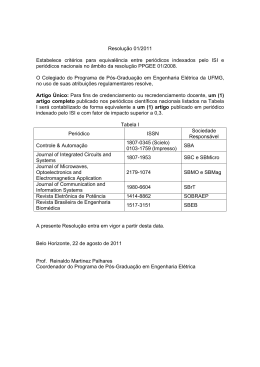
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical Programa de Pós-graduação Stricto Senso Tese de Doutorado Tecnologia de melhoramento de produção de Kappaphycus alvarezii (Doty) Doty ex P.C. Silva. Rafael Rodrigues Loureiro Rio de Janeiro 2013 Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical Programa de Pós-graduação Stricto Senso Tecnologia de melhoramento de produção de Kappaphycus alvarezii (Doty) Doty ex P.C. Silva Rafael Rodrigues Loureiro Tese apresentada ao Programa de Pós-Graduação em Botânica, Escola Nacional de Botânica Tropical, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Doutor em Botânica. Orientadora: Renata Perpetuo Reis Rio de Janeiro 2013 ii Tecnologia de melhoramento de produção Kappaphycus alvarezii (Doty) Doty ex P.C. Silva de Rafael Rodrigues Loureiro Tese submetida ao corpo docente da Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro - JBRJ, como parte dos requisitos necessários para a obtenção do grau de Doutor. Aprovada por: Prof.a Dr.a Renata Perpetuo Reis (Orientadora) _________________________________ Prof.a Dr.a Leila Hayashi _________________________________ Prof.a Dr.a Nair Sumie Yokoya __________________________________ Prof.a Dr.a Valéria Laneuville Teixeira ___________________________________ Prof.a Dr.a Yocie Yoneshigue-Valentin ___________________________________ em 22/02/ 2013 Rio de Janeiro 2013 iii Loureiro, Rafael Rodrigues. L892t Tecnologia de melhoramento de produção de Kappaphycus alvarezii (Doty) Doty ex P.C. Silva / Rafael Rodrigues Loureiro. – Rio de Janeiro, 2013. XIII, 79f. : il. Tese (doutorado) – Instituto de Pesquisas Jardim Botânico do Rio de Janeiro/Escola Nacional de Botânica Tropical, 2013. Orientadora: Renata Perpetuo Reis. Bibliografia. 1.Algicultura. 2.Kappaphycus alvarezii. 3.Tecnologia de melhoramento. I. Título. II. Escola Nacional de Botânica Tropical. CDD 589.3 . iv “Oxalá adquiras - queres alcançá-las! - as virtudes do burrico!: humilde, duro para o trabalho e perseverante, teimoso!, fiel, seguríssimo no seu passo, forte e - se tiver bom dono - agradecido e obediente” S. Josemaria Escrivá v AGRADECIMENTOS Gostaria de manifestar meus sinceros agradecimentos a todas as pessoas que de forma direta ou indireta me ajudaram a realizar este trabalho e foram responsáveis pelo final de tão longa jornada: À minha orientadora Dra Renata Perpetuo Reis, pelos ensinamentos divididos durante longos anos de amizade e por sempre exigir o melhor de seus alunos. Ao Dr. Alan T. Critchley, por durante esses quatro anos ter agido como um coorientador honorário incentivando-me sempre a excelência. À Dra Cristina A. G. Nassar, por ter sido a primeira a acreditar em meu potencial e por continuar a fazê-lo. A todos os funcionários da Escola Nacional de Botânica Tropical e do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. Aos colaboradores: Dr. Daniel Robledo, Dra Gloria Batista Veja, Dr. Jonas Collén, Dra Lynn Cornish, Miguel Sepulveda e Dr. Raúl Ernesto Rincones pelas informações sobre cultivos, bibliografias e críticas feitas ao longo deste trabalho. Aos heróis sem medalha: Ana Carolina Calheiros, Beatriz Castelar, Flávia Berrogain, Gian Berardinelli, Hanna Brum, Henrique Geromel Góes, Íris Menezes, Guillermo Ivanov, Pedro Mattos, Rafael “Garrafa” Marroig e tantos outros; por tornarem esse trabalho possível e por me ensinarem muito durante os anos de convivência no LALGIC. Aos meus pais, pelo incentivo e por sempre questionarem minha profissão, instigando minha teimosia em “Quixoter” pelo caminho profissional que escolhi. Ao meu irmão, pelas conversas, risadas e amor com que me presenteia nos momentos bons e ruins. À minha amada esposa Anne, por ser uma mulher fantástica a qual me força, diariamente, tentar ser um homem melhor. – To my beloved wife Anne, for being an amazing woman, compelling me daily to try to be a better man. vi RESUMO Apesar do sucesso da indústria do cultivo de espécies de Eucheumatoides (Kappaphycus spp. Doty e Eucheuma spp. J. Agardh), esta sofre inúmeras dificuldades, não só devido à infestação por epífitas (goose bumps), mas também por doenças causadas pelo sintoma conhecido como ice-ice. Os benefícios do uso de Acadian Marine Plant Extract Powder (AMPEP), obtido a partir da macroalga marrom Ascophyllum nodosum Mont., no melhoramento de cultivos agrícolas motivou esse trabalho no intuito de, não somente, propor uma nova tecnologia de cultivo para Kappaphycus alvarezii (Doty) Doty ex. P. C. Silva, através do uso do AMPEP, no incremento de biomassa, rendimento de carragenana e mitigação do estresse causado por agentes abióticos ou epífitos, mas também o entendimento dos processos envolvidos em tais respostas. Após testes realizados in vitro, no qual foi estabelecido um protocolo de uso do AMPEP em mudas de K.alvarezii (20g.L-1 e 1h de exposição) e confirmados in situ, a eficiência do extrato comercial de A. nodosum, as quais validouo as seguintes hipóteses:maior taxa de crescimento diário quando expostas ao AMPEP e incremento no rendimento de carragenana. Neste experimento foi verificado que apesar de menor força e viscosidade do gel, estes estiveram dentro da faixa considerada comercial. Além destes resultados, o provável processo responsável pelaa ação do AMPEP em K.alvarezii foi proposto como um “efeito vacina”, no qual o extrato comercial ilicita as defesas do próprio K.alvarezii contra agentes externos, sendo eles abióticos (i.e. temperaturas de cultivo mais baixas do que o ótimo considerado para a espécie) ou bióticos (i.e eliminação de epífitas não cortiçadas e de ramos jovens não cortiçados de filamentosas cortiçadas). No final deste estudo foi comprovada, portanto, a eficiência do extrato em pó comercial da alga marrom A. nodosum (AMPEP) como uma técnica de incremento produtivo para a algicultura de K.alvarezii e também em protocolos de translocação de mudas, para evitar a introdução de alguns organismos acompanhantes, que podem ser potenciais invasores, além de aumentar a resistência da alga perante o estresse do transporte e aclimatação em um possível novo ambiente. vii ABSTRACT Despite the success of the industry of the cultivation of Eucheumatoid (Kappaphycus spp. Doty and Eucheuma spp. J. Agardh) it suffers from many difficulties, not only because of the infestation of epiphytes (goose bumps), but also from diseases caused by the condition known as ice-ice. The benefits of using Acadian Marine Plant Extract Powder (AMPEP), obtained from the brown seaweed Ascophyllum nodosum Mont. on agricultural crops has motivated this work, in order to not only, propose a new technology of cultivation to Kappaphycus alvarezii (Doty) Doty ex. P. C. Silva, through the use of AMPEP, incrementing its biomass, carrageenan yield and mitigation of stress caused by abiotic agents or epiphytes, but also on the understanding of the processes involved in these responses. After tested in vitro, where a protocol for the use of AMPEP with K.alvarezii seedlings was established (20g.L-1 and 1h exposure) and in situ the efficiency of the commercial extract of A. nodosum was confirmed validating the hypothesis: of higher daily growth rate when exposed to AMPEP as well as an increased carrageenan yield without changing its rheological properties. Moreover, the possible processes responsible for the interaction between AMPEP and K.alvarezii were also proposed as a "vaccine effect", where the commercial extract elicits K.alvarezii own defenses against external agents, such as abiotic factors (growth temperatures lower than the optimum considered for the species) or biotic factors (elimination of noncorticated epiphytes and young branches of filamentous macroalgae that were yet noncorticated). At the end of this study, we confirmed the efficiency of the commercial powdered extract of the brown algae A. nodosum (AMPEP) as a new technique to increase the production of K. alvarezii, in translocation protocols of seedlings to prevent the introduction of some escort organisms, that can which can be a potential invading species and to increase the resistance of the algae to the stress of transportation and acclimatization in a possible new environment. viii SUMÁRIO 1. INTRODUÇÃO .......................................................................................................... 1 1.1. HISTÓRICO DA UTILIZAÇÃO DE MACROALGAS ......................................................... 1 1.2. INDÚSTRIA NOS DIAS ATUAIS ................................................................................... 2 1.2.1. Agar ................................................................................................................ 4 1.2.2. Alginatos ......................................................................................................... 4 1.2.3 Carragenana.................................................................................................... 5 1.3. CULTIVO DE KAPPAPHYCUS ALVAREZII (DOTY) DOTY EX P.C.SILVA ......................... 7 1.3.1. Cultivos em ilhas do Pacífico ......................................................................... 9 1.3.2 Cultivos em países africanos ........................................................................... 9 1.3.3. Cultivos em países asiáticos ..................................................................... 10 1.3.4 Cultivos em países americanos (norte, central e sul) ..................................... 11 1.3.4.1. Brasil .......................................................................................................11 1.3.4.2 Colômbia ................................................................................................ 14 1.3.4.3 Equador................................................................................................... 14 1.3.4.4 México .................................................................................................... 15 1.3.4.5 Panamá ................................................................................................... 15 1.3.4.6 Santa Lúcia, Saint Martin e Granadinas ................................................. 16 1.3.4.7 Venezuela ................................................................................................ 16 1.4. TECNOLOGIAS DE MELHORAMENTO PRODUTIVO NA ALGICULTURA DE K. ALVAREZII .................................................................................................................................... 17 1.5. O USO DE ASCOPHYLLUM NODOSUM E PERSPECTIVAS NA MARICULTURA ................ 18 2. OBJETIVO................................................................................................................ 21 3. HIPÓTESES ............................................................................................................. 21 4. CAPÍTULOS ............................................................................................................. 21 4.1 CAPÍTULO I ............................................................................................................ 21 4.2 CAPÍTULO II ........................................................................................................... 22 4.3 CAPÍTULO III ......................................................................................................... 22 4.4 CAPÍTULO IV ......................................................................................................... 22 CAPÍTULO I: CULTIVO IN VITRO DE TRÊS VARIANTES (VERDE, VERMELHA E MARROM) DE KAPPAPHYCUS ALVAREZII (RHODOPHYTA, ARESCHOUGIACEACE) EXPOSTAS AO EXTRATO COMERCIAL DA ALGA MARROM ASCOPHYLLUM NODOSUM (FUCACEAE, OCHROPHYTA). ....... 23 1- INTRODUÇÃO ........................................................................................................... 24 2- MATERIAIS E MÉTODOS ........................................................................................... 25 3- RESULTADOS ........................................................................................................... 26 4- DISCUSSÃO .............................................................................................................. 27 5- REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................... 28 CAPÍTULO II: O EFEITO DE “VACINA” EM KAPPAPHYCUS ALVAREZII (DOTY) DOTY EX. P.C. SILVA PROMOVIDO PELO EXTRATO EM PÓ DA ALGA MARROM ASCOPHYLLUM NODOSUM (LINNAEUS) LE JOLIS (AMPEP). ...................................................................................................................... 31 1 - INTRODUÇÃO .......................................................................................................... 32 2 -MATERIAIS E MÉTODOS ........................................................................................... 33 Condições laboratoriais e aclimatação .................................................................. 33 Desenho experiemental........................................................................................... 34 Crescimento de K. alvarezii e redimento de carragenana...................................... 35 ix Análise estatística dos dados .................................................................................. 35 3- Resultados .......................................................................................................... 36 4- Discussão ........................................................................................................... 37 CAPÍTULO III: EFEITO DO EXTRATO COMERCIAL DA ALGA MARROM ASCOPHYLLUM NODOSUM NA PRODUÇÃO DE BIOMASSA DE KAPPAPHYCUS ALVAREZII (DOTY) DOTY EX PC SILVA E NO SEU RENDIMENTO E A QUALIDADE DO GEL DE CARRAGENANA CULTIVADA NO BRASIL .................................................................................................................. 45 1. INTRODUÇÃO ........................................................................................................... 46 2. MATERIAIS E MÉTODOS ........................................................................................... 47 Área de estudo e desenho experimental .................................................................. 47 Taxa de crescimento diário de K. alvarezii............................................................. 48 Rendimento da carragenana semirefinada de K. alvarezii .................................... 48 Reologia de K. alvarezii ......................................................................................... 48 Análise estatística dos dados .................................................................................. 49 4- DISCUSSÃO .............................................................................................................. 50 5. CONCLUSÃO ............................................................................................................ 53 6- REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................... 53 CAPÍTULO IV: EFEITO DO EXTRATO COMERCIAL DA ALGA MARROM ASCOPHYLLUM NODOSUM MONT. EM CULTIVO DE KAPPAPHYCUS ALVAREZII (DOTY) DOTY EX P.C. SILVA SUBMETIDO À TEMPERATURAS LETAIS .......................................................................................................................... 58 1. INTRODUÇÃO ........................................................................................................... 59 2. MATERIAL E MÉTODOS ............................................................................................ 60 Condições laboratoriais ......................................................................................... 60 Desenho experiemental........................................................................................... 60 Crescimento, rendimento e qualidade da carragenana de K. alvarezii ................. 60 Análise estatística ................................................................................................... 61 3. RESULTADOS ............................................................................................................ 62 4. DISCUSSÃO .............................................................................................................. 63 5- CONCLUSÃO ............................................................................................................ 66 6. REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................... 66 7. CONCLUSÕES FINAIS .......................................................................................... 70 8. REFERÊNCIAS BIBLIOGRÁFICAS GERAIS ................................................... 71 x LISTA DE TABELAS Tabela 1. Valores da indústria de hidrocolóides nos últimos 10 anos. Adaptada de Bixler & Porse (2011). ................................................................................................................. 3 Tabela 2 Distribuição geográfica da produção de agar. Adaptada de Bixler & Porse (2011). ............................................................................................................................... 4 Tabela 3 Distribuição geográfica da produção de alginato em 10 anos. PGA=propylene glycol alginate. Adaptada de Bixler & Porse (2011). ....................................................... 5 Tabela 4 Distribuição geográfica da produção de carragenana em 10 anos, onde álcoll – CR (carragenana refinada); PES (carragenana semi-refinada para consumo humano); SRC (carragenana semi-refinada para consumo animal). Adaptada de Bixler & Porse (2011). ............................................................................................................................... 7 Tabela 5. Uma década de produção e preços de K. alvarezii “cottonii” nas Filipinas e Indonésia. Adaptada de Bixler & Porse (2011). ............................................................. 10 Tabela 6. Cultivos de Kappaphycus alvarezii nas Américas e Caribe (adaptada em colaboração com Hurtado et al. – Kappaphycus Cultivation: Problems and Impacts of Acadian Marine Plant Extract Powder – em preparo). ................................................... 17 Tabela 7. Média±desvio padrão da taxa de crescimento diário e do rendimento de carragenana de Kappaphycus alvarezii cultivado em diferentes tratamentos após 14 dias in vitro. Tratamentos são representados por (KC) = K. alvarezii; (KA) = K. alvarezii banhado em solução de AMPEP; (KO) = K. alvarezii submetido a induzir pulso oxidativo; (KAO) = K. alvarezii banhado em solução de AMPEP previamente induzido a produzir pulso oxidativo; (KE) = Epífita amarrada em K. alvarezii; (KAE) = Epífita amarrada em K. alvarezii previamente banhada em solução de AMPEP; (KOE) = Epífita amarrada em K. alvarezii induzido ao pulso oxidativo (KAOE) = Epífita amarrada em K. alvarezii banhada em solução de AMPEP previamente induzido ao pulso oxidativo. As letras indicam a aparência de K. alvarezii; K. alvarezii (A) = amostras saudáveis sem branqueamento; (B) = amostras sem cor característica, porém sem branqueamento e (C) = amostras com branqueamento. Asteriscos indicam a aparência de Polysiphonia xi subtilissima (*) = amostras saudáveis; (**) = amostras com as porções apicais (não corticadas) branqueadas e (***) = amostras totalmente branqueadas. ........................... 37 Tabela 8 Resultados de taxa de crescimento diario (TCD), redimento de carragenana (RC), força do gel (FG) e viscosidade do gel (VG) de amostras de K. alvarezii (n=12) submetidas a dois tratamentos (controle e AMPEP) em dois periodos de cultivo (20 e 40 dias). ............................................................................................................................... 50 xii LISTA DE FIGURAS Figura 1. Aspecto de uma muda de Kappaphycus alvarezii proveniente de cultivo comercial em Paraty Mirin, RJ. Foto R.G. Marroig. A- Corte transversal do talo; B- Talo tetraspórico e C- Aspecto geral do cistocarpo. Fonte: FAO ............................................. 8 Figura 2. Região entre as latitudes ±10o na qual existe o potencial mundial para o cultivo de K. alvarezii. Retirado de Hayashi et al. (2010). .............................................. 9 Figura 3. Desenho de balsas flutuantes com (a) a técnica rede tubular e (b) e com a técnica tie-tie. Fonte: Góes & Reis, 2011. Foto com a dimensão do módulo da balsa de de cultivo. Foto: Renata P. Reis. ..................................................................................... 13 Figura 4. Detalhe de um banco natural de Ascophyllum nodosum em Halifax, Canadá. Foto: R.R.Loureiro. ........................................................................................................ 19 Figura 5. Taxa de crescimento de três variantes de Kappaphycus alvarezii submetidas à diferentes concentrações do extrato de Ascophyllum nodosum. Símbolos - quadrado representa a mediana; retângulo, os quartis, superior e inferior; barras, a projeção dos non-outliers e círculo, os outliers. .................................................................................. 27 Figura 6. (A) Taxa de crescimento diário, (B) redimento de carragenana, (C) força do gel e (D) viscosidade do gel de amostras de K. alvarezii previamente banhadas em solução de AMPEP (TA) ou não reatadas (TC) (n=8) e submetidas a três temperaturas (T1 =16±1oC; T2 =19±1oC e T3 = 22±1oC). .................................................................. 63 xiii 1. Introdução 1.1. Histórico da utilização de macroalgas A utilização de macroalgas para consumo humano tem registros escritos desde o século 4o D.C. no Japão e 6 D.C. na China. Porém, acredita-se que esta prática seja muito mais antiga, pelos registros de tentativas de cultivo, utilizando métodos simples, como a disposição de hastes de bambu próximas aos bancos naturais. A explotação de macroalgas pode datar de muito antes, uma vez que na região da Serra do Bussaco (Portugal), existem vestígios desta prática em cavernas. O documento de medicina conhecido como o Tratado Zheng Hi, documento Chinês do século 6 D.C., descreve o manejo e cultivo de macroalgas para fins medicinais. Neste, são detalhados: a qualidade do bambu que será utilizado, os númerosos riscos em cada haste (com o intuito de deixar a superfície mais rugosa para captar mais esporos), as luas ideais para cultivo (fluxo das marés e esporulação) e que as estruturas de cultivo deveriam ser colocadas próximas aos bancos naturais (Craigie 2011). Nas Ilhas Britânicas e na Escandinávia, espécies nativas também foram utilizadas, pelas populações locais, durante séculos para inúmeras finalidades. Em um poema escrito em 523 D.C., atribuído a São Columbo foi relatado que os monges de Iona coletavam dulse (Palmaria palmata (Linnaues) Weber & Mohr) nas rochas, que eram fervidas e, posteriormente, misturadas no mingau de aveia. Na Edda, coletânea de sagas viking, escrita no século IX, foi narrada a história de um herói ferido em combate, que foi tratado com uma “poção mágica” feita a partir de algas marinhas. A porção foi confeccionada por uma bruxa que vivia em uma cabana perto do mar, que colheu e ferveu as “plantas da água” para aplicar o elixir no ferimento. Provavelmente, o efeito foi devido à alta concentração de iodo contida nas algas pardas, comuns em regiões de água fria (Celb 2000). Nos séculos XVII e XVIII, novas utilizações para macroalgas foram descobertas, o que deu início ao processo de produção industrial conhecido atualmente. No final do século XVII, indústrias de extração e beneficiamento de iodo e compostos de potássio (Potash), a partir de espécies dos gêneros Fucus Linnaues, Laminaria J.V. Lamouroux e Ascophyllum Stackhouse foram instaladas pelo Império Britânico no arquipélago formado pelas Ilhas do Oeste e Orkney, com a intensão de quebrar o monopólio espanhol. Este produzia um composto chamado barilla, rico em iodo e potássio, que era usado na indústria de vidraria, fabricação de sabão e lavagem e 1 branqueamento de tecidos e, assim, se livrar do bloqueio imposto pelas guerras napoleônicas. Contudo, a indústria britânica teve seu declínio após a descoberta de minas de compostos de potássio na região de Stassfurd, Alemanha e de cristais de iodo no Chile no início do século XIX (Craigie 2011). Após a primeira guerra mundial, um novo grupo de produtos químicos derivados de macroalgas cresceu em interesse comercial, os hidrocolóides. Em 1881, o cientista escocês E.C. Stanford trabalhou com extratos de Laminaria sp. que aumentaram o interesse pelo uso dos hidrocolóides, principalmente, devido a sua capacidade gelatinizante, espessante e emulsificante. Estes extratos eram utilizados em rações militares e em produtos médicos. Contudo, tais produtos já eram utilizados de maneira impirica e tradicional por inúmeras comunidades. Anos mais tarde, em 1949, com a descoberta de Kathleen Drew-Baker da fase conchocelis do ciclo de vida de Porphyra sp. foi possível estabelecer a técnica de cultivo desta espécie, que alavancou a indústria de Porphyra e sua comercialização mundial (Celb 2000). 1.2. Indústria nos dias atuais O consumo de macroalgas é mundial. Indústrias associadas aos cultivos são responsáveis por um arcabouço de produtos que gera valores anuais de US$ 4,5 a 5 bilhões de dólares por ano com foco em agar, carragenana e alginatos. Além disso, compostos biologicamente ativos são vendidos como suplemento alimentar, a alga é usada no consumo direto, ou no uso indireto. Corantes e flavorizantes vem ganhando espaço no mercado. Dados recentes mostram o aumento no consumo e na produção em países da America Latina e Caribe (FAO 2010). O cultivo de algas marinhas se expandiu rapidamente incentivado pela crescente demanda da indústria a qual, inicialmente, contava apenas com a explotação de bancos naturais. Esta demanda motivou a busca de tecnologias para o melhoramento da produção que resultassem em maneiras mais efetivas para o incremento de biomassa e de indivíduos mais resistentes (Zemke-White & Ohno 1999). Cultivos comerciais estão presentes em 35 países, distribuídos em ambos os hemisférios, e são responsáveis por 90% da produção mundial de macroalgas para a indústria (Lüning & Pang 2003; FAO 2010; Bixler & Porse, 2011). A estrutura industrial de macroalgas início na década de 30 com pequenos produtores. A partir da década de 60, as companhias de maior porte, como a Algin Corp e Marine Colloids/FMC, se interessaram em incentivar a produção de algas em larga 2 escala e a investir mais nesta indústria. Com o passar do tempo o inverso ocorreu em países como as Filipinas e a Malásia, onde pequenos produtores se organizaram em pequenas cooperativas e originaram as grandes corporações (e.g. Shemberg Corp., TBK e Marcel Trading Corp.) e atrairam a atenção de empresas ocidentais. Empresas como a Cargill, a CPKelko e FMC Corporation atualmente compram o material bruto de empresas locais (alga seca ou carragenana semirefinada) ou instalam suas sedes em locais de compra de matéria prima, a fim de reduzir os custos em transporte, produção e mão de obra (Zemke-White & Ohno 1999; Bixler & Porse 2011; Bindu & Levine 2011). Entre os maiores produtores mundiais, destacam-se os países asiáticos. A China é a maior produtora de algas para consumo direto, com uma produção de 7,5 milhões de toneladas fresca.ano-1 de seu total de produção de 11,092 milhões no ano de 2010 (FAO 2010; Bixler & Porse 2011). A maior parte da produção está voltada para o cultivo de kombu [Saccharina japonica (Areschoug) C.E.Lane, C.Mayes, Druehl & G.W.Saunders], cultivada em centenas de hectares em sistemas de cordas suspensas (Tseng, 2001; McHugh 2003). A República da Coréia produz anualmente cerca de 500.000 toneladas fresca.ano -1 com destaque para os cultivos de Porphyra sp. e wakame [Undaria pinnatifida (Harvey) Suringar]. O Japão é o segundo maior produtor de algas para consumo direto, com 700.000 toneladas fresca.ano-1 e o nori (Porphyra spp.) é responsável por 65% do total desta produção. Dentre as algas cultivadas para consumo direto, o nori é o mais valorizado, podendo atingir US$ 16.000 por tonelada seca, dependendo da origem e da qualidade do produto, quando comparado ao wakame US$ 6.900 por toneladas secas e ao kombu US$ 2.800 toneladas secas (FAO 2010) (Tab. 1). Tabela 1. Valores da indústria de hidrocolóides nos últimos 10 anos. Adaptada de Bixler & Porse (2011). Hidrocolóide 1999 (US$ milhões) 2009 (US$ milhões) Agar 128 173 Alginato 225 318 Carragenana 291 527 Total 644 1018 3 1.2.1. Agar Dois gêneros, Gracilaria sp. Greville e Gelidium sp. J.V. Lamouroux, são as principais fontes de matéria-prima para indústria de agar no mundo (McHugh 2003; Bixler & Porse 2011). Contudo, apenas Gracilaria sp. obteve sucesso em cultivos no Chile e na Indonésia. Atualmente, Gelidium sp. é preferencialmente utilizado na fabricação de géis para culturas bacteriológicas, sendo explotado em bancos naturais de países como a França, Indonésia, República da Coréia, México, Marrocos, Portugal e Espanha (Bixler & Porse, 2011). No Chile, bancos naturais de Gracilariopsis sp. E.Y. Dawsom também são explotados em menor quantidade, assim como na Índia, onde Gelidiella sp. Feldmann & G.Hamel é retirada de bancos naturais e utilizada exclusivamente no mercado interno (Becker & Rotmann 1990; Jensen 1993). A indústria de agar gera US$ 132 milhões.ano-1 e companhias como Algas Marinas (Chile) e Agarindo Bogatama (Indonésia) produzem cerca de 3.600 toneladas.ano-1, o que representa 58% da produção mundial. Empresas como a Setexam (Marrocos), MSC Co. (República da Coreia), Hispanagar (Espanha) e Huey Shyang Seaweed Industrial Company (China) somam uma produção de 5.500 toneladas ano-1 (57% do mercado mudial). Contudo, apenas 77% da produção mundial (12.500 tonelada seca.ano-1) é utilizada devido à competição entre os produtores (McHugh 2003; Bixler & Porse 201,Tab.2). Tabela 2. Distribuição geográfica da produção de agar. Adaptada de Bixler & Porse (2011). Região Volume (t) 1999 Volume (t) 2009 Europa 1.000 700 900 800 Américas 2.600 2.800 Ásia-Pacífico 3.000 5.300 Produção total 7.500 9.600 Total estimado 9.000 12.500 83 77 África % Utilizado 1.2.2. Alginatos A indústria de alginatos produz ácido algínico e sais alcalinos. Dentre seus 4 derivados, o PGA (propylene glycol alginate) destaca-se em preço de mercado e é o insumo mais barato utilizado na indústria alimentícia (Turan & Neori, 2010; Bixler & Porse, 2011). O cultivo de macroalgas para a produção de alginato é excessivamente custoso, portanto, a indústria utiliza a exploração controlada de bancos naturais e, principalmente, com os excedentes das produções de espécies de Laminaria e Undaria Suringar, as quais são muito utilizadas no consumo direto (McHugh 2003). A partir da na década de 80, a China tornou-se a maior produtora de alginato e quebrou o domínio europeu, norte americano e japonês, com a implementação das indústrias de beneficiamento de alginato, as quais também usam o excedente dos cultivos de algas utilizadas no consumo direto (Vasquez 1995; Tseng 2001; Vauchel et al. 2008). O valor da produção mundial de alginato não é preciso, uma vez que na elaboração de blends, que são misturas de substâncias químicas ao alginato, a quantidade de alginato utilizada, por motivo de segredo industrial, não é descrita e a China ser o maior produtor mundial, que por sua vez não fornece dados precisos de produção (Bixler & Porse, 2011). Estima-se que a produção de alginato a partir da espécie de maior consumo, a Saccharina japônica, seja de 22.000 toneladas ano-1 (FAO 2010). A indústria de alginato norte americana é praticamente inexistente quando comparada com outros produtores mundiais. Na Europa, dois terços da produção de alginato é creditado à FMC Corporation. Os países do Indo-pacífico são a verdadeira força motriz desta indústria (McHugh, 2003; Bixler & Porse 2011, Tab.3). Tabela 3. Distribuição geográfica da produção de alginato em 10 anos. PGA=propylene glycol alginate. Adaptada de Bixler & Porse (2011). Região Alginato (t) 1999 PGA (t) 1999 Alginato (t) 2009 PGA (t) 2009 Europa 12.000 0 10.000 0 Américas 4.000 1.000 1.000 100 Ásia-Pacífico 9.000 1.000 15.500 1.900 Subtotal 25.000 2.000 26.500 2.000 Total estimado 28.000 2.300 35.000 - 89 - 76 - % Utilizado 1.2.3 Carragenana A maior indústria de hidrocolóides é a indústria de carragenana, que gera anualmente US$ 240 milhões e está difundida em todos os continentes (Bindu & Levine 5 2011). Inicialmente, esta indústria dependia da explotação de bancos naturais de Chondrus crispus Stackhouse, alga nativa do Atlântico Norte. No início da década de 70, a indústria da carragenana expandiu-se de maneira exponencial, devido à descoberta do potencial de explotação e cultivo de carragenófitas em locais de clima quente e do avanço de tecnologias de cultivo fomentadas pela pesquisa básica (Bixler & Porse 2011; Bindu & Levine 2011). A carragenana é outro ficocolóide difícil de ser quantificado e caracterizado. Uma das dificuldades de se caracterizar a carragenana é que, apesar de ser conhecido que a carragenana kapa e iota formam géis, sendo a iota mais elástica, e a carragenana lambda é viscosa e de difícil obtenção, muitas vezes esta última é obtida de misturas de carragenanas mu/nu extraídas de espécies de Gigartina sp. Stackhouse e de Sarcothalia sp. Kützing e são feitas misturas de kappa e iota (McHugh 2003, Bixler & Porse 2011). Além disso, de maneira semelhante ao alginato são elaborados blends nos quais a quantidade e o tipo de carragenana não citados (H.G. Góes comunicação pessoal). No passado, a maior produção de carragenana era realizada por indústrias européias e norte americanas e o restante pelos países da Ásia-Pacífico e latinoamericanos. A partir da década de 70, com a permissão do uso da carragenana semirefinada (PES) para consumo humano, aumentou a demanda por carragenana e os locais de sua produção (McHugh 2003; Bixler & Porse 2011, Tab. 4). Anteriormente, nos Estados Unidos, a carragenana semirefinada (SRC) usada em alimentos de animais de estimação era limpa de microorganismos no processo de esterelização e purificada na industrialização para consumo humano e não era exigida a diferenciação entre a carragenana refinada e a semirefinada, enquanto que as agências européias e a FAO/WHO (Food and Agriculture Organization of the United Nations/World Health Organization) Codex Alimentarius exigiam a clasificação de carragenana refinada como E 407 e a semirefinada PES, E 407a. A partir da liberação da carragenana semirefinada (PES) para consumo humano, a indústria indo-Pacífica e a chinesa cresceram (McHugh 2003; Bixler & Porse 2011). 6 Tabela 4. Distribuição geográfica da produção de carragenana em 10 anos, onde álcoll – CR (carragenana refinada); PES (carragenana semi-refinada para consumo humano); SRC (carragenana semi-refinada para consumo animal). Adaptada de Bixler & Porse (2011). Região (ano) Álcool (t) gel (t) PES (t) SRC (t) Total (t) 1999 Europa 7.700 2.500 200 - 10.400 Américas 4.800 2.000 - - 6.800 Ásia-Pacífico 1.000 3.000 8.000 11.000 23.000 - 1.000 1.000 - 2.000 Produção total 13.500 8.500 9.200 11.000 42.000 Total estimado 15.200 11.000 12.000 13.000 55.200 88 77 77 85 76 Europa 6.000 1.000 100 - 7.100 Américas 4.500 3.500 1.400 - 9.400 Ásia-Pacífico 1.000 4.000 16.000 5.000 26.000 - 4.500 3.000 - 7.500 Produção total 11.500 13.000 20.500 5.000 50.000 Total estimado 13.500 16.500 27.000 8.000 65.000 85 78 76 65 76 China %Utilizado 2009 China %Utilizado 1.3. Cultivo de Kappaphycus alvarezii (Doty) Doty ex P.C.Silva Indivíduos do gênero Kappaphycus Doty podem atingir mais de 100 cm de altura. Seu talo principal é multiaxial podendo conter uma medula central composta por células isodiamétricas. Contudo, algumas espécies apresentam estrutura de talo do tipo pseudoparenquimatosa formando uma mistura de células grandes e pequenas variando em forma e quantidade (Fig.1) (Areces 1995) 7 Figura 1. Aspecto de uma muda de Kappaphycus alvarezii proveniente de cultivo comercial em Paraty Mirin, RJ. Foto R.G. Marroig. A- Corte transversal do talo; B- Talo tetraspórico e C- Aspecto geral do cistocarpo. Fonte: FAO Estágios iniciais de reprodução do gênero ainda não foram totalmente descritos, porém assemelham-se ao gênero Eucheuma, do qual faziam parte até 1987, quando Doty segregou o gênero Kappaphycus pela presença de kappa carragenana. O cistocarpo maduro tem a tendência de formar-se no talo principal (Areces 1995; Guiry & Guiry 2011). A espécie tipo é oriunda do arquipélago de Sulu, localizado no mar de Sulu, sudoeste das Filipinas. Devido à sua importância para a maricultura, o gênero foi introduzido em vários países e nomeada em honra ao biólogo pesquisador filipino Vicente Alvarez. Populações nativas podem ser encontradas no Pacífico oeste e central, no leste africano, na Indonésia, na Malásia, na Ilha Hainan, nas Filipinas e na Micronésia (Guiry & Guiry 2011). Registros dos primeiros cultivos de K. alvarezii datam de 1960, na região sul de Mindanao, Filipinas e informam a utilização de variantes derivadas de indivíduos coletados em bancos naturais (Bindu & Levine, 2011). Após quatro décadas, os cultivos de K. alvarezii e de Eucheuma denticulatum (N.L.Burman) F.S.Collins & Hervey representavam 88% da produção de kappa (K. alvarezii) e iota (E. denticulatum) carragenana, o que corresponde a 120.000 toneladas secas.ano-1. Os principais países produtores são as Filipinas, Indonésia e Tanzânia (Ask et al. 2001; Hayashi et al. 2010; 8 Bindu & Levine 2011). Contudo, cultivos de K. alvarezii são encontrados em mais de 20 países localizados entre as latitudes de ±10o (Fig.2), a qual possui as condições abióticas ideais para o cultivo da espécie (Hayashi et al. 2010). Figura 2. Região entre as latitudes ±10o na qual existe o potencial mundial para o cultivo de K. alvarezii. Retirado de Hayashi et al. (2010). 1.3.1. Cultivos em ilhas do Pacífico Cultivos comerciais de K. alvarezii começaram a ser difundidos na região, principalmente após o sucesso dos cultivos nas Filipinas, em meados da década de 70, indicado como uma alternativa de geração de recursos e postos de trabalho para populações costeiras e ao desenvolvimento econômico da região (Pickering 2006). Pickering & Forbes (2002) apontaram o crescimento do cultivo de K. alvarezii como o responsável pela consolidação da algicultura em países como Fiji e Nova Guiné. As pesquisas básicas associadas à práticas locais, geraram manuais práticos de assistência aos maricultores nativos (Teitelbaum 2003) e workshops instrutivos organizados pelos governos, a fim de solucionar problemas como a herbívora excessiva, macroalgas epífitas e o ice-ice, sintoma observado pelo branqueamento do talo e enfraquecimento do indivíduo (Bindu & Levine 2011) 1.3.2 Cultivos em países africanos Os primeiros cultivos de K. alvarezii registrados em países africanos ocorreram em meados dos anos 80, associados aos cultivos de E. denticulatum, na Tanzânia e mais tardiamente na costa leste de Zanzibar e nas Ilhas Pemba expandindo-se na década de 9 90 (Ask et al. 2001). O método utilizado, diferente dos long-lines usados nas Filipinas, foi composto por cercados retangulares. Mtolera (2003) fez uma associação entre a baixa produção do método com a cobertura de gramíneas marinhas que promoveram a limitação da captação de recursos pelas algas (e.g. luz e nutrientes) com consequente diminuição da produção. 1.3.3. Cultivos em países asiáticos As Filipinas mantém a maior concentração de refinarias de carragenana da Ásia, e compra matéria prima de outros países produtores asiáticos, a Indonésia, a Malásia, o Japão, o Vietnã, a China e a Índia (McHugh 2003; Bindu & Levine 2011). K. alvarezii é responsável por 80% das exportações de macroalgas das Filipinas. A alga é vendida úmida ou seca, sendo a última mais valorizada no mercado externo. A alga úmida (87%) é consumida nos países asiáticos (Hurtado 2003; Hayashi et al. 2010). A Indonésia é a principal produtora mundial K.alvarezii, após quebrar o domínio filipino que durou mais de 35 anos (Tab.5). Isto ocorreu, principalmente, por perda de produção ocasionada por problemas decorrentes de mudanças climáticas, epifitismo e ice-ice, assim como políticas governamentais desfavoráveis aos pequenos produtores (Bindu & Levine 2011). Tabela 5. Uma década de produção e preços de K. alvarezii “cottonii” nas Filipinas e Indonésia. Adaptada de Bixler & Porse (2011). Ano Produção Filipina (t seca) Produção Indonésia (t seca) Produção total de cottonii (t seca) Preço Filipino (US$ t-1 seca) Preço Indonésio ( US$ t-1 seca) 2000 113.000 27.000 140.000 710 650 2001 97.000 28.000 125.000 575 525 2002 91.000 30.000 121.000 420 535 2003 90.000 44.000 134.000 770 615 2004 97.000 49.000 146.000 850 750 2005 89.000 56.000 145.000 774 650 2006 84.000 68.000 142.000 748 617 2007 81.000 74.000 155.000 947 811 2008 73.000 79.000 152.000 2.342 2.166 2009 61.000 85.000 146.000 1.280 1.208 10 Além do crescimento na Indonésia no cenário mundial do cultivo de K. alvarezii, outros países, como a Índia, que tradicionalmente não usavam o cultivo de macroalgas como uma atividade comercial, passaram a se destacar como produtores, com promissores programas geridos pelos governos locais, que vislumbraram no cultivo de macroalgas um meio de desenvolvimento econômico sustentável, além de gerar novos postos de emprego para populações de baixa renda (Bindu 2006; 2007; 2009; Bindu & Levine 2011). 1.3.4 Cultivos em países americanos (norte, central e sul) Ask & Azanza (2002) citam a incipiência de cultivos comerciais de K. alvarezii nas américas, que apresentam potencial de produção para atingir o patamar dos grandes produtores mundiais. No entanto, a maior preocupação da introdução nesses países é o potencial de risco ambiental. Algumas complicações foram relatadas na introdução de K. alvarezii para fins comerciais no Havaí (Rodgers & Cox 1999; Conklin & Smith 2005), com relatos posteriores na Venezuela (Barrios 2005) e na Índia (Chandrasekaran et al. 2008). Contudo, outros pesquisadores comentaram que a invasão desta espécie é restrita à locais que sofreram ações antropogênicas (Pickering et al. 2007). Além disso, a falta de cultura e tradição no cultivo de macroalgas marinhas na região e poucos incentivos governamentais deixa para a academia a tarefa de gerar alternativas biotecnológicas e estudos de riscos ambientais da introdução para fazer o cultivo Eucheumatoides nas Américas e no Caribe tornar-se sustentável. 1.3.4.1. Brasil Tradicionalmente as algas são colhidas por mulheres, jovens ou pescadores em bancos naturais na região Nordeste. As macroalgas são uma fonte de renda alternativa para as comunidades costeiras que as vendem como matéria-prima para a indústria de carragenana. Hypnea musciformis (Wulfen) J.V. Lamouroux é a carragenófita explorada em bancos naturais, como matéria prima para a indústria de carragenana brasileira, entretanto não supre a demanda nacional. Devido à falta de manejo de colheita, os bancos naturais estão se esgotando (Castelar et al. 2009). Atualmente, a espécie exótica K. alvarezii é a principal macroalga cultivada comercialmente no Brasil. O seu sucesso é explicado por sua simples técnica de manejo, ou seja, através de crescimento vegetativo a partir da fragmentação do talo, a resiliência da espécie a fatores abióticos e as suas elevadas taxas de crescimento (Góes & Reis 11 2011; Pellizari & Reis 2011). A introdução de K. alvarezii no Brasil ocorreu na baía de Ubatuba, SP, por Edison José de Paula e seu grupo de pesquisa da Universidade de São Paulo. Foi utilizado um clone do Japão, originário das Filipinas. Após uma quarentena de 10 meses, mudas foram introduzidas no mar (Paula et al. 2002; Reis et al. 2007). Também, foram introduzidas mudas de Kappaphycus striatus (F. Schmitz) Doty ex P.C. Silva, que foram retiradas do mar por produzirem tetrásporos viáveis (Bulboa et al. 2007; Reis et al. 2007). A primeira algicultura no Brasil foi implantada na baía da Ilha Grande, RJ, por Miguel Sepúlveda, em 1998, após um período de quarentena em tanques, em SC, com mudas trazidas da Venezuela (Castelar et al. 2009). Em 2004, outro cultivo foi instalado na baía de Sepetiba, RJ, pela Ondas Biomar Cultivo de Algas Marinhas Ltda. Esta empresa possuiu o maior cultivo de K.alvarezii na América Latina, com mais de 20 hectares. No entanto, em 2008, esta empresa finalizou suas atividades, porcausa de altos custos operacionais. O que se seguiu, foram as implantações de pequenos cultivos comerciais espalhados no sul da baía de Sepetiba (Góes & Reis 2011). Atualmente, a maioria dos cultivos comerciais está localizada na baía da Ilha Grande, no sul do estado do Rio de Janeiro e pertence a empresas privadas sem qualquer ligação com o governo. Na legislação brasileira para a liberação do cultivo desta espécie foram criadas duas instruções normativas pelo IBAMA/MMA. A primeira foi a Instrução Normativa no165, de 17 de julho de 2007, permitiu a implantação de cultivos comerciais que tivessem requisição para implantação enviada à Secretaria de Aquicultura e Pesca SEAP, atual Ministério de Pesca e Aquicultura, até 29 de novembro de 2005 e o empreendedor deveria apresentar após um ano um relatório sobre os riscos ambientais em suas fazendas. Através da parceria entre o Instituto de Pesquisas Jardim Botânico do Rio de Janeiro e a empresa Ondas Biomar Cultivo de Algas Ltda. foi iniciado um estudo que avaliou o potencial invasor das fazendas de K. alvarezii nas baías de Sepetiba e Ilha Grande (Reis et al. 2007). Foi constatada a ausência de recrutamento de esporos, a falta de estrutura reprodutiva em mudas e a ausência de recrutamento ou o estabelecimento de mudas fora dos sítios de cultivo, que comprovou ausência de risco ambiental (Castelar et al. 2009). Uma vez que estes estudos foram iniciados em 2006, foi enviado ao IBAMA, um relatório técnico com base neste estudo e em outros desenvolvidos no estado de São Paulo, que subsidiaram a elaboração da Instrução Normativa no185, 22 de 12 julho de 2008, que permitiu o cultivo comercial de K. alvarezii na costa sudeste do Brasil entre a baía de Sepetiba, ao sul do estado do Rio de Janeiro e a Ilhabela, no norte do estado de São Paulo (Reis, em preparo). No Brasil, as técnicas de cultivo utilizadas para K. alvarezii são compostas de balsas flutuantes feitas com tubos de PVC (Tab. 6; Fig. 3), diferindo das técnicas mais utilizadas no cultivo da espécie em outros países, como os sistemas off-bottom (Hayashi et al. 2010; Góes & Reis 2011). Este fato se deve principalmente pela maior turbidez da água e fundo lodoso nos locais de cultivo, que tornou as técnicas usuais de cultivo utilizadas em países asiaticos impossíveis de serem estabelecidas nestes locais (Reis, em preparo). Góes & Reis (2011) proposurem o uso de uma nova técnica de cultivo, a de rede tubular em balsas flutuantes. Nesta técnica, as balsas flutuantes de PVC são mantidas, no entanto a tradicional técnica tie-tie é substituída por redes tubulares recheadas com mudas de K. alvarezii. Nesta técnica não ocorreu diferença na taxa de crescimento diário das mudas e no rendimento e na qualidade da carragenana, quando comparada com a técnica tie-tie, entretanto, o tempo de manejo foi reduzido e foi mais fácil e o custo foi menor. Ressaltaram inclusive que, o maior custo em uma produção é com o pagamento de pessoal, que com o uso desta técnica é reduzido. Figura 3. Desenho de balsas flutuantes com (a) a técnica rede tubular e (b) e com a técnica tie-tie. Fonte: Góes & Reis 2011. Foto com a dimensão do módulo da balsa de cultivo. Foto: Renata P. Reis. Marroig & Reis (2011) afirmaram que nos cultivos de K. alvarezii na costa sul 13 fluminense, apesar das balsas flutuantes serem incrustadas por vários organismos, a incrustação por organismos sem carbonato de cálcio não afetaram a produção. As balsas incrustadas por animais com conchas podem ser afundadas com a massa destes organismos. Por este motivo, incentiva-se a necessidade de limpeza constante destas estruturas de cultivo. Foi constatado que, não foram observadas mudas estabelecidas nos costões rochosos localizados próximos aos cultivos. No litoral sul, no estado de Santa Catarina, a viabilidade do cultivo desta alga é restrita nos meses do inverno, quando a temperatura da água pode dimuir até 16 oC, que é letal para esta espécie (Hayashi et al. 2010). 1.3.4.2 Colômbia K. alvarezii foi introduzida na Colômbia, em 1991 (Tab. 6), através de um programa financiado pela FAO, orientado pelo Prof. Bula Germnán Meyer. Mudas cubanas, originadas da Venezuela, tiveram um processo de quarentena e aclimatização em tanques fechados (Raúl Ernesto Rincones, comunicação pessoal). Atualmente, programas pilotos oriundos do programa original da FAO, vem sendo financiados pela organização não governamental Fundação Terrazul, em Cabo de La Vela na península de La Guarija. Trabalham com comunidades nativas americanas, como os Wayuu, em fazendas experimentais, e utilizam balsas flutuantes com redes tubulares em tanques, anteriormente utilizados na criação de camarão. No entanto, tais tentativas de criar uma cultura semifechada para diminuir os riscos de invasões biológicas foram ineficazes, uma vez que, K. alvarezii foi relatada em 2004, ao longo da costa de La Guarija devido ao descaso no manejo de descarte de mudas (Rincones 2006; Hurtado et al. em preparo). Diversas ocorrências de ice-ice, seguidas da infestação das mudas por epífitas foram observadas. Contudo, o que parece ser a maior preocupação é a herbívoria, que causa perdas significativas na maioria dos cultivos (Miguel Sepúlveda e Raúl Ernesto Rincones, comunicação pessoal, Hurtado et al. em preparo). 1.3.4.3 Equador Introduzida por Miguel Sepúlveda (consultor de algas marinhas do Brasil), as mudas oriundas do Brasil foram trazidas para o Equador para fazer parte de uma empresa chamada Ecualgas SA, que recebeu incentivo do governo local através do Ministério da Pesca e Aquicultura (MAGAP). O cultivo ainda está restrito em tanques 14 de criação de camarão (camaroeiras), ativos ou desativados, onde a aclimatação de K. alvarezii e seu processo de quarentena ocorreu com mudas cultivadas no método offbottom (Tab. 6). Nestes tanques, K. alvarezii apresentou o sintoma de ice-ice e infestação por epífitas, sem solução aparente (Miguel Sepulveda, comunicação pessoal; Hurtado et al. em preparo). 1.3.4.4 México Muñoz et al. (2005) propôs o cultivo de K. alvarezii na península de Yucatán após a obtenção de resultados promissores. Esta tentativa não teve apoio do governo e foi voltada para um programa de cultivo sustentável de K. alvarezii realizado por pescadores e comunidades costeiras por meio de cooperativas sociais. As mudas utilizadas foram originárias do Brasil e foram colocadas em quarentena, seguindo as regras da Norma Oficial Mexicana (NOM). Não há relatos de complicações nos cultivos, tanto com herbívoros quanto com a infestação por epífitas. Tal fato, foi atribuído às características abióticas locais, aliadas à técnica de cultivo offbottom (Tab. 6). A principal causa da perda de colheitas foi observada pelo sintoma de ice-ice, o qual foi atribuído a alta temperatura da água, acima de 30°C, um problema que pode ser resolvido por meio da utilização de balsas flutuantes (Daniel Robledo, comunicação pessoal; Hurtado et al. em preparo). 1.3.4.5 Panamá Juntamente com o Brasil e os países do Caribe, Santa Lúcia, Saint Martin e Granadinas, o Panamá é um dos poucos que mantém fazendas comerciais de K. alvarezii nas américas e exporta sua produção para a Europa e Ásia (Glória Batista Vega e Raúl Ernesto Rincones, pessoal comunicação; Hurtado et al., em preparo). Mudas de K.alvarezii da Venezuela foram originalmente introduzidas no Panamá pelo Smithsonian Tropical Research Institute, sem procedimentos de quarentena para fins experimentais, na província de Colón, um lugar que hoje detém o único cultivo comercial no Panamá, e que tem apoio de empresas privadas, como a Seafarms Bansistemas S.A. e é supervisionado pelo governo local, através da Autoridade de Recursos Aquáticos do Panamá (ARAP). Além disso, na província de Bocas del Toro, estudos experimentais estão sendo realizados com balsas flutuantes e tie-tie, sistema também utilizado na península de Colón (Batista 2006; 2009; Hurtado et al. em preparo). 15 Nos primeiros anos dos cultivos na península Colón (Tab. 6), as infestações de epífitas foram uma grande preocupação, por influenciar as taxas de crescimento de K. alvarezii e causar perdas na colheita (Batista 2009). Atualmente, não existem relatos de epífitas, no entanto, a ocorrência de ice-ice e de herbívoros é a preocupação dos algicultores locais (Raúl Ernesto Rincones, comunicação pessoal; Hurtado et al., em preparo). 1.3.4.6 Santa Lúcia, Saint Martin e Granadinas Sem o apoio do governo e contando com empresas privadas e organizações não governamentais, tais como: Ashton Multiusage Cooperative AMCO e Projeto Sustentável Granadinas, esses países do Caribe são os únicos a manterem cultivos funcionais de K.alvarezii na região, com mudas oriundas da Venezuela (Tab. 6). Essas mudas foram introduzidas em 2011, sem procedimento de quarentena (Raúl Ernesto Rincones e Miguel Sepulveda, comunicação pessoal; Hurtado et al. em preparo). Através de várias técnicas de cultivo disponíveis, que variaram de acordo com o sítio de cultivo (i.e. balsas flutuantes e sistema off-bottom, ambos com tie-tie), não houve relatos de infestações por epífitas. Contudo, complicações ocasionadas por herbívoros e por ice-ice também foram observadas nestes países, bem como na maioria dos países das Américas (Raúl Ernesto Rincones, comunicação pessoal; Hurtado et al. em preparo). 1.3.4.7 Venezuela K. alvarezii foi legalmente introduzida na Venezuela com mudas das Filipinas, em 1996, após aclimatação de seis semanas e processo de quarentena. Teve apoio do governo, através do Instituto Nacional de Pesquisas Agropecuária (INIA) e da Fundação do Desenvolvimento Rural, Inovação e Capacitação da Venezuela, estudos experimentais foram realizados com K.alvarezii na península de Araya e Ilha Cubagua (Rincones & Rubio 1999; Smith & Rincones 2006; Hurtado et al. em preparo). Esses estudos deram as informações necessárias para o governo permitir o cultivo de espécies exóticas ao longo da península de Araya (estado Sucre), através da utilização de balsas flutuantes com redes tubulares ou tie-tie, dependendo do local de cultivo. Redes tubulares são mais utilizadas em locais com maior movimentação de água para evitar a perda de mudas (Tab. 6). Até o momento não foram relatadas complicações causadas pela infestação de epífitas (Hurtado et al. em preparo). 16 Tabela 6. Cultivos de Kappaphycus alvarezii nas Américas e Caribe (adaptada em colaboração com Hurtado et al. – Kappaphycus Cultivation: Problems and Impacts of Acadian Marine Plant Extract Powder – em preparo). País Cultivo Técnica de cultivo Origem da muda Processo de quarentena /inspeção Complicações Brasil Comercial Balsa flutuante; rede tubular. Japão e Venezuela Sim/Sim Herbívoros. Colômbia Experimental Balsa flutuante; rede tubular/tie-tie. Caribe; Cuba Sim (mudas Cubanas) Herbívoros; Ice-Ice; Epífitas. Equador Experimental off-bottom; camaroeiras. Brasil Sim/Sim Ice-Ice; Epífitas. México Experimental off-bottom; tie-tie. Brasil Sim/Não Ice-Ice. Panamá Experimental; (Bocas del Toro) comercial (Colón) Balsas flutuantes; tietie. Venezuela Não Herbívoros; Ice-Ice. Santa Lucia, San Martin & Granadinas. Comercial Balsas flutunates; tie-tie e off-bottom. Venezuela Não Herbívoros; Ice-Ice. Venezuela Experimental Balsas flutuantes; redes tubulares; tietie Filipinas Sim/Sim Herbívoros; Ice-Ice. 1.4. Tecnologias de melhoramento produtivo na algicultura de K. alvarezii Além das alternativas indicadas ao combate do ice-ice e do goose bump, muitos esforços têm sido realizados, especialmente por cientistas e empresas de processamento, para combater os problemas de declínio da produção em cultivo e aumentar o número de mudas geradas. Um desses esforços foi o desenvolvimento dos sistemas de cultivo em balsas flutuantes em contraste com o método off-bottom tradicionalmente utilizado nos países que são os maiores produtores de K.alvarezii (Góes & Reis 2011). Considerando que o método off-bottom é trabalhado durantes as marés baixas, em locais mais profundos, onde não existe esta possibilidade, a técnica com linhas flutuantes, pode ser usada e de maneira constante, não dependendo da maré baixa. No entanto, é necessário o uso de barcos e canoas, para a manutenção e colheita que encarece a 17 produção. Esta técnica pode ser uma solução para as comunidades costeiras pobres, que na verdade, constituem a maioria dos produtores (Msuya et al. 2007). Outros esforços foram focados na utilização de extratos de algas para mitigar a proliferação de epífitas e aumentar a produtividade da alga. Experimentos utilizaram o extrato comercial da alga marrom Ascophyllum nodosum (Linnaeus) Le Jolis, o AMPEP (Acadian Marine Plant Extract), utilizado em horticultura e agricultura, que testaram a eficiência deste extrato através de sua aplicação em Kappaphycus alvarezii (Hurtado et al. 2009; Loureiro et al. 2010; Yunque et al. 2011; Borlongan et al. 2011). Estes autores observaram a eficiência do uso do AMPEP para reduzir a incidência de algumas epífitas e para aumentar a taxa de crescimento diário de K. alvarezii. Em outro estudo, o uso de AMPEP em cultura de tecidos de K. alvarezii var. tambalang obtec=veve sucesso na micropropagação de mudas para plantio (Hurtado et al. 2008). Em 2005, na Tanzânia, os compradores das algas, em resposta ao baixo retorno de seus investimentos com perda de K. alvarezii, com sintmas de ice ice, introduziram K. striatus (F.Schmitz) Doty ex P.C.Silva em colaboração com o governo, devido a restrições a importação e cautela na introdução de novas espécies. Neste local, esta alga é conhecida como kikarafuu pela semelhança do seu talo com o cravo, que é chamada de karafuu em Swahili, a língua local da Tanzânia. As taxas de crescimento de K. striatus foram menores do que as de K. alvarezii. No entanto, são mais resistentes e não apresentam os sintomas de ice-ice (Hayashi et al. 2010) 1.5. O uso de Ascophyllum nodosum e perspectivas na maricultura As macroalgas e as microalgas são utilizadas na agricultura como adubo e agentes condicionadores de solo (Chapman & Chapman 1980; Guiry & Blunden, 1981; Metting et al. 1988; 1990; Hong et al. 2007; Craigie, 2010). Os benefícios observados no crescimento, vigor e rendimento das culturas vegetais foram atribuídos ao fornecimento por nutrientes, melhora na textura do solo e maior retenção de água. 18 O desenvolvimento de produtos liquefeitos ou em pó de algas, que são vendidos como extratos, e seus resultados promissores pelo uso de pequenas quantidades do produto por hectare (<15 L.ha-1) induziram o exame mais detalhado dos componentes das algas (nutrientes), conhecidos por promover o crescimento de plantas. A alga marrom Ascophyllum nodosum (Fig.4) destaca-se por ser a espécie com o maior número de artigos científicos sobre bioensaios de produção agrícola (Craigie 2011). Figura 4. Detalhe de um banco natural de Ascophyllum nodosum em Halifax, Canadá. Foto: R.R. Loureiro. Dominante na zona de entremarés nas costas do Atlântico Norte, onde a temperatura da água não ultrapassa 270C, na região da Nova Escócia, A. nodosum produz safra de 71,3 tonelada umida.ano-1. As frondes são penduculares e fixam-se nas rochas. No apressório basal crescem vários filóides e geralmente a planta regenera novas frondes da base, quando uma destas é danificada, o que é de interesse para a indústria desde que a colheita seja cuidadosamente administrada para garantir a viabilidade deste recurso renovável (Ugarte et al. 2010). As frondes são perenes e capazes de sobreviver mais de 20 anos e a porção basal pode sobreviver por um século ou mais (Xu et al. 2008; Guiry & Guiry 2011). Produtos derivados de A. nodosum têm sido usados na agricultura como agentes de crescimento, melhorando a capacidade de absorção de nutrientes, através do aumento de tecido radicular e proteção contra infecções causadas por fungos (Jayaraj et al. 2008; 19 Khan et al. 2008; 2009; Parys et al. 2009; Fan, 2010; Jayaraman et al. 2011). Partindo do pressuposto desta melhora, estudos foram realizados sobre o efeito do uso do extrato em pó obtido de A. nodosum, comercializado como AMPEP (Acadian Marine Plant Extract), no rendimento produtivo de K. alvarezii. Os resultados foram promissores em cultura de tecidos, no aumento de taxas de crescimento diário e na mitigação na ocorrência de epífitas (Hurtado et al. 2009; Loureiro et al. 2010; Yunque et al. 2011; Borlongan et al. 2011). Em relação aos mecanismos de ação do AMPEP em K. alvarezii, Khan et al. (2009) citaram a presença de elicitores de citoquinina em outro produto fabricado a partir de A. nodosum macerado, oriundo do leste do Canadá. Esta descoberta indicou que estes elicitores podem atuar no crescimento de mudas de K. alvarezii quando expostas ao AMPEP. De fato, é conhecido que as algas vermelhas apresentam um sistema de defesa complexo e eficaz contra patógenos e bioincrustantes, sendo capazes de perceber a presença de epibiontes e agentes patogênicos e os seus efeitos negativos e que respondem de maneira específica para cada ameaça (Weinberger & Potin 2010). A maioria destas informações foi obtida através de ensaios realizados com espécies de Gracilaria Greville e com Chondus crispus Stackhouse, que indicaram a presença de caminhos moleculares de sinalização em algas vermelhas (Weinberger et al. 1999; 2001; Weinberger 2007). Outra substância presente em Rhodophyta, são os compostos orgânicos halogenados (Kladi et al. 2004) e seu possível papel nos mecanismos de defesa. Entretanto, pouco foi investigado sobre tais compostos, que podem estar relacionados aos processos de sinalização (Weinberger & Potin 2010). Os compostos orgânicos halogenados voláteis são produzidos por algas vermelhas após um período de estresses ambientais, como a forte movimentação da água e as variações na temperatura e na salinidade, e estão relacionados ao estresse oxidativo (Mtolera et al. 1996; Pedersen et al. 1996a; Potin et al. 2002; Gachon et al. 2010). Weinberger et al. (1999) observaram que a resposta da halogenação, causada por um pulso oxidativo provocado por uma fonte de estresse externa, pode desencadear a liberação de peróxido de hidrogênio que causa o aumento na atividade de bromação em Gracilaria spp.. Este fato pode provocar uma reação que culmina em sintomas semelhantes ao de ice-ice, como o branqueamento do talo (Pedersen et al. 1996b). 20 2. Objetivo Examinar o efeito do extrato em pó da alga marrom A.nodosum (AMPEP) em K.alvarezii, quanto à sua taxa de crescimento diário, ao seu rendimento de carragenana, no controle de epífitas, em suas propriedades reológicas e na sua resistência à variação de temperatura e avaliar os processos envolvidos nas respostas obtidas. 3. Hipóteses 1. Amostras de K. alvarezii previamente expostas ao extrato AMPEP antes de cultivadas in vitro e in situ apresentarão maior crescimento quando comparadas as amostras não expostas ao AMPEP; 2. Amostras de K. alvarezii previamente exposta ao extrato AMPEP in vitro e in situ apresentarão maior rendimento de carragenana quando comparadas as amostras não expostas ao AMPEP; 3. Amostras de K. alvarezii previamente expostas ao extrato AMPEP apresentarão variações em suas propriedades reológicas (viscosidade e dureza do gel) quando comparadas as amostras não expostas ao AMPEP; 4. O uso do AMPEP apresenta efeito de vacina em K. alvarezii que estimula defesas naturais dos indivíduos expostos e 5. O uso do AMPEP em amostras de K. alvarezii in vitro promove efeito mitigatório aos possíveis danos causados por temperatura baixas consideradas como letais para esta espécie. 4. Capítulos A tese foi dividida em quatro capítulos, cada um representa uma etapa das análises sobre o efeito do AMPEP no beneficiamento do cultivo de K. alvarezii e corresponde a um artigo científico aceito ou submetido. As conclusões de cada artigo estão reunidas na conclusão geral da tese. 4.1 Capítulo I Título: Cultivo in vitro de três variantes (verde, vermelha e marrom) de Kappaphycus alvarezii (Rhodophyta, Areschougiaceace) expostas ao extrato comercial da alga marrom Ascophyllum nodosum (Fucaceae, Ochrophyta). Objetivo: Testar a eficiência do extrato de A. nodoum no aumento do crescimento 21 diário e no combate a epífitas em três variantes de Kappaphycus alvarezii em experimentos in vitro. 4.2 Capítulo II Título: Efeito “vacina” em Kappaphycus alvarezii (Doty) Doty ex. P.C. Silva promovido pelo extrato em pó da alga marrom Ascophyllum nodosum (Linnaeus) Le Jolis (AMPEP). Objetivo: Sugerir os processos envolvidos no beneficiamento do uso do extrato de A. nodosum em cultivos de K. alvarezii relacionandos ao aumento do crescimento e combate ao epifitismo . 4.3 Capítulo III Título: Efeito do extrato comercial da alga marrom Ascophyllum nodosum na produção de biomassa de Kappaphycus alvarezii (Doty) Doty ex P.C. Silva e no seu rendimento e qualidade do gel de carragenana cultivada no Brasil Objetivo: Verificar o efeito do uso do AMPEP em cultivos de K. alvarezii no mar, através da taxa de crescimento diário, rendimento e qualidade de carragenana. 4.4 Capítulo IV Título: Efeito do extrato comercial da alga marrom Ascophyllum nodosum Mont. em cultivo de Kappaphycus alvarezii (Doty) Doty ex P.C. Silva submetido a temperaturas letais. Objetivo: Analisar a taxa de crescimento diário, o rendimento e a qualidade do gel obtido em mudas de K.alvarezii in vitro, cultivadas em três temperaturas (16±1; 19±1 e 22±1 oC) quando expostas previamente à uma solução do extrato em pó da alga marrom A. nodosum. 22 Capítulo I: Cultivo in vitro de três variantes (verde, vermelha e marrom) de Kappaphycus alvarezii (Rhodophyta, Areschougiaceace) expostas ao extrato comercial da alga marrom Ascophyllum nodosum (Fucaceae, Ochrophyta). Rafael R. Loureiro; Renata P. Reis e Alan T. Critchley. Artigo publicado em no Journal of Applied Phycology, 22: 101-104. 23 1- Introdução O sucesso mundial da indústria de carragenana está baseado em espécies de Eucheuma J. Agardh e Kappaphycus Doty referenciadas na literatura como Eucheumatoids (Ask & Azanza 2002). Um dos maiores problemas enfrentados pelos cultivos comerciais é a doença conhecida como ice-ice, responsável por causar o branqueamento (Vairappan 2006) e fragmentação do talo (Largo et al. 1997). Vairappan et al. (2008) comenta que ice-ice foi primeiramente reportada por Trono (1974) nas Filipinas e Uyenco et al. (2008) relacionadam à condições ambientais não favoráveis para o cultivo nos sítios onde foram cultivadas. Largo et al. (1995a; 1995b; 1997; 1998) relataram a presença de bactérias dos complexos Vibrio–Aeromonas e Cytophaga–Flavobacterium nos indivíduos afetados e relacionaram à presença de bactérias nos indivíduos afetados por ice-ice, previamente submetidos a estresses de alta irradiância e temperatura, assim como, à baixa salinidade. Outro problema crítico é o goose bumps causado pela alga filamentosa Neosiphonia sayateri (Hariot) M.S. Ki et I.K. Lee (Vairappan et al. 2008), antes identificada como Polysiphonia sp. Mont, em trabalhos pretéritos sobre o mesmo problema, no qual a epífita se estabelece na camada cortical do hospedeiro e ocasiona rupturas no talo, propiciando um ambiente ideal para a infecção por bactérias (Vairappan 2006; Huratado et al. 2006; Vairappan et al. 2008). Ambos ice-ice e goose bumps são responsáveis por decréscimos significativos na produção e qualidade de carragenana destas espécies em países como as Filipinas, Indonésia, Malásia, e Tanzânia (Vairappan et al. 2008). Estudos recentes mostraram que o epifitismo causado por Neosiphonia (tida como Polysiphonia) provocou mais de 70% de perda na produção de Kappaphycus nas Ilhas Calaguas, Filipinas (Hurtado et al. 2006). O método mais utilizado para sanar o problema foi retirar todas as algas do cultivo atingido e descartar os indivíduos afetados ou não, que causou a perda do ciclo de produção e de mudas para futuros cultivos (Hurtado et al. 2005). Algas marinhas vêm sendo utilizadas como fertilizantes naturais por séculos, contudo, seu verdadeiro potencial nunca foi totalmente explorado (Guiry & Blunden 1991). Estudos sobe o uso do extrato da alga marrom Ascophyllum nodosum (Linnaeus) Le Jolis mostraram sua eficácia no aumento da produção (biomassa) e redução de pragas (fungos) em vegetais superiores, consequentemente melhorando a qualidade final 24 do produto (Blunden et al. 1996; Leach et al. 1999). Tais efeitos positivos em vegetais superiores motivaram os objetivos desse trabalho que tem como foco a tentativa de aumentar a taxa de crescimento diário e mitigar a ocorrência de epífitas e doenças em K.alvarezii, através da utilização do extrato comercial de A. nodosum. 2- Materiais e Métodos Três variantes da alga Kappaphycus alvarezii (e.g. morfotipos da cor verde, vermelha e marrom) foram cultivadas in vitro após serem expostas a diferentes concentrações de um extrato alcalino, preparado com a alga marrom A. nodosum. O extrato produzido pela Acadian Seaplants Limited (Dartmouth, Nova Scotia, Canada) é composto por: 3-5% de proteínas/aminoácidos; 1% lipídios; 12-18% acido algínico; 1215% polímeros contendo fucose; 5-6% manitol e 10-15% de outros carboidratos. As amostras de K. alvarezii utilizadas no experimento, foram obtidas em cultivos localizados na Praia Grande, Ilha de Itacuruçá, norte da baía de Sepetiba, estado do Rio de Janeiro, Brasil (22°57′02″S e 43°54′22″W). O voucher do material utilizado foi incluído no Herbário do Jardim Botânico do Rio de Janeiro (RB 425.507). As variantes foram previamente limpas de epífitas, com o uso de pinças e toalhas de papel, com o intuito de não lesionar o talo, e foram aclimatadas durante 30 dias, em aquários de 5 litros (temperatura da água-22±2°C, irradiância da superfície da água-130±5 μmol fótons.m−2·s−1, fotoperíodo-12 h, salinidade (PSS)-35±2 e pH-8.1±0.3). Para testar o efeito do extrato de A.nodosum nas epífitas, porções apicais de K. alvarezii (controle e testadas) não foram limpas após o período de aclimatação, foram mantidas nas mesmas condições, e suas epífitas, identificadas através de literatura taxonômica apropriada e adotanda a nomenclatura descrita no Algaebase (Guiry & Guiry, 2008). Após o período de aclimatação, porções apicais foram banhadas em diferentes concentrações do extrato de A.nodosum (e.g., 5, 10, 15, 20, 25 e 30 g.L−1 de água do mar) pelo período de 1 h (período escolhido para tornar possível a utilização do extrato em cultivos comerciais). Seis repetições de cada variante de K. alvarezii com 5cm (com 5 g±0.2 cada) foram dispostas em frascos Erlenmeyer com 1 L de água do mar filtrada em membrana de ester celulose (0,45 μm-Millipore), com aeração constante por duas semanas nas mesmas condições de aclimatação. No tratamento controle, as porções apicais não foram expostas ao extrato. O crescimento das porções apicais foi estimado em taxa crescimento diário (TCD), calculado de acordo com a fórmula TCD=100×[ln(Mf /Mo)/t], onde Mo é o massa inicial, Mf o massa final e t o tempo de 25 cultivo (Reis et al. 2006). A normalidade (teste de Shapiro-Wilk) e a homogeneidade (teste de Cochran) dos dados foram inicialmente testadas. A diferença entre as taxas de crescimento diário das variantes de K.alvarezii submetidas a diferentes concentrações do extrato de A. nodosum e seus controles foram analisadas pelo teste de Mann-Whitney, uma vez que os dados não apresentaram homogeneidade. A interação entre as variantes de K. alvarezii e as concentrações do extrato de A. nodosum em relação as suas taxas de crescimento diário foram testadas utilizando o teste de ANOVA bifatorial e, em seguida, o teste posterior de Tukey, a fim de distinguir as diferenças significativas entre as médias. Todas as análises estatísticas foram realizadas no programa Statistica 6.0, da StatSoft®, em usado o intervalo de confiança de 95% (p<0,05). Os dados foram expressos em média±desvio padrão. 3- Resultados Após 14 dias de cultivo em laboratório, as porções apicais de K. alvarezii banhadas no extrato de A. nodosum apresentaram quase o dobro do valor de suas taxas de crescimento diário, quando comparadas aos seus controles (controle, Z=2,88, p<0,001, Fig. 5). Foi bservada uma interação positiva entre as três variantes de K. alvarezii (ANOVA bifatorial, F=4,50, p<0,001) e as diferentes concentrações do extrato de A .nodosum (ANOVA bifatorial, F=219.00, p<0.001). A maior taxa de crescimento de K. alvarezii obtida durante o período de cultivo, foi observada nas porções apicais banhadas nas concentrações de 15 e 20 g.L-1, seguidas por 25 g.L-1, 30 g.L-1 e os menores valores do crescimento diário nas concentrações de 10 g.L-1, seguida por 5 g.L-1 (ANOVA bifatorial, F=32,71, p<0,001). As variantes verdes e marroms obtiveram os maiores resultados em crescimento diário quando comparadas com a vermelha (Fig. 5). No período de duas semanas Cladophora sp. Kützing e Ulva sp. Linnaeus não foram encontradas nas porções epifitadas de K. alvarezii submetidas às concentrações do extrato (e.g., 5, 10, 15, 20, 25, and 30 g.L−1), contudo, Polysiphonia subtilissima Mont, permaneceu como pequenos tufos ao longo do talo em todas as concentrações testadas. 26 Figura 5. Taxa de crescimento de três variantes de Kappaphycus alvarezii submetidas à diferentes concentrações do extrato de Ascophyllum nodosum. Símbolos - quadrado representa a mediana; retângulo, os quartis, superior e inferior; barras, a projeção dos non-outliers e círculo, os outliers. 4- Discussão Robert-Anderson et al. (2006) examinaram o efeito do extrato comercial de Ecklonia maxima (Osbeck) Papenfuss, Kelpak® na alga vermelha Gracilaria gracilis (Stackhouse) Steentoft, Irvine et Farnham em cultivos in vitro e em cultivos comerciais de Ulva lactuca Linnaeus e obtiveram resultados promissores na taxa de crescimento dos indivíduos submetidos ao extrato (controle - 2,8%.dia-1 e tratadas com Kelpak® 6,1%.dia-1). Diferente desse estudo, os experimentos conduzidos com o extrato de E. maxima tiveram o acréscimo de PES (Provasoli Enriched Seawater) uma vez que o Kelpak® apresenta baixa concentração de nutrientes. O crescimento de 5,9%.dia-1 nas amostras de K. alvarezii submetidas ao extrato, quando comparadas ao de 2,4%.dia-1 de seus controles, mostra-se promissor para a utilização dessa tecnologia em cultivos comerciais, uma vez que o maior crescimento em menor tempo reduz a exposição das algas a agentes que possam ser danosos ao 27 produção final. As altas taxas de crescimento obtidas pelas amostras banhadas em solução de A. nodosum podem estar relacionadas à presença de hormônios vegetais (e.g. citocinina) em K. alvarezii, no qual o extrato age como um indutor de tal fator, aumentando assim a produção de antioxidantes e promovendo a divisão celular (Salisbury & Ross, 1992). Provavelmente, esse efeito pode promover a resistência de K. alvarezii contra fatores abióticos estressantes como temperatura e irradiância e as porções previamente banhadas ficam menos suscetíveis aos efeitos danosos como o ice-ice. A eliminação das epífitas Ulva e Cladophora nas concentrações de 15 a 25 g.L-1 de A. nodosum, pode estar relacionada ao limite de tolerância de tais espécies as concentrações testadas do extrato, uma vez que em concentrações mais baixas (i.e. 5 a 10 g.L-1) os dois gêneros estavam presentes ao final dos 14 dias de cultivo. P. subtilissima provou ser mais tolerante, por estar presente em concentrações elevadas de 25 e 30 g.L-1. Nessas concentrações, K. alvarezii apresentou menores valores de taxas de crescimento diário quando comparados com a concentração de 15 g.L-1, contudo, estes valores foram maiores do que os do controle, ou seja, os não banhados em A. nodosum. Esses resultados mostram que o extrato de A. nodosum foi eficiente no aumento da taxa de crescimento diário de K. alvarezii e, em cultivos comerciais, também pode ser utilizado na mitigação de epífitas pertencentes à classe Ulvophyceae. 5- Referências bibliográficas Ask, E.I., Azanza, R.V. 2002. Advances in cultivation technology of commercial eucheumatoid species: a review with suggestion for future research. Aquaculture 206: 257–277. Blunden, G., Jenkin, T., Yan-Wen, L. 1996. Enhanced leaf chlorophyll levels in plants treated with seaweed extract. Journal of Applied Phycology 8 (6):535–554. Guiry, M.D., Blunden, G. 1991. Seaweed resources in Europe: uses and potential. John Wiley & Sons Canada, Ltd., pp xi, 432 Guiry, M.D., Guiry, G.M. 2008. AlgaeBase. Worldwide electronic publication, National University of Ireland, Galway. http://www. algaebase.org. Accessado 17 Julho 2008. 28 Hurtado, A.Q., Bleicher-Lhonneur, G., Crichley, A.T. .2005. Kappaphycus‘cottonii’ farming. Integrated Services for the Development of aquaculture and Fisheries, Philippines, p 26. ____________, Crichley, A.T., Bleicher-Lhonneur, G. 2006. Occurrence of Polysiphonia epiphytes in Kappaphycus farms at Calaguas Is., Camarines Norte, Philippines. Journal of Applied Phycology 18:301-306. Largo, D.B., Fukami, F., Nishijima, T., Olino, M. 1995a. Laboratory induced development of the ice-ice disease of the farmed red algae Kappaphycus alvarezii and Eucheuma denticulatum (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 7:539–543. ___________, Fukami, F., Nishijima, T. 1995b. Occasional bacteria promoting ice-ice disease in the carrageenan-producing red algae Kappaphycus alvarezii and Eucheuma denticulatum (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 7: 545–554. ___________, Fukami, K., Adachi, M., Nishijima, T. 1997. Direct enumeration of total bacteria from macroalgae by epifluorescence microscopy asapplied to the fleshy red algae Kappaphycus alvarezii and Gracilaria spp. (Rhodophyta). Journal of Phycology 33: 554–557. ___________, Fukami, M.A.K., Nishijima, T. 1998. Immunofluorescent detection of ice-ice disease-promoting bacterialstrain Vibrio sp. P11 of the farmed macroalga, Kappaphycus alvarezii (Gigartinales, Rhodophyta). Journal of Marine Biotechnology 6: 178–182. Leach, W.L., Plunkett, B.A., Blunden, G. 1999. Reduction of nitrate leaching from soil treated with an Ascophyllum nodosum based soil conditioning agent. Journal of Applied Phycology 11: 593–594. Reis, R.P., Caldeira, A.Q., Miranda, A.P.S.M., Barros-Barreto, M.B. 2006 Potencial para maricultura da carragenófita Hypnea musciformis (Wulfen) J. V. Lamour. 29 (Gigartinales—Rhodophyta) na Ilha da Marambaia, Rio de Janeiro, Brasil. Acta Botanica Brasica 20: 763–769. Robertson-Andersson, D.V., Leitao, D., Bolton, J.J., Anderson, R.J., Njobeni, A., Ruck, K. 2006 Can kelp extract (KELPAK®) be useful in seaweed mariculture? Journal of Applied Phycology 18: 315–321. Salisbury, F.B., Ross, C.W. 1992. Plant physiology. Wadsworth, California, 234 p. Trono, G.C. Jr. 1974. Eucheuma farming in the Philippines. U.P. Natural Science Research Centre, Quezon City Uyenco FR, Saniel LS, Jacinto GS (1981) The ‘iceice’ problem in seaweed farming. Proceedings of the International Seaweed Symposium 10:625-630 Vairappan, C.S. 2006. Seasonal occurrences of epiphytic algae on the commercially cultivated red alga Kappaphycus alvarezii (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 18: 611-617. ____________, Chung, C.S., Hurtado, A.Q., Soya, F.E., Bleicher-Lhonneur, G., Critchley, A. 2008. Distribution and symptoms of epiphyte infection in major carrageenophyte-producing farms. Journal of Applied Phycology 20:477-483. 30 Capítulo II: O efeito de “vacina” em Kappaphycus alvarezii (Doty) Doty ex. P.C. Silva promovido pelo extrato em pó da alga marrom Ascophyllum nodosum (Linnaeus) Le Jolis (AMPEP). Rafael R. Loureiro; Renata P. Reis; Flávia D. Berrogain e Alan T. Critchley. Artigo publicado em 2012, no Journal of Applied Phycology 24: 427-432. 31 1 - Introdução A indústria mundial de carragenana tem como base o cultivo de Eucheuma J. Agardh (para iota carragenana) e Kappaphycus Doty (para Kappa carragenana) sendo estas as principais fontes de renda e emprego para muitas familías em países em desenvolvimento como a Indonésia, Filipinas, Malásia e Tanzania (Pickering 2006; Bixler & Porse 2001). Doenças como o ice-ice, que provoca o branqueamento do talo de Kappaphucys, e o goose bumps causado pelo epifitismo da alga vermelha Neosiphonia apiculata (Hollenberg) Masuda & Kogame, são responsáveis pela perda de mais de 70% da produção em cultivos comerciais de Kappaphycus alvarezii (Doty) Doty ex P.C. Silva (Largo et al. 1995a; 1995b; 1997; 1998; Vairappan 2006; Vairappan et al. 2008). A maneira mais efetiva encontrada para contornar tais complicações é a retirada do cultivo e a não utilização de mudas oriundas de áreas de cultivo infectadas. Contudo, tal prática vem afeta drasticamente a cadeia de fornecimento de novas mudas para cultivo (Hurtado et al. 2006). Embora, ocorreram progressos na tentativa de compreender e gerenciar tais problemas (Goecke et al. 2010; Solid et al. 2010; Vairappan et al. 2010), não existe solução viável para evitar a perda de cultivos inteiros. Extratos de algas marinhas têm sido usados na agricultura como agentes de crescimento, melhorando a capacidade de absorção de nutrientes através do aumento de tecido radicular e de proteção contra infecções causadas por fungos (Jayaraj et al. 2008; Khan et al., 2008; 2009; Fan 2010). Umas das macroalgas mais utilizadas em aplicações agrícolas é a alga marrom Ascophyllum nodosum (Linnaeus) Le Jolis (Parys et al. 2009; Craigie 2011; Jayaraman et al. 2011). Estudos recentes que utilizaram o Acadian Marine Plant Extract (AMPEP), extrato comercial em pó da alga A. nodosum em Kappaphycus alvarezii, apresentaram resultados promissores na melhoria de mudas em cultura de tecidos, no aumento de taxas de crescimento diário e na mitigação na ocorrência de epifitas (Hurtado et al. 2009; Loureiro et al. 2010; Yunque et al. 2011; Borlongan et al. 2011). No entanto, os mecanismos de ação do AMPEP em K. alvarezii ainda não foram discutidos ou testados. Experimentos conduzidos por Khan et al. (2009) citaram a presença de elicitores de citoquinina em outro produto fabricado a partir de Ascophyllum nodosum macerado, oriundo do leste do Canadá. Esta evidência indica o que pode ocorrer quando amostras de K. alvarezii forem expostas ao AMPEP, especialmente no tocante ao aumento das 32 taxas de crescimento diário. É conhecido que algas vermelhas apresentam um complexo e eficaz sistema de defesa contra patógenos e bioincrustações e são capazes de perceber a presença de epibiontes e agentes patogênicos e os seus efeitos negativos e, consequentemente, respondem de maneira específica a cada ameaça (Weinberger & Potin 2010). A maioria destas informações foi obtida através de ensaios realizados com espécies do gênero Gracilaria Greville e de Chondus crispus Stackhouse, que indicaram a presença de caminhos moleculares de sinalização em algas vermelhas (Weinberger et al. 1999; 2001; Weinberger 2007). Outro aspecto presente em Rhodophyta são os compostos orgânicos halogenados (Kladi et al. 2004) e seu possível papel nos mecanismos de defesa, embora pouco tenha sido investigado sobre tais compostos, que podem estar relacionados aos processos de sinalização (Weinberger & Potin, 2010). Os compostos orgânicos halogenados voláteis, até onde se sabe, são produzidos por algas vermelhas após um período de estresse (e.g. fatores ambientais, como a movimentação abrupta e violenta de água e variações em temperatura e salinidade) e estão ligados diretamente ao estresse oxidativo (Mtolera et al. 1996; Pedersen et al. 1996a; Potin et al. 2002; Gachon et al. 2010). Weinberger et al. (1999) observaram que a resposta da halogenação, causada por um pulso oxidativo provocado por uma fonte de estresse externa, pode desencadear a liberação de peróxido de hidrogênio que causa aumento na atividade de bromação em Gracilaria spp. Tal fato pode provocar uma reação que culmina em sintomas semelhantes ao de ice-ice, como o branqueamento do talo (Pedersen et al. 1996b). O presente estudo examina as relações entre os mecanismos de defesa presentes em algas vermelhas e as respostas obtidas em K. alvarezii através da utilização do AMPEP. 2 -Materiais e Métodos Condições laboratoriais e aclimatação Amostras de K. alvarezii foram obidas em um cultivo comercial localizado na Praia Grande, Ilha Itacuruça, ao norte da baía de Sepetiba, Rio de Janeiro, Brasil (22°57′02″S e 43°54′22″W). O voucher do material utilizado foi incluído no Herbário do Jardim Botânico do Rio de Janeiro (RB 425.507). As amostras foram previamente limpas de epífitas com o uso de pinças e toalhas de papel, com o intuito de não lesionar o talo, e aclimatadas durante 30 dias em aquários com 5 litros de ágia do mar e foi 33 utilizado as mesmas condições sugeridas por Loureiro et al. (2010), tais como: temperatura da água - 22±2°C, irradiância da superfície da água - 130±5 μmol photons m−2·s−1, fotoperíodo – 12:12h, salinidade (PSS) - 35±2 e pH-8.1±0.3. Todas as variantes de cor foram tratadas de maneira única, sendo que porções apicais de cada variante foram coletadas em proporções iguais a fim de que todas as variantes fossem representadas em cada frasco de cultivo. Desenho experiemental Após o período de aclimatação, durante 1h, amostras de 5g de K. alvarezii (n=6), obtidas a partir de porções apicais de 5cm, foram banhadas em solução de AMPEP (20 g.L-1 em água do mar filtrada) sugerida por Loureiro et al. (2010) como a concentração ideal para aumentar o crescimento de K. alvarezii, ou submetidas ao tratamento de indução de pulso oxidativo, através de agitação mecânica por haste de metal por 3 minutos, ou por uma combinação de ambos (amostras induzidas a produzir pulso oxidativos e banhadas em AMPEP). Para testar o efeito do AMPEP nas epifitas de K. alvarezii, a alga Polysiphonia subtilissima Mont. foi escolhida devido a sua semelhança morfológica com Neosiphonia spp. M.-S. Kim & I.K, Lee. De acordo com Guiry & Guiry (2011), ambas as espécies têm organização de talo semelhante (e.g., talo formado por regiões medulares e corticais) e são membros da familia Rhodomelaceae. As amostras foram limpas com ajuda de um microscópio estereoscópico e aclimatadas por 30 dias, sob as mesmas condições de K. alvarezii. Após as amostras de K. alvarezii terem sido banhadas em AMPEP e submetidas aos outros tratamentos, cada P. subtilissima (um indivíduo com apressório, com 2±0,2 g coletada no Canal do Jardim de Alah, Rio de Janeiro, RJ, Brasil) foi amarrada com uma linha nas porções apicas de K. alvarezii. Para induzir K. alvarezii a produzir pulso oxidativo como uma resposta de defesa, as amostras de porções apicais foram submetidas ao procedimento descrito por Mtolera et al. (1996) em Eucheuma denticulatum (NL Burman) FS Collins & Hervey de indução de estresse para produzir compostos orgânicos halogenados voláteis. Os tratamentos foram: KC= K. alvarezii; KA= K. alvarezii banhado em solução de AMPEP; KO= K. alvarezii submetido à indução de pulso oxidativo; KAO= K. alvarezii banhado em solução de AMPEP previamente submetido à indução de pulso oxidativo; KE= K. alvarezii com epífita amarrada; KAE= epífita amarrada em K. 34 alvarezii previamente banhada em solução de AMPEP; KOE= epífita amarrada em K. alvarezii previamente submetida à indução de pulso oxidativo; KAOE= epífita amarrada em K. alvarezii previamente submetida à indução de pulso oxidativo e banhada em solução de AMPEP. Cada tratamento foi realizado em Erlenmeyer com 1L de água do mar filtrada e mantido por duas semanas, nas mesmas condições em que as amostras haviam sido aclimatadas. Crescimento de K. alvarezii e redimento de carragenana O crescimento das amostras foi estimado em taxa de crescimento diário (TCD) e calculado de acordo com a fórmula: TCD=100×[ln(Mf /Mo)/t], onde Mo é a massa inicial, Mf a massa final e t o tempo de cultivo (Góes & Reis 2011). Foi utilizado o método para obtenção de carragenana semirefinada. Neste método, as amostras de K. alvarezii são secas em estufa (60 ºC) por 48 horas e pesadas. Após duas horas em solução de hidróxido de potássio em banho maria (80 °C), as amostras foram lavadas em água corrente e colocadas em solução de hipoclorito de sódio, em temperatura ambiente, durante 25 minutos e, posteriormente, lavadas em água corrente por aproximadamente 5 minutos a fim de retirar resíduos. Ao fim deste procedimento, as algas foram secas em estufa (60 oC), pesadas e determinado o percentual de rendimento da carragenana semi-refinada (RC), através da razão entre a massa seca da alga tratada e sua massa seca inicial. O pH de 8,4 foi mantido durante o procedimento (Góes & Reis 2011). O rendimento da carragenana (RC) das amostras de K. alvarezii foi expresso como o percentual de carragenana por amostra de alga seca (Reis et al. 2008; Góes & Reis 2011) obtido pela formula: RC= (Mc/Ma)× 100, onde Mc é o massa seca final da carragenana seca (seca em estufa a 60ºC até obtenção de massa constante) e Ma o massa da alga seca (seca em estufa a 60ºC até obtenção de massa constante). Análise estatística dos dados A normalidade dos dados (teste de Shapiro-Wilks) e a homogeneidade das variâncias (teste de Cochran) foram previamente testadas para o uso de testes de análise de variância paramétricos ou não paramétricos. A diferença da taxa de crescimento diário de K. alvarezii submetida aos diferentes tratamentos foi testada pela análise variância unifatorial (ANOVA) e para distinguir as médias significativamente diferentes foi usado o teste posterior LSD Fisher. A diferença do rendimento de carragenana de K. 35 alvarezii sob diferentes tratamentos foi analisada pela Krushal-Wallis ANOVA. As análises estatísticsa dos dados foram realizadas no programa Statistica 6.0 da Statsoft em um intervalo de confiança de 95% (p<0,05). Os dados foram expressos em média±desvio padrão e foram usadas seis repetições para cada tratamento (n=6). 3- Resultados Depois de duas semanas de experimento, diferenças significativas foram observadas na taxa de crescimento diário de K. alvarezii (p<0,001, Tab.7) e o rendimento carragenana (p<0,001, Tab.7) em cada tratamento. A maior taxa de crescimento diário foi obtida em amostras expostas ao AMPEP (KA e KAE, exceto KAOE). A taxa de crescimento diário das amostras que foram tratadas com AMPEP e também sofreram indução de pulso oxidativo (KAO) não diferiram entre as que não foram submetidos ao AMPEP (KE, p=0,641 e KO, p=0,753). Quando os tratamentos não foram estimulados a produzir um pulso oxidativo (KE) ou, na ausência de Polissiphonia subtilissima (KC) e quando expostos ao AMPEP (KA) houve diferença significativa entre as taxas de crescimento diário das amostras de K. alvarezii (KA>KC>KE). No entanto, quando elas foram estimuladas a produzir o pulso oxidativo (KO e KAO), não diferiram entre si. Os maiores valores de rendimento carragenana de K. alvarezii foram observados em amostras que foram estimuladas a produzir pulso oxidativo, na presença de epífitas ou em sua ausência (KO, KAO, KAE, KOE; KAEO; Tab.7). O rendimento de carragenana foi afetado pela presença de epífitas. Em amostras expostas à epífitas e estimuladas a produzir pulso oxidativo (KOE <KAOE), os valores foram maiores do que as que não estavam na presença de P. subtilissima (KC<KA<KO=KAO). O branqueamento do talo das epífitas foi observado em amostras que foram estimuladas a produzir pulso oxidativo (KOE, KAOE) e P. subtilissima morreu após 14 dias. Amostras de K. alvarezii banhadas em AMPEP e estimuladas a produzir pulso oxidativo (KAO) não apresentaram branqueamento do talo no período de cultivo. No entanto, as amostras as quais as epífitas foram fixadas e não foram estimuladas a produzir pulso oxidativo (KAE) tiveram seus ramos apicais jovens esbranquiçados e morreram após duas semanas. 36 Tabela 7. Média±desvio padrão da taxa de crescimento diário e do rendimento de carragenana de Kappaphycus alvarezii cultivado em diferentes tratamentos após 14 dias in vitro. Tratamentos são representados por (KC) = K. alvarezii; (KA) = K. alvarezii banhado em solução de AMPEP; (KO) = K. alvarezii submetido a induzir pulso oxidativo; (KAO) = K. alvarezii banhado em solução de AMPEP previamente induzido a produzir pulso oxidativo; (KE) = Epífita amarrada em K. alvarezii; (KAE) = Epífita amarrada em K. alvarezii previamente banhada em solução de AMPEP; (KOE) = Epífita amarrada em K. alvarezii induzido ao pulso oxidativo (KAOE) = Epífita amarrada em K. alvarezii banhada em solução de AMPEP previamente induzido ao pulso oxidativo. As letras indicam a aparência de K. alvarezii; K. alvarezii (A) = amostras saudáveis sem branqueamento; (B) = amostras sem cor característica, porém sem branqueamento e (C) = amostras com branqueamento. Asteriscos indicam a aparência de Polysiphonia subtilissima (*) = amostras saudáveis; (**) = amostras com as porções apicais (não corticadas) branqueadas e (***) = amostras totalmente branqueadas. Treatmento Taxa de Rendimento de K. alvarezii P. subtilissima Crescimento Diário carragenana após 14 dias após 14 dias -1 (% dia ) (% PS) KC 3,0±0,43 28,1±2,05 A - KA 5,6±0,23 33,3±1,10 A - KO 2,2±0,71 39,4±1,59 C - KAO 2,0±0,26 38,0±1,50 B - KE 2,3±0,22 30,6±1,69 B * KAE 4,7±0,34 37,0±1,70 A ** KOE 1,6±0,45 42,1±3,51 C *** KAOE 1,9±0,41 46,6±2,30 C *** 4- Discussão A eficiência do uso de AMPEP em K. alvarezii no aumento da taxa de crescimento diário e na prevenção contra o estabelecimento de epífitas foi demonstrado por Hurtado et al. (2009), Loureiro et al. (2010), Yunque at al. (2010) e Borlongan et al. (2011). Resultados semelhantes foram obtidos em todos os tratamentos no qual o AMPEP foi utilizado neste trabalho (KA e KAE) quando estas foram comparadas com as amostras induzidas a produzir pulso oxidativo (KAO e KAOE). No entanto, os 37 mecanismos responsáveis por estas respostas não foi totalmente investigado. É evidente em bioensaios que o AMPEP apresentou respostas positivas quando utlizado em K.alvarezii, mas não está claro como ou por quê. Os resultados apresentados neste trabalho abrem uma discussão sobre alguns dos possíveis mecanismos e também amplia a nossa compreensão sobre as interações funcionais de um extrato de alga marrom com o crescimento de algas vermelhas e sua resistência a epífitas. Com base nos resultados da melhoria na taxa de crescimento de K. alvarezii banhada na solução de AMPEP (KA) e dos valores de rendimento de carragenana em amostras induzidas a produzir pulso oxidativo (KAO, KAO, KOE, e KAOE), sugerimos que o AMPEP pode ter um efeito de vacina em K. alvarezii. Isto é um tanto semelhante a uma função já atribuída ao extrato conhecido como Vacciplant® da alga marrom Laminaria Lamouroux e seus efeitos sobre vegetais superiores (Gachon et al. 2010). A maioria dos resultados obtidos com o Vacciplant® é baseada sobre os efeitos do β-1, 3 glucanos e sua indução à liberação de H2O2 em plantas de tabaco (Klarzynski et al. 2000). Jayaraman et al. (2011) também destaca a capacidade do extrato de elicitar enzimas de defesa, incluindo β-1, 3 glucanase conhecida por estar presente em algas vermelhas com propriedades antifúngicas (Cordeiro et al. 2006). Isto, combinado as atividades elicitoras de hormônios do extrato (principalmente citocinina) discutidas por Reitz & Trumble (1996) e Khan et al. (2009) e a presença de betaínas demonstrada por Mackinnon et al. (2009) e Blunden et al. (2009) em extratos de Ascophyllum, assim como a influência do aumento no teor de clorofila discutido por Whapham et al. (1993), o extrato de A. nodosum pode funcionar como um indutor de estresse que culmina na produção de compostos halogenados voláteis que, como descritos por Mtolera et al. (1996), atuam no processo de defesa da alga. Estes compostos seriam parte do caminho que induz o pulso oxidativo e a produção de peróxido de hidrogênio que acarreta o branqueamento do talo das epífitas, como observado por Pedersen et al. (1996a; 1996b) e por Weinberger et al. (1999) em Gracilaria conferta (Schousboe ex Montagne) Montagne. Também, pode explicar o não estabelecimento ou sobrevivência dos propágulos das epífitas, como descrito por Borlongan et al. (2010). Este processo, também explica o efeito do branqueamento causado pelo pulso oxidativo nas amostras K. alvarezii induzidas a produzi-lo e o efeito do branqueamento de P. subtilissima, bem como a baixa taxa de crescimento diário e alto valor do rendimento de carragenana destas amostras de K. alvarezii quando comparadas com as amostras não induzidas a produzir o pulso oxidativo. 38 A cor do talo de K. alvarezii das amostras expostas à solução de AMPEP (KAO e KAEO) que não sofreu o efeito de branqueanto pode ser explicado pela presença de betaínas e seus efeitos sobre K. alvarezii. As betaínas são conhecidas por aumentar o conteúdo de clorofila, que por sua vez, aumentam a taxa fotossintética (Whapham et al. 1993). Estas, também, podem ser responsáveis por bloquear o efeito de branqueamento produzido pelo estresse causado pela liberação de peróxido de hidrogênio, portanto, não interferindo nas taxas de crescimento de K. alvarezii, como observado nos tratamentos. O aumento das taxas de crescimento de K. alvarezii em amostras expostas a solução de AMPEP (KA e KAE) em relação aos outros que não foram expostos a este extrato (KC, KO, KE, e KOE) tornam-se mais claras quando considerarmos os efeitos da presença de betaínas, como descrito por Mackinnon et al. (2010) e da elicitação da atividade da citocinina promovida pela presença do extrato (Reitz & Trumble 1996; Khan et al. 2009). A citocinina é conhecida por promover a divisão celular (Salisbury & Ross 1992). Yokoya et al. (2010) discutiram a presença de citocininas em Rhodophyta e afirmaram que a complexidade dos perfis hormonais encontrados em macroalgas desempenham um papel fundamental na regulação dos processos fisiológicos das algas vermelhas. A administração do extrato em amostras induzidas ao pulso oxidativo reduziu os efeitos do estresse oxidativo (produção de peróxido de hidrogênio), que pode ser extremamente agressiva para um indivíduo e sua epífita. O branqueamento das porções jovens do talo de P. subtilissima que foram cultivadas simulando epífitas em K.alvarezii banhado em solução de AMPEP (KAE) confirmou que a reação foi evidente, na qual o AMPEP protegeu K. alvarezii dos efeitos do peróxido de hidrogênio. A eficácia do extrato também pode ser confirmada nos talos de K. alvarezii que foram protegidos do clareamento (KAO) quando comparadas com as amostras que não foram banhadas em solução de AMPEP (KO). Com as evidências apresentadas neste trabalho, uma hipótese pode ser construída para começar a entender os caminhos complexos envolvidos nesta nova abordagem do uso de um extrato de uma alga marrom como uma potencial vacina em K. alvarezii, que provoca a ativação de suas defesas naturais contra patógenos e ameniza os efeitos negativos da exposição em longo prazo ao pulso oxidativo (i.e. a exposição ao peróxido de hidrogênio que culmina no branqueamento do talo). 5- Referências bibliográficas Bixler, H.J., Porse, H. 2011. A decade of change in the seaweed hydrocolloids industry. 39 Journal of Applied Phycology 23: 321-335. Blunden, G. Currie, M., Máthé, I., Hohmann, J., Critchley, A. 2009 Variation in betaine yields from marine algal species commonly utilized in the preparation of seaweed extracts used in agriculture. The Phycologist 76: 14. Borlongan, I.A.G., Tibudos, K.R., Yunque, D.A.T., Hurtado, A.Q., Critchley, A.T. 2011. Impact of AMPEP on the growth and occurrence of epiphytic Neosiphonia infestation on two varieties of commercially cultivated Kappaphycus alvarezii grown at different depths in the Philippines. Journal of Applied Phycology 23:615– 621 Cordeiro, R.A., Gomes, M.V., Carvalho, A.F.U., Melo, V.M.M. 2006 Effect of proteins from the red seaweed Hypnea musciformis (Wulfen) Lamouroux on the growth of human pathogen yeasts. Brazilian Archives of Biology and Technowloge 49: 915– 921 Craigie, J.S. 2011. Seaweed extracts stimuli in plant science and agriculture. Journal of Applied Phycology 23: 371 393. Fan, D. 2010. Ascophyllm nodosum extracts improve shelf life and nutritional quality of spinach (Spinacia oleracea L.). Dissertação de mestrado, Dalhousie University Gachon, C.M.M., Sime-Ngando, T., Strittmatter, M., Chambouvet, A., Kim, G.H. 2010. Algal diseases: spotlight on a black box. Trends in Plant Science 15: 633-640. Goecke, F., Labes, A., Wiese, J., Imhoff, J.F. 2010. Chemical interactions between marine macroalgae and bacteria. Marine Ecology Progress Series 409: 267–299. Góes, H.G., Reis, R.P. 2011. An initial comparison of tubular netting versus tie–tie methods of cultivation for Kappaphycus alvarezii (Rhodophyta, Solieriaceae) on the south coast of Rio de Janeiro State, Brazil. Journal of Applied Phycology 23: 607–613. 40 Hurtado, A.Q., Critchley, A.T., Bleicher-Lhonneur, G. 2006. Occurrence of Polysiphonia epiphytes in Kappaphycus farms at Calaguas Island., Camarines Norte, Philippines. Journal of Applied Phycology 18: 301–306. ____________, Yunque, D.A., Tibudos, K., Critchley, A.T. 2009. Use of Acadian marine plant extract powder from Ascophyllum nodosum in tissue culture of Kappaphycus varieties. Journal of Applied Phycology. 21: 633–639. Jayaraj, J., Wan, A., Rahman, M., Punja, Z.K. 2008. Seaweed extract reduces foliar fungal diseases on carrots. Crop. Prot. 27:1360–1366. Jayaraman, J., Norrie, J., Punja, Z.K. 2011. Commercial extract from the brown seaweed Ascophyllum nodosum reduces diseases in greenhouse cucumber. Journal of Applied Phycology 23: 353–361. Khan, W., Palanisamy, R., Hankins, S.D., Critchley, A.T., Smith, D.L., Papadopoulos, Y., Prithiviraj, B. 2008. Ascophyllum nodosum (L.) Le Jolis extract improves root nodulation in alfalfa. Canadian Journal of Plant Science 88: 728–728 ________, Rayirath, U.P., Subramanian, U.P., Jitesh, M.N., Rayorath, P., Hodges, D.M., Critchley, A.T, Craigie, J.S., Norrie, J., Prithiviraj, B. 2009 Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant. Growth Regulators 28: 386–399. Kladi, M., Vagias, C., Roussis, V. 2004. Volatile halogenated metabolites from marine red algae. Phytochemestry 3: 337-366. Klarzynski, O., Plesse, B., Joubert, J.M., Yvin, J.C., Kopp, M., Kloareg, B., Fritig, B. 2000. Linear β-1,3 glucans are elicitors of defense responses in tobacco. Plant Physiology 124: 1027–1037. Largo, D.B., Fukami, F., Nishijima, T. 1995a. Occasional bacteria promoting ice-ice disease in the carrageenan-producing red algae Kappaphycus Eucheuma denticulatum (Solieriaceae, Gigartinales, alvarezii and Rhodophyta). Journal of 41 Applied Phycology 7: 545-554. __________, Olino, M. 1995b. Laboratory-induced development of the ice-ice disease of the farmed red algae Kappaphycus alvarezii and Eucheuma denticulatum (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 7: 539– 543. __________, Fukami, K., Adachi, M., Nishijima, T. 1997 Direct enumeration of total bacteria from macroalgae by epifluorescence microscopy as applied to the fleshy red algae Kappaphycus alvarezii and Gracilaria spp. (Rhodophyta). Journal Phycology 33: 554– 557. ___________, Fukami, M.A.K., Nishijima, T. 1998 Immuno fluorescent detection of ice-ice disease-promoting bacterial strain Vibrio sp. P11 of the farmed macroalga, Kappaphycus alvarezii (Gigartinales, Rhodophyta). Journal of Marine Biotechnologe 6: 178–182. Leach, W.L., Plunkett, B.A., Blunden, G. 1999. Reduction of nitrate leaching from soil treated with an Ascophyllum nodosum based soil conditioning agent. Journal of Applied Phycology 11: 593–594. Loureiro, R.R., Reis, R.P., Critchley, A.T. 2010. In vitro cultivation of three Kappaphycus alvarezii (Rhodophyta, Areschougiaceae) variants (green, red and brown) exposed to a commercial extract of the brown alga Ascophyllum nodosum (Fucaceae, Ochrophyta). Journal of Applied Phycology 22: 101–104. Mackinnon, S.L., Hiltz, D., Ugarte, R., Craft, C.A. 2010 Improved methods of analysis for betaines in Ascophyllum nodosum and its commercial seaweed extracts. Journal of Applied Phycology 22: 489–494. Mtolera, M.S.P., Collén, J., Pedersén, M., Ekdahl, A., Abrahamsson, K., Semesi, A.K. 1996. Stress-induced production of volatile halogenated organic compounds in Eucheuma denticulatum (Rhodophyta) caused by elevated pH and high light intensities. European Journal of Phycology 31: 89–95. 42 Parys, S., Kehraus, S., Pete, R., Kupper, F.C., Glombitza, K.W., Konig, G.W. 2009. Seasonal variation of polyphenolics in Ascophyllum nodossum (Phaeophyceae). European Journal of Phycology 44: 331–338. Pedersén, M., Collén, J., Abrahamsson, K., Ekdahl, A. 1996a. Production of halocarbons from seaweeds: an oxidative stress reaction? Marine Science 60: 257– 263. ________________________________________, Mtolera, M., Semesi, A., Garcia Reina, G. 1996b. The ice-ice disease and oxidative stress of marine algae. In: Björk M, Semesi AK, Pedersén M, Bergman B(eds) Current trends in marine botanical research in the East African region. Ord & Vetande AB, Uppsala, Sweden, pp 11– 24. Pickering, T. 2006. Advances in seaweed aquaculture among Pacific Island countries. Journal of Applied Phycology 18: 227–234. Potín, P., Bouarab. K., Salaün, J-P., Pohnert, G., Kloareg, B. 2002 Biotic interactions of marine algae. Current Opinion Plant Biology 5: 308–317 Reis, R.P., Yoneshigue-Valentin, Y., Santos, C.P. 2008. Spatial and temporal variation of Hypnea musciformis carrageenan (Rhodophyta-Gigartinales) from natural beds in Rio de Janeiro State, Brazil. Journal of Applied Phycology 20:1–8. Weinberger, F. 2007. Pathogen-induced defense and innate immunity in macroalgae. Biological Bullutin 213: 290–302. ____________, Potin, P. 2010. Red algal defenses in the genomics age. In: Chapman DJ, Seckbach J (eds) Red algae in the genomic age, 1st edn. Springer, New York, pp 457–472. ____________, Friedlander, M., Hoppe, H.G. 1999. Oligoagars elicit a physiological response in Gracilaria conferta (Rhodophyta). Journal Phycology 35: 747–755. 43 ____________, Richard, C., Kloareg, B., Kashman, Y., Hoppe, H.G., Friedlander, M. 2001 Structure activity relationships of oligoagar elicitors toward Gracilaria conferta (Rhodophyta). Journal of Phycology 37: 418–426. Whapham, C.A., Blunden, G., Jenkins, T., Hankins, S.D. 1993. Significance of betaines in the increased chlorophyll content of plants treated with seaweed extract. J Journal of Applied Phycology 5: 231–234. Yokoya, N.S., Stirk, W.A., van Staden, J., Novák, O., Turecková, V., Pňnčík, A., Strnad, M. 2010. Endogenous cytokinins, auxins, and abscisic acid in read algae from Brazil. Journal of Phycology 46:1198–1205 Yunque, D.A.T., Tibudos, K.R., Hurtado, A.Q., Critchley, A.T. 2011. Optimization of culture conditions for tissue culture production of young plantlets of carrageenophyte Kappaphycus. Journal of Applied Phycology 23: 433–438. 44 Capítulo III: Efeito do extrato comercial da alga marrom Ascophyllum nodosum na produção de biomassa de Kappaphycus alvarezii (Doty) Doty ex PC Silva e no seu rendimento e a qualidade do gel de carragenana cultivada no Brasil Rafael R. Loureiro; Renata P. Reis; Flávia D. Berrogain e Alan T. Critchley. Artigo submetido ao Journal of Applied Phycology. 45 1. Introdução Eucheumatoides é um termo que inclui espécies do género Kappaphycus Doty e Ecuheuma J.Agardh, que são valiosos produtos cultivados para a indústria de hidrocolóides e, juntos, contribuem com 88% da produção mundial de carragenana. Entre as eucheumatoides, Kappaphyccus alvarezii Doty ex (Doty) P.C. Silva é a principal fonte de matéria-prima para a indústria sendo cultivada em mais de 20 países (Bindu e Levine 2011). Apesar de seu sucesso, o cultivo de K. alvarezii vem sofrendo uma série de complicações agronômicas que são comumente encontradas em cultivos na terra (i.e. infestações de pragas e doenças), o que gerou uma grande preocupação para o cultivo de algas marinhas mundial, principalmente relacionada a indústria de carragenana. O epifitismo, causado principalmente por um pequeno número de macroalgas filamentosas vermelhas e outras espécies: Neosiphonia savatieri (Hariot) M.S.Kim & I.K.Lee; N. apiculata (Hollenberg) Masuda & Kogame, Ceramium sp. Roth, Acanthophora sp. J.V. Lamouroux, Ulva sp. Lineu e Centroceras sp. Kützing afeta a produção de biomassa, uma vez que compromete a porção cortical, abrindo feridas nos talos, permitindo infecções bacterianas que causam necrose do talo (Vairappan 2006; Vairappan et al. 2008). Problemas adicionais, que começam com por trocas nosfatores ambientais, resultando em uma condição chamada ice-ice, sintoma que resulta no clareamento do talo, culminando na menor produção de biomassa e de carragenana. Mendonza et al. (2002) observaram uma depolimerização da carragenana nas porções apicais do talo que diminuiu a qualidade da carragenana produzida por mudas afetadas (diminuindo a viscosidade e a resistência do gel). Surtos de ice-ice têm sido mais dominantes nas Filipinas e Indonésia, com relatos recentes no leste da África, mas as perdas nas colheitas parece ser um fenômeno causado pelo epifitismo e, depois de estabelecido, mais difícil de ser mitigado. A remoção completa dos cultivares e a destruição das mudas são as alternativas atualmente usadas para resolver rapidamente esses problemas (Mendonza et al. 2002; Hurtado et al. 2006; Vairappan et al. 2006). Ao considerar o cultivo de K. alvarezii como uma "cultura global", por serem usadas mudas de origem genética semelhante nos cultivos espalhados pelo mundo, todos os cultivos estão suscetiveis a tais surtos. 46 Esforços têm sido realizados na tentativa de solucionar este problema com o uso do extrato comercial da macroalga marrom Ascophyllum nodosum (Linnaeus) Le Jolis, chamado AMPEP, nas mudas cultivadas de K. alvarezii. O AMPEP aumentou a taxa de crescimento diário e o rendimento de carragenana de K. alvarezii bem como atenuou os efeitos prejudiciais de epífitas (Hurtado et al. 2009; Loureiro et al. 2010; Yunque et al. 2011; Borlongan et al. 2012). O uso de AMPEP em K. alvarezii tem se mostrado promissor e foi proposto por Loureiro et al. (2012), o "efeito de vacina", para explicar as vias bioquímicas envolvidas nas respostas positivas observadas da ação do extrato nas mudas de K. alvarezii. No entanto, nenhum dos estudos analisou o efeito da AMPEP sobre a qualidade (propriedades reológicas, tais como a força e viscosidade do gel) da carragenana extraída K. alvarezii. Este estudo avaliou o efeito do AMPEP na taxa de crescimento diário, no rendimento da carragenana, na força e na viscosidade do gel de amostras de K. alvarezii tratadas com AMPEP. 2. Materiais e Métodos Área de estudo e desenho experimental O domínio amostral foi composto por três cultivos comerciais produtivos na baía de Sepetiba, litoral sul do estado do Rio de Janeiro, Brasil: Prainha (22°57'17''S 43°54'31''W), Jardim (22°56'25''S - 43° 55'08'’W) e Praia Leste (22°56'01''S 43°52'20''W) de modo a incluir diferentes sítios submetidos a diferentes condições ambientais. A baía tem um clima tropical (verões chuvosos e invernos secos) com alta irradiação no verão que diminui no inverno (Góes & Reis 2011). Neste experimento foram usadas as balsas flutuantes de cultivos comerciais, que usavam a técnica de rede tubular por ser a principal técnica de cultivo usada em cultivos brasileiros (Góes & Reis 2011). Cada balsa era composta por 10 a 15 módulos (3x5m) e cada módulo feito com dois tubos de PVC (as extremidades fechadas) ligados uns aos outros por cordas de polipropileno com 6 milímetros de diâmetro. Para a montagem do experimento, oito balsas foram selecionadas aleatoriamente em cada local de cultivo, que continham de 15 a 20 balsas flutuantes. Quatro receberam o tratamento AMPEP e as outras atuaram como o tratamento controle. De cada balsa comercial (compostas com 10 a 15 módulos), um módulo foi escolhido aleatoriamente. Quatro redes tubulares foram colocadas em cada módulo, duas para serem removidas em 20 dias e as outras duas em 40 dias. Em cada rede tubular foram colocadas oito 47 mudas. O tratamento com AMPEP consistiu em banhar amostras de K. alvarezii (indivíduos com 100±10g cada) em uma solução de 20 g.L-1 do extrato AMPEP com água do mar durante 1h, adaptando o protocolo utilizado por Loureiro et al. (2010). Para assegurar a cobertura total do talo, as amostras foram colocadas em recipientes plásticos com capacidade de 60 L com a solução. O período de 40 dias foi escolhido uma vez que é semelhante ao do ciclo de produção de 45 dias usado no Brasil (Góes & Reis 2011; 2012). Taxa de crescimento diário de K. alvarezii No final de cada período de cultivo, o crescimento das amostras foi estimado como taxa de crescimento diário (TCD) e calculada de acordo com a fórmula: TCD = 100 x [ln (Mt/Mo)/t] onde Mt foi a massa final , Mo a massa inicial e t o tempo de cultivo (Góes & Reis 2011, 2012). Rendimento da carragenana semirefinada de K. alvarezii Doze amostras, de cada tratamento, foram levadas para o laboratório para a obtenção da carragenana semirefinada de K. alvarezii através do processo de transformação alcalina com solução de hidróxido de potássio 6% em banho-maria (80°C) durante 2 h com lavagem sucessiva com água fresca. As amostras foram então mantidas em solução de cloreto de sódio 0,06%, durante 30 min (25°C) e lavadas com água doce para remoção dos resíduos. As amostras foram secas a 60°C em estufa até obtenção de massa constante e o pH 8,4 foi mantido durante o procedimento (Loureiro et al. 2010; Góes & Reis 2011; 2012). O rendimento de carragenana de K. alvarezii (RC) foi expresso como a percentagem de carragenana em relação à massa seca das algas (Reis et al. 2008; Góes & Reis 2011; 2012), derivado da fórmula: RC (Mc/Ms)x100, onde Mc é a massa seca da carragenana e Ms é a massa seca da alga (secas em estufa a 60°C até massa constante). Reologia de K. alvarezii Para preparar o gel da carragenana semirefinada, cada amostra foi macerada em moinho de bola (Mixer Mill MM 400, Retsch) e misturada em uma solução de água destilada na proporção de 1,5% de carragenana semirefinada (1,5 g de carragenana semirefinada e 98,5 mL de água destilada). A solução foi mantida durante 1 h em 48 temperatura ambiente (25°C), depois aquecida (80ºC) durante 10 minutos em agitação constante e posteriormente resfriada (7±2°C) durante 24 h. A viscosidade do gel foi analisada em reômetro de cone DV-III Ultra (Brookfield Engineering Labs, Inc.) em 75ºC com sonda CP-40. A força de gel das amostras foi medida com um texturometro, TexturePro CT (Brookfield Engineering Labs, Inc.) com uma sonda cilíndrica de 2,5cm de largura e 5cm de altura. As medições reológicas foram realizadas após 40 dias, o que reflete um ciclo de cultivo completo, o que permite a investigação de qualquer efeito do AMPEP sobre K. alvarezii relacionados à qualquer possível transformação molecular da carragenana. Análise estatística dos dados A normalidade (Teste de Shapiro-Wilks) e a homogeneidade da variância (teste de Levene) foram testadas para o uso de análises paramétricas ou não paramétricas dos dados. As diferenças entre as amostras do controle e do AMPEP em cada período de cultivo (taxa de crescimento diário e rendimento da carragenana em 20 dias e viscosidade e força do gel) foram testadas utilizando o teste de Mann-Whitney, uma vez que, mesmo após a transformação, os dados não satisfizeram os critérios para testes paramétricos. A taxa de crescimento diário e rendimento de carragenana em 40 dias foram analisados por meio de teste de Student. As análises foram realizadas utilizando o programa Statistica 6.0, da StatSoft, no nível de confiança de 95% (p <0,05). Os dados foram expressos em valores (mínimo) média±desvio padrão (máximo) com 12 repetições (n = 12) para cada tratamento. 3. Resultados Após 20 dias de cultivo, não houve diferenças significativas entre os tratamentos (p=0,43, Tab.8) das taxas de crescimento diário, entretanto, o rendimento de carragenana foi maior nas amostras tratadas com AMPEP (p<0,001, Tab.8). Em 40 dias, tanto a taxa de crescimento diário quanto o rendimento de carragenana das amostras tratadas com AMPEP foram maiores quando comparadas com seus controles (p<0,001, Tab.8). A resistência do gel da carragenana semirefinada de K. alvarezii tratada com AMPEP foi menor do que as amostras controle (p=0,03, Tab. 8), semelhante a viscosidade do gel (p<0,001, Tab.8) que também foi menor nas amostras que receberam o tratamento AMPEP. 49 Tabela 8 Resultados de taxa de crescimento diario (TCD), redimento de carragenana (RC), força do gel (FG) e viscosidade do gel (VG) de amostras de K. alvarezii (n=12) submetidas a dois tratamentos (controle e AMPEP) em dois periodos de cultivo (20 e 40 dias). Variavel Tempo 20 TCD (%.dia-1) 40 RC Treatmento (mín)media±desvio padrão(max) Controle (3,6) 5,9±2,1 (8,9) AMPEP (4,1) 6,2±1,0 (7,5) Controle (1,9) 5,6±1,8 (7,9) AMPEP (4,7) 7,3±1,7 (10,2) Controle (17,5)31,6±6,2(46,6) 20 AMPEP (17,0)38,0±6,7(48,3) 40 Controle (22,1)35,1±8,9(50,3) AMPEP (34,2)42,0±5,9(53,8) Controle (255,3)479,2±150,58(750,1) AMPEP (205,8)369,0±133,6(533,5) Control (610,3)923,7±211,34(1228,9) AMPEP (270,3)527,2±133,6(879,7) Teste p Resultado Z= -0,79 0,43 C = AMPEP Z= -0,44 0,01 C < AMPEP Z= -3,64 0,55 C = AMPEP Z= -2,40 0,02 C < AMPEP Z= 1,45 0,03 C > AMPEP t= -563,21 <0,001 C > AMPEP (%) FG (g.cm-2) VG 40 40 (cP) 4- Discussão As taxas de crescimento diário de K. alvarezii cultivadas no mar, em condições de produção comercial, durante 20 e 40 dias, foram, em gera, maiores do que o que é esperado para a baía de Sepetiba. Na porção sul da baía de Sepetiba, Góes & Reis (2011; 2012) relataram taxas médias de crescimento de K. alvarezii de 3,3 a 3,8 %. dia-1. Os resultados obtidos neste estudo estão dentro da faixa obtida para mudas de K. alvarezii cultivadas em balsas flutuantes no Brasil e em outros países, variando entre 0,8 a 7,2% dia-1 (Muñoz et al. 2004; Hayashi et al. 2007a; 2007b; Hung et al. 2009). Os resultados das taxas de crescimento diário de K. alvarezii obtidos neste estudo e as in vitro (5,5 %.dia-1 amostra AMPEP dia-1 e 2,5 % dia-1 amostra controle) por Loureiro et al. (2010), que usaram o mesmo protocolo para tratar as amostras com solução de AMPEP podem ser consideradas promissoras quando comparadas com os resultados obtidos por by Borlongan et al. (2011). Eles examinaram a eficiência do AMPEP em duas variantes de K.alvarezii em diferentes profundidades em cultivo usando uma baixa concentração de extrato (0.1 gL-1 de AMPEP) e exposição das amostras de K. alvarezii a um menor período (30 min). Eles também obtiveram 50 resultados positivos no implemento do crescimento (1,3 a 4,1 %.dia-1 em amostras tratadas com AMPEP e 0,9 a 3,0 %.dia-1 nas amostras controle), entretanto, quando seus resultados foram comparados com os nossos, as taxas de crescimento nas águas brasileiras foram maiores. Yokoya et al. (2010) relataram a presença de citocininas em algumas algas vermelhas e discutiram a importância dos hormônios na fisiologia de crescimento das algas vermelhas. Maiores concentrações de AMPEP induziram a maiores taxas de crescimento de K. alvarezii, que pode estar relacionado aos compostos estimulatórios presentes no extrato relacionado com a síntese de citocinina (Khan et al. 2008; 2009) e pela presença de certas betaínas (Mackinnin et al. 2009) como observados em plantas superiores. O rendimento da carragenana de K. alvarezii em 20 e 40 dias (amostras AMPEP e controle) estiveram dentro da faixa ideal para os padrões industriais, próximas a 30% (Hung et al. 2009). Hayashi et al. (2007a), Reis et al. (2008) e Góes & Reis (2012) discutiram a importância das mudanças ambientais como fatores de estresse, tais como mudanças de salinidade e movimentação da água, na conformação da carragenana, que consequentemente causam alterações nas suas propriedades reológicas, assim como as respostas a esses estresses. AMPEP pode atuar com elicitor para as respostas de K.alvarezii as condições de estresse, no aumento de suas defesas e causando uma maior produção de carragenana (Loureiro et al. 2012). Isto pode explicar o resultado de maior rendimento de carragenana das amostras de K.alvarezii expostas ao AMPEP, em 20 dias de cultivo quando comparado com o resultado das amostras do tratamento controle. No entanto, em longo prazo, este fator não é prejudicial para K. alvarezii uma vez que o extrato parece transportar uma série de indutores e moléculas de protecção (isomorfos específicos de enzimas antioxidantes, como o superóxido dismutase [MnSOD-1, ~ 150 kDa], ascorbato peroxidase [APX-4, ~ 55 kDa], glutationa peroxidase [GSH-Px-2, ~ 55 kDa] e glutationa redutase [GR-1, ~ 180 kDa] as quais especificamente responderam à suplementação AMPEP) que, mesmo sob estresse e a exposição a pulsos oxidativos (como uma resposta a condições desfavoráveis), são capazes de sustentar e proteger as amostras, mesmo depois de um breve período de exposição (Loureiro et al. 2012; Kumar et al. 2012). Além de melhorar a taxa de crescimento e de produção de carragenana, AMPEP parece ter sido eficaz na redução de algumas epífitas (Loureiro et al. 2010). Este fator 51 não foi testado, mas foi observado nas mudas que foram tratadas por AMPEP. Uma vez que os experimentos foram realizados em balsas flutuantes de cultivos comerciais, quando os módulos das balsas que continham amostras tratadas com AMPEP foram comparados com os módulos das balsas do cultivo comercial, aparentemente os módulos experimentais continham cerca de 20% menos de bioincutação do que os módulos que não foram usados no experimento (observação pessoal). AMPEP é rico em potássio solúvel (17-22%) em sua composição (Hurtado et al. 2009; Loureiro et al. 2010) e K. alvarezii em kappa carragenana (Pereira et al. 2013). Como kapa carragenana é conhecida por ser sensível aos íons de potássio, a carragenana extraída das amostras tratadas com AMPEP deveriam produzir géis mais rígidos. Isto pode ser explicado pela troca na sinerese (extrusão espontânea de água através da superfície do gel quando está em repouso) das mudas quando expostas a soluções aquosas, ricas em potássio solúvel. Altas concentrações de potássio deveriam exibir maior sinerese resultar em maior força de gel. No entanto, a força do gel de K. alvarezii foi maior nas amostras controle, mas dentro da faixa obtida por Góes e Reis (2011, 2012) em cultivos na baía de Sepetiba e em outros locais (Ohno et al. 1994;. Hayashi et al. 2007a; 2007b), que leva à ideia de que o potássio disponível pela exposição ao tratamento AMPEP que foi absorvido por K.alvarezii, quando a alga foi cultivada no mar não foi suficiente para alterar a estrutura molecular do carragenana extraída. Apesar da viscosidade da carragenana derivada das amostras de K. alvarezii tratadas com AMPEP terem sido menores do que as amostras controle, estes valores foram semelhantes aos registrados para a Baía de Sepetiba por Góes & Reis (2011; 2012). Quanto maior a viscosidade, esta pode ser creditada às diferentes técnicas de análise aplicadas no processo de confecção do pó de carragenana semirefinado (Montolalu et al. 2008). Basmal et al. (2009) analisou o efeito de um novo agente, KCl, precipitando carragenana em diferentes concentrações e do seu impacto sobre as leituras de força e viscosidade do gel. As amostras testadas demonstram que as técnicas que utilizam diferentes concentrações de KCl podem produzir diferentes leituras de força e viscosidade de gel mesmo usando as mesmas amostras. Qaundo avaliando o rendimento e qualidade de carragenana, Mendoza et al. (2002) encontraram que as amostras saudáveis forneciam um extrato com melhor qualidade do gel (maior força gel e maior viscosidade) quando comparado com amostras estressadas que apresentavam sintomas de ice-ice. As amostras afetadas por 52 ice-ice mostraram alterações na massa molecular, impedindo a formação correta de pontes de hidrogénio, o que refleteu diretamente na capacidade para formar a dupla hélice que ocorre no processo regular de gelificação. A carragenana é constituida de unidades alternadas de D-galactose e 3,6-anidro-galactose (3,6-AG), unidas por α-1, 3 e β-1, 4-glucosídicas. Níveis mais altos de sulfato de éster no extrato em pó podem resultar em menor força e viscosidade do gel, devido a uma alteração da massa molecular (Craigie e Wong 1979; Craigie 1990; Knutsen et al. 2001). Os resultados do presente estudo corroboram com os de Loureiro et al. (2012), no qual o AMPEP atua como um elicitor das próprias defesas de K. alvarezii. 5. Conclusão O uso de AMPEP no cultivo de K.alvarezii in situ é recomendado para aumentar o crescimento de K. alvarezii, uma vez que após 40 dias a taxa de crescimento diário aumentou e foi superior aos valores obtidos para a região da baía de Sepetiba. Além disso, mais carragenana foi produzida quando comparada com amostras não submetidas ao AMPEP. Apesar do efeito negativo na qualidade da carragenana semirefinada os valores estivram dentro dos valores comerciais. 6- Referências bibliográficas Bindu, M.S., Levine, I.A. 2011. The commercial red seaweed Kappaphycus alvarezii an overview on farming and environment. Journal of Applied Phycology 23: 789796. Borlongan, I.A.G. , Tibudos, K.R. , Yunque, D.A.T. , Hurtado, A.Q. , Critchley, A.T. 2011. Impact of AMPEP on the growth and occurrence of epiphytic Neosiphonia infestation on two varieties of commercially cultivated Kappaphycus alvarezii grown at different depths in the Philippines. Journal of Applied Phycology 23: 615621. Craigie, J.S.1990. Cell walls. In: Cole K M and Sheath R G, editors. Biology of the Red Algae, Cambridge: Cambridge University Press p.221–25. _________, Wong, K.F.1979. Carrageenan biosynthesis. Proceeding of the International Seaweed Symposium 9: 369–377. 53 Góes, H.G., Reis, R.P. 2011. An initial comparison of tubular netting versus tie–tie methods of cultivation for Kappaphycus alvarezii (Rhodophyta, Solieriaceae) on the south coast of Rio de Janeiro State, Brazil. Journal of Applied Phycology 23: 607-613. _________, ________ 2012. Temporal variation of the growth, carrageenan yield and quality of Kappaphycus alvarezii (Rhodophyta, Gigartinales) cultivated at Sepetiba bay, southern Brazilian coast. Journal of Applied Phycology 24:173-180 Hayashi, L., Oliveira, E.C., Bleicher-Lhonneur, G., Boulenguer, P., Pereira, R.T.L., Seckendorff, R., Shimoda, V.T., Leflamand, A., Vallée, P. & Critchley, A.T. 2007a. The effects of selected cultivation conditions on the carrageenan characteristics of Kappaphycus alvarezii (Rhodophyta,Solieriaceae) in Ubatuba Bay, São Paulo, Brazil. Journal of Applied Phycology 19: 505–511. _________, Paula, E.J. & Chow, F. 2007b. Growth rate and carrageenan analyses in four strains of Kappaphycus alvarezii (Rhodophyta, Gigartinales) farmed in the subtropical waters of São Paulo State, Brazil. Journal of Applied Phycology. 19:393–399. ___________, Santos, A.A., Faria, G.S.M. , Nunes, B.G. , Souza, M.S. , Fonseca, A.L.D. , Barreto, P.L.M., Oliveira, E.C., Bouzon, Z.L. 2011. Kappaphycus alvarezii (Rhodophyta, Areschougiaceae) cultivated in subtropical waters in Southern Brazil. Journal of Applied Phycology 23: 337-343. Hung, L.D., Hori, K., Nang, H.Q., Kha, T. & Hoa, L.T. 2009. Seasonal changes in growth rate, carrageenan yield and lectin content in the red alga Kappaphycus alvarezii cultivated in Camranh Bay, Vietnam. Journal of Applied Phycology 21: 265-272. Hurtado, A.Q., Critchley, A.T., Bleicher-Lhonneur, G. 2006. Occurrence of Polysiphonia epiphytes in Kappaphycus farms at Calaguas Is., Camarines Norte, Philippines. Journal of Applied Phycology 18:301-306. 54 ___________, Yunque, D.A., Tubudos, K., Critchley, A.T. 2009. Use of Acadian marine plant extract powder from Ascophyllum nodosum in tissue culture os Kappaphycus varieties. Journal of Applied Phycology 21: 633-639. Khan, W., Palanisamy, R., Hankins, S.D., Critchley, A.T., Smith, D.L., Papadopoulos, Y., Prithiviraj, B. 2008. Ascophyllum nodosum (L.) Le Jolis extract improves root nodulation in alfalfa. Canadian Journal of Plant Science 88:728-728. ______, Rayirath, U.P., Subramanian, U.P., Jitesh, M.N., Rayorath, P., Hodges, D.M., Critchley, A.T., Craigie, J.S., Norrie, J., Prithiviraj, B. 2009. Seaweed Extracts as Biostimulants of Plant Growth and Development. Journal of Plant Growth Regulators 28:386-399. Knutsen, S.H., Sletmoen, M., Kristensen, T., Barbeyron, T., Kloareg, B. and Potin, P. 2001. A rapid method for the separation and analysis of carrageenan oligosaccharides released by iota- and kappa-carrageenase. Carbohydrates 331: 101-106. Kumar, M., Reddy, C.R.K., Jha, B. 2012. The ameliorating effect of Acadian marine plant extract against ionic liquids-induced oxidative stress and DNA damage in marine macroalga Ulva lactuca. Journal of Applied Phycology 10.1007s10811-0129871-8. Loureiro, R.R., Reis, R.P., Critchley, A.T. 2010. In vitro cultivation of three Kappaphycus alvarezii (Rhodophyta, Areschougiaceae) variants (green, red and brown) exposed to a commercial extract of the brown alga Ascophyllum nodosum (Fucaceae, Ochrophyta). Journal of Applied Phycology 22: 101-104. ____________, Reis, R.P., Berrogain, F.D., Critchley, A.T. 2012. Extract powder from the brown alga Ascophyllum nodosum (Linnaeus) Le Jolis (AMPEP): A "vaccinelike" effect on Kappaphycus alvarezii (Doty) Doty ex P.C. Silva. Journal of Applied Phycology 24:427-432. 55 Mackinnon, S.L., Hiltz, D., Ugarte, R., Craft, C.A. 2009. Improved methods of analysis for betaines in Ascophyllum nodosum and its commercial seaweed extracts. Journal of Applied Phycology 22:489-494. Mendonza, W.G., Montaño, E.N., Gazon-Fortes, E.T., Villanueva, R.D. 2002. Chemical and gelling profile of ice-ice infected carrageenan from Kappaphycus striatum (Schmitz) Doty “sacol” strain (Solieriaceae,Gigartinales, Rhodophyta Journal of Applied Phycology 14: 409-418. Muñoz, J., Freile-Pelegrín, Y., Robledo, D. 2004. Mariculture of Kappaphycus alvarezii (Rhodophyta, Solieriaceae) color strains in tropical waters of Yucatán, México. Aquaculture 239:161–177. Ohno, M., Largo, B.D., Ikumoto, T. 1994. Growth rate, carrageenan yield and gel properties of cultured kappa-carrageenan producing red alga Kappaphycus alvarezzi (Doty) Doty in the subtropical waters of Shikoku, Japan. J. Journal of Applied Phycology 6:1-5. Pereira L, Gheda SF, Ribeiro-Claro PJA (2013) Analysis by vibrational spectroscopy of seaweed polysaccharides with potential use in food, pharmaceutical and cosmetic industries. International Journal of Carbohydrate Chemestry 2013:1-7 Reis, R.P., Yoneshigue-Valentin, Y., Santos, C.P. 2008. Spatial and temporal variation of Hypnea musciformis carrageenan (Rhodophyta-Gigartinales) from natural beds in Rio de Janeiro State, Brazil. Journal of Applied Phycology 20:1-8. Vairappan, C. S. 2006. Seasonal occurrences of epiphytic algae on the commercially cultivated red alga Kappaphycus alvarezii (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 18: 611-617. ___________, Chung, C.S., Hurtado, A.Q., Flower, E.S., Lhonneur, G.B. & Critchley, A.T. 2008. Distribution and symptoms of epiphyte infection in major carrageenophyte-producing farms. Journal of Applied Phycology 20:477-483. 56 Yokoya, N.S., Stirk, W.A., van Staden, J., Novák, O., Turecková, V., Pňnčík, A., Strnad, M. 2010. Endogenous cytokinins, auxins, and abscisic acid in read algae from Brazil. Journal of Phycology 46:1198–1205. Yunque, D.A.T., Tibudos, K.R. , Hurtado, A.Q., Critchley, A.T. 2011. Optimization of culture conditions for tissue culture production of young plantlets of carrageenophyte Kappaphycus. Journal of Applied Phycology 23: 433-438. 57 Capítulo IV: Efeito do extrato comercial da alga marrom Ascophyllum nodosum Mont. em cultivo de Kappaphycus alvarezii (Doty) Doty ex P.C. Silva submetido à temperaturas letais Rafael R. Loureiro; Renata P. Reis & Rafael G. Marroig Artigo submetido ao Journal of Applied Phycology. 58 1. Introdução Kappaphyccus alvarezii Doty ex (Doty). P.C. Silva é a principal fonte de matéria prima para a indústria de carragenana (Bixler e Porse 2011; Bindu & Levine 2011). Para suprir a demanda brasileira por carragenana, esta espécie foi introduzida experimentalmente no Brasil na década de 90, seguido pela introdução em fazendas comerciais. Entretanto, existe a necessidade de adaptar as técnicas de cultivo aos diferentes fatores ambientais inadequados ao cultivo desta espécie neste novo ambiente. A solução para o cultivo comercial de K. alvarezii foi o uso do sistema de balsas flutuantes feitas com PVC e o uso da técnica de rede tubular em locais com fundo lodoso e maior movimentação de água. Este sistema de cultivo foi espalhado por toda a área permitida para cultivo comercial de K. alvarezii pelo Governo brasileiro na costa sudeste (Góes & Reis 2011). Hayashi et al. (2011) teve sucesso com cultivo experimental de K. alvarezii, usando a mesma técnica no sul do Brasil, no estado de santa catarina. Entretanto, no inverno a temperatura da água do mar (16-180C) na sua área amostral foi letal para as amostras cultivadas. Para a viabilidade do cultivo de K. alvarezii nesta área existe a necessidade de permitir o crescimento desta espécie em baixas temperaturas. Hayashi (comunicação persoal) comentou que a melhor maneira de fazer isto é cultivar esta alga em tanques durante os meses de baixas temperaturas. Outra opção que poderia ser estabelecida em sítios com baixas temperaturas consiste em obter mudas saudáveis para serem cultivadas até o final da estacao fria. Entretanto, partino do pressuposto que a perda de mudas deve ser usada no futiros cultivos, existe a necessidade de translocar mudas de outros plantios, que aumentará os custos para previnir a introdução de organismos acompanhantes das mudas translocadas, que apesar de serem considerados organismos nativos para a costa brasileira, eles podem ser tratados como organismos exóticos em locais nos quais a translocação das mdas ocorrerá. Processos de quarentena devem ser estabelecidos, entretanto até o momento ainda nao foram implantados. Com base no beneficiamento do uso do extrato comercial em pó de Ascophyllum nodosum, (Acadian Marine Plant Extract - AMPEP), nos cultivos de K. alvarezii in vitro e no mar, o qual tornou a alga mais resistente à algumas epífitas e aumentou seu crescimento e rendimento de carragenana (Hurtado et al. 2009; Loureiro et al. 2010; Yunque et al. 2011; Borlongan et al. 2011; Loureiro et al. 2012). Foi levantada a 59 hipótese de que mudas previamente submetidas à solução de AMPEP serão mais resistentes à algumas das temperaturas consideradas letais ao crescimento desta espécie. Para testar esta hipótese, nos avaliamos o efeito do uso da solução de AMPEP em K. alvarezii in vitro nas baixas tenmperaturas que ocorrem no sul do Brasil entre o outono e a primavera (Hayashi et al. 2011). Os parâmetros escolhidos para a avaliação da produção e desenvolvimento de K. alvarezii sob estresse de temperatura foi a taxa de crescomento diário eo rendimento e qualidade de carragenana (viscosidade e rigidez do gel). 2. Material e Métodos Condições laboratoriais Mudas de K. alvarezii foram obtidas em cultivo comercial em Paraty Mirim, Município de Paraty, estado do Rio de Janeiro, Brasil (23°14'52"S e 44°39'09" W). As amostras foram limpas de epífitas e aclimatadas durante 30 dias em aquários de 5 litros (temperatura da água - 22±2°C; irradiância -130±5 μmol.fótons m−2·s−1; fotoperíodo 12:12h; salinidade (PSS); 35±1 e pH - 8.1±0,3). Desenho experiemental Após o período de aclimatação, as algas foram banhadas em solução de AMPEP (20 gL-1 de água do mar filtrada , 0.45µm), durante 1h, em recipientes com 12 litros de água do mar filtrada (0,45 Milipore), com aeração constante. As amostras que não foram banhadas em solução de AMPEP foram denominadas como controle. Cada amostra foi composta por porções apicais de K. alvarezii (5 cm de comprimento, em um total de 5g de massa úmida) e foi cultivada em Erlenmeyer, com 500 ml de água filtrada (0.45µm) e aeração constante. Oito repetições de amostras AMPEP e de amostras controle foram submetidas a três temperaturas T1 (16±1oC); T2 (19±1oC) e T3 (22±1oC). As temperaturas T1 e T2 correspondem às consideradas letais ao crescimento de K. alvarezii. A temperatura T3 é a que ocorre nos cultivos comerciais de K. alvarezii no sul fluminense (Góes & Reis, 2011). As temperaturas foram manipuladas com coolers (ventiladores), marca BAYO FS-120, em recipientes plásticos contendo os Erlenmeyers. O experimento teve duração de 20 dias. Crescimento, rendimento e qualidade da carragenana de K. alvarezii O crescimento das amostras foi estimado como taxa de crescimento diário 60 (TCD), que corresponde à TCD (% day-1) =[(Mt /M0)1/t−1]×100% onde M0 é a massa úmida inicial, Mt é a massa úmida final e t os dias de cultivo (Yong et al. 2013). Para calcular o rendimento da carragenana das amostras de K. alvarezii, pelo processo de transformação alcalina foi obtida a carragenana semirefinada das amostras. Neste processo as amostras permaneceram em solução KOH a 6% em banho-maria (80°C), durante 2 h, com posterior lavagem em água corrente e manutenção em solução de NaClO de 0,06%, durante 30 min, em temperatura ambiente (25°C). As amostras foram lavadas em água corrente para remoção dos resíduos e foram secas em estufa (60°C) até obtenção de massa seca constante. O pH 8,4 foi mantido durante o procedimento (Góes & Reis 2011; 2012; Loureiro et al. 2012). O rendimento da carragenana (RC) de cada amostra de K. alvarezii (n=8) foi estimado através do cálculo da percentagem de carragenana semirefinada obtida em relação à massa seca da amostra de alga (Góes & Reis 2011; 2012), pela fórmula RC = (Mc/Ma) x 100, onde Mc é a massa seca de carragenana e Ma é a massa seca da amostra de K. alvarezii. A qualidade da carragenana de K. alvarezii foi estimada pela força do gel (FG) e viscosidade do gel (VG) da carragenana de oito amostras de gel (n=8). Para o preparo do gel da carragenana semirefinada, cada amostra foi macerada em moinho de bola (Mixer Mill MM 400, Retsch) e misturada em 100 mL de água destilada na proporção de 1,5%. Esta solução ficou em repouso durante 1h, em temperatura ambiente (25°C), e foi aquecida (80ºC), durante 10 minutos em agitação constante e posteriormente foi resfriada (7±2°C) durante 24 h. A viscosidade do gel foi analisada em reômetro de cone, DV-III Ultra (Brookfield Engineering Labs, Inc.) em 75ºC, com sonda CP-40. A força de gel foi medida em texturômetro, TexturePro (Brookfield Engineering Labs, Inc.), com uma sonda cilíndrica em frasco com 2,5cm de largura por 5cm de altura. Análise estatística As premissas da normalidade (teste de Shapiro-Wilks) e da homogeneidade (teste de Cochran) dos dados foram previamente testadas. A diferença da taxa de crescimento diário e da viscosidade do gel das amostras de K. alvarezii (AMPEP e controle) cultivadas nas três temperaturas (T1, T2 e T3) foram obtidas Kruskal-Wallis ANOVA por nao encontraram as premissas para o uso de testes paramétricos meso após serem transformados. As medianas significativamente distintas foram identificadas 61 através de teste de comparações múltiplas. As diferenças estatísticas do rendimento da carragenana e da forca do gel das amostras de K. alvarezii nas três temperaturas foram obtidas pela análise de variância unifatorial (ANOVA unifatorial) e diferenças obtidas pelo teste LSD Fisher. As análises foram realizadas no programa Statistica 6.0, da StatSoft Inc. e foi usado o intervalo de confiança de 95% (p<0.05). Foram usadas oito repetições por tratamento (n=8). Os dados foram expressos como mádia ± desvio padrão. 3. Resultados Após o período de cultivo (20 dias), houve diferença significativa entre os tratamentos (H=43.83, p <0.001, Fig. 6). As maiores taxas de crescimento diário de K. alvarezii foram obtidas nas amostras submetidas ao tratamento AMPEP, quando comparadas com seus controles em todas as temperaturas (p<0,001, Fig. 6A). Na temperatura TI (16±1°C), as amostras do controle apresentaram branqueamento do talo, enquanto que este branqueamento não foi observado nas amostras submetidas ao tratamento AMPEP. Entretanto, ocorreram diferenças significativas no rendimento de carragenana das amostras controle e das AMPEP (F = 271.94, p < 0.001, Fig. 6B). Quando as amostras AMPEP foram comparadas com as amostras controle, os resultados nas temperaturas T1 (16±1°C) e T2 (19±1°C) foram diferentes das amostras cultivadas na temperatura T3 (22±1°C). Os maiores rendimentos foram obtidos nas amostras do controle (p<0.001, teste LSD) enquanto que na temperature T3 (22±1°C), o rendimento da carragenana das amostras AMPEP foi maior do que a das amostras controle (p <0.001, teste LSD). Resultados semelhantes foram obtidos com a viscosidade do gel de K. alvarezii cultivado nas diferentes temperaturas (H=33.30, p < 0.001, Fig. 6C), com maior viscosidade de gel nas amostras controle cultivadas nas temperaturas T1 e T2 e viscosidade de gel semelhantes (AMPEP and Control) nas cultivadas na temperature T3. Entretanto, a força do gel de K. alvarezii das amostras cultivadas nas diferentes temperaturas (F=65.67, p<0.001, Fig. 6D) mostraram respostas diferentes, com menor força de gel nas amostras controle quando cultivadas nas temperaturas T1 e T2 e sem diferenca significativa na temperature T3. 62 Figura 6. (A) Taxa de crescimento diário, (B) redimento de carragenana, (C) força do gel e (D) viscosidade do gel de amostras de K. alvarezii previamente banhadas em solução de AMPEP (TA) ou não tratadas (TC) (n=8) e submetidas a três temperaturas (T1 =16±1 oC; T2 =19±1 oC e T3 = 22±1 oC). 4. Discussão As taxas de crescimento diário das mudas submetidas a solução de AMPEP e cultivadas em baixas temperaturas estiveram dentro da faixa de valores encontrados em cultivos comerciais no estado do Rio de Janeiro (Góes & Reis 2011; 2012) e na faixa encontrada em outros estudos in vitro e no mar sobre o efeito do AMPEP nesta espécie (Loureiro et al. 2010; 2012; Borlongan et al. 2011; Yunque et al. 2011). Altas taxas de crescimento das mudas de K. alvarezii foram obtidas em todas as tempertauras quando submetidas ao tratamento AMPEP, até em temperaturas consideradas como adversas para o crescimento desta espécie (Ohno et al. 2004; Hayashi et al. 2011). Isto talvez ocorra devido à presença de betaínas que promovem proteção a interrupção de transporte de elétrons pelo fotosistema II e evita a degradação de pigmentos, como descrito por Mackinnon et al. (2010). Entretanto, a presença de citocininas também poderia atuar como uma atividade elicitora no estrato de AMPEP e suas propriedades (Reitz & Trumble 1996; Khan et al. 2009). Trabalhos com vegetais superiores mostraram que o extrato de A. nodosum 63 administrado no solo ou como spray foliar foi um eficaz antioxidante no combate a pragas ou na proteção dos efeitos adeversos causados pela longa exposição a respostas de defesa própria [ROS – Reactive Oxigen Species] (Jayaraj et al. 2008; Khan et al. 2008; 2009; Parys et al. 2009; Jayaraman et al. 2011). O efeito benéfico do uso do AMPEP em outros organismos que foram submetidos ao estresse térmico também mostrou resultados promissores. Rayirath et al. (2008) observaram a eficiência deste extrato no auxílio a sobrevivência de Arabidopsis thaliana (L.) Heynh. in vitro (-7.5°C) e in situ (-5.5°C) e em geral atribuíram este benefício a presença de compostos lipofílicos (LPC). Nair e colaboradores (dados não publicados) também verificaram o efeito positivo do uso de extrato de A.nodosum em A. thaliana em baixas temperaturas e comentaram que a expressão de genes mostrou a acumulação de prolina, que foi mediada pelo aumento da expressão dos genes de síntese de prolina (P5CS1 P5CS2) e pela redução na expressão do gene da prolina desidrogenase (PRODH). Além disso, a presença elevada de LPCs aumentou significativamente a concentração de açúcares solúveis no citosol em resposta ao estresse de congelamento, comprovando o modo de ação dos LPCs na proteção contra baixas temperaturas e seus efeitos. Barros et al. (2006) quando submeteram K.alvarezii ao estresse por congelamento, observaram a exsudação de H2O2 (prevenindo a ação efetiva da catalase – CAT e a indução do gene APX (Ascorbate Peroxidase Gene). Foi sugerido que a exudação foi uma estratégia fisilógica da alga vermelha para suportar a produção rápida e elevada do altamente tóxico H2O2 em seus tecidos. Loureiro et al. (2012), observaram resultado semelhante quando induziram K.alvarezii a produzir H2O2 para avaliar o efeito do AMPEP em tais condições. É sugerido que o gradual (porém constante) acúmulo de H2O2 no entorno do cultivo pode ser responsável pelo atraso adequado nas respostas apropriadas de CAT e APX, como sugerido por Barros et al. (2006), que garante uma aclimatação em longo prazo, que permite que K. alvarezii aumente sua taxa de crescimento (aumento de biomassa) sob condições ambientais adversas combinadas com compostos favoráveis presentes no AMPEP e a sua ação antioxidante. Os valores de rendimento de carragenana das mudas de K. alvarezii nas três temperaturas (amostras AMPEP e controle) estiveram dentro da faixa considerada ideal para os valores da indústria, perto de 30% (Hayashi et al. 2007; Hung et al. 2009, Góes & Reis, 2011; 2012). Provavelmente, os maiores valores observados nas amostras em 64 baixas temperaturas (T1 e T2) quando comparadas com a temperatura T3, foi causado pelo conhecido aumento na produção de polissacarídeo (agar, carragenana e alginato) como a primeira defesa da alga em resposta aos estresses abióticos (Craigie & Wen 1984; Knutsen et al. 2001). A força e a viscosidade do gel de amostras de K. alvarezii foram independentes da faixa de temperatura que as amostras foram submetidas ou o efeito do tratamento AMPEP e estão dentro da faixa de valores encontrados em outros trabalhos com esta espécie em cultivos na baía de Sepetiba, RJ (Góes & Reis 2011; 2012) e em outros locais (Ohno et al. 1994; Hayashi et al. 2007). Os valores também estão incluídos nos esperados para a indústria de carragenana, força de gel superior a 200g.cm2 e viscosidade entre 30 e 800 cP (Góes & Reis 2011; 2012). Craigie e Wen (1984) comentaram que algas submetidas a baixas temperaturas apresentam alta força do gel. Isto está de acordo com os resultados deste estudo, independente das amostras terem sido banhadas ou não na solução de AMPEP. Além disso, a força do gel da carragenana das amostras AMPEP cultivadas em temperaturas baixas (T1 e T2) foram maiores do que as do controle. Possivelmente, pelo fato do AMPEP ser rico em potássio solúvel (17-22%) em sua composição (Hurtado et al. 2009; Loureiro et al. 2010) e a kappa carragenana ser sensível aos íons de potássio, que permitem a produção de géis mais rígidos quando as amostras são previamente expostas à soluções aquosas ricas em potássio solúvel. A temperatura também afetou a proporção de sulfato e 3,6 anidrogalactose na conformação final da carragenana, fato que está diretamente relacionado às propriedades reológicas do polissacarídeo. Em temperaturas baixas, a concentração de sulfato na carragenana é menor do que a de 3,6 anidrogalactose. Esta configuração molecular confere maior rigidez ao gel quando estes são comparados com géis de indivíduos cultivados em temperaturas mais altas (Craigie & Wong 1979; Craigie 1990; Craigie & Wen 1994). Alterações negativas na massa molecular da carragenana ocasionadas por fatores estressantes (nesse estudo a baixa temperatura) também impedem a formação correta de pontes de hidrogênio, que são responsáveis pela formação da dupla hélice, a qual aumenta o poder de gelificação do polissacarídeo. Esta deficiência acarreta a menor viscosidade do gel (Mendonza et al. 2002). Neste experimento, este padrão foi observado apenas nas amostras tratadas com a solução de AMPEP. 65 5- Conclusão A utilização do extrato comercial em extrato de A. nodossum, AMPEP, em amostras de K. alvarezii in vitro em baixas temperaturas atuou como um remediador ao estresse térmico, através do aumento nas taxas de crescimento diário e na força de gel. Apesar do menor rendimento de carragenana e de viscosidade do gel quando usado o AMPEP, os valores estão entre os utilizados na indústria. Desta maneira, o tratamento de mudas de K. alvarezii com solução de AMPEP pode ser uma alternativa como ação preventiva para o cultivo de mudas em tanques e no mar em períodos de baixas temperaturas. 6. Referências Bibliográficas Barros, M.P.; Necchi Jr.; O. Colepicolo P., & Pedersén, M. 2006. Kinetic study of the plastoquinone pool availability correlated with H2O2 release in seawater and antioxidant responses in the red alga Kappaphycus alvarezii exposed to single or combined high light, chilling and chemical stresses. Biochimica et Biophysica Acta 1757: 1520–1528 Bindu, M.S. & Levine, I.A. 2011. The commercial red seaweed Kappaphycus alvareziian overview on farming and environment. Journal of Applied Phycology 23: 789-796 Bixler, H.J. & Porse, H. 2011. A decade of change in the seaweed hydrocolloids industry. Journal of Applied Phycology 23:321–335 Borlongan, I.A.G.; Tibudos, K.R.; Yunque, D.A.T.; Hurtado, A.Q. & Critchley, A.T. 2011. Impact of AMPEP on the growth and occurrence of epiphytic Neosiphonia infestation on two varieties of commercially cultivated Kappaphycus alvarezii grown at different depths in the Philippines. Journal of Applied Phycology 23, 615-621. Craigie, J.S.1990. Cell walls. In: Cole K M and Sheath R G, editors. Biology of the Red Algae, Cambridge: Cambridge University Press p.221–25. Craigie, J.S. & Wen, Z.C. 1984. Effects of temperature and tissue age on gel stregth and composition of agar from Gracilaria tikhaviae (Rhodophyta). Canadian Journal of Botany 62: 1665-1670 66 Craigie, J.S. & Wong, K.F. 1979. Carrageenan biosynthesis. Proceedings of the International Seaweed Symposyum 9: 369–377. Góes, H.G. & Reis, R.P. 2011. An initial comparison of tubular netting versus tie–tie methods of cultivation for Kappaphycus alvarezii (Rhodophyta, Solieriaceae) on the south coast of Rio de Janeiro State, Brazil. Journal of Applied Phycology 23:607-613 Góes, H.G. & Reis, R.P. 2012. Temporal variation of the growth, carrageenan yield and quality of Kappaphycus alvarezii (Rhodophyta, Gigartinales) cultivated at Sepetiba bay, southeastern Brazilian coast. Journal of Applied Phycology 24:173-180 Hayashi L.; Oliveira, E.C.; Bleicher-Lhonneur, G.; Boulenguer, P.; Pereira, R.T.L.; von Seckendorff, R.; Shimoda, V.T.; Leflamand, A.; Vallée, P. & Critchley, A.T. 2007. The effects of selected cultivation conditions on the carrageenan characteristics of Kappaphycus alvarezii (Rhodophyta, Solieriaceae) in Ubatuba Bay, São Paulo, Brazil. Journal of Applied Phycology 19: 505–511. ___________, Santos, A.A.; Faria, G.S.M.; Nunes, B.G.; Souza, M.S.; Fonseca, A.L.D.; Barreto, P.L.M.; Oliveira, E.C. & Bouzon, Z.L. 2011. Kappaphycus alvarezii (Rhodophyta, Areschougiaceae) cultivated in subtropical waters in Southern Brazil. Journal of Applied Phycology 23, 337-343. Hung, L.D.; Hori, K.; Nang, H.Q.; Kha, T. & Hoa, L.T. 2009. Seasonal changes in growth rate, carrageenan yield and lectin content in the red alga Kappaphycus alvarezii cultivated in Camranh Bay, Vietnam. Journal of Applied Phycology 21, 265-272 Hurtado, A.Q.; Yunque, D.A.; Tibudos, K. & Critchley, A.T. 2009. Use of Acadian marine plant extract powder from Ascophyllum nodosum in tissue culture of Kappaphycus varieties. Journal of Applied Phycology 21:633–639 Jayaraj, J.; Wan, A.; Rahman, M. & Punja, Z.K. 2008. Seaweed extract reduces foliar fungal diseases on carrots. Crop Prot 27:1360–1366 67 Jayaraman, J.; Norrie, J. & Punja, Z.K. 2011. Commercial extract from the brown seaweed Ascophyllum nodosum reduces diseases in greenhouse cucumber. Journal of Applied Phycology 23: 353–361 Khan, W.; Palanisamy, R.; Hankins, S.D.; Critchley, A.T.; Smith, D.L.; Papadopoulos Y. & Prithiviraj, B. 2008. Ascophyllum nodosum (L.) Le Jolis extract improves root nodulation in alfalfa. Canadian Journal of Plant Science 88:728–728 ________; Rayirath, U.P.; Subramanian, U.P.; Jitesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J. & Prithiviraj, B. 2009. Seaweed Extracts as Biostimulants of Plant Growth and Development. Journal of Plant Growth Regulultors 28:386–39 Knutsen, S.H.; Sletmoen, M.; Kristensen, T.; Barbeyron, T.; Kloareg, B. & Potin, P. 2001. A rapid method for the separation and analysis of carrageenan oligosaccharides released by iota- and kappa-carrageenase. Carbohydrates 331, 101-106. Loureiro, R.R.; Reis, R.P.; Berrogain, F.D. & Critchley, A.T. 2012. Extract powder from the brown alga Ascophyllum nodosum (Linnaeus) Le Jolis (AMPEP): A "vaccinelike" effect on Kappaphycus alvarezii (Doty) Doty ex P.C. Silva. Journal of Applied Phycology 24: 427-432. _____________; Reis, R.P. & Critchley, A.T. 2010. In vitro cultivation of three Kappaphycus alvarezii (Rhodophyta, Areschougiaceae) variants (green, red and brown) exposed to a commercial extract of the brown alga Ascophyllum nodosum (Fucaceae, Ochrophyta). Journal of Applied Phycology 22: 101-104. Mendonza, W.G.; Montaño, N.E.; Ganzon-Fortes, E.T. & Villanueva, R.D. 2002 Chemical and gelling profile of ice-ice infected carrageenan from Kappaphycus striatum (Schmitz) Doty “sacol” strain (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 14: 409–418. Ohno, M.; Largo, D.B. & Ikumoto, T. 2004. Growth rate, carrageenan yield and gel 68 properties of cultured kappacarrageenan producing red alga Kappaphycus alvarezii (Doty) Doty in the subtropical waters of Shikoku, Japan. Journal of Applied Phycology 6: 1-5. Parys, S., Kehraus, S.; Pete, R..; Kupper, F.C., Glombitza, K.W. & Konig, G.W .2009. Seasonal variation of polyphenolics in Ascophyllum nodossum (Phaeophyceae). Europena Journal of Phycology 44:331–338 Rayirath, P.; Benkel, B.; Hodges M.D.; Allan-Wojitas, P.; MacKinnon, S.; Critchely A.T. & Prithiviraj B. 2009. Lipophilic components of the brown seaweed, Ascophyllum nodosum, enhance freezing tolerance in Arabidopsis thaliana. Planta 230:135-147 Reitz, S.R. & Trumble, J.T. 1996. Effects of cytokinin-containing seaweed extract on Phaseolus lunatus L.: influence of nutrient availability and apex removal. Botanica Marina 39: 33–38. Yong, Y.S.; Yong, W.T.L. &Anton, A. 2013. Analysis of formulae for determination of seaweed growth rate. Journal of Applied Phycology DOI 10.1007/s10811-013-00227 Yunque, D.A.T.; Tibudos, K.R.; Hurtado, A.Q. & Critchley, A.T. 2011. Optimization of culture conditions for tissue culture production of young plantlets of carrageenophyte Kappaphycus. Journal of Applied Phycology 23: 433-438. 69 7. Conclusões Finais Os resultados apresentados nos quatro capítulos evidenciam a eficiência do extrato em pó comercial da alga marrom A. nodosum (AMPEP) na algicultura de K. alvarezii, que é respaldada pelos seguintes resultados: 1 - In vitro foi constatado que o crescimento e o rendimento de carragenana das algas tratadas com AMPEP (em concentrações de 15-20 gL-1, expostas por 1h) foram maiores do que as não tratadas, assim como eliminou a presença de algumas epífitas; 2 - O extrato de A. nodosum induziu um efeito similar ao de uma vacina em mudas de K. alvarezii, uma vez que, elicitou suas defesas contra agentes estressores; 3 - Após 40 dias de cultivo no mar (tempo de um ciclo de produção de K. alvarezii nos cultivos brasileiros, quando as algas são coletadas de 40 a 50 dias), mudas de K. alvarezii tratadas com AMPEP quando comparadas com as não expostas, apresentaram maiores taxas de crescimento diário e de rendimento de carragenana e sem alteração em sua reologia (força e viscosidade do gel de carragenana); 4 - O AMPEP pode ser usado como um agente remediador em mudas de K. alvarezii afetadas por estresse térmico (baixas temperaturas), por promover o aumento de suas taxa de crescimento diário e força de gel da carragenana; 5 - O uso do extrato de A. nodosum é recomendado em processos de quarentena de mudas para futuro cultivo, no intuito de combater algumas epífitas e tornar as mudas mais resistentes a baixas temperaturas da água do mar e com maior produção de biomassa e 6 - Em protocolos para translocação de mudas de K. alvarezii, o tratamento com AMPEP poderá evitar a introdução de alguns organismos acompanhantes, que podem ser potenciais invasores, além de aumentar a resistência da alga perante o estresse do transporte e aclimatação ao novo ambiente. 70 8. Referências bibliográficas gerais Areces A.J. 1995. Cultivo comercial de carragenófitas del genero Kappaphycus Doty. In: Alveal K., Ferrario M.E., Oliveira E.C. & Sar E. (eds). Manual de Métodos Ficológicos. Universidad de Concepción, Concepción, Chile. Pp 529- 549 Ask, E.l. 2001. Creating sustainable commercial Eucheuma cultivation industry: the importance and necessity of human factor. Proceedings of the 17th International Seaweed Symposium. Chapman ARO, Anderson RJ, Vreeland VJ, Davison IR (eds), Cape Town, Pp.13–18 ________ Azanza, R.V. 2002 Advances in cultivation technology of commercial eucheumatoid species: a review with suggestions for future research. Aquaculture 206:257-277 Barraca, R.T. & Neish, I.C. 1978. A survey of Eucheuma farming practices in TawiTawi. I. The Sitangkai, Sibutu, Tumindao Region. Report Marine Colloids (Phils.) Inc. 65 pp. Barrios, J.E. 2005. Spread of exotic algae Kappaphycus alvarezii (Gigartinales: Rhodophyta) in the northeast region of Venezuela. Boletin do Instituto Oceanografico da Venezezuela 44:29–34. Becker, K.J. & Rotmann K.W.G 1990. A marketing approach to agar. Journal of Applied Phycology 2:105-110. Bixler, H.J. & Porse, H. 2011. A decade of change in the seaweed hydrocolloids industry. Journal of Applied Phycology 23:321–335 Bindu, M.S. 2007. Emplowerment of coastal communities in cultivation of economically important seaweeds of Kerala. Project Report, Science & Society Division, Department of Science & Technology, Government of India, 25 p. ____________ 2009 Empowerment of coastal communities in cultivation and 71 processing of Kappaphycus alvarezii—a case study at Vizhinjam village, Kerala, India. Abstract, 7th Asia-Pacific Conference on Algal Biotechnology, New Delhi, 21 ____________, Kumaraswamy; Achary, G.P. 2006. Experimental field cultivation of Kappaphycus alvarezii (Doty) Doty at VizhinjamcBay, south west coast of Kerala, Abstracts, International Conference on Applied Phycology, Algae in Biotechnology and Environment, New Delhi, 58 Borlongan, I.A.G.; Tibudos, K.R.; Yunque, D.A.T.; Hurtado, A.Q.; Critchley, A.T. 2011. Impact of AMPEP on the growth and occurrence of epiphytic Neosiphonia infestation on two varieties of commercially cultivated Kappaphycus alvarezii grown at different depths in the Philippines. Journal of Applied Phycology 23:615–621 Bulboa, C.R. & Paula, E.J. 2005. Introduction of non-native species of Kappaphycus (Rhodophyta) in sub-tropical waters: comparative analysis on growth rates of Kappaphycus alvarezii and Kappaphycus striatum in vitro and in the seas in southeastern Brazil. Phycological Research 53: 183–188. Doty, M.S. 1984. The production and use of Eucheuma. In: Doty M.S., Caddy J.F. & Santelices B. (eds). Case Studies of Seven Commercial Seaweed Resources. FAO Fish Tech Pap 281: 123 161. ________ & Norris J.N. 1985. Eucheuma species (Solieriaceae, Rhodophyta) that are major sources of carrageenan. In: Abott I.A. & Norris J.N. (eds). Taxonomy of economic seaweeds: with reference to some Pacific and Caribbean species. California Sea Grant College Program, La Jolla, California. pp. 47-61. Castelar, B.; Reis, R.P.; Moura, A.L.; Kirk, R. 2009. Invasive potential of Kappaphycus alvarezii off the south coast of Rio de Janeiro State, Brazil: a contribution to environmentally secure cultivation in the tropics. Botanica Marina 52: 283-289 Chandrasekaran, S.; Nagendran, N.A.; Pandiaraja, D.; Krishnankutty, N. & Kamalakannan, B. 2008. Bioinvasion of Kappaphycus alvarezii on corals in the Gulf of Mannar, India. Current Science 94: 1167–1172. 72 Chapman, V.J. & Chapman, D.J. 1980. Seaweeds and their Uses, 3rd edition. Chapman and Hall, London, 334 pp. Craigie, J.S. 2011. Seaweed extract stimuli in plant science and agriculture Journal of Applied Phycology 23:371 393 Celb, E.B. 2000. History of the use of seaweed, 2nd edition. Flora Celtica. Royal Botanical Garden of Edinburg, 50 pp. Fan, D. 2010. Ascophyllm nodosum extracts improve shelf life and nutritional quality of spinach (Spinacia oleracea L.). Dissertation, Dalhousie University FAO 2010. FAO Yearbook of Aquaculture Production. Disponível em <www.fao.org> Acesso em 11 de outubro de 2011. Góes, H.G., Reis, R.P. 2011. An initial comparison of tubular netting versus tie–tie methods of cultivation for Kappaphycus alvarezii (Rhodophyta, Solieriaceae) on the south coast of Rio de Janeiro State, Brazil. Journal of Applied Phycology 23:607-613 Guiry, M.D. & Blunden, G. 1981. The commercial collection and utilization of seaweeds in Ireland. Proceeding of the International Seaweed Symposium 10:675680 _________, Guiry GM (2011). AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org. Acessado 12 Janeiro 2011 Hayashi, L.; Oliveira, E.C.; Bleicher-Lhonneur, G.; Boulenguer, P.; Pereira, R.T.L.; von Seckendorff, R.;Shimoda, V.T.; Leflamand, A.; Vallée, P. & Critchley, A.T. 2007. The effects of selected cultivation conditions on the carrageenan characteristics of Kappaphycus alvarezii (Rhodophyta, Solieriaceae) in Ubatuba Bay, São Paulo, Brazil. Journal of Applied Phycology 19: 505–511. __________, Hurtado, A.Q;. Msuya F.E.; Bleicher-Lhonneur, G. & Critchley A.T. 2010. 73 A review of Kappaphycus farming: propspects and constraints. In: Israel, A.; Einav, R. & Seckbach, J. (eds) Seaweeds and their role in globally changing environments. Springer Publishing Company. Pp. 255-279. Hong, D.D.; Hien, H.M. & Son PN 2007. Seaweeds from Vietnam used for functional food, medicine and biofertilizer. Journal of Applied Phycology 19:817-826 Hurtado, A. 2003. Farming the seaweed Kappaphycus. SEAFDEC Asian Aquaculture 25:23 _________; Critchley, A.T.; Bleicher-Lhonneur, G. 2006. Occurrence of Polysiphonia epiphytes in Kappaphycus farms at Calaguas Island., Camarines Norte, Philippines. Journal of Applied Phycology 18:301–306 __________, Yunque, D.A. Tibudos, K. & Critchley, A.T. 2009. Use of Acadian marine plant extract powder from Ascophyllum nodosum in tissue culture of Kappaphycus varieties. Journal of Applied Phycology 21:633–639 Jayaraman, J.; Norrie, J. & Punja, Z.K. 2011. Commercial extract from the brown seaweed Ascophyllum nodosum reduces diseases in greenhouse cucumber. Journal of Applied Phycology 23:353–361. Jayaraj, J.; Wan, A.; Rahman, M. & Punja, Z.K. 2008. Seaweed extract reduces foliar fungal diseases on carrots. Crop Protoplasma 27:1360–1366. Jensen, G.I 1990. Present and future needs for algae and algal products. Hydrobiologia 6: 261-266, Khan, W.; Palanisamy, R.; Hankins, S.D.; Critchley, A.T.; Smith, D.L.; Papadopoulos, Y. & Prithiviraj, B. 2008 Ascophyllum nodosum (L.) Le Jolis extract improves root nodulation in alfalfa. Canadian Journal of Plant Science 88:728–728. _______; Rayirath, U.P.; Subramanian, U.P.; Jitesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.C; Craigie, J.S.; Norrie, J. & Prithiviraj, B. 2009. Seaweed Extracts as 74 Biostimulants of Plant Growth and Development. Journal of Plant Growth Regululators 28:386–399. Largo, D.B.; Fukami, K. & Nishijima, T. 1995. Occasional pathogenic bacteria promoting ice–ice disease in the carrageenan-producing red algae Kappaphycus alvarezii and Eucheuma denticulatum (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 7: 545–554. __________; Fukami, K. & Nishijima, T. 1999. Time-dependent attachment mechanisms of bacterial pathogen during ice-ice infection in Kappaphycus alvarezii (Gigartinales, Rhodophyta). Journal of Applied Phycology. 11: 129–136. Loureiro, R.R.; Reis, R.P. & Critchley, A.T. 2010. In vitro cultivation of three Kappaphycus alvarezii (Rhodophyta, Areschougiaceae) variants (green, red and brown) exposed to a commercial extract of the brown alga Ascophyllum nodosum (Fucaceae, Ochrophyta). Journal of Applied Phycology 22:101-104. Lüning, K. & Pang, S. 2003. Mass cultivation of seaweeds: current aspects and approaches. Journal of Applied Phycology 15: 115-119. Marroig, R.G. & Reis, R.P. 2011. Does biofouling influence Kappaphycus alvarezii (Doty) Doty ex Silva farming production in Brazil? Journal of Applied Phycology 23:925–931. Mendonza, W.G.; Montaño, N.E.; Ganzon-Fortes, E.T. & Villanueva, R.D. 2002. Chemical and gelling profile of ice-ice infected carrageenan from Kappaphycus striatum (Schmitz) Doty “sacol” strain (Solieriaceae, Gigartinales, Rhodophyta). Journal of Applied Phycology 14: 409–418. Metting, B.; Rayburn, W.R. & Reynaud, P.A. 1988 Algae and agriculture. In Lembi C and Waaland JR Algae and Human Affairs. Cambridge University Press, New York. Pp. 335-370. McHugh, D.J. 2003 A Guide to the Seaweed Industry. FAO Fisheries Technical Paper. 75 Food and Agriculture Organization of the United Nations, Rome, Pp.105. Msuya, F.E. & Salum, D. 2007 Effect of cultivation duration, seasonality, nutrients, air temperature and rainfall on carrageenan properties and substrata studies of the seaweeds Kappaphycus alvarezii and Eucheuma denticulatum in Zanzibar, Tanzania. WIOMSA/MARG I n° 2007-06. 36. ___________& Kyewalyanga, M.S. 2006 Quality and quantity of phycocolloid carrageenan in the seaweeds Kappaphycus alvarezii and Eucheuma denticulatum as affected by grow out period, seasonality, and nutrient concentration in Zanzibar, Tanzania. Report submitted to Cargill Texturizing Solutions, 46 pp. Paula, E.J.; Pereira, R.T.L. & Ohno, M. 2002 Growth rate of the carrageenophyte Kappaphycus alvarezii (Rhodopyta, Gigartinales) introduced in subtropical waters of Sao Paolo State, Brazil. Phycological Resouch 50: 1–9. Mtorela, M.S.P. 2003. Effect of seagrass cover and mineral content on Kappaphycus and Eucheuma productivity in Zanzibar, West Indian Ocean. Journal of Marine Science 2:163–170. Munoz, J.; Freile-Pelegrin, Y., Robledo, D. 2005. Mariculture of Kappaphycus alvarezii (Rhodophyta, Solieriaceae) color strains in tropical waters of Yucatan, Mexico. Aquaculture 239:161–177. ________& Sahoo, D. 2007. Impact of large scale Kappaphycus alvarezii cultivation in coastal water of India, XIXth International Seaweed Symposium, Kobe, Japan, Program and Abstracts, 121 pp. Pedersén, M.; Collén, J.; Abrahamsson, K. & Ekdahl, A. 1996a. Production of halocarbons from seaweeds: an oxidative stress reaction? Science Marine 60: 257– 263 _________________________________; Mtolera, M.; Semesi, A. & Garcia Reina, G. 1996b. The ice–ice disease and oxidative stress of marine algae, In: Björk M, Semesi 76 AK, Pedersén M, Bergman B (eds.) Current Trends in Marine Botanical Research in the East African Region. Ord & Vetande AB, Uppsala, Sweden, pp 11–24. Pickering, T. 2006. Advances in seaweed aquaculture among Pacific Island countries. Journal of Applied Phycology 18:227-234. __________ & Forbes, A. 2002. The progress of aquaculture development in Fiji. Technical Report, Marine Studies Programme, University of South Pacific 2002/1: 40. Paula, E.J.; Pereira, R.T.L. & Ohno, M. 2002. Growth rate of the carrageenophyte Kappaphycus alvarezii (Rhodopyta, Gigartinales) introduced in subtropical waters of Sao Paolo State, Brazil. Phycological Research 50: 1–9. Parys, S.; Kehraus, S.; Pete, R.; Kupper, F.C.; Glombitza, K.W. & Konig, G.W. 2009 Seasonal variation of polyphenolics in Ascophyllum nodossum (Phaeophyceae). European Journal of Phycology 44:331–338 Potín, P.; Bouarab, K.; Salaün, J.P.; Pohnert, G. & Kloareg, B. 2002. Biotic interactions of marine algae. Current Opinian in Plant Biology 54:308–317. Rebello, J.; Ohno, M.; Ukeda, H. & Sawamura, M. 1997. Agar quality of commercial agarophytes from different geographical origins: Physical and rheological properties. Journal of Applied Phycology 8:517-52. Reis, R.P. 2007. Monitoramento ambiental da alga exótica Kappaphycus alvarezii cultivada comercialmente nas Baías de Sepetiba e da Ilha Grande, RJ. Relatório técnico para o Centro de Pesquisa e Gestão de Recursos Pesqueiros do litoral Sudeste e Sul (CEPSUL – Instituto Chico Mendes de Conservação da Biodiversidade). 70pp. Weinberger, F. 2007. Pathogen-induced defense and innate immunity in macroalgae. Biological Bulletin 213:290–302. ___________& Potin, P. 2010. Red algal defenses in the genomics age. In: Chapman 77 DJ, Seckbach J (eds) Red algae in the genomic age, 1st edn. Springer, New York, Pp 457–472. ____________; Friedlander, M. & Hoppe, H.G. 1999. Oligoagars elicit a physiological response in Gracilaria conferta (Rhodophyta). Journal of Phycology 35:747–755. ____________; Richard, C.; Kloareg, B.; Kashman, Y.; Hoppe, H.G. & Friedlander, M. 2001. Structure activity relationships of oligoagar elicitors toward Gracilaria conferta (Rhodophyta). Journal of Phycology 37:418–426. Potín, P.; Bouarab, K.; Salaün, J.P.; Pohnert, G. & Kloareg, B. 2002. Biotic interactions of marine algae. Current Opinian in Plant Biology 5:308–317. Rodgers, S.K. & Cox, E.F. 1999. Rate of spread of introduced rhodophytes Kappaphycus alvarezii, Kappaphycus striatum, and Gracilaria salicornia and their current distributions in Kane’ohe Bay, O’ahu, Hawai’i. Pacific Science 53: 232–241. Teitelbaum A 2003. Farming seaweed in Kiribati: a practical guide for seaweed farmers. Handbook, SPC, 38. Tseng, C.K. 2001. Algal biotechnology industries and research activities in China. Journal of Applied Phycology 13:375–380. Turan, G. & Neori, A. 2010. Intensive seaweed aquaculture: A potent solution against global warming. In: Israel, A.; Einav, R. & Seckbach, J. (eds) Seaweeds and their role in globally changing environments. Springer Publishing Company. Ugarte, R.; Craigie, J.S. & Critchley, A.T. 2010. Fucoid flora of the rocky intertidal of the Canadian Maritimes: Implications for the future with rapid climate change. In: Israel, A.; Einav, R. & Seckbach, J. (eds) Seaweeds and their role in globally changing environments. Springer Publishing Company. Uyengco, F.R., Saniel, L.S. & Jacinto, G.S. 1981. The ‘ice-ice’ problem in seaweed farming. Procedings of the International Seaweed Sympoisium 10: 625–630. 78 ____________; Chung, C.S.; Hurtado, A.Q.; Soya, F.E.;, Bleicher-Lhonneur, G. & Critchley, A. 2008. Distribution and symptoms of epiphyte infection in major carrageenophyte-producing farms. Journal of Applied Phycology 20:477–483. Vasquez J.A. (1995) Ecological Effects of Brown Seaweed Harvesting. Botanica Marina 38: 251–258. Vauchel, P.; Kaas, R.; Arhaliass, A.; Baron R. & Legrand J. 2008. A New Process for Extracting Alginates from Laminaria digitata: Reactive Extrusion. Food and Bioprocess Technology 1:297-300. Xu, H.; Deckert, R.J. & Garbary, D.J. 2008. Ascophyllum and its symbionts. X. Ultrastructure of the interaction between A. nodosum (Phaeophyceae) and Mycophycias ascophylli (Ascomycetes). Botany 86:185-193. Yunque, D.A.T.; Tibudos, K.R.; Hurtado, A.Q. & Critchley, A.T. 2011. Optimization of culture conditions for tissue culture production of young plantlets of carrageenophyte Kappaphycus. Journal of Applied Phycology 23:433–438. Zemke-White, W.L. & Ohno M. 1999. World seaweed utilisation: An end-of-century summary. Journal of Applied Phycology 11:369-376. ___________ & Smith, J.E. 2006. Environmental impacts of seaweed farming in the tropics, In: A.T. Critchley, M. Ohno and D. Largo (eds.) World Seaweed Resources. Expert Centre for Taxonomic Identification (ETI), University of Amsterdam (CDROM series). 79
Baixar