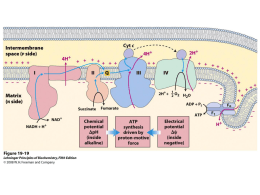
Uso efetivo da irrigação no mamoeiro (Carica papaya L): aspectos fisiológicos e produtividade Prof. Eliemar Campostrini Plant Physiology Lab Northern Rio de Janeiro State University Campos dos Goytacazes, RJ Brazil www.uenf.br [email protected] ORCID number: 0000-0002-1329-1084 Research ID C-4917-2013 DAT 0 FI PRD RDI NI DAT 6 DAT 14 My Lab A ação humana tem influenciado significativamente a atmosfera durante a era industrial -uso de combustíveis fósseis (CO2, NO) -Desmatamento (CO2) -Agricultura (CO2 e NO produzido pelas industrias de fertilizantes e CH4 pela fermentação entérica de ruminantes, cultivo de arroz inundado, aterros sanitarios) O que está provocando a escassez de água no planeta -A partir da era pré-industrial até 2009, a ação antropogênica causou um incremento na concentração de CO2 de 280 para 390 µL L-1, e a concentração deste gás atingiu o valor de 400 µL L-1 em 2013. Nesta condição, observa-se um elevação de 2 µL L-1 por ano (Rodrigues et al., 2015). A partir desta informação, espera-se que em 2100 a concentração de CO2 no ar possa atingir em torno de 800 ou 1150L L-1 O que está provocando a escassez de água no planeta -A partir da era pré-industrial até 2009, a ação antropogênica causou um incremento na concentração de CO2 de 280 para 390 µL L-1, e a concentração deste gás atingiu o valor de 400 µL L-1 em 2013. Nesta condição, observa-se um elevação de 2 µL L-1 por ano (Rodrigues et al., 2015). Segundo o IPCC (2014), esta elevação na concentração pode causar um incremento na temperatura do ar em torno de 3,7 a 4,8ºC Tal elevação pode provocar extremos na variação de pressão do ar, com consequentes alterações no deslocamento de massa de ar, provocando zonas de extrema escassez de água e locais com chuvas de elevadas intensidades Segundo o IPCC (2014), esta elevação na concentração pode causar um incremento na temperatura do ar em torno de 3,7 a 4,8ºC Tal elevação pode provocar extremos na variação de pressão do ar, com consequentes alterações no deslocamento de massa de ar, provocando zonas de extrema escassez de água e locais com chuvas de elevadas intensidades TORONTO, Canadá (Thomson Reuters Foundation) - Novos dados de satélite mostram que a seca no Brasil é pior do que se pensava, com o Sudeste perdendo 56 trilhões de litros de água em cada um dos últimos três anos A pior seca do país nos últimos 35 anos também tem levado o Nordeste brasileiro, região maior, mas menos povoada, a perder 49 trilhões de litros de água a cada ano nos últimos três anos http://br.reuters.com/article/topNews/idBRKCN0SO2P220151030 -Semi-lenhosa (uma posição intermediária entre uma herbácea e uma árvore) -Precocidade de produção (3-4 meses) -Elevada taxa fotossintética(≈26 mol m-2 s-1 – field condition in 2000 mol m-2 s-1 PAR) -café: 8- 9 mol m-2 s-1 -milho: 40-50 mol m-2 s-1 -feijão: 12 mol m-2 s-1 -arroz: 13 mol m-2 s-1 -cana de açúcar: 40-50 mol m-2 s-1 -crescimento rápido -produção de várias sementes ‘Solo’: 270 sementes por fruto (5g) ‘Formosa’ : 350 sementes por fruto (17g) -Reduzido custo de construção do tronco -Produção contínua de frutos (crescimento indeterminado) Porque a água é importante para o mamoeiro -estrutura semilenhosa (biologia) -constante produção de frutos (agronomia)/Produtor rural Tronco - C. papaya possui uma anatomia diferencial - Densidade do tronco é somente 0.13 g cm3 -eucalipto: 1 g cm3 -brauna: 1.16 g cm3 -angico: 0.78 g cm3 -Apesar de não ser uma planta lenhosa, o mamoeiro pode crescer até 10 m de altura, -Cada planta do grupo ‘Solo’ pode produzir 50 frutos por planta com 0.5kg cada fruto (25kg total de fruto). Grupo ‘Formosa’ pode produzir 25 frutos por planta com 1.5kg cada fruto (Total de suporte de peso: 45kg) Como pode explicar esta sustentação se a planta não é lenhosa! Explicação para compensar a falta de lenho: 1) Pressão de turgencencia nas células do parênquima 2) Fibras lignificadas do floema em forma de rede (parte externa do tronco) ou a combinação dos dois fatores Cada planta com 20cm de diâmetro!! ‘Solo’ pode suportar um peso total 25kg “Formosa’ 45kg The construction of fibers and high-pressure matrix parenchyma is unique in tree-like plants, and guarantees adequate flexural rigidity with a minimum fibre content!! A pressão de turgência foi medida em mamoeiro e ficou entre 0.82 e 1.25 MPa, O que indicou que esta pressão é essencial para rigidez do tronco!!!! Kempe et al (2014) Um pneu de carro cheio tem uma pressão de 0.2 MPa A água que dentro de um cano numa casa tem em torno de 0.2-0.3 MPa Tronco -Para compensar o tronco oco, na base deste tronco não existe o espaço vazio. -Ainda nesta base da planta, os internós são pouco espaçados entre eles (compactos!) Uma estratégia para suportar mecanicamente o peso dos frutos e da planta!! - O xilema é pobremente lignificado (Fisher 1980 ) -Este xilema é composto de vasos largos e pode ser visto a olho nu. Estes vasos estão embebidos em um parênquima não lignificado Xylem vessel O mamoeiro apresenta uma alta relação área foliar/área do xilema 0,10m2 de área foliar por cada cm2 de área ativa do xilema A planta de papaya perde 460 mL de H2O por cada cm2 de xilema ativo Isto faz com que a planta apresenta uma maior sensibildade ao grau de secura do ar ( maior sensibilidade ao déficit de pressãod e vapor do ar (DPVar) Ao que tudo indica a planta de mamoeiro possui uma resistência hidráulica elevada no continum raiz-caule-folha-atmosfera Possivelmente o caule seria o ponto limitante no movimento de água mamoeiro Como a falta de água afeta o mamoeiro? -O mamoeiro é sensível a falta de água no ar e no solo Valor SPAD 41,2 Valor SPAD 39,8 Valor SPAD 26,6 Valor SPAD 6,6 Falta de água no ar! Alte temperatura do ar e baixa umidade relativa elevam o DPVair DPVair = 0,61137 * exp (17,502 * T° / 240,97 + T°) * (1,0 – (UR% / 100)) Valores máximos de DPV para o papaya: Elevado DPVair e DPVleaf-air reduz gs DPVair = < 1kPa (es air – eair) DPVleaf-air= < 2kPa (es leaf –eair) DPVleaf-air 10 9 d i ffe r e n c e s (k P a ) L e a f-to -a i r v a p o r p r e ssu r e O decréscimo de A após a saturação luminosa é devido em parte ao decréscimo na condutãncia estomática (gs) por meio do aquecimento da folha causado pela elevada energia luminosa (limitação estomática) 8 7 6 5 4 3 2 1 0 0 500 1000 1500 2000 2500 3000 P h o to sy n th e ti c a l l y a c ti v e r a d i a ti o n ( m o l m -1 -2 Tleaf s ) PPF (mol m-2 s-1) The photosynthetic response of papaya is strongly linked to environmental conditions through stomatal behavior soil field capacity High light (2200 µmol m-2 s-1) High leaf temperature (40ºC) High VPD (≈ 6 kPa) Low g (0.10 mol m-2 s-1) s Low A (5 µmol m-2 s-1) Midday depression of photosynthesis can reduce yield soil field capacity Papaya leaves showed paraheliotropic movement associated with reduced leaf turgor (Reis 2008). Paraheliotropic movement can be translated into decreased radiation load per unit leaf area which, in turn, could prevent the photoinhibition Falta de água no solo! Ψsoil = - 20 kPa Ψsoil = - 68 kPa Em relação ao déficit hídrico no solo, o mamoeiro mostra uma limitação estomática e nãoestomática Como relação a limitação estomática, esta limitação pode ser controlada por natureza hidráulica e não-hidraulica Valor SPAD 41,2 Valor SPAD 39,8 Valor SPAD 26,6 Valor SPAD 6,6 Baixinho de Santa Amália genotype 70L pots Greenhouse Maximum PAR 1200µmol m-2 s-1 Severe water stress: ψleaf =-0,8MPa Regular irrigated plants: ψleaf =-0,6MPa Estratégia para tentar escapar do estresse hídrico sem afetar a produtividade -Elevar o uso efetivo de água! H 2O evaporação H2O a) Estratégias práticas de manejo: 1) Evitar vazamento em tubulações 0.6 day-1 2) Aplicação de água no horário de maior transpiração 10 L H2 O plant-1 day-1 Summer aplicou 10L dia-1 no solo aplicou 16L dia-1 no solo The crop was irrigated with a drip/fertigation system providing supplemental irrigation of 10 (winter) and 16 L per plant per day (summer) 0.4 0.3 0.2 0.1 Summer Winter 0 8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 4.5 -0.1 Summer Hour Winter 4 77,09 g CO2 plant-1 day-1 Winter 3.5 CO 2 Assimilation (g m-2 h-1 ) Transpirtion (L m-2 h-1 ) 0.5 -1 L H2 O plant Luz15 Winter 3 2.5 2 1.5 1 50,74 g CO2 plant-1 day-1 Summer 0.5 0 8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 -0.5 Hour 3) Uso de cobertura plástica sobre o solo Na Austrália, 40% da disponibilidade total da água no solo é perdida por evaporação! 4) Remoção de folhas mais velhas 4) Remoção de folhas mais velhas Em geral, cada folha madura pode prover fotoasimilados para 3 frutos (Zhou et al., 2000) A fotossíntese pode afetar a qualidade dos frutos (Salazar, 1978). Remoção de 75% das folhas reduziu significativamente a produção de novas flores, o pegamento de frutos, e decresceu o teor de solidos soluveis (TSS) dos frutos 50% de remoção das folhas não reduziu o pegamento de frutos ou TSS (Zhou et al, 2000) In general: (Brazil) Solo group: < leaf area High yield: 95 t ha-1 95 fruits 1 leaf support 4 fruits 24 leaves Average yield : 61 t ha-1 62 fruits 1 leaf support 3 fruits 19 leaves Formosa group: > leaf area High yield: 150 t ha-1 73 fruits 1 leaf support 3 fruits 25 leaves Average yield : 85 t ha-1 42 fruits 1 leaf support 2 fruits 21 leaves 4) Uso de irrigação de subsuperfície 5) Técnicas alternativas de irrigação: -Irrigação parcial do sistema radicular e deficit de irrigação controlado -Estas técnicas podem aumentar a “pegada hídrica” Sai cerca de 2000 a 3000 µmol H2O m-2 s-1 e entra cerca de 20 a 30 µmol CO2 m-2 s-1 Transpiration (mmol m-2 s-1) 12 Planta C3 y = 10.871x + 1.6225 R² = 0.722 RDI y = 12.69x + 0.72 R² = 0.5369 FI y = 11.092x + 1.4987 R² = 0.7368 PRD 14 10 y = 10.543x + 0.7411 R² = 0.715 NI 8 6 4 FI 2 0 0 0,2 0,4 0,6 0,8 Stomatal conductance (mol m-2 s-1) 1 NI PRD 14 dias após a aplicaçao dos tratamentos -maximo estresse gs (mol H2O m-2 s-1) 0,8 CIFI 0,7 PRD IPSR RDI 0,6 NI 0,5 0,4 0,3 0,2 0,1 0 0 3 6 9 12 14 16 Days after treatment 17 21 0,6 gs (mol m-2 s-1) 0,5 VPD 0,4 0,3 0,2 0,1 0 DAT CI IPSR RDI NI 14 High VPD 12 Transpiration (mmol m-2 s-1) Low VPD High VPD 12 FI CI PRD IPSR 10 y = 10.871x + 1.6225 R² = 0.722 RDI y = 12.69x + 0.72 R² = 0.5369 FI y = 11.092x + 1.4987 R² = 0.7368 PRD 10 y = 10.543x + 0.7411 R² = 0.715 NI 8 6 4 FI PRD RDI NI E (mmolH2O m-2 s-1) RDI 2 NI 8 0 0 6 0,2 0,4 0,6 Stomatal conductance (mol m-2 s-1) 4 2 0 0 3 6 9 12 14 16 Days after treatment 17 21 0,8 1 Condition de campo July 80 60 50 70% 40 1nd gas exchange measurements 30 20 10 70% 0 3 days after rainfall 13 days after rainfall Rainfall (mm) Rainfall (mm) 70 80 70 60 50 40 30 20 10 0 October 2nd gas exchange measurements root dead (7 days without irrigation) young root 5 days with irrigation after 7 days without irrigation FI NI FI Dry side of PRD 5) Melhoramento genético a) Busca de genótipos com maior agressividade de raiz (maior aprofundamento do sistema radicular) -Raízes com maior condutividade hidráulica Root 70 a 75% in 30cm 1 year old ≈ 3.5 m 5) Melhoramento genético b) Busca de genótipos de mamoeiro com menor transpiração noturna c) Genótipos com maior espessura de cutícula d) Genótipos com maior capacidade de ajustamento osmótico Conclusão: -A água é um fator do ambiente extremamente importante para o mamoeiro, uma vez que esta planta não possui uma estrutura anatômica completa para caracterizá-la como lenhosa. -Tal característica anatômica da planta, associada à produção contínua dos frutos, bem como a grande sensibilidade da assimilação fotossintética do carbono à limitação hídrica do solo e do ar faz que com esta molécula seja importante para a cultura - Devido a crise hídrica, torna-se de fundamental importãncia usar estratégias de manejo e ações no melhoramento genético da planta para elevar o uso efetivo de água e assim tentar manter a taxa fotossintética e a produtividade com menos água aplicada na planta - My team: Bárbara dos Santos Esteves (Pos Doc) José Altino Machado Filho (PhD student) Weverton Pereira Rodrigues (PhD student) Luciene de Sousa Ferreira (PhD student) Jefferson Ranjel da Silva (PhD student) Katherine Fraga Ruas (PhD student) Emile Caroline Silva Lopes (MSc student) Deivisson Peregrino de Abreu (undergraduate) Luan Baritiello da Silva Bezerra (undergraduate) Leticia Cespom (undergraduate) Wallace Bernado (undergraduate) Raynan de Souza Aguliar (undergraduate) Colaboração Projects: Ecophysiology of tropical tree crops -Effects of enviromental factors (light, water, mineral nutrients) on photosynthesis, water use efficiency and yield in papaya, coffee, grape, sugarcane, soybean Whole-canopy gas exchange in coffee Projects: Ecophysiology of tropical tree crops -Effects of enviromental factors (light, water, mineral nutrients) on photosynthesis, water use efficiency and yield in papaya, coffee, grape, soybean Whole-canopy gas exchange in papaya Partial rootzone drying in papaya Partial rootzone drying in sugarcane -Hydraulic conductivity and gas exchange in coffee, eucaliptus and soybean Root hydraulic conductivity in different coffee clones -Ecophysiology of photoautotrophic micropropagation in papaya and banana grown in bioreactors -Ecophysiology of photoautotrophic micropropagation in papaya and banana grown in bioreactors -Photoautotrophic micropropagation in papaya -Photoautotrophic micropropagation in papaya -Photoautotrophic micropropagation in papaya 400 ppm CO2 5000 ppm CO2 Papaya (Carica papaya L.) is a principal horticultural crop of tropical and subtropical regions (Campostrini et al, 2010). Papayas are produced in about 60 countries, with the bulk of production occurring in developing economies! Indonesia Peru Thailand Democratic Republic of the Congo Mexico Faostat 2015: Area harvested in the world: 441,042 ha Brazil: 31,989 ha (7.3%) Bangladesh Brazil Nigeria Asia is the largest papaya-producing continent, providing 55.5% of the total production, followed by South America (23.0%) and Africa (13.2%) India 0 40000 80000 120000 Area harvested (ha) 160000 Venezuela (Bolivarian Republic of) Thailand Democratic Republic of the Congo Dominican Republic Mexico Nigeria Indonesia Production in the world: 12,420,584 tonnes Brazil:1,582, 638 ton (13%) Brazil India 0 2000000 4000000 Production (ton) 6000000 Papaya (Carica papaya L.) is a principal horticultural crop of tropical and subtropical regions. Faostat 2015: Yield in the world: 28,161 kg ha-1 Brazil: 43,333 kg ha-1 (65% higher!) China, mainland China Chile Brazil Bolivia (Plurinational State of) Belize Bangladesh Bahamas Australia Argentina 14000000 Productin (tonnes) 12000000 0 y = 371.852x - 7E+08 R² = 0.9257 40000 80000 Yield kg ha-1 10000000 8000000 World 6000000 Brazil 4000000 2000000 y = 20.290x - 4E+07 R² = 0.2756 0 1996 1998 2000 2002 2004 2006 2008 2010 2012 2014 Year Faostat 2015: World-wide production of papaya has been increasing approximately 372,000 tonnes year−1(FAOSTAT, 2015) Brazil has been increasing 20.290 tonnes per year!! 120000 Gaining in popularity worldwide, papaya is now ranked third with 11.22 Mt, or 15.36 percent of the total tropical fruit production, behind mango with 38.6 Mt (52.86%) and pineapple with 19.41 Mt (26.58%) Total tropical fruit production: Mango – 52.86% Pineapple – 26.58% Papaya – 15.36% (Evans e Ballen, 2012) Papaya in Europe: -Tropical fruits are grown in very limited countries in Europe due to the climatic constraints -However, some of them are grown due to a favorable climate on the Mediterranean coast and Atlantic Islands for local consumption (Voth, 2000) -Papaya is cultivated under protected cultivation in subtropical country such as the Canary Islands (Spain) (Galan Sauco et al. 2008). -However, global climate changing should cause an increase in air temperature, and will be expected that cultivation of papaya will be expanded in the subtropical regions in Europe and Asia Morphological analysis and photosnthetic performance of improved papaya genotypes Brazilian Journal plant Physiology 21:209-222. 2009 5 genotypes: -Sunrise Solo -JS12 -Golden -Tainung 6L pots Greenhouse Maximum PAR 1000 µmol m-2 s-1 Temp max 32ºC Maximum yield 100 to 150 t ha-1 Maximum SPAD reading = 60 Sunrise Solo Portable Chlorophyll Measurement (PCM) Maximum yield 100 to 150 t ha-1 54 52 50 48 46 44 42 40 38 36 34 32 30 28 26 24 A A A A Golden Híbrido JS12 Solo 7212 Tainung A A A A B B A A A B B B B B 14 B B 28 35 B Maximum yield 90 t ha-1 B 21 42 49 56 63 Days after seedling (DAS) Golden Maximum SPAD reading = 36 Maximum yield 90 t ha-1 70 77 84 Sunrise Solo: -High yield -High soluble solids (more sugar) -450g weight -red pulp -green leaf (high chlorophyll concentration) -100 ton ha-1 Tainung: -High yield -High soluble solids (more sugar) -900 -1200 g weight -red/orange pulp -green leaf (high chlorophyll concentration) - High production (150 ton ha-1) First Brazilian Hybrid UENF/Caliman 01 Red pulp fruit weight 1200 g per fruit, fruit diameter 9,9 cm, Fruit length 21,5 cm Length/diameter 2,2, 10 g per 100 fresh seeds Others Hybrids: UENF/Caliman 02, 03, 04, 05, 06, 07 and 08 More informations: contact [email protected] Papaya in Spain www.freshfruitportal.com There are currently 310 planted hectares of papaya on the Canary Islands, the Spanish publication wrote. 90% are located in greenhouses on Tenerife and Gran Canaria. Production reaches around 16,000 tons (MT) a year. Yield: 51,600 kg ha-1 high yield!!! Yield in the world: 28,161 kg ha-1 Brazil: 43,333 kg ha-1 Almeria Papaya is starting!! Spain: Cajamar demonstrates viability of papaya cultivation in Almeria According to Juan Carlos, papaya plants develop very quickly and start producing eight to nine months after being planted; much earlier than most other fruit crops, which usually take four to six years. They are also very productive. Most papaya growers in the Iberian Peninsula introduced the fruit last year with small plantations (500 to 1,000 square metres), and they are only just harvesting their first papayas. "Some growers dared starting off with larger plantations and the crop's future is promising," stated Juan Carlos Gázquez. Source: Elalmeria.es 2/26/2014 Papaya commercialization significantly contributes to the economy of developing countries Papaya is considered one of the most important fruits because it is a rich source of antioxidant nutrients (e.g., carotenes, vitamin C, and flavonoids), the B vitamins (e.g., folate and pantothenic acid), minerals, and fiber. Papaya fruit is known for its high content of vitamins A, and C (approaches or exceeds USDA minimum daily requirements for adults) and is a good source of calcium, potassium. On an average, 100 g of ripe papaya contains 2500 I.U. of Vitamin A and 70 mg of vitamin C (ascorbic acid) In addition, papaya is a source of the digestive enzyme papain, which is used as an industrial ingredient in brewing, meat tenderizing, pharmaceuticals, beauty products, and cosmetics. Papain is a commercially valuable proteolytic enzyme that is produced in the milky latex of green, unripe papaya fruits (Dunne and Horgan, 1992). Evolutionarily, papain may be associated with protection from frugivorous (fruit-eating) predators and herbivores (El Moussaoui et al 2001). Papain yield: 245 -700 kg ha-1 ≈250 kg fruit produce 1kg crude papain!! Papain is a commercially valuable proteolytic enzyme that is produced in the milky latex of green, unripe papaya fruits (Dunne and Horgan, 1992). Evolutionarily, papain may be associated with protection from frugivorous (fruit-eating) predators and herbivores (El Moussaoui et al 2001). Papain yield: 245 -700 kg ha-1 ≈250 kg fruit produce 1kg crude papain!! Papain : is a cysteine protease E-64: a cysteine protease specific inhibitor Papaya plant -Semi-woody (an intermediate position between herbs and trees) -Latex-producing -Usually single-stemmed -Reproductive precocity (3-4 months) -High photosynthetic rates (≈26 mol m-2 s-1 - field condition in 2000 mol m-2 s-1 PAR) -fast growth -production of many seeds ‘Solo’ group: 270 seeds per fruit (5g) ‘Formosa’ group: 350 seeds per fruit (17g) -Low construction cost of hollow stems -Continuous fruit production (indeterminate growth) Papaya plant Root Root 70 a 75% in 30cm 1 year old ≈ 3.5 m Fruit -Papaya fruit ripening is climacteric, and high ethylene production may start only hours after harvest at the recommended stage (appearance of one to two yellow stripes on the fruit) physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent and lumpy pulp (Oliveira and Vitória 2011 ). Skin freckles Fruit -Papaya fruit ripening is climacteric, and high ethylene production may start only hours after harvest at the recommended stage (appearance of one to two yellow stripes on the fruit) Physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent and lumpy pulp (Oliveira and Vitória 2011 ). Skin freckles Fruit -Papaya fruit ripening is climacteric, and high ethylene production may start only hours after harvest at the recommended stage (appearance of one to two yellow stripes on the fruit) Physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent and lumpy pulp (Oliveira and Vitória 2011 ). High diurnal temperature variation induces latex exudation and increased variation in cell t pressure Fruit (Downton, 1981). Increased pressureisisclimacteric, expected with temperature -Papayaturgor fruit ripening andhigh highdiurnal ethylene production fluctuations. may start only hours after harvest at the recommended stage (appearance of one to two yellow Thestripes exuded on the fruit surface may dehydrate the cells around the exudate, thus form onlatex the fruit) different sized spots that are proportional to the volume of exuded latex (Reyes and Paull, 1994) Physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent and lumpy pulp (Oliveira and Vitória 2011 ). Fruit -Papaya fruit ripening is climacteric, and high ethylene production may start only hours after harvest at the recommended stage (appearance of one to two yellow stripes on the fruit) Physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent and lumpy pulp (Oliveira and Vitória 2011 ). 16 Fruit set2000 jan2001 Pedunc casca i casca e 7 14 -1 Teor de Cálcio(g.kg ) Amplitude Térmica( o 6 -Papaya fruit ripening is climacteric, and high ethylene production may start only 12 hours after harvest at the recommended stage5 (appearance of one to two yellow 10stripes on the fruit) 4 3 Physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent and lumpy pulp (Oliveira and Vitória 2011 ). 2 6 8 1 4 jun2000 Semana fev2001 0 set2000 época jan2001 Campostrini et al. (2005) proposed that the high calcium contents associated with high temp and large diurnal temperature fluctuations in September contributed to greater cell wall rigid facilitating increased turgor pressure resulting in the rupture of the latex vessels and latex le Potencial hídrico do solo média diária (kPa) Fruit 200 180 -Papaya fruit production may start only 160ripening is climacteric, and high ethylene113,6 hours after140 harvest at the recommended stage (appearance of one to two yellow 50,1 stripes on the 120 fruit) 100 Physiological80disorders of papaya fruits are bruising (“skin freckles”) and translucent and lumpy pulp 60 (Oliveira and Vitória 2011 ). 40 20 25/09/2003 08/09/2003 22/08/2003 05/08/2003 19/07/2003 04/07/2003 17/06/2003 31/05/2003 14/05/2003 27/04/2003 11/04/2003 25/03/2003 08/03/2003 19/02/2003 02/02/2003 16/01/2003 01/01/2003 15/12/2002 28/11/2002 11/11/2002 25/10/2002 08/10/2002 0 data Reis et al. (2003) demonstrated that in the months that preceded the greatest SF incidence, a severe soil water deficit (ψsoil mean of -113.6 kPa) in association with a minimal potential ET demand were the factors most related to SF incidence Fruit High density < water content -Papaya fruit ripening is climacteric, and high ethylene production may start only hours after harvest at the recommended stage (appearance of one to two yellow Reduced rigidity of stripes on the fruit) these cell wall Physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent Reduced efficiency of and lumpy pulp (Oliveira and Vitória 2011 ). the tonoplast in maintaining the turgor pressure in the cell in the tissue of pulp flesh translucency (PFT) Water did not enter the cells due to disruption of the membrane pumps that generate the proton gradient driving water permeability! In healthy fruit: the dry seed wei 6.24±0.88 g fruit-1 and the calcium concentration was 7.53±2.58 g kg PFT fruit: had a greater dry seed (8.02±1.75 g fruit-1) and the calci content was 9.0±1.04 g kg-1 Fruit High density -Papaya fruit ripening is climacteric, and high ethylene production may start only hours after harvest at the recommended stage (appearance of one to two yellow The mass of calcium extracted by th stripes on the fruit) with PFT was 72.18 g fruit-1, w calcium mass extracted from the Physiological disorders of papaya fruits are bruising (“skin freckles”) and translucent the healthy fruit was 46.98 g fruit-1 and lumpy pulp (Oliveira and Vitória 2011 ). More research is needed to unders role of calcium transport in papaya Leaves arranged in a spiral pattern > light interception > Number of sun leaves Male Hermaphrodite Female Leaf -plant produce large palmate leaves (≈0,6m2) -Five to nine pinnate lobes of various widths (40-60 cm) -Leaf epidermis and the palisade parenchyma are composed of a single cell layer, while spongy mesophyll consists of four to six layers of tissue Male Hermaphrodite Female Stomata -Papaya leaves are hypostomatics -stomatal density of sunlight leaves is 400 to 800 stomata mm-2 Male Hermaphrodite Female Male Hermaphrodite Female 1.60m Stem (trunk) -The single stem provides structural support -Stem diameters of adult plants vary from 10 to 30 cm at base to 5 -10 cm at the crown. Latex - Damage to any aerial part of the papaya plant, where laticifers are widely distributed, elicits latex release, which is very typical for this species (Azarkan et al. 2003 ) This milky latex is a slightly acidic fluid composed of 80 % water (Rodrigues et al. 2009 ). It contains sugars, starch grains, minerals (S, Mg, Ca, K, P, Fe, Zn), alkaloids, isoprenoids, lipidic substances, and proteins, including enzymes like lipases, cellulases, and cysteine proteases (papain, chymopapain), important in defense against insect herbivores (Sheldrake 1969 ; El Moussaoui et al. 2001 ; Azarkan et al. 2003 ; Konno et al. 2004 ) Ecophysiology of papaya (Carica papaya L) Prof. Eliemar Campostrini Plant Physiology Lab Northern Rio de Janeiro State University Campos dos Goytacazes, RJ Brazil www.uenf.br [email protected] ORCID number: 0000-0002-1329-1084 Research ID C-4917-2013 DAT 0 FI PRD RDI NI DAT 6 DAT 14 Stomata Single-leaf photosynthesis -Papaya leaves are hypostomatics -stomatal density of sunlight leaves is 400 to 800 stomata mm-2 Male Hermaphrodite Female Single-leaf photosynthesis Papaya is classified as a plant with C3 metabolism (Imai et al., 1982; Marler et al., 1994; Campostrini, 1997; Marler and Mickelbart, 1998; Jeyakumar et al., 2007) The absence of margin cell formation in the vascular bundles of papaya leaves (Buisson and Lee, 1993) is a characteristic associated with C3 metabolism C4 leaf Bundle sheath Single-leaf photosynthesis The average C13 value of C4 plants is -14‰ C3 plants is -23 to -36 ‰ Campostrini, 1997 Torres-Netto 2005 Single-leaf photosynthesis Ψsoil = - 20 kPa Ψsoil = - 68 kPa Marler e Mickelbart, 1998 Maximum net carbon assimilation (A) rates of 25 to 30 μmol m-2 s-1 are achieved at 2000 μmol m-2 s-1 photosynthetic photo flux (PPF) in soil field capacity (Marler and Mickelbart, 1998; Campostrini and Yamanishi, 2001; Reis, 2007) Morphological analysis and photosnthetic performance of improved papaya genotypes Brazilian Journal plant Physiology 21:209-222. 2009 6L pots Greenhouse Maximum PAR 1000 µmol m-2 s-1 Temp max 32ºC Maximum SPAD reading = 60 Sunrise Solo Maximum yield 100 to 150 t ha-1 Portable Chlorophyll Measurement (PCM) 5 genotypes: -Sunrise Solo -JS12 -Golden -Tainung 54 52 50 48 46 44 42 40 38 36 34 32 30 28 26 24 A A A A Golden Híbrido JS12 Solo 7212 Tainung A A A A B B A A A B B B B B 14 B B 28 35 B Maximum yield 90 t ha-1 B 21 42 49 56 63 Days after seedling (DAS) Golden Maximum SPAD reading = 36 70 77 84 CVL Days after seeding CVL Central vein length 40 38 A AB -2 SLA (g m ) 36 AB 34 AB 32 B 30 28 Golden Hibrido JS12 Solo 7212 Tainung SPAD reading = 60 0,22 Sunrise Solo 0,21 A A 2 Leaf area - LA (m ) 0,20 0,19 A SPAD reading = 30 A 0,18 A 0,17 Golden 0,16 0,15 Golden Hibrido JS12 Solo Tainung 24 15 14 A 22 SDM (g) 20 13 A A A A A 12 11 A 18 10 RDM (g) A 9 16 B 8 B 7 14 6 12 5 Golden Híbrido JS 12 Solo7212 Tainung Golden Híbrido JS 12 Solo7212 Tainung 3,0 0,012 A A 0,011 B 2,0 B B B 0,010 AB AB LAR SDM / RDM 2,5 AB 1,5 B 1,0 0,009 0,008 Golden Híbrido JS12 Solo7212 Tainung Golden Híbrido JS 12 Solo7212 Tainung 0,60 0,50 A A 0,58 0,48 A A A A 0,46 TMR 0,54 AB AB 0,52 0,44 B 0,50 B 0,42 0,48 0,46 0,40 Golden Híbrido JS12 Solo7212 Tainung Golden Híbrido JS12 Solo7212 Tainung Shoot dry mass (SDM), Root dry mass (RDM), Leaf area ratio (LAR)(leaf area/total dry mass), Leaf dry mass ratio (LMR)(total leaf dry mass/total dry mass), Trunk dry mass ratio (TMR)(trunk dry mass/total dry mass) and Shoot / root dry mass ratio (SDM/RDM) of Carica papaya L. plants with 92 days after seedling (DAS). Each column represents the average of 8 replicates. Average follow by the same letter doesn’t show statistic differences (Tukey 5%). 6L pots , greenhouse, maximum PAR 1000 µmol m-2 s-1 and max temp 32ºC LMR 0,56 68000 A 64000 However, Golden genotype had high daily carbon gain! AB -2 -1 Adaily ( mol m dia ) 66000 A 62000 The reduction of growth and yield in Golden papaya can be due: B B 60000 58000 56000 54000 Golden Híbrido JS12 Solo7212 Tainung -high photorespiration -high dark respiration In general, each mature leaf can provide photoassimilate for about three fruits (Zhou et al., 2000) The photosynthetic capacity also influences papaya fruit quality (Salazar, 1978). Defoliation by 75% significantly reduced new flower production and fruit set, decreased ripe fruit total soluble solids (TSS) 50% defoliation did not reduce new fruit set or ripe fruit TSS (Zhou et al, 2000) In general: (Brazil) Solo group: < leaf area High yield: 95 t ha-1 95 fruits 1 leaf support 4 fruits 24 leaves Average yield : 61 t ha-1 62 fruits 1 leaf support 3 fruits 19 leaves Formosa group: > leaf area High yield: 150 t ha-1 73 fruits 1 leaf support 3 fruits 25 leaves Average yield : 85 t ha-1 42 fruits 1 leaf support 2 fruits 21 leaves Increase net photosynthetic rate (A) is very important to high papaya production While high A rates are possible in papaya, environmental factors as PPF (light), water (soil and air), soil compaction, wind, VPD, temperature, soil oxygen, often limit A. 1) Light (PPF) Sun leaf Ψsoil = - 20 kPa Ψsoil = - 68 kPa With rapid reductions in irradiance, photosynthesis declines due to rapid biochemical adjustments. With the return to high irradiance, and in the absence of any stomatal lmitation, photosynthesis quickly increase Ligh compensation point in papay single leaf. Field condition 60 mol m-2 s-1 Dark respiration 1.75 to 2.5 mol CO2m-2 s-1 The PPF response of papaya may also decline with PPF above saturating levels 10 9 d i ffe r e n c e s (k P a ) L e a f-to -a i r v a p o r p r e ssu r e The decrease in A that begins at light saturation is due, in part, to the decrease in stomatal conductance (gs) through the direct action of radiant energy on leaf heating (stomatal limitation) 8 7 6 5 4 3 2 1 0 0 500 1000 1500 2000 2500 3000 P h o to sy n th e ti c a l l y a c ti v e r a d i a ti o n ( m o l m -1 -2 Tleaf s ) PPF (mol m-2 s-1) The photosynthetic response of papaya is strongly linked to environmental conditions through stomatal behavior soil field capacity High light (2200 µmol m-2 s-1) High leaf temperature (40ºC) High VPD (≈ 6 kPa) Low g (0.20 mol m-2 s-1) s Low A (5 µmol m-2 s-1) Midday depression of photosynthesis can reduce yield soil field capacity Papaya leaves showed paraheliotropic movement associated with reduced leaf turgor (Reis 2008). Paraheliotropic movement can be translated into decreased radiation load per unit leaf area which, in turn, could prevent the photoinhibition Tainung Cloudy days overcast Sunny days sunny day Red lady Although midday Fv/Fm of well-watered plants was sgnificantly lower than early morning Fv/Fm, the values of ≈0.75 are within the range of typical values and thus do not indicate the substantial decline in photochemical efficiency However, chronic photoinhibition also decreases A rates at light levels above saturation, in this case through damage and replacement of the D1 protein in the reaction center of PSII by excess PPF (Critchley, 1998)(non stomatal limitation) Sunrise Irradiance can control stomatal conductance and can reduce photosynthesis Papaya stomata are able to track rapid changes in irradiance!! (Clemente and Marler, 1996) Most plants experience continuous fluctuations in light under natural conditions rather that long periods of uniform irradiance!! Even individuals of pioneer and other species growing in open habitats experience high and low light periods in the order of several minutes each due to broken cumulus cloud cover (Knapp and smith, 1990). hour of the day 15:56 15:06 14:16 13:26 12:36 11:46 10:56 10:06 09:16 08:26 07:36 3,00 1500 1000 2,00 1,00 500 0,00 0 -1 PAR (mol m s ) 5,00 -2 PAR external air 06:46 05:56 05:06 04:16 03:26 02:36 01:46 00:56 00:06 VPD air (kPa) 6,00 2500 2000 4,00 2000 mol m-2 s-1 2000 mol m-2 s-1 320mol m-2 s-1 durante 3min 1600 mol m-2 s-1 250 mol m-2 s-1 durante 2.5 min 1600 mol m-2 s-1 3.6m x 1.5m = 5.4m2 field condition in Brazil Almeria greenhouse: 3.0m x 1.6m = 4.8 m2 The papaya photosynthetic capacity can be linked to non stomatal limitation in soil field capacity condition. As exemple: -Nitrogen leaf concentration 15 g kg-1DM 18 g kg-1DM 35 g kg-1DM 37 g kg-1DM 15 g kg-1DM 18 g kg-1DM 35 g kg-1DM 37 g kg-1DM http://www.specmeters.com/weathermonitoring/weather-stations/2000mini-stations/watchdog-2475-plantgrowth-station/ 1200 1000 y = 0,4365x R2 = 0,9986 W m -2 800 600 400 200 0 0 500 1000 1500 um ol m -2 s -1 2000 2500 3000 Whole-canopy photosynthesis 25.2 mol m-2 s-1 4.5 Summer Winter 4 77,09 g CO2 plant-1 day-1 Winter CO 2 Assimilation (g m-2 h-1 ) 3.5 3 2.5 2 1.5 1 50,74 g CO2 plant-1 day-1 Summer 0.5 0 8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 -0.5 Hour g CO2 m-2 h-1 to mol m-2 s-1 conversion factor is 6.30 The crop was irrigated with a drip/fertigation system providing supplemental irrigation of 10 (winter) and 16 L per plant per day (summer) Summer: (clear sky, during 4 days) PPFmax: 2400 mol m-2 s-1 Tmax: 38ºC VPDmax: 4 kPa Winter: (clear sky during 4 days) PPFmax: 1400 mol m-2 s-1 Tmax: 33ºC VPDmax: 3.5 kPa Each chamber had a volume of 3.4 m3 Under the environmental conditions evaluated : (4 sunny days) Winter: Maximum vapor pressure deficit (VPDair)=3.5 kPa Air maximum temperature of 33°C Maximum PPF: 1400 mol m-2 s-1 Summer Maximum VPDair=4.0 kPa Air maximum temperature of 38ºC Maximum PPF : 2400 mol m-2 s-1 Leaf area each plant 5 months old Winter :3.5m2 Summer: 4 m2 Tair inside ballon -Tair outside ºC Luz 44 42 40 38 Tleaf (°C) 36 34 32 30 28 Winter 26 Summer 24 22 20 08:0009:0010:0011:0012:0013:0014:0015:0016:0017:00 Time Air temperature Optimal temperature: 21 a 33ºC (Knight, 1980) 22 A 26ºC (Lassoudiere, 1968) 30ºC (Allan, 1978) Allan and Jager (1978) reported that A increased when air temperature rose from 16 to 30°C, and then A decreased linearly at temperatures above 30°C, the value at 41°C being half that at 30°C However, air temperature acts indirectly on papaya photosynthesis via increases in leaf-toair VPD! (stomatal limitation) When air temperature increased from 20° to 40°C for ‘Sunrise Solo’ papaya growing in Linhares, southeastern Brazil, the VPDleaf air increased from 2 to 6 kPa and A decreased from 20 to 5 μmol m-2 s-1 (Torres-Netto, 2000) Air temperature High air temperature and low Relative Humidity increase VPDair VPDair = 0,61137 * exp (17,502 * T° / 240,97 + T°) * (1,0 – (UR% / 100)) Threshold VPD values to papaya: VPDair = < 1kPa (esair – eair) VPDleaf-air= < 2kPa (esleaf –eair) High VPDair and VPDleaf-air reduce gs VPDleaf-air In addition, air temperature acts directly on papaya leaves photosynthesis via increases in leaf temperature (non stomatal limitation) Nonstomatal limitation: -inhibition of Rubisco, -heat stress results in a loss of the oxygen evolving complex activity (Enami et al., 1994; Yamane et al., 1998), -dissociation of the peripheral antenna complex of PSII from its core complex (Gounaris et al., 1984; Srivastava et al., 1997) -inhibition of electron transfer from primary/secondary electron-accepting plastoquinone of PSII at the acceptor side (Bukhov et al., 1990; Cao and Govindjee, 1990), High temperature increases the membrane fluidity and electron transport is blocked!! Than Fo increase Carica papaya is heat tolerant! Table 1. Maximum quantum efficiency of open Photosystem II reaction centers-quantum yield (Fv/Fm), photoinhibition (%), and canopy temperature (°C) in Golden papaya inside whole canopy chambers during four days of measurements during winter and summer. winter summer Fv/Fm 08:00 0.759 (539)x Aaz ± 0.004y 0.741(1000) Aaz ± 0.007y Fv/Fm 13:00 0.643 (1440) Ab ± 0.012 0.674 (2440) Ab ± 0.013 y Fv/Fm 17:00 0.764 (30) Aa ± 0.004 0.740 (200) Aa ± 0.008 y Photoinhibition (%) 13 A± 3.14 8.89 B ± 1.75 y Leaf temperature (ºC) Maximum Average Minimum 37.5 43.9 31.0 ± 1.12 36.95 ± 0.67 y 26.4 30 followed by the same letter are not significantly different using the Turkey’s mean separation test (P=0.05). Capital letters compare columns and lower case letters compare rows. yStandard error of the mean. xnumbers inside the parentheses represent the photosynthetic photon flux density (PPFD, µmol m-2 s-1) zMeans 0.6 -1 L H2 O plant Luz15 Winter day-1 10 L H2 O plant-1 day-1 Summer The crop was irrigated with a drip/fertigation system providing supplemental irrigation of 10 (winter) and 16 L per plant per day (summer) 0.4 0.3 0.2 0.1 Summer Winter 0 8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 4.5 -0.1 Summer Hour Winter 4 O-1 WUEwinter =77/15 = 5gCO2 kg H2 WUEsummer = 50/10 = 5gCO2 kg H2O-1 77,09 g CO2 plant-1 day-1 Winter 3.5 CO 2 Assimilation (g m-2 h-1 ) Transpirtion (L m-2 h-1 ) 0.5 3 2.5 2 1.5 1 50,74 g CO2 plant-1 day-1 Summer 0.5 0 8:00 9:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 -0.5 Hour Reduced whole-canopy photosynthesis and transpiration in papaya in the summer were due to high leaf temperatures (Tmax=43.9ºC)(due high radiation and high air temperature) and high VPDair (4.0kPa) that caused stomatal closure. However, high temperature did not affect photochemical efficiency (Fv/Fm ) when assessed by chlorophyll fluorescence (Fv/Fm ) 40 y = 2.1493x + 3.4249 R² = 0.7081 winter summer winter 35 Carbon dioxide assimilation (µ mol m-2 s-1 ) 30 25 20 15 10 y = 1.9155x + 2.1847 R² = 0.4961 summer 5 0 0 2 4 6 8 10 12 14 -5 -10 -15 Transpiration (mmol m-2 s-1 ) The water use of field-grown papaya plants during four days: Summer: 2.5 L H2O m-2 leaf area day-1 plant-1 Winter: 4.2 L H2O m-2 leaf area day-1 plant-1 16 Papaya (summer and winter) 2 µmol CO2 mmol H2O-1 4.8 g CO2 kg H2O-1 0.208 L H2O g CO2-1 0.0020 mol CO2 mol H2O-1 496 mol H2O-1 mol CO2 C3: 2.48 µmol CO2 mmol H2O-1 6.1 g CO2 kg H2O-1 0.164 to 0.5 L H2O g CO2-1 0.0025 mol CO2 mol H2O-1 400 mol H2O-1 mol CO2 C4: 2 to 5 g CO2 kg H2O-1 What strategies to reduce negative effects of high PPF on photosynthesis in summer Particle film What is a ‘Particle Film’? • A microscopic layer of mineral particles Particle Film Plant Surface • allows water and carbon dioxide to pass through the film Source: Dr. David Michael Glenn USDA, ARS, West Virginia, USA Stomates are not blocked After initial application After 24 hours Reflective surfaces are common plant adaptation • Plants “use” pubescence and cuticular waxes to reduce environmental stresses and reduce disease and insect damage • Particle film technology builds on this strategy of a reflective plant surface that repels insects Licania tomentosa 3% kaolin Source: Dr. David Michael Glenn USDA, ARS, West Virginia, USA 12% kaolin Source: Dr. David Michael Glenn USDA, ARS, West Virginia, USA Untreated control 26.0 C Infrared image of apple trees Surround treated 24.4 C 40 Temperature (Cº) 35 30 25 Surround WP control air temperature 20 15 900 1100 1300 1500 Hour 1700 1900 Source: Dr. David Michael Glenn USDA, ARS, West Virginia, USA 12% Surround Control 3% Surround Source: Dr. David Michael Glenn USDA, ARS, West Virginia, USA 15 11 Control irrigated #1 3% Surround irrigated 12% Surround irrigated control irrigated #2 3% Surround irrigated #2 12% Surround irrigated #2 9 7 5 3 1 -1 -3 900 1800 300 1200 2100 600 1500 2400 900 1800 300 2000 500 1400 2300 800 1700 200 1100 Photosynthesis (g CO2/m2/hr) 13 Irrigated September 13-20 2007 Hour In summer the use particle film (kaolin particles) applied in high light condition (clear sky) -Increase reflection of UV and IR -Reduce leaf temperature -Increase stomatal conductance -Increase whole-canopy photosynthesis and transpiration Concentration: 10%/12% 12kg = 38US$ Microspray irrigation applying above the canopy during summer (12h to 14h). Evaporative cooling reduce VPDleaf-air, increase stomatal conductance, increase transpiration and photosynthesis When the air temperature inside the canopy reached 35ºC the microspray irrigation system was turned on controle microaspersão 13,0 controle microaspersão transpiração (mmol m -2 s -1) déficit de pressão de vapor (kPa) 4,5 12,5 4,0 12,0 11,5 3,5 11,0 10,5 3,0 10,0 8:50 10:12 hora controle 23 12:39 microaspersão 0,50 10:12 hora controle 12:39 microaspersão condutância estomática (mol m-2 s -1) taxa fotossintética líquida ( mol m-2 s -1) 22 8:50 0,45 21 20 5 fruits plant-1 0,40 19 0,35 18 17 0,30 8:50 10:12 hora 12:39 Microspray irrigation increased 8:50 10:12 hora 12:39 Greenhouse can provides adequate environmental conditions to cultivate papaya. -protection against excessive light (sunburn, photoinhibition, high leaf temperature, high VPDleaf-air) -protection against wind and hail - Exclusion of PRV vector Baixinho de Santa Amália genotype Full sunlight ● Greenhouse E SOL SOL Full sunlightE ● Greenhouse 2000 1500 P P F 1000 winter 500 fluxo de fótons fotossintéticos (mol m-2 s-1) P P F fluxo de fótons fotossintéticos (mol m-2 s-1) 2000 09:15 10:22 11:46 winter summer 7 VPDleaf-air (kPa) 0900 h 1200 h 4 3 2 1 0 greenhouse full sunlight greenhouse full sunlight Treatments 9:44 hora 30% Light reduction 5 summer 500 8:09 13:45 hora 6 1000 0 0 8 1500 12:42 8 25 winter summer winter 7 5 20 0900 h 1200 h Rate of net photosynthesis (mol m-2 s-1) VPDleaf-air (kPa) 6 4 3 2 1 0 greenhouse full sunlight greenhouse full sunlight Stomatal conductance (mol m-2 s-1) 0,60 winter 0,50 0900 h 0,40 1200 h summer 0,30 0,20 0,10 0,00 greenhouse full sunlight Treatments 0900 h 1200 h 10 5 0 greenhouse full sunlight greenhouse full sunlight Treatments Treatments 0,70 15 summer greenhouse full sunlight 55 53 Spad reading 51 49 47 45 E greenhouse SOL full sunlight Full sunlight ● Greenhouse 320 300 0,80 260 Fv/Fm 0,75 240 0,70 0,65 220 7:56 9:35 12:32 0,60 hora 0,55 07:56 09:35 hora 360 330 300 ET/CS RC/CS 0,85 280 270 240 210 180 150 07:56 09:35 hora 12:32 12:32 Commercial production 2004/2005 Greenhouse Full sunlight Plant Height (cm) 183.8 a 144.2 b Trunk diameter (cm) 13 a 10 b Leaf number plant-1 35.3 a 29.4 b Leaf area plant (m2) 0.2 a 0.15 b Number of fruits plant- 9.7 a 6.5 b Fruit weight plant-1 3.53 kg a 2.12 kg b Average fruit weight 364.7 g a 326.1g b 1 Martelleto et al 2008 Light quality and intensity can affect leaf anatomy in papaya plant < R/FR Shade 50% Full sunlight Buisson e Lee, 1993 Buisson e Lee, 1993 Sun leaf Shade leaf Leaf thickness (µm) 137 119 Specific leaf mass (mg cm-2) 4,7 2,8 Leaf area (cm-2) 292 162 Chlorophyll content (µg cm-2) 3.57 5.16 Petiole lenght (mm) 207 148 Stomata density (mm-2) 465 330 Degree air space 0.29 0.33 Under conditions of low light intensity and low red:far red light , leaf lobing was dramatically reduced. Change in spectral quality also resulted in a reduction in the ratio chlorophyll a to b High Leaf lobing can reduce leaf temperature in sun leaf Motorcycle radiator The papaya photosynthetic capacity can be linked to non stomatal limitation in soil field capacity condition. As exemple: -Nitrogen leaf concentration 15 g kg-1DM 18 g kg-1DM 35 g kg-1DM 37 g kg-1DM Valor SPAD 41,2 Valor SPAD 39,8 Valor SPAD 26,6 Valor SPAD 6,6 15 g kg-1DM 18 g kg-1DM 35 g kg-1DM 37 g kg-1DM SPAD reading = 60 Sunrise Solo SPAD reading = 30 Golden Eliemar Campostrini [email protected] Fisiologia Vegetal UENF/CCTA 1 Spad reading 45 Spad reading Solo Golden 16 30 14 20 3-4 leaf 15 6-7 leaf 10 5 Apot (µmol O2 m-2 s-1) Apot (mol O2 m-2.s-1) 25 12 10 3-4 leaf 8 6 6-7 leaf 4 2 0 0 10 20 30 40 Spad reading 50 60 70 0 0 10 20 30 40 Spad reading 50 60 70 K+ ATP H+ ADP + Pi ClH2O Field-grown papaya (-20kPa) (--68kPa) Tainung Red Lady Sunrise Marler e Mickelbart, 1998 Strategies to increase effective use of water in papaya -Regulated deficit irrigation (RDI) -Partial rootzone drying (PRD) Strategies to increase effective use of water in papaya -Regulated deficit irrigation (RDI) -Partial rootzone drying (PRD) 15L pots Substrate soil, sand and cattle manure (2:1:2) The plants were kept at field capacity (FC) until they were 96 days old. -Gas exchange -Chlorophyll fluorescence -Growth (central vein lenght, root dry biomass, stem dry biomass, leaf dry biomass, total dry biomass, root volume) -Proline -Carbon isotope discrimination -Agronomic water use efficiency -Thermal imaging Max Av PAR RH Min Temp VPD The plants were kept at field capacity (FC) until they were 96 days old. NI PRD 14 days after planting maximum stress NI FI NI 14 days after planting maximum stress gs (mol H2O m-2 s-1) 0,8 CIFI 0,7 PRD IPSR RDI 0,6 NI 0,5 0,4 0,3 0,2 0,1 0 0 3 6 9 12 14 16 Days after treatment 17 21 0,6 gs (mol m-2 s-1) 0,5 VPD 0,4 0,3 0,2 0,1 0 DAT CI IPSR RDI NI 14 High VPD 12 Transpiration (mmol m-2 s-1) Low VPD High VPD 12 FI CI PRD IPSR 10 y = 10.871x + 1.6225 R² = 0.722 RDI y = 12.69x + 0.72 R² = 0.5369 FI y = 11.092x + 1.4987 R² = 0.7368 PRD 10 y = 10.543x + 0.7411 R² = 0.715 NI 8 6 4 FI PRD RDI NI E (mmolH2O m-2 s-1) RDI 2 NI 8 0 0 6 0,2 0,4 0,6 Stomatal conductance (mol m-2 s-1) 4 2 0 0 3 6 9 12 14 16 Days after treatment 17 21 0,8 1 20 Photosynthesis (mol m-2 s-1) 20 10 FI 5 16 12 10 8 6 4 2 0 RDI 0 NI 0 0 3 FI RDI NI 14 PRD 6 9 12 14 16 Days after treatment 17 50 100 150 Soil water potential (kPa) 200 16 21 14 A (µmol m-2 s-1) A (µmol CO2 m-2 s-1) 15 18 12 10 8 6 4 FI PRD RDI NI Photochemical efficiency of PSII PI=(RC/ABS) x (TR/DI) x (ET/(TR-ET)) (RC/ABS): Active RC density on a Chl basis (FV/F0): Performance due to trapping probability Fv/F0=TR/DI (ET/(TR-ET): Performance due to electron-transport probability Fv/Fm= TR/ABS 85 20 FI NI PRD RDI NI 80 16 75 14 70 High number Leaf number 18 FI y = 0.281x + 12.816 R² = 0.9695 FI 12 y = 0.8018x + 61.052 R² = 0.9566 FI 65 60 10 8 y = 0.2068x + 13.101 R² = 0.8998 RDI y = 0.1678x + 12.876 R² = 0.8736 PRD 55 y = 0.7248x + 58.33 R² = 0.9617 PRD y = 0.5561x + 61.358 R² = 0.9666 RDI 50 6 0 5 10 15 Days after treament 20 25 0 5 10 15 Days after treament 20 25 700 Root volume (cm3) 600 500 400 300 200 100 0 CI IPSR RDI FI FI NI RDI PRD NI 1,60 Leaf area (m2) 1,40 1,20 1,00 0,80 0,60 0,40 FI NI PRD RDI FI NI FI Dry side of PRD root dead (7 days without irrigation) young root 5 days with irrigation after 7 days without irrigation Carbon isotope discrimination CI IPSR RDI NI -27,5 >Ci/Ca (≈0,7) = < δ‰ -28 b b b <Ci/Ca (≈0,3) = > δ‰ -29 -29,5 -30 0,9 a -30,5 0,8 Ca 0,7 Ci/Ca δ13C -28,5 a 0,6 b 0,5 bc 0,4 Ci c 0,3 0,2 CI IPSR RDI NI Agronomic water use efficiency AWUE (g total DW.L-1) 5 a a 4 b 3 b 2 Treatment FI PRD RDI NI Water economy in relation FI (%) 50% 50% 55.14% AWUE (g DM L-1) 3.12 4.55 4.57 2.46 Transpiration rate (L H2O g DM-1) 0.322 0.217 0.217 0.400 1 0 FI PRD RDI NI C3 crops 1 to 6 g DM L-1 H2O C4 grasses 10 to 30 g DM L-1 H2O Arkley (1982) Treatment L H2O m-2 day-1 FI 1.63 PRD 0.84 RDI 0.78 NI 1.17 Treatment Volume water applied per plant per day Transpiration L H2O per m2 leaf per day per plant Transpiration L H20 per plant per day Leaf area m2 age Whole canopy summer 16.0 2.5 10 4.0 5 months Whole canopy winter 10.0 4.2 15 3.5 5 months FI 2.3 1.63 2.3 1.41 3 months PRD 1.1 0.84 1.1 1.30 3 months RDI 1.1 0.78 1.1 1.40 3 months NI 1.0 1.17 1.0 0.85 3 months Thermal imaging 20 months Caliman company Brazil http://www.caliman.com.br/pt/ Field condition Field condition ET0 Field condition July 80 60 50 70% 40 1nd gas exchange measurements 30 20 10 70% 0 3 days after rainfall 13 days after rainfall Rainfall (mm) Rainfall (mm) 70 80 70 60 50 40 30 20 10 0 October 2nd gas exchange measurements Dr. Thomas Marler University of Guam Guam Flooding Papaya is considered a species sensitive to low oxygen availability in the soil (hypoxia), which is commonly caused by waterlogging (Ogden et al., 1981; Malo and Campbell, 1986) Reduced oxygen can occur as a result of tropical storms that saturate the soil for several days, flood irrigation, as well as microirrigation practices that create microenvironments of reduced soil oxygen A completely flooded soil can cause death to papaya plants in 2 d (Wolf and Lynch, 1940; Khondaker and Ozawa, 2007) or 3 to 4 d (Samson, 1980) Khondaker and Ozawa (2007) constructed chambers that controlled soil gas composition at ambient (20%), 18% and 11% oxygen; under soil oxygen at and below 18%, A, chlorophyll content, large and small roots, and shoot dry matter were all decreased 20% O2 18% O2 11% O2 20% O2 18% O2 11% O2 Box 1: 20% O2 Box 2: 18% O2 Box 3: 11% O2 Papaya, considered sensitive to hypoxia, responds with accentuated senescence (chlorotic leaves), leaf fall and does not recover after hypoxic conditions are removed (Marler et al., 1994). These studies indicate that papaya is sensitive to small reductions in soil oxygen content and it is likely that micro-irrigation saturation of a small portion of the soil is having some negative effects. Consequently, a welldrained soil is essential for high productivity. Salinity Papaya seed germination is inhibited by very low levels of salinity (Kottenmeier et al., 1983), yet seedling growth can be stimulated by 1/10 seawater salinity levels (8 mS cm-1) when compared to a Hoagland’s nutrient solution control (Kottenmeier et al., 1983) Maas (1993), however, classified papaya production as moderately sensitive with salinity effects at 3 mS cm-1 Similarly Elder et al. (2000) found that moderately saline water (1.4 to 4 mS cm-1) applied in trickle or under-tree mini-sprinkler irrigation had no adverse affect on productivity but when overhead applied, there was leaf damage and reduced growth. seawater: 3.5% (35 g/L, or 599 mM) 50-80 mS cm-1 Hoagland solution: 2.7 mS cm-1 1 mS cm-1 = 1 dS m-1 The experiment was conducted in a greenhouse between March and October 2010, at UENF, in Campos dos Goytacazes, RJ 2 genotypes: Golden and UENF/Caliman 100L pots EC 1; 1.6; 2.2; 2.8; and 3.4 dS m-1 96 to 126 Days after transplanting * Control. The experiment was conducted in a greenhouse between March and October 2010, at UENF, in Campos dos Goytacazes, RJ 2 genotypes: Golden and UENF/Caliman 100L pots EC 1; 1.6; 2.2; 2.8; and 3.4 dS m-1 * Control. The experiment was conducted in a greenhouse between March and October 2010, at UENF, in Campos dos Goytacazes, RJ 2 genotypes: Golden and UENF/Caliman 100L pots EC 1; 1.6; 2.2; 2.8; and 3.4 dS m-1 Treat. Treat. Treat. Treat. Treat. Solution A Solution B 1 2* 3 4 5 Fertilizers (g) x 0.5 x1 x 1.5 x2 x 2.5 Urea 23.7 47.5 71.3 95.1 118.8 MAP 11.8 23.6 35.4 47.3 59.1 K2SO4 29.6 59.3 88.9 118.6 148.3 MgSO4 29.6 59.2 88.8 118.4 148 Micronutrients 3.5 7.0 10.5 14 17.5 CE (dS m-1) 1.0 1.6 2.2 2.8 3.4 Ca(NO3)2 56.2 CE (dS m-1) 1.0 112.4 168.6 224.8 1.5 2.0 2.6 Maximum 3 L each treatment per day per plant. After each nutrient solution were applied 1.5 to 3L water in each plant per day; 3 times per day) 281 3.2 Φe = is the initial slope of the light response curve ETR versus PAR(quantum efficiency) PI=(RC/ABS) x (TR/DI) x (ET/(TR-ET)) (RC/ABS): Active RC density on a Chl basis (FV/F0): Performance due to trapping probability Fv/F0=TR/DI (ET/(TR-ET): Performance due to electron-transport probability 75 DAP Golden 25ºC 1,6 dS m-1 35,9ºC 3,4 dS m-1 UENF/ Caliman 25ºC 1,6 dS m-1 25,4ºC 1,6 dS m-1 32,4ºC 3,4 dS m-1 Relationships between sap-flow measurements, whole-canopy transpiration, and reference evapotranspiration in field-grown papaya (carica papaya l.) Summer: (clear sky, during 4 days) PPFmax: 2400 mol m-2 s-1 Tmax: 38ºC VPDmax: 4 kPa Winter: (clear sky during 4 days) PPFmax: 1400 mol m-2 s-1 Tmax: 33ºC VPDmax: 3.5 kPa The crop was irrigated with a drip/fertigation system providing supplemental irrigation of 10 (winter) and 16 L per plant per day (summer) Under the environmental conditions evaluated : (4 sunny days) Winter: Maximum vapor pressure deficit (VPDair)=3.5 kPa Air maximum temperature of 33°C Maximum PPF: 2400 mol m-2 s-1 Summer Maximum VPDair=4.0 kPa Air maximum temperature of 38ºC Maximum PPF : 1400 mol m-2 s-1 Leaf area each plant 5 months old Winter :3.5m2 Summer: 4 m2 A B Effects on sap flow heated probe non-heated probe Sap flow measure differences between heated and non-heated probe H2O Water reduce temperature Xylem vessel [reviewer1]is night time relevant? 0,40 y = -1,2861x2 + 1,0287x + 0,1222 R² = 0,7867 0,35 K=[(ΔTmax/ΔT)-1] 0,30 0,25 winter 8-9 winter 10-11 winter 12-13 winter 14-15 winter 16-17 summer 8-9 summer 10-11 summer 12-13 summer 14-15 summer 16-17 0,20 0,15 0,10 0,05 0,00 0 0,2 0,4 0,6 Transpiration (L m-2 leaf h-1) K is the heat coefficient: Tm : the maximum temperature difference (°C) between sensors in active xylem (night time), and T is the temperature difference (°C) between sensors in active xylem 0,8 Mycorrhizal fungi effects on papaya productivity The beneficial effects of arbuscular mycorrhizal (AM) fungi in the plant kingdom and agricultural cropping systems are well documented, and include increased P, water, and nutrient uptake as well as improved pest resistance (Harley and Smith, 1983; Bethlenfalvay and Linderman, 1992) Arbuscular mycorrhizal fungi colonize papaya under natural conditions. Papaya appears to be very dependent on AM since plants in sterilized soil, as compared to inoculated, showed poor growth and particularly P uptake (Habte, 2000) Mohandas (1992) reported that AM inoculation of papaya seedlings increased growth, P concentration and acid phosphatase activity in leaves 20 days of water-stress treatment Treatments were applied 3 months after planting 20 days of waterstress treatment Mycorrhiza establishment may result in the control of ethylene levels as one mechanism of reducing damage by water stress in papaya plants. Besmer and Koide (1999) showed that mycorrhizal colonization can decrease ethylene concentration in flowers, which might explain the increased vase-life of cut flowers. AM colonization may act as an inhibitor of ethylene biosynthesis by influencing ACC conversion to ethylene
Baixar