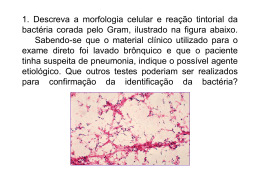
Faculdade de Medicina da Universidade do Porto Microbiologia 2006/2007 ANATOMIA BACTERIANA 3 de Outubro de 2006 Aula leccionada por Doutor Acácio Gonçalves Rodrigues SUMÁRIO DA AULA T2 Aspectos ultrastruturais e bioquímicos dos vários componentes da célula bacteriana. Cápsula, camada mucóide ou glicocálice. Parede celular: mucopeptídeo e componentes especiais nas bactérias Gram positivo e Gram negativo. Degradação da parede celular: protoplastos, esferoplastos e formas L. Membrana citoplasmática: ultrastrutura e composição bioquímica. Compartimento intracelular. Flagelos. Fímbrias. Esporo bacteriano. Metabolismo energético e assimilação bacteriana. Análise cinética do crescimento e morte dos microrganismos (Tópico da aula T3 abordado nesta aula). Nesta aula apresentamos todos os assuntos abordados na teórica, complementados com as imagens e quadros mais relevantes da aula. Colocamos informação adicional do livro (Murray) e da sebenta de microbiologia (letra verdana). Apesar da aula ser muito extensa não se assustem com o tamanho. A aula tem muitas imagens!! Se detectarem qualquer erro agradecíamos que nos informassem: [email protected] ou [email protected]. Bom estudo e boa sorte para as frequências!! Susana Sá Tânia Borges Turma 13 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Hoje é uma aula comprida e é uma aula em que eu vos vou chamar à atenção daquilo que considero os aspectos essenciais de uma matéria bastante longa que tem a ver com a citologia da bactéria na sua constituição, da sua anatomia, da sua fisiologia ou metabolismo. Vamos lá à anatomia da bactéria. CONSTITUIÇÃO DA BACTÉRIA ⇒ Nucleóide. ⇒ Algumas inclusões. ⇒ Ribossomas. Ela tem que ser capaz de efectuar a síntese proteica. Estes ribossomas são diferentes dos das células eucarióticas: subunidades e coeficiente de sedimentação de 70s. ⇒ Plasmídeos. Vamos abordar isto mais tarde e com mais detalhe na próxima aula sobre genética microbacteriana. Mas são fragmentos de material genético, de DNA, que podem estar extra ou intracromossómicos, neste caso estão extracromossómicos. ⇒ Membrana celular. É o principal órgão metabólico da bactéria, é o sítio onde se dá a respiração celular. ⇒ Parede celular. Aqui chama muito à atenção, o destaque deste peptidoglicano. ⇒ Está aqui também representado um flagelo, que é um órgão de locomoção. Passando por alto rapidamente, sobre as estruturas intracitoplasmáticas, temos o tal cromossoma nucleóide, RNA mensageiro, obviamente ribossomas e proteínas. Proteínas estas que têm funções diversas. Acabaram de ser sintetizadas e estão em trânsito até chegarem à parede ou até chegarem à membrana. Existem sempre algumas proteínas presentes num substrato solúvel. Obviamente que o principal constituinte intracitoplasmático da bactéria é a água (80 a 90% do peso seco de uma bactéria). E depois metabolitos e inclusões. Estas inclusões podem ser por exemplo inclusões lipídicas, que servem como substrato energético; inclusões de carbohidratos, nomeadamente por exemplo o glicogénio; inclusões de fosfatos. Quando algumas bactérias cresciam em meios com fósforo incluíam esse fósforo no seu interior e formavam grânulos de fósforo e que depois deram no passado origem a métodos e descrições muito fantasiosas: Para que é que serviam estas inclusões, por exemplo? E que mais tarde se veio a verificar, em microscopia electrónica, que eram inclusões de artefacto, talvez pelo método de fixação das bactérias. ANATOMIA BACTERIANA 2/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO E agora vamos do interior para o exterior, para os invólucros bacterianos. Vamos começar pela membrana citoplasmática e parede citoplasmática. Ora bom, nós estamos aqui em microbiologia médica a falar nas tais eubactérias, não é nas arqueobactérias. E em todas as bactérias (eubactérias), com excepção dos micoplasmas e de alguns homófilos que ainda podem ter importância em microbiologia médica, quase todas essas bactérias têm uma parede citoplasmática. Membrana, obviamente que todos têm de ter membrana. Sem membrana não há limite celular. MEMBRANA Este é o modelo clássico de Singer Nicholson que também não é novo para vocês: ⇒ Uma bicamada de fosfolipídeos. Os fosfolípidos são polares, com uma extremidade hidrofílica e uma extremidade hidrofóbica (voltada para o interior); ⇒ Proteínas (proteínas periféricas intrínsecas-transmembranares). ou extrínsecas e proteínas integrais ou Os fosfolípidos são parecidos com os das membranas celulares dos eucariotas, mas têm alguma diferença em termos de constituição: têm mais fosfatidilglicerol e mais fosfatidiletanolamina. De um modo geral, as membranas celulares de uma bactéria, têm também um teor proteico maior do que têm as membranas dos eucariotas, porque estas proteínas têm que efectuar muitas funções metabólicas, por exemplo: ⇒ Produção de energia; ⇒ Respiração, que nos eucariotas, nas células mais evoluídas, já são feitas por organelos especializados. Portanto em peso seco comparativamente a um eucariota, uma bactéria tem mais proteínas na sua membrana. Funções As funções são muitas: ⇒ Permeabilidade; ⇒ Transporte de iões e substâncias dos mais diversos tipos que a bactéria precisa; ⇒ Transporte de electrões e fosforilação oxidativa; ⇒ Excreção de exoenzimas e de outras proteínas; ⇒ Funções biossintéticas das mais variadas, inclusivamente vamos ver que parte da síntese da parede celular é feita a um passo importante que só ocorre a nível membranar; ⇒ Funções de quimiotaxia (despertando a resposta pelo organismo que está a ser agredido). Transporte Ora bom, de uma forma muito simplificada, como é que se faz o transporte através destas membranas? Ou se faz por difusão passiva ou se faz por transporte activo. Um modelo muito recente sobre fisiologia da membrana, baseia-se também no transporte através destas proteínas, mas segundo 3 modelos: ⇒ Modelo uniporte: passa uma única substância num só sentido; ⇒ Modelo simporte: passam 2 substâncias no mesmo sentido (uma digamos a reboque); ⇒ Modelo antiporte: passam 2 substâncias mas em direcções opostas. ANATOMIA BACTERIANA 3/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Segundo este modelo, a mesma proteína pode funcionar apenas de 1 destas 3 maneiras; de 2 destas 3 maneiras; ou ainda das 3 maneiras em simultâneo (não é bem em simultâneo, mas é pouco tempo após). A membrana é o grande órgão metabólico da bactéria. PAREDE Temos aqui 2 tipos de parede. Gram positivo Aqui temos sobretudo uma substância que é o peptidoglicano. E cada uma destas “linhas” representa uma camada e como vocês estão a ver, estão aqui várias camadas. Esta parede é essencialmente constituída por peptidoglicano. Peptidoglicano, mucopeptídeo, mureína, são três termos que são usados indistintamente nos vários livros, para designar a mesma coisa. Aparecem aqui também representados estes ácidos teicóicos. E estes ácidos basicamente são lipídeos, têm glicerol ou ribitol, e podem ser classificados em dois tipos: ⇒ Ácidos teicóicos da Parede celular unidos por ligações covalentes ao peptidoglicano; ⇒ Ácidos teicóicos da Membrana celular (ácido lipoteicóico) unidos por ligações covalentes aos glicolípidos da membrana celular. Mas o principal constituinte é o peptidoglicano. Gram negativo Aqui temos uma membrana citoplasmática que é igual à das gram positivo, mas já agora devo dizer-vos, que em ultraestrutura, em microscopia electónica (e isto era uma coisa que classicamente era considerado muito importante, mas ninguém sabia dizer porquê), a membrana citoplasmática da bactéria gram positiva tem um perfil assimétrico (o folheto externo é mais espesso), enquanto que a membrana da gram negativo tem um perfil simétrico. Isto hoje em dia tem muito pouca relevância. ANATOMIA BACTERIANA 4/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Constituição: ⇒ O peptidoglicano, resume-se a uma ou duas camadas. Não quer dizer que seja exactamente uma ou duas, mas é muito pouco. ⇒ Temos aqui um espaço – espaço periplasmático – que não é virtual, mas é um espaço laxo digamos assim, pouco denso, comparativamente a este e está preenchido sobretudo por proteínas que vocês estão aqui a ver, tem aqui algum fluido e existem aqui alguns nutrientes solúveis. ⇒ Depois existe aqui outra vez uma membrana, que se chama outer membrane (tradução para português é membrana externa). Não é semelhante à membrana citoplasmática, porque no folheto interno ela tem essencialmente fosfolípidos, mas no folheto externo tem uma coisa que se chama lipopolissacáridos – tem lipídeos, mas a esses lipídeos estão ligados carbohidratos. ⇒ Também não é sempre assim, podem haver zonas da membrana do gram negativo, quer no folheto interno, quer no folheto externo onde existam fosfolípidos. Aliás, há bactérias gram negativa que não têm, ou têm muito pouco, lipopolissacáridos e têm no folheto externo sobretudo fosfolípidos. Mas este é um padrão clássico. ⇒ Também aqui nesta membana externa (outer membrane) existem proteínas longas que formam poros, as chamadas porinas. Obviamente que são sistemas de transporte através desta membrana externa. PEPTIDOGLICANO O peptidoglicano é constituído acima de tudo por dois radicais: N-acetil-glicosamina e ácido N-acetil-muramico, ligados por ligações β-1,4. Tem alguns radicais de ácidos aminados, que são depois quebrados. Ele é sintetizado nestas unidades e é montado assim na parede da bactéria, de modo a formar quase que uma macromolécula única. A bactéria gram negativa tem muito pouco peptidoglicano, por isso vamos focar agora a atenção na bactéria gram positiva. Esta é envolvida, quer seja um coco, quer seja um bacilo, quase como uma macromolécula única, porque isto é montado com unidades repetitivas. É quebrado aqui (ver figura 3.7), são feitas estas ligações, estes cross-linkings com outros radicais e iões e fica uma camada única à volta da bactéria. A ligação β1,4 é quebrada pela lisozima. ANATOMIA BACTERIANA 5/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Como é que se faz a síntese da parede, e do peptidoglicano? Começa no interior do citoplasma, na fase citoplasmática e pela formação do UDPácido N-acetil-muramico e N-acetil-glicosamina. Vem até à membrana citoplasmática. Aqui liga-se a um carregador, que é o undecaprenol, dito de uma forma muito simplificada. Atravessa a membrana citoplasmática, e é já na membrana externa da membrana citoplasmática, que são montados os resíduos, dando-se aquela quebra e o cross-linking. Portanto fica depois tudo isto montado como se fosse umas placas de betão armado, qualquer coisa assim desse género, que surge hoje nas engenharias. Materiais que são pré-fabricados e que depois são montados assim ali e ligados de uma forma contínua no exterior. Obviamente, devo-vos dizer que existem, e isto está muito bem caracterizado nas grampositivo, autolisinas que também degradam o excesso de mucopeptídeo. É o ANATOMIA BACTERIANA 6/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO mucopeptídeo, a parede, que vai dar forma à bactéria, portanto ela para ficar redonda, ou para ficar mais alongada, tem que ser degradada nalguns pontos. O que é fabuloso em termos de aspectos genéricos da síntese desta parede, é que a bactéria não acumula percursores, a bactéria não tem reservatórios por exemplo de ácido N-acetil-murânico ou N-acetil-glicosamina. Ela começa a sintetizá-los logo que precisa deles, mas a síntese acaba por ser contínua, porque também a parede está continuamente a degradar-se, aliás como a membrana, porque está a ser constantemente atacada. Quando o metabolismo energético da bactéria não lhe permite acudir a tantos pontos de agressão e começa a ter problemas de recuperação da parede e sobretudo da membrana, a bactéria acaba por morrer. É o peptidoglicano que confere rigidez estrutural à bactéria gram positivo. É uma substância que se torna insolúvel, quando são retirados aqueles resíduos de ácidos aminados. Enquanto está na fase citoplasmática e na fase membranar é hidrossolúvel, mas depois torna-se insolúvel na água. Estes ácidos teicóicos, obviamente que são solúveis, são hidrossolúveis. Não conferem propriamente rigidez, funcionam muitas vezes como antigénios, têm funções ainda muito pouco esclarecidas. Resumo da biossíntese do peptidoglicano: ⇒ Fase citoplasmática: o Produção dos percursores peptídicos ⇒ Fase membranar o Transporte daquelas substâncias citoplasmática ⇒ Fase parietal o Interligações interpeptídicas através da membrana Vocês têm o hábito de dizer que a bactéria gram negativo tem uma parede menos espessa que a bactéria gram positivo, e isso não é verdade! Ela (gram negativo) tem menos mucopeptídeo, mas a espessura não tem só a ver com o peptidoglicano ou mucopeptídeo. Ela tem muito pouco peptidoglicano, mas tem um espaço periplasmático, que aliás o próprio peptidoglicano já está contido no espaço periplasmático, como tem proteínas e depois tem uma membrana externa, constituída por dois folhetos, e que sobretudo nas enterobacteriácea têm uma espessura grande, porque têm lipopolissacáridos bastante longos. Devo-vos dizer que estes lipopolissacáridos, este LPS, é um dos principais mecanismos de patogenicidade das bactérias. E é um mecanismo de patogenicidade porquê? De um modo geral, ele é genericamente igual em todas as bactérias de gram negativo, obviamente existem algumas especificidades nalgumas espécies e nalguns casos concretos, mas disperta por parte do hospedeiro basicamente o mesmo tipo de reacção: ⇒ Ele interfere com a cascata de coagulação; ⇒ Activa a cascata do complemento; ⇒ Causa leucopenia. Liberta-se sobretudo quando as bactérias se estão a multiplicar rapidamente, ou então liberta-se em grande quantidade quando as bactérias são lisadas. E é por isso que quem tem sépsis, uma infecção sistémica por um bacilo de gram negativo, tem febre, tem leucopenia (por leucopenia, não é por toxicidade medular directa), mas tem por exemplo um síndrome de coagulação intravascular disseminada, por acção deste lipopolissacárido. ANATOMIA BACTERIANA 7/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Constituição do lipopolissacárido: ANATOMIA BACTERIANA 8/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO O lipopolissacárido (LPS) é constituído por várias partes. A parte mais interna é o lipídeo A, reparem que não é exactamente igual aos fosfolípidos. E agora aqui vê-se mais um detalhe daquilo que eu já disse: a membrana externa tem um folheto interno, que são fosfolípidos semelhantes aos da membrana, e que pensa-se que migram e podem substituir rapidamente a partir da membrana citoplasmática. Eles podem sair daqui intercalando-se ali. Mas na membrana externa, sobretudo nas enterobacteriáceas, o LPS é constituído por este lipídeo A e depois tem a porção polissacarídica. Este lipídeo A considera-se que é do LPS, a fracção mais responsabilizada por estas acções que eu vos estive a dizer: toxicidade sobre o endotélio, sobre o complemento, de activação do sistema das cinases. Mas depois tenho aqui duas substâncias bastante estranhas, que não estão presentes em lado nenhuma membrana, que é ácido desoxioctanóico e esta heptose que constituem o chamado inner core. E depois tem vários radicais de carbohidratos. Isto vão funcionar como antigénios, quando nós falarmos mais tarde nas enterobacteriáceas e falarmos na tipagem das enterobacteriáceas, vão ver que os antigénios que são utilizados para a tipagem são estes. O que é importante vocês reterem a partir daqui? Se a bactéria perder o outer core do seu lipopolissacárido sobrevive, aliás há bactérias de gram negativo que não têm este outer core. Agora se a bactéria perder este ácido desoxioctanóico, lipídeo A, se perder o inner core já não sobrevive. Portanto, mas como estão a ver, esta parede celular (da gram negativa), é uma parede celular bastante espessa. Não se esqueçam disto! A membrana citoplasmática tem cerca de 8 nm, é igual nas duas; a camada de peptidoglicano tem 15 a 80 nm numa bactéria gram positivo (representa entre 50 a 90% do peso seco da parede); enquanto que uma bactéria gram negativo são 3 nm. Mas depois tem aqui este folheto externo que ocupa cerca de 8nm. Queria chamar a atenção do seguinte: há pouco mostrei aquelas ligações β1,4, onde actuava a lisozima. Isto tem interesse porquê? A lisozima é uma enzima que existe nas lágrimas, na saliva, na clara do ovo, nas nossas excreções, e essa enzima é capaz de quebrar essas ligações. Ora bom, quando a lisozima actua sobre a parede celular de uma bactéria de gram positivo, dá-se a lise dessa parede e forma-se um protoplasto. Mas esta lisozima, não é capaz de atingir esta camada de peptidoglicano da bactéria de gram negativo, por causa desta membrana externa, por causa daqueles fosfolípidos. Então se eu fizer actuar lisozima, mas juntamente um agente quelante de iões, por exemplo o EDTA, e um tampão Tris, estes vão favorecer a entrada através desta camada acima de tudo lipídica. A lisozima atinge o peptidoglicano da parede da bactéria de gram negativo, degrada e portanto a bactéria fica com uma forma esférica, mas passa a chamar-se não protoblasto, mas esferoblasto. Isto porque a lisozima não faz desaparecer a membrana externa, e portanto fica uma célula sem peptidoglicano, mas rodeada de membrana externa, acima de tudo constituída por proteínas e lipídeos. ANATOMIA BACTERIANA 9/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Portanto, sintetizando, qual é a grande diferença, de uma forma muito simplista, em termos de parede entre uma bactéria gram positiva e uma bactéria gram negativa? É o teor de peptidoglicano, que é muito menor na bactéria gram negativa e o teor de lipídeos que é muito maior nas bactérias gram negativa. Excepções a isto? Nem todas as bactérias são assim, e insisto mais uma vez, não estamos aqui a falar de arqueobactérias, estamos a falar de eubactérias. Mas há bactérias muito importantes em patologia humana, que eu destaco o mycobacterium, mycobacterium Leprae, mycobacterium tuberculosis (agente da tuberculose), que muito embora tendo ultra-estruturalmente, a parede semelhante a uma parede de gram positiva (tem bastante peptidoglicano), tem no seu exterior uma camada de lipídeos completamente diferentes daqueles que nós vimos na bactéria de gram negativo. Ele tem no seu exterior, acima de tudo, lípidos de alto peso molecular, que são estes ácidos micólicos; Ele tem como que “bolas de cera” no seu exterior e é isto que o torna completamente insensível aos métodos de coloração, que são utilizados para corar a maior parte das bactérias. Este tipo de bactéria não cora pelo método de gram, porque os corantes não conseguem penetrar aqueles lipídeos, aquelas ceras. A síntese destes lipídeos é bastante complexa, é bastante demorada, demora muitas horas. Eu disse-vos que uma Escherichia Coli se dividia em 20 minutos e uma bactéria destas divide-se em várias horas. Depois havemos de ver mais tarde, que a resposta imunológica para estas bactérias é completamente distinta daquela que é genericamente semelhante para a grande maioria das bactérias que nós vamos estudar. Pensa-se que essa resposta imunológica, que tem a ver com este tipo de lipídeos, concretamente com estes ácidos micólicos. Depois havemos de ver, mais adiante, com mais detalhe, o mycobacterium, a corar muito fracamente com o método Ziehl-Neelsen. Como estão a ver aqui nesta imagem o mycobacterium, apesar de serem bacilos, têm uma forma irregular, que tem a ver com os ácidos micólicos que têm. Relevância prática da parede: ⇒ É a parede, que independentemente de ser gram positiva, ou gram negativa, dará forma à bactéria. O facto de haver cocos, de haver bacilos alongados, de haver bactérias espiraladas ou fusiformes, isto tem a ver com a parede. A parede é que confere forma e rigidez estrutural à bactéria, não é a membrana. ⇒ E por outro lado é a parede e a sua riqueza ou deficiência em peptidoglicano, que vai levar a que as bactérias corem de maneira diferente no método de Gram. Isso vai ser abordado com detalhe nas aulas práticas, mas isto tem a ver com a constituição da parede e no caso daquelas excepções, aquelas que têm ácidos micólicos, não coram pelo método de Gram. ANATOMIA BACTERIANA 10/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO ANATOMIA BACTERIANA 11/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO OUTRAS ESTRUTURAS EXTERNAS QUE PODEM EXISTIR Pili Flagelos Pili ou Fímbrias: ⇒ São pequenas projecções proteicas (proteína classicamente chamada de pilina) que não são visíveis em microscopia óptica, só em microscopia electrónica. ⇒ São muito importantes em termos de adesão. Não uma adesão inespecífica, mas uma adesão a receptores. Primeiro conheceram-se os chamados F pili ou pili de fertilidade, pili factores sexuais. Na próxima aula vamos falar sobre trocas de material genético e vamos ver que as bactérias podem contactar entre elas e transferir material genético, através desses pili. Tem a ver com a adesão das bactérias, há um pili que funciona como um factor de fertilidade masculino e portanto essa foi a primeira adesão que foi conhecida por estes pili. Mas eles são muito importantes para aderir por exemplo a células do epitélio, que têm determinados receptores, determinadas leptinas e isto por exemplo, está muito bem caracterizado para algumas enterobacteriáceas, alguns bacilos de gram negativo, que são agentes importantes de infecção urinária. ⇒ Quem tem o azar de ter uma infecção urinária baixa, uma cistite, por uma bactéria que tem o mesmo tipo de adesinas e tem uma boa interacção específica com as proteínas que existem à superfície do epitélio, tem o azar de facilmente ver essa cistite, essa infecção urinária baixa, transformada numa pielonefrite. A bactéria ascende pelo ureter e atinge o rim. Precisamente por isso, não é só porque ela se multiplica rapidamente ou que seja móvel. Uma bactéria para infectar a mesa tem primeiro que colonizar a mesa. Por aqui passam fluidos, vento e, portanto, a bactéria terá que se “agarrar” à mesa. Imaginem a árvore brônquica: a bactéria para chegar aos pulmões tem que se “agarrar” aos brônquios caso contrário será facilmente eliminada. A mesma coisa acontece num endotélio de um vaso. Tem que haver uma adesão que lhe permita permanecer ali algum tempo para depois se replicar. É um processo muito importante para garantir a capacidade de colonização e multiplicação da bactéria. Os cílios distribuem-se uniformemente à volta da célula. Os flagelos são: ⇒ Órgãos ⇒ ⇒ ⇒ ⇒ ⇒ ANATOMIA BACTERIANA muito importantes na mobilidade; São muito maiores que os cílios; São projecções para o exterior da célula; Estão ancorados na membrana citoplasmática; Precisam de muita energia, tendo um aparelho energético próprio; São de natureza proteica (flagelina). 12/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO A bactéria Gram – tem um flagelo obviamente muito mais complexo, porque estando só ancorado na membrana (não tendo o suporte do peptidoglicano), o flagelo é facilmente removido. Portanto, tem vários anéis que o fixam, quer no peptidoglicano (que é pequeno relativamente à bactéria Gram +), quer na membrana. O flagelo está ligado a uma região denominada gancho que, por sua vez, se liga ao corpo basal, que está ancorado na membrana citoplasmática e parede celular. O corpo basal consiste num sistema de anéis e de um eixo central que passa através dos anéis. Desta forma, o flagelo insere-se de forma diferente consoante a bactéria seja Gram + (um par de anéis no corpo basal) e Gram – (dois pares de anéis no corpo basal). Os flagelos podem ser visualizados directamente por microscopia óptica por colorações especiais (técnica de Laisson em que os corantes acrescentam alguma espessura ao flagelo). São grandes e portanto são passíveis de serem visualizados em microscopia óptica ou, obviamente, em microscopia electrónica. As bactérias classificam-se quanto à distribuição dos flagelos: ⇒ Monótricas – um flagelo polar; ⇒ Anfítricas – um flagelo em cada pólo; ⇒ Lofótricas – tufos ou grupos de flagelos polares; ⇒ Perítricas – flagelos distribuídos à volta da bactéria. A distribuição dos flagelos vai influenciar a mobilidade da bactéria. ANATOMIA BACTERIANA 13/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO ESTRUTURA EXTERNA Cápsula [polissacarídica, polipeptídica (bacillus anthracis)] Slime Layer Glicocálice S-layer CÁPSULA De modo geral a cápsula é de natureza polissacarídica (carboidratos) mas existem algumas excepções, como a cápsula do bacillus anthracis (tem uma cápsula polipeptídica de ácido glutâmico). A cápsula pode ser visualizada quer positivamente (corando-a por um método que cora a cápsula), quer negativamente (corando tudo o que está à volta ficando um halo) pelo método de tinta-da-china. O que é que estão a ver aqui? Nem todas as bactérias têm cápsula. Raros fungos têm cápsula. Estão aqui a ver um fungo que tem uma cápsula bastante espessa. Há bactérias que tipicamente são capsulares e podem perder a cápsula. As bactérias quando estão a crescer num meio de cultura numa placa de petri formam colónias e essas bactérias têm, de modo geral, uma cápsula lisa com um aspecto mucóide (“parece aquele açúcar com que se fazem bolos”). Quando ficam com um aspecto mais baço, quando perdem aquele brilho, dizemos que a bactéria fica com um aspecto rugoso perdeu a cápsula. Então pelo aspecto das colónias conseguimos prever se as bactérias são ou não capsulares. Mas para que é que serve a cápsula? Sob o ponto de vista primário da sobrevivência da bactéria, a cápsula acumula muita água e portanto, a cápsula é uma boa protecção contra a desidratação. Tem carboidratos e como vocês sabem os carboidratos atraem e fixam a água. A cápsula é um mecanismo de patogenicidade bacteriana ou fúngica, conforme o caso. A cápsula impede a fagocitose. Os neutrófilos e os bacteriófagos não conseguem fagocitar uma bactéria que tenha cápsula, a não ser que a cápsula esteja recoberta por anticorpos (ou seja, que esteja opsonizada). A opsonização ocorre no baço. Importância prática disso? Um doente que tenha sido esplenectomizado (remoção do baço por causas médicas/ traumatismos abdominais devido a acidentes de viação/ fractura do baço) deve receber periodicamente (todos os anos, no início do Inverno) uma vacina anti-bactérias capsuladas (streptococcus pneumoniae). Eu já vi um politraumatrizado ter sobrevivido muitos meses a longas complicações e depois morreu com uma sepsis fulminante provocada por uma bactéria capsulada (como o streptococcus pneumoniae). Portanto, um dos mecanismos mais importantes de patogenicidade bacteriana é a cápsula. Para além de carboidratos têm, em alguns casos (as bactérias Gram + e Gram – não são as únicas), polissacarídeos com especificidade antigénica. Assim servem como antigénios, podendo ser utilizados para a produção de anticorpos que identificam e detêm as bactérias. Evidentemente se tivermos um anticorpo contra um determinado antigénio que é específico, por exemplo, do streptococcus pneumoniae, eu já nem preciso de cultivar a bactéria e esperar longas horas e longos dias. Eu encontro especificamente aquela bactéria com esta técnica e podemos até identificar vários tipos de bactérias. ANATOMIA BACTERIANA 14/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO SLIME LAYER E GLICOCÁLICE A slime layer e o glicocálice têm carboidratos e polissacarídeos extra-parede. Enquanto que a cápsula está muito bem organizada acompanhando a forma da bactéria, a slime layer é um mucopolissacarídeo muito difuso (mais espesso aqui, mais espesso ali). Tem mucopolissacarídeos no exterior da parede mas que não apresentam uma forma muito organizada. Para que é que isto serve? Têm algumas propriedades anti-fagocíticas não tão consistentes como a da cápsula e também favorecem a adesão. Algumas células podem ser atingidas por microbianos. O glicocálice tem projecções melhor organizadas. Estão dispostas à volta da bactéria mas estão de uma forma totalmente organizada. Contudo, há pontos da parede que não há glicocálice. Concluindo, é exactamente do mesmo material da slime layer mas está disposto de uma forma melhor organizada. Eu não sei até que ponto não será, em alguns casos, artefactos resultantes da coloração e do tratamento bioquímico a que as células são submetidas para serem visualizadas em microscopia electrónica. Acredita-se que este glicocálice, como tem uma forma de projecção mais consistente, não está tão desorganizado como a slime layer, mas não é tão uniforme como é a cápsula. S-LAYER A S-layer é visível e é de natureza proteica. Tem uma camada muito fininha periódica que está onde não se devia encontrar. Não digam que é uma cápsula proteica mas sim uma camada fininha de revestimento de natureza periódica que está à superfície de determinadas bactérias. ESPOROS Na última aula falamos das diferenças entre uma bactéria, um fungo e uma célula eucariótica. O fungo reproduz-se sempre por esporos e as bactérias reproduzem-se por divisão binária. Mas há bactérias que produzem esporos. Contudo, as bactérias que produzem esporos não os produzem para se reproduzir mas sim para assegurar a sobrevivência da espécie. A bactéria divide-se sempre por divisão binária! Bactérias que produzem esporos: Alguns bacilos Gram + : géneros Bacillus e Clostridium; Cocos Gram + : Sporosarcina ureae (não é patogénica); Rickettsiae: Coxiella burnetti (agente da febre Q). Algumas bactérias Gram + têm na parede coxielas e vão corar como uma bactéria Gram – . Têm uma forma cocoide e produzem uma coisa semelhante a esporos. Estas bactérias apesar de corarem Gram – têm DNA e biologia molecular característicos de uma bactéria Gram +. De modo geral, só alguns bacilos Gram + produzem esporos. E este esporo é uma célula que está perfeitamente quiescente. ANATOMIA BACTERIANA 15/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Forma-se em condições adversas: Ambiente com falta de nutrientes; Desidratação; Alterações térmicas – não podem ser muito súbitas, porque se não a célula morre. É preciso que a célula tenha tempo para esporolar, processo que demora algumas horas (7-8 horas dependendo do tipo de bactéria). Durante este período, a célula tem que suportar estas condições adversas. Os esporos podem permanecer quiescente horas, dias, dezenas, centenas, milhares de anos! E ficam perfeitamente quiescentes sem qualquer função metabólica (permanecendo viáveis). Notas: ⇒ Cerca de 15% do peso seco do esporo é constituído por ácido dipicolínico complexado com iões cálcio, localizados no core; ⇒ Termo-resistência devido a: o Estabilização do DNA pela presença de cálcio-dipicolinato e proteínas solúveis em ácido; o Desidratação do protoblasto; o Presença do coat; o Enzimas capazes de reparar o DNA; o Grande estabilidade das proteínas celulares; o Adaptadas ao crescimento a elevadas temperaturas. ⇒ As cápsulas e as slimes não são necessárias para o crescimento da bactéria mas são muito importantes para a sobrevivência no hospedeiro. A cápsula constitui uma barreira a moléculas hidrofóbicas. Quando esta célula encontrar condições para que possa esporolar/ germinar (a célula tem sensores), isto é, voltar à forma vegetativa, ela quebra a parede do esporo e desenvolve-se tal como ela era, tendo as mesmas características genéticas. Esquema de um esporo ANATOMIA BACTERIANA 16/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Constituição do esporo: ⇒ Core – protoblasto do esporo contendo DNA e os restantes elementos da bactéria; zona central, “coração”; ⇒ Membrana interna ⇒ Parede do esporo – formada por peptidoglicano normal que irá originar a futura parede celular da célula vegetativa; camada interna da bactéria; ⇒ Córtex – camada mais espessa constituída por um tipo especial de peptidoglicano com menos peptídeos “crosslinks” (são ligações finais que vão ser quebradas na célula vegetativa); é muito sensível à lisozima e às autolisinas; ⇒ Membrana externa ⇒ Protein “coat” – formada por uma proteína “keratin-like”; impermeável e confere aos esporos a sua relativa resistência aos agentes químicos; ⇒ Exosporo – membrana lipoproteica contendo também carboidrato. Como se forma o esporo? Forma-se naturalmente no interior de uma bactéria. Forma-se a partir de uma extremidade para onde é transferido material genético idêntico ao da célula mãe, obviamente condensado. São transferidos ribossomas e quase tudo que existe na célula mãe mas de uma forma extremamente condensada. Vai invaginar na parede e na membrana da célula mãe. Formação do Esporo 1. Duplicação do DNA; 2. Uma cópia do DNA e conteúdo citoplasmático é envolvido pela membrana citoplasmática, pelo peptidoglicano e pela membrana do septo; 3. As duas camadas estão envolvidas pelo córtex, que é constituído pela membrana interna do peptidoglicano “crosslinker” e uma camada externa de peptidoglicano; 4. O cortéx é envolvido por uma camada externa formada por uma proteína “keratin-like” (coat); 5. Libertação do esporo. ANATOMIA BACTERIANA 17/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Porque é que a célula-mãe não resiste ao calor, não resiste ao frio, não resiste à desidratação e o esporo é capaz de resistir? A célula-mãe tem também uma parede celular! Contudo, no esporo, a parede sofre transformações. O invólucro torna-se mais espesso, resultando da sobreposição de duas membranas celulares e de duas paredes celulares, tendo muito peptidoglicano. Esta parede é enriquecida com ácido dipicolínico e este ácido confere uma termo-resistência e uma resistência mecânica às células. No interior do esporo visualizam-se muitas inclusões de glicogénio que lhe vão permitir, após a germinação, sobreviver nas primeiras horas. Germinação do esporo ⇒ Activação – processo reversível e que pode resultar de vários tratamentos como a acção do calor; ⇒ Germinação – é o despertar do esporo adormecido. Há um aumento do volume do esporo, ruptura da parede externa, perda das resistências ao calor, perda da refractibilidade, libertação de componentes do esporo e aumento da actividade metabólica; ⇒ Crescimento – o protobasto produz novos componentes e emerge do resto da camada externa (coat), desenvolvendo-se em seguida como uma bactéria activa. Posição do esporo na célula vegetatica No interior da forma vegetativa mas que não deformam o corpo da bactéria Deformam o corpo bacteriano Para além desta resistência química, a resistência física/ rigidez deste esporo é tão grande que é extremamente difícil obter um corte de um esporo para microscopia electrónica. Tal como os tecidos, as bactérias têm que ser aglutinadas, têm que ser incluídas em Epon e cortada com uma lâmina de diamante. Este esporo é tão rígido que quando a lâmina de diamante toca na parede do esporo, muitas vezes ele salta e libertase do Epon. ANATOMIA BACTERIANA 18/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO METABOLISMO ENERGÉTICO BACTERIANO A cinética do crescimento e a morte dos microorganismos MUITO IMPORTANTE PARA O PROFESSOR! ☺ A bactéria precisa de O2, fonte de carbono, azoto, sais minerais (cobre, potássio, magnésio, cálcio, ferro, sódio e cloro) e alguns micronutrientes. Não vou estar a descrever ao pormenor as vias de catabolismo e anabolismo mas quero que tenham presente o seguinte: as bactérias não se podem “dar ao luxo” de ter uma função metabólica que não precisam. De um modo geral, a bactéria está preparada para metabolizar em primeiro lugar carboidratos metaboliza primeiro a glicose porque é muito mais difícil estar a degradar a frutose e a lactose pois são carboidratos mais complexos. Contudo, ela é capaz de degradar muitos carboidratos diferentes. A bactéria sofre um fenómeno a que se chama repressão metabólica, isto é, tem muitas das suas vias metabólicas reprimidas porque não necessita delas. A bactéria produz pouca energia para aquilo que precisa de fazer: sintetizar substâncias, reparar a membrana, a parede celular, a cápsula e as suas proteínas. Portanto, de um modo geral, a bactéria obtém energia e consegue converter tudo através do ciclo de Krebs: quer a partir de proteínas, quer a partir de polissacáridos, quer a partir de lípidos… Reparem na importância do piruvato e da acetil-CoA. Não vou falar destas vias mas façam o favor de as ler e compreender (ver em anexo imagens sobre o tema). É assim que a bactéria consegue sintetizar outros aminoácidos que são essenciais (através do ciclo de Krebs) e que não os obtém a partir do meio. ANÁLISE DA CINÉTICA DOS MICROORGANISMOS. Curva de crescimento bacteriano. Neste eixo temos o logaritmo da concentração/ número de bactérias, ou seja, aqui está uma concentração/ número de bactérias por mL. Vamos ver como é que a concentração das bactérias varia ao longo do tempo. Na curva de crescimento bacteriano podemos distinguir 4 fases essenciais: a fase lactente, a fase de crescimento exponencial, a fase estacionária e a fase de declínio. As bactérias crescem de uma forma exponencial, isto é, uma dão 2, 2 dão 4… A bactéria tem que crescer rapidamente se não morrem. ANATOMIA BACTERIANA 19/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO Mas o que é que nós representamos aqui? Temos o tempo e temos o logaritmo da concentração/ número de bactérias. Temos portanto, a taxa de crescimento bacteriano (concentração em função do tempo). A taxa de crescimento (Tc) é dada pelo declive da recta: fase lactente (Tc=0); fase de crescimento exponencial (Tc=positiva), a fase estacionária (Tc=0) e a fase de declínio (Tc=negativa). Este processo demorou entre oito a nove horas. As bactérias dividiram-se e esta representação é obtida a partir de uma cultura de bactérias naquilo que se chama cultura fechada. Fechada porquê? Temos um meio de cultura, os nutrientes, introduzse algumas bactérias (figurativamente duas ou três bactérias) e não acrescento mais nada. Isto é um meio líquido e é a forma que as bactérias assimilam melhor os nutrientes. As bactérias, como por exemplo a escherichia coli, têm que se dividir em 20 minutos (no máximo 22 ou 25) porque se não vão morrer. Vamos imaginar que a bactéria caiu num meio de cultura que tinha como fonte de carbono a lactose e ela estava habituada a metabolizar glicose. A bactéria tem que activar o seu sistema enzimático para degradar a lactose, ou seja, ela tem uma fase de adaptação ao meio. Vai adaptar o seu aparelho enzimático ao substrato nutritivo que encontra. Portanto, nesta fase, elas estão-se a dividir mas não estão a crescer muito em número. Entretanto a bactéria está completamente adaptada ao meio e então a escherichia coli já se consegue dividir exponenciamente. Tem uma nova geração a cada 20 minutos. Interpretação da curva: Inicialmente temos a fase inicial de adaptação e fase de crescimento exponencial (Tc=positiva). Entretanto começam a escassear nutrientes (carboidratos) ou o O2 (que é essencial para as bactérias aeróbias) – Por isso a cultura é mantida em agitação (facilita a dissolução do oxigénio). Então, a bactéria entra numa fase de crescimento estacionário, ou seja, a bactéria leva um bocado mais de tempo a dividir-se até parar por completo a sua divisão (Tc=0). Entretanto o meio não suporta mais o crescimento. Inicialmente, a Tc ainda não é negativa, porque muitas bactérias, apesar de não terem nutrientes no exterior, podem ter reservas metabólicas (carboidratos ou lipídeos) que assegurem a sua sobrevivência. Contudo, esgotadas todas as reservas, a taxa de crescimento torna-se francamente negativa porque o número de bactérias que nasce é significativamente inferior ao número de bactérias que morrem. A taxa de crescimento diminui até que chega ao zero. Esta curva é diferente para o mesmo meio de cultura, é diferente de bactéria para bactéria, é diferente da salmonella para a escherichia coli. Para a mesma escherichia coli é diferente de meio de cultura para meio de cultura. Como calcular ao fim de 4, ao fim de 8 e ao fim de 12 horas quantas bactérias estão ali? E como é que eu sei quantas bactérias viáveis estão ali? Como conto o número de bactérias que estão presentes num meio líquido? Ou as conto numa câmara de contagem ou num aparelho próprio. Ao contar essas células eu não sei se estão viáveis ou estão mortas, sei é que elas não estão lisadas. Contudo, se eu fizer a sementeira num meio sólido eu conto o número de unidades formadoras de colónias. Aquelas que estavam viáveis são capazes de se reproduzir e isto permite-me que, para a mesma bactéria, consiga saber num determinando momento, ANATOMIA BACTERIANA 20/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO quantas bactérias existe naquele meio de cultura. Eu consigo saber qual o tempo de duplicação da bactéria que estou a estudar. É a principal forma de estudar a acção de uma substância, de um fármaco sobre a bactéria, isto é, como é que essa substância afecta a curva de crescimento. Pode afectar sob o ponto de vista positivo ou negativo. Importância da análise da cinética dos microorganismos (exemplos): Permite estudar a influência de determinadas medicações, administradas a doentes que estão nos cuidados intensivos, no desenvolvimento de bactérias, aumentando ou diminuindo a susceptibilidade a infecções. Significa que muitas vezes eles podem acelerar esta fase de crescimento exponencial. Estudos de sensibilidade Quando eu quero estudar células numa população bacteriana que estejam metabolicamente activas e que sejam representantes desse microorganismo colheita na fase de crescimento exponencial (onde eu encontro a maior parte das células viáveis, jovens, com o seu metabolismo perfeitamente expresso). Resumo o que a bactéria precisa para se formar – macro e micro nutrientes. ANATOMIA BACTERIANA 21/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO ANEXOS As imagens apresentadas em anexo visam uma compreensão global das vias de catabolismo e anabolismo presentes nas bactérias. O professor não deu grande relevância a este tema. O importante é ficar com umas noções básicas! Grande parte das imagens foi apresentada na aula. ANATOMIA BACTERIANA 22/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO ANATOMIA BACTERIANA 23/24 FACULDADE DE MEDICINA DA UNIVERSIDADE DO PORTO ANATOMIA BACTERIANA 24/24
Baixar