☰
Explorar
Assinar em
Inscrever-se
Envio
×
Baixar
Sem categoria
Olivia Morgan Lapenta - Universidade Presbiteriana Mackenzie
Prática Mental
Slide 1 - Faculdade de Motricidade Humana
Diapositive 1
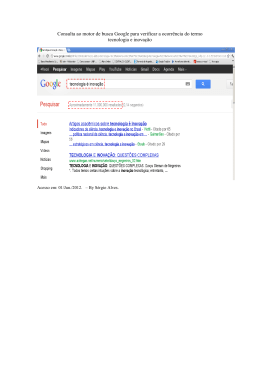
Consulta ao motor de busca Google para verificar a ocorrência do
CONTROLE SOCIAL
navioemmamaersk
Nauta Solar
a importância da educação física escolar no desenvolvimento
A INFLUÊNCIA DA PRÁTICA SISTEMÁTICA DE ATIVIDADE FÍSICA