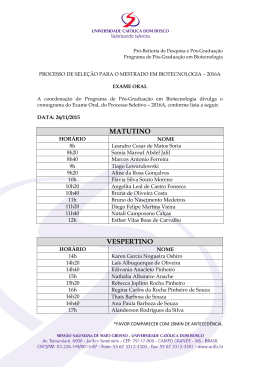
R$ 5,00 ano III número 19 março/abril de 2001 BIOTECNOLOGIA/KL3 (Repete Fotolito 2º capa última edição) 2 Biotecnologia Ciência & Desenvolvimento BIOTECNOLOGIA/KL3 (Repete Fotolito pág. 3 da última edição) Biotecnologia Ciência & Desenvolvimento 3 A CLONAGEM DE BOVINOS ENTREVISTA NO BRASIL Entrevista concedida a Lucas Tadeu Ferreira E-mail do Dr. Rodolfo Rumpf: [email protected] Fotos: Cláudio Bezerra Os segredos da Vitória A EMBRAPA surpreende o mundo e anuncia que obteve o primeiro nascimento de um clone bovino, da raça simental, na sua fazenda experimental de Brasília, no mês de março de 2001. Apelidada simpaticamente de Vitória, pelo seu próprio criador e líder do projeto de pesquisa de clonagem, o médico veterinário Ph.D, Rodofo Rumpf, que explica que para a obtenção Rodolfo Rumpf da Vitória foram utilizados óvulos imaturos de vacas mestiças das raças nelore e simental e que foram feitas vinte e nove tentativas e só uma deu certo, culminando com o nascimento da Vitória, que teve a gestação e parto normais. De acordo com Rodolfo Rumpf, a técnica de clonagem da Vitória foi muito parecida com a empregada na produção da ovelha Dolly, em 1997, na Escócia. A diferença básica dos métodos empregados na obtenção dos dois animais é a origem da célula doadora do núcleo, onde fica o material genético extraído e transferido. As que deram origem à ovelha Dolly foram obtidas de glândulas mamárias de uma fêmea de sete anos, enquanto que as células do núcleo que originaram a Vitória foram extraídas de um embrião de apenas cinco dias e transferidas para outro embrião da mesma idade e raça. Vitória nasceu perfeita, o que deixou Rodolfo e sua equipe muito confiantes quanto à continuidade do projeto de clonagem na Embrapa. Os objetivos pretendidos pela Embrapa com este projeto de clonagem, no médio e longo prazos, são principalmente a regeneração de bancos genéticos, a multiplicação de animais com boas características genéticas, a otimização e maximização do potencial genético das raças de interesse zootécnico, além de possibilitar o resgate e a multiplicação de raças silvestres ou comerciais em risco de extinção, incluindo aí o melhoramento genético de todas elas. Para falar um pouco mais deste projeto de pesquisa que resultou na Vitória e do estado da arte da clonagem animal, no Brasil, Rodolfo Rumpf, que é pesquisador da Embrapa Recursos Genéticos e Biotecnologia, em Brasília, concedeu esta entrevista a Lucas Tadeu Ferreira para esta edição da revista BIOTECNOLOGIA Ciência & Desenvolvimento. 4 Biotecnologia Ciência & Desenvolvimento BC&D - Qual foi a técnica empregada na obtenção da bezerra Vitória? Rodolfo Rumpf - A técnica empregada na obtenção da Vitória é conhecida como transferência nuclear. Utilizamos uma célula doadora de núcleo oriunda de um embrião que, efetivamente, transferiu a herança genética para um ovócito receptor previamente nucleado, de uma outra vaca. Trabalhamos com o núcleo de um material genético elite e o citoplasma que recebeu este núcleo de um outro animal que não era de elevado mérito genético. Um ponto importante temos que esclarecer: neste citoplasma havia também o DNA mitocondrial o qual pode ser responsável por características fenotípicas, inclusive de produção. Com a Vitória, temos um clone, mas não é um clone verdadeiro, como se diz. Ele tem algumas características de DNA externo àquele de seu núcleo oriundo da célula do embrião doador. BC&D - Neste caso, a Vitória é clone de qual animal e pertence à que raça? Rodolfo Rumpf Vitória é clone de um embrião que não chegou a nascer, que era oriundo de um acasalamento da raça simental. Isto foi muito questionado quando anunciamos o nascimento dela: se ela não é igual a outro animal, não é clone. Na verdade, foi a própria mídia que divulgou que a tecnologia de transferência nuclear é igual à clonagem. Mas, não é exatamente assim. A melhor técnica de que dispomos hoje para viabilizar a clonagem - cópia de indivíduos idênticos em larga escala - é de fato a transferência nuclear. Por exemplo: se daquele embrião que nós utilizamos na transferência nuclear e obtivemos a Vitória conseguíssemos ainda produzir outros três embriões que, infelizmente, não prosperaram, tivéssemos obtido mais de um nascimento, esses indivíduos seriam idênticos - seriam clones. Este é um ponto importante que cabe esclarecer porque ficou a dúvida na mídia. De fato, a técnica da transferência nuclear permite obter clones, com a ressalva das questões relativas à herança citoplasmática já discutidas. já conduzia a parte inicial das pesquisas na área animal juntamente com o Dr. Teodore Romano Vaske. Escrevemos um programa de pesquisa de reprodução animal, que existe até hoje, onde prevíamos por etapas o que iríamos fazer na área da biotecnologia da reprodução e com a conservação de recursos genéticos animais. Vitória e sua mãe Sempre trabalhamos para criopreservar sêmen, óvulos, emCabe ainda uma explicação adicional a brião ou células, quer dizer, o banco esta pergunta: O clone verdadeiro só genético de animais, mas também viseria obtido a partir da separação das sando regenerar este banco genético, células de um embrião muito jovem - trazer de volta o germoplasma e dispo3 a 4 dias - , as quais ainda possuem a nibilizá-lo nos rebanhos. Todas as noscapacidade de se desenvolverem isola- sas tecnologias foram dirigidas para damente e cada uma podendo dar este objetivo. Em 1993, fiz um treinaorigem a um novo embrião. Neste mento de pós-doutoramento no Canacaso, o número de possibilidades inici- dá, na Universidade de Montreal, no ais não passaria de 8 cópias se tudo Centro de Pesquisa em Reprodução fosse 100% para a espécie bovina. A Animal sob a orientação do Dr. Laoutra alternativa seria a clonagem de wrence Smith, que é um brasileiro naturalizado canadense e trabalhou em sua tese de doutorado com o Dr. Ian Wilmult - quem produziu a ovelha A técnica empregada na Dolly, na Escócia. Neste ano, ao retorobtenção da Vitória é conhenar para Brasília, iniciamos a aquisição cida como transferência de equipamentos, construção de laboratórios, formação de recursos humanuclear. Trabalhamos com o nos, e tínhamos também que melhorar núcleo de um material os índices técnicos da fecundação in genético elite e o citoplasma vitro num patamar técnico-científico que justificasse a clonagem. Efetivaque recebeu este núcleo de mente, em 1999/2000 é que, de fato, a um outro animal que não era clonagem ganhou espaço no nosso laboratório, porque as outras tecnolode elevado mérito genético gias já haviam atingido a maturação científica necessária para dar suporte a uma vaca adulta em que tanto a célula ela. Gastamos, nos últimos três anos, doadora do núcleo como o ovócito perto de 300 mil reais em custeio, receptor fossem obtidos dela mesma. porque já tínhamos a infra-estrutura. No novo indivíduo tanto o DNA mito- Quem financiou as pesquisas foi a condrial como o nuclear possuem a Embrapa, além da Fundação de Apoio mesma origem. O macho sempre vai à Pesquisa FAP-DF que aprovou um depender de um citoplasma de alguma projeto de produção de embriões e fêmea e por isso não seria verdadeiro. clones in vitro e nos ajudou a impulsionar este trabalho. Como parceiros, BC&D - Em que ano foram iniciados tivemos basicamente a minha formaos primeiros trabalhos desta pes- ção no Canadá e, depois, não tivemos quisa na Embrapa Recursos Genéti- muita interação com o pessoal de lá. cos e Biotecnologia, quem finan- No Brasil, contribuímos para estruturar ciou, quanto custou, e quais foram o laboratório de transferência nuclear os seus principais parceiros? da USP já que eu co-orientava um estudante de mestrado do professor Rodolfo Rumpf - Em 1989, quando Visintin; as parcerias efetivamente vão cheguei à Embrapa Recursos Genéti- se constituir daqui pra frente. Espero. A cos e Biotecnologia, o Dr. Assis Rober- equipe específica envolvida com os to De Bem, que faleceu precocemente, experimentos de transferência nuclear Biotecnologia Ciência & Desenvolvimento 5 foi constituída por mim, dois estudantes de mestrado e o técnico Regivaldo Vieira de Sousa, da Embrapa. Uma andorinha só não faz verão; é necessário o apoio das outras biotécnicas para viabilizar com eficiência a clonagem, e aí a equipe é bem maior. BC&D - Qual a principal diferença da técnica empregada na obtenção da bezerra Vitória e da ovelha Dolly? Rodolfo Rumpf Basicamente, a técnica de transferência nuclear é a mesma. Existem peculiaridades com relação ao momento da nucleação do citoplasma, mas a grande diferença é que a Dolly vai ter este mérito pelo resto da história, já que foi concebida de uma célula de um indivíduo adulto, extraída da glândula mamária, ou seja, de uma célula somática diferenciada. Essa sinalização de Dolly nos abriu muitos outros caminhos e a nossa Vitória seria um passo anterior à Dolly, porque trabalhamos com uma célula embrionária que antecede a sua diferenciação. Vitória foi obtida de uma célula que, do ponto de vista da reprogramação nuclear, provavelmente é muito mais reprogramável do que uma célula somática. BC&D - Quantas tentativas de clonagem foram realizadas para obtenção da Vitória? Rodolfo Rumpf - Foi na terceira gestação que obtivemos êxito. As outras perdemos por excesso de zelo. Neste experimento estávamos olhando muito mais para o citoplasma receptor do que para o núcleo. Utilizamos um embrião de cinco dias e meio congelado, que coletamos pela técnica de transferência de embriões clássica; ele foi descongelado e suas células separadas. No citoplasma estávamos avaliando outros aspectos relativos a qualidade. Por que o citoplasma é importante? Ele contém todas mensagens que permitem a reprogramação do núcleo. Alguns anos atrás atribuíamos todo este mérito ao núcleo. Hoje sabemos que se não for um bom citoplasma e se ele não estiver devidamente sintonizado com o núcleo não vamos ter suces6 Biotecnologia Ciência & Desenvolvimento so. Este é o segredo. Produzimos neste experimento da Vitória mais de vinte embriões; quinze foram inovulados, ou seja, transferidos para mães de aluguel distintas e destes quinze resultou apenas a Vitória. A Vitória teve origem num embrião, o qual após o descongelamento conseguimos isolar vinte e nove células viáveis que resultaram em quatro embriões no estágio de blastocisto, que é o estágio esperado de um embrião de sete dias. Estes quatro embriões foram transferidos para diferentes receptoras e nasceu apenas a Vitória. Com a Vitória, temos um clone, mas não é um clone verdadeiro, como se diz. Ele tem algumas características de DNA externo àquele de seu núcleo oriundo da célula do embrião doador BC&D É correto afirmar que a Dollly já nasceu velha com idade do doador das células adultas -, e que a Vitória nasceu totalmente jovem, com a idade do embrião doador do núcleo? Rodolfo Rumpf Sim, isto é um fato. Mas é um conceito que está hoje um pouco ultrapassado, porque esta questão da idade está relacionada à atividade de uma enzima, a telomerase, que produz os telômeros, localizados nas extremidades dos cromossomos e que ajudam na hora da divisão celular, para que não ocorram translocações, erros na divisão celular. À medida que a idade avança, a ação da telomerase vai diminuindo e o indivíduo vai ficando mais exposto a doenças de fundo genético. Depois desta constatação, de que a Dolly teria a idade biológica da doadora do núcleo, se provou em outros ensaios que é possível interferir na atividade da telomerase, e inclusive aumentar a expectativa de vida do futuro clone. Acredito que a questão da idade do clone de indivíduos adultos, logo, logo será resolvida, se é que já não foi. A Vitória teria biologicamente a idade de uma concepção normal. BC&D - Numa próxima etapa das pesquisas da Embrapa, o senhor pretende empregar técnicas semelhantes às da ovelha Dolly para obtenção de novos clones? Rodolfo Rumpf Sim, sem dúvida. Estamos iniciando agora pesquisas com células somáticas, fibroblastos da pele. Tiramos um pedacinho da orelha do animal e este pedaço de pele é capaz de produzir muitas células pelo cultivo em laboratório e se tornar uma fonte doadora de núcleos quase inesgotável. Esta técnica nos interessa mais pelo seu potencial em termos de regeneração de raças em vias de extinção, tanto domésticos como silvestres. Será possível no futuro regenerar animais silvestres atropelados, encontrados mortos nas rodovias. Vamos pegar um pedaço de pele de um indivíduo, mandar para um laboratório, fazer um banco de células e guardar para no futuro regenerar animais. O que tem que ficar bem claro é que partes desta tecnologia já foram patenteadas em outros países e que o uso é restrito para trabalhos científicos. A partir do momento em que formos trabalhar em escala comercial vamos nos inserir no contexto legal que protege a propriedade intelectual. BC&D - Além do avanço científico, que vantagens comparativas a clonagem pode oferecer em relação à reprodução convencional? Rodolfo Rumpf - Temos que deixar bem claro que a clonagem ainda não está suficientemente madura para ser utilizada na multiplicação de animais de interesse zootécnico. Temos uma picada aberta. Falta ainda abrir a estrada, pavimentá-la etc., e chegar a uma relação custo/ benefício compatível com a atividade. Existe um estudo dos melhoristas franceses que mostra que, por simulação, a partir do momento em que se usa estas modernas biotécnicas de multiplicação animal, aí incluída a fecundação in vitro, a punção folicular, a sexagem do sêmen, a própria clonagem, o congelamento etc., enfim, todas estas tecnologias devidamente articuladas, é possível conseguir em um ano o avanço genético para determinada característica que, normalmente, pelos métodos convencionais, demoraria doze anos. BC&D - A clonagem pode contribuir com a produção de animais transgênicos? Rodolfo Rumpf - Temos relacionado três grandes áreas de utilização da clonagem: a regeneração de recursos genéticos, a questão da produção animal, o apoio que ela dará aos programas de melhoramento genético animal, e o suporte cientifico para a própria embriologia e para a produção de animais transgênicos. Por que isso? De um pedaço de pele, por exemplo, conseguimos fazer talvez milhões de células. Transformamos estas células in vitro, fazemos uma transfecção, colocamos junto a um gene marcador, e depois de alguns dias identificamos as células que efetivamente foram transformadas geneticamente, e usamos estas células no sistema de reconstrução nuclear. Com isso, conseguimos melhorar em muito a eficiência na produção dos transgênicos. Portanto, tem tudo a ver a técnica da clonagem com os transgênicos, e esse é o próximo passo que vamos dar. BC&D - Quais vantagens e benefícios podem ser incorporados aos animais transgênicos clonados? Rodolfo Rumpf O que pensamos em fazer em termos de transgênicos? No início, estamos preocupados em desenvolver um modelo porque temos de um lado um projeto de conservação e caracterização de recursos genéticos, e um segundo que faz prospecção de genes; além de um terceiro projeto que desenvolve ferramentas biotecnológicas, que é o nosso. Os esforços devem andar juntos e o dia em que for encontrado genes de interesse, a tecnologia de obtenção de animais transgênicos deverá já estar dominada. Assim poderemos transferir genes associados a resistência a endo e ectoparasitas, por exemplo, de animais comerciais. Com isso estaríamos diminuindo o custo de pro- Na verdade, foi a própria mídia que divulgou que a tecnologia de transferência nuclear é igual à clonagem. Mas, não é exatamente assim. A melhor técnica de que dispomos hoje para viabilizar a clonagem - cópia de indivíduos idênticos em larga escala - é de fato a transferência nuclear dução, melhorando a qualidade da carne ou do leite, beneficiando tanto o produtor como, e, principalmente, o consumidor. Este é apenas um exemplo que ainda não sabemos se efetivamente será possível, mas existe muitas outras possibilidades de uso de animais transgênicos. A produção de biomoléculas, a melhoria na eficiência de produção, a qualidade de produtos e a produção de modelos animais para estudos de doenças ou até mesmo para a doação de Por que o citoplasma é importante? Ele contém todas mensagens que permitem a reprogramação do núcleo. Alguns anos atrás atribuíamos todo este mérito ao núcleo. Hoje sabemos que se não for um bom citoplasma e se ele não estiver devidamente sintonizado com o núcleo não vamos ter sucesso. Este é o segredo órgãos (os xenotransplantes), entre outras possibilidades de uso. Em última análise, agregar valor ao produto final. O que é bom para o consumidor deve ser bom para o produtor também. BC&D Qual é objetivamente a finalidade principal do seu trabalho? Rodolfo Rumpf É desenvolver ferramentas biotecnológicas que viabilizem o banco genético e que permitam regenerar qualquer animal de interesse zootécnico e/ou em risco de extinção. Regenerar significa tirar a célula, o ovócito, o sêmen, o embrião que está lá no botijão congelado, fazer indivíduos a partir deste material, e trazer este material genético de volta para o rebanho. Esta é a motivação inicial de todo o trabalho. Estamos hoje no modelo bovino. Deveremos começar a trabalhar com ovino, caprino, enfim, o importante é que cada espécie tem suas individualidades fisiológicas e que a inserção da tecnologia no modelo específico só terá sucesso se for efetuado segundo a base fisiológica. É claro que estas tecnologias possuem uma enorme demanda do setor produtivo e, dentro do possível, procuramos atendê-lo. BC&D - Na sua opinião, como a comunidade científica reagiu ao anúncio do clone Vitória? Rodolfo Rumpf - Pelo que temos recebido de manifestação, a comunidade científica se mostrou muito satisfeita com a Vitória. É importante frisar que, hoje, conseguimos a Vitória e que vamos conseguir mais ainda no Brasil. Já existem aqui quatro ou cinco laboratórios trabalhando com transferência nuclear, e isso é muito bom porque imagino que vamos ter um grande progresso, desta técnica específica, em um futuro próximo. E, como eu disse, temos muito a fazer ainda; o que fizemos foi apenas abrir uma picada na selva. Tem toda uma maturação da tecnologia ainda pela frente. Se tivermos várias equipes, com várias cabeças e em ecossistemas distintos, trabalhando com objetivos semelhantes, então vamos ter, seguramente, um grande progresso científico nessa área. Gostaria de mencionar que hoje temos equipe de pesquisadores na Unesp de Jaboticabal, na Faculdade de Medicina Veterinária da USP/SP, na Faculdade de Veterinária, da URGS, na Faculdade de Veterinária da Norte-Fluminense e também no estado do Pará. Todos, se ainda não estão trabalhando, estão montando seus laboratórios, e, seguramente, logo vamos ouvir falar dos feitos de todos eles. BC&D - Podemos afirmar que, a partir da Vitória, o Brasil encontra-se hoje ao lado dos países do primeiro mundo no domínio da tecnologia da clonagem animal? Biotecnologia Ciência & Desenvolvimento 7 Rodolfo Rumpf - É difícil fazer esta afirmação. A grande diferença é a agilidade que se tem lá fora nos países desenvolvidos em relação ao que temos aqui, no Brasil. Agilidade desde a importação de um determinado reagente, à rapidez com que a ciência básica responde a demandas específicas. No Brasil temos que abrir espaço e fomentar cada vez mais a ciência básica; é ela que nos dá o suporte necessário às pesquisas aplicadas. Quando se busca o desenvolvimento tecnológico tem-se que parar muitas vezes, e voltar, fazer parcerias, buscar respostas que não se tem disponíveis naquele momento. Isto acontece em qualquer lugar do mundo, mas eu diria que lá fora eles têm mais agilidade exatamente porque investem mais em pesquisa básica. BC&D - Do ponto de vista biológico e veterinário, quais as características deletérias potenciais que podem se expressar na Vitória a longo prazo? Rodolfo Rumpf - Isto nós não sabemos. Estamos atualmente extremamente satisfeitos porque os nove meses de gravidez foram de aflição, e as duas últimas semanas, antes do nascimento, foram altamente estressantes para nós; porque tínhamos um plantão de 24 horas, não queríamos interferir de jeito algum, aguardamos mesmo o limite para o parto ocorrer naturalmente, e, para nossa surpresa, tudo ocorreu dentro da maior normalidade possível. Há que se dizer também que a mãe de aluguel é uma mãe de primeira. Ela não só pariu como fez todos os procedimentos necessários, como massagear, limpar a bezerra, também amamentou no período desejável e, por fim, eliminou a sua placenta. Nós acompanhamos à distância e registramos todas as informações possíveis. Contudo, até o momento, estamos muito contentes e felizes. Nada nos garante que daqui a alguns dias ela não tenha um problema imunológico qualquer e que venha a falecer como já aconteceu com outros clones nascidos no mundo. Vitória se tornou um pequeno laboratório de estudos e vai ser um indicador eterno para nós como a Dolly está sendo para o mundo. BC&D - É possível a clonagem hu8 8 Biotecnologia Ciência & Desenvolvimento Biotecnologia Ciência & Desenvolvimento mana usando as mesmas técnicas? Rodolfo Rumpf - Este é um terreno complicado e eu não me considero em condições de comentar muito sobre isto. Antes de comentar o assunto é importante voltarmos para a questão Estamos iniciando agora pesquisas com células somáticas, fibroblastos da pele. Tiramos um pedacinho da orelha do animal e este pedaço de pele é capaz de produzir muitas células pelo cultivo em laboratório e se tornar uma fonte doadora de núcleos quase inesgotável conceitual da clonagem e da transferência nuclear. A clonagem humana não tem nada que possa justificá-la, já a transferência nuclear poderia encontrar indicação pontual na clínica reprodutiva, ou ainda nas pesquisas de doenças hereditárias, por exemplo. Se existir uma indicação clínica pontual, não vejo razão no futuro para que ela A grande diferença é a agilidade que se tem lá fora nos países desenvolvidos em relação ao que temos aqui, no Brasil. Agilidade desde a importação de um determinado reagente, à rapidez com que a ciência básica responde a demandas específicas não venha a ser utilizada, mas muito antes de discutirmos isso, temos que ouvir o que a sociedade deseja. Somos favoráveis à criação de instrumentos legais e éticos que permitam o desenvolvimento da ciência e monitorem o bom uso das tecnologias. BC&D Quais os limites da legislação brasileira à clonagem humana? Rodolfo Rumpf - De fato, a legislação brasileira deve ser revista neste aspecto e eu gostaria que tivéssemos uma legislação muito clara, que criasse mecanismos de regulamentação do uso da transferência nuclear e que não proibisse tudo simplesmente. Neste sentido, já existem os conselhos de ética, mas também deveria existir, a exemplo da Comissão Técnica Nacional de Biossegurança CTNBio, um fórum em âmbito nacional, com todos os segmentos da sociedade lá representados e que pudesse opinar sobre questões éticas e científicas. Este fórum deveria exigir o cadastramento de todos os pesquisadores que trabalham com transferência nuclear, com bom curriculum acadêmico, condições de trabalho adequadas, descrição detalhada do que estão fazendo e para que estão fazendo, se não existe outro modelo científico que possa trazer as mesmas informações e se existe coerência entre a metodologia e os objetivos, entre outras informações importantes. Todos os projetos passariam obrigatoriamente por um crivo científico e ao mesmo tempo ficariam sujeitos a uma análise dos outros segmentos da sociedade que, em última instância, é que têm que nos dizer o que é que temos que fazer. BC&D - Na sua opinião, a sociedade brasileira, de um modo geral, está preparada para receber os clones animais? Rodolfo Rumpf Até agora, temos tido um retorno muito bom das pessoas para as quais temos proferido palestras sobre a clonagem e as modernas biotécnicas de reprodução animal, em todo o país. A própria mídia tem assimilado de forma muito satisfatória esta questão. Basta ler a cobertura que os jornais, as revistas e a televisão deram ao anúncio da Vitória. É claro que sempre vai haver os mais diversos questionamentos dos segmentos representativos da sociedade, e é importante e bom que seja assim. Temos que aproximar, cada vez mais, os cientistas da sociedade e a sociedade dos cientistas. E a Vitória, de alguma forma, está contribuindo para esta aproximação. Em última análise trabalhamos para melhorar a qualidade de vida dos nossos cidadãos e quando explicamos de que forma isso irá acontecer, as coisas são muito bem assimiladas. PORTAL WWW.BIOTECNOLOGIA.COM.BR (Repete Fotolito pág. 9 da última edição) Biotecnologia Ciência & Desenvolvimento 9 Carta ao Leitor BIOTECNOLOGIA Ciência & Desenvolvimento KL3 Publicações Fundador Henrique da Silva Castro Direção Geral e Edição Ana Lúcia de Almeida Diretor de Arte Henrique S. Castro Fº Departamento Comercial, Redação e Edição: SRTV/Sul - Quadra 701 Ed. Palácio do Rádio II Sala 215 - CEP 70340-902 Brasília - DF Tel.: (061) 225-1512 (061) 225-0976 Fax: (061) 224-2830 E-mail [email protected] Home-Page www.biotecnologia.com.br Projeto Gráfico Agência de Comunicação IRIS www.agenciairis.com.br [email protected] Impressão: Gráfica São Francisco Fotolito: Ribelito Assinaturas O pedido de assinatura é realizado através da carta resposta-comercial encartada em cada revista, por telefonema ou fax diretamente à KL3 ou pela Internet através dos nossos endereços eletrônicos. A revista não tem vendedores autorizados. Os artigos assinados são de inteira responsabilidade de seus autores. ISSN 1414-4522 10 Biotecnologia Ciência & Desenvolvimento O Brasil entra na era do domínio tecnológico da clonagem animal. Com o simpático nome de Vitória, nasceu neste mês de março de 2001, na fazenda da Embrapa, em Brasília, o primeiro bezerro clonado a partir da técnica de transferência nuclear, pesquisa de ponta que é dominada por poucas nações desenvolvidas no mundo. A Vitória é fruto de pesquisa liderada pelo cientista da Embrapa Recursos Genéticos e Biotecnologia, Rodolfo Rumpf, médico veterinário formado pela Universidade Federal de Pelotas com doutoramento na Áustria e pós-doutoramento no Canadá, que vem sendo realizada desde 1984. O processo de clonagem da bezerra Vitória foi muito parecido com o da ovelha Dolly produzida em 1997, na Escócia. Com este feito, o Brasil passa a ser o quinto país do mundo que conseguiu, até hoje, clonar um animal. Antes, somente o Reino Unido, os Estados Unidos, a Austrália e o Japão conseguiram. A ovelha Dolly foi gerada a partir de uma célula extraída da glândula mamária de uma ovelha doadora adulta, de sete anos, que introduzida num embrião foi capaz de gerar um outro indivíduo geneticamente idêntico. No caso da Vitória, os cientistas da Embrapa extraíram as células do núcleo de um embrião de cinco dias e as transferiram para o núcleo de um outro embrião da mesma idade e raça. Aparentemente o processo é simples. Requer contudo muitos estudos, investimentos e, principalmente, recursos humanos que sejam capazes de dominar esta técnica. Felizmente, o nosso Brasil tem cientistas tão bem formados e preparados como os dos países do primeiro mundo, a despeito de enfrentar todas as adversidades e falta de investimentos em ciência e tecnologia. Que a Vitória sirva de alerta e desperte as autoridades da área econômica para que o País invista mais recursos nas pesquisas científicas neste novo milênio. Dr. Henrique da Silva Castro Conselho Científico Dr. Dr. Dr. Dr. Dr. Dr. Dr. Dr. Dr. Dr. Aluízio Borém - Genética e Melhoramento Vegetal Henrique da Silva Castro - Saúde; Ivan Rud de Moraes - Saúde - Toxicologia; João de Deus Medeiros - Embriologia Vegetal; Maçao Tadano - Agricultura; Naftale Katz - Saúde; Pedro Juberg - Ciências; Sérgio Costa Oliveira - Imunologia e Vacinas; Vasco Ariston de Carvalho Azevedo - Genética de Microorganismos; William Gerson Matias - Toxicologia Ambiental. Conselho Brasileiro de Fitossanidade - Cobrafi Dr. Luís Carlos Bhering Nasser - Fitopatologia Fundação Dalmo Catauli Giacometti Dr. Eugen Silvano Gander - Engenharia Genética; Dr. José Manuel Cabral de Sousa Dias - Controle Biológico; Dra. Marisa de Goes - Recursos Genéticos Instituto de Pesquisas Energéticas e Nucleares - IPEN Dr. José Roberto Rogero Colaboraram nesta edição: Adriana Ferreira Uchoa; Andréia Rachel Ramos Cruz; Antônia Elenir Amâncio de Oliveira; Bárbara Barreto Andrade Dias; Cassiana Rocha Azevedo; Cristiane Paulain Cavalcanti; Eliana Abdelhay; Francisco José Lima Aragão; Francisco Ricardo Ferreira; Giovanni Rodrigues Vianna; Janete Vetorazzi; Jean Matos; Jerusa de Souza Andrade; João Batista Teixeira; José Renato Santos Cabral; José Xavier-Filho; Josias Corrêa Faria; Kátia Valevski Sales Fernandes; L. Pedro Barrueto Cid; Lilian Maria Mazzuco; Lílian Pantoja de Oliveira; Luciana Belarmindo da Silva; Luciano Hammes; Luís Carlos de Souza Ferreira; Marco Antônio Lopes Cruz; Margareth das Mercês Cerqueira Albino; Maria da Graça Nascimento; Maura da Cunha; Melissa Ang Simões; Mônica Pereira; Nei Pereira Júnior; Olga Lima Tavares Machado; Paulo Naud; Pedro Ramos da Costa neto; Roberto Maeda; Sônia Maria da Silva Carvalho; Spartaco Astolfi Filho; Thiago Motta Venâncio; Valdirene Moreira Gomes; Viviane de Oliveira Santos. Entrevista Rodolpho Rumpf pág. 04 Pesquisa Promotores complexos Transgênico resistente a geminivirus Biotransformação de óleos e gorduras Insulina de plantas Processo fermentativo para produção de bebida pág. 12 pág. 22 pág. 28 pág. 36 pág. 50 Cultura de tecidos A propagação in vitro de plantas. O que é isso? Biotecnologia aplicada à produção de mudas pág. 16 pág. 42 Saúde Adjuvantes de mucosas pág. 32 BioNotícias pág. 48 Biotecnologia Ciência & Desenvolvimento 11 PESQUISA Promotores Complexos Transgênese animal na dissecção in vivo de promotores complexos Eliana Abdelhay Dra. - Instituto de Biofísica Carlos Chagas Filho. UFRJ. [email protected] Mônica Pereira Dra. - Instituto de Biofísica Carlos Chagas Filho. UFRJ. [email protected] Fotos cedidas pelas autoras s últimos anos viram uma revolução na biologia molecular derivada do alto grau de automação na análise dos genomas e seus produtos. No entanto, a liberação do primeiro rascunho das seqüências presentes no genoma humano relembrou a todos que o gigantesco passo é somente o início de uma longa expectativa até que possamos entender corretamente a função dos diferentes genes e como eles são regulados nas diferentes células e processos. O camundongo tem sido o organismo modelo para a análise de função de genes individuais devido à alta homologia entre os dois genomas. Várias técnicas que incluem geração de alelos nulos (nocaute), mutagênese química e introdução de genes in vivo têm sido utilizadas. Essas técnicas, apesar de muito potentes, são ainda utilizadas de forma artesanal. O mesmo acontece no que diz respeito ao estudo da regulação dos genes. Organismos eucarióticos possuem genes bastante complexos, cuja regulação ocorre em diversos níveis. Genes responsáveis por codificar proteínas mantenedoras (housekeeping) mostram regiões reguladoras (promotores) pequenas (± 500 pb), enquanto genes envolvidos com processos dinâmicos como o desenvolvimento podem ter regiões reguladoras que ultrapassem em várias 12 Biotecnologia Ciência & Desenvolvimento Figura 1 dezenas o tamanho de sua região codificante. Um bom exemplo é o gene engrailed, cuja unidade gênica (região codificante + região reguladora) possui 100.000 pb para produzir um RNA mensageiro de 3000 pb. A pergunta que vem imediatamente após esse conhecimento é: Por que tantas bases de DNA são necessárias para regular o gene? A resposta só pode ser dada se entendermos a complexidade do gene. No homem, duas proteínas engrailed são produzidas a partir dos genes En1 e En2. Essas proteínas têm papel fundamental na delimitação do sistema nervoso, onde En2 define o metaencéfalo e En1, a medula espinhal. No entanto, elas também são essenciais durante vários outros processos, como formação de membros (En1 determina região ventral dos membros), e determinação da região ventral do tubo neural, que dará origem aos motoneurônios, entre outros. Portanto, diversos elementos regulatórios distintos são necessários para ativar ou inibir a expressão do gene em células diferentes ao longo do desenvolvimento embrionário. Além disso, sendo o gene tão essencial na vida do organismo, existem sistemas em duplicata, ou seja, elementos regulatórios estão representados em duplicata no promotor. Poucos são os genes eucarióticos que já têm toda sua região reguladora (promotor) mapeada. Isso porque essa análise exige estudos in vitro e in vivo para que não restem dúvidas sobre qual elemento (seqüência) regula qual atividade. O Gene Msx-1 e seu nocaute: um modelo animal para malformações craniofaciais Em nosso laboratório, nos últimos anos, temos analisado a regulação do gene Msx-1 de camundongo utilizando todas as tecnologias até hoje disponíveis. O gene Msx-1 tem todas as características de um gene de promotor complexo.Ele tem uma expressão dinâmica espaço-temporal, durante a embriogênese. Inicialmente sua expressão ocorre nas células ectodérmicas e mesodérmicas da fenda primitiva durante a gastrulação do embrião (6,5 dpc). Essa expressão, posteriormente, torna-se restrita ao ectoderma e ao mesoderma lateral e ao neuroepitélio, que vai formar a parte dorsal do tubo neural, incluída a região que dará origem às células da crista neural (7,5 a 8,5 dpc), e, nos colcinetes cardíacos, nas células que darão origem à válvula átrioventricular. Nos estágios subseqüentes, sua expressão ocorre nos brotos dos membros na região de proliferação celular, nos gânglios intercostais, na região dorsal dos somitos e do tubo neural, nos gânglios trigeminais, no ectoderma e mesênquima da maxila e mandíbula, plexus coróide e, finalmente, nas papilas dos bulbos capilares (Figura 1). Tem sido sugerido um papel fundamental para o gene Msx-1 durante o desenvolvimento, tanto por sua conservação filogenética, como pelos defeitos encontrados no camundongo nulo para esse gene. O camundongo mutado (homozigoto nulo) não consegue formar os dentes e exibe diversas anormalidades craniofaciais que incluem ausência do processo alveolar da mandíbula e maxila, palato secundário fendido, nos ossos frontal, parietal e nasal. Esse fenótipo é bastante relacionado ao que ocorre em humanos que possuem uma mutação dominante no gene levando a uma craniosinostose. Interessantemente, a perda total do produto do gene parece não afetar outros sítios de expressão do gene, com exceção da região crâniofacial, o que tem sido relacionado ao potencial que o gene parálogo Msx2 tem para substituir a função de Msx1 nesses sítios. Recentemente, a produção de um camundongo nulo para os dois genes mostrou um fenótipo muito mais grave, no qual falta toda a parte superior do crânio, associada aos defeitos já demonstrados no mutante Msx-1. Apesar disso nenhum defeito de membros ou outro órgão foi identificado nesses animais.(Figura 2) A importância dos genes da família Msx para o desenvolvimento, assim como seu padrão complexo de expressão, indica que sua regulação deve envolver um número grande de seqüências e fatores protéicos que a elas se ligam para realizar um contro- Figura 2 le de célula específico da expressão do gene. O promotor do gene Msx-1 Os esforços para desvendar essas seqüências e seus fatores ligantes iniciaram pela clonagem e seqüenciamento de uma região de 5kb a montante da região codificante (ATGiniciador). A análise dessa seqüência e sua comparação com seqüências homólogas em outros organismos, como o homem, permitiu identificar regiões conservadas de reconhecimento de fatores de transcrição. Essas regiões foram classificadas como região promotora basal, que corresponde a 130 bp localizados logo acima do sítio inicial da transcrição, e de quatro caixas contendo putativos sítios ativadores e inibidores capazes de regular o gene Msx-1 (Figura 3). Para verificar a funcionalidade dessas regiões foram efetuados ensaios de transfecção em células F12, e Cos utilizando-se construções de fragmen- tos do promotor ligados ao gene da luciferase e co-transfecção com RSVβ gal. Os resultados demonstraram que a eliminação dos sítios das caixas 3 e 4, assim como os da caixa 2, diminuem sensivelmente o nível de expressão do gene repórter, enquanto a perda da região promotora basal elimina completamente a expressão do gene. Apesar de indicativos, esses resultados não eram capazes de correlacionar uma região específica reguladora com uma das expressões do gene, o que só passou a ser possível com a análise In vivoatravés da transgênese. Identificação de um elemento regulatório capaz de ligar o gene nas células da crista neural cefálica Num primeiro experimento, animais transgênicos que continham os 5 kb de seqüência a montante da região codificante dirigindo a expressão do gene repórter Lac Z mostraram que essa região é suficiente para regular a expressão completa do gene. Em experimentos sucessivos, foram geradas linhagens transgênicas, cuja seqüência do promotor utilizada era representada por deleções na região 5. Portanto, foram obtidos animais transgênicos, nos quais o gene repórter LacZ tinha sua expressão dirigida por 4kb, 3kb, 2kb e 500 pb. Figura 3 Biotecnologia Ciência & Desenvolvimento 13 A comparação entre o padrão de expressão de lac Z nos animais transgênicos que contêm 5 Kb ou 4Kb de seqüência reguladora mostrou que, apesar das expressões serem relativamente mais baixas nos transgênicos contendo 4kb de região regulatória, estas eram mantidas em todos os locais de expressão do gene Msx-1, exceto pela expressão na crista neural cefálica, que estava ausente em todos os animais analisados (Figura 4). Esse resultado sugere que a expressão do gene Msx-1 na crista neural cefálica depende de elementos regulatórios presentes na seqüência de 1kb deletada. A análise dessa seqüência sugere a presença de um consenso para ligação de proteínas contendo domínios bicoid. Entre essas, a melhor candidata a regular a expressão de Msx-1 na cabeça é a proteína codificada pelo gene Otx-2. Este gene é expresso durante a gastrulação do embrião de camundongo na região anterior e, posteriormente mostra um padrão de expressão na cabeça muito semelhante ao do gene Msx-1. Para testar essa hipótese, foram realizados experimentos de modificação de mobilidade eletroforética (EMSA) (Figura 5 ) utilizando-se fragmentos do promotor contendo esse sítio e testando-os para ligação de proteínas de extrato nucleares de células da cabeça ou do tronco de embriões em vários estágios de desenvolvimento. A especificidade da ligação foi garantida por experimentos de competição e pela utilização de mutações introduzidas na seqüência em questão (Figura 6 ). A identificação de uma 14 Biotecnologia Ciência & Desenvolvimento Figura 4 - Perda do padrão de expressão de Lac Z cefálico no animal transgênico contendo 4kb de região promotora do gene MSX-1 Figura 5 Figura 6 região responsável por ligar a expressão do gene num sítio tão importante como a crista neural cefálica, que dará origem a várias estruturas crânio-faciais, como ossos, gânglios e mesênquima, é um passo importante para possíveis manipulações da atividade gênica na embriogênese. Bibliografia Robert, B.; Sasson, D.; Jack, B.; Ghering, W.; Buckingham, M. (1989) Hox-7, a mouse gene with a novel pattern of expression during embryogenesis. EMBO J. 8, 91-100. Lyons, G.E.; Houzelstein, D.; Sassoon, D.; Robert, B. and Buckingham, M. (1992) Multiple sites of Hox 7 expression during mouse embryogenesis: comparison with retinoic acid receptor mRNA localization. Mol. Reprod. Dev., 32, 303-314. Gonzalez, S.M.C.; Ferland, L.H.; Robert B. and Abdelhay E. (1998) Structural and functional analysis of mouse Msx-1 gene promoter sequence conservation with human Msx-1 promoter points at potential regulatory elements. DNA and Cell Biology 17(6) : 561-572. Houzelstein, D.; Cohen, A.; Buckingham, M. and Robert, B. (1997). Insertional mutation of the mouse Msx-1 homeobox gene by an nlac Z reporter gene. Mech.Dev.65: 123133. Satokata, I.; Ma, L.; Ohshima H. et al (2000) Msx-2 deficiency in mice causes pleiotropic defects in bone growth and ectodermal organ formation. Nature Genetics 24: 391-395. Pereira, M.; Houzelstein, D.; Vinci , G.; Cohen, A.; Abdelhay, E. and Robert, B. (1999). Analysis of the mouse Msx1 gene promoter using embryonic stem cell mediated transgenesis. Transgenic 2: 403-416. ANÚNCIO UNISCIENCE (Fotolito enviado anexo) Biotecnologia Ciência & Desenvolvimento 15 A propagação in vitro de plantas. CULTURA DE TECIDOS O que é isso? Cultura de tecidos vegetais - uma ferramenta fundamental no estudo da biologia moderna de plantas. L. Pedro Barrueto Cid Biólogo M.Sc. Ph.D Embrapa/Cenargen - Área de Biologia Celular Autor do livro "O método científico, o cientista e a sociedade", pela editora da Universidade do Amazonas. [email protected] Fotos do autor Definição Antecedentes históricos A propagação in vitro de plantas, Figs.1 e 2, chamada também micropropagação, é uma técnica para propagar plantas dentro de tubos de ensaios ou similares de vidro (por isso, o termo in vitro), sob adequadas condições de assepsia, nutrição e fatores ambientais como luz, temperatura, 02 e CO2. É uma parte importante de biotecnologia, conjuntamente com outras duas áreas: DNA recombinante e fermentação. A cultura in vitro, apresenta diferentes modalidades conforme os objetivos de sua aplicação, como por exemplo, cultura de protoplastos; anteras; calos (Fig. 3); células em suspensão (Fig. 4); sementes (Figs. 5 e 6),etc. A teoria da totipotencialidade formulada por Matthias Schleiden & Theodor Schwann, em 1838, pode ser dita que constitui um dos primeiros fundamentos da cultura in vitro, embora seus formuladores nem tenham imaginado uma metodologia como essa. A teoria afirma que a célula é autônoma, portanto, que contém o potencial necessário para originar um organismo completo; nesse caso, uma planta completa. É claro que essa capacidade deve manifestar-se sob especiais condições de estímulo. Em decorrência desta teoria, células com diferentes fenótipos dentro da planta têm idéntico genótipo. Haberlandt, um fisiólogo vegetal austro-húngaro, por volta de 1902, imbuído dessa teoria, foi o primeiro a manipular um sistema de cultura in vitro de plantas, procurando estabelecer e consolidar um sistema de micropropagação. Infelizmente, por limitações técnicas da época, seus esforços falharam. Contudo, alguns anos mais tarde a partir dos trabalhos de Robbin (1922) e White (1934) em ponta de raízes; cultura de embriões, por La Rue (1936); cultura de calos, por Gautheret Nobécourt (1939); enriquecimento de meios nutritivos com leite de coco, por van Overbeek, 1941; uso de plantas de tabaco como modelo experimental para estudo de morfogênese, por Skoog, desde 1944, e uso de meristemas apicais na obtenção de plantas livres de vírus, por Morel & Martin, 1952, abriram-se as estradas que a cultura de tecido de plantas percorreria triunfalmente ao longo de todo o século XX, com mais e mais descobertas e aplicações. Importância Fig.1: Plântula de café cv Rubi, crescida in vitro e obtida a partir 16 Biotecnologia Ciência & Desenvolvimento de gema axilar de uma outra plântula similar a ela. Assim por diante, outros clones podem ser obtidos A cultura in vitro de plantas, é uma técnica que não apenas apresenta importância prática na área florestal e agrícola, mas, também na científica básica. Dentro do campo da biologia de plantas talvez seja uma das técnicas mais polivalentes. Assim, através da cultura de protoplastos, podem-se hibridizar variedades diferentes, vencendo barreiras genéticas. Através da cultura de anteras, podem-se obter plantas haplóides, que logo depois podem-se diploidizar e transformar-se em homozigotos, isto é, indivíduos que produ- zem um só tipo de gameta para um determinado locus. Com a cultura de células em meio líquido, podem-se obter mutantes, isto é, genótipos que ganharam ou perderam alguma característica específica. Com a cultura de embriões e meristemas, podem-se fazer trabalhos de criopreservação para conservar materiais em bancos de germoplasmas, com economia de espaço e dinheiro, especialmente em espécies de reprodução assexuada como batata, mandioca, abacaxi etc. Com a cultura de meristemas apicais, pode-se pensar em obter plantas livres de vírus, e com a cultura de gemas axilares (Fig.7), propagar milhares de plantas, com genótipos superiores, por exemplo resistentes a nematóides, Fusarium etc. (Fig.8). Na mesma linha de raciocínio, a embriogênese somática (Fig.9), pode fornecer grandes quantidades de plântulas que podem servir de base para plantios no campo, tanto na área florestal quanto na agronômica e na hortigranjeira. Por outro lado, a cultura de tecidos dá suporte técnico a trabalhos de transformação na área da genética e na obtenção de plantas transgênicas, hoje, assunto da moda e muito controvertido. No campo da aplicação básica, a cultura de tecidos dá suporte técnico também à bioquímica, à fisiologia vegetal, à fitopatologia e à citogenética. Na bioquímica, para estudo e dilucidação de rotas metabólicas. Na fisiologia, para estudos de crescimento e desenvolvimento, efeito de metais pesados etc., na fitopatologia, para estudos de toxinas; e na citogenética, para estudos de cromossomos ou aberrações cromossômicas (quebras cromossômicas). Apesar de toda essa diversidade de técnicas, a cultura de tecido é uma só, e o denominador comum de todas elas é: a assepsia, o explante, o meio nutritivo e os fatores ambientais: luz, temperatura, C02 e 02. Assepsia Será entendida como um conjunto de procedimentos para tornar um explante livre de microrganismos (bactérias, fun- Fig.2: Plântula de banana obtida in vitro e transferida para terra em casa de vegetação gos filamentosos, leveduras etc). A respeito de como evitar microrganismos que possam contaminar o explante, é inevitável o uso de antissépticos, sejam estes bacteriostáticos ou germinicidas. Esses antissépticos podem ser antibióticos ou de outra natureza, como álcoois (álcool etílico); halogênios (hipoclorito de sódio); sais de metais pesados (bicloreto de mercúrio), fungicidas orgânicos etc. Em relação à vidraria e aos meios de cultura, estes devem ser esterilizados para que se destruam todos os microrganismos, por calor seco (forno, ar quente) ou úmido (autoclave). Pinças bisturis e Fig.3: Calo friável oriundo de raízes de plântulas de eucalipto in vitro (Eucalyptus grandis x E.urophylla) demais utencilios metálicos para a manipulação do tecido podem ser esterilizados por flambagem direta, como por exemplo: lamparina com álcool ou bico de Bunsen na câmara de fluxo laminar, ambiente este axênico (livre de germes) portanto, adequado para o trabalho in vitro. Explante Dentro da terminologia da cultura de tecidos, em geral, explante é qualquer segmento de tecido oriundo de uma planta para iniciar uma cultura in vitro, geralmente com as vistas a estabeler um protocolo de plantas de genótipo superior. Assim, o explante pode ser um ápice radicular ou caulinar, uma gema axilar (Fig.7), um segmento de folha jovem (Fig. 10), uma antera, um ovário, um embrião zigótico, etc. Esses explantes poderão devir plantas diretamente, ou passar por uma etapa intermediária de calo, antes de a planta ser obtida. Um calo é uma massa de células não diferenciadas do ponto de vista organogênico (brotos, raízes, frutos etc), que está em contínua proliferação celular, podendo ser compactos, friáveis, esbranquiçados ou amarelados Fig. 3, etc. Podemos realizar indução de plântulas a partir de um calo, com hormôBiotecnologia Ciência & Desenvolvimento 17 Fig.4: Aspecto microscópico de um conglomerado de células em suspensão, obtidas a partir de calos friáveis de raízes de plântulas de eucalipto in vitro nios vegetais ou reguladores do crescimento (estes últimos de ação hormonal, mas de origem sintética). Assim, numa reviravolta espetacular (morfogênese) o calo começa a induzir brotos, mas a base molecular do fenômeno na célula, ainda não é especificamente conhecida, por isso também a dificuldade de extrapolar protocolos de micropropagação de uma espécie para outra, ou de uma variedade para outra. Contudo, muitas vezes, a incapacidade de induzir brotos pode estar relacionada com a poliploidia ou com aberrações cro- mossômicas. Por outro lado, discute-se muito se a obtenção de plantas via calo é um bom sistema para clonar plantas com fins comerciais. Embora a preocupação seja pertinente, há também nessa preocupação muita carga especulativa. Dentro da biologia, clone é um termo de origem botânica e representa a idéia de plantas que se originam de uma mesma matriz e, supostamente, possuem o mesmo genótipo. Em cultura de tecidos, isso não necessariamente pode ser verdade, por isso, então, essa dificuldade de aceitar sem suspeitas plantas via calo. Quando essa variabilidade morfogenética acontece a denominamos variação somaclonal, embora ela seja indesejável para a propagação clonal, ela possui muito potencial no estudos de melhoramento de plantas. Meio nutritivo a) sais minerais Se uma planta no solo precisa para crescer de elementos minerais que são absorvidos pelas raízes, quanto mais um explante, que é, por definição, um pedaço de tecido separado da planta mãe. E que elementos minerais são esses? Os estudos da nutrição de plantas provenientes do âmbito da fisiologia vegetal informam que os elementos que compõem o meio nutritivo da cultura in vitro devem pertencer à categoria dos essenciais, isto é, a planta não se desenvolve na ausência deles. Existem dois grupos deles: os macronutrientes e os micronutrientes. Entre os primeiros, podem-se citar: fósforo, magnésio, nitrogênio etc.; entre os segundos, boro, manganês, cobre etc. Os primeiros são adicionados em forma de sal, acima de 100 mg/l até o máximo de 2.500, como no caso de KNO3 em B5, enquanto os segundos, na quantidade de fração de miligramas, uns poucos miligramas por litro (máximo 27 mg no caso do Fe em FeSO4. 7H2O). Não sendo o selênio, rubídio e outros, essenciais para a planta, não formam parte dos meios nutritivos mais utilizados: tais como MS, B5, White, Heller. Embora a manipulação desses meios constitua um processo de receituário, existe muito fundamento fisiológico na sua utilização; por exemplo, balanço líquido de cátions e ânions na solução; antagonismo iônico (o aumento da absorção de um diminui a do outro); percentagem crítica de um elemento (consumo de luxo); influência do pH na disponibilidade dos sais para a planta; absorção diferencial de íons entre tipos de planta, exemplo: árvores e gramíneas, estas últimas mais exigentes em bases; transporte ativo de íons a nível celular; enfim, são conhecimentos que podem auxiliar de modo importante no estabelecimento de um protocolo de cultura de tecidos para uma espécie. b) componentes orgânicos: Fig.5: Fruto de mamão (Carica papaya L. cv tainung 1) mostrando aspectos e quantidade de sementes por fruto 18 Biotecnologia Ciência & Desenvolvimento sacarose Um meio nutritivo também possui componentes orgânicos, entre eles, sacarose, vitaminas e inositol, para citar os mais utilizados. A sacarose é importante como fonte de carbono para alimentar a glicólise e o ciclo de Krebs, devido a que, inicialmente, o tecido, explante, não é suficientemente autotrófico. De um outro ponto de vista, a sacarose, quando em presença de auxina, pode contribuir para a diferenciação do câmbio em benefício de novas formações de xilema e floema. Em alho, altas con- centrações de sacarose (6%) têm melhorado ostensivamente sua bulbificação. Sua concentração normalmente varia entre 1% a 3 %, mas, por efeito da autoclavagem e do pH, sua concentração no meio pode variar, já que é hidrolizada em glicose e frutose, sendo que esses limites não são claros e esse problema pode tornar-se limitante para o explante ou para a planta. vitaminas São substâncias requeridas por animais e plantas, embora estas últimas sejam mais autônomas quanto à sua Fig.6: Germinação de sementes de mamão sob condições in vitro, após síntese. As vitaminas são importantes ter-se retirado a sarcotesta e realizado sua assepsia fatores catalíticos de rotas metabólicas na célula. Nos trabalhos pioneiros da cultura de tecido, os requerimentos de ser encontrado como pertencente ao mentos variam segundo as plantas, senvitaminas foram satisfeitos mediante su- complexo vitamínico B e necessário ao do que, no meio de cultura, as monocoplementos de composição pouco co- crescimento de leveduras e de células tiledôneas são muito sensíveis à sua nhecidos, como extrato de leveduras, de animais (mamíferos) in vitro. falta. No MS, o meio mais universalmenleite de côco, hidrolizado de proteínas, Na área de cultura de tecidos vege- te utilizado, sua prescrição é 100 mg/ extrato de malte, etc. Hoje, apesar de tais, muitos o consideram uma fonte litro. essa tendência não haver desaparecido, complementar de carboidrato, mais o mais freqüente, é que, os meios nutri- que um composto vitamínico, e que agentes gelificantes tivos incorporem vitaminas específicas desempenha um papel importante na São necessários, considerando que o como B1 (tiamina), B6 (piridoxina), áci- formação de pectinas e hemicelulose explante e as plantas obtidas devem do pantotênico, ácido nicotínico, ácido na parede celular. Seu uso data, apro- ficar sobre um suporte, para não afundaascórbico etc. ximadamente, de 1951, e é decorrente rem. Em geral, os meios sólidos são Não obstante as plantas sepreferidos aos de cultura líquirem capazes de sintetizar suas da, sendo que estes últimos são próprias vitaminas, fica a dúvida mais caros pelo fato de exigirem se a incorporação no meio é um agitador para evitar o afogasempre necessária. Contudo, em mento ou a hipoxia do explante. se tratando de drenos, raízes, por O agente gelificante forma exemplo, a suposição de que com a água um gel que funde a nem todos os seus requerimen100°C e que solidifica por volta tos nutricionais orgânicos sejam dos 45°C; não é hidrolizado por sintetizados ficou evidenciada já enzimas e, aparentemente, não em 1930, quando pesquisadores reage com o resto de ingrediencomo Bonner, Robbins, White tes do meio. etc, observaram que, na presença O ágar-ágar é um tipo de de algumas vitaminas (tiamina, agente gelificante de natureza Fig.7: piridoxina, ác. nicotínico, etc.), o polissacarídica produzido por Prévia assepsia, gema caulinar de abacaxi (Ananas algas (Gelidium amansii), sencrescimento das raízes (tomate, ervilha e rabanete) melhorava comosus L. Merr. Cv Pérola) cultivada in vitro do que sua composição em posensivelmente. lisacarídeo pode variar de 50% a Com respeito à quantidade a ser do fato de que, em muitos casos, sua 90 %, por isso, a procedência do mesmo incorporada, ela varia desde fração de incorporação ao meio de cultura pro- é importante. Por outro lado, a autoclamiligramas até 10 miligramas por litro, moveu diferentes eventos: formação vagem pode hidrolizá-lo se o pH do podendo ser esterilizada por filtração ou de gemas, crescimento de calos etc. meio está ácido, fazendo com que perca autoclavagem. Convém lembrar que, em algumas plan- firmeza e, dependendo da marca, o tas, o myo-inositol está conjugado com agente gelificante pode apresentar immyo-inositol a auxina, sendo essa uma forma de purezas, como cloro, bário, sulfato etc, Este composto também descrito como estocar ou transportar aquele hormô- que podem afetar o crescimento dos meso-inositol, é um isômero do inositol nio, ou, conjugado, em alguns tipos de explantes. o que, apresenta importância biológica, sementes, com o ácido fosfórico, forGelrite (Merck) é um outro tipo de já que participa da internalização de mando o ácido fítico. agente gelificante. Produzido pela bacestímulos externos a partir de receptores A respeito da sua aplicação, a infor- téria Pseudomonas elodea, é usado em de membrana. Na literatura médica, pode mação disponível é que os requeri- menor quantidade (2,5 g/l); uma vez Biotecnologia Ciência & Desenvolvimento 19 solidificado, é transparente, o que facilita a observação da raiz e focos de contaminação. Com íons, como Ca++ e Mg++, forma um elo mais firme do que com cátions monovalentes (K+ e Na+). Devem-se ter cuidados porque, às vezes, os agentes gelificantes, incluída a agarose, podem induzir hiperhidricidade (vitrificação) nos brotos ou plântulas obtidas. A hiperhidricidade é uma anomalia do tecido, na qual este fica rígido e quebradiço (glassy shoot), havendo malformação de estômatos na plântula e dificuldade para enraizar. É um fenômeno complexo e depende também de outros fatores como umidade, amônia, citocinina, pH etc. hormônios Estes representam o alma mater da cultura de tecido, porque são eles que direcionam o processo morfogenético. Agrupam-se tradicionalmente em 5 grupos: auxinas, citocininas, giberelinas, etileno e ac. absícico, sendo, os três primeiros os mais usados na micropropagação. Por exemplo, sob condições in vitro, uma planta de abacaxi pode ser clonada através de uma gema axilar, a qual, é induzida a produzir inúmeros brotos com citocinina, que, depois de alongados com giberelinas, podem ser enraizados com auxina e transferidos para terra em casa de vegetação. Quando se fala de hormônios, é sempre oportuno registrar uma diferenciação terminológica importante. Trata-se dos chamados reguladores de crescimento, que são substâncias que têm ação similar aos hormônios, porém, têm origem sintética. Assim, o ácido indol acético (AIA) e o Picloram são duas auxinas, mas, enquanto o primeira é hormônio, a segunda é um regulador de crescimento, que, na prática laboratorial, terminam sendo chamados de hormônios. Em geral, tanto uns como outros são usados na ordem de uns poucos µM até 40 µM. No caso do meio ter carvão ativado, pode-se aumentar a concentração em até 100 µM. Por outro lado, as auxinas e giberelinas 20 Biotecnologia Ciência & Desenvolvimento tidas em freezer ou refrigerador. Fatores ambientais: Fig.8: Plântula de abacaxi oriunda de uma gema caulinar. Por micropropagação milhares dessas plântulas (clones) podem ser obtidas podem ser dissolvidas em NaHO (0,1 1 N) e as citocininas em HCl também na faixa de 0,1 - 1 N. Em doses maiores produzem efeitos tóxicos ou teratológicos ou ainda inibirem a fotosíntese, como é o caso da auxina sintética 2,4D ( ácido 2,4-diclorofenoxiacético). No caso de seu uso em quantidades pequenas, é aconselhável obtê-los de soluções estoques que devem ser man- Fig.9: Embriões somáticos de café, obtidos a partir de explantes foliares de cv Rubi luz Luz e temperatura são dois fatores importantes nas salas de cultura, onde devem ser controlados para que as plantas ali mantidas se desenvolvam adequadamente. São raras as salas de cultura com iluminação constante. A luz é importante para a planta sob três pontos de vista. Do ponto de vista da fotossíntesse, da fotomorfogênese e do fototropismo; por isso, a sua inclusão numa sala de cultura. Nessa sala, a luz deve seguir um determinado fotoperíodo, por exemplo, 16 h luz e 8 de escuro, sendo que, nas prateleiras a irradiância (densidade de fluxo radiante por superfície, W m-2, ou densidade de fluxo de fótons, µmol m-2 s-1)1 pode variar de 8 a 15 W m2 . Embora nas salas seja usada luz branca fluorescente, a composição espectral pode variar conforme as marcas comerciais oferecidas. Algumas podem apresentar mais irradiação na região do azul / violeta (perto de 71 kcal/Einstein ); outras, na região do laranja / vermelho ( cerca de 43 kcal/Einstein) Essas variações podem representar impactos diferentes na cultura in vitro, segundo a espécie em questão. Assim, em alguns casos, uma luz mais rica em vermelho que azul pode, por exemplo, estimular melhor a indução de raízes adventícias. Já em outros casos, a luz azul pode estimular mais a brotação de calos que outros comprimentos de onda, mas esta também pode contribuir a quebrar a molécula de AIA. De modo geral, a luz branca estimula a indução de brotos, porém, tende a inibir a indução de raízes. Entretanto, é bom lembrar que o nível de irradiância é importante na resposta morfogenética. Sendo assim, uma irradiância baixa (3 W m-2) pode ser mais efetiva que uma irradiância alta (12 W m-2) na indução de brotos a partir de calos, ou vice-versa. Pode-se contornar o problema de se obterem maiores ou menores irradiâncias com o número de lâmpadas por pra- teleira ou determinando que altura deverá ter a prateleira. Para a cultura in vitro, irradiâncias maiores que 15 W m-2 podem reduzir a fotosíntesse das plantas, o que não necessariamente pode acontecer em condições de campo. Irradiâncias maiores na sala de cultura obviamente contribuirão para elevar a temperatura dos frascos, embora se saiba que, em café, irradiâncias altas não reduziram a fotosíntesse quando a temperatura foliar permaneceu constante a 25 °C. temperatura Em geral, nas salas de cultura, a temperatura a ser usada varia entre 24º e 27°C. É importantíssimo contar com um sistema de refrigeração acionado por termostato para manter fixa a temperatura. O fato de os reatores das lâmpadas ficarem fora da sala de cultura, ajuda, ou senão, o uso de reatores eletrônicos, que não esquentem. Mesmo com todas as precauções de manter as variações de temperatura sob controle, sempre temos nas placas ou tubos de ensaios problemas de condensação da água nas paredes desses materiais. Existem câmaras ou incubadoras que evitam esse problema, mas seu preço é proibitivo. Com respeito às necessidades eventuais de temperaturas alternadas entre o dia e a noite, é preferível usar incubadoras para lograr isso. 02 e C02 Existe pouca informação sobre o microclima gasoso dentro do tubo de ensaio. De qualquer maneira, a presença deles é importante para a respiração do explante ou para a fotossíntesse da plântula. No que diz respeito à respiração aeróbica-(glicólise + ciclo de Kreb), considerando apenas um tecido heterotrófico, e em se tratando de tubos de ensaio, podemos supor que, no volume disponível acima do meio sólido, existe, inicialmente, uma concentração e uma pressão parcial do 02 equivalente à atmosférica (25 °C e 1 atmosfera); isso é:, 8,6 mM e 0,2 atmosfera, respectivamente, embora esse valor, no decorrer do tempo, possa Fig.10: Explante foliar de café, mantido no escuro, insinuando, após 30 dias, formação de calo pela ação de um regulador de crescimento presente no meio nutritivo diminuir no interior do tubo e a respiração anaeróbica, supostamente, não começará a operar antes da repicagem seguinte do material. É provável que, em níveis de 0,3 mM e 0,07 atmosfera de pressão parcial do 02 no espaço interior do tecido (explante, calo etc.) essa respiração rapidamente comece a operar e, com isso, iniciar-se a morte do explante. Isso levanta a questão sobre a transferência periódica do material, o volume do frasco e a permeabilidade ou não da tampa. A concentração de C02 de 350 µl / l no ambiente externo não necessariamente reflete a concentração de C02 no ambiente interno do tubo de ensaio. Muitas vezes ela pode ser maior no tubo, devido à respiração da plântula no período noturno. Por outro lado, no caso de uma plântula com irradiância de 60 µm m-2 s-1 e aquela quantidade de C02 rapidamente deveria ultrapassar o ponto de compensação. Contudo, parece que essas interações são mais complexas do que parecem, porque existem evidências de que a presença de sacarose no meio pode deprimir a rubisco, reduzir a fotosíntesse e, conseqüentemente, a utilização de C02 e, dessa forma, a plântula não apresentará sinais de crescimento satisfatório e provocará reações de desconcerto no observador, sobre as possíveis causas de seu não crescimento. Tentou-se aqui apresentar uma visão sinóptica do que seja a cultura de tecido in vitro e sua importância para a biologia de plantas. No campo econômico, sem ir muito longe, basta lembrar que, através dessa técnica, a floricultura movimenta milhões de dólares e de plantas anualmente. Na Embrapa /Cenargen (Brasília DF) usamos essa técnica em quatro frentes: na micropropagação , na criopreservação, na conservação de germoplasma e na transformação, todas elas, visando a uma agricultura mais moderna e competitiva para o país. Bibliografia BARRUETO CID, L.P. (Ed.). Introdução aos hormônios vegetais. Brasília: Embrapa Recursos Genéticos e Biotecnologia,2000. 205 p. No prelo. BARRUETO CID, L.P.; MACHADO, A. C.G.; CARVALHEIRA, S.B.C.; BRASILEIRO, A.C.M. Plant regeneration from seedling explants of Eucalyptus grandis x E. urophylla. Plant Cell, Tissue Organ Culture, v.56:17-23,1999. BUCHANAN, B.B.; GRUISSEM, W.; JONES, R.L. Biochemistry & molecular biology of plants. Rockville: American Society of Plant Physiologist, 2000. 1367 p. DODDS,J.H.; ROBERTS, L. Experiment in plant tissue culture. London: Cambridge Unv. Press, 1982. 178 p. FOSKET, D.E. Plant growth and development. A molecular approach. San Diego: Academic Press, 1994. 580 p. GEORGE, E.F. Plant propagations by tissue culture. Part 1-2. Edington: Exegetics Ltd, 1993/1996. 1361 p. PIERIK, R.L.M. In vitro culture of higher plants. Drodrecht: Martinus Nijhoff Pub.,1987. 344 p. REDLIN, S.C.; CARRIS, L.M. Endophytic fungi in grasses and woody plants. Minnesota: APS Press, 1996. 223 p. Biotecnologia Ciência & Desenvolvimento 21 Transgênico resistente a PESQUISA GEMINIVIRUS Feijoeiro geneticamente modificado imune ao vírus do mosaico dourado Francisco José Lima Aragão PhD em Biologia Molecular [email protected] Giovanni Rodrigues Vianna MsC. , Doutorando em Biologia Molecular Margareth das Mercês Cerqueira Albino Graduanda em Biologia Bárbara Barreto Andrade Dias Graduanda em Biologia Embrapa Recursos Genéticos e Biotecnologia Brasília, DF Josias Corrêa Faria PhD em Fitopatologia Embrapa Arroz e Feijão Santo Antônio de Goiás, GO [email protected] Fotos cedidas pelos autores s plantas são, direta ou indiretamente, a principal fonte de combustíveis, de remédios, de material de construção e, principalmente, de alimentos. Talvez, por essa importância vital, não se deva ficar surpreso que o homem, desde tempos remotos, tenha se preocupado em desenvolver os tipos que melhor satisfaçam às suas necessidades. A sistematização dos métodos de obter tais plantas resultou na ciência do Figura 1: A) Estrutura da região intergênica (RI) do BGMV mostrando o elemento iteron (→→), o TATA box e a estrutura em stem-loop. B) Detalhe da origem de replicação do BGMV, os sítios de clivagem e ligação da proteína viral Rep. A seqüência nanomérica comum a todos os geminivirus está indicada pelo semi-círculo pontilhado. O último A do nanonucleotídio (em vermelho) é a base onde ocorre o corte do DNA durante a replicação 22 Biotecnologia Ciência & Desenvolvimento melhoramento genético de plantas. Problemas como ausência da característica de interesse dentro da espécie e incompatibilidade sexual sempre foram empecilhos para se obterem plantas ou organismos com as combinações genéticas desejadas pelos pesquisadores para satisfazer as crescentes demandas da sociedade; em outras palavras: a variabilidade genética existente na natureza não poderia ser explorada em todo o seu potencial. Gradativamente, os pesquisadores foram selecionando as melhores raças de plantas e de microorganismos. Portanto, a ação do homem foi a de retirar da natureza os organismos que tinham um melhor conjunto de genes capaz de produzir, eficientemente, produtos para a alimentação, a saúde humana e o uso industrial. Quando o rendimento ainda é baixo, o homem utiliza-se de novos conhecimentos científicos para fazer o melhoramento genético, através de cruzamentos, indução de mutações no genoma dos organismos, etc. Em tempos mais recentes, com os avanços na cultura de tecidos, biologia molecular, bioquímica, etc., o homem passou a melhor entender os organismos e a poder trabalhar mais intensamente o potencial do material genético (genes) disponível. Com a tecnologia do DNA recombinante, o homem pôde manipular os genes de interesse, e utilizando-se de várias tecnologias, transferílos para a espécie desejada, sem ter que passar pela fecundação. Resistência derivada do patógeno Sanford & Johnson (1985) foram os primeiros a propor a obtenção de resistência a patógenos em plantas geneticamente modificadas, pela utilização de seqüências genômicas dos próprios patógenos. Na verdade, esse conceito havia sido empregado há várias décadas. Antes mesmo de se conhecer a composição dos vírus de plantas, por volta de 1929, foram feitas observações de que inoculando-se plantas de fumo com uma estirpe fraca de vírus do mosaico do fumo, ao tentar reinocular com uma estirpe que causava sintomas mais severos, as plantas encontravam-se protegidas contra a super infecção (McKinney, 1929). Isto veio a se chamar proteção cruzada. O mecanismo de tal proteção nunca foi completamente desvendado. Nos anos 80, com o desenvolvimento de técnicas moleculares, foi possível testar a hipótese de que a proteção era mediada pela capa protéica (CP) do vírus, e que a resistência era válida para vírus homólogo ao que forneceu a capa. Sem se importar com qual que fosse o mecanismo, a tecnologia recebeu a denominação de pathogen derived resistance, que traduzimos como resistência derivada do patógeno. Uma seqüência lógica do desdobramento desse conceito foi o desenvolvimento de estratégias para resistência a viroses, uma vez que os vírus, apesar de terem biologia molecular complexa, são, na maioria dos casos, estruturalmente mais simples que outros organismos causadores de doenças em plantas. A primeira estratégia empregada foi a expressão da CP em plantas geneticamente modificadas (PGM) através da tecnologia do DNA recombinante. O primeiro caso de sucesso foi a expressão da CP do vírus do mosaico do fumo (TMV) em plantas de fumo, gerando linhagens resistentes ao vírus (Powell et al 1986). Desde então, uma série de outras tentativas de utilização dessa estratégia foram realizadas, com a utilização de genes estruturais e não estruturais (para uma revisão, ver Souza & Gonsalves, 1999). Brevemente pôde-se listar: (1) expressão da capa protéica, (2) uso de satélites, (3) RNA senso e antisenso, (4) RNAs defectivos, (5) expressão da replicase, (6) expressão de proteínas do movimento, (7) expressão de anticorpos (plantbodies). Em nossos laboratórios (Embrapa Arroz e Feijão & Embrapa Recursos Genéticos e Biotecnologia), temos como objetivo introduzir resistência ao vírus do mosaico dourado do feijoeiro (BGMV) em Phaseolus vulgaris, através de métodos de biologia celular e molecular. Não existe imunidade nem mesmo alto grau de resistência a essa virose em germoplasma de Phaseolus spp. Paulo (Costa 1965). Seu agente transmissor, a mosca branca (Bemisia tabaci Gennadius), foi também identificado. Subseqüentemente, um vírus de partículas geminadas foi identificado, associado às plantas mostrando sintomas de mosaico. Esse vírus foi então denominado vírus do mosaico dourado do feijoeiro - VMDF (em inglês: bean golden mosaic virus - BGMV), um geminivirus. No início dos anos 70, as plantações de feijoeiro nos estados de São Paulo, Paraná e Minas Gerais foram severamente atingidas pelo mosaico dourado. Esse fato foi atribuído ao avanço da cultura da soja e de outras culturas hospedeiras da mosca branca. Em algumas regiões, devido à grande incidência da doença, os agricultores tiveram como única opção parar com o plantio do feijão. Essa doença está hoje disseminada por todas as áreas produtoras de feijão do Brasil. Doença semelhante é encontrada.em outros países das Américas, tais como Cuba, República Dominicana, Porto Rico, Jamaica, Estados Unidos, México, Belize, Guatemala, El Salvador, Honduras, Nicarágua, Costa Rica, Panamá, Venezuela e Colômbia. No Brasil, em condições de campo, as perdas ficam em torno de 40% a 85%, podendo chegar a 100% (Menten et al 1980, Gálvez & Morales 1989), dependendo da cultivar, do estágio das plantas quando infectadas e do isolado do vírus. Quanto aos sintomas, inicia-se pelas nervuras das folhas, que, numa etapa mais avançada, exibem um amarelo brilhante na maior parte do limbo foliar, formando um mosaico. Ocorrem ainda distorções foliares, nanismo, malformação de vagens e sementes. Além disso, as sementes provenientes de plantas infectadas têm a germinação afetada. O mosaico dourado do feijoeiro O Vírus O mosaico dourado do feijoeiro foi inicialmente descrito pelo Dr Álvaro Santos Costa como uma doença que, inicialmente não teria importância econômica no Estado de São O BGMV consiste numa partícula icosaédrica, que contém DNA de fita simples, circular, como material genético. Trata-se de vírus com genoma dividido em dois componentes, de- Figura 2: Plantas transgênicas de feijoeiro cultivadas in vitro Biotecnologia Ciência & Desenvolvimento 23 Figura 3: Linhagem imune ao vírus do mosaico dourado do feijoeiro. Esquerda: planta não transgênica inoculada com o virus mostrando os sintomas de mosaico dourado. Direita: planta transgênica inoculada com o vírus sem qualquer evidência de sintoma nominados A e B, sendo encapsidados independentemente. Replica-se no núcleo de células do floema do hospedeiro, através de um mecanismo conhecido como círculo rolante, tendo DNA de fita dupla como intermediário de replicação. O DNA A contém os genes necessários para a replicação e encapsidação da progênie viral, enquanto o DNA B contém os genes requeridos para o movimento célula-a-célula e a longa distância, gama de hospedeiros e desenvolvimento de sintomas (Timmermans et al 1994). Ambos os componentes são necessários para a infecção sistêmica de plantas. Exceto por uma seqüência de aproximadamente 200 nucleotídeos, denominada de região comum, os dois componentes não apresentam similaridade significativa em suas seqüências de nucleotídeos. O DNA A contém o gene da CP (ORF AV1, gene cp) e três no sentido complementar (ORFs AC1, AC2 e AC3), correspondentes aos genes rep (que codifica o gene para uma proteína associada à replicação); trap (transactivation protein, que é o fator de transcrição que atua in trans no promotor de genes de sentido viral 24 Biotecnologia Ciência & Desenvolvimento (cp e mp), - Sunter & Bisaro 1991, 1992; na presença de trap, a expressão do promotor da proteína capsidial foi aumentada em cerca de 60 a 90 vezes Brough et al 1992), e ren (que é um fator de amplificação da replicação viral, que, embora não seja essencial para que a replicação ocorra, provoca um acúmulo de DNA viral muito maior quando está presente). No DNA B, estão localizadas as ORFs BV1 e BC1, correspondentes aos genes ns e mp, respectivamente (Palmer & Rybicki 1998). A proteína ns (nuclear shuttle), anteriormente BR1 ou BV1, é necessária para o tráfico intracelular de DNA viral do núcleo para o citoplasma, enquanto que a mp (movement protein), anteriormente BL1 ou BC1, está envolvida no movimento do DNA viral célula-a-célula, via plasmodesmas. A proteína Rep é a única essencial para a replicação. O DNA de vários isolados do BGMV foram completamente seqüenciados. O vírus do mosaico dourado amarelo do feijoeiro (BGYMV) da República Dominicana e da Guatemala (Faria et al 1994); de Porto Rico (Howarth et al., 1985) e (BGMV); e o do Brasil (Gilbertson et al., 1993 - Acesso do GenBank M88686 e M88687). O BGMV é transmitido na natureza pela mosca branca, de uma forma circulativa. Os isolados do BGYMV podem ser transmitidos por inoculação mecânica, isto é, através do extrato de uma planta infectada, friccionado sobre a folha de uma planta sadia. Entretanto, o isolado brasileiro, BGMVBR, não é transmitido dessa forma (Costa, 1965). Recentemente, pode-se transmitir um isolado brasileiro do vírus, clonado no vetor pBSKS+ (Stratagene), através do processo de biobalística (Aragão et al., 1995). O BGMV não é transmitido através das sementes (Costa, 1965). Cerca de 151 espécies e/ou isolados de Geminividae foram inteiramente seqüenciados. Um exame da seqüência dos vários geminivirus revela a existência de um nanômero conservado em todas as espécies, localizado na região intergênica tanto do DNA-A quanto do DNA-B. Esse nanômero (TAATATTAC), chamado de SCE (elemento estruturalmente conservado), foi logo reconhecido como a origem de replicação (ori) e como o sítio de clivagem do DNA pela proteína associada à replicação. Esse elemento SCE está localizado dentro de uma seqüência conservada de 30 nucleotídeos, com potencial para formação de uma estrutura em forma de grampo. A manutenção conformacional dessa estrutura é fundamental para sua função de origem de replicação (Orozco & Hanley-Bowdoin, 1996). Outro elemento, chamado de iteron, foi localizado a montante dessa estrutura de grampo, repetido duas vezes (no caso do BGMV), ao lado da região TATA (Figura 1), que foi caracterizada como a região de ligação da Rep ao DNA (Walker et al 1982; Lazarowitz et al., 1992; Fontes et al., 1994). Os iterons são específicos para cada vírus. Por exemplo, os iterons do BGMVBR são diferentes daqueles em BGYMV-GA e em BGYMV-PR No entanto, sua posição relativa e sua orientação são conservados dentro de Geminiviridae filogeneticamente relacionados. A proteína associada à replicação (Rep), codificada pelo gene rep, tem sido alvo de um grande número de estudos nos últimos anos. Essa proteína possui várias funções associadas à replicação: (1) dirigir o complexo re- plicativo para a origem de replicação; (2) desenovelar o DNA molde (atividade de helicase); (3) clivar o DNA e iniciar o mecanismo de círculo rolante; e (4) separar os genomas após a replicação (atividade de nuclease e ligase). Além da sua principal atividade na replicação, a Rep está envolvida na sua autoregulação: repressão de sua própria síntese ao nível de transcrição. Uma seqüência consenso (NTPbinding motif) (EGX4GKTX22DD) foi encontrada na replicase de doze geminivírus analisados (Hanson et al 1995). Experimentos feitos in vivo mostraram que uma simples mutação na replicase do BGMV [de lisina (K) para histidina (H), ou de ácido aspártico (D) para arginina (R)], anulou a replicação viral e o aparecimento dos sintomas nas plantas de feijoeiro inoculadas (Hanson et al 1995). Mutações de lisina (K) para histidina (H) na Rep do tomato yellow leaf curl geminivirus (TYLC) reduziram significativamente a atividade de ATPase dessa enzima. Outro motivo conservado (DVKXYXXKD) no domínio amino terminal da Rep dos geminivirus foi identificado como sítio de clivagem e ligação ao DNA, sendo a tirosina (Y) presente nesse consenso o aminoácido ativo. A estratégia de RNA antisenso foi empregada para vírus de diferentes grupos, tais como comovirus, potyvirus, tobamovirus e outros. Na maioria dos casos, usou-se o mRNA antisenso para o gene da CP. Em geral, obtevese apenas um nível limitado de proteção, isto é, a proteção ficou limitada a baixos níveis de inóculo viral. Entretanto, o uso de RNA antisenso tem sido bastante eficiente no bloqueio da expressão de genes nucleares em plantas. Esses resultados serviram como suporte para a hipótese de que essa estratégia fosse útil no bloqueio de vírus com parte de seu ciclo de vida e de replicação ocorrendo no núcleo, tais como geminivírus e caulimovírus (Wilson 1993). Os mecanismos de ação de seqüências antisenso ainda são apenas parcialmente compreendidos. Em ge- Estratégias para resistência Em geminivirus, a expressão da capa protéica não tem apresentado resultados satisfatórios. De fato, plantas transgênicas de fumo expressando a CP do abutilon mosaic geminivirus mostraram sintomas parecidos com os da infecção pelo vírus, e proporcionais à expressão do gene (Wilson 1993). Provavelmente isso se deve ao fato de a CP não ser essencial à infecção e ao desenvolvimento de sintomas em plantas infectadas por geminivirus. Assim, têm sido propostas outras estratégias para obtenção de plantas transgênicas resistentes a geminivírus, tais como o RNA antisenso e a expressão da CP e proteína associada a replicação mutada. Figura 4: Vagens e sementes de plantas transgênicas (T) e de plantas não transgênicas (NT), ambas inoculadas com o vírus do mosaico dourado do feijoeiro ral, é proposto que essas seqüências interfeririam, em nível traducional, de forma direta ou indireta, podendo ser em nível nuclear ou citoplasmático. A nível nuclear, a hibridização RNAantisenso e mRNA pode interferir no processamento do pré mRNA, inibindo o splicing, ou ainda o transporte do mRNA do núcleo para o citoplasma. Esse dúplex formado pode ser reconhecido pela RNase H a nível nuclear ou citoplasmático, e, subseqüentemente, o mRNA é degradado. No nível citoplasmático, a interação do RNA antisenso e do mRNA pode interferir na ligação de fatores de iniciação de tradução ou inibir diretamente a tradução do mRNA pelos ribossomos. Interações RNA antisenso e DNA podem também ocorrer, sendo esse híbrido também substrato para a RNase, que hidrolisa a fita de RNA. Os fenômenos de co-supressão, podem também ser responsáveis pela resistência a vírus em plantas transgênicas. A presença de uma seqüência de DNA no genoma da planta poderá suprimir a expressão (silenciar) do próprio gene e de um gene homólogo presente. Assim, a presença de uma seqüência do vírus integrado ao genoma da planta poderá silenciar sua expressão por parte do próprio vírus, quer por interações entre os genes, quer por metilação ou ativação de mecanismos específicos de degradação de RNA. Seqüências que englobam os genes rep, trap, ren, e mp do BGMV-BR foram posicionadas em antisenso, sob controle do promotor 35S do cauliflower mosaic virus (35S CaMV). Essa construção foi então utilizada para obtenção de plantas transgênicas de feijoeiro (Aragão et al 1996). Isolados do BGMV de alguns estados do Brasil (Goiás, São Paulo e Pernambuco) foram caracterizados no nível molecular e achou-se que havia uma grande homologia entre suas seqüências (75-100%) (Faria e Maxwell, 1999). Portanto, considerase que o isolado utilizado nesse estudo representa os demais isolados exisBiotecnologia Ciência & Desenvolvimento 25 tentes no país. As plantas transgênicas com as seqüências do BGMV foram autofecundadas durante 4-5 gerações e, então, inoculadas com o vírus. A inoculação foi feita com a utilização de moscas brancas virulíferas. Algumas linhagens transgênicas não apresentaram diferença significativa nos sintomas em relação às plantas não transgênicas. Entretanto, duas linhagens mostraram um retardamento no aparecimento dos sintomas, além desses serem mais fracos que aqueles normalmente apresentados pelas plantas controle. Além disso, uma titulação do vírus através de análises por Southern blot nas plantas inoculadas mostrou que havia uma quantidade inferior de DNA viral nas plantas transgênicas (Aragão et al., 1998). Transdominantes letais Embora os resultados tenham sido bastante animadores, o nosso objetivo principal ainda era a obtenção de plantas imunes ao vírus, isto é, linhagens nas quais não ocorresse replicação viral. Assim, outra estratégia foi proposta, denominada transdominância letal. Essa estratégia envolve a criação de uma Rep não funcional que interferiria com a ligação do tipo normal de Rep produzido pelo vírus (Hanson et al., 1995). A mutagênese da proteína Rep de BGMV mostrou que a mutação de um códon no sítio envolvido na etapa de corte do DNA (Hoogstraten et al., 1996) ou no motivo de ligação e transferência de nucleosídeo trifosfato - NTP-binding motifs (Hanson et al., 1995) são letais. Esses dois motivos são conservados em todas as proteínas Rep, e são sítios atrativos para construir plantas transdominantes letais (Hanson et al., 1999). Obtivemos plantas transgênicas contendo o gene rep com a mutação D262R (ácido aspártico da posição 262 para arginina), que inibiu eficientemente a replicação do DNA A, em trans, em experimentos com células de fumo. Entre as plantas transgênicas de feijão obtidas, foi possível conseguir a completa resistência ao vírus. Essas plantas estão, no momento, na terceira geração, 26 Biotecnologia Ciência & Desenvolvimento apresentando o mesmo comportamento. Na próxima fase será realizado um estudo mais detalhado do comportamento das plantas transgênicas de feijoeiro em condições de campo. Nesses estudos, seriam avaliadas as interações das plantas transgênicas com outras plantas do ambiente agrícola, e também a estabilidade da expressão dos genes introduzidos. Dessa forma, devem ser avaliadas as questões relativas à biossegurança. Além disso, há ainda necessidade de estudo do comportamento desses genes no que diz respeito a fatores relacionados com a interação destes e a complexa fisiologia dessas plantas submetidas a estresse natural nas condições agroclimáticas tropicais. Entre outras conclusões do nosso trabalho, pode-se afirmar que: a) É possível a transformação consistente de feijoeiro via método de biobalística; b) O gene rep mutagenizado no motivo relacionado com a ligação de nucleosídio trifosfato, expresso em trans, inibiu completamente a infecção das plantas, pelo menos quanto ao vírus homólogo ao de onde fora extraído o gene. Agradecimentos Os autores agradecem o excelente suporte técnico dado pelas seguintes pessoas: Elsa O. P. L. Nogueira, Vanderlino M. Santana., Warley Almeida e Luiz Lemos. Este trabalho foi financiado pela EMBRAPA, PADCT, CNPq. Referências Bibliográficas Aragão FJL, Brasileiro ACM, Ribeiro SG, Faria JC & Rech EL 1995 Fitopatologia Brasileira 20:642-644. Aragão FJL, Barros LMG, Brasileiro ACM, Ribeiro SG, Smith FD, Sanford JC, Faria JC & Rech EL 1996 Theoretical Applied Genetics 93:142150. Aragão FJL, Ribeiro SG, Barros LMG, Brasileiro ACM, Maxwell DP, Rech EL, Faria JC 1998 Molecular Breeding 4:491-499. Brough C L, Sunter G, Gardiner W E & Bisaro D M 1992 Virology 187:1-9 Costa A S 1965 FAO Plant Prot Bull 13:121-130. Faria JC & Maxwell DP 1999 Phytopathology 89:262-268. Faria J C, Gilbertson R L, Hanson S F, Morales F J, Ahlquist P, Loniello A O & Maxwell D P 1994 Phytopathology 84:321-329. Fontes EPB, Gladfelter H J, Schaffer RL, Petty ITD & Hanley-Bowdoin L 1994 Plant Cell 6:405-416. Gálvez G E & Moráles F J 1989 Bean Production in the Tropics CIAT Cali, Colombia. Gilbertson R L, Faria J C, Ahlquist P & Maxwell D P 1993 Phytopathology 83:709-715. Hanson S F, Hoogstraten R A, Ahlquist P, Gilbertson R L, Russell D R & Maxwell D P 1995 Virology 211:1-9. Hanson S F & Maxwell D P 1999 Phytopathology 89:480-486. Hoogstraten R A, Hanson S F & Maxwell D P 1996 Mol Plant-Microbe Interact 9:594-599. Howarth A J, Caton J, Bossert M & Goodman R M 1985 PNAS USA 82:3572-3576. Lazarowitz S G 1992 CRC Crit Rev Plant Sci 11:327-349. McKinney HH 1929 Journal Agricultural Research 39:557-578. Menten JOM, Tulmann Neto A & Ando A 1980 Turrialba 30:173-176. Orozco BM & Hanley-Bowdoin L 1996 Journal of Virology 70:148-158. Palmer KE & Rybicki EP 1998 Advances Virus Research 50:183-234. Powell PA, Nelson C, De B, Hoffmann N, Rogers SG, Fraley RT & Beachy RN 1986 Science 232:738743. Sanford JC & Johnson SA 1985 Journal of Theoretical Biology 115:395-405. Souza Jr MT & Gonsalves D 1999 Fitopatologia Brasileira 24:485-506. Sunter G & Bisaro D M 1991 Virology 180:416-419. Sunter G & Bisaro D M 1992 Plant Cell 4:1321-1331. Timmermans M C P, Das O P & Messing J 1994 Annu Rev Plant Physiol Plant Mol Biol 45:79-112. Walker JE, Saraste M, Runswick MJ & Gay NJ 1982 EMBO Journal 1:945-951. Wilson TMA 1993 PNAS USA 90:3134-3141. ANÚNCIO UNISCIENCE (Repete Fotolito) (Repete Fotolito pág. 41 última edição) Atenção: são dois anúncios da Uniscience: Um inédito (fotolito novo enviado em anexo) que entra na página 15 desta edição. O outro (desta página) que repete o que saiu na página 41 da última edição. Biotecnologia Ciência & Desenvolvimento 27 Pesquisa Biotransformação de ÓLEOS E GORDURAS Utilização de lipases para obtenção de biocombustível Maria da Graça Nascimento Doutora em Química Orgânica pela UNICAMP e professora do DQ-UFSC [email protected] Pedro Ramos da Costa Neto Doutorando no DQ-UFSC Licenciado pelo CEFET/PR [email protected] Lilian Maria Mazzuco Iniciação Científica no DQ-UFSC [email protected] Departamento de Química Universidade Federal de Santa Catarina - SC Fotos cedidas pelos autores Aspectos Gerais As lipases, E.C.3.1.1.3., fazem parte de um grupo de enzimas hidrolíticas, com cerca de 300 resíduos de aminoácidos, que catalisam a quebra de ligações ésteres de acil gliceróis. Elas não requerem cofatores, são de baixo custo, regioespecíficas, atuam em uma larga faixa de pH e, além de efetuar reações de hidrólise, podem também exercer atividade catalítica (Figura 1). nesse seguimento ainda não está suficientemente difundido (Bon e Pereira 1999). Segundo alguns trabalhos, que fazem parte da literatura, as lipases têm sido empregadas em processos de extração de óleos, melhoramento das propriedades físicas e nutricionais de gorduras por interesterificação (Macrae 1983, Pecnik e col.1992, Konishi e col. 1995, Hayes 1996, Glosh e col.1997, Facioli e col. 1998), limpezas de tripas em abatedouros bovinos (Burin e col.1994), hidrólise e degomagem de óleos vegetais na industria (Bon e Pereira 1999). Biocombustíveis Figura 1. Reações catalisadas por lipases 28 Biotecnologia Ciência & Desenvolvimento As lipases são dotadas de uma especificidade pelo substrato que supera todas as outras enzimas conhecidas. Isso lhes confere uma aplicação de fronteiras. Elas podem ser empregadas na produção de fármacos, cosméticos, detergentes, alimentos, perfumaria, diagnósticos médicos, síntese de compostos opticamente ativos, resolução de racematos, produção de aromas e fragrâncias, modificações de gorduras e tratamento de couros (Basheer 1995, Bon e Pereira 1999, Faber 1997). Considerando-se as transformações de óleos e gorduras em derivados ou matérias-primas industriais, o potencial de aplicações de enzimas Com exceção de hidrelétricas e da energia nuclear, a maior parte de toda a energia consumida no mundo vem do petróleo, do carvão e do gás natural. Como essas fontes são limitadas e deverão se esgotar no futuro, a visão de uma fonte alternativa de energia é de vital importância. A produção de biocombustíveis alternativos para o óleo diesel, provenientes de óleos vegetais brutos, tem sido tema de diversos estudos. Historicamente, relata-se que o próprio Rudolf Diesel desenvolveu pesquisas utilizando óleos vegetais em seus motores. Apesar de favorável energeticamente, o uso direto desse produto em motores é problemático devido à sua alta viscosidade (11 a 17 vezes maior que a do óleo diesel) e baixa volatilidade, o que impede a sua queima completamente, formando depósitos nos bicos injetores dos motores. Deve-se considerar também, que a decomposição do glicerol produz acroleína (substância altamente tóxica). Portanto, visando a reduzir a alta viscosidade dos óleos vegetais, diferentes alternativas têm sido consideradas, tais como: diluição; microemulsão com metanol ou etanol; decomposição térmica; craqueamento catalítico; e reação de transesterificação com etanol ou metanol. Entre essas alternativas, a transesterificação é a melhor escolha. Os ésteres de ácidos graxos obtidos são conhecidos como biodiesel e apresentam características físicas semelhantes às do óleo diesel, podendo ser utilizados em motores do ciclo diesel sem nenhuma modificação (Schuchardt e col. 1998, Costa Neto e col. 2000). A transesterificação de óleos e gorduras pelo processo químico, embora seja simples, rápido e com alto rendimento, apresenta algumas desvantagens. A primeira refere-se ao catalisador (ácido ou base), que, ao final do processo, permanece misturado com o principal subproduto da reação, que é a glicerina, dificultando sua separação e purificação. Utilizando um processo enzimático, essas dificuldades podem ser minimizadas e existirá a possibilidade de obter-se tanto biodiesel como glicerina, com maior grau de pureza e possibilidade de reutilização do biocatalisador. Uma segunda desvantagem do processo químico está relacionada com o tipo de álcool utilizado, que, de modo geral, utiliza-se metanol. Nesse caso, a alcoólise enzimática com etanol hidratado apresenta grandes vantagens em relação ao metanol, tais como custo e toxidade. Além disso, o etanol é um produto obtido através de biomassas e, dessa maneira, o processo se torna totalmente independente do petróleo (tabela 1). Embora os processos de transesterificação enzimática para obtenção de biodiesel ainda não sejam comercialmente desenvolvidos, novos resultados têm sido reportados em artigos e patentes (Schuchardt, et. al. 1998). O aspecto comum desses estudos consiste na otimização das condições de reação (solvente, temperatura, pH, tipo de microorganismo que gera a enzima, etc.), a fim de estabelecer as características para aplica- Tabela 1. Principais vantagens e desvantagens enzimático para a produção de biodiesel Processos Vantagens Químico Simplicidade Alto rendimento Curto tempo de reação Enzimático Facilidade de separação do catalisador (suporte) Obtenção de produtos com maior grau de pureza Possibilidade de utilizar etanol hidratado na reação dos processos químico e Desvantagens Dificuldade de separação do catalisador Impossibilidade de reutilização do catalisador Dificuldade de utilização de etanol hidratado Obtenção de produtos com menor grau de pureza Longo tempo de reação Custo das enzimas Baixo rendimento Tabela 2. Rendimentos (%) dos ésteres obtidos através da reação de transesterificação enzimática do óleo de girassol Lipases C/Solvente Álcoois P. fluorescens Mucor mirhei Candida sp 25 79 53 MeOH 82 99 79 EtOH (96%) 80 81 29 n-PrOH 3 MeOH S/Solvente EtOH (anidro) 70 EtOH (96%) 82 n-ButOH 76 Tabela 3. Rendimentos (%) dos ésteres obtidos através da reação de transesterificação enzimática de triglicerídeos com álcoois primário e secundário Lipases C. antarctica P.cepacia Álcoois M. miehei Sebo 25,7 MeOH 94,8 13,9 / EtOH (95%) 98,3 13,7 / EtOH (anidro) 68,0 / 61,2 l-PrOH 24,3 44,1 83,8 2-BuOH 19,6 41,0 Soja / MeOH 75,4 14,5 EtOH 97,4 Colza 77,3 MeOH ções industriais. Contudo, tanto o rendimento como o tempo de reação ainda são desfavoráveis se comparados com o sistema de reação por catálise básica. A seguir são apresentados alguns desses estudos. A alcoólise de óleo de girassol, na presença ou ausência de solvente (éter de petróleo) foi investigada por Mittelbach (1990), que utilizou três lipases obtidas de diferentes fontes. Os melhores rendimentos foram obtidos com a lipase de Pseudomonas sp e etanol hidratado. Por outro lado, na ausência de solvente, o rendimento foi menor e o tempo de reação maior (tabela 2). Nelson e col. (1996) investigaram a reação de transesterificação de alguns óleos vegetais e sebo bovino, utilizando álcoois primários, secundários e diversas lipases. Os melhores resultados foram obtidos na alcoólise do sebo com metanol e de etanol com a lipase de Mucor miehei. Os rendimentos Biotecnologia Ciência & Desenvolvimento 29 obtidos com etanol hidratado foram superiores ao do anidro. Utilizando-se álcoois secundários, a lipase de Candida antartica foi a mais eficiente nesse estudo FIGURA 2. Óleo de soja aquecido a 180-200 oC, em diferentes tempos (0-81h), para (tabela 3). estu-dos de reações de transesterificação com lipases A atividade hidrolítica representa uma característica básica das lipases e rendimento total de ésteres de 98,4%. dos foram usados em reações de normalmente se associa com a habi- A estabilidade da lipase usada nas três esterificação do ácido láurico, oléico, lidade sintética. Wu e col. (1996) etapas da metanólise foi verificada linoléico e esteárico com n-pentanol, investigaram essas características uti- pela repetição de sua aplicação em etanol e metanol, além da etanólise lizando nove lipases. De acordo com novos substratos, que em cinquenta do óleo de soja. Em geral, os produos resultados obtidos, a atividade ciclos de reutilizações apresentaram tos (ésteres) foram obtidos com bons hidrolítica da lipase pode ter pouco rendimentos superiores a 95%. rendimentos (> 90%) quando a CCL valor na predição da atividade sintéfoi imobilizada em gel de ágar. tica. Ao contrário, a lipase pode não Uso de CCL em Síntese Os resultados obtidos para a reaexibir atividade sintética, mas pode ção de esterificação enzimática do apresentar alta atividade hidrolítica. A preparação de suportes com ácido láurico foram comparados com Em outro estudo, Wu e col. lipase de Candida rugosa (CCL, Sigma os da enzima livre por meio do tempo (1999) empregaram a metodologia 905 u/mg sólido) para posterior utili- de reação. A reutilização dos suporde superfície de resposta para otimi- zação em reações de transesterifica- tes carvão de coco e gel de ágar zar parâmetros de reação de transes- ção de óleo de soja novo, e degradado também foram avaliados para essa terificação de banha usada em restau- termicamente visando à obtenção de reação. rantes com etanol hidratado, utilizan- biodiesel por via enzimática, tem sido A Figura 4 mostra os rendimentos do do as lipases de P. cepacia e C. realizada no Laboratório de Biocatáli- laurato de n-pentila obtidos pela utiantarctica. O melhor rendimento da se da UFSC, por Nascimento e col. lização da CCL livre e imobilizada nos reação (96%) foi obtido quando utili- 1999. (Figura 2). diferentes suportes. O melhor resulzaram-se simultaneamente, as duas Os suportes avaliados para esse tado foi obtido com a CCL imobilizalipases. fim foram os carvões ativos de casca da em gel de ágar, após 24 horas. A otimização da reação de de coco, pinus e bauxita ativada, to- Utilizando-se a CCL livre (método de transesterificação de óleo de soja e dos granulados e com áreas superfici- Klibanov), obtém-se o éster em rencolza com metanol, utilizando-se li- ais de 854, 1.038 e 100 m2/g, respec- dimento quantitativo, após 165 hopase de Candida antartica imobili- tivamente. Com a mesma finalidade, ras. Com os demais suportes, os renzada em cerâmica foi estudada por outro suporte que vem sendo estuda- dimentos dos produtos foram relatiShimada e col. (1999). Nessa reação, do é o de gel de ágar. (Figura 3). vamente baixos (< 40%) e o tempo de necessita-se de três equivalentes de Os biocatalisadores assim prepara- reação muito longo (> 400 horas). A metanol para um de óleo; foi reutilização desses suportes observado que o equivalenmostra que o gel de ágar pode te molar de metanol, superiser reutilizado pelo menos três or a 1,5, provocou a inativavezes sem perda da atividade ção da enzima. Portanto, a catalítica da enzima. Porém, reação foi conduzida com a quando a CCL foi imobilizada adição do álcool em etapas. em carvão de coco, houve Na primeira, a reação foi diminuição no rendimento de conduzida com um equivaéster, que foi de 40%, 18% e lente de MeOH, por 10 ho11%. (Figura 5). ras. Após a conversão de Os resultados preliminares 95% do metanol colocado obtidos para a reação de traninicialmente, foi acrescentasesterificação do óleo de soja do mais um equivalente e comercial com etanol, usando deixado em reação por 14 Figura 3. Suportes com CCL imobilizada (bauxita a CCL imobilizada nos referihoras. O terceiro equivalen- ativada, carvão ativo de pinus, de casca de coco e dos suportes, tem apresentado te foi finalmente adicionado de gel de ágar) comportamento similar ao das e a reação continuou por reações de esterificação. mais 24 horas, obtendo-se As reações têm sido conduzi30 Biotecnologia Ciência & Desenvolvimento Figura 4. Influência do tempo no rendimento da reação de esterificação do ácido láurico com n-pentanol em hexano, com a CCL livre e imobilizada em carvão de coco, pinus, bauxita e em gel de ágar, a 35 oC, (massa de CCL em cada suporte, 27 mg/g) Os transésteres derivados do etanol foram obtidos com rendimento quantitativo, após 36 h de reação a 35 o C com a CCL imobilizada em gel de ágar. Utilizando-se a CCL livre nas mesmas condições experimentais, o rendimento foi inferior a 50 %. Usando-se a lipase imobilizada em carvão de coco, nas mesmas condições experimentais descritas anteriormente, não foi verificada formação do produto. Considerações Finais A busca de novas alternativas para produção de energia deve ser considerada e avaliada. Assim, a investigação do uso de lipases imobilizadas para produção de biodiesel é de relevante importância, considerando-se o crescimento da utilização desse biocombustível em âmbito mundial, não somente pelo aspecto de meio ambiente, mas, principalmente, por se tratar de uma fonte de energia renovável. Agradecimentos Os autores agradecem a UFSC, CAPES, CEFET/PR, UNICAMP, FBCPR e NOVO NORDISK. REFERÊNCIAS BIBLIOGRÁFICAS Basheer, S.; Mogi, K.; and Nakajima, M. Interesterification Kinetics of Figura 5. Variação do rendimento do laurato de n-pentila em sucessivas reutilizações com a CCL imobilizada em carvão de coco e gel de ágar, em hexano a 35 oC Triglycerides and Fatty Acids with Modified Lipase in n-Hexane, J. Am. Oil Chem. Soc. vol. 72, no. 5, 1995, p. p.511-518. Bon, E. P. S. e Pereira Jr, N. Tecnologia Enzimática, Rio de Janeiro, 1999, 113p. Burin, A. M. e Fontoura, P. S. G. Extração da gorgura intestinal de bovinos por meio enzimático, Bol. CEPPA, vol. 12, no.2, 1994, p. 89-94. Costa Neto, P. R.; Rossi, L. F. S.; Zagonel, G. F.; Ramos, L. P. Produção de Biocombustíveis Alternativo ao Óleo Diesel Através da Transesterificação de Óleo de Soja Usado em Frituras. Química Nova. Vol. 23, n. 4, 2000, p. 531-537. Faber, K. Biotransformation in Organic Chemistry. 3a. ed., Springer, 1997, 402 p. Facioli, N. L. e Gonçalves, L. A. G. Modificação por via Enzimática da Composição Triglicéridica do Óleo de Piqui, Química Nova, 21(1), 1998, p.16-19. Ghosh, S. and Bhattacharyya, D. K. Utilization of High-Melting Palm Stearin in Lipase-Catalyzed Interesterification with Liquid Oils, J. Am. Oil Chem. Soc. vol. 74, no. 5, 1997, p. 589-592. Hayes, D. G. The Catalytic of Lipases Toward Hydroxy Fatty Acids A Review, J. Am. Oil Chem. Soc. vol. 73, no. 5, 1996, p. 543-549. Konishi, H.; Neff, W. E. and Mounts, T. L Oxidative Stability of Soybean Oil Products Obtained by Regioselective Chemical Interesterification, J. Am. Oil Chem. Soc. vol. 72, no. 11, 1995, p.1393-1398. Macrae, A. R. Lipase-Catalyzed Interesterification of Oils and Fats. J. AM. Oil Chem. Soc. v. 80, n.2, 1983, 291-294. Mittelbach M. Lipase Catalyzed Alcoholyses of Sunflower Oil, J. Am. Oil Chem. Soc. vol. 67, no.3, 1990, p. 168-170. Nascimento, M. G. e Costa Neto, P. R. Uso de Carvão Ativo e Bauxita para Imobilização de Lipase Candida cilyndracea; 22a. Reunião Anual da SBQ; 1999. V.1.(FQ-054). Nelson, L. A .; Foglia, T. A.; and Marmer, W. N.; Lipase-Catalyzed Production of Biodiesel; J. Am. Oil Chem. Soc;. v. 73; n. 8; 1996; 1191-1195. Pecnik, S. and Knez, Z. J. Am. Oil Chem. Soc. v.69, n.3, 1992, 261-265. Schucharddt, U.; Serchel, R. and Vargas, R. M. Transesterification of Vegetable Oils: a Review, J. Braz. Chem. Soc. , vol. 9, no. 1, 199-210, 1998, p.199-210. Wu, X. Y.; Jaaskelainen, S. and Linko, Y. Y. An investigation of crude lipases for hydrolysis, esterification, and transesterification, Enzyme and Microbial Technology, 19:226-231, 1996. Wu, W. H.; Foglia, T. A.; Marmer, W. N. and Phillips, J. G.; Optimizing Production of Ethyl Esters of Grease Using 95% Ethanol by Response Surface Methodology. J. Am. Oil Chem. Soc. vol. 76, no. 4, 1999, p. 517-521. Biotecnologia Ciência & Desenvolvimento 31 Saúde Adjuvantes de MUCOSAS Melissa Ang Simões Mestranda do programa de pós- graduação em biotecnologia Luís Carlos de Souza Ferreira Dr., PhD, Professor Titular do Dep. de Microbiologia da USP. Departamento de Microbiologia, Instituto de Ciências Biomédicas, Universidade de São Paulo - SP [email protected] Aliados na busca de vacinas orais mais eficazes maioria dos patógenos, sejam eles bactérias, vírus, fungos ou parasitas, penetra no organismo humano através de uma superfície de mucosa, como as que limitam os tratos respiratório, gastrointestinal e geniturinário. Entre outros exemplos, podemos citar doenças causadas por microrganismos, que penetram no organismo através de mucosas, como a AIDS, a gonorréia, meningites bacterianas e virais; a tuberculose, diarréias infecciosas, febre tifóide, esquistossomose, hepatites, câncer de colo uterino causado pelo papilomavírus; a poliomielite e as gripes e resfriados. Uma alternativa eficaz e econômica para o controle dessas doenças, além de medidas que possam melhorar as condições educacionais, sanitárias e higiênicas da população, seria o desenvolvimento de vacinas capazes de induzir proteção imunológica local, isto é, na própria superfície da mucosa em contato com o patógeno. Nos mamíferos, a proteção imunológica em mucosas depende, em grande parte, da produção de anticorpos do isotipo IgA secretor (sIgA) presentes na saliva, muco gastrointestinal e pulmonar, secreções vaginais e leite materno (Ver quadro - sistema imune comum de mucosas). Vacinas convencionais (vírus e bactérias mortas ou atenuadas, frações acelulares como toxóides ou componentes de superfície) são eficazes na prevenção de doenças infecciosas que necessitam de anticorpos séricos circulantes (por exemplo, o tétano, a difteria e a coqueluche, assim como algumas doenças virais como febre amarela, hepatite A e B, cachumba e varíola). Entretanto, essas vacinas são incapazes de gerar resposta imune local, isto é, a produção de sIgA em mucosas, o que explica, em parte, a dificuldade de se desenvolver em vacinas eficazes contra a AIDS e doenças entéricas. Para que uma resposta imune local seja gerada, a vacina deve ser administrada preferencialmente no sistema imune de mucosas, como aquelas administradas por via oral, e deve ser preparada de preferência com microrganismos atenuados capazes de se replicarem nos tecidos do hospedeiro. A vacina oral contra a pólio (Sabin) é um exemplo bem sucedido de vacina de mucosas. Vacinas de mucosas também são capazes de ativar respostas imunológicas sistêmicas, o que as tornam instrumentos valiosos para a proteção imunológica contra inúmeros patógenos. Entretanto, embora as vacinas orais ou nasais apresentem uma série de vantagens em relação às 32 Biotecnologia Ciência & Desenvolvimento vacinas de administração parenteral, o número de vacinas de mucosas atualmente disponíveis para uso no homem ou em animais domésticos ainda é muito limitado. Portanto, a pesquisa e o desenvolvimento de novas vacinas de mucosas mais seguras e eficazes é uma prioridade para a saúde pública mundial. Figura 1 : Diagrama da estrutura cristalográfica da enterotoxina termo-lábil de E. coli (LT). As subunidades A1 (amarelo) e A2 (vermelho) formam uma estrutura triangular, que se encaixa no anel pentamérico formado pelas cinco subunidades B (azul/verde) que delimitam um poro central, através do qual o fragmento A1 atinge o interior da célula alvo Nos últimos dez anos, observamos um grande avanço nas pesquisas de novas vacinas, as chamadas vacinas de nova geração, que resultaram em diversas estratégias vacinais promissoras como aquelas constituídas por peptídeos sintéticos, vacinas de DNA e vacinas recombinantes bi ou multivalentes baseadas em vírus ou bacté- rias atenuadas. A pesquisa voltada para vacinas de mucosas também experimentou avanços importantes como o uso de veículos baseados em lipossomos e microesferas, microrganismos atenuados e plantas transgênicas, utilizados como veículos para antígenos diversos. Novas estratégias baseadas em vacinas de DNA também trazem boas perspectivas para uma nova geração de vacinas de mucosas mais eficazes (Lásaro & Ferreira, 2000). No entanto, uma das contribuições mais importantes para a pesquisa de novas vacinas de mucosas foi a descoberta de adjuvantes capazes de atuar sobre o sistema imune de mucosas. O termo adjuvante é utilizado para designar uma substância que, usada em combinação com um antígeno de natureza protéica ou polissacarídica, resulta em resposta imune maior do que aquela produzida pelo antígeno administrado isoladamente. Entretanto, os efeitos dos adjuvantes não se restringem apenas ao aumento da imunogenicidade de antígenos fracos, já que também podem promover a Tabela 1: Principais propriedades dos adjuvantes. 1 - Diminuição do período necessário para a indução da resposta imune e aumento da duração da resposta de memória imunológica; 2 - Capacidade de modular a avidez, a especificidade, o isotipo e/ou a distribuição dos anticorpos gerados; 3 - Aumento da resposta imunológica em indivíduos idosos ou imaturos imunologicamente; 4 - Em alguns casos, promover a indução de imunidade em mucosas (adjuvantes de mucosas); 5 - Aumento da potência imunológica de peptídeos recombinantes ou sintéticos; 6 - Capacidade de modular a resposta imune, tanto celular como humoral. ativação de diferentes vias do sistema imunológico (Tabela 1). Existe uma enorme diversidade de adjuvantes na natureza. Alguns adjuvantes têm origem mineral, outros são derivados de vegetais ou produtos bacterianos. Existem ainda compostos com propriedades adjuvantes produzidos pelo próprio metabolismo como alguns mediadores químicos da resposta imunológica (citocinas) e hormônios (Tabela 2) (Edelman, 1997). O sistema imune comum de mucosas A produção de sIgA em algumas secreções glandulares e no muco que reveste os epitélios de membranas do corpo humano está a cargo de células especializadas, os plasmócitos. As células precursoras de plasmócitos produtores de sIgA estão localizadas nos tecidos linfóides localizados ao longo do trato respiratório, geniturinário e gastrointestinal. No intestino, o tecido linfóide é representado pelas placas de Peyer, formadas por células epiteliais diferenciadas, as células M, capazes de capturar e transportar antígenos presentes no lúmen, e folículos subjacentes ricos em linfócitos e macrófagos. A partir desses folículos, os linfócitos B precursores, sensibilizados por um antígeno, migram por meio de linfonodos mesentéricos, duto linfático torácico e corrente sangüínea para vários sítios efetores espalhados pelo corpo, como glândulas salivares, lacrimais e mamárias, além da lâmina própria subjacente aos epitélios do intestino, pulmão e trato geniturinário. Dessa forma, a administração de um antígeno a um sítio de mucosa pode levar à produção de sIgA específico em diferentes mucosas e glândulas. Esse conjunto de elementos do sistema imunológico foi, portanto, denominado sistema imune comum de mucosas. Os anticorpos do isotipo IgA produzidos por plasmócitos diferenciados ligam-se a uma porção secretora, sintetizada por células epiteliais, responsável pelo trânsito intracelular e secreção da molécula de IgA para o ambiente extracelular, além de conferir resistência à ação de proteases. Um indivíduo adulto, com 70 kg, produz cerca de 3 g de sIgA por dia, o que representa 80% da produção total de anticorpos. A função básica da sIgA é prevenir a interação inicial do patógeno com a superfície de mucosa pelo bloqueio da colonização e/ou invasão de células do hospedeiro e neutralização de toxinas. Os únicos adjuvantes até hoje licenciados para uso clínico são os sais de alumínio, principalmente hidróxidos e fosfatos. Entretanto, a maioria dos adjuvantes, de uso clínico ou não, apresentam como principais limitações a necessidade de administração parenteral e efeitos restritos às respostas imunológicas sistêmicas. Os adjuvantes de mucosas, administrados por via oral ou nasal, por sua vez, promovem respostas sistêmicas e locais. Entre os compostos adjuvantes até hoje testados, dois se destacam pelo efeito sobre o sistema imune de mucosas, a toxina colérica (CT) de Vibrio cholerae e a toxina termo-lábil (LT), produzida por algumas linhagens de Escherichia coli enterotoxigênica (ETEC). CT e LT são enterotoxinas do tipo A-B, isto é, toxinas que apresentam uma subunidade A, responsável pela atividade tóxica, e uma subunidade B, envolvida com a ligação a receptores específicos em células do hospedeiro. Embora LT e CT compartilhem aspectos estruturais e funcionais, essas moléculas apresentam diferenças bioquímicas e imunológicas importantes que se refletem em suas propriedades tóxicas e no potencial de utilização biotecnológica. LT e CT apresentam 80% de identidade em suas seqüências de aminoácidos; entretanto, CT possui maior toxicidade do que LT, em humanos. Além disso, em mamíferos, CT leva à ativação de respostas imunológicas que se caracterizam pela produção de IgE, o que pode resultar em choque anafilático em animais reexpostos à toxina. Por outro lado, LT induz respostas imunológicas sem o acúmulo de IgE, o que reduz o risco de efeitos colaterais. Biotecnologia Ciência & Desenvolvimento 33 Vários grupos de pesquisa dedicaram-se ao isolamento de derivados atóxicos de LT, sendo que alguns destes apresen(A) A toxina LT se liga pelas taram propriedades imunomosubunidades B a receptores presentes duladoras preservadas. Uma no enterócito; uma vez interiorizado, o forma mutante de LT foi gerafragmento A1 modifica o componente da pela modificação do sítio α da proteína G (Gsα), que regula a sensível à ação de proteases atividade de adenil ciclase (AC) a qual, na subunidade A. Esse derivapor sua vez, provoca aumento do do, denominado LT(R192G), conteúdo de cAMP e conseqüente possui a arginina na posição perda de íons-cloro e água; (B) 192 substituída por uma glicina, e mantém suas propriedaMecanismo de mono ADP-ribosilação des adjuvantes quando commediado pela toxina LT parada à forma nativa de LT (Dickinson & Clements, 1996). O isolamento de derivados atóxicos de LT permitiu a elaboração de diversas formulações vacinais baseadas em antígenos purificados, extratos celulares e microrganismos inativados ou atenuados. Entre as vacinas de mucosas preparapelo bloqueio da ativação de resposta das com LT destacam-se aquelas voltaimune contra antígenos ingeridos das para o controle do vírus influenza, (como aqueles presentes na alimentação). Por fim, LT age como adjuvante HIV, rotavírus, Shigella flexneri, Helide mucosas e promove o aumento da cobacter pylori, Campylobacter pylori, resposta de anticorpos, local e sistêmi- Salmonella typhimurium e Candida ca, para antígenos co-administrados albicans. Os resultados obtidos depor via oral ou nasal. Tais proprieda- monstraram que todas as vacinas fodes conferem à LT um grande poten- ram capazes de induzir resposta de cial de utilização biotecnológica como anticorpos sistêmicos e secretores conadjuvante para vacinas, sejam elas tra os antígenos co-administrados e, constituídas de antígenos purificados em alguns casos, foi possível demonsou de microrganismos mortos ou ate- trar a proteção conferida em modelos nuados, administradas por via de mu- animais. Tais resultados levam a crer cosas (Dickinson & Clements, 1996). que um efetivo programa de imunizaO efeito adjuvante de LT, assim ção baseado em vacinas de mucosas como o de CT, ainda não está esclare- acrescidas de LT modificada possa ser cido, mas evidências experimentais implementado em futuro próximo para demonstraram que as propriedades o controle de diversas doenças virais, imunoestimuladoras dessa toxina po- bacterianas ou parasitárias. Nosso grupo realiza pesquisas voldem envolver o aumento da permeatadas para o desenvolvimento de nobilidade do epitélio intestinal, a ativação de células apresentadoras de an- vas estratégias vacinais contra diarréitígenos por meio da maior expressão as causadas por Salmonella e ETEC de moléculas co-apresentadoras de (Lásaro e Ferreira, 2000). Entre as aborsuperfície e produção de citocinas dagens utilizadas, está a incorporação que favorecem a produção de anticor- de formas atóxicas de LT a vacinas pos sistêmicos e secretores. Entretan- bivalentes de administração oral baseto, a atividade tóxica, tanto de LT adas em linhagens atenuadas de Salcomo de CT, inviabilizaria o uso des- monella typhimurium modificadas gesas toxinas em formulações de vaci- neticamente para expressar antígenos nas destinadas a humanos. Uma alter- de ETEC. Como protótipo desses exnativa para superar essa dificuldade perimentos, utilizamos células vivas foi a obtenção, através de técnicas de de uma linhagem recombinante de S. mutagênese sítio dirigida, de formas typhimurium (HG3) capaz de expressar a fímbria CFA/I de ETEC, uma mutantes não tóxicas de CT e LT. Figura 2 : Modelo esquemático do mecanismo de ação da toxina LT. A molécula de LT é formada por uma subunidade A (29kDa) e cinco subunidades B (11,5 kDa); estas últimas formam uma estrutura em forma de barril, que acomoda; no seu interior, a subunidade tóxica (Figura 1). A ligação do componente B a receptores nas células do hospedeiro permite a entrada da subunidade A, que, em seguida, sofre uma clivagem proteolítica para gerar um peptídeo enzimaticamente ativo (A1) e outro menor (A2), que estabiliza o complexo. A subunidade A1 promove a mono ADPribosilação de uma proteína reguladora (GSα) da adenilato ciclase de enterócitos, o que resulta no aumento dos níveis intracelulares de AMP cíclico (cAMP), com conseqüente aumento na secreção de íons-cloro. Esse desequilíbrio iônico reverte o fluxo normal de líquidos através do epitélio intestinal, o que resulta em perda de água e o quadro típico da diarréia. Curiosamente, além da ação tóxica associada ao aumento nos níveis de cAMP, LT (assim como CT) é capaz de ativar uma série de efeitos que resultam em marcante atividade imunoreguladora. Ao contrário da maioria dos antígenos administrados por via oral, LT resiste à passagem pelo ambiente gastrointestinal e induz resposta de anticorpos, local e sistêmica, contra si mesma. LT também impede o estabelecimento da tolerância, responsável 34 Biotecnologia Ciência & Desenvolvimento adesina responsável pela aderência da ETEC ao epitélio intestinal, em conjunto com a LT(R192G). Animais imunizados por via oral com a formulação vacinal formada pela combinação de 1010 células vivas da linhagem HG3 com 25 µg de LT(R192G) resultou em marcante resposta de anticorpos, sistêmicos (IgG) e secretores (sIgA) contra o vetor (S. typhimurium) e o antígeno heterólogo de ETEC (CFA/I) (Figura 3). Além disso, todos os animais imunizados desenvolveram resposta de anticorpos contra LT, o que torna a vacina mais eficaz contra a infecção pela ETEC, pois abrange também o componente tóxico do patógeno. Experimentos adicionais revelaram que resultados semelhantes podem ser alcançados com linhagens Salmonella inativadas, alternativa mais segura para a imunização de indivíduos imunodeprimidos. Esses resultados mostraram, pela primeira vez, que os efeitos adjuvantes de LT podem ser combinados a vacinas bivalentes baseadas em linhagens atenuadas de Salmonella recombinante (Guillobel et al., 2000). O principal objetivo da pesquisa em vacinas é alcançar uma formulação ideal que atue em dose única, gere proteção duradoura contra diversos patógenos, dispense o uso de seringas, tenha baixo custo de produção, estabilidade térmica, e seja segura mesmo para indivíduos imunocomprometidos ou recémnascidos. Ainda não podemos prever quando tais vacinas estarão disponíveis para o uso humano, mas o contínuo e acelerado ritmo de desenvolvimento dos adjuvantes de mucosas e novas estratégias vacinais contribuirá para que, em futuro próximo, possamos usufruir desses produtos. Referências bibliográficas 1 Edelman, R. (1996). Adjuvants for the future. In. Levine, MM, Woodrow, GC, Kaper, JB, & Cobon, GS (Eds.). New Generation Vaccines, pp. 173-192. Marcel Dekker, Inc., New York. Tabela 2: Principais tipos de adjuvantes segundo a origem ou natureza química Origem ou Natureza química Principais adjuvantes Compostos minerais Sais de cálcio e/ou alumínio; Produtos bacterianos Compostos derivados de micobactérias (adjuvante completo de Freund, Detox, BCG, peptídeos derivados da parede, dimicolato de trealose); Lipídeo A de Salmonella, extratos ou frações derivadas de Corynebacterium parvum, Bordetella pertussis, e Klebisiella pneumonia, toxina colérica de V. cholerae e toxina termo-lábil de ETEC; Produtos vegetais Glucanas extraídas de algas, Saponinas e glicosídeos triterpênicos de Quillaja saponaria; Vitaminas Vitamina A, D3, E; Citocinas IFN-α, IFN-γ, GM-CSF, IL-1, IL-2, IL-12; Hormônios Hormônio de crescimento, DHEA. Segundo R. Edelman (1997) 2 Dickinson, BL & Clements, JD (1996). Use of Escherichia coli heatlabile enterotoxin as an oral adjuvant. In Kiono, H, Ogra, PL, & McGhee, JR (Eds.) Mucosal Vaccines, pp 73-87. Academic Press, San Diego. 3 - Guillobel, HCR., Carinhana JI, Cárdenas, L, Clements, JD, Almeida, DF & Ferreira, LCS (2000). Adjuvant activity of a non toxic mutant of Escherichia coli heat-labile enterotoxn on systemic and mucosal immune responses elicited against a heterologous antigen carried by a live Salmonella enterica Serovar Typhimurium vaccine strain. Infection and Immunuty, 68 (7):4349-4353. 4 Lásaro, MO, & Ferreira, LCS. (2000). Vacinas contra diarréias. Novas perspectivas baseadas em vacinas de DNA. Biotecnologia Ciência e Desenvolvimento, 3(14):28-32. Figura 3: Produção de IgG sérico e sIgA em fezes para o CFA/I de ETEC expresso por uma linhagem vacinal de Salmonella após administração oral em camundongos. Camundongos BALB/c foram imunizados com três doses de 1010 células vivas de HG3 por via oral, na presença (HG3 + LT) ou não (HG3) de LT. Amostras de animais não imunizados (NI) foram utilizadas como controles negativos. (A) Resposta de anticorpos sitêmicos de isotipo IgG específicos para o CFA/I; (B) resposta de anticorpos secretados em fezes do isotipo IgA especifícos para o CFA/I Biotecnologia Ciência & Desenvolvimento 35 INSULINA DE Pesquisa PLANTAS Hormônio peptídico em plantas Antonia Elenir Amâncio de Oliveira (Doutoranda, UENF) Cassiana Rocha Azevedo ( Iniciação Científica, UENF) Thiago Motta Venâncio (Iniciação Científica, UENF) Luciana Belarmindo da Silva (Mestranda, UENF) Marco Antônio Lopes Cruz (Mestrando, UENF) Olga Lima Tavares Machado (Professora Associada, UENF [email protected]) Maura da Cunha (Professora Associada, UENF [email protected]) Viviane de Oliveira Santos (Iniciação Científica, UENF) Cristiane Paulain Cavalcante (Iniciação Científica, UA) Adriana Ferreira Uchoa (Doutoranda, UENF) Kátia Valevski Sales Fernandes (Professora Associada, UENF [email protected]) Valdirene Moreira Gomes (Professora Associada, UENF [email protected]) Spartaco Astolfi Filho (Professor Titular, UA [email protected]) José Xavier-Filho (Professor Titular, UENF [email protected]) Fotos cedidas pelos autores 36 Biotecnologia Ciência & Desenvolvimento betes. Os Gregos antigos não tinham tratamento para a doença e, em várias partes do mundo, acrediO diabetes, caracterizado por tava-se que ela fosse causada por elevados níveis de glicose no sancalor excessivo nas vísceras, por gue e excesso de urina com sabor variações humorais ou pelo excesadocicado é, atualmente, uma das so de bebidas. No fim do século doenças mais importantes que afeXVIII, Matthew Dobson provou que tam a humanidade. Segundo a Oro sabor da urina no diabetes era ganização Mundial de Saúde (OMS), devido à presença de açúcar e mostrou o excesso de açúcar no sangue; na mesma época, John Rollo observou o odor de acetona nos pacientes com diabetes (Pike, 1999). Posteriormente, foram feitas descobertas importantíssimas para o melhor entendimento do diabetes. Figura 1 Planta e sementes de Canavalia Entre elas a de Von ensiformis Mering e Oscar Minkowski, em 1899, de que a retirada do pâncreas do existem 142 milhões de diabéticos cachorro promovia o aparecimenno mundo. A estimativa é que, até to da doença naquele animal. Em 2005, o número alcance 300 mi1hões 1921, uma equipe liderada por Fre(O Globo, 1998). A doença é coderick Banting descobriu, no Cananhecida desde a antiguidade. há dá, que um peptídeo, a insulina, mais de 1500 anos A.C. os antigos secretado pelas ilhotas de Lanegípcios tinham alguns remédios gerhans do pâncreas, causava o para combater o excesso de urina, abaixamento dos níveis de açúcar e os hindus observaram que insetos do sangue. Em 1926, esse peptídeo e moscas eram atraídos para a urina foi cristalizado por J. J. Abel e, em de algumas pessoas e que isso 1955, Frederick Sanger determinou estava associado a certas doenças. sua seqüência de aminoácidos Mil anos A.C. o pai da medicina na (Turkenburg-van Diepen, 1996). Índia, Susruta, diagnosticou o diaDiabetes e Insulina Sabe-se, atualmente, que a insulina exerce seus efeitos quando se liga ao seu receptor na parte externa da membrana celular. Essa ligação inicia uma cascata de fosforilação no interior da célula, que, entre outras coisas, promove a fosforilação e conseqüente ativação de transportadores de glicose. Isso faz com que essa molécula penetre na célula e participe dos processos metabólicos em que está envolvida. São conhecidos também outros efeitos da ação de insulina sobre o metabolismo e ativação de genes na célula animal. Tratamento do diabetes Diabetes não tem cura. Seu tratamento é feito, principalmente, à base de injeções de insulina, sendo sua ingestão ineficaz. O hormônio tem uma importância enorme no controle dessa afecção. Entretanto, o sofrimento decorrente das múltiplas injeções diárias a que são submetidos os pacientes tem sido um estímulo para a busca de substitutivos, principalmente os que exerçam seus efeitos por via oral. Alguns desses são pequenas moléculas que, de uma forma ou de outra, mimetizam a ação da insulina em pacientes diabéticos. Exemplo de composto anti-hiperglicêmico do tipo acima é o metformin (N, N-dimetil guanidina) originalmente detectado em uma planta, Galega officinalis, da família das leguminosas. Extratos de partes de plantas, como folhas, raízes ou sementes, têm sido utilizados, através dos tempos, pela medicina popular de, praticamente, todas as populações humanas no tratamento do diabetes. Entretanto, após a descoberta da insulina e com sua extensa comercialização, estudos sobre a utilização de remédios populares originados de plantas diminuíram seja por não serem considerados de valor comercial seja por estarem, aparentemente, associados a uma ciência de segunda categoria. Vale ressaltar que muito poucos remédi- Figura 2 Seqüências de aminoácidos de insulinas bovinas e de plantas os ditos naturais para o tratamento do diabetes tiveram seu valor terapêutico comprovado. Insulina de Plantas Logo após a realização dos estudos que levaram à descoberta da insulina no pâncreas de cães, dois dos cientistas envolvidos (Collip, 1923; Best et al., 1924) com a descoberta apresentaram resultados nos quais sugeriam a presença de substâncias possivelmente similares à insulina em extratos das mais diversas plantas. Collip chegou a dar ao seu produto o nome de glucocinina, pois imaginou que um produto derivado de plantas não poderia ter o nome de insulina (originado da palavra latina que significa ilhota, referência às ilhotas de Langerhans do pâncreas). Em 1976, Khann et al. forneceram indícios mais concretos sobre a presença de insulina em plantas. Eles isolaram de frutos e de sementes de Momordica charantia (melão-de-São Caetano) uma fração protéica com massa molecular de, aproximadamente, 6,0 kDa, que reagia com anticorpo contra a insulina humana. Os estudos desse grupo foram baseados nas experiências da medicina popular indiana, que indica serem os frutos e sementes dessa cucurbitácea de grande valor no tratamento do diabetes. Posteriormente, Collier et al. (1987) relataram o isolamento de proteínas de folhas de espinafre e de centeio e de plantas de Lemna gibba G3, que apresentaram pesos moleculares semelhantes aos das insulinas animais, reagiram com anticorpo antiinsulina suína e exibiam a propriedade de ligação ao receptor de insulina humana. Nenhuma informação estrutural foi fornecida na ocasião ou posteriormente. Não há registros posteriores a esses relatos na literatura científica sobre a presença de insulina em plantas. Mas há, no entanto, um registro contínuo na literatura, dos efeitos benéficos de uma enorme quantidade de extratos de folhas e de sementes das mais variadas plantas sobre o diabetes, e cujas propriedades sugerem a presença de proteínas ativas. A despeito dessa grande quantidade de informação, que sugere a presença de insulina em plantas, parece haver um paradigma que estabelece que as plantas não se valem de hormônios peptídicos, como insulina, para seus processos metabólicos. Há, conseqüentemente, descrença de que os vegetais possam se valer de insulina, por exemplo, para o controle da metabolização de açúcar em suas células. Como exemplo dessa descrença, há afirmações facilmente encontradas, como a que se segue (Brach, 2000): Biotecnologia Ciência & Desenvolvimento 37 Uma vez que insulina é um hormônio em animais, as plantas certamente não teriam um uso para ela; assim ela pode não ter qualquer efeito sobre as mesmas (......) . Há um enorme interesse em se utilizarem plantas como produtoras de proteínas animais de difícil preparação ou daquelas requeridas em grandes quantidades, a preços acessíveis. Como exemplo disso, há tentativas de produção de hormônio do crescimento em plantas transformadas adequadamente (Staub et al, 2000). As famosas plantas produtoras de vacinas estão freqüentemente anunciadas na literatura (Yu & Langridge, 2000). No caso da insulina, sua produção utilizando-se a chamada tecnologia do DNA recombinante já é uma indústria importante. A indústria brasileira BIOBRÁS, de Montes Claros, Minas Gerais, produz insulina humana, não propriamente utilizando células de vegetais, mas a partir de células de Escherichia coli transformadas com um gene da proinsulina humana (BIOBRÁS, 1998). Há uma iniciativa do Professor Adilson Leite, da Universidade de Campinas, que procura transformar plantas de fumo com um gene de insulina humana para produção do hormônio em sementes (FAPESP, 2000), além de registro sobre transformação de batata com um gene de insulina humana (Arakawa et al, 1998). Nossos resultados Acaso A descoberta de uma proteína com características da insulina bovina foi feita no Laboratório de Química e Função de Proteínas e Peptídeos, Centro de Biociências e Biotecnologia, Universidade Estadual do Norte Fluminense, em Campos dos Goytacazes, Rio de Janeiro, por acaso, durante investigações sobre o efeito de proteínas do tegumento de sementes de feijão-de38 Biotecnologia Ciência & Desenvolvimento Figura 3 Efeitos de insulinas sobre os níveis de glicose no sangue porco (Canavalia ensiformis) (Figura 1) na sobrevivência do caruncho ou gorgulho (Callosobruchus maculatus) de feijão-de-corda (Vigna unguiculata). Resultados já publicados (Oliveira et al., 1999) mostram que a proteína tem massa molecular idêntica e seqüência primária exatamente igual à da insulina bovina (Figura 2). Além disso, a proteína isolada reage com anticorpos antiinsulina (humana e bovina) da mesma maneira que insulina bovina. Mostramos também que a insulina isolada tem os mesmos efeitos que a animal no abaixamento dos níveis de glicose em animais diabéticos (Figura 3). Demonstramos, posteriormente, que a insulina de sementes de feijão-deporco localiza-se exclusivamente no tegumento e, neste, em uma camada de células que delimita o espaço entre esse tecido e os cotilédones (Figura 4). Sabe-se que essa camada de células está envolvida no transporte de açúcar das partes aéreas da planta para a semente durante a formação desta última. Juntamente com a molécula de insulina, detectamos um fragmento de proteína que apresentou homologia de seqüência com o receptor de insulina humana. Em experimentos feitos com a utilização de anticorpos antiinsulina e anti-receptor de insulina, mostramos que essas duas proteínas exercem um efeito marcante durante as primeiras etapas da germinação, quando se dá a embebição de água pela semente. A presença de insulina estimula a emersão da radícula que é bloqueada quando se adiciona anticorpo antiinsulina ao meio (Figura 5). Os experimentos sugerem fortemente que a insulina possa estar exercendo papel de sinalizador para a entrada de glicose nas células. Posteriormente, mas ainda não publicados, resultados obtidos mostram que as sementes de Canavalia braziliensis (feijão-de-boi), uma espécie relacionada a C. ensiformis, também contêm insulina e seu receptor. A idéia de que o papel da insulina em plantas possa estar ligado a processos de sinalização necessários para a metabolização de glicose semelhantes aos existentes em animais nos levou à procura da molécula em diferentes plantas. Plantas antidiabéticas As vagens verdes de feijões são recomendadas como sendo úteis no tratamento do diabetes. As vagens são utilizadas para a preparação de chás ou consumidas como saladas. Folhas de diversas plantas são também bastante utilizadas para o tratamento dessa afecção. Diante dos resultados obtidos com as sementes de feijão-de-porco, passamos a investigar as vagens de feijão-decorda (Vigna unguiculata) e as folhas de patade-vaca (Bauhinia forficata), duas plantas bastante utilizadas no Brasil (Panizza, 1997). Além de evidenciada a presença de insulina em vagens e tegumentos de sementes de feijão-de-corda em formação, mostramos sua presença também nas folhas dessa leguminosa (Venâncio et al, 2000). A presença do hormônio também foi comprovada nas folhas de patade-vaca. Neste último caso, conseguimos mostrar que a insulina se encontra predominantemente associada aos cloroplastos que são as organelas responsáveis pelos processos fotossintéticos que se dão nas folhas verdes (Azevedo, 2000). Preparação de insulinas de plantas A descoberta de insulina em tegumentos de feijão-de-porco foi feita utilizando-se preparações obtidas a partir de extratos feitos com tampões fosfato de potássio (50 mM, pH 7,6). Essas preparações eram cromatografadas em colunas de troca iônica e de filtração molecular e a sua pureza verificada por eletroforese em gel de poliacrilamida. Após a constatação de que nossa preparação continha insulina, feita inicialmente pelo seqüenciamento de amino ácidos, passamos a utilizar técnicas de imunoquímica para a detecção da proteína. Utilizamos um anticorpo poli- mostrada por ELISA ou por eletroforese em gel de poliacrilamida, seguida de Western blot. Conservação evolucionária Figura 4 Imunolocalização de insulina no tegumento de Canavalia ensiformis clonal contra insulina humana e empregamos técnicas de ELISA e de Western blot. As insulinas de folhas das diversas plantas estudadas foram detectadas seja por extração com tampões alcalinos (fosfato ou borato) seja por uma metodologia de extração que emprega uma mistura de etanol e água e acidificação com ácido forte. Essa metodologia é semelhante à que se emprega para preparação de insulina a partir de tecido pancreático (bovino ou de outro animal). Em alguns casos, lança-se mão do método desenvolvido por Khann et al. (1976), que utiliza extrações com misturas de álcool e clorofórmio, nas etapas iniciais. As etapas posteriores de purificação, quando foram necessárias, foram feitas por meio de técnicas comuns de química de proteínas, como cromatografia em troca iônica e filtração molecular. A identidade da insulina foi sempre Diante dos resultados obtidos no LQFPP com plantas da família das leguminosas, e tendo em vista os resultados de outros, com plantas de outras famílias (ver Collier et al., 1987, e Khann et al., 1976), passamos a examinar, utilizando técnicas de imunoquímica (ELISA, Western blot), folhas de plantas de um maior número de espécies. Essas foram selecionadas não somente por sua conhecida ação antihiperglicêmica relatada na medicina popular, mas para cobrir um grande número de espécies dos diferentes grupos de vegetais. Foram também incluídas plantas típicas da Região Amazônica, conhecida por sua grande biodiversidade. Os resultados obtidos mostram que a maioria dos extratos de folhas examinados contêm moléculas imunoreativas, seja em um ensaio em meio líquido (ELISA) seja sobre membranas (Western blot). Neste último caso, os resultados apontam para a presença de moléculas de insulina que apresentam a mesma massa molecular das insulinas animais. Grande parte dos resultados aponta também para a complexação da molécula de insulina de plantas com compostos de natureza glicídica. A fim de tentar estabelecer a presença de insulina (ou antígenos que se associam a anticorpos antiinsulina) em grupos mais primitivos, mostramos que essa molécula está presente em uma cianobactéria (Spirulina sp). Esses organismos tiveram origem em época anterior à formação dos cloroplastos das plantas verdes (Cavalier-Smith, 2000). Biotecnologia Ciência & Desenvolvimento 39 O conhecimento que se tem da extensão da presença de insulina em animais (Chan & Steiner, 2000) associado aos resultados obtidos por nós sobre a presença de insulina em plantas e cianobactérias e àqueles resultados dos que relatam a presença desse hormônio em bactérias (LeRoith et al., 1985) e fungos (LeRoith et al., 1980), nos levam a crer que essa molécula foi conservada durante a evolução, sugerindo que ela esteja associada aos processos de metabolização de glicose em todos os seres vivos. As investigações realizadas por nós, além de sugerirem que a insulina está presente em plantas e é uma molécula evolucionariamente conservada, também apontam para a validação de produtos da medicina popular utilizados no tratamento do diabetes. Esforços estão sendo dirigidos para compreender porque as insulinas de folhas de plantas são eficazes mesmo quando administradas oralmente. Referências Bibliográficas Arakawa T., Yu J., Chong D.K., Hough J., Engen P.C. & Langridge W.H. - A plant-based cholera toxin B subunit-insulin fusion protein protects against the development of autoimmune diabetes. Nat Biotechnol. 16: 934-938 (1998) Azevedo, C. R. -Investigações sobre a presença de insulina em folhas de Bauhinia forficata (patade-vaca) utilizadas no tratamento do diabetes (Monografia de graduação, UENF) 2000 Best C. H., Smith R. G. & Scott D. A. An insulin-like material in various tissues of the normal and diabetic animal. Amer. J. Physiol. 68: 161-182 (1924) BIOBRÁS - Uma Nova Era no Tratamento do Diabético Brasileiro. http://www.biobras.com.br/ releases_04.htm (1998) Brach, A.R. - Plants and Insulin http://www.herbaria.harvard.edu/ ~brach/ (2000) Cavalier-Smith, T. - Membrane heredity and early chloroplast evo40 Biotecnologia Ciência & Desenvolvimento Figura 5 Influência de insulina e anticorpo sobre a germinação de Canavalia ensiformis lution. Trends Plant Sci. 5: 174182 (2000) Chan, S. J. & Steiner, D. Insulin through the ages: Phylogeny of a growth promoting and metabolic regulatory hormone. Amer. Zool. 40: 213-222 (2000) Collier E., Watkinson A., Cleland C. F. & Roth J. - Partial purification and characterization of an insulin-like material from spinach and Lemna gibba G3. J. Biol. Chem. 262: 6238-6247 (1987). Collip J. B. Glucokinin. A new hormone present in plant tissue. Preliminary paper. J. Biol. Chem. 56, 513-543 (1923). FAPESP-Tabaco produz pró-insulina. PESQUISA FAPESP 54, http:/ /www.fapesp.br/lab54.htm (2000) Khann P., Nag T. N., Chandrajain S. & Mohan S. United States Patent. 3: 945-988 (1976). LeRoith D., Shiloach J., Heffron R., Rubinovitz C, Tannebaum R & Roth, J Insulin-related material in microbes: similarities and differences from mammalian insulins. Can. J. Biochem. Cell Biol. 63: 839849 (1985) LeRoith D., Shiloach J., Roth J & Lesniak M. A. Evolutionary origins of vertebrate hormones: Substances similar to mammalian insulins are native to unicellular euka- INSULINA (ANTÍGENO) ESTÁ PRESENTE EM CIANOBACTÉRIAS E PLANTAS VERDES Cianobactéria Briófitas Pteridófitas Gimnospermas Angiospermas Monocotiledôneas Dicotiledôneas Spirulina sp + Psilotaceae Equisetaceae Selaginelaceae Cycadaceae Ginkgoaceae Pinaceae Cupressaceae Nymphaeales Ceratoophyllales Piperales Aristolochiales Magnoliales Laurales Illiciales Acorales Alismatales Asparagales Liliales Dioscoreales Nartheciales Pandanales Arecales Bromeliales Philydrales Commelinales Typhales Juncales Poales Zingiberales Ranunculales Proteales Vitales Caryophyllales Polygonales Saxifragales Santalales Zygophyllales Geraniales Celastrales Malpighiales Oxalidales Fabales Rosales Cucurbitales Fagales Myrtales Brassicales Malvales Sapindales Cornales Ericales Garryales Solanales Gentianales Lamiales Aquifoliales Apiales Dipsacales Asterales + + + + + + + + + + + + + + + + + + + + + + + + + + + + + ryotes. Proc. Natl. Acad. Sci. 77: 6184-6188 (1980) Oliveira A. E. A., Machado O. L. T., Gomes V. M., Xavier Neto J., Pereira A. C., Vieira J. G. H., Fernandes K. V. S. & Xavier-Filho J. Jack bean seed coat contains a protein with complete sequence homology to bovine insulin. Prot. Pept. Lett. 6: 15-21 (1999). Organização Mundial da Saude (OMS) - Obesidade está associada a 80% dos casos de diabetes. O Globo, Rio de Janeiro (6/12/1998) Panizza S. Plantas que curam (Cheiro de mato). 15a. ed. IBRASA, São Paulo, 279 pp (1997) Pyke D.A. - Preamble: the History of Diabetes. John Wiley & Sons, Ltd. http://www.wiley.com/ wileychi/diabetes/sample.html (1999) Staub JM, Garcia B, Graves J, Hajdukiewicz PT, Hunter P, Nehra N, Paradkar V, Schlittler M, Carroll JA, Spatola L, Ward D, Ye G & Russell DA - High-yield production of a human therapeutic protein in tobacco chloroplasts. Nat Biotechnol 18: 333-338 (2000) Turkenburg-van Diepen M. G. W. - Crystallographic studies of modified insulin (A Thesis Submitted to the University of York for the Degree of Doctor of Philosophy Department of Chemistry) -http:// www.yorvic.york.ac.uk/~mgwt/ thesis-tth/chapter1.html (1996) Venâncio T. M., Azevedo C. R., Oliveira A.E.A., Machado O.L.T., Cunha M. Da, Gomes V. M., Fernandes K. V. S. & Xavier-Filho, J. Presença de insulina durante o desenvolvimento de feijão-de-corda [Vigna unguiculata (L.) Walp.], em sementes de Canavalia ensiformis (L.) DC. e folhas de pata-devaca (Bauhinia forficata Link, mororó). 51 Congresso Nacional de Botânica, Brasília. Resumos Pg 14. (2000) Yu J, Langridge WH - Novel Approaches to Oral Vaccines: Delivery of Antigens by Edible Plants. Curr Infect Dis Rep 2: 73-77 (2000). + indica presença de insulina (antígeno). Biotecnologia Ciência & Desenvolvimento 41 CULTURA DE TECIDOS Biotecnologia aplicada à PRODUÇÃO DE MUDAS Produção de mudas micropropagadas de abacaxi 1. Importância da cultura do abacaxi João Batista Teixeira, Ph.D., Biologia Celular [email protected] Andréa Rachel Ramos Cruz, M.Sc., Fruticultura [email protected] Francisco Ricardo Ferreira, D.Sc. Fruticultura [email protected] Embrapa-Recursos Genéticos e Biotecnologia, Brasília, DF. José Renato Santos Cabral, M.Sc., Melhoramento de Plantas [email protected] Embrapa Mandioca e Fruticultura, Cruz das Almas, BA Fotos cedidas pelos autores 42 Biotecnologia Ciência & Desenvolvimento portadores de abacaxi são os Estados Unidos, a França, Alemanha e Holanda, que importaram, em 1998, 1.370.025, 181.449, 162.479 e 136.002 toneladas, respectivamente (FAO, 2000). O abacaxi (Ananas comosus (L.) Merril) é uma fruta tropical apreciada mundialmente pelo seu aroma e sabor acentuados. Além de apresentar propriedades medicinais, tem alto valor 2. Métodos de propagação nutritivo, sendo particularmente rico do abacaxizeiro em sais minerais e vitaminas. O consumo pode ser feito in natura ou procesA planta do abacaxi pode ser prosado na forma de compota, geléia, pagada de diversas formas, contudo sorvete, diferentes tipos de sobremesa sua propagação é predominantemene na indústria de confeitaria. te assexuada. A produção de mudas Essa cultura ocupa o nono lugar no ranque mundial das frutas, com uma produção de 13.444.203 toneladas de frutos. O maior produtor mundial de abacaxi é a Tailândia, com uma produção de 2.353.037 toneladas, em 1999. O Brasil está em segundo lugar, com uma produção de 1.740.840 toneladas de frutos, numa área plantada de 51.191 ha (FAO, 2000). O mercado mundial de exportação de abacaxi movimentou, em 1999, um volume de recursos da ordem de um bilhão de dólares, sendo as Filipinas, a Tailândia e a Costa Rica os principais países exportadores, com um volume de exportação de 582.691; 398.626 e 276.680 toneladas, respectivamente. O Brasil ocupou o sétimo lugar, com um voluFIGURA 1. Tipos de mudas produzidas me exportado de 15.796 topela planta de abacaxi, utilizadas na neladas. Os principais países impropagação convencional via semente, embora possível, é utilizada basicamente por melhoristas para fins de obtenção de híbridos entre diferentes genótipos, em programas de melhoramento genético. O método convencional de propagação do abacaxizeiro é feito por meio de mudas formadas a partir de brotações laterais da planta, denominadas filhote, filhote-rebentão ou rebentão (Figura 1). A coroa dos frutos pode constituir material propagativo, embora não seja muito utilizada, já que acompanha o fruto na época da comercialização. É possível também obter mudas através de métodos de seccionamento do caule, destruição do meristema apical, tratamento químico durante a diferenciação floral e por cultura de tecidos (Reinhardt & Cunha, 1999). brevipes, a broca do fruto (Thecla basalides), a broca do talo (Castnia icarus) e o ácaro Dolichotetranychus floridanus são as pragas que têm causado maiores danos à cultura (Sanches, 1999). Outras pragas como insetos, nematóides e sínfilos podem atacar o abacaxizeiro causando danos nas raízes, hastes, folhas e frutos, que contribuem para a redução da produtividade e/ou depreciação do fruto. Entre as principais, podem ser citadas o sínfilo (Hanseniella sp.), nematóides (Meloidogyne javanica, Meloidogyne incognita, Smoth Cayenne, Jupy e Pérola, são suscetíveis, essa doença constitui uma ameaça constante à abacaxicultura nacional. O fungo Fusarium subglutinans (Wollenw. & Reinking) Nelson, Tousson & Marasas comb. Nov. (Nelson et al., 1983), causador da fusariose, é capaz de atacar diferentes partes da planta do abacaxizeiro. No material propagativo (coroa, filhote, filhoterebentão e rebentão), o ataque ocorre na haste caulinar, causando lesões, exsudação de goma e infecção na base das folhas. Entretanto, em sua fase inicial, a fusariose pode passar despercebida e ser introduzida em áreas novas de plantio com mudas contaminadas. Assim, a movimentação de mudas infectadas constitui a principal forma de disseminação da doença dentro de uma mes3. Demanda por ma região ou mesmo mudas de alta entre regiões distanqualidade tes (Matos, 1987). Uma vez introduzido O uso de mudas numa determinada convencionais de bairegião, o patógeno é xa qualidade pode acardisperso pelo vento, retar problemas para a pela chuva, por imFIGURA 2. A-Haste caulinar de muda tipo filhote sem as folhas, lavoura a ser estabeleplementos agrícolas e com diâmetro da base de 3 cm e comprimento de 5 cm, aproxicida em conseqüência veículos, pelo próprio madamente; B-Segmentos da haste contendo uma gema cada; Cdo baixo vigor, ocasiohomem, e também Gemas em desenvolvimento, após 1 a 2 meses de cultivo nado principalmente por insetos, que, ao pela contaminação por pragas e doen- Pratylenchus brachyurus, Rotylenchu- visitarem uma planta infectada, acaças. lus reniformis), cupins (Cornitermes bam por levar em seu corpo o fungo e, A cochonilha (Dysmicoccus brevi- striatus, Syntermes silvestrii), formigas ao posarem em outras plantas, por pes) representa a principal praga da (Atta bisphaerica), percevejo (Lybin- inocular a doença. O solo, felizmente, cultura em todo o mundo e sua infes- dus dichrous), broca do colo (Paradi- não constitui um repositório de persistação leva ao aparecimento da mur- ophorus crenatus), caruncho (Parisos- tência do Fusarium, uma vez que o cha-do-abacaxizeiro. Embora a associ- choenus ananasi) e cochonilha pe- fungo não produz clamidosporos, o ação dessa cochonilha com a murcha quena (Diapsis bromeliae) (Sanches, que restringe a sobrevivência naquele tenha sido detectada no começo dos 1999). ambiente por períodos relativamente anos 30 (Carter, 1933), somente no Entre as enfermidades, a fusariose, curtos (Maffia, 1980; Matos & Cunha, início dos anos 60 foi que a etiologia da por sua vez, é considerada a principal 1980). Além disso, não tem sido consdoença começou a ser esclarecida (Car- doença do abacaxizeiro no Brasil, ten- tatada infecção de mudas sadias quanter, 1963). Esse autor demonstrou que do sido relatada inicialmente em frutos do plantadas em solo contaminado apenas cochonilhas retiradas de plan- da cultivar Smooth Cayenne (Kimati & pelo Fusarium (Matos & Cunha, 1980). tas doentes eram capazes de induzir os Tokeshi, 1964). Há evidências de que Várias outras doenças atacam o sintomas de murcha. Mais recente- a fusariose tenha sido introduzida no abacaxizeiro, como a mancha negra mente, foi isolado um virus a partir de Brasil através de mudas vindas do do fruto (Penicillium funiculosum), plantas com sintomas de murcha e Uruguai e da Argentina (Laville, 1980). podridão negra do fruto, podridão da acredita-se que esse virus associado à Atualmente, a doença causa danos base da muda e mancha branca das cochonilha seja o agente causal da elevados à cultura nas principais regi- folhas (Chalara paradoxa); podridão murcha (Gunasinghe & German, 1986; ões produtoras do país, com exceção do olho (Phytophthora nicotiana var. 1987; 1989; Ullman et al., 1989; San- do Tocantins, onde ela ainda não foi parasitica), podridão das raízes e poches & Matos, 1999). introduzida. Considerando que as prin- dridão do fruto verde (Phytophthora Além da cochonilha Dysmicoccus cipais cultivares plantadas no Brasil, cinnamomi); mancha amarela da planBiotecnologia Ciência & Desenvolvimento 43 FIGURA 3. Gemas em meio de multiplicação na presença de benzilaminopurina e ácido naftaleno acético, após três meses de cultivo ta e do fruto (vírus denominado Tomato Spotted Wilt Virus, transmitido por várias espéceis de Thrips); podridão rósea (causada por várias espécies de bactérias, como Acetobacter aceti, Erwinia herbicola e Gluconobacter oxydans); colapso do fruto e podridão bacteriana da roseta foliar (Erwinia chrysanthemi), e marbling desease (Acetobacter sp.) (Matos, 1999). Em 2001,o Brasil deverá ter uma área plantada de, aproximadamente, 55 mil ha, o que resultará numa demanda aproximada de 2 bilhões de mudas, para uma densidade de plantio de 36.000 mudas por ha. O sucesso da cultura do abacaxi depende, entre outros fatores, da qualidade da muda utilizada pelos agricultores. A sanidade do material propagativo constitui num dos pré-requisitos básicos para que possam ser obtidas altas produtividades e frutos de excelente qualidade. 3.1. Produção de mudas via cultura de tecidos A produção de mudas de abacaxi via cultura de tecidos consiste na regeneração de plantas completas (caule, folhas e raízes) a partir de gemas axilares de plantas matrizes selecionadas no campo em plantios comerciais. 44 Biotecnologia Ciência & Desenvolvimento O processo envolve várias etapas, começando com a coleta das mudas de plantas selecionadas no campo, passa pela extração das gemas axilares, cultivo e regeneração das plântulas, as quais são, numa segunda etapa, inoculadas em meio de multiplicação. Após essa fase, os brotos são cultivados em meio próprio para alongamento/enraizamento. Finalmente, as plântulas são transferidas para casa de vegetação para aclimatação, crescimento e desenvolvimento. Os detalhes desse processo são descritos a seguir. Mudas do tipo filhote, filhote-rebentão, rebentão ou coroa, retiradas da planta mãe, passam por uma poda das folhas e são levadas para o laboratório, onde podem ser armazenadas por vários dias até o momento da extração das gemas. Para retirada das gemas, faz-se um poda de dois a três centímetros da base da haste. Em seguida, são retiradas as folhas, deixando apenas a haste, a qual contém de dez a quinze gemas axilares bem visíveis (Figura 2-A). As hastes são colocadas em frasco autoclavado e imediatamente levadas para a câmara de fluxo laminar de ar estéril onde se procederá à desinfestação superficial para eliminar fungos e bactérias presentes na superfície da haste e das gemas. Caso haja contaminação elevada, além da desinfestação superficial recomenda-se adicionar antibióticos e/ou fungicidas ao meio de cultura para se obterem gemas livres de contaminação. Eventualmente, altos índices de contaminação bacteriana e/ou fúngica podem ser indícios de presença de contaminantes endógenos, o que é um indicador de que a qualidade da muda é insatisfatória, ou seja, a muda foi colhida de plantas matrizes doentes, debilitadas, excessivamente úmidas ou cuja manipulação e estocagem após a colheita tenham sido inadequadas. As mudas devem ser colhidas após um período de estiagem suficiente para que a água presente na superfície das folhas evapore, permitindo melhor armazenamento sem que ocorra proliferação superficial de fungos e bactérias. Caso o crescimento de fungos e bactérias seja muito intenso e as condições de umidade e temperatura, adequados, os feixes vasculares podem ser contaminados, o que irá dificultar ou mesmo impossibilitar o processo de assepsia das hastes e gemas. Nesse caso, é preferível utilizar outras fontes de mudas, associando a uma melhorara no processo de colheita, manipulação e estocagem do material antes do processamento em laboratório. A desinfestação é feita em duas etapas: primeiramente com álcool etílico comercial, na concentração de 50% a 70%, por 1 a 2 minutos. Em seguida, o álcool é drenado e adiciona-se uma solução de hipoclorito de sódio na concentração de 0,5% a 1%, por um período de 10 a 20 minutos. Não se deve utilizar água sanitária comercial por ela apresentar alto teor de hidróxido de sódio e, conseqüentemente, alto pH, o que pode acarretar a morte das gemas. Após a drenagem dessa solução, o material é lavado com água destilada estéril por 3 a 5 vezes, por um período mínimo de 5 minutos por lavagem. Ao final da última lavagem, o material é deixado imerso em água estéril até o momento da excisão das gemas, a qual é feita sob microscópio estereoscópico ou mesmo a olho nu. Com a ajuda de pinças e bisturis devidamente estéreis, é feita a retirada de um tetraedro irregular de 2 a 4 mm de aresta de tecido da haste contendo a gema (Figura 2-B). Esse fragmento é imediatamente colocado sobre a su- perfície do meio de cultura em um recipiente que pode ser um tubo de ensaio ou qualquer outro tipo de frasco de vidro ou plástico autoclavável. O meio de cultura tem a função de nutrir a gema e é constituído de macro e micronutrientes, vitaminas, aminoácidos, sacarose, agente gelificante e reguladores de crescimento vegetal. O meio básico utilizado é o de Murashige & Skoog (1962), suplementado com dois reguladores de crescimento, benzilaminopurina (BAP), também chamada benziladenina (BA), na concentração de 0,5 mg/L, e ácido naftalenoacético (ANA), na concentração de 0,125 mg/L. A primeira substância pertence ao grupo das citocininas e está relacionada com a diferenciação celular, formação e multiplicação de gemas e crescimento da haste caulinar e folhas; enquanto que a segunda, o ANA, faz parte das auxinas e age na mitose, formação e crescimento das raízes. Em combinação, essas substâncias são benéficas ao pegamento e desenvolvimento das gemas (Figura 2C). Após de dois a três meses de cultivo in vitro, há formação de uma plantinha de abacaxi completa, ou seja, folhas, haste e raízes, com tamanho aproximado de 8 a 10 cm de altura. A gema, inicialmente colocada em meio de cultura, permanece em sala de crescimento com temperatura em torno de 24 o C durante a noite e 28 o C durante o dia, sob a luminosidade correspondente a quatro lâmpadas fluorescentes de 40 W, tipo luz do dia. O fotoperíodo, i.é., o período diário de iluminação, deve ser de 14 a 16 horas. A cada quatro semanas, o material é transferido para meio fresco, até que se obtenha número desejado de plântulas, com três a cinco folhas, com raiz bem desenvolvida e um diâmetro do talo de 0,5 a 0,8 cm. Essas plântulas podem ser mantidas em meio de cultura por tempo indeterminado, desde que, a intervalos de quatro a seis meses, seja feita a renovação do meio nutritivo. Para cada cem gemas inoculadas, são obtidas, na melhor das hipóteses, o mesmo número de plântulas no final do processo. Entretanto, isso é raro acontecer, uma vez que ocorrem perdas por morte e contaminação. Normalmente, uma muda tipo filhote resulta em dez a quinze gemas isoladas e entre seis a dez plântulas estabelecidas. Portanto, a taxa de multiplicação nesse processo é de seis a dez por muda tipo filhote. Essa taxa de multiplicação é muito baixa e não compensa os custos de manipulação via laboratório. Dessa forma, é necessário lançar mão de um processo adicional, no qual as plântulas existentes no estoque in vitro possam ser multiplicadas centenas ou mesmo milhares de vezes. O processo de multiplicação consiste na poda das folhas e raízes das plântulas estocadas in vitro, com conseqüente redução do porte da haste para um a dois centímetros de comprimento. Esse material é inoculado em meio nutritivo gelificado ou líquido e mantido aí por vários meses para multiplicação das gemas pré-existentes na plântula (Figura 3). Para que ocorra uma intensa multiplicação das gemas, o meio nutritivo é enriquecido com BAP e ANA, na concentração de 2 e 0,5 mg/L, respectivamente, o que corresponde a quatro vezes a concentração do meio de estabelecimento. O intervalo de tempo de cultivo nessa fase varia de três a seis meses, com renovação do meio nutritivo a cada quatro semanas. Após esse período, procedese à transferência das gemas para meio sem reguladores de crescimento, denominado meio de alongamento/enraizamento. Nesse meio, as gemas, que se encontravam em processo ativo de multiplicação vão dar origem, após 45 a 60 dias, a pequenas mudas de abacaxi, com comprimento que varia entre 5 e 7 cm (Figura 4). Nessa fase, as mudinhas estão aptas a ser transferidas para telados ou casa de vegetação. Para isso, as mudas são tratadas com uma suspensão de Benlate a 0,1% por 1 h e imediatamente plantadas em substratos adequados em telados ou casa de vegetação. Diferentes tipos de substratos podem ser utilizados como mistura de solo e areia na proporção de 2:1, vermiculita pura, substratos comerciais do tipo Plantmax, solo turfoso ou qualquer outro tipo de substrato que seja propício ao crescimento das mudas. Durante essa fase, a luminosidade máxima deve ser de 40% da luz solar direta e a umidade relativa entre 70% e 80%. A adubação deve ser adequada, com um nível maior de fósforo e potássio em comparação com o nitrogênio. A adubação foliar com macro e micronutrientes pode dar bons resultados principalmente quando empregada de forma complementar. Após 6 a 8 meses em casa de vegetação, as mudas estão prontas para ser levadas para o campo (Figura 5). A taxa de multiplicação do abacaxi varia, principalmente, em função da cultivar, bem como do tipo de meio de cultura utilizado. Na Embrapa Recursos Genéticos e Biotecnologia, foi desenvolvida uma metodologia de produção de mudas in vitro para oito genótipos, sendo quatro comerciais, Pérola, Perolera, Smooth Cayenne e Primavera e quatro não comerciais FIGURA 4. Mudas de abacaxi com 5 a 7 cm de comprimento após cultivo em meio de alongamento/enraizamento por 45 a 60 dias Biotecnologia Ciência & Desenvolvimento 45 te a fase de multiplicação conduzidos em nosso laboratório mostraram que é possível reduzir o período de multiplicação para três meses, o que representa uma substancial redução do tempo total do processo. A produção de mudas de abacaxi em laboratório apresenta vantagens e desvantagens. Entre as vantagens, podemos citar as seguintes: - alto vigor e uniformidade; - ausência de pragas e doenças; - mudas enraizadas e prontas para ser cultivadas no campo; - disponibilidade de acordo com a demanda em termos de época e local de plantio; FIGURA 5. Mudas de abacaxi micropropagadas medindo entre 20 a 25 cm de comprimento após 6 a 8 meses em casa de vegetação, prontas para ser levadas ao campo (FRF-820, FRF-168, FRF-632 e Comum), clones procedentes do banco de germoplasma da Embrapa Mandioca e Fruticultura. A fase de indução de multibrotação das gemas foi feita em meio gelificado enquanto o alongamento/enraizamento foi conduzido em dois diferentes tipos de meio, apenas variando o grau de consistência, i. é, em meio líquido e em meio gelificado, para fins de comparação. A taxa de multiplicação média para os oito genótipos ao longo de cinco meses de cultivo em meio de multiplicação gelificado, seguido de dois meses em meio de alongamento/enraizamento gelificado foi de 482, i.é, foram produzidas 482 mudas a partir de cada plântula estabelecida in vitro. Quando a fase de alongamento/enraizamento foi conduzida em meio líquido, a taxa subiu para 1.676, um aumento de 348%. O uso de meio líquido na fase de alongamento/enraizamento tem outras vantagens adicionais como maior facilidade para retirar as mudas do meio de cultura por ocasião da transferência das mudas para a casa de vegetação. Portanto, considerando-se que, de 10 a 15 gemas inoculadas inicialmente a partir de cada muda tipo filhote, apenas 6 se desenvolvem e que a taxa de multiplicação seja apenas de 1.000 plântulas por gema, serão obtidas, ao final do processo, algo em torno de 6.000 mudas micropropagadas por muda convencional utilizada, conside46 Biotecnologia Ciência & Desenvolvimento rando uma perda próxima de zero durante a fase de aclimatação. Caso as 6.000 mudas sejam mantidas in vitro para uma nova rodada de multiplicação, algo próximo a 6.000.000 de mudas podem ser obtidas num período aproximado de 2 anos. Portanto, o potencial de multiplicação in vitro é muito elevado comparado com os métodos tradicionais. O cronograma do processo é outro ponto muito importante e pode ser resumido da seguinte forma: Entre as desvantagens, as mais importantes são as seguintes: - o custo da muda, algo entre 20 a 30 centavos, é bem superior ao da muda convencional, que está entre 3 a 5 centavos; - a produção depende de infraestrutura relativamente sofisticada; - o investimento inicial para montagem do laboratório, casas de vegetação e telados é alto; - o processo requer mão-de-obra especializada, conseqüentemente melhor remunerada; - a metodologia está em constante evolução, o que requer atualização freqüente do processo de produção. A. estabelecimento do estoque de plântulas a partir das gemas axilares 2 a 3 meses; B. multiplicação das gemas em meio rico em BAP e ANA 3 a 5 meses; C. alongamento/enraizamento 2 meses; D. aclimatação e crescimento em casa de vegetação 6 a 8 meses. Embora as mudas de abacaxi obtidas via cultura de tecidos ainda tenham preços elevados, principal fator que tem limitado a sua utilização em lavouras comerciais, esse tipo de muda pode ser recomendado com os seguintes objetivos: Assim o processo pode demorar de 13 a 18 meses para se obter mudas com 20 a 30 cm de altura, prontas para serem transferidas para o campo, partindo do início da fase de estabelecimento do estoque de plântulas in vitro. Da mesma forma que o meio líquido foi muito superior ao meio gelificado na fase de alongamento/enraizamento, é provável que o emprego do meio líquido na fase de multiplicação poderá acelerar o processo. Testes preliminares com meio líquido duran- 1. na introdução da cultura em novas regiões de plantio, onde ainda não existem problemas fitossanitários; 2. na introdução/substituição de novas cultivares, quando não se dispõe de mudas convencionais dessas cultivares para iniciar de grandes plantios; 3. na multiplicação rápida de genótipos selecionados pelos programas de melhoramento genético, antes do lançamento de novas cultivares; 4. na produção de material básico para atender a programas de produção de mudas certificadas de abacaxi; 5. no intercâmbio de germoplasma para se evitar a introdução de pragas e doenças exógenas. Conclusão A produção comercial de mudas de laboratório das variedades mais comuns de abacaxi plantadas no Brasil, como Smoth Cayenne, Pérola e Jupy, tem sido inviabilizada, como já foi dito, pelo alto custo de produção. Embora a taxa de multiplicação possa estar próxima a 10.000 por muda convencional, o custo final da muda é muito superior ao da muda convencional, cerca de 4 a 5 vezes. Na composição desse custo, estima-se que a maior parte, entre 60% a 70%, seja devido ao gasto com mão-deobra. Desta forma, para viabilizar este tipo de muda, protocolos mais eficientes no uso de mão-de-obra precisam ser desenvolvidos. Deve-se levar em consideração, entretanto, que, para outras aplicações listadas anteriormente, o protocolo está pronto para ser utilizado, como, por exemplo, para produção de mudas de novas variedades derivadas do melhoramento genético. Mesmo para variedades tradicionais, é possível aplicar esta tecnologia através da comercialização de mudas de laboratório para estabelecimento de jardins clonais, em regiões e locais estratégicos para essa finalidade. Nesse caso, as mudas poderiam ser adquiridas por viveiristas, que multiplicariam as matrizes no campo, sob condições de controle rigoroso de pragas e doenças. As mudas matrizes adquiridas de laboratórios credenciados poderiam resultar em uma taxa de multiplicação mínima de 5 vezes por ciclo, além da produção de um fruto por planta. Desde que se tomem os devidos cuidados, o material poderia passar ainda por, pelo menos, uma segunda multiplicação no campo. Dessa forma, por exemplo, 50.000 mudas matrizes de laboratório renderiam, ao longo de dois ciclos de multiplicação no campo, algo em torno de 1.250.000 mudas, além de produzir, no mesmo período, 300.000 frutos (50.000 fru- tos no primeiro ciclo e 250.000, no segundo), numa área de cinco a sete hectares e num período de tempo aproximado de trinta e seis meses. Portanto, a presente metodologia poderá contribuir, em futuro próximo, para melhorar a qualidade da muda para cultura do abacaxi, em âmbito nacional, o que resultará num aumento de produtividade e qualidade do fruto produzido. Pelas características da planta, que pode suportar períodos temporários de seca, adapta-se a diferentes condições de clima e solo, e pelas características do fruto, quanto a sabor, aroma, riqueza nutricional, a cultura do abacaxi deverá se firmar como a terceira mais importante fruta para o mercado interno, depois da laranja e da banana, com excelentes condições para se tornar mais um importante produto de exportação da agricultura brasileira. Referências 1. FAO. Disponível no site da FAO (2000): http://apps.fao.org/page/ form? collection = Production. Crops. Primary&Domain= Production & servlet = 1&language = EN&hostname= apps.fao.org&version=default; 2. Carter, W. A wilt of pineapple similar to mealybug wilt but caused by drought. Pineapple Quarterly, v.3, p.181-184, 1933; 3. Carter, W. Mealybug wilt of pineapple a reappraisal. Annals of the New York Academy of Science, v.105, n.13, p.741-764, 1963; 4. Reinhardt, D.H.R.C. & Cunha, G.A.P.da. Métodos de Propagação. In: Cunha, J.R.S. & Souza, L.F. da S., eds. O Abacaxizeiro. Cultivo, Agroindústria e Economia. Brasília: Embrapa Comunicação para Transferência de Tecnologia, p.105-138, 1999; 5. Gunasinghe, U.B. & German, T.L. Association of virus particle with mealybug wilt of pineapple. Phytopathology, v.76, p.1073 (Abstract), 1986; 6. Gunasinghe, U.B. & German, T.L. Further characterization of a virus associated with mealybug wilt of pineapple. Phytopathology, v.77, p.1776 (Abstratct), 1987; 7. Gunasinghe, U.B. & German, T.L. Purification and parcial characterization of a virus from pineapple. Phytopathology, v.79, n.12, p.13371341, 1989; 8. Kimati, H.; Tokeshi, H. Nota sobre a ocorrência de Fusarium sp. causando resinose fúngica em abacaxi. Revista de Agricultura, v.39, n.3, p.131-133, 1964; 9. Laville, E. La fusariose de l´ananas au Brésil. I-synthèse des connaissences actuelles. Fruits, v.35, n.2, p.101-113, 1980; 10. Nelson, P.E.; Tousson, T.A.; Marasas, W.F.O. Fusarium species na illustrated manual for identification. [S.l.]:The Pennsylvania State University Press, University Park and London, 1983. 193p; 11. Maffia, L.A. Sobrevivência de Fusarium moniliforme Sheld. var. subglutinans WR & RG. no solo e em restos culturais e sua erradicação de mudas de abacaxi (Ananas comosus L.) Merril) através de tratamento térmico. Fruits, v.35, n.4, p.217-243, 1980; 12. Matos, A.P. de. Pineapple fusariosis in Brazil: an overview. Fruits, v. 41, n.7/8, p.417-422, 1987; 13. Matos, A.P. de. Doenças e seu controle. In: Cunha, J.R.S. & Souza, L.F. da S., eds. O Abacaxizeiro. Cultivo, Agroindústria e Economia. Brasília: Embrapa Comunicação para Transferência de Tecnologia, p.269-305, 1999; 14. Matos, A.P. de; Cunha, G.A.P. Persistência e capacidade infectante de Fusarium moniliforme no solo. Pesquisa Agropecuária Brasileira, v. 15, n.2, p.163-165, 1980; 15. Murashige, T. & Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant., n.15, p.473-497, 1962 ; 16. Sanches, N.F. Pragas e seu controle. In: Cunha, J.R.S. & Souza, L.F. da S., eds. O Abacaxizeiro. Cultivo, Agroindústria e Economia. Brasília: Embrapa Comunicação para Transferência de Tecnologia, p.307-341, 1999; 17. Sanches, N.F. & Matos, A.P. Murcha associada à Cochonilha Dysmicoccus brevipes (Cockerel, 1893). In: Cunha, J.R.S. & Souza, L.F. da S., eds. O Abacaxizeiro. Cultivo, Agroindústria e Economia. Brasília: Embrapa Comunicação para Transferência de Tecnologia, p.343-366, 1999; 18. Ullman. D.E.; German, T.L.; Gunasinghe, U.B.; Ebesu, R.H. Serology of a closteroviruslike particle associated with mealybug wilt of pineapple. Phytopathology, v. 79, n.12, p.13411345, 1989; Biotecnologia Ciência & Desenvolvimento 47 Vacina para o HPV Uma alternativa para a erradicação do câncer de colo do útero Grupo de Detecção Precoce e Prevenção do Câncer de Colo do Útero Hospital de Clínicas de Porto Alegre (HCPA) Coordenador: Dr. Paulo Naud Participantes: Dr. Paulo Naud, Dr. Jean Matos, Dr. Luciano Hammes e Dra. Janete Vetorazzi [email protected] [email protected] O câncer de colo uterino é o 3°câncer mais para cada 100.000 mulheres, com 6.900 vidas percomum em mulheres, no mundo todo. Anualmente, didas naquele ano por essa patologia. Em países cerca de 700.000 novos casos são diagnosticados. Em como a Finlândia, a incidência de câncer de colo países desenvolvidos, o risco de uma mulher desenuterino alcança apenas a marca de 4,4 casos / volver câncer cervical durante sua vida, sem nunca ter 100.000 mulheres /ano. participado de um programa de prevenção, é de cerca de 1%. Em países em desenvolvimento, esse risco Nos últimos anos, identificou-se um aumento na aumenta para 5%. Esses números refletem claramente incidência e na mortalidade por câncer de colo a diferença das políticas de saúde públicas adotadas uterino em nosso meio. Estudos demonstram uma e reforçam a importendência crescente tância dos programas dessa patologia e de prevenção para a enfatizam a necessiConsideramos que o câncer de colo do útero erradicação dessa dodade tanto de proença. gramas adequados seja uma doença com um potencial de de screening, como prevenção que pode chegar a 100% dos A introdução do exade programas de me citopatológico há identificação e tratacasos, e que nossa atuação - dos profissionais 50 anos, como métomento precoce de do de rastreamento, lesões precursoras de saúde, do governo e da população em provocou uma espedo câncer de colo tacular redução dos uterino. geral - pode mudar a história dessa doença. casos de morte por câncer de colo uterino. Novas alternativas de Em países onde se ofereceu uma efetiva cobertura tratamento devem ser incorporadas à prática de pelo screening, deu uma redução de até 70% dos todos os profissionais de saúde envolvidos na casos. prevenção do câncer de colo do útero.Também seria fundamental iniciarmos nossa atuação precoO Brasil, sendo um país em desenvolvimento, possui cemente, antes do surgimento das lesões precursouma incidência alarmante de câncer cervical. Em ras do câncer de colo do útero. Devemos impedir 1999, o INCA (Instituto Nacional do Câncer do Brasil) que nossas pacientes se infectem pelo papiloma estimou que ocorreram 30,34 casos de câncer cervical vírus humano (HPV), um DNA vírus que é o grande 48 Biotecnologia Ciência & Desenvolvimento 48 Biotecnologia Ciência & Desenvolvimento responsável pelo desenvolvimento do câncer de colo do útero. Podemos afirmar, baseados em estudos de biologia molecular, que não existe câncer de colo do útero sem que exista a infecção pelo HPV. Até o momento, conseguimos identificar, por métodos de captura híbrida e por PCR, cerca de 100 subtipos de HPV, que possuem capacidade de produzirem diferentes tipos de lesões, desde lesões exofíticas externas, verrucosas, até lesões cervicais precursoras a lesões invasivas do câncer de colo do útero. O estudo envolverá cerca de 1000 mulheres em centros selecionados nos Estados Unidos, no Canadá e no Brasil, e avaliará a resposta imunológica das pacientes vacinadas contra o HPV. Foram selecionadas pacientes que procuraram espontaneamente nosso serviço e que não tivessem qualquer indício de infecção pelo HPV. Depois de exames iniciais realizados fora do país, as pacientes foram selecionadas para receberem a vacina ou placebo. Todas as pacientes serão acompanhadas por exames que rastreiam a infecção pelo HPV, tanto por exames citopatológicos quanto por PCR e por sorologia que identifica anticorpos específicos contra o HPV, durante um período de cerca de 18 meses. O HPV pode infectar tanto a pele como as mucosas, Ressaltamos que as pacientes serão vacinadas com uma porém ele tem um grande potencial de atuação na partícula que imita o DNA dos subtipos de HPV 16 e 18, mucosa de colo do chamada de VLP ( vírus like útero, que possui uma particle), que estimula o velocidade alta de organismo a produzir Informamos, com satisfação, que estamos desenturn over celular. anticorpos específicos a essa volvendo, em parceria com SmithKline Beecham Dos cerca de 100 partícula que imita o HPV subtipos identificados 16 e 18. Imagina-se que, Biologicals, um estudo multicêntrico, que testará a existe um grupo de quando as pacientes entraHPV que possui a carem em contato com as eficácia de uma vacina para o HPV. pacidade de invadir o partículas virais verdadeigenoma celular, os ras, o organismo produzirá quais são classificados como HPV do grupo B ou de anticorpos imediatamente, impedindo que essas pacienalto potencial oncogênico, tendo a capacidade de tes se infectem pelo HPV 16 e 18 e talvez, por reação produzir alterações celulares que podem levar, sob cruzada, também possam vir a produzir anticorpos contra determinadas condições, ao câncer de colo do outros subtipos virais. útero. Lembramos que as pacientes não receberão nenhuma Sabemos que desse grupo, considerado de alto partícula viral e que, por isso, não correm nenhum risco potencial oncogênico (subtipos 16, 18, 33, 35, 51) de desenvolverem qualquer lesão cervical.Todas as pacios subtipos 16 e 18 são os principais responsáveis entes serão acompanhadas por 18 meses e a eficácia da pelo desenvolvimento do câncer de colo do útero, vacina será avaliada através da formação de anticorpos tanto pela sua prevalência quanto pela sua específicos para o HPV e também através de outros agressividade. exames que detectem possíveis lesões causadas pelo HPV. Baseados em estudos de biologia molecular, podemos dizer que nossa ação tem como objetivo a prevenção da infecção desses subtipos virais. Vários estudos estão sendo desenvolvidos para se chegar a uma vacina eficaz para prevenir a infecção pelo HPV dos subtipos 16 e 18. Podemos dizer que a melhor alternativa de tratamento para infeções virais seria a estimulação do sistema imunológico de nossas pacientes contra o vírus do HPV. Informamos, com satisfação, que estamos desenvolvendo, em parceria com SmithKline Beecham Biologicals, um estudo multicêntrico, que testará a eficácia de uma vacina para o HPV e que cerca de 100 pacientes serão acompanhadas no Hospital de Clínicas de Porto Alegre. Consideramos que o câncer de colo do útero seja uma doença com um potencial de prevenção que pode chegar a 100% dos casos, e que nossa atuação - dos profissionais de saúde, do governo e da população em geral - pode mudar a história dessa doença. Talvez possamos num futuro próximo, dizer que, assim como o exame papanicolau mudou a história dessa patologia, a vacina contra o HPV foi um marco para a erradicação desse câncer, câncer de mulheres e, principalmente, câncer de países pobres, como o Brasil. Informamos que a Revista Biotecnologia Ciência & Desenvolvimento está sendo indexada para a AGROBASE - Base de Dados Bibliográfica da Agricultura Brasileira. A AGROBASE é gerenciada pela CENAGRI - Ministério da Agricultura e do Abastecimento. Biotecnologia Ciência & Desenvolvimento 49 Biotecnologia Ciência & Desenvolvimento 49 PESQUISA Processo fermentativo para PRODUÇÃO DE BEBIDA Processo Fermentativo para Produção de Bebida Alcoólica de Pupunha (Bactris gasipaes Kunth) Resumo Lílian Pantoja de Oliveira Bióloga, MSc Laboratório de Bioquímica e Fisiologia Pós-colheita/ CPTA/INPA Roberto Maeda Engenheiro Agrônomo Laboratório de Bioquímica e Fisiologia Pós-colheita/ CPTA/INPA Jerusa de Souza Andrade Engenheira Agrônoma, DSc Laboratório de Bioquímica e Fisiologia Pós-colheita/CPTA/INPA Nei Pereira Junior [email protected] Engenheiro Químico, PhD Laboratório de Desenvolvimento de Bioprocessos do Departamento de Engenharia Bioquímica da EQ/UFRJ Sônia Maria da Silva Carvalho Farmacêutica, MSc Laboratório de Micologia do Departamento de Parasitologia do ICB/UA. Spartaco Astolfi Filho [email protected] Biólogo, DSc Laboratório de Genética do Centro de Apoio Multidisciplinar do ICB/UA. Fotos cedidas pelos autores 50 Biotecnologia Ciência & Desenvolvimento A viabilidade técnica de produzir de bebida alcoólica a partir de pupunha (Bactris gasipaes Kunth), fruto amiláceo da Região Amazônica, foi investigada por fermentação semi-sólida. A polpa do fruto foi caracterizada quimicamente, sendo exibido alto conteúdo amiláceo (44,3%, em base úmida). Após a hidrólise enzimática prévia do amido, com α-amilase e glucoamilase comerciais, o meio foi inoculado com células de Saccharomyces cerevisiae (levedura de panificação) e o processo fermentativo monitorado em intervalos de 24 horas, para acompanhar a viabilidade celular, o consumo de substrato, a produção de etanol, a acidez total e o teor de carotenóides totais. O alto rendimento em polpa amilácea (71,2%), as elevadas conversões de hidrólise (90%) e eficiência de fermentação (93,5%), assim como a boa aceitabilidade do produto (80%), sugerem a aplicação da pupunha na produção de bebida alcoólica fermentada, gerando oportunidades para novos desenvolvimentos econômicos na região amazônica. Palavras chave: pupunha, composição química, α-amilase, glucoamilase, fermentação, Saccharomyces cerevisiae, análise sensorial. Introdução A pupunheira (Bactris gasipaes Kunth) é uma palmeira domesticada pelos ameríndios desde épocas pré-colombianas. Seu cultivo é feito em toda a Amazônia e constitui-se em valiosa planta de subsistência (Figura 1). Tem ampla distribuição, sendo encontrada por toda a Amazônia e América Central a partir de Honduras (Figura 2). Seus frutos têm grande potencial econômico devido à sua composição química, produtividade agrícola (13.500 ton/ano) e amplo consumo regional (Clement, 2000). A pupunha é um fruto rico em minerais e pró-vitamina A e, dependendo da raça, em carboidratos ou lipídios (Arkoll & Aguiar, 1984). A partir da pupunha é preparada a caiçuma, bebida consumida pelos índios em datas comemorativas. É feita artesanalmente de diferentes modos, de acordo com a etnia. Algumas tribos mastigam a pupunha e a deixam fermentar, geralmente por até sete dias, e, dessa forma, a ptialina contida na saliva Figura 1. Pupunheira amazônica Foto: C. R. Clement é incorporada à massa, e hidrolisa o amido, conseqüentemente. Apesar do enorme potencial da pupunha na Região para a produção e comercialização de bebidas, esse aspecto tem sido pouco explorado, pois a maioria dos produtos oriundos daquele fruto é processada de forma artesanal. O presente trabalho teve como objetivo desenvolver a tecnologia para produzir uma bebida alcoólica, por processo Figura 2. Distribuição geográfica da pupunheira fermentativo, utilizando-se a pupunha como matéria-prima. Materiais e Métodos Matéria-prima e caracterização Foram utilizados frutos da pupunheira no estádio maduro, pertencentes à raça Solimões, colhidos em safra secundária e adquiridos no mercado Adolpho Lisboa, em Manaus. Os cachos foram pesados e, após a despenca, os frutos foram selecionados quanto à sanidade e à maturação, avaliados em relação ao número, peso, diâmetro longitudinal e transversal e peso do descarte (semente e casca) e lavados em água corrente. Os frutos foram, então, submetidos à cocção por 5 minutos em tacho de aço inoxidável com capacidade para 100 litros, aquecido a vapor, com agitação mecânica. Após a drenagem da água e o Figura 3. Pré-tratamento da matéria prima resfriamento, foram acondicionados em sacos plásticos de alta densidade e estocados a 10 ºC. Para a caracterização centesimal e o preparo do meio de fermentação, os frutos foram descongelados e após descascamento e retirada de semente, a polpa foi caracterizada (em triplicata), quanto às proteínas solúveis, pelo método de Biureto (Harris & Angal, 1994) e carotenóides totais, por espectrofotometria (Higby, 1962). Os lipídios (extração com éter de petróleo em Soxhlet), fibra total (tratamento ácido-básico), umidade (65ºC) e cinzas (incineração em mufla a 550 ºC) seguiram metodologias descritas pelas Normas Analíticas do Instituto Adolfo Lutz (1985). O amido foi extraído e hidrolisado segundo Ranganna (1996) e a glicose liberada, quantificada pelo método de Somogyi-Nelson (Southgate, 1976). Enzimas e Microrganismo A hidrólise do amido presente na pupunha foi realizada com as enzimas comerciais αamilase e glucoamilase (NOVO Nordisk S/A) e a levedura Saccharomyces cerevisiae (fermento seco Fleischmann) foi empregada como agente do processo fermentativo. Preparo do Meio de Fermentação Analogamente à caracterização da matéria-prima, a massa de fermentação foi preparada pelo descongelamento dos frutos, seguido de cocção por 2,5 horas, resfriamento, despolpa, trituração e esterilização em autoclave por 15 minutos, a 121 ºC. Para otimização das condições de hidrólise, foram realizados testes preliminares, com diferentes proporções de α-amilase (70ºC) e glucoamilase (55 ºC) em banho termostatizado, por 30 minutos. Os graus de hidrólise foram avaliados mediante a quantificação dos açúcares redutores, pelo método de Somogyi-Nelson (Southgate, 1976). A fim de avaliar a fermentabilidade do meio hidrolisado, inoculou-se o mesmo com células de levedura em fermentômetros (recipiente amplamente utilizado em indústria viti-vinícola para se medir a atividade fermentativa de linhagens de leveduras), possibilitando o acompanhamento do processo por meio de pesagens sucessivas do conjunto, sendo a perda de peso observada, decor- rente do desprendimento de CO2. As condições de hidrólise enzimática, eleitas como ótimas, foram empregadas para o preparo do meio de fermentação e o teor de açúcar inicial, corrigido com a adição de xarope de sacarose estéril para se obter a concentração desejada de etanol na bebida. Processo Fermentativo A fermentação semi-sólida foi conduzida por batelada simples com três repetições, em cubas cilíndricas de acrílico, geometricamente iguais, com capacidade nominal de 4,5 litros, a 29 oC e o processo monitorado a intervalos de 24 horas. De cada cuba, amostras foram retiradas para determinações analíticas (triplicatas) das variáveis de interesse do processo fermentativo. Determinações Analíticas Além das análises físico-químicas para a determinação de amido e carotenóides, empregadas na caracterização da matéria-prima, foram ainda utilizadas as seguintes técnicas para o acompanhamento do processo fermen- Tabela 1. Composição química da pupunha (Bactris gasipaes Kunth) in natura, pertencente a raça Solimões Constituintes* Umidade (%) Proteínas (%) Lipídios (%) Amido (%) Fibra Total (%) Cinza (%) Carotenóide (mg/100g) in natura 46,1 ± 0,4 1,3 ± 0,1 5,9 ± 0,4 44,3 ± 0,3 0,7 ± 0,1 0,7 ± 0,2 2,5 ± 0,3 * base úmida Tabela 2. Análises físico-químicas da bebida fermentada Constituintes PH Acidez (g/L) Açúcares redutores (g/L) Sólidos solúveis (Brix) Etanol (g/L) Carotenóides (µg/L) Média* 3,9 2,7 0,16 6,4 75,1 7,0 * Os valores representam média de triplicatas e os desvios não ultrapassaram 1,8 % das medidas realizadas Biotecnologia Ciência & Desenvolvimento 51 tativo: avaliação da viabilidade celular, por contagem de células em Câmara de Neubauer, empregandose azul de metileno como corante vital (Alves e Morais, 1998); teor alcoólico, após destilação de amostras em fase aquosa e posterior medida, por densimetria (Normas Analíticas do Instituto Adolfo Lutz, 1985); pH, por potenciometria; acidez, por titulometria com NaOH 0,01N; sólidos solúveis, por refratometria e açúcares redutores, pelo método de Somogyi-Nelson (Southgate, 1976). Variáveis de Resposta Os valores de extensão da hidrólise enzimática do amido (EH), fator de conversão de substrato consumido em etanol (YP/S), e a produtividade em etanol (QP) foram calculados de acordo com as seguintes expressões: EH (%) = [(Ao – Af)/Ao] x 100 YP/S = (∆P)/(-∆S) = (Pf – Po)/(So – Sf) [=] gP/gS QP = (Pf – Po)/ tF [=] gP/L.h Onde: A: amido; P: concentração de etanol; S: concentração de subtrato; tF: tempo de fermentação. Os índices o e f representam as condições inicial e final, respectivamente. Preparo da Bebida e Análises Sensoriais Após sete dias de fermentação, procedeu-se à decantação, filtração, acondicionamento em garrafas de vidro de 1 litro, fechamento hermético, pasteurização por imersão das garrafas em água a 85 ºC por dez minutos, resfriamento em banho de água com gelo, e estocagem a, aproximadamente, 6 ºC. Ao final de 30 dias, a bebida foi avaliada quanto ao pH, açúcares redutores residuais (Somogyi-Nelson), sólidos solúveis, grau alcoólico e carotenóides totais (Higby, 1972). Foram realizados testes preliminares para se determinar o grau de doçura ideal na bebida. Essas formulações (0, 3, 6, 9 e 12% (p/v) de sacarose) foram submetidas à análise sensorial, utilizando-se escala hedônica (Monteiro, 1984). A bebida com o grau de edulcoração eleito foi analisada sensorialmente por 35 pessoas não treinadas, avaliando-se as 52 Biotecnologia Ciência & Desenvolvimento ção como matéria prima para produção de bebidas alcoólicas. De acordo com Clement (1987), há uma grande variabilidade em relação à composição de pupunhas encontradas na região Amazônica, havendo raças que produzem frutos pequenos, fibrosos, com elevados teores de lipídeos e baixos conteúdos de amido ou vice-versa. Hidrólise do Amido Figura 4. Efeito das combinações de α-amilase (a) e glucoamilase (g) na hidrólise do amido de pupunha características de sabor, aparência, cor e aroma, pelo método descrito por Monteiro (1984). Os resultados foram analisados estatisticamente, aplicando-se a análise de variância e teste de Tukey, com nível de significância de 5%. Resultados e Discussão Características da matéria-prima Os cachos de pupunha pesavam entre 0,9 a 5,0 kg e continham de 25 a 150 frutos, com peso médio de 36 g e diâmetros longitudinal e transversal de 4,5 e 4 cm, respectivamente. O rendimento em polpa, considerado elevado, foi de 71,2% em relação ao fruto integral. A Tabela 1 mostra a composição da polpa in natura, considerando-se os principais componentes. O alto conteúdo de amido e o baixo teor de lipídeos dos frutos indicam sua utiliza- Para a hidrólise do amido, utilizaram-se as enzimas α-amilase e glucoamilase, como descrito na seção de Materiais e Métodos. Os resultados estão exibidos no gráfico da Figura 4, onde se observa, inicialmente, a imprescindibilidade da α-amilase para uma hidrólise mais eficiente do amido. As melhores quantidades de enzimas para o processo hidrolítico foram atingidas com 10 µL e 50 µL de α-amilase e glucoamilase, respectivamente, tendo sido essa proporção utilizada para o preparo do meio de fermentação. A extensão da hidrólise do amido presente na pupunha foi de, aproximadamente, 90 %. A alta eficiência da hidrólise foi também confirmada em experimento paralelo, conduzido em fermentômetro, onde se observou expressiva evolução de CO2 (Figura 5). Apesar do curto tempo de fermentação (em torno de 4 horas), a alta concentração de células e as condições de anaerobiose no fermentômetro, contribuíram para que o etanol fosse produzido com apreciável fator de conversão do substrato consumido no produto (0,38 g/g), considerando-se ser esse um ensaio preliminar conduzido em condições de fermentação rápida (com inóculo maciço). Processo fermentativo Figura 5. Evolução de CO2 na fermentação do meio hidrolisado enzimaticamente O meio de fermentação submetido à hidrólise prévia do amido foi adicionado de xarope de sacarose, nas cubas de fermentação, formando-se um sistema semi-sólido de massa homogênea e pastosa. A formação de bolhas de gás (CO2) no interior da massa foi observada cerca de três horas após a inoculação e tornou-se menos intensa após 48 horas. A coloração alaranjada do meio permaneceu inalterada e a consistência da massa tornou-se mais fluida. O teor de sólidos solúveis, compos- to em sua maioria por açúcares provenientes da hidrólise do amido e da adição do xarope, decresceu em função do consumo de açúcares fermentáveis. A produção máxima de etanol (12% v/v) foi atingida no período de 72 horas após o início da fermentação, diminuindo ligeiramente até o final do processo (Figura 6), sugerindo a possibilidade de redução no tempo de fermentação, desde que essa estratégia não interfira nas características de sabor e aroma da bebida. Com 72 h de fermentação, o rendimento de substrato consumido em Figura 6. Perfil cinético da produção etanol foi de 0,49 g/g, corresponden- de etanol em meio de fermentação do a uma eficiência de fermentação com amido de pupunha hidrolisado de 95,5 %. A produtividade máxima foi alcançada no tempo de 24 horas, apresentando o valor de 2,6 g de etanol/L.h. O perfil de consumo de açúcares redutores totais (Figura 7) coadunase com o de produção de etanol e com a variação de sólidos solúveis (Figura 6). Observa-se que o maior consumo de açúcar ocorre nas primeiras 24 horas do processo, diminuindo, posteriormente devido às elevadas concentrações de etanol e a escassez de substrato. Figura 7. Cinética do consumo de O decréscimo do pH e, conseaçúcares redutores totais (ART) na qüentemente, o aumento da acidez fermentação alcoólica de hidrolisado do meio de fermentação (Figura 8) de pupunha estão associados à formação de ácidos orgânicos (ácidos succínico, lático, acético e outros), como amplamente reportada na literatura (Ribéreau-Gayon & Peynaud, 1966). É importante ressaltar que a evolução da acidez durante a fermentação influencia a estabilidade e a coloração de bebidas fermentadas (Rizzon et al,1998), assim como valores de pH entre 3 e 4 dificultam contaminações bacterianas (Aquarone et al., 1986). Os carotenóides totais mantiveram-se inalterados (4,7 mg/100g) durante todo o curso da fermenta- Figura 8. Variação da acidez do meio na fermentação alcoólica de hidrolisado de ção. A taxa de mortandande celular pupunha aumentou com o curso da fermentação, tornando-se mais acentuada após seqüência ocorre a liberação de meta96 horas, devido à alta concentração de bólitos importantes para o meio exteretanol no meio fermentado (12,2% v/ no, bem como a entrada de substânciv). Na fermentação alcoólica, o princi- as, através da membrana, de forma não pal responsável pela diminuição da seletiva (Duarte et al., 1996; Jones, viabilidade celular é o seu próprio 1988; Pereira Jr., 1999). Isso resulta na produto, cuja ação tóxica reflete-se na perda da viabilidade celular (Figura 9). desorganização da membrana citoplasÉ importante ressaltar que o micrormática, de composição fosfolipídica, ganismo agente do processo fermentaalterando sua integridade. Como con- tivo foi uma levedura de panificação, que não era a mais adequada à produção de etanol, tendo em vista que o processo industrial para a sua produção, a batelada alimentada, é conduzida em condições de elevada aeração e baixas concentrações de glicídeos no meio, a fim de minimizar os clássicos fenômenos de repressão catabólica. Como conseqüência às condições impostas no presente trabalho, que visavam a produção de etanol por batelada simples com elevadas concentrações de glicídeos em sistema não aerado, a levedura apresentou bom desempenho no que tange à atividade fermentativa, ainda que apresentasse elevadas taxas de mortandade, decorrentes do efeito deletério de seu próprio produto. Características gerais da bebida A bebida fermentada apresentou sabor e aroma agradáveis, aspecto límpido e coloração amareloalaranjado (Figura 10). As características físico-químicas (Tabela 2) mostram que a concentração de etanol (75,11 g/L) foi superior à encontrada (11,83 g/L) por Sotero (1996), estudando a produção de caiçuma de pupunha por fermentação espontânea. A maioria dos carotenóides totais presentes na massa foi removida juntamente com os lipídios durante o processo de filtração para obtenção da bebida, o que seguramente está atrelado às características lipossolúveis desses terpenóides. O teor de açúcar na bebida foi bastante reduzido, decorrente do consumo praticamente total durante o processo de fermentação. Considerando não haver um grau de edulcoração definido, a escolha da proporção de açúcar efetuada através de análise sensorial mostrou que a proporção de sacarose de maior preferência foi de 9 %, com 85,7 % de aceitação. A fim de se verificar a aceitabilidade da bebida já edulcorada, procedeu-se a uma nova análise sensorial cobrindo-se os requisitos aparência, aroma, sabor e cor. Os resultados desse ensaio estão apresentados na Figura 11, na qual se registram graus satisfatórios, superiores a 3,5. Alguns provadores afirmaram que o aroma exalado pela bebida asseBiotecnologia Ciência & Desenvolvimento 53 Figura 9. Viabilidade celular de S. cerevisiae durante o processo de produção da bebida alcoólica do hidrolisado de pupunha de, e de um bom retorno de investimento de capital. O presente trabalho demonstra ser factível produzir bebida alcoólica de pupunha, fruto abundante na região Amazônica e amplamente consumido pela população do Norte brasileiro. O desenvolvimento dessa tecnologia exigiu hidrólise enzimática prévia, tendo em vista a expressiva composição amilácea do fruto, seguida de processo fermentativo. A bebida resultante apresentou alto teor alcoólico e excelentes propriedades organolépticas. Tais características sinalizam um futuro promissor para esse inexplorado mercado. Figura 10. Bebida alcoólica de pupunha melha-se ao de mel e manga. De uma forma geral, a bebida obteve um bom nível de aceitabilidade; em torno de 80%. Conclusões O Brasil é a maior reserva continental de solos agricultáveis de potencial ainda não integralmente aproveitado. Mesmo assim o nosso país é o terceiro maior exportador mundial de alimentos. Sabe-se, ainda, que o mercado de bebidas convencionais responde por uma parcela significativa da economia brasileira, encontrando-se nesse segmento industrial as maiores empresas da área de alimentos. No entanto, apesar de se constatar uma crescente demanda por distintos sabores de origem natural, a comercialização de novas bebidas constitui um mercado pouco explorado. Nesse contexto, a produção de bebidas fermentadas, oriunda de frutos tropicais da região Amazônica, desponta como uma interessante alternativa para novos mercados por apresentar perspectivas de alta lucrativida54 Biotecnologia Ciência & Desenvolvimento Figura 11. Diagrama gráfico do teste classificatório do perfil da bebida Agradecimentos À CAPES, pela concessão de bolsa de mestrado para a primeira autora e ao BASA (Banco da Amazônia S.A.), que acreditou em nossa proposta, aprovando um projeto que recentemente se iniciou no âmbito do PROBEM/BIOAMAZÔNIA, para continuidade deste trabalho. Referências Bibliográficas ALVES, S. B. & MORAES, S. A. (1998). Quantificação de inóculo de patógenos de insetos. In: ALVES, S. B. Controle Microbiano de Insetos. Ed. FEALQ, Piracicaba-SP, 765-774. AQUARONE. E; LIMA, U. A. L. & BORZANI, W. (1986). Alimentos e Bebidas Produzidos por Fermentação. Ed. Edgard Blücher Ltda, São Paulo-SP. v. 5, 243p. ARKCOLL. D.B. & AGUIAR, J.P.L. (1984). Peach palm (Bactris gasipaes) a new source of vegetable oil from the tropics. J. Sci. Food Agric., 35, 520-526,. CLEMENT, C. R. & MORA-URPI, J. E.. Pejibaye Palm (Bactris gasipaes H. B. K. Aracaceae): Multi-use potencial for the lowland Humid Tropics. (1987). Jour. Econ. Bot., 42 (2), 302-311. CLEMENT, C. R. (2000). Comunicação pessoal, Instituto Nacional de Pesquisas da Amazônia, Manaus-AM. DUARTE, M. C. T., SERZEDELLO, A.; SERRA, G. E.; FAVERI, M. C.; OLIVEIRA, L ; PONEZI, A. N. & SARTORATTO, A. (1996). Effect of lecithin and soy oil the fermentative performance of Saccharomyces uvarum 1904. Journal of the Brazilian Society for Microbiology, 27, 255-262. HARRIS, E. L.V. & ANGAL, S. (1994). Protein purification methods. Ed. IRL PRESS, 6a ed., Oxford, England, 317p. HIGBY, W. K. (1962). A simplified method for determination of some aspects of the carotenoid distribuition in natural and carotene-fortified orange fuice. Journal Food Science, 27 (1), 42-49. INSTITUTO ADOLPHO LUZT. (1985). Métodos químicos e físicos para análise de alimentos. Normas Analíticas v. 1. 3ª ed. São Paulo-SP., 317p. JONES, R. P. (1988). Intracellular etanol accumulation and from yeast and other cell. FEMS Microbiol. Rev., 54, 239258. PEREIRA JR., N. (1999). Bioprocessos industriais. In: Tecnologia Enzimática. eds. BON, E. P. S. & PEREIRA JR., N. Rio de Janeiro-RJ. 1ª ed.. 110p MONTEIRO, C. L. B. (1984). Técnicas de avaliação sensorial. Curitiba-PR, CEPPA, 2a. ed., 101p. RANGANNA, A. (1986). Handbook of Analysis and Quality Control for Fruit and Vegetable Products. 27-30. RIBÉREAU-GAYON, J. & PEYNAUD, E. (1966). Analise e controllo dei vini. Edizioni Agricoli Bologna, 543 p. RIZZON, L. A; ZANUZ, M. C. & MIELE, A. (1998). Evolução da acidez durante a vinificação de uvas tintas de três regiões vitícolas do Rio Grande do Sul. Ciênc. Tecnol. Aliment., 18 (2), 179-183. SOTERO, V. E.; GARCIA, D. & LESSI, E. (1996). Bebida fermentada a partir de pujuayo (Bactris gasipaes H.B.K.) parametros y evolucion. Iquitos-Peru. Folia Amazônica, 8(1), p. 5-18. SOUTHGATE, D. A. T. (1976). Determination of food carbohydrates. Ed. Applied Science Publishers LTD., London-UK., 28-47. "O que é Biossegurança?" Associação Nacional de Biossegurança - ANBio (Repete Fotolito pág. 49 última edição) Biotecnologia Ciência & Desenvolvimento 55 FUNDAÇÃO GIACOMETTI (Repete Fotolito pág. 60 última edição) 56 Biotecnologia Ciência & Desenvolvimento EMBRAPA (Repete Fotolito 3º capa última edição) Biotecnologia Ciência & Desenvolvimento 57 MONSANTO (Repete Fotolito 4º capa última edição)
Baixar