig. 5.32). Lal e Edwards (1996) encontraram valores superiores de activação para Zea mays (cerca de 80%). Os valores observados neste trabalho poderão reflectir as condições de crescimento das plantas, uma vez que o estado de activação da RuBisCO está dependente da PPFD ambiente (Perchorowicz et al., 1981; Zhang e Portis, 1999). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 132 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB269 PB369 60 Activação da RuBisCO (%) Activação da RuBisCO (%) 60 40 20 0 25 50 75 40 20 0 25 100 50 THR (%) 75 100 THR (%) Fig. 5.32 – Variação da activação da RuBisCO em função do teor hídrico relativo (THR), ao longo das experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Desta forma, em ambas as variedades, verificam-se diferenças nas actividades da RuBisCO quando sujeitas a diferentes tipos de stresse. No stresse lento, há uma progressiva diminuição das actividades da RuBisCO com a diminuição do THR, enquanto que no stresse rápido, a enzima não sofre alteração nas suas actividades. No entanto, verificou-se na fase I deste trabalho que ocorria uma diminuição da A com a diminuição do THR, no stresse rápido (Fig. 4.13). Tal facto indica que se deverá a limitações estomáticas da fotossíntese (Fig. 4.14) ou a outro tipo de limitações não-estomáticas, nomeadamente a elementos da cadeia transportadora de electrões (Fig. 4.25) ou a alterações das actividades de outras enzimas do ciclo fotossintético do carbono (Fig.5.35 e Fig. 5.37). 5.3.5.2. 5.3.5.2. Fosfoenolpiruvato Carboxilase (PEPC) Na Fig. 5.33 está representada a variação da actividade inicial da PEPC em função do THR. Perto da hidratação total, a variedade PB269 apresenta uma actividade inicial da PEPC de cerca de 30 µmol PEP.hr-1.mg-1 proteína solúvel, inferior à observada na variedade PB369 (cerca de 40 µmol PEP.hr-1.mg-1 proteína solúvel). No entanto, verifica-se que, com a diminuição do THR, a actividade inicial da PEPC vai diminuindo até atingir o valor de 10 µmol PEP.hr-1.mg-1 proteína solúvel para THR na ordem dos 30%. Isto não está de acordo com o observado para o milho, em que a actividade da PEPC se mantém ou chega a aumentar em condições de stresse moderado, a qual tem sido relacionada com o aumento da concentração de prolina, através do aumento dos seus percursores (Jeanneau et al., 2002). Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 133 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico PB369 50 Actividade inicial da PEPC µ mol PEP.hr-1.mg -1 proteína) (µ Actividade inicial da PEPC µ mol PEP.hr-1.mg-1 proteína) (µ PB269 40 30 20 10 0 25 50 75 100 50 40 30 20 10 0 25 50 THR (%) 75 100 THR (%) Stresse (∆; - -): y = 4,161 + 0,2432x (r2 = 0,59 ; p < 0,05) Stresse (∆; - -): y = 0,06901 + 0,4057x (r2 = 0,83 ; p < 0,01) Recuperação () Recuperação () Fig. 5.33 – Variação da actividade inicial da PEPC em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Durante a recuperação, tal como já foi verificado nas actividades da RuBisCO, a actividade inicial da PEPC de ambas as variedades permanece constante no valor observado no final do período de stresse. Na Fig. 5.34 está representada a variação da actividade máxima da PEPC em função do THR. PB369 70 Actividade máxima da PEPC µ mol PEP.hr-1.mg -1 proteína) (µ Actividade máxima da PEPC µ mol PEP.hr-1.mg-1 proteína) (µ PB269 60 50 40 30 20 10 0 25 50 75 100 THR (%) 70 60 50 40 30 20 10 0 25 50 75 100 THR (%) Stresse (∆; - -): y = 2,992 +0,3476x (r2 = 0,89 ; p < 0,01) Stresse (∆; - -): y = 7,812 + 0,4423x (r2 = 0,85 ; p < 0,01) Recuperação () Recuperação () Fig. 5.34 – Variação da actividade máxima da PEPC em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. A variedade PB269 apresenta, perto da hidratação total, uma actividade máxima da PEPC de cerca de 40 µmol PEP.hr-1.mg-1 proteína solúvel. Na variedade PB369, o valor da actividade total da PEPC é superior (50 µmol PEP.hr-1.mg-1 proteína solúvel). Em ambas as variedades, há uma diminuição da actividade total com a diminuição do THR, atingindo os 15 Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 134 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico µmol PEP.hr-1.mg-1 proteína solúvel na variedade PB269 e os 20 µmol PEP.hr-1.mg-1 proteína solúvel na variedade PB369, para THR de 30%. No período de recuperação, mais uma vez, a actividade máxima da PEPC permaneceu inalterada nos valores registados no final do período de stresse. Nos ensaios de stresse rápido, verificam-se, tal como nas actividades da RuBisCO, diferenças significativas em relação aos ensaios de stresse lento. Na Fig. 5.35 está representada a variação das actividades inicial e máxima da PEPC em função do THR, no ensaio de stresse rápido. PB369 Actividade da PEPC µ mol PEP.hr-1.mg -1 proteína) (µ Actividade da PEPC µ mol PEP.hr-1.mg -1 proteína) (µ PB269 50 40 30 20 10 0 25 50 75 100 THR (%) 50 40 30 20 10 0 25 50 75 100 THR (%) Actividade inicial (∆) Actividade inicial (∆) Actividade máxima () Actividade máxima () Fig. 5.35 – Variação das actividades inicial (∆) e máxima () da PEPC em função do teor hídrico relativo (THR), ao longo das experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Embora as diferenças observadas entre os valores das actividades da PEPC, entre as duas variedades, sejam notórias, em nenhuma há alteração com a diminuição do THR. No entanto, mais uma vez se relembra que as actividades observadas perto da hidratação total são inferiores às observadas nas plantas do stresse lento devido ao facto de terem passado por dois dias de temperaturas elevadas (38ºC) devido a avaria da câmara climatizada. A regulação da enzima PEPC é efectuada essencialmente pela resposta a metabolitos efectores, como a inibição pelo malato e a activação por açúcares fosfatados (Ting e Osmond, 1973; Doncaster e Leegood, 1987). Os efeitos destes metabolitos são modulados pela fosforilação da enzima, a qual é reversível, dependente da luz e levada a termo pela enzima PEPCK. Esta fosforilação leva a uma diminuição da sensibilidade à inibição pelo malato e a um aumento da sensibilidade à activação pela glucose-6-fosfato (Jao e Chollet, 1992; Bailey et al., 2000). Por outro lado, a actividade da enzima PEPCK está dependente da quantidade de Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 135 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico ATP presente, a qual diminui durante períodos de stresse hídrico (Tezara et al., 1999). Também as curvas A/Ci (não representadas, mas muito semelhantes às curvas A/CO2a da Fig. 5.9), através da análise do seu declive inicial (eficiência de carboxilação), indicam algumas propriedades cinéticas da enzima (Bailey et al., 2000). Desta forma, observa-se uma diminuição do declive da curva à medida que o stresse hídrico aumenta, reflectindo uma diminuição da actividade da PEPC. Na variedade PB269, existe alguma recuperação após irrigação, enquanto que na variedade PB369 a recuperação não existe, continuando o declive inicial do gráfico a diminuir. 5.3.5.3. Enzima Málica Dependente de NADP (EM(EM-NADP) Na Fig. 5.36 está representada a variação da actividade da enzima málica dependente de NADP em função do THR, no ensaio de stresse lento. PB369 Actividade da EM-NADP µ mol malato.hr-1.mg -1 proteína) (µ Actividade da EM-NADP µ mol malato.hr-1.mg -1 proteína) (µ PB269 20 15 10 5 0 25 50 75 100 THR (%) 15 10 5 0 25 50 75 100 THR (%) Stresse (∆; - -): y = – 3,609 + 0,1982x (r2 = 0,92 ; p < 0,01) Recuperação () 20 Stresse (∆) Recuperação (; –): y = 16,84 – 0,1537x (r2 = 0,66 ; p < 0,05) Fig. 5.36 – Variação da actividade máxima da EM-NADP em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Ambas as variedades apresentam valores de actividade desta enzima de cerca de 15 µmol malato.h-1.mg-1 proteína solúvel, na hidratação total. No entanto, na variedade PB269, há uma clara diminuição da actividade com a redução do THR até 5 µmol malato.hr-1.mg-1 proteína solúvel em THR de cerca de 30%. Em PB369, a variação não é significativa. Durante a recuperação, PB269 mantém a actividade em 5 µmol malato.hr-1.mg-1 proteína solúvel, enquanto que em PB369 há uma diminuição até atingir o mesmo valor de Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 136 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico actividade a THR de 75%. No entanto, estes dados não permitem discutir a recuperação, dadas as circunstâncias em que esta ocorreu. A variação da actividade no ensaio do stresse lento contrasta claramente com a variação obtida no ensaio de stresse rápido (Fig. 5.37). No ensaio de stresse rápido, a actividade não sofre alterações estatisticamente significativas, mantendo-se constante em ambas as variedades. No entanto, a actividade em PB269, na hidratação total, é cerca de metade da verificada nos ensaios de stresse lento, o que pode indicar alguma perturbação nesta variedade durante o crescimento das plantas. PB369 20 Actividade da EM-NADP µ mol malato.hr-1.mg -1 proteína) (µ Actividade da EM-NADP µ mol malato.hr-1.mg -1 proteína) (µ PB269 15 10 5 0 25 50 75 100 20 15 10 5 0 25 THR (%) 50 75 100 THR (%) Fig. 5.37 – Variação da actividade máxima da EM-NADP em função do teor hídrico relativo (THR), ao longo das experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Esta manutenção da actividade da enzima málica no milho, em condições de stresse rápido, já havia sido verificada por outros autores (Saccardy et al., 1996), e a expressão do gene desta enzima em plantas de tabaco, melhorou consideravelmente a sua tolerância ao stresse hídrico (Laporte et al., 2002). No entanto, as actividades observadas perto da hidratação total são inferiores às observadas nas plantas do stresse lento pelas razões já referidas. 5.3.6. 5.3.6. Determinação de Solutos Compatíveis A quantificação de aminoácidos livres e prolina, para averiguar a possível presença de solutos compatíveis, foi realizada em três ensaios, dois de stresse lento e um de stresse rápido. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 137 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico 5.3.6.1. 5.3.6.1. Aminoácidos Livres A concentração de aminoácidos livres, na variedade PB269, permaneceu relativamente constante ao longo da experiência de stresse lento (cerca de 3 µg.mg-1 peso seco), ao contrário da variedade PB369 que, para THR semelhantes (30 %), duplicou a sua concentração (de cerca de 3-4 µg.mg-1 peso seco para cerca de 8 µg.mg-1 peso seco) (Fig. 5.38), possivelmente devido a efeitos de senescência, resultante de uma maior actividade endoproteolítica, característica de espécies susceptíveis ao stresse hídrico (Arad e Richmond, 1976; BuchananWollaston, 1997). No período de recuperação do stresse, a concentração pareceu indicar uma leve diminuição em PB269 e uma diminuição marcada em PB369, regressando a valores próximos dos iniciais. Estes dados sugerem a utilização dos aminoácidos na reparação dos danos provocados pelo stresse hídrico, na variedade PB369. PB269 PB369 15 Aminoácidos livres (µ µ g.mg-1 peso seco) Aminoácidos livres (µ g.mg-1 peso seco) 15 10 5 0 0 25 50 75 10 5 0 100 0 THR (%) 25 50 75 100 THR (%) Stresse (∆) Stresse (∆; - -): y = 11,07 – 0,07952x (r2 = 0,76 ; p < 0,01) Recuperação () Recuperação (; –): y = 11,96 – 0,1102x (r2 = 0,79 ; p < 0,01) Fig. 5.38 – Variação dos aminoácidos livres em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Contudo, o período de recuperação foi realizado a uma temperatura diferente da inicial (38ºC e não os 25ºC programados), devido a uma avaria na câmara climatizada, pelo que procedeu-se à realização de um ensaio independente (Fig. 5.39). Neste ensaio, não se atingiram valores de THR tão baixos como no primeiro ensaio (cerca de 60%). Na variedade PB269, a concentração de aminoácidos livres permaneceu relativamente constante (cerca de 3 µg.mg-1 peso seco) (ver caixa da Fig. 5.39) mas no período de recuperação desceu até valores mais baixos (cerca de 1 µg.mg-1 peso seco). Na variedade PB369, a concentração de aminoácidos livres manteve-se relativamente constante até THR de 80%, embora tenha sido obtido um ponto com THR inferior (cerca de 65%) com Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 138 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico uma concentração de aminoácidos livres de cerca de 4 µg.mg-1 peso seco, e durante o período de recuperação diminuiu para cerca de 2 µg.mg-1 peso seco (ver caixas da Fig. 5.39). Este ensaio veio confirmar os dados e reforçar a hipótese colocada no ensaio anterior. PB269 PB369 5 Aminoácidos livres (µ g.mg-1 peso seco) Aminoácidos livres (µ g.mg-1 peso seco) 5 4 3 2 1 0 60 70 80 90 4 3 2 1 0 60 100 70 THR (%) 80 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação (; –): y = 7,603 – 0,06901x (r2 = 0,91 ; p < 0,01) Recuperação () Fig. 5.39 – Variação dos aminoácidos livres em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Procedeu-se também à quantificação dos aminoácidos livres em experiências de stresse rápido (Fig. 5.40). PB269 PB369 6 Aminoácidos livres (µ g.mg-1 peso seco) Aminoácidos livres (µ g.mg-1 peso seco) 6 5 4 3 2 1 0 30 40 50 60 70 80 90 5 4 3 2 1 0 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) y = 6,363 – 0,0308x (r2 = 0,66 ; p < 0,01) Fig. 5.40 – Variação dos aminoácidos livres em função do teor hídrico relativo (THR), ao longo das experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Na variedade PB269, a concentração de aminoácidos livres manteve-se constante ao longo do stresse rápido, enquanto que em PB369 sofreu um aumento significativo. O stresse hídrico provoca uma degradação dos componentes celulares, nomeadamente a nível das membranas celulares (Gigon et al., 2004). No entanto, poderão também estar Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 139 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico relacionadas com a mobilização de proteínas de reserva (Raymond e Smirnoff, 2002). De uma forma ou de outra, é possível verificar as diferenças entre as duas variedades, em ambas as experiências de stresse lento e de stresse rápido. A variedade PB269 parece possuir mecanismos que evitam uma maior degradação dos seus componentes celulares ou manter a sua actividade sem aumento da mobilização das suas reservas proteicas, em comparação com PB369. 5.3.6.2. 5.3.6.2. Prolina Nas duas variedades, a concentração de prolina, durante o período de stresse lento, não sofreu uma variação significativa (Fig. 5.41). Tal facto poderá estar relacionado com a diminuição da actividade da enzima PEPC, a qual já foi apontada como responsável pelo aumento da prolina, conduzindo a um ajustamento osmótico (Jeanneau et al., 2002). Contudo, durante o período de recuperação, a concentração de prolina diminuiu de forma marcada na variedade PB269. Em PB369 mostrou alguma tendência para diminuir mas não de forma significativa. PB269 PB369 10 Prolina (µ g.mg-1 peso seco) Prolina (µ g.mg-1 peso seco) 10 8 6 4 2 0 0 25 50 75 8 6 4 2 0 100 0 THR (%) 25 50 75 100 THR (%) Stresse (∆) Stresse (∆) Recuperação (; –): y = 8,639 – 0,06283x (r2 = 0,65 ; p < 0,01) Recuperação () Fig. 5.41 – Variação da prolina em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. As amostras utilizadas nesta experiência também estiveram sujeitas a temperaturas elevadas durante o período de recuperação (38ºC), pelo que se repetiu o ensaio numa experiência independente. Os resultados foram semelhantes, com ambas as variedades a não sofrerem variações significativas na concentração de prolina ao longo do período de stresse (Fig. 5.42). Em PB269, a concentração de prolina diminuiu de forma marcada durante o período de Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 140 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico recuperação para cerca de 30% da concentração inicial. Esta diminuição pode ser resultado da indução da prolina desidrogenase, cuja actividade aumenta durante a recuperação do stresse hídrico (Hare et al., 1999). A variedade PB369 demonstra uma certa tendência para ocorrer a diminuição da concentração de prolina durante a recuperação, embora não seja significativa. PB269 PB369 10 Prolina (µ g.mg-1 peso seco) Prolina (µ g.mg-1 peso seco) 10 8 6 4 2 0 60 70 80 90 8 6 4 2 0 60 100 70 80 THR (%) 90 100 THR (%) Stresse (∆) Stresse (∆) Recuperação (; –): y = 16,18 – 0,147x (r2 = 0,78 ; p < 0,01) Recuperação () Fig. 5.42 – Variação da prolina em função do teor hídrico relativo (THR), ao longo das experiências de stresse lento. Os pontos foram obtidos de 3 folhas de diferentes plantas. Nas experiências de stresse rápido, também se verificam diferenças nas duas variedades (Fig. 5.43). Em PB269, ocorreu uma diminuição da concentração de prolina para cerca de metade da inicial, enquanto que, em PB369, a concentração de prolina se manteve constante ao longo do stresse. PB269 PB369 10 Prolina (µ g.mg-1 peso seco) Prolina (µ g.mg-1 peso seco) 10 8 6 4 2 0 30 40 50 60 70 80 90 8 6 4 2 0 30 100 THR (%) 40 50 60 70 80 90 100 THR (%) y = – 0,2655 + 0,01805x (r2 = 0,73 ; p < 0,05) Fig. 5.43 – Variação da prolina em função do teor hídrico relativo (THR), ao longo das experiências de stresse rápido. Os pontos foram obtidos de 3 folhas de diferentes plantas. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 141 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Os dados parecem indicar que ambas as variedades possuem uma reserva de prolina, a qual tem sido indicada como sendo um soluto compatível que permite manter actividade celular a stresses hídricos elevados (Hare et al., 1999; Heuer, 2003; Nayyar, 2003; Larher et al., 2003) e também com função na remoção de radicais hidroxilo (Bohnert e Shen, 1999), os quais afectam a actividade das enzimas do ciclo de Calvin (Shen et al., 1997a). Além disso, mantém a relação NADP+/NADPH em valores compatíveis com o metabolismo e é essencial no desenvolvimento da planta em condições sem stresse (Hare et al., 1999; Raymond e Smirnoff, 2002). Estes ensaios parecem indicar que a reserva de prolina é utilizada como fonte de carbono, azoto e energia rapidamente mobilizável durante o período de recuperação (Hare et al., 1999), mais facilmente utilizada pela variedade PB269 do que pela variedade PB369, provavelmente compensando as baixas taxas de recuperação das actividades da RuBisCO e da PEPC, durante esse período. Tal facto, pode observar-se nas experiências de stresse rápido em que PB269 rapidamente inicia a mobilização das reservas de prolina, ao contrário de PB369, cujas reservas não são tão facilmente mobilizáveis. 5.3.7. Conclusões da Fase II As variedades escolhidas na primeira fase e analisadas mais detalhadamente na segunda fase apresentam, claramente, diferenças na resposta ao stresse hídrico. A variedade PB269 apresenta valores mais reduzidos de rigidez da parede celular e de THR no ponto de perda de turgescência, os quais permitem a manutenção do Ψw compatível com o metabolismo fotossintético do carbono. A sua diminuição é efectuada de uma forma progressiva, com um fecho dos estomas controlado, mantendo, desta forma, uma WUE mais elevada para THR mais baixos. Por outro lado, a variedade PB369 apresentou sempre limitações estomáticas mais elevadas e uma menor resposta estomática ao CO2, durante a recuperação. Esta variedade poderá também sofrer uma maior inibição do funcionamento do PSII com a imposição do stresse hídrico, que poderá ser consequência da degradação da proteína D1. Durante o período de stresse, PB269 apresentou alguma degradação da clorofila a, não recuperando o conteúdo desse pigmento após irrigação, o que poderá ter contribuído para a dificuldade em regressar aos valores iniciais de taxa fotossintética. No entanto, apresenta um mecanismo de manutenção dos valores de qN (em especial de qE) até níveis de stresse hídrico moderados. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 142 5. Fase II – Estudo Comparativo Aprofundado das Duas Variedades Mais e Menos Susceptíveis ao Stresse Hídrico Nas actividades da RuBisCO, observou-se uma diminuição progressiva, na variedade PB269, durante o stresse lento, em contraste com a diminuição de “tudo-ou-nada” da variedade PB369, já observada durante a primeira fase deste trabalho. Durante o stresse rápido, as variedades mantiveram as suas actividades constantes até THR de 50%, mas a variedade PB269 apresentou uma ligeira descida para valores de THR inferiores a este. Nas actividades da PEPC, os valores diminuíram durante o stresse lento, mas ficaram inalteradas durante o stresse rápido, um pouco à semelhança do que acontece com a actividade da EM-NADP. Relativamente à prolina, observou-se que ambas as variedades possuem uma reserva deste aminoácido, que parece ser rapidamente mobilizável, talvez como fonte de energia, pela variedade PB269, durante o stresse rápido e na recuperação do stresse lento. Os dados recolhidos permitem afirmar que PB269 é uma variedade de milho que tolera o stresse hídrico (stress endurer) mas que apresenta algumas dificuldades em recuperar os parâmetros hídricos. Por outro lado, a variedade PB369 é mais susceptível ao stresse hídrico, mas as plantas conseguem recuperar um pouco melhor os parâmetros hídricos, embora o metabolismo fotossintético do carbono fique mais comprometido do que na variedade PB269. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 143 Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 144 6. Conclusões Gerais 6. Conclusões Conclusões Gerais As variedades de milho analisadas apresentaram diferenças entre si, na forma como respondem ao stresse hídrico, comprovando que uma das dificuldades dos estudos de stresse reside na variabilidade inter-varietal das respostas. A esta acrescem, naturalmente, as que são recorrentemente referidas na bibliografia, nomeadamente o efeito da velocidade de imposição do stresse, as condições ambientais em que ocorre e a idade e espécie das plantas. As técnicas de determinação das trocas gasosas e de fluorescência modulada da clorofila a confirmaram a sua utilidade na identificação de diferenças intra-específicas. O stresse hídrico afecta fortemente os processos fotossintéticos, o crescimento e a sobrevivência das espécies vegetais. O conjunto de todos os processos, desde a captação da luz até à fixação do carbono, formam uma rede complexa de interacções, as quais serão afectadas pelo stresse hídrico em diferentes graus, contribuindo de formas diferentes para a redução global da fotossíntese. Nas variedades analisadas, observaram-se variações dos parâmetros hídricos, das trocas gasosas e dos parâmetros fotoquímicos, corroborando os resultados de outros ensaios efectuados noutras plantas sujeitas à seca. Das diferenças entre os dois sistemas de indução de stresse, destacam-se a rapidez com que os parâmetros se alteram no stresse rápido, enquanto no stresse lento variam mais suavemente. No entanto, as diferenças entre variedades em stresse rápido eram menos significativas do que em stresse lento, pelo que foi este último que permitiu a distinção de duas variedades com respostas qualitativamente diferentes ao stresse hídrico. A análise mais pormenorizada das variedades seleccionadas, PB269 como a mais tolerante e PB369 como a mais susceptível ao stresse hídrico, permitiu recolher dados sobre os diferentes mecanismos subjacentes. A elasticidade mais elevada da parede celular da variedade PB269 permite-lhe manter a turgescência até THR mais reduzidos, permitindo a manutenção do Ψw compatível com o metabolismo fotossintético do carbono, durante mais tempo, sendo uma estratégia importante na tolerância ao stresse hídrico. Por outro lado, a variedade PB369 apresenta limitações estomáticas mais elevadas na ausência de stresse mas, à medida que o stresse lento se vai intensificando, as limitações nãoestomáticas começam a desempenhar um papel importante nas duas variedades. Nas actividades da RuBisCO, observaram-se diferenças entre as duas variedades: uma diminuição progressiva, na variedade PB269, durante o stresse lento, semelhante ao observado nas plantas do grupo de resposta ao stresse do tipo I, em contraste com a diminuição de “tudo-ouIndução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 145 6. Conclusões Gerais nada” da variedade PB369, que passa de um patamar de valores para outro, semelhante ao observado nas plantas do grupo de resposta ao stresse do tipo II (Lawlor e Cornic, 2002). No entanto, no stresse rápido, as variedades mantiveram as suas actividades constantes até THR de 50%. Nas actividades da PEPC, os valores diminuíram durante o stresse lento, mas ficaram inalterados durante o stresse rápido, tal como acontece com a actividade da EM-NADP. Desta forma, durante a indução lenta do stresse hídrico, a diminuição da fotossíntese, em stresse moderado, deve-se, essencialmente, a limitações estomáticas, enquanto que, a partir de stresse intenso, tomam lugar as limitações não-estomáticas. Também durante a imposição do stresse hídrico ocorreram variações nos parâmetros da fluorescência da clorofila a, de forma mais suave durante o stresse lento e mais marcadas durante o stresse rápido. Estes dados, conjuntamente com os dados das actividades enzimáticas, indicam alterações na estrutura fotoquímica e enzimática, durante o stresse lento, e uma maior incidência na fase fotoquímica, durante o stresse rápido. Tal facto poderá estar relacionado com mecanismos de protecção induzidos ao longo da imposição do stresse lento, ao contrário do que acontece no stresse rápido, em que não existe uma janela temporal suficiente para desencadear esses mecanismos. Destaca-se PB269, com uma manutenção de qE até THR de cerca de 50%, que poderá traduzir-se num mecanismo de tolerância a níveis de stresse hídrico moderados, sem activação de processos de amortecimento não-fotoquímicos. Também se observou, em PB269, alguma degradação da clorofila a, não recuperando após irrigação, e a existência de uma reserva de prolina rapidamente mobilizável, talvez como fonte de energia, durante o stresse rápido e na recuperação do stresse lento. O conjunto de todos estes dados permitem afirmar que PB269 é uma variedade de milho que tolera o stresse hídrico (stress endurer) mas que apresenta algumas dificuldades em recuperar. Por outro lado, a variedade PB369 é mais susceptível ao stresse hídrico, mas as plantas conseguem recuperar um pouco melhor os parâmetros hídricos, embora o metabolismo fotossintético do carbono fique comprometido. O rápido desenvolvimento e melhoramento de plantas agrícolas, como o milho, é um dos objectivos a atingir, recorrendo a técnicas de Fisiologia Vegetal, de modo a compreender as respostas da planta ao stresse e, ulteriormente, isolarmos os factores responsáveis. Os conhecimentos adquiridos neste trabalho fornecem dados consistentes com os observados noutros trabalhos, reforçando ideias que se têm vindo a estabelecer sobre o efeito do stresse hídrico no processo fotossintético, fornecendo pistas para compreender as respostas das plantas e as formas de enfrentarem este problema. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 146 7. Referências Bibliográficas 7. Referências Bibliográficas Bibliográficas Abraham, E.M., Huang, B., Bonos, S.A., Meyer, W.A. (2004). Evaluation of drought resistance for Texas Bluegrass, Kentucky Bluegrass, and their hybrids. Crop Science 44: 1746–1753. Alberte, R.S., Thornber, J.P. (1977). Water stress effects on the content and organization of chlorophyll in mesophyll and bundle sheath chloroplasts of maize. Plant Physiology 59: 351-353. Andrade, F.H., Vega, C., Uhart, S., Cirilo, A., Cantarero, M., Valentinuz, O. (1999). Kernel number determination in maize. Crop Science 39: 453-459. Aphalo, P.J., Sánchez, R.A. (1986). Stomatal responses to light and drought stress in variegated leaves of Hedera helix. Plant Physiology 81: 768-773. Arad, S., Richmond, A.E. (1976). Leaf cell water and enzyme activity. Plant Physiology 57: 656-658. Arakawa, T., Timasheff, S.N. (1985). The stabilization of proteins by osmolytes. Biophysical Journal 47: 411414. Araus, J.L., Slafer, G.A., Reynolds, M.P., Royo, C. (2002). Plant breeding and drought in C3 cereals: What should we breed for? Annals of Botany 89: 925-940. Asner, G.P., Nepstad, D., Cardinot, G., Ray, D. (2004). Drought stress and carbon uptake in an Amazon forest measured with spaceborne imaging spectroscopy. PNAS 101 (16): 6039-6044. Augé, R.M., Hickok, L.G., Stodola, A.J.W. (1989). Psychrometric pressure-volume analysis of osmoregulation in roots, shoots, and whole sporophytes of salinized Ceratopteris. Plant Physiology 91: 322-330. Bailey, K.J., Battistelli, A., Dever, L.V., Lea, P.J., Leegood, R.C. (2000). Control of C4 photosynthesis: effects of reduced activities of phosphoenolpyruvate carboxylase on CO2 assimilation in Amaranthus edulis L.. Journal of Experimental Botany 51: 339-346. Baker, N.R., Rosenqvist, E. (2004). Applications of chlorophyll fluorescence can improve crop production strategies: an examination of future possibilities. Journal of Experimental Botany 55 (403): 1607-1621. Bänziger, M., Edmeades, G.O., Lafitte, H.R. (1997). Selection for drought tolerance increases maize yields across a range of nitrogen levels. Crop Science 39: 1035-1040. Bartholomew, D.M., Bartley, G.E., Scolnik, P.A. (1991). Abscisic acid control of rbcS and cab transcription in tomato leaves. Plant Physiology 96: 291-296. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 147 7. Referências Bibliográficas Beardsell, M.F., Cohen, D. (1975). Relationships between leaf water status, abscisic acid levels, and stomatal resistance in maize and sorghum. Plant Physiology 56: 207-212. Björkman, O., Demmig-Adams, B. (1995). Regulation of photosynthetic light energy capture, conversion, and dissipation in leaves of higher plants. In: Ecophysiology of Photosynthesis. E.-D. Schulze, M.M. Caldwell (eds.), Springer, Germany, págs. 17-47. Bohnert, H.J., Nelson, D.E., Jensen, R.G. (1995). Adaptations to environmental stresses. The Plant Cell 7: 1099-1111. Bohnert, H.J., Shen, B. (1999). Transformation and compatible solutes. Scientia Horticulturae 78: 237-260. Borel, C., Audran, C., Frey, A., Marion-Poll, A., Tardieu, F., Simonneau, T. (2001). N. plumbaginifolia zeaxanthin epoxidase transgenic lines have unaltered baseline ABA accumulations in roots and xylem sap, but contrasting sensitivities of ABA accumulation to water deficit. Journal of Experimental Botany 52: 427-434. Bradford, M. (1976). A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Annals of Biochemistry 72: 248-254. Bruce, W.B., Edmeades, G.O., Barker, T.C. (2002). Molecular and physiological approaches to maize improvement for drought tolerance. Journal of Experimental Botany 53 (366): 13-25. Buchanan-Wollaston, V. (1997). The molecular biology of leaf senescence. Journal of Experimental Botany 48 (307): 181-199. Burghardt, M., Riederer, M. (2003). Ecophysiological relevance of cuticular transpiration of deciduous and evergreen plants in relation to stomatal closure and leaf water potential. Journal of Experimental Botany 54 (389): 1941-1949. Casas, A.M., Kononowicz, A.K., Zehr, U.B., Tomes, D.T., Axtell, J.D., Butlers, L.G., Bressan, R.A., Hasegawa, P.M. (1993). Transgenic sorghum plants via microprojectile bombardment. PNAS 90: 1121211216. Casper, B.B., Forseth, I.N., Kempenich, H., Seltzer, S., Xavier, K. (2001). Drought prolongs leaf life span in the herbaceous desert perennial Cryptantha flava. Functional Ecology 15: 740–747. Catsky, J. (1960). Determination of water deficit in discs cut out from leaf blades. Biologia Plantarum 2: 76-77. Chapman, S.C., Edmeades, G.O. (1999). Selection improves drought tolerance in tropical maize populations: II. Direct and correlated responses among secondary traits. Crop Science 39: 1315-1324. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 148 7. Referências Bibliográficas Chaves, M.M., Pereira, J.S., Maroco, J., Rodrigues, M.L., Ricardo, C.P.P., Osório, M.L., Carvalho, I., Faria, T., Pinheiro, C. (2002). How plants cope with water stress in the field. Photosynthesis and growth. Annals of Botany 89: 907-916. Chaves, M.M., Maroco, J.P., Pereira, J.S. (2003). Understanding plant responses to drought – From genes to the whole plant. Functional Plant Biology 30: 239-264. Chaves, M.M., Oliveira, M.M. (2004). Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture. Journal of Experimental Botany 55 (407): 2365-2384. CIMMYT (2001). CIMMYT 1999–2000 World Maize Facts and Trends. Meeting World Maize Needs: Technological Opportunities and Priorities for the Public Sector. Pingali, P.L. (ed.), México. Cleland, D., Krader, P., McCree, C., Tang, J., Emerson, D. (2004). Glycine betaine as a cryoprotectant for prokaryotes. Journal of Microbiological Methods 58 (1): 31-38. Collinson, S.T., Clawson, E.J., Azam-Ali, S.N., Black, C.R. (1997). Effects of soil moisture deficits on the water relations of bambara groundnut (Vigna subterranea L. Verdc.). Journal of Experimental Botany 48 (309): 877-884. Cornic, G. (2000). Drought stress inhibits photosynthesis by decreasing stomatal aperture – not by affecting ATP synthesis. Trends in Plant Science 5 (5): 187-188. Cornic, G., Fresneau, C. (2002). Photosynthetic carbon reduction and carbon oxidation cycles are the main electron sinks for photosystem II activity during mild drought. Annals of Botany 89: 887-894. Costa, J.B. (1985). Caracterização e constituição do solo. Fundação Calouste Gulbenkian, Lisboa. Crafts-Brandner, S.J., Salvucci, M.E. (2000). Rubisco activase constraints the photosynthetic potential of leaves at high temperature and CO2. PNAS 97 (24): 13430-13435. Crafts-Brandner, S.J., Salvucci, M.E. (2002). Sensitivity of photosynthesis in a C4 plant, maize, to heat stress. Plant Physiology 129: 1773-1780. DaCosta, M., Wang, Z., Huang, B. (2004). Physiological adaptation of Kentucky Bluegrass to localized soil drying. Crop Science 44: 1307-1314. Dai, N., Schaffer, A., Petreikov, M., Shahak, Y., Giller, Y., Ratner, K., Levine, A., Granot, D. (1999). Overexpression of Arabidopsis hexokinase in tomato plants inhibits growth, reduces photosynthesis, and induces rapid senescence. Plant Cell 11: 1253-1266. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 149 7. Referências Bibliográficas Dallmier, K.A., Stewart, C.R. (1992). Effect of exogenous abscisic acid on proline dehydrogenase activity in maize (Zea mays L). Plant Physiology 99: 762-764. Delauney, A.J., Verma, D.P.S. (1993). Proline biosynthesis and osmoregulation in plants. The Plant Journal 4 (2): 215-223. Doncaster, H.D., Leegood, R.C. (1987). Regulation of phosphoenolpyruvate carboxylase activity in maize leaves. Plant Physiology 84: 82-87. Donkin, M., Martin, S.E. (1980). Studies on the properties of carboxylating enzymes in the epidermis of Commelia communis. Journal of Experimental Botany 31: 357-363. Duvick, D.N. (2001). Biotechnology in the 1930s: The development of hybrid maize. Nature Reviews Genetics 2: 69-74. Edmeades, G. O., Bolaños, J., Chapman, S.C., Lafitte, H.R., Bänziger, M. (1999). Selection improves drought tolerance in tropical maize populations: I. Gains in biomass, grain yield, and harvest index. Crop Science 39: 1306-1315. Edwards, G., Jenkins, C., Andrews, J. (1988). CO2 assimilation and activities of photosynthetic enzymes in high chlorophyll fluorescence mutants of maize having low levels of ribulose 1,5-bisphosphate carboxilase. Plant Physiology 86: 533-539. Edwards, G.E., Franceschi, V.R., Ku, M.S.B., Voznesenskaya, E.V., Pyankov, V.I., Andreo, C.S. (2001). Compartimentation of photosynthesis in cells and tissues of C4 plants. Journal of Experimental Botany 52 (356): 577-590. Ehleringer, J.R., Cerling, T.E. (2002). C3 and C4 Photosynthesis. In: Encyclopedia of Global Environmental Change – Volume 2, The Earth system: biological and ecological dimensions of global environmental change. H.A. Mooney, J.G. Canadell, T. Munn (eds). John Wiley & Sons, Ltd., págs. 186-190. Enquist, B.J., Niklas, K.J. (2002). Global allocation rules for patterns of biomass partitioning in seed plants. Science 295: 1517-1520. Flexas, J., Medrano, H. (2002). Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Annals of Botany 89: 183-189. Floyd, N., Horst, W.J., Puigdomenech, P., Riveill, N. (2002). Maize. [online]: http://www.ienica.net/crops/maize.pdf. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 150 7. Referências Bibliográficas Fracheboud, Y., Haldimann, P., Leipner, J., Stamp, P. (1999). Chlorophyll fluorescence as a selection tool for cold tolerance of photosynthesis in maize (Zea mays L.). Journal of Experimental Botany 50 (338): 15331540. Franks, P.J., Farquhar, G.D. (2001). The effect of exogenous abscisic acid on stomatal development, stomatal mechanics, and leaf gas exchange in Tradescantia virginiana. Plant Physiology 125: 935-942. Furbank, R.T., Chitty, J.A., Caemmerer, S., Jenkins, C.L.J. (1996). Antisense RNA inhibition of rbcS gene expression reduces Rubisco level and photosynthesis in the C4 plant Flaveria bidentis. Plant Physiology 111: 725-734. Gallais, A., Hirel, B. (2004). An approach to the genetics of nitrogen use efficiency in maize. Journal of Experimental Botany 55 (396): 295-306. Gebbing, T., Schyder, H. (1999). Pre-anthesis reserve utilization for protein and carbohydrate in grains of wheat. Plant Physiology 121: 871-878. Geber, M.A., Dawson, T.E. (1997). Genetic variation in stomatal and biochemical limitations to photosynthesis in the annual plant, Polygonum arenastrum. Oecologia 109: 535-546. Genty, B., Briantais, J.-M., Baker, N.R. (1989). The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta 990: 87-92. Ghannoum, O., Conroy, J.P., Driscoll, S.P., Paul, M.J., Foyer, C.H., Lawlor, D.W. (2003). Nonstomatal limitations are responsible for drought-induced photosynthetic inhibition in four C4 grasses. New Phytologist 159: 599-608. Gigon, A., Matos, A.-R., Laffray, D., Zuily-Fodil, Y., Pham-Thi, A.-T. (2004). Effect of drought stress on lipid metabolism in the leaves of Arabidopsis thaliana (ecotype Columbia). Annals of Botany 94 (3): 345-351. Giles, K.L., Beardsell, M.F., Cohen, D. (1974). Cellular and ultrastructural changes in mesophyll and bundle sheath cells of maize in response to water stress. Plant Physiology 54: 208-212. Giménez, C., Mitchell, V.J., Lawlor, D.W. (1992). Regulation of photosynthetic rate of two sunflower hybrids under water stress. Plant Physiology 98: 516-524. Goodger, J.Q.D., Sharp, R.E., Marsh, E.L., Schachtman, D.P. (2005). Relationships between xylem sap constituents and leaf conductance of well-watered and water-stressed maize across three xylem sap sampling techniques. Journal of Experimental Botany 56 (419): 2389-2400. Griffiths, H., Parry, M.A.J. (2002). Plant responses to water stress. Annals of Botany 89: 801-802. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 151 7. Referências Bibliográficas Gunasekera, D., Berkowitz, G.A. (1993). Use of transgenic plants with ribulose-1,5-bisphosphate carboxylase/oxygenase antisense DNA to evaluate the rate limitation of photosynthesis under water stress. Plant Physiology 103: 629-635. Hammond, E.T., Andrews, T.J., Woodrow, I.E. (1998). Regulation of ribulose-1,5-bisphosphate carboxylase/oxygenase by carbamylation and 2-carboxyarabinitol 1-phosphate in tobacco: insights from studies of antisense plants containing reduced amounts of rubisco activase. Plant Physiology 118: 1463-1471. Hanson, A.D., May, A.M., Grumet, R., Bode, J., Jamieson, G.C., Rhodes, D. (1985). Betaine synthesis in chenopods: Localization in chloroplasts. PNAS 82: 3678-3682. Hare, P.D., Cress, W.A., van Staden, J. (1999). Proline synthesis and degradation: a model system for elucidating stress-related signal transduction. Journal of Experimental Botany 50 (333): 413-434. Haupt-Herting, S., Fock, H.P. (2002). Oxygen exchange in relation to carbon assimilation in water-stressed leaves during photosynthesis. Annals of Botany 89: 851-859. Haupt-Herting, S., Klug, K., Fock, H.P. (2001). A new approach to measure gross CO2 fluxes in leaves. Gross CO2 assimilation, photorespiration, and mitochondrial respiration in the light in tomato under drought stress. Plant Physiology 126: 388-396. Havaux, M. (1992). Stress tolerance of photosystem II in vivo: Antagonistic effects of water, heat, and photoinhibition stresses. Plant Physiology 100: 424-432. Heuer, B. (2003). Influence of exougenous application of proline and glycinebetaine on growth of salt-stressed tomato plants. Plant Science 165: 693-699. Holbrook, G.P., Campbell, W.J., Rowland-Bamford, A., Bowes, G. (1994). Intraspecific variation in the light/dark modulation of ribulose 1,5-bisphosphate carboxylase-oxygenase activity in soybean. Journal of Experimental Botany 45 (8): 1119-1126. Holmström, K.-O., Somersalo, S., Mandal, A., Palva, T.E., Welin, B. (2000). Improved tolerance to salinity and low temperature in transgenic tobacco producing glycine betaine. Journal of Experimental Botany 51 (343): 177-185. Horton, P., Ruban, A.V., Walters, R.G. (1994). Regulation of light harvesting in green plants: Indication by nonphotochemical quenching of chlorophyll fluorescence. Plant Physiology 106: 415-420. Hunt, S. (2003). Measurements of photosynthesis and respiration in plants. Physiologia Plantarum 117: 314325. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 152 7. Referências Bibliográficas Hymus, G.J., Ellsworth, D.S., Baker, N.R., Long, S.P. (1999). Does free-air carbon dioxide enrichment affect energy use by evergreen trees in different seasons? A chlorophyll fluorescence study of mature loblolly pine. Plant Physiology 120: 1183-1191. Iturbe-Ormaetxe, I., Escuredo, P.R., Arrese-Igor, C., Becana, M. (1998). Oxidative damage in pea plants exposed to water deficit or paraquat. Plant Physiology 116: 173-181. Ivandic, V., Hackett, C.A., Zhang, Z.J., Staub, J.E., Nevo, E., Thomas, W.T.B., Forster, B.P. (2000). Phenotypic responses of wild barley to experimentally imposed water stress. Journal of Experimental Botany 51 (353): 2021-2029. Jackson, M.B. (2002). Long-distance signalling from roots to shoots assessed: the flooding story. Journal of Experimental Botany 53: 175-181. Jeanneau, M., Vidal, J., Gousset-Dupont, A., Lebouteiller, B., Hodges, M., Gerentes, D., Perez, P. (2002). Manipulating PEPC levels in plants. Journal of Experimental Botany 53 (376): 1837-1845. Jefferies, R.L., Rudmik, T., Dillon, E.M. (1979). Responses of halophytes to high salinities and low water potentials. Plant Physiology 64: 989-994. Jiang, M., Zhang, J. (2002). Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. Journal of Experimental Botany 53 (379): 2401-2410. Jiao, J., Chollet, R. (1992). Light activation of maize phosphoenolpyruvate carboxylase protein-serine kinase activity is inhibited by mesophyll and bundle sheath-directed photosynthesis inhibitors. Plant Physiology 98: 152-156. Jones, H.G. (1985). Partitioning stomatal and non-stomatal limitations to photosynthesis. Plant, Cell and Environment 8: 95-104. Jones, H.G. (1998). Stomatal control of photosynthesis and transpiration. Journal of Experimental Botany 49: 387-398. Kanechi, M., Uchida, N., Yasuda, T., Yamaguchi, T. (1996). Non-stomatal inhibition associated with inactivation of Rubisco in dehydrated coffee leaves under unshaded and shaded conditions. Plant and Cell Physiology 37 (4): 455-460. Kang, S., Zhang, J. (2004). Controlled alternate partial root-zone irrigation: its physiological consequences and impact on water use efficiency. Journal of Experimental Botany 55 (407): 2437–2446. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 153 7. Referências Bibliográficas Khan, S., Andralojc, P.J., Lea, P.J., Parry, M.A.J. (1999). 2'-Carboxy-D-arabitinol 1-phosphate protects ribulose 1,5-bisphosphate carboxylase/oxygenase against proteolytic breakdown. European Journal of Biochemistry 266: 840-847. Kim, J.-Y., Mahé, A., Brangeon, J., Prioul, J.-L. (2000). A maize vacuolar invertase, IVR2, is induced by water stress. Organ/tissue specificity and diurnal modulation of expression. Plant Physiology 124: 71-84. Kim, S.-H., Lieth, J.H. (2003). A coupled model of photosynthesis, stomatal conductance and transpiration for a rose leaf (Rosa hybrida L.). Annals of Botany 91: 771-781. Kingston-Smith, A.H., Foyer, C.H. (2000). Bundle sheath proteins are more sensitive to oxidative damage than those of the mesophyll in maize leaves exposed to paraquat or low temperatures. Journal of Experimental Botany 51 (342): 123-130. Kishor, P.B.K., Hong, Z., Miao, G.-H., Hu, C.-A.A., Verma, D.P.S. (1995). Overexpression of ∆1-pyrroline5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiology 108: 1387-1394. Knight, H., Trewavas, A.J., Knight, M.R. (1997). Calcium signalling in Arabidopsis thaliana responding to drought and salinity. The Plant Journal 12 (5): 1067-1078. Kramer, P.J., Boyer, J.S. (1995). Transpiration and the ascent of sap. In: Water relations of plants and soils, 2ª edição, Academic Press, California, págs. 201-256. Laisk, A., Eichelmann, H., Oja, V., Rasulov, B., Rämma, H. (2006). Photosystem II cycle and alternative electron flow in leaves. Plant and Cell Physiology 47 (7): 972-983. Lal, A., Edwards, G.E. (1996). Analysis of inhibition of photosynthesis under water stress in the C4 species Amaranthus creuntus and Zea mays: Electron transport, CO2 fixation and carboxylation capacity. Australian Journal of Plant Physiology 23: 403-412. Lamberts, H., Chapin III, F.S., Pons, T.L. (1998). Plant water relations. In: Plant Physiological Ecology, 1ª edição, Springer-Verlag Berlin Heidelberg, New York. Laporte, M.M., Shen, B., Tarczynski, M.C. (2002). Engineering for drought avoidance: expression of maize NADP-malic enzyme in tobacco results in altered stomatal function. Journal of Experimental Botany 53 (369): 699-705. Larher, F.R., Aziz, A., Gibon, Y., Trotel-Aziz, P., Sulpice, R., Bouchereau, A. (2003). An assessment of the physiological properties of the so-called compatible solutes using in vitro experiments with leaf discs. Plant Physiology and Biochemistry 41: 657-666. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 154 7. Referências Bibliográficas Larkindale, J., Knight, M.R. (2002). Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiology 128: 682-695. Lawlor, D.W. (2002). Limitation to photosynthesis in water-stressed leaves: Stomata vs. metabolism and the role of ATP. Annals of Botany 89: 871-885. Lawlor, D.W., Cornic, G. (2002). Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant, Cell and Environment 25: 275-294. Lazár, D. (1999). Chlorophyll a fluorescence induction. Biochimica et Biophysica Acta 1412: 1-28. Leakey, A.D.B., Uribelarrea, M., Ainsworth, E.A., Naidu, S.L., Rogers, A., Ort, D.R., Long, S.P. (2006). Photosynthesis, productivity, and yield of maize are not affected by open-air elevation of CO2 concentration in the absence of drought. Plant Physiology 140: 779-790. Lichtenthaler, H.K. (1987). Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods in Enzymology 148: 350-382. Lin, C.C., Kao, C.H. (2001). Regulation of ammonium-induced proline accumulation in detached rice leaves. Plant Growth Regulation 35 (1): 69-74. Lizana, C., Wentworth, M., Martinez, J.P., Villegas, D., Meneses, R., Murchie, E.H., Pastenes, C., Lercari, B., Vernieri, P., Horton, P., Pinto, M. (2006). Differential adaptation of two varieties of common bean to abiotic stress: I. Effects of drought on yield and photosynthesis. Journal of Experimental Botany 57 (3): 685-697. Loggini, B., Scartazza, A., Brugnoli, E., Navari-Izzo, F. (1999). Antioxidative defense system, pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physiology 119: 1091-1099. Long, S.P., Hallgren, J.-E. (1985). Measurements of CO2 assimilation by plants in field and laboratory. In: Techniques in bioproductivity and photosynthesis. J. Coombs, D.O. Hall, S.P. Long, J.M.O. Scurlock (eds.), 2nd ed., Pergamon Press, New York. Long, S.P., Hallgren, J.-E. (1993). Measurements of CO2 assimilation by plants in field and laboratory. In: Photosynthesis and production in a changing environment – a field and laboratory manual. D.O. Hall, J.M.O. Scurlock, H.R. Bolhar-Nordenkampf, R.C. Leegood, S.P. Long (eds.), Chapman & Hall, London. Louis, P., Trüper, H.G., Galinski, E.A. (1994). Survival of Escherichia coli during drying and storage in the presence of compatible solutes. Applied Microbiology and Biotechnology 41 (6): 684-688. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 155 7. Referências Bibliográficas Lucas, P.W., Turner, I.M., Dominy, N.J., Yamashita, N. (2000). Mechanical defences to herbivory. Annals of Botany 86: 913-920. Ludlow, A.E. (1991). Ochna pulchra Hook: Leaf growth and development related to photosynthetic activity. Annals of Botany 68: 527-540. Lüttge, U. (2004). Ecophysiology of Crassulacean acid metabolism (CAM). Annals of Botany 93: 629-652. Manetas, Y., Petropoulou, Y., Karabourniotis, G. (1986). Compatible solutes and their effects on phosphoenolpyruvate carboxylase of C4-halophytes. Plant, Cell and Environment 9: 145-151. Manter, D.K., Kerrigan, J. (2004). A/Ci curve analysis across a range of woody plant species: influence of regression analysis parameters and mesophyll conductance. Journal of Experimental Botany 55 (408): 25812588. Maroco, J., Ku, M., Lea, P., Dever, L., Leegood, R., Furbank, R., Edwards, G. (1998). Oxygen requirement and inhibition of C4 photosynthesis – An analysis of C4 plants deficient in the C3 and the C4 cycles. Plant Physiology 116: 823-832. Maroco, J.P., Edwards, G.E., Ku, M.S.B. (1999). Photosynthetic acclimation of maize to growth under elevated levels of carbon dioxide. Planta 210: 115-125. Maroco, J.P., Rodrigues, M.L., Lopes, C., Chaves, M.M. (2002). Limitations to leaf photosynthesis in grapevine under drought – Metabolic and modelling approaches. Functional Plant Biology 29: 1-9. Marques da Silva, J. (1999). Aspectos da fotossíntese de Setaria sphacelata var. splendida (Stapf) Clayton em condições de deficit hídrico. Tese de Doutoramento, Faculdade de Ciências da Universidade de Lisboa. Marques da Silva, J., Arrabaça, M.C. (2004a). Contributions of soluble carbohydrates to the osmotic adjustment in the C4 grass Setaria sphacelata: A comparison between rapidly and slowly imposed water stresse. Journal of Plant Physiology 161: 551-555. Marques da Silva, J., Arrabaça, M.C. (2004b). Photosynthetic enzymes of the C4 grass Setaria sphacelata under water stress: a comparison between rapidly and slowly imposed water deficit. Photosynthetica 42 (1): 4347. Marshall, J.G., Dumbroff, E.B. (1999). Turgor regulation via cell wall adjustment in white spruce. Plant Physiology 119: 313-319. Matos, M. (1990). A economia da água da macieira – Aspectos fisiológicos e aplicação ao controlo da rega. Tese de Doutoramento, Faculdade de Ciências da Universidade de Lisboa. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 156 7. Referências Bibliográficas Matsubara, S., Chow, W.S. (2004). Populations of photoinactivated photosystem II reaction centers characterized by chlorophyll a fluorescence lifetime in vivo. PNAS 101 (52): 18234-18239. Mattos, E.A., Herzog, B., Lüttge, U. (1999). Chlorophyll fluorescence during CAM-phases in Clusia minor L. under drought stress. Journal of Experimental Botany 50 (331): 253-261. Maxwell, K., Borland, A.M., Haslam, R.P., Helliker, B.R., Roberts, A., Griffiths, H. (1999). Modulation of Rubisco activity during the diurnal phases of the crassulacean acid metabolism plant Kalanchoë daigremontiana. Plant Physiology 121: 849–856. Maxwell, K., Johnson, G.N. (2000). Chlorophyll fluorescence – A pratical guide. Journal of Experimental Botany 51 (345): 659-668. Medrano, H., Escalona, J.M., Bota, J., Gulías, J., Flexas, J. (2002). Regulation of photosynthesis of C3 plants in response to progressive drought: stomatal conductance as a reference parameter. Annals of Botany 89: 895905. Meyer, S., Genty, B. (1998). Mapping intercellular CO2 mole fraction (Ci) in Rosa rubiginosa leaves fed with abscisic acid by using chlorophyll fluorescence imaging. Plant Physiology 116: 947-957. Modarres, A.M., Hamilton, R.I., Dijak, M., Dwyer, L.M., Stewart, D.W., Mather, D.E., Smith, D.L. (1998). Plant population density effects on maize inbred lines grown in short-season environments. Crop Science 38: 104-108. Müller, P., Li, X.-P., Niyogi, K.K. (2001). Non-photochemical quenching. A response to excess light energy. Plant Physiology 125: 1558-1566. Nayyar, H. (2003). Accumulation of osmolytes and osmotic adjustment in water-stressed wheat (Triticum aestivum) and maize (Zea mays) as affected by calcium and its antagonists. Environmental and Experimental Botany 50: 253-264. Niu, X., Bressan, R.A., Hasegawa, P.M., Pardo, J.M. (1995). Ion homeostasis in NaCl stress environments. Plant Physiology 109: 735-742. Nogués, S., Baker, N.R. (2000). Effects of drought on photosynthesis in Mediterranean plants grown under enhanced UV-B radiation. Journal of Experimental Botany 51 (348): 1309-1317. Ögren, E. (1990). Evaluation of chlorophyll fluorescence as a probe for drought stress in willow leaves. Plant Physiology 93: 1280-1285. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 157 7. Referências Bibliográficas Parry, M.A.J., Andralojc, P.J., Parmar, S., Keys, A.J., Habash, D., Paul, M.J., Alred, R., Quick, W.P., Servaites, J.C. (1997). Regulation of Rubisco by inhibitors in the light. Plant, Cell and Environment 20: 528– 534. Parry, M.A.J., Andralojc, P.J., Khan, S., Lea, P.J., Keys, A.J. (2002). Rubisco activity: effects of drought stress. Annals of Botany 89: 833-839. Perchorowicz, J.T., Raynes, D.A., Jensen, R.G. (1981). Light limitation of photosynthesis and activation of ribulose bisphosphate carboxylase in wheat seedlings. PNAS 78 (5): 2985-2989. Perchorowicz, J.T., Raynes, D.A., Jensen, R.G. (1982). Measurement and preservation of the in vivo activation of ribulose 1,5-bisphosphate carboxilase in leaf extracts. Plant Physiology 69: 1165-1168. Pieters, A.J., Tezara, W., Herrera, A. (2003). Operation of the xanthophyll cycle and degradation of D1 protein in the inducible CAM plant, Talinum triangulare, under water deficit. Annals of Botany 92: 393-399. Pinheiro, H.A., DaMatta, F.M., Chaves, A.R.M., Loureiro, M.E., Ducatti, C. (2005). Drought tolerance is associated with rooting depth and stomatal control of water use in clones of Coffea canephora. Annals of Botany 96: 101-108. Platt, T., Gallegos, C.L., Harrison, W.G. (1980). Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. Journal of Marine Research 38: 687-701. Poorter, H., Nagel, O. (2000). The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review. Australian Journal of Plant Physiology 27: 595-607. Price, A.H., Cairns, J.E., Horton, P., Jones, H.G., Griffiths, H. (2002). Linking drought-resistance mechanisms to drought avoidance in upland rice using a QTL approach: progress and new opportunities to integrate stomatal and mesophyll responses. Journal of Experimental Botany 53 (371): 989-1004. Quick, W.P., Stitt, M. (1989). An examination of factors contributing to non-photochemical quenching of chlorophyll fluorescence in barley leaves. Biochimica et Biophysica Acta 977: 287-296. Raschke, K. (1970). Stomatal responses to pressure changes and interruptions in the water supply of detached leaves of Zea mays L.. Plant Physiology 45: 415-423. Raymond, M.J., Smirnoff, N. (2002). Proline metabolism and transport in maize seedlings at low water potential. Annals of Botany 89: 813-823. Richards, R.A., Rebetzke, G.J., Condon, A.G., van Herwaarden, A.F. (2002). Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Science 42: 111-121. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 158 7. Referências Bibliográficas Richardson, S.G., McCree, K.J. (1985). Carbon balance and water relations of sorghum exposed to salt and water stress. Plant Physiology 79: 1015-1020. Rizhsky, L., Liang, H., Mittler, R. (2002). The combined effect of drought stress and heat shock on gene expression in tobacco. Plant Physiology 130: 1143-1151. Rodrigues, M.L., Pacheco, C.M.A., Chaves, M.M. (1995). Soil-plant water relations, root distribution and biomass partitioning in Lupinus albus L. under drought conditions. Journal of Experimental Botany 46 (8): 947956. Rodriguez, M.M., Heyser, J.W. (1988). Growth inhibition by exogenous proline and its metabolism in saltgrass (Distichlis spicata) suspension cultures. Plant Cell Reports 7 (5): 305-308. Roitsch, T., Balibrea, M.E., Hofmann, M., Proels, R., Sinha, A.K. (2003). Extracellular invertase: key metabolic enzyme and PR protein. Journal of Experimental Botany 54 (382): 513-524. Saccardy, K., Cornic, G., Brulfer, J., Reyss, A. (1996). Effect of drought stress on net CO2 uptake by Zea mays L. cv. Adonis. Planta 199: 589-595. Sakamoto, A., Murata, N. (2000). Genetic engineering of glycinebetaine synthesis in plants: current status and implications for enhancement of stress tolerance. Journal of Experimental Botany 51 (342): 81-88. Schmidt, C.N.G., Gutteride, S., Parry, M.A.J., Keys, A.J. (1984). Inactive forms of wheat ribulose bisphosphate carboxilase. Biochemichal Journal 220: 781-785. Schurr, U., Heckenberger, U., Herdel, K., Walter, A., Feil, R. (2000). Leaf development in Ricinus communis during drought stress: dynamics of growth processes, of cellular structure and of sink–source transition. Journal of Experimental Botany 51 (350): 1515-1529. Schwartz, S.H., Tan, B.C., Gage, D.A., Zeevaart, J.A.D., McCarty, D.R. (1997). Specific oxidative cleavage of carotenoids by VP14 of maize. Science 276: 1872-1874. Servaites, J.C., Parry, M.A.J., Gutteridge, S., Keys, A.J. (1986). Species variation in the predawn inhibition of ribulose-1,5-bisphosphate carboxylase/oxygenase. Plant Physiology 82: 1161-1163. Sharp, R.E., LeNoble, M.E. (2002). ABA, ethylene and the control of shoot and root growth under water stress. Journal of Experimental Botany 53 (366): 33-37. Sharp, R.E., Poroyko, V., Hejlek, L.G., Spollen, W.G., Springer, G.K., Bohnert, H.J., Nguyen, H.T. (2004). Root growth maintenance during water deficits: physiology to functional genomics. Journal of Experimental Botany 55 (407): 2343-2351. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 159 7. Referências Bibliográficas Shen, B., Jensen, R.G., Bohnert, H.J. (1997a). Increased resistance to oxidative stress in transgenic plants by targeting manitol biosynthesis to chloroplasts. Plant Physiology 113: 1177-1183. Shen, B., Jensen, R.G., Bohnert, H.J. (1997b). Mannitol protects against oxidation by hydroxyl radicals. Plant Physiology 115: 527-532. Siddique, M.R.B., Hamid, A., Islam, M.S. (1999). Drought stress effects on photosynthetic rate and leaf gas exchange of wheat. Botanical Bulletin of Academia Sinica 40: 141-145. Simon, P. (1987). Different effects of chilling on the activity of C4 enzymes in two ecotypes of Echinocloa crusgalli from sites of contrasting climates. Physiologia Plantarum 69: 205-210. Smeekens, S., Rook, F. (1997). Sugar sensing and sugar-mediated signal transduction in plants. Plant Physiology 115: 7-13. Solomon, M., Belenghi, B., Delledonne, M., Menachem, E., Levine, A. (1999). The involvement of cysteine proteases and protease inhibitor genes in the regulation of programmed cell death in plants. Plant Cell 11: 431444. Steudle, E. (2000). Water uptake by roots: effects of water deficit. Journal of Experimental Botany 51 (350): 1531-1542. Steudle, E., Peterson, C.A. (1998). How does water get through roots? Journal of Experimental Botany 49 (322): 775-788. Stoop, J.M.H., Williamson, J.D., Pharr., D.M. (1996). Mannitol metabolism in plants: a method for coping with stress. Trends in Plant Science 1 (5): 139-144. Stuhlfauth, T., Sultemeyer, D.F., Weinz, S., Fock, H.P. (1988). Fluorescence quenching and gas exchange in a water stressed C3 plant, Digitalis lanata. Plant Physiology 86: 246-250. Syngenta (2004). Milho. [online]: http://www.syngenta.pt/producao_desc.asp?culturaID=3. Taiz, L., Zeiger, E. (2002). Plant Physiology, 3ª edição, Sinauer Associates, Inc.. Tallman, G. (2004). Are diurnal patterns of stomatal movement the result of alternating metabolism of endogenous guard cell ABA and accumulation of ABA delivered to the apoplast around guard cells by transpiration? Journal of Experimental Botany 55 (405): 1963-1976. Tang, A.-C., Boyer, J.S. (2002). Growth-induced water potentials and the growth of maize leaves. Journal of Experimental Botany 53 (368): 489-503. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 160 7. Referências Bibliográficas Tang, A.-C., Boyer, J.S. (2003). Root pressurization affects growth-induced water potentials and growth in dehydrated maize leaves. Journal of Experimental Botany 54 (392): 2479-2488. Tang, A.-C., Kawamitsu, Y., Kanechi, M., Boyer, J.S. (2002). Photosynthetic oxygen evolution at low water potential in leaf discs lacking an epidermis. Annals of Botany 89: 861-870. Tezara, W., Mitchell, V.J., Driscoll, S.D., Lawlor, D.W. (1999). Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 401: 914-917. Tezara, W., Martínez, D., Rengifo, E., Herrera, A. (2003). Photosynthetic responses of the tropical spiny shrub Lycium nodosum (Solanaceae) to drought, soil salinity and saline spray. Annals of Botany 92: 757-765. Ting, I.P., Osmond, C.B. (1973). Photosynthetic phosphoenolpyruvate carboxylases – Characteristics of alloenzymes from leaves of C3 and C4 plants. Plant Physiology 51: 439-447. Tollenaar, M., Wu, J. (1999). Yield improvement in temperate maize is attributable to greater stress tolerance. Crop Science 39: 1597-1604. Torrecilla, I., Leganés, F., Bonilla, I., Fernández-Piñas, F. (2000). Use of recombinant aequorin to study calcium homeostasis and monitor calcium transients in response to heat and cold shock in cyanobacteria. Plant Physiology 123: 161-175. Tuberosa, R., Salvi, S., Sanguineti, M.C., Landi, P., Maccaferri, M., Conti, S. (2002). Mapping QTLs regulating morpho-physiological traits and yield: Case studies, shortcomings and perspectives in droughtstressed maize. Annals of Botany 89: 941-963. Turner, N.C. (1974). Stomatal behavior and water status of maize, sorghum, and tobacco under field conditions: II. At low soil water potential. Plant Physiology 53: 360-365. van Wijk, K.J., Nilsson, L.O., Styring, S. (1994). Synthesis of reaction center proteins and reactivation of redox components during repair of photosystem II after light-induced inactivation. The Journal of Biological Chemistry 269 (45): 28382-28392. Vermaas, W., Madsen, C., Yu, J., Visser, J., Metz, J., Nixon, P.J., Diner, B. (1995). Turnover of the D1 protein and of Photosystem II in a Synechocystis 6803 mutant lacking Tyrz. Photosynthesis Research 45 (2): 99104. Vijn, I., Smeekens, S. (1999). Fructan: More than a reserve carbohydrate? Plant Physiology 120: 351-359. von Caemmerer, S., Farquhar, G.D. (1981). Some relationships between the biochemistry and the gas exchange of leaves. Planta 153: 376-387. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 161 7. Referências Bibliográficas Vu, J.C.V., Allen, L.H., Bowes, G. (1987). Drought stress and elevated CO2 effects on soybean ribulose bisphosphate carboxylase activity and canopy photosynthetic rates. Plant Physiology 83: 573-578. Walker, R.P., Chen, Z.-H., Acheson, R.M., Leegood, R.C. (2002). Effects of phosphorylation on phosphoenolpyruvate carboxykinase from the C4 plant Guinea Grass. Plant Physiology 128: 165-172. Wei, C., Tyree, M.T., Steudle, E. (1999). Direct measurement of xylem pressure in leaves of intact maize plants. A test of the cohesion-tension theory taking hydraulic architecture into consideration. Plant Physiology 121: 1191-1205. Werker, E. (2000). Trichome diversity and development. Advances in Botanical Research 31: 1-35. Wilson, K.B., Baldocchi, D.D., Hanson, P.J. (2000). Quantifying stomatal and non-stomatal limitations to carbon assimilation resulting from leaf aging and drought in mature deciduous tree species. Tree Physiology 20: 787-797. Wingler, A., Walker, R.P., Chen, Z.-H., Leegood, R.C. (1999). Phosphoenolpyruvate carboxykinase is involved in the decarboxylation of aspartate in the bundle sheath of maize. Plant Physiology 120: 539-545. Xin, Z., Li, P.H. (1993). Relationship between proline and abscisic acid in the induction of chilling tolerance in maize suspension-cultured cells. Plant Physiology 103: 607-613. Xiong, L., Schumaker, K.S., Zhu, J.-K. (2002). Cell signaling during cold, drought, and salt stress. The Plant Cell 14: S165-S183. Yang, J., Zhang, J., Huang, Z., Zhu, Q., Wang, L. (2000). Remobilization of carbon reserves is improved by controlled soil-drying during grain filling of wheat. Crop Science 40: 1645-1655. Ying, J., Lee, E.A., Tollenaar, M. (2002). Response of leaf photosynthesis during the grain-filling period of maize to duration of cold exposure, acclimation, and incident PPFD. Crop Science 42: 1164-1172. Yordanov, I., Velikova, V., Tsonev, T. (2003). Plant responses to drought and stress tolerance. Bulgarian Journal of Plant Physiology (Special Issue): 187-206. Young, A.J., Phillipa, D., Rubanb, A.V., Hortonb, P., Frank, H.A. (1997). The xanthophyll cycle and carotenoid-mediated dissipation of excess excitation energy in photosynthesis. Pure and Applied Chemistry 69 (10): 2125-2130. Zhang, N., Portis, A.R. (1999). Mechanism of light regulation of Rubisco: A specific role for the larger Rubisco activase isoform involving reductive activation by thioredoxin-f. PNAS 96: 9438–9443. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 162 7. Referências Bibliográficas Živković, T., Quartacci, M.F., Navari-Izzo, F. (2005). Low-molecular weight substances in the poikilohydric plant Romanda serbica during dehydration and rehydration. Plant Science 168: 105-111. Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 163 Indução e Recuperação do Stresse Hídrico em Variedades Portuguesas de Milho 164
Baixar