Fisiologia do sistema reprodutor feminino
João Soares Fernandes
Prof. Doutor JM Soares Fortunato
Dr. Jorge Correia-Pinto
Universidade do Minho
SOF-Fisiologia
ÍNDICE
INTRODUÇÃO
página 3
ANATOMIA FISIOLÓGICA DO APARELHO GENITAL FEMININO
página3
EMBRIOLOGIA E BIOLOGIA DAS CÉLULAS GERMINATIVAS.
OVOGÉNESE E FOLICULOGÉNESE.
página 4
OVULAÇÃO E FORMAÇÃO DO CORPO AMARELO
página 6
REGULAÇÃO HORMONAL DA FUNÇÃO OVÁRICA.
O EIXO HIPOTÁLAMO-HIPÓFISE-OVÁRIO
página 7
AS FASES INICIAIS DO CRESCIMENTO FOLICULAR
página 11
O CICLO MENSTRUAL
página 12
FASE FOLICULAR
página 13
MECANISMO DA OVULAÇÃO
página 15
FASE LUTEÍNICA
página 17
HORMONAS OVÁRICAS
página 18
BIOSSÍNTESE, ACÇÕES CELULARES E METABOLISMO DOS ESTROGÉNIOS
página 18
BIOSSÍNTESE E METABOLISMO DA PROGESTERONA
página 19
ACÇÕES DOS ESTROGÉNIOS E PROGESTERONA SOBRE O APARELHO GENITAL página 19
ACÇÕES SISTÉMICAS DOS ESTROGÉNIOS E PROGESTERONA
FISIOPATOLOGIA DA FUNÇÃO OVÁRICA
página 21
página 22
PERTURBAÇÕES MENSTRUAIS
página 22
SÍNDROMA DO OVÁRIO POLIQUÍSTICO
página 22
TUMORES OVÁRICOS
página 23
FISIOLOGIA DO ACTO SEXUAL
página 24
FISIOLOGIA DA GRAVIDEZ
Módulo reprodução e manutenção da espécie
Página
1
Universidade do Minho
SOF-Fisiologia
FERTILIZAÇÃO
página 26
AS PRIMEIRAS FASES DA DIVISÃO DO ZIGOTO. IMPLANTAÇÃO
página 27
A FORMAÇÃO DA PLACENTA. FISIOLOGIA DO LÍQUDO AMNIÓTICO
página 28
FUNÇÕES DA PLACENTA
página 30
MECANISMOS DE TRANSPORTE ATRAVÉS DA PLACENTA
página 31
FUNÇÃO ENDÓCRINA DA PLACENTA
página 31
MODIFICAÇÕES FISIOLÓGICAS INDUZIDAS PELA GRAVIDEZ
página 34
SISTEMA CARDIOVASCULAR
página 34
SANGUE
página 35
APARELHO RESPIRATÓRIO
página 35
APARELHO DIGESTIVO
página 36
RIM E VIAS URINÁRIAS
página 36
SISTEMA ENDÓCRINO. METABOLISMO.
página 37
FISIOLOGIA DO PARTO
página 37
LACTAÇÃO
página 39
DESENVOLVIMENTO DA MAMA
página 39
A PRODUÇÃO DE LEITE.
página 40
PROLACTINA E SUA REGULAÇÃO.
A EJECÇÃO DO LEITE
BIBLIOGRAFIA
página 41
página 43
Módulo reprodução e manutenção da espécie
Página
2
Universidade do Minho
SOF-Fisiologia
INTRODUÇÃO
A função reprodutora feminina pode ser dividida em duas fases principais: primeira, a preparação do
corpo feminino para a concepção e gestação; segunda, o período da própria gestação. A primeira parte
deste trabalho reporta-se à preparação do corpo feminino para a gestação e a segunda apresenta a
fisiologia da gravidez.
ANATOMIA FISIOLÓGICA DO APARELHO GENITAL FEMININO
A figura 1 representa o aparelho reprodutor feminino. Destacam-se os ovários, trompas de Falópio
(trompas uterinas) e o útero, que constituem os orgãos reprodutores internos, localizados na pelve.
Figura 1: Aparelho reprodutor feminino
O ovário é constituído por 3 zonas distintas:
-
o córtex, que constitui a maior parte do ovário, é delimitado pelo epitélio germinativo e contém o
estroma e os folículos. O estroma contém células tecais, células contrácteis e tecido conjuntivo. As
estruturas foliculares são constituídas por ovócitos rodeados de células da granulosa (uma ou mais
camadas, consoante o estadio de desenvolvimento)
-
a medula, formada por um agregado celular heterogéneo
-
o hilo, por onde entram e saem vasos sanguíneos.
O ovário é parcialmente recoberto por extensões (fímbrias) da trompa uterina, cujo lúmen é contínuo com
o do útero, orgão muscular composto por três camadas:
-
perimétrio, camada externa, conjuntiva;
-
miométrio, camada média, músculo liso;
Módulo reprodução e manutenção da espécie
Página
3
Universidade do Minho
-
SOF-Fisiologia
endométrio, epitélio escamoso estratificado não-queratinizado, composto por um stratum basale e
um stratum functionale, mais superficial, renovado ciclicamente, crescendo sob a acção dos
estrogénios e progesterona e desagregando-se, posteriormente, na menstruação.
O útero apresenta um estreitamento inferior, o colo uterino ou cérvix, que se abre na vagina. A única
barreira física entre a vagina e o útero é um rolhão de muco cervical. A abertura vaginal localiza-se
posteriormente à abertura uretral. Estes orifícios são recobertos longitudinalmente por pregas, os
pequenos e os grandes lábios. O clítoris é uma pequena estrutura formada por tecido eréctil, situada na
extremidade anterior dos pequenos lábios.
EMBRIOLOGIA E BIOLOGIA DAS CÉLULAS GERMINATIVAS. OVOGÉNESE E FOLICULOGÉNESE.
Às 5-6 semanas de gestação, as células germinativas primordiais (ovogónias), migram do saco vitelino do
embrião para a crista genital, onde o ovário se encontra em desenvolvimento. Multiplicam-se por mitoses
sucessivas até às 20-24 semanas de gestação, atingindo os 6-7 milhões de células. A partir das 8-9
semanas de gestação, algumas ovogónias iniciam a primeira divisão da meiose (que suspendem na
profase I), convertendo-se em ovócitos primários. Este processo continua até aos 6 meses de vida extrauterina, altura em que todas as ovogónias se converteram em ovócitos. A primeira divisão da meiose
apenas se completa aquando da ovulação, portanto os ovócitos primários podem viver até 50 anos, isto é,
até à menopausa.
Figura 2: Ovogénese e foliculogénese
Módulo reprodução e manutenção da espécie
Página
4
Universidade do Minho
SOF-Fisiologia
Em simultâneo com a ovogénese tem lugar um processo de degenerescência dos ovócitos - atrésia - pelo
que o seu número se reduz drasticamente para 1-2 milhões (ao nascimento) e cerca de 300 a 400.000 (no
início da puberdade). A atrésia é causada por apoptose (morte celular programada), que se caracteriza
pelo aumento da produção de radicais livres e pela degradação do DNA por endonucleases. O ovócito
torna-se necrótico, a cromatina sofre picnose e as células da granulosa também degeneram. Em contraste
com o sexo masculino, no sexo feminino o número de células reprodutoras diminui continuamente até
que, na menopausa, poucos ovócitos restarão e a capacidade reprodutora terminará.
A formação de folículos ováricos inicia-se, no ovário fetal, pelas 12-16 semanas de gestação. Quando o
ovócito entra na meiose, é rodeado por 1 camada de células fusiformes provenientes do estroma ovárico,
constituindo-se o folículo primordial. Pelas 20-30 semanas, estas células fusiformes transformam-se em
células cubóides (já chamadas células da granulosa) e o folículo passa a designar-se por folículo primário.
As células da granulosa dividem-se e constituem diversas camadas, criando-se o folículo secundário.
Segregam mucopolissacarídeos que constituem um halo protector do ovócito - a zona pelúcida. O folículo
secundário continua a crescer e atinge um diâmetro de 150 µm, enquanto o ovócito atingiu o seu diâmetro
máximo (80 µm). Nesta altura, ocorrem 2 outros fenómenos:
-
é recrutada mais uma camada de células do interstício, que se diferenciam e constituem a teca interna
(células epitelióides semelhantes às da granulosa, segregam hormonas esteróides) e a teca externa
(cápsula de tecido conjuntivo altamente vascularizado).
-
As células da granulosa segregam um líquido folicular que se acumula em vesículas (folículo
vesicular). O líquido vesicular tem, na sua composição, mucopolissacarídeos, electrólitos,
glicosaminoglicanos, hormonas esteróides, oxitocina, activina, inibina, FSH, LH, vasopressina e
proteínas do plasma.
Um destes folículos vai prosseguir o seu desenvolvimento e, neste, o líquido das vesículas coalesce numa
única área central, o antro - é o folículo maduro, folículo antral ou folículo de Graaf.
Figura 3: Estrutura do folículo de Graaf
Módulo reprodução e manutenção da espécie
Página
5
Universidade do Minho
SOF-Fisiologia
À medida que o folículo se desenvolve, o ovócito primário completa a primeira divisão da meiose, de que
resultam o ovócito secundário (maior, contendo todo o citoplasma do ovócito primário) e o primeiro
corpo polar, que se fragmenta e acaba por desaparecer. O ovócito secundário inicia, depois, a segunda
divisão meiótica, que é interrompida, por acção de um factor inibidor da meiose (provavelmente a
inibina), em metafase II, completando-se apenas se ocorrer fertilização.
O ovócito secundário está contido num folículo de Graaf. As células da granulosa deste folículo formam
um anel à volta do ovócito e um pedículo que o suporta. O anel designa-se por corona radiata e o
pedículo é o cumulus oophorus. Entre o ovócito e a corona radiata mantém-se a zona pelúcida que vai
funcionar como barreira à penetração dos espermatozóides.
OVULAÇÃO E FORMAÇÃO DO CORPO AMARELO
Cinco a sete dias após o primeiro dia de uma menstruação, é seleccionado um folículo maduro, que se
torna o folículo dominante desse ciclo. Os restantes folículos secundários sofrem atrésia, um processo que
é estimulado pelos androgénios e pelo ligando do FAS e inibido pelas gonadotrofinas. O folículo
dominante cresce exponencialmente nas 48 horas anteriores à ovulação, atingindo os 20 mm de diâmetro
e fazendo saliência macroscópica à superfície do ovário. A basal das células da granulosa é degradada
proteoliticamente e liberta-se um ovócito secundário, rodeado pela zona pelúcida e corona radiata. Se
não for fecundado, degenera em 12-24 horas. Se houver fecundação, o ovócito completa a segunda
divisão meiótica, em que o citoplasma é, novamente, dividido de modo desigual: a maior parte permanece
no zigoto (ovo fertilizado) e o restante vai para o segundo corpo polar que acaba por degenerar.
As transformações no ovário continuam após a ovulação (figura 4). O folículo que rompeu enche-se
Figura 4: Visão geral do ciclo ovárico
Módulo reprodução e manutenção da espécie
Página
6
Universidade do Minho
SOF-Fisiologia
imediatamente de sangue, formando o chamado corpus hemorrhagicum, que, por vezes, está na origem de
pequenas hemorragias para o interior da cavidade abdominal, causadoras de irritação peritoneal e
consequente dor hipogástrica ("mittelschmerz"). As células da granulosa e tecais iniciam imediatamente a
sua multiplicação e o sangue coagulado é rapidamente substituído por uma nova estrutura endócrina, o
corpo amarelo, composto por células da granulosa (80%), células tecais (20%), capilares e fibroblastos.
As células da granulosa aumentam marcadamente a sua secreção de esteróides, um processo designado
por luteinização, que se traduz, em termos ultra-estruturais, pelo desenvolvimento mitocondrial (matriz
densa, cristas tubulares), aparecimento de gotículas lipídicas no citoplasma e proliferação do retículo
endoplasmático liso.
O corpo amarelo segrega estrogénios e progesterona. Persiste, se houver fertilização; caso contrário,
começa a degenerar cerca de 4 dias antes da menstruação seguinte (luteólise). Sofre necrose, sendo
invadido por leucócitos, macrófagos e fibroblastos, resultando, no final, uma cicatriz avascular - o corpus
albicans.
REGULAÇÃO HORMONAL DA FUNÇÃO OVÁRICA. O EIXO HIPOTÁLAMO-HIPÓFISE-OVÁRIO.
A Hormona Luteo-estimulante (LH) e a Hormona Foliculo-estimulante (FSH) são glicoproteínas cuja
função é regular o crescimento, maturação pubertária, processos reprodutivos e secreção de esteróides
sexuais pelas gónadas de ambos os sexos.
A secreção de FSH e LH é estimulada por uma única hormona hipotalámica, a Hormona Libertadora das
Gonadotrofinas (GnRH). Esta hormona provoca muito maior libertação de LH do que de FSH, pelo que é
também designada LHRH.
A GnRH é um decapeptídeo produzido predominantemente ao nível do núcleo arqueado e na área préóptica do hipotálamo, sendo posteriormente transportada até à eminência mediana e armazenada em
grânulos de secreção.
Antes da puberdade, a secreção de GnRH é contínua e não consegue despertar a secreção de
gonadotrofinas pela hipófise. Na puberdade, por maturação do sistema límbico, a secreção de GnRH
torna-se pulsátil, restringindo-se, primeiro, à fase REM do sono e generalizando-se, depois, ao sono nãoREM e finalmente às 24 horas.
A libertação de GnRH é modulada por influências inibitórias (dopaminérgicas, endorfinérgicas) e
excitatórias (noradrenérgicas) (figura 5).
Módulo reprodução e manutenção da espécie
Página
7
Universidade do Minho
SOF-Fisiologia
Figura 5: O eixo hipotálamo-hipófise-ovário.
O mecanismo de activação da libertação de GnRH é desencadeado pela estimulação de terminais
noradrenérgicos na região da eminência mediana, por terminações glutamatérgicas que libertam
glutamato. Este actua via receptores NMDA situados no neurónio noradrenérgico. A noradrenalina
libertada actua, através de receptores α1, em neurónios NOérgicos (figura 6), onde leva à produção de
NO. Este difunde-se para neurónios produtores de GnRH, que se encontram entremeados com os
neurónios NOérgicos e estimula a libertação de GnRH para os vasos porta-hipófisários pela activação das
enzimas guanil-cíclase solúvel e cicloxigenase-1. A resposta das terminações neuronais produtoras de
GnRH ao NO é inibida pelo etanol.
-
NAn
Libertação de GnRH
por acção do NO
+
α 1r
GM-CSF
IL-1
+
GnRHn
NO
+
GABAn
NO
GABA-A r
-
GnRH
S. PortaHipofisário
Figura 6. Controlo da libertação de GnRH pelo NO.
Módulo reprodução e manutenção da espécie
Página
8
Universidade do Minho
SOF-Fisiologia
A melatonina é produzida pela glândula pineal em resposta às variações da luminosidade do meio. A
secreção de melatonina, que inibe a produção de gonadotrofinas, é suprimida pela luminosidade e
estimulada pela escuridão. Nalgumas espécies, a melatonina funciona como mediador das variações
sazonais da produção de gonadotrofinas e da actividade reprodutora. No homem, embora os níveis de
melatonina se relacionem inversamente com a secreção de gonadotrofinas, não está, ainda, bem
esclarecido o papel deste mediador na regulação da função reprodutora.
A ansiedade e o stress físico e emocional influenciam reconhecidamente a função reprodutora. Na mulher
tem-se verificado a perda da função menstrual e, no homem, a diminuição da produção de
espermatozóides, efeitos que são, provavelmente, mediados pela Hormona Libertadora da Corticotrofina
(CRH) que inibe a libertação de GnRH.
As feromonas, partículas estimulantes ou inibitórias de natureza química, são registadas pelo olfacto e
chegam ao hipotálamo através de conexões deste com o bolbo olfactivo. Transmitem, assim, ao Sistema
Nervoso Central (SNC), sinais provenientes do meio e de outros indivíduos.
A GnRH atinge a hipófise anterior (adeno-hipófise) através dos vasos do sistema porta hipofisário e ligase à membrana plasmática dos gonadotropos. Geram-se segundos mensageiros (cálcio-calmodulina,
fosfatidilinositol e derivados), estimulando-se a exocitose dos grânulos de gonadotrofinas.
Uma infusão intravenosa de GnRH causa uma resposta bifásica na concentração plasmática de LH: há um
pico inicial aos 30 minutos, seguido por uma subida gradual que se inicia aos 90 minutos e continua por
várias horas. Relativamente às concentrações plasmáticas de FSH, a GnRH provoca um crescimento
unifásico e progressivo.
A pulsatilidade da secreção de LH mantém-se à custa da pulsatilidade da GnRH e não depende dos
esteróides gonadais (os indivíduos eunucos e as mulheres pós-menopáusicas têm picos de LH). Por isso, a
amplitude dos picos de LH aumenta durante a puberdade, primeiro apenas durante a noite e
generalizando-se às 24 horas ao fim de 1-2 anos de maturação.
A característica fundamental da secreção de LH na mulher é a sua variação cíclica mensal. O ciclo
menstrual resulta de uma complexa interacção entre o eixo hipotálamo-hipofisário e as alterações
sequenciais na secreção de esteróides pelo ovário, como é descrito adiante.
A infertilidade feminina pode resultar de perturbações na regulação hipotalámica levando à incapacidade
de produção de gonadotrofinas e à impossibilidade de ocorrer ovulação. Administrando GnRH em pulsos,
de amplitude e frequência semelhantes ao normal (a administração contínua de GnRH resulta em downregulation do receptor de GnRH nos gonadotropos e perda das funções gonadais), é possível restaurar a
normalidade da ovulação e dos ciclos menstruais. A GnRH exógena é ainda utilizada no tratamento das
dores menstruais em mulheres com endometriose.
Módulo reprodução e manutenção da espécie
Página
9
Universidade do Minho
SOF-Fisiologia
A secreção de FSH é, também, pulsátil e exibe um padrão semelhante ao da secreção de LH mas a
amplitude dos picos é muito menor.
A secreção de LH e FSH é regulada pelos produtos de secreção ovárica e rege-se, genericamente, por um
mecanismo de retrocontrolo negativo, isto é, um aumento dos esteróides ováricos suprime a libertação de
gonadotrofinas. A ablação cirúrgica ou a inactivação funcional dos ovários (hipogonadismo) resulta em
aumento das concentrações plasmáticas de FSH e LH, mas a FSH aumenta proporcionalmente mais que a
LH. Os produtos de secreção ovárica que regulam a secreção de gonadotrofinas estão representados na
figura 5.
O 17-β-estradiol (E2) é o estrogénio mais importante sob o ponto de vista funcional. É produzido pelas
células da granulosa e inibe a secreção de LH, diminuindo a amplitude e a frequência dos seus pulsos, o
que indica que o estradiol actua tanto a nível hipotalámico como a nível hipofisário. A administração de
estradiol exógeno bloqueia a resposta dos gonadotropos a um pulso de GnRH. Em contraste, quando há
deficiência de estradiol, a produção de LH em resposta à GnRH aumenta muito.
A secreção de FSH é também inibida pelo estradiol, que bloqueia a resposta dos gonadotropos produtores
de FSH à GnRH. Todavia, o retrocontrolo negativo da produção de FSH é especificamente efectuado por
uma glicoproteína, a inibina, produzida pelo corpo amarelo (inibina A) e pelas células da granulosa
(inibina B). Esta glicoproteína
inibe a síntese da subunidade β da FSH, a exocitose de FSH e,
possivelmente, a secreção de GnRH. Tem efeitos muito pouco significativos sobre a secreçao de LH.
Porém, o estradiol pode exercer retrocontrolo positivo sobre a secreção de gonadotrofinas. Quando se
administra estradiol, em dose apropriada e durante um período suficiente, a secreção de LH, em resposta à
GnRH, aumenta. Além disso, administrando pulsos de GnRH a mulheres previamente expostas ao
estradiol, verifica-se que a resposta secretora de LH é muito maior. Isto significa que o tratamento com
estradiol aumenta a sensibilidade dos gonadotropos (porque induz nestes a síntese de receptores para a
GnRH) e aumenta, também, as reservas de LH.
A progesterona é outro esteróide ovárico que regula a libertação de LH. A administração aguda de
progesterona aumenta as concentrações plasmáticas de LH, nas 24-48 horas seguintes. Esta hormona
pode, ainda, amplificar ou atenuar o efeito de retrocontrolo positivo do estradiol, dependendo do
momento da administração. Continuamente, tem um efeito inibidor sobre a secreção de gonadotrofinas.
Os contraceptivos orais são compostos por uma forma sintética de estrogénio combinada com um
progestativo sintético. Os comprimidos são tomados diariamente, durante 3 semanas, após o último dia de
um período menstrual. Há um aumento imediato e sustentado dos níveis plasmáticos dos esteróides
ováricos, que exercem feedback negativo sobre a secreção de gonadotrofinas, impedindo a ovulação.
Módulo reprodução e manutenção da espécie
Página
10
Universidade do Minho
SOF-Fisiologia
Todo o ciclo se assemelha, então, a uma falsa fase luteínica, com altos níveis de progesterona e
estrogénios e baixos níveis de gonadotrofinas.
A secreção e síntese de FSH é, ainda, estimulada pela activina (estruturalmente semelhante à inibina,
produzida pelas célula da granulosa). A activina é, também, sintetizada pelas células da hipófise, onde
actua por auto e paracrinia. A foliculostatina liga-se à activina, inactivando-a. A prolactina também inibe
a libertação de GnRH, diminuindo a secreção basal de LH e FSH. Finalmente, a LH pode inibir a
secreção de GnRH, através da denominada via curta de retrocontrolo.
AS FASES INICIAIS DO CRESCIMENTO FOLICULAR
A possível existência de um factor desencadeante do início do crescimento folicular é uma questão que
permanece por esclarecer.
Na mulher, há uma correlação forte entre a reserva de folículos em repouso (constituída pelos folículos
primários e pelos folículos primários pequenos) e o número de folículos em crescimento: quanto maior a
reserva (ovário da criança), maior o número de folículos em crescimento. Com o envelhecimento, à
medida que os folículos abandonam a reserva, o número absoluto de folículos em crescimento diminui
mas a proporção aumenta. Este aumento pode ser despoletado pela desaparecimento progressivo de uma
influência inibidora exercida pelos próprios folículos em repouso, actuando por um mecanismo
desconhecido e de magnitude directamente proporcional ao tamanho da reserva.
Esse hipotético mecanismo pode representar um processo adaptativo, permitindo ao ovário promover o
crescimento de um número suficiente de folículos, de modo que, um deles, atinja o completo
desenvolvimento e a fase de ovulação, apesar do número, cada vez menor, de folículos disponíveis.
Por outro lado, os estudos animais sugerem que o início do crescimento folicular é independente dos
efeitos das gonadotrofinas, até porque, em situações de hipopituitarismo, não se encontra completamente
abolido.
Recentemente, foi proposto que a TSH pode actuar em sinergia com a FSH, promovendo o início do
crescimento dos folículos em repouso.
Finalmente, sugeriram-se diferenças na expressão da proteína do gene do retinoblastoma (pRb), nas
células da granulosa e no ovócito. Nos folículos primordiais, a expressão de pRb é baixa nas células da
granulosa e alta no nucléolo do ovócito, onde induz a produção de factores inibidores da proliferação das
células da granulosa. Com o início do crescimento folicular, aumenta a expressão de pRb na granulosa,
diminuindo no ovócito. Estes dados sugerem que o estado de repouso se mantém à custa do ovócito e que
o início do crescimento se deve às células da granulosa.
Módulo reprodução e manutenção da espécie
Página
11
Universidade do Minho
SOF-Fisiologia
CICLO MENSTRUAL
As variações cíclicas na secreção de gonadotrofinas estão na base das transformações que ocorrem no
ovário durante um ciclo mensal (ciclo menstrual). O ciclo ovárico faz-se acompanhar de variações
cíclicas na secreção de estradiol e progesterona que, interactuando com o hipotálamo e hipófise, regulam
a secreção de gonadotrofinas.
Figura 7. O ciclo menstrual.
A duração de um ciclo menstrual típico é de 28 dias, podendo variar entre os 21 e os 35 dias.
Convencionou-se designar o primeiro dia de menstruação por "dia 1" do ciclo, porque o fluxo de sangue
menstrual é a mais evidente das alterações que acontecem.
O CICLO OVÁRICO
O ciclo ovárico divide-se, fisiologicamente, em 3 fases sequenciais:
-
a fase folicular, que se inicia com a hemorragia menstrual e se prolonga por 15 dias (variando entre
os 9 e os 23 dias).
-
a fase ovulatória, com 1 a 3 dias de duração, culminando na ovulação.
-
a fase luteínica, com uma duração mais constante de, aproximadamente, 13 dias, terminando com o
início da hemorragia menstrual.
Módulo reprodução e manutenção da espécie
Página
12
Universidade do Minho
SOF-Fisiologia
Fase folicular
Durante esta fase, verifica-se o crescimento de alguns folículos primários, o desenvolvimento de vesículas
e a transformação em folículos secundários, um dos quais será seleccionado para atingir a maturidade
(folículo dominante). À medida que os folículos crescem, aumenta a secreção de estradiol pelas células da
granulosa, atingindo a sua concentração máxima pelo dia 12 do ciclo, 2 dias antes da ovulação (figura 8).
Figura 8. Variação das secreções de LH,
FSH, estradiol e progesterona ao longo do
ciclo ovárico.
No final da fase luteínica do ciclo anterior, vamos encontrar as concentrações plasmáticas de FSH e LH
nos seus níveis mais baixos. Um a dois dias antes da menstruação, a concentração de LH começa a subir,
seguindo-se, mais tardiamente, a subida da concentração de LH. Os estrogénios (estradiol, produzido
predominantemente pelo folículo dominante e estrona, produzida perifericamente, a partir de estradiol e
androstenediona) aumentam gradualmente, estimulados pelas concentrações crescentes de FSH na metade
inicial da fase folicular. Na segunda metade da fase folicular, a concentração de FSH cai moderadamente,
enquanto a concentração de LH continua a subir lentamente.
Como se explicam estas variações?
A acção central da FSH é estimular a hiperplasia e hipertrofia das células da granulosa dos folículos
primários, induzindo nestas a síntese dos seus próprios receptores (FSH-R) e sensibilizando-as, assim, à
sua própria acção. Induz, ainda, a produção de aromatase, que converte precursores androgénicos em
estrogénios, de IGF-1 e IGF-2 (que estimulam a proliferação e esteroidogénese folicular), de factor de
crescimento epidérmico (EGF) e de factor de transformação de crescimento α (TGF α) (próproliferativos, mas inibidores da esteroidogénese).
O aumento da concentração local de estradiol induz, nas células da granulosa, a síntese de FSH-R e de
receptores para o próprio estradiol, sensibilizando-as à acção destas hormonas. Daqui resulta que, embora
a concentração plasmática de FSH não aumente durante a segunda metade da fase folicular (de facto,
pode, até, diminuir), o seu efeito estimulante aumentará sempre.
Módulo reprodução e manutenção da espécie
Página
13
Universidade do Minho
SOF-Fisiologia
O FSH-R pertence à família dos receptores ligados a proteínas G. Quando uma molécula de FSH se liga
ao receptor, a subunidade Gsα dissocia-se e, com o GTP, activa a adenilciclase, sintetizando-se AMPc. O
AMPc promove a entrada de Ca++ e activa a proteína cínase A (PKA), que se dissocia em subunidades
catalítica (C) e reguladora (R). A subunidade C vai actuar a nível nuclear, activando, por fosforilação,
factores de transcrição (CRE), levando à transcrição de RNAm e consequente síntese proteica (figura 9).
Figura 9. Transducção de sinal pelo receptor da FSH.
Por outro lado, a FSH em conjunto com o estradiol induz a síntese de receptores para a LH nas células da
granulosa. Os níveis crescentes de estradiol actuam a nível hipotalamo-hipofisário, aumentando os pulsos
de GnRH e a secreção e reservas de LH.
O estradiol também induz a síntese de receptores para a LH nas células tecais, sensibilizando-as à acção
desta hormona. A LH estimula, nas células tecais, a produção de androgénios (predominantemente
androstenediona), que difundem através da lâmina basal para as células da granulosa, onde servem de
substrato para a síntese de estradiol. Nas células da granulosa, a LH estimula a síntese de progesterona,
que pode difundir para a teca e servir de substrato à síntese de androgénios. Esta "cooperação celular" em
resposta às acções da LH e FSH tem sido designada por "Teoria das Duas Células - Duas Gonadotrofinas"
(figura 10).
Na regulação da esteroidogénese folicular intervêm, ainda, a inibina, activina e foliculostatina. Por um
lado, a inibina estimula a produção de androgénios pelas células tecais, enquanto a activina a inibe. Por
outro lado, está provado no animal de experiência (rato), que a inibina é um potente inibidor da
proliferação das células da granulosa, enquanto a activina estimula essa proliferação. A foliculostatina
estimula a síntese de progesterona e inibe as acções da activina.
Módulo reprodução e manutenção da espécie
Página
14
Universidade do Minho
SOF-Fisiologia
Figura 10. Teoria bicelular da esteroidogénese folicular.
Pelo 5º-7º dias da fase folicular estabelece-se a dominância folicular, isto é, um folículo suplanta os
outros, em termos de velocidade de crescimento. Este folículo dominante é o melhor vascularizado, pelo
que se encontra mais acessível à acção da FSH. Tem, por isso, mais receptores para a FSH, maior
capacidade de aromatização e, logo, de síntese de estradiol.
O aumento da síntese de estradiol devido ao folículo dominante, permite que se atinjam durante, pelo
menos, dois dias, concentrações plasmáticas da ordem das 200 pg/ml. Nestas condições, o estradiol
desencadeia um feedback positivo explosivo e de curta duração pelo qual:
-
a nível hipotalámico são bloqueadas as influências inibitórias dopaminérgicas e endorfinérgicas,
resultando em aumento dos pulsos de GnRH.
-
a adenohipófise, sensibilizada pela exposição prévia ao estradiol, responde lançando em circulação as
reservas de LH acumuladas. A concentração plasmática de LH sobe em pico. Como a GnRH também
estimula a secreção de FSH, há também um pico de concentração desta hormona, se bem que menor.
Mecanismo da ovulação
Na preparação da ovulação, ocorrem no ovário uma série de acontecimentos finamente regulados. Antes
de mais, o folículo pré-ovulatório tem de sofrer maturação, seguindo-se a expansão do cumulus oophorus
e, finalmente, a rotura da parede folicular.
A maturação folicular deve-se às acções combinadas da FSH, do estradiol e de vários factores de
crescimento (figura 11). O folículo pré-ovulatório expressa hormonas esteroidogénicas, necessárias à
síntese de estradiol que, como vimos, despoleta o pico de LH.
A LH em altas concentrações, actuando no folículo maduro, bloqueia a expressão dos genes associados à
foliculogénese, isto é, dos genes que controlam a proliferação da granulosa - IGF-1, receptor da FSH,
Módulo reprodução e manutenção da espécie
Página
15
Universidade do Minho
SOF-Fisiologia
receptor β dos estrogénios e ciclina D2. Como consequência do aumento do AMPc intracelular causado
pela acção da LH, termina, também, a expressão dos genes codificantes das enzimas esteroidogénicas.
Figura 11. Expressão genética nos folículos pré-ovulatórios e ovulação.
Em contraponto, a LH induz a expressão de genes de proteínas envolvidas na ovulação:
-
Nas células da granulosa, é induzida a cicloxigenase-2 (COX-2), enzima determinante na síntese de
prostaglandinas, fundamentais na sinalização celular e na criação de uma reacção pseudoinflamatória local.
-
Nas células do cumulus oophorus, induz a síntase do ácido hialurónico, permitindo a síntese deste
proteoglicano, determinante da expansão da matriz extra-celular do cumulus.
-
Nas células tecais, é induzido o receptor da progesterona (PR), um membro da super-família de
receptores nucleares, que regula numerosas funções nos orgãos reprodutores, incluindo o útero,
glândula mamária e ovário. Recentemente, identificaram-se os alvos do PR, de que se destaca a
ADAMTS-1 (figura 12). Trata-se de uma proteína multi-funcional, com diversos domínios:
metaloproteinase, desintegrina e trombospondina-like. O domínio metaloproteinase degrada os
proteoglicanos e proteínas específicas da parede folicular, os domínios desintegrina e
trombospondina-like permitem-lhe a activação de vias de sinalização celular, resultando,
nomeadamente, num potente efeito anti-angiogénico, impedindo a formação de neo-vasos neste
passo específico da ovulação. Outro dos alvos do PR é a catepsina L, uma enzima da família da
papaína. É uma protease lisosómica que também contribui para a degradação da parede folicular e
libertação do ovócito.
Módulo reprodução e manutenção da espécie
Página
16
Universidade do Minho
SOF-Fisiologia
A ovulação acontece, portanto, como resultado dos efeitos sequenciais da FSH e LH nos folículos
ováricos. Pelo feedback positivo do estradiol sobre a secreção de LH, o folículo como que determina o
momento da sua própria ovulação. Isto porque a ovulação é desencadeada por um pico de LH que, por
sua vez, resulta do aumento da secreção de estradiol que ocorre com o crescimento folicular e maturação
do folículo dominante. Este não pode, pois, entrar no processo de ovulação enquanto não atingir o
tamanho e maturidade necessários.
Figura 12. Potenciais alvos de acção da ADAMTS-1 durante a ovulação.
Fase luteínica
Depois da ovulação, o folículo vazio é transformado pela LH numa nova estrutura - o corpo amarelo
(corpus luteum), ocorrendo, simultaneamente, uma transformação funcional - enquanto os folículos
produzem estradiol, o corpo amarelo produz estradiol e progesterona (17-hidroxiprogesterona).
As altas concentrações de progesterona exercem, em conjunto com o estradiol, feedback negativo sobre a
secreção de LH e FSH. O corpo amarelo produz, ainda, inibina A, que exerce a mesma função. A
supressão da secreção de FSH retarda o desenvolvimento de novos folículos, impossibilitando novas
ovulações nos dias seguintes do ciclo.
A perda do suporte gonadotrófico e a secreção, pelo útero, de uma hormona designada por luteolisina
(provavelmente PGF2α) levam à involução e atrésia do corpo amarelo, caindo as concentrações de
estrogénios e progesterona para níveis muito baixos. Se houver fertilização e gravidez, a gonadotrofina
coriónica (HCG) e a prolactina, pelo seu efeito luteotrófico, mantêm funcionante o corpo amarelo.
Módulo reprodução e manutenção da espécie
Página
17
Universidade do Minho
SOF-Fisiologia
HORMONAS OVÁRICAS
Biossíntese, acções celulares e metabolismo dos estrogénios
Os estrogénios mais importantes são o 17-β-estradiol (E2), a estrona e o estriol. O E2 é o mais potente dos
três, encontrando-se em equilíbrio com a estrona, na circulação. A estrona é metabolizada em estriol, que
é o menos potente dos três.
Os estrogénios são produzidos pela teca interna e granulosa dos folículos ováricos, pelo corpo amarelo,
placenta e, em pequenas quantidades, pelo córtex da suprarrenal e pelo testículo. A biossíntese dos
estrogénios inclui a sua formação a partir dos androgénios e a síntese por aromatização da
androstenediona, na circulação (a aromatase é a enzima que cataliza a conversão de androstenediona a
estrona e de testosterona a estradiol.
Figura 13. Biossíntese dos estrogénios.
É importante recordar aqui a cooperação entre as células da granulosa e a teca na síntese de estradiol
(teoria bicelular da esteroidogénese folicular), que já foi descrita anteriormente.
Nas células alvo, os estrogénios actuam a três níveis distintos:
-
a nível membranar: em receptores semelhantes aos clássicos e aos não-clássicos e em canais iónicos
dependentes de ligandos e dependentes da voltagem.
-
a nível citoplasmático: activam cínases e induzem a NO síntase endotelial, aumentando a síntese de
NO.
Módulo reprodução e manutenção da espécie
Página
18
Universidade do Minho
-
SOF-Fisiologia
a nível nuclear: actuam sobre receptores clássicos, Erα, e não-clássicos, Erβ (estes de menor afinidade
para o E2). O complexo estrogénio-receptor vai, depois, modular a expressão de genes e a síntese
proteica.
O estradiol em circulação encontra-se nas formas livre (3%) e ligado a proteínas (60% ligado à albumina
e 37% ligado à globulina de ligação das hormonas sexuais, SHBG, que também se liga à testosterona). Os
estrogénios são metabolizados a nível hepático, por oxidação, glicuronoconjugação e sulfoconjugação.
Sofrem secreção biliar e reabsorção intestinal (circulação entero-hepática).
Biossíntese e metabolismo da progesterona
A progesterona é produzida pelo corpo amarelo e pela placenta. É um importante intermediário da síntese
de todas as hormonas esteróides. A progesterona circula ligada à globulina de ligação dos esteróides
sexuais (SSBG) (45%), à albumina (50%) e 1-2% na forma livre. Tem uma semi-vida curta e é convertida
no fígado em pregnanediol que é conjugado com o ácido glicurónico e excretado na urina.
Acções dos estrogénios e progesterona a nível genital
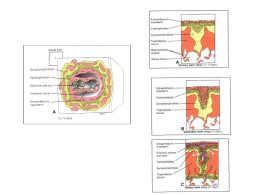
Endométrio: o ciclo endometrial
As modificações no endométrio devem-se às variações cíclicas na secreção de estradiol e progesterona
pelo ovário. Divide-se habitualmente em 3 fases: proliferativa, secretora e menstrual (Figura 14).
Figura 14. O ciclo endometrial
A fase proliferativa ocorre quando o ovário está em fase folicular. O aumento da secreção de estradiol
estimula o crescimento do stratum functionale do endométrio (atinge os 10 mm de espessura).
Módulo reprodução e manutenção da espécie
Página
19
Universidade do Minho
SOF-Fisiologia
Desenvolvem-se vasos tortuosos, as artérias espiraladas. O estradiol também estimula a síntese de
receptores para a progesterona, preparando a fase seguinte do ciclo.
A fase secretora coincide com a fase luteínica do ovário. O aumento da secreção de progesterona estimula
o desenvolvimento de glândulas uterinas e a acumulação de glicogénio. Pelas acções combinadas do
estradiol e progesterona, o endométrio torna-se espesso, bem vascularizado e de aspecto esponjoso,
preparado para acolher um embrião, caso ocorra fertilização.
A fase menstrual resulta da queda da secreção hormonal ovárica, na parte final da fase luteínica. Há
necrose e desagregação do stratum functionale do endométrio, devido à constrição das artérias
espiraladas, por acção das prostaglandinas produzidas localmente (particularmente PGF2α).
A menstruação normal contém sangue (predominantemente arterial, apenas 25% do sangue menstrual é
venoso), células endometriais descamadas, prostaglandinas e fibrinolisina. A fibrinolisina, de origem
endometrial, lisa os coágulos, pelo que, a menos que o fluxo seja anormalmente volumoso, o sangue
menstrual não contém coágulos. A duração habitual do fluxo menstrual é de 3-5 dias (variando entre 1 e
8 dias em mulheres normais). O volume varia entre pequenas perdas não-quantificáveis (spotting) até aos
80 mL (30 mL, em média). Obviamente, o volume do fluxo é condicionado por factores como a espessura
do endométrio, medicação e coagulopatias.
Colo uterino
A mucosa do colo uterino não sofre descamação cíclica, mas as características do muco cervical alteramse regularmente. Sob influência estrogénica, o muco torna-se abundante, aquoso, alcalino, transparente,
pouco viscoso mas muito elástico. Esta elasticidade (ou filância, ou spinnbarkeit) é máxima na ovulação,
de tal forma que, nessa altura, uma gota de muco pode ser distendida até aos 8-12 cm de comprimento. Os
estrogénios aumentam, ainda, o diâmetro do orifício cervical externo e do endocérvix, o que,
conjuntamente com as características do muco, facilita a ascensão dos espermatozóides.
Depois da ovulação e durante a gravidez, sob influência da progesterona, o muco torna-se espesso e
opaco e o diâmetro do orifício cervical externo diminui de diâmetro.
Vagina
Os estrogénios têm uma acção mitogénica e pró-proliferativa no epitélio vaginal, estimulando, ainda, a
acumulação de glicogénio. A progesterona provoca uma diminuição da espessura do epitélio vaginal,
promovendo a descamação das camadas superficiais.
Módulo reprodução e manutenção da espécie
Página
20
Universidade do Minho
SOF-Fisiologia
Mama
Nos estudos efectuados nos últimos anos verificou-se a existência de um verdadeiro ciclo na mama. Este
é constituído por uma fase de repouso pré-ovulatória e uma fase de proliferação pós-ovulatória, a qual
conduz a um aumento de volume da mama, que é máximo nos dias que antecedem a menstruação. O
aumento da mama é condicionado pela acção da progesterona que provoca hiperemia, aumento do tecido
conjuntivo e aumento do tecido glandular (alvéolos mamários), o qual leva a um aumento do produto de
secreção. Também ocorre um aumento dos ductos glandulares e dilatação dos ductos excretores,
provavelmente induzido pelos estrogénios. Depois da menstruação a mama entra num período de
involução (fase de regressão).
Acções sistémicas dos estrogénios e progesterona
Para além do efeito sobre o aparelho genital, os estrogénios actuam, a nível sistémico, no
desenvolvimento do fenótipo feminino: ombros estreitos e ancas largas, convergência das coxas e
divergência dos braços (amplo ângulo de carregador), concentração adiposa nas mamas e nádegas,
manutenção das proporções pré-pubertárias da laringe e, logo, do tom de voz mais agudo, distribuição do
pelo púbico em forma de triângulo de base superior.
A nível renal, os estrogénios promovem a reabsorção de sódio, contribuindo para a retenção hídrica
observada ciclicamente nalgumas mulheres.
No esqueleto, os estrogénios inibem a reabsorção óssea, inibindo os osteoclastos e aumentam a formação
de tecido ósseo, induzindo localmente a síntese de factores de crescimento. Promovem, ainda, a fusão das
epífises dos ossos longos, mais precocemente do que a testosterona no sexo masculino, motivo pelo qual
as mulheres têm, em geral, uma estatura inferior à dos homens.
A nível hepático, os estrogénios promovem a síntese de diversas proteínas: globulina de ligação da
tiroxina (TBG), globulina de ligação do cortisol (CBG), globulina de ligação das hormonas sexuais
(SHBG), angiotensinogénio, lipoproteínas de muito baixa densidade (VLDL) e lipoproteínas de alta
densidade (HDL). Baixam, contudo, os níveis de lipoproteínas de baixa densidade (LDL).
A nível vascular, os estrogénios têm uma acção vasodilatadora e anti-vasoconstrictora, pois aumentam a
síntese de substâncias vasodilatadoras (NO, via receptores ERα e por activação directa da NO síntase
endotelial; PGE2 e PGI2) e inibem a formação de endotelina-1 (vasoconstrictor).
A progesterona é termogénica e é responsável pela subida de cerca de 0.5 ºC na temperatura corporal que
ocorre logo após a ovulação. Estimula a respiração (aumenta a sensibilidade do centro respiratório à
estimulação pelo CO2), pelo que a PCO2 no sangue arterial da mulher em fase luteínica é inferior à do
Módulo reprodução e manutenção da espécie
Página
21
Universidade do Minho
SOF-Fisiologia
homem. A progesterona bloqueia a acção da aldosterona, produzindo natriurese. A nível central, aumenta
o apetite e induz uma tendência à sonolência.
FISIOPATOLOGIA DA FUNÇÃO OVÁRICA
Perturbações menstruais
A infertilidade feminina pode dever-se a ciclos anovulatórios: embora haja períodos menstruais regulares,
não ocorre ovulação. Estes ciclos anovulatórios são muito frequentes nos primeiros meses após a menarca
e, novamente, antes da menopausa.
O termo amenorreia refere-se à ausência de períodos menstruais. Se nunca houve uma hemorragia
menstrual, a perturbação designa-se por amenorreia primária e pode dever-se a anomalias anatómicas do
aparelho genital (p.ex. hímen imperfurado) ou a síndromas de imaturidade sexual. A cessação dos ciclos
numa mulher com períodos, até então, normais é chamada amenorreia secundária. A causa mais frequente
de amenorreia secundária é a gravidez. Outras causas de amenorreia incluem estímulos emocionais,
doenças hipotalámo-hipofisárias, doenças ováricas e diversas doenças sistémicas. Nalgumas mulheres
com amenorreia de causa hipotalámica, a frequência dos pulsos de GnRH encontra-se diminuida por
excesso de actividade opióide (endorfinérgica). Tem-se conseguido a restauração da frequência dos
pulsos com a administração de bloqueadores dos receptores opióides, como a naltrexona.
Os termos oligomenorreia e menorragia referem-se, respectivamente, a fluxos menstruais escasso e
anormalmente abundante, em ciclos regulares. Metrorragia é a hemorragia genital fora do período
menstrual. Dismenorreia é a menstruação dolorosa. É mais frequente em mulheres jovens e desaparece
frequentemente após a primeira gravidez. O alívio sintomático consegue-se com a administração de antiinflamatórios não-esteróides, que inibem a síntese de prostaglandinas, cuja acumulação é responsável
pelos fenómenos dolorosos.
Algumas mulheres apresentam o chamado síndroma pré-menstrual, durante os últimos 7-10 dias do ciclo.
O quadro sintomático inclui: irritabilidade e labilidade emocional, dificuldade de concentração, cefaleias,
obstipação e edemas.
Síndroma do ovário poliquístico
O síndroma do ovário poliquístico (Síndroma de Stein-Leventhal) é uma causa de infertilidade e
amenorreia e caracteriza-se pelo espessamento da cápsula ovárica e desenvolvimento de múltiplos quistos
foliculares (figura 15). É habitualmente bilateral. Cursa com níveis elevados de testosterona, estradiol e
LH e níveis baixos de FSH. Tem sido sugerido que esta perturbação se deve a um aumento da frequência
dos pulsos de GnRH, o que favorece a secreção de LH.
Módulo reprodução e manutenção da espécie
Página
22
Universidade do Minho
SOF-Fisiologia
Figura 15. Síndroma do ovário poliquístico
Tumores ováricos
Do ponto de vista fisiopatológico, a divisão mais importante consiste em agrupá-los em tumores
secretores de androgénios, que causam masculinização e tumores secretores de estrogénios, que
promovem o desenvolvimento sexual precoce.
Módulo reprodução e manutenção da espécie
Página
23
Universidade do Minho
SOF-Fisiologia
O ACTO SEXUAL
O desejo sexual feminino é aumentado por factores:
-
endócrinos: o desejo atinge o máximo próximo da ovulação, provavelmente devido aos altos níveis
de secreção de estrogénios. Os androgénios também exercem, na mulher, um efeito estimulante do
desejo sexual.
-
sensoriais: as feromonas, sinais químicos emitidos por um indivíduo e percepcionados pelo olfacto.
-
psíquicos: os pensamentos eróticos, a líbido.
Durante a excitação sexual, as terminações nervosas parassimpáticas provenientes do plexo sagrado
libertam, nas estruturas genitais, acetilcolina, peptídeos vasodilatadores e NO, tendo como resultado, a
dilatação e congestão venosas, causando a erecção do clítoris e a turgescência dos pequenos lábios que
fazem saliência e alongam funcionalmente a vagina. Estimulam a secreção das glândulas de Bartholin,
responsáveis pela lubrificação durante o acto sexual.
Figura 16. Excitação.
A congestão venosa origina passagem de líquido para o espaço intersticial e consequente edema. Ocorre
transudação de líquido através das paredes vaginais. Os dois terços superiores da vagina distendem-se e o
útero eleva-se para o interior da falsa pelve. Abaixo, encontra-se uma plataforma de tecidos distendidos
pela congestão e edema pélvicos - a plataforma orgásmica (figura 17).
O orgasmo varia de um episódio para outro, mas, em geral, consiste em 15-18 contracções rítmicas dos
músculos transverso do períneo e elevador do ânus, que convergem nas paredes laterais do terço inferior
da vagina. As 5-6 primeiras contracções são as mais intensas. Resultam de reflexos da medula espinal que
provocam, também, o aumento da motilidade tubar e contracções uterinas semelhantes às do parto. O
canal cervical permanece dilatado durante cerca de 30 minutos, permitindo, assim, o fácil transporte dos
Página 24
Módulo reprodução e manutenção da espécie
Universidade do Minho
SOF-Fisiologia
espermatozóides. Nalguns casos, o sangue pode refluir às estruturas distendidas, o que explica que
algumas mulheres respondam a novas estimulações, segundos depois de um orgasmo inicial e tenham
orgasmos múltiplos.
Figura 17. Planalto ou plataforma orgásmica.
A resposta orgásmica não se limita ao orgãos genitais. As mamas aumentam de volume e há erecção
mamilar, em consequência da congestão durante a fase de excitação. Durante as fases de elevada
excitação sexual podem também produzir-se espasmos da musculatura abdominal. A pele de algumas
mulheres adquire uma tonalidade rosada, que é mais evidente na face, pescoço e parte superior do tórax.
Costuma designar-se por "asfixia sexual" e desaparece, como todas as outras modificações, na fase de
resolução.
Figura 18. Orgasmo.
Módulo reprodução e manutenção da espécie
Página
25
Universidade do Minho
SOF-Fisiologia
FISIOLOGIA DA GRAVIDEZ
FERTILIZAÇÃO
Após a ovulação, o ovócito secundário é captado pela ampola da trompa uterina (o cumulus oophorus
adere aos cílios das fímbrias). É viável durante 12-24 horas, enquanto os espermatozóides podem
sobreviver até 48 horas. Todavia, dos cerca de 300 milhões de espermatozóides ejaculados, a esmagadora
maioria morre e apenas 100 chegam a entrar nas trompas uterinas.
Os espermatozóides possuem, na cabeça, uma vesícula (tipo lisossoma) de conteúdo enzimático, situada
logo acima do núcleo - o acrossoma. A interacção do espermatozóide com receptores específicos da zona
pelúcida (p. ex. a proteína ZP3) desencadeia a reacção acrossómica (figura 19), que consiste na fusão da
membrana do acrossoma com a membrana plasmática do espermatozóide, criando-se poros que permitem
a exocitose das enzimas: acrosina (uma protease, semelhante à tripsina), neuraminidase e hialuronidases
que digerem a zona pelúcida. É assim possível a fusão do espermatozóide com a membrana plasmática do
ovócito, despolarizando-a e levando à exocitose de proteases e glicosidases que alteram as glicoproteínas
da superfície celular (como a ZP3), impedindo a fertilização por outros espermatozóides - polispermia.
Figura 19. Reacção acrossómica e fertilização.
Com a fertilização, o ovócito secundário é estimulado a completar a segunda divisão da meiose, em que o
óvulo maduro fica com praticamente todo o citoplasma e o segundo corpo polar acaba por sofrer
fertilização e degenerescência. Em cerca de 12 horas, a membrana nuclear do óvulo desaparece,
permitindo a fusão com os 23 cromossomas do espermatozóide. Forma-se, assim, o zigoto, diplóide, com
46 cromossomas.
Módulo reprodução e manutenção da espécie
Página
26
Universidade do Minho
SOF-Fisiologia
Note-se que o espermatozóide contribui para o zigoto com mais do que os 23 cromossomas paternos. De
facto, pensa-se que o centrossoma, necessário para a organização do fuso mitótico do zigoto, deriva
também do espermatozóide e não do ovócito.
AS PRIMEIRAS FASES DA DIVISÃO DO ZIGOTO. IMPLANTAÇÃO.
Cerca de 30-36 horas após a fecundação, e ainda na trompa de Falópio, o zigoto divide-se, por mitose, em
duas células mais pequenas (figura 20). A clivagem do zigoto continua, produzindo: 4 células às 40 horas;
8 células às 50-60 horas, formando a mórula que entra no útero aos 3 dias. Aí permanece por 2-3 dias
durante os quais atinge as 32-64 células e se converte no blastocisto. O blastocisto consiste em duas
partes: (1) a massa celular interna, que dará origem ao feto, rodeada pelo (2) córion, que fará parte da
placenta.
Figura 20. Clivagem e evolução do zigoto.
As células que formam o córion são as células trofoblásticas. Estas apresentam, à superfície,
microvilosidades que se interdigitam com as células endometriais, formando complexos juncionais, o que
é facilitado pela existência de moléculas como a laminina e a fibronectina à superfície endometrial. As
células trofoblásticas produzem enzimas que lisam a matriz intercelular endometrial e fagocitam as
células endometriais mortas, permitindo a consolidação da implantação ou nidação (figura 21), que
habitualmente ocorre na parede dorsal do útero e se completa entre o 7º e o 10º dias.
Sob o efeito da progesterona, as células de tipo fibroblástico do estroma uterino crescem e acumulam
glicogénio, transformando-se em células deciduais, que nutrem o embrião até que a implantação produza
as necessárias conexões vasculares entre a mãe e o embrião. Funcionam, ainda, como barreira mecânica e
Módulo reprodução e manutenção da espécie
Página
27
Universidade do Minho
SOF-Fisiologia
imunológica que impede a excessiva invasão da parede uterina e como orgão endócrino, produtor de
prolactina, relaxina, prostaglandinas e factores de crescimento.
Figura 21. Implantação do blastocisto.
A implantação é muito susceptível de sofrer perturbações: em cerca de 70% das concepções há
perturbações da implantação. O bloqueio dos receptores da progesterona por fármacos como o RU 486
(mifepristona) causa aborto por impedir a implantação e a decidualização do estroma uterino.
A FORMAÇÃO DA PLACENTA. FISIOLOGIA DO LÍQUIDO AMNIÓTICO.
Entre o 7º e o 12º dias, o blastocisto é completamente envolvido pelo endométrio e o córion diferencia-se
em duas camadas celulares: o citotrofoblasto (internamente) e o sinciciotrofoblasto (externamente). A
massa celular interna também desenvolve duas camadas: a ectoderme (que dará origem à pele e sistema
nervoso) e a endoderme (que origina o sistema digestivo). A mesoderme surge mais tarde. Nesta altura, o
embrião apresenta-se sob a forma de um disco com duas camadas, separado do citotrofoblasto pela
cavidade amniótica.
À medida que o sinciciotrofoblasto invade o endométrio, segrega enzimas proteolíticas que originam
cavidades, cheias de sangue, no tecido uterino. O citotrofoblasto forma projecções, ou vilosidades
(coriónicas) que crescem para o interior dessa cavidades de sangue venoso, produzindo uma estrutura de
aparência digitiforme - o chorion frondosum. Com o crescimento do embrião, só a parte do córion voltada
para a parede uterina mantém esta estrutura. O lado oposto vai crescendo para o interior da cavidade
uterina, perde as vilosidades, tornando-se liso (chorion laeve).
Módulo reprodução e manutenção da espécie
Página
28
Universidade do Minho
SOF-Fisiologia
Uma vez que a membrana coriónica deriva do embrião que contém genes paternos que produzem
proteínas estranhas ao sistema imunitário da mãe, tem sido sugerido que o citotrofoblasto, pela produção
de ligando do FAS (um pró-apoptótico) induz a apoptose dos linfócitos T, impedindo-os, desta forma, de
atacarem a placenta. A placenta é, pois um verdadeiro local de "privilégio imunitário".
Entretanto, a nível endometrial, prossegue a decidualização, sob a estimulação da progesterona. O tecido
uterino, em contacto com o chorion frondosum é a decidua basalis. Estas duas estruturas formam, em
conjunto, a unidade funcional designada por placenta.
A placenta é contínua com a parte lisa do córion, que faz protrusão na cavidade uterina. Internamente à
membrana coriónica encontra-se o âmnios, que envolve a totalidade do embrião, encontrando-se este,
com o seu cordão umbilical, dentro de uma cavidade cheia de líquido, o saco amniótico (figura 22).
Figura 22. Estrutura do embrião e anexos
O líquido amniótico é qualitativamente semelhante ao plasma, mas diferente em termos quantitativos,
pois, relativamente a este, contém apenas 5% das proteínas e menos glicose. Os 700 mL (500-1200) de
líquido amniótico renovam-se continuamente (a cada 2 horas) e, para a sua dinâmica, concorrem:
-
Urina fetal: é um dos contribuintes mais importantes. Estudando a creatinina e o ácido úrico do
líquido amniótico obtêm-se informações sobre a maturação funcional renal.
-
Deglutições fetais: são importantes na renovação do líquido, que, desta forma, contribui também para
a nutrição fetal, pois contém proteínas e glicose.
-
Corrente traqueobrônquica: é um mecanismo tipo maré e não uma deglutição activa. Permite a
utilização da lecitina do líquido amniótico como indicador da maturidade pulmonar fetal.
Módulo reprodução e manutenção da espécie
Página
29
Universidade do Minho
SOF-Fisiologia
O líquido amniótico contém células descamadas do feto, placenta e saco amniótico. Um dos maiores
interesses do estudo dessas células (por aspiração de líquido amniótico, amniocentese, efectuada
habitualmente às 16 semanas) consiste na possibilidade de detecção de doenças genéticas, como a
trissomia 21 (síndroma de Down).
A biópsia da vilosidades coriónicas permite a detecção mais precoce (12 semanas) de doenças genéticas.
Nesta técnica, insere-se um catéter através do colo uterino até ao córion, recolhendo uma amostra das
vilosidades. Esta amostra contém muito mais células do que as obtidas por amniocentese, ultrapassando a
necessidade de cultura celular e consequente atraso na obtenção de resultados.
FUNÇÕES DA PLACENTA
As artérias umbilicais fazem chegar o sangue fetal às vilosidades do chorion frondosum. Este sangue,
depois de circular nas vilosidades, regressa ao feto via veias umbilicais. O sangue materno é transportado
e drenado das cavidades da decidua basalis, localizadas entre as vilosidades coriónicas. Desta forma, o
sangue materno e o fetal aproximam-se sem nunca se misturarem, interpondo-se uma barreira constituída
por sinciciotrofoblasto e mesênquima (figura 23).
Figura 23. Estrutura da placenta.
Módulo reprodução e manutenção da espécie
Página
30
Universidade do Minho
SOF-Fisiologia
Mecanismos de transporte através da placenta
A difusão simples através da barreira placentar permite o movimento dos gases respiratórios (O2 para o
sangue fetal e CO2 para o sangue materno), electrólitos (Na+, K+, Cl-), água e corpos cetónicos. Este
mecanismo permite apenas a passagem de moléculas com peso molecular inferior a 600-800 dalton. Dois
exemplos ilustram a importância deste facto:
-
nas mães hipocoaguladas, deve-se substituir a varfarina pela heparina que é uma molécula muito
grande, não passa a placenta, não acarretando o risco de hemorragias fetais.
-
nas mães diabéticas, a terapêutica com anti-diabéticos orais deve substituir-se pela insulina que não
passa a placenta, prevenindo-se, assim, hipoglicemias fetais.
Os catabolitos azotados são também excretados por difusão passiva, com grande facilidade: a ureia, por
ser uma molécula pequena; a creatinina devido ao elevado gradiente de concentração existente, através da
barreira placentar. Cumpre-se, desta forma, a função de emunctório placentar.
A difusão de glicose e aminoácidos através do trofoblasto é levada a cabo por moléculas transportadoras
da superfície destas células - é uma difusão facilitada. Como se trata de um mecanismo de transporte a
favor do gradiente de concentração, deve recomendar-se à mulher grávida que evite os jejuns
prolongados, prevenindo, assim, as hipoglicemias, que se repercutiriam directamente sobre o feto.
O cálcio, o fósforo, o ferro e as vitaminas são transportados activamente através da placenta. Se a mãe
sofrer uma hipocalcemia, o feto fica protegido, pois o cálcio não passará do lado fetal para o lado
materno. No caso de hipercalcemia materna, só passará para o feto o cálcio estritamente necessário,
independentemente da calcemia materna.
A pinocitose permite a passagem de substâncias de peso molecular superior a 600-800 dalton, por
exemplo, anticorpos: a IgG passa a placenta por este mecanismo, a IgM não passa e a IgA também parece
não passar.
Função endócrina da placenta
A placenta segrega hormonas esteróides e peptídeos hormonais, essenciais à gravidez normal. São de
destacar a produção de:
Gonadotrofina coriónica humana (HCG)
Estimulado pelo GnRH produzido pelo citotrofoblasto, o sinciciotrofoblasto produz HCG, uma
glicoproteína composta por subunidades α (idêntica à da TSH, LH e FSH) e β (homóloga em 80% à da
LH). A HCG pode detectar-se no plasma e urina maternos 7 dias após a concepção (a sua detecção na
urina é o teste de gravidez mais utilizado). A concentração plasmática de HCG aumenta
Módulo reprodução e manutenção da espécie
Página
31
Universidade do Minho
SOF-Fisiologia
exponencialmente até às 9-12 semanas, diminuindo posteriormente até um plateau que permanece estável
o resto da gravidez (figura 24).
Figura 24. Variações hormonais durante a gestação.
A HCG mantém a viabilidade do corpo amarelo para além dos 14 dias habituais (quando não ocorre
fertilização), conservando a secreção de estradiol e progesterona e impedindo a menstruação. Quando a
placenta se torna capaz de sintetizar esteróides, a secreção de HCG diminui.
A HCG actua sobre o feto, estimulando, a nível das supra-renais, a síntese de sulfato de
dehidroepiandrosterona (DHEA-S) e, a nível testicular, a síntese de testosterona.
No organismo materno, a HCG inibe a secreção de LH pela hipófise e, dada a sua homologia com a TSH,
estimula a actividade tiroideia, podendo induzir hipertiroidismo. A nível miometrial, a HCG inibe as
contracções produzidas pela oxitocina, mantendo a quiescência uterina.
Somatomamotrofina coriónica humana (HCS)
O sinciciotrofoblasto também segrega, logo a partir das 4 semanas de gestação, a HCS, uma hormona
estruturalmente semelhante à hormona de crescimento e com actividade lactogénea (tal como acontece
com a prolactina, também os níveis de HCS aumentam ao longo da gravidez.
A HCS estimula a lipólise materna e antagoniza as acções da insulina sobre o metabolismo glicídico,
provocando hiperglicemia. Promove, ainda, a excreção de grandes volumes de urina - poliúria. O jejum
materno e a hipoglicemia estimulam a produção de HCS, aumentando a sua concentração no plasma
materno. No sangue fetal, todavia, as concentrações de HCS permanecem muito inferiores.
Módulo reprodução e manutenção da espécie
Página
32
Universidade do Minho
SOF-Fisiologia
Hormonas esteróides
Depois das primeiras 5 semanas de gravidez, quando o corpo amarelo regride, a placenta torna-se no
principal orgão de produção de estrogénios. A concentração plasmática de estrogénios eleva-se cerca de
100 vezes relativamente aos níveis do início da gravidez. A placenta também segrega grandes quantidades
de progesterona, de forma que o ratio estrogénios/progesterona, que é de 100:1 no início da gravidez,
aproxima-se de 1 no termo.
É placenta é, contudo, um "orgão endócrino incompleto", uma vez que necessita de precursores formados
pela mãe e pelo feto para sintetizar estrogénios e progesterona. O colesterol, necessário à síntese de
progesterona, é fornecido pela circulação materna; os androgénios, necessários à síntese de estrogénios,
são fornecidos pelo feto, pois a placenta carece de 17-hidroxilase e de 17,20-desmolase, imprescindíveis à
síntese de androgénios. A placenta utiliza o DHEA-S produzido pelas supra-renais materna e fetal e
sintetiza estradiol e estrona. O estriol é quase todo sintetizado à custa de precursores fetais.
Figura 25. Síntese de esteróides placentários.
A progesterona é a hormona mais directamente responsável pelas condições de estabilidade do feto na
cavidade uterina. Estimula, nos epitélios tubar e endometrial, a secreção de nutrientes (glicogénio)
necessários à nurição do embrião - decidualização. Ao inibir as contracções uterinas (inibindo a produção
de prostaglandinas e diminuindo a sensibilidade do miométrio à oxitocina), impede a expulsão prematura
do feto. Finalmente, estimula o centro respiratório, facilitando a eliminação do CO2 materno e fetal.
Módulo reprodução e manutenção da espécie
Página
33
Universidade do Minho
SOF-Fisiologia
Tal como a progesterona, os estrogénios são inicialmente produzidos pelo corpo amarelo, sob estimulação
da HCG, até que a placenta assuma esta função. Os estrogénios estimulam o crescimento contínuo do
miométrio, preparando-o para o parto. A nível mamário, estimulam o crescimento do sistema ductal, a
partir do qual, sob estimulação da progesterona, se efectuará o crescimento alveolar. Cerca de 90% dos
estrogénios excretados na urina materna são estriol, um estrogénio pouco potente, de produção
exclusivamente placentar, pelo que, o seu doseamento, pode constituir um índice de bem-estar fetal.
Relaxina
Para além de produzir esteróides, o corpo amarelo da gravidez sintetiza, sob estimulação da HCG,
relaxina. Esta hormona inibe as contracções do miométrio (inibindo a fosforilação da cadeia leve da
miosina), relaxa os ligamentos pélvicos e a sínfise púbica, amolece e dilata o colo uterino. Permite, pois,
uma melhor acomodação da estrutura pélvica ao útero em crescimento e previne o abortamento
espontâneo, embora, na fase final da gravidez, facilite a expulsão do feto.
MODIFICAÇÕES FISIOLÓGICAS INDUZIDAS PELA GRAVIDEZ NOS DIVERSOS ORGÃOS E SISTEMAS
Sistema cardiovascular
A gravidez induz uma circulação hiperdinâmica, aumentando a frequência cardíaca (dos 70 bat./min. para
80-90 bat./min.) e aumentando o débito cardíaco em 30-50%. Este aumento do débito inicia-se pelas 6
semanas da gravidez e atinge o máximo entre as 16 e as 28 semanas. Permanece elevado até às 30
semanas, quando diminui ligeiramente porque o útero em crescimento causa obstrução da veia cava
inferior, diminuindo o retorno venoso.
À medida que a placenta e o feto se desenvolvem, o útero requer um aumento do fluxo sanguíneo, de tal
modo que, na gravidez de termo, o fluxo uterino atinge 1 L/minuto, ou seja 20% do débito cardíaco do
indivíduo normal.
A pressão arterial média desce ligeiramente durante a gravidez. A pressão arterial sistólica quase não se
modifica, mas a pressão arterial diastólica desce 5-10 mmHg, entre as 12 e as 26 semanas, voltando aos
níveis anteriores à gravidez pelas 36 semanas.
Aumentam, ainda, a frequência de sopros funcionais e a frequência de contracções auriculares e
ventriculares prematuras, que não devem ser confundidas com doença cardíaca. O coração assume uma
posição mais horizontalizada, rodando para a esquerda, o que aumenta o seu diâmetro transverso em RX
torácicos e desloca o eixo do QRS do ECG para a esquerda.
Módulo reprodução e manutenção da espécie
Página
34
Universidade do Minho
SOF-Fisiologia
Sangue
A volémia aumenta proporcionalmente ao aumento do débito cardíaco, sendo o aumento do volume
plasmático (perto de 50%) superior ao aumento da massa eritrocitária (cerca de 25%), resultando numa
diminuição da concentração de hemoglobina, por diluição - "anemia fisiológica da gravidez". A
concentração dos leucócitos aumenta (de 5000-7000/mm3 para 9000-12000/mm3) e atinge o máximo
durante o parto e primeiros dias do pós-parto.
Com o aumento da massa de glóbulos rubros, aumentam as necessidades de ferro para a produção de
hemoglobina. As necessidades de ferro podem atingir os 5-6 mg/dia na segunda metade da gravidez e, se
não se prescreverem suplementos, surge anemia por deficiência de ferro. Se não houver disponibilidade
imediata de ferro, o feto utiliza as reservas maternas, pelo que a produção de hemoglobina fetal é
preservada. A anemia fetal é, pois, uma situação rara. Acontece, porém, que a deficiência materna de
ferro pode causar parto pré-termo, abortamento espontâneo e aumentar a incidência de recém-nascidos de
baixo peso. Relativamente ao sistema da coagulação, são de salientar a hiperfibrinogenemia e o aumento
dos níveis de factor VIII, características da gravidez. Ocorrem aumentos menores dos níveis dos factores
VII, IX, X, XII. A actividade fibrinolítica encontra-se caracteristicamente deprimida.
Aparelho respiratório
A gravidez produz alterações anátomo-fisiológicas que condicionam o desempenho respiratório. Muito
precocemente, há dilatação capilar por todo o aparelho respiratório, levando ao engorgitamento da
nasofaringe, laringe, traqueia e brônquios, dificultando a respiração nasal. À medida que o útero aumenta
de volume o diafragma eleva-se (até 4 cm) e a grade costal é deslocada superiormente e alargada,
aumentando o diâmetro torácico inferior (cerca de 2 cm) e a circunferência torácica (até 6 cm). De referir
que a elevação do diafragma não impede o seu movimento. De facto, como o tónus dos músculos
abdominais diminui durante a gravidez, a respiração tende a ser, sobretudo, diafragmática.
Relativamente aos volumes e capacidades pulmonares, são de destacar: o aumento do espaço morto,
devido ao relaxamento da musculatura das vias aéreas; o aumento progressivo do volume corrente (até
35-50%); a redução das capacidades pulmonar total (devido à elevação do diafragma) e residual funcional
e dos volumes residual e de reserva expiratória.
Como resultado do aumento do volume corrente e da frequência respiratória, produz-se a chamada
"hiperventilação da gravidez", que tem como consequências a diminuição do CO2 alveolar e, logo, a
diminuição da PCO2 no sangue materno, enquanto o O2 alveolar se mantém em valores normais. A
hiperventilação materna é considerada um factor de protecção contra a exposição do feto a níveis
excessivos de CO2.
Módulo reprodução e manutenção da espécie
Página
35
Universidade do Minho
SOF-Fisiologia
Aparelho digestivo
Durante a gravidez, aumentam as necessidades vitamínicas e minerais, aumenta o apetite (embora
algumas mulheres possam referir diminuição do apetite, náuseas e vómitos - hiperemese gravídica),
sintomas que podem ser devidos à HCG.
A motilidade digestiva pode diminuir, devido aos elevados níveis de progesterona, que diminui a
produção de motilina. O aumento do tempo de trânsito dos alimentos pelo tubo digestivo promove a
reabsorção de água, provocando obstipação.
A salivação pode diminuir devido às dificuldades na deglutição associada às náuseas. Se o pH da
cavidade oral diminuir podem surgir cáries dentárias, que não se devem à falta de cálcio, pois o cálcio
dentário não é mobilizado durante a gravidez, ao contrário do que acontece com o cálcio ósseo. As
gengivas podem hipertrofiar-se, tornando-se hiperémicas e friáveis, o que se deve aos elevados níveis de
estrogénios circulantes. A deficiência de vitamina C também pode causar dor e hemorragias gengivais.
A nível esofago-gástrico, verifica-se uma diminuição do peristaltismo esofágico e uma tendência ao
refluxo gastro-esofágico, devido à lentificação do trânsito e ao relaxamento do cárdia. Para além dos
sintomas de refluxo (azia, pirose, regurgitação), estas alterações aumentam o risco anestésico pela
possibilidade de aspiração do conteúdo gástrico. O refluxo tende a piorar com o avanço da gestação, pela
compressão exercida pelo útero sobre o estômago. A produção de gastrina aumenta, levando à diminuição
do pH gástrico.
Rim e vias urinárias
O comprimento renal aumenta 1-1.5 cm, acompanhando-se do respectivo aumento de peso. Os ureteres
dilatam ao nível da porção localizada acima do rebordo pélvico. Há, pois, tendência à estase urinária e,
logo, à infecção. As causas de estase não estão bem esclarecidas, mas pensa-se que os altos níveis de
progesterona podem contribuir para a hipotonia do músculo liso do ureter. Outra hipótese sustenta que a
veia ovárica dilatada (e o ligamento suspensor do ovário, que a contém) podem exercer compressão sobre
o ureter ao nível do bordo da pelve, causando dilatação a montante.
Em termos de função renal, são de destacar o aumento da taxa de filtração glomerular (cerca de 50%),
sem grande aumento do volume urinário diário. A glicosúria não é necessariamente anormal, podendo
explicar-se pelo aumento da filtração e saturação da capacidade de reabsorção. Tem importância por
aumentar a susceptibilidade às infecções. A proteinúria altera-se pouco durante a gravidez, e adquire
importância patológica se maior do que 500 mg/dia.
Módulo reprodução e manutenção da espécie
Página
36
Universidade do Minho
SOF-Fisiologia
Sistema endócrino. Metabolismo.
Durante a gravidez normal, a mãe ganha 11-12 Kg de peso, necessitando de um acréscimo de 250-300
Kcal/dia, no aporte calórico.
Do ponto de vista metabólico, a gravidez pode ser dividida em duas fases: a primeira metade, a fase
anabólica e a segunda metade (especialmente o último trimestre), a fase catabólica em que o feto e a
placenta aumentam rapidamente de peso.
A fase anabólica caracteriza-se por uma sensibilidade normal ou aumentada à insulina, que tem como
consequências a diminuição das concentrações plasmáticas de glicose, aminoácidos, ácidos gordos e
glicerol. São inibidas a glicogenólise, a neoglicogénese e a lipólise e estimuladas a lipogénese e a
constituição de reservas hepáticas de glicogénio. Estas modificações são essenciais ao crescimento do
útero, mamas e músculo, preparando o organismo materno para as exigências da fase seguinte.
Durante a fase catabólica, há resistência à acção da insulina (estado semelhante à diabetes). A assimilação
de lipídeos, protídeos e glicídeos diminui, aumentando os níveis pós-prandiais de glicose e aminoácidos
que difundem, através da placenta, para o feto. Também a lipólise é estimulada, aumentando os níveis
plasmáticos de ácidos gordos, glicerol e cetoácidos que podem ser utilizados pela mãe e pelo feto, em vias
oxidativas, como metabolitos geradores de energia.
Outras alterações endócrinas de relevo que ocorrem durante a gravidez são: o aumento do cortisol
plasmático (como resultado do aumento de produção da globulina de ligação do cortisol, estimulada pelos
estrogénios), contribuindo para o aumento de peso e desenvolvimento mamário; o aumento da
concentração das hormonas tiroideias (por aumento da síntese da respectiva globulina de ligação), que
provoca a aceleração do metabolismo basal e da cinética cardiovascular; finalmente, o aumento dos níveis
de 1,25-(OH2)-vitamina D resultando no aumento da absorção de cálcio e supressão da paratormona.
FISIOLOGIA DO PARTO
Assim que o feto atinge um tamanho crítico, a distensão do útero e o estiramento das fibras musculares
aumentam a sua contractilidade. Várias semanas antes do fim da gravidez, o útero apresenta, pois,
episódios de contracções rítmicas, fracas e de baixa frequência - contracções de Braxton-Hicks.
Para a expulsão do feto, no trabalho de parto, são necessárias contracções mais potentes, que são
estimuladas por 2 agentes: (1) a oxitocina, uma hormona peptídica produzida pelo hipotálamo e libertada
pela neurohipófise e (2) prostaglandinas, particularmente a PGE2 e a PGF2α. (figura 26)
Embora se conheçam os estímulos para as contracções do trabalho de parto, desconhece-se o factor
responsável pelo seu início. Uma das hipóteses mais consistentes, resultante de estudos em mamíferos
Módulo reprodução e manutenção da espécie
Página
37
Universidade do Minho
SOF-Fisiologia
não-primatas, sustenta que o parto se inicia com a activação do córtex da supra-renal fetal e que o eixo
hipotálamo-hipófise-supra-renal determina o momento do início. Os corticosteróides provenientes da
supra-renal fetal estimulam a placenta a converter progesterona em estrogénios, o que é significativo,
pois, enquanto a progesterona inibe a actividade contráctil do miométrio, os estrogénios estimulam-na.
No homem, o início do trabalho de parto é mais complexo, pois a placenta não converte progesterona em
estrogénios, apenas o conseguindo fazer a partir de androgénios fetais.
Figura 26. Fisiologia do parto
A glândula supra-renal fetal não possui medula, mas o córtex divide-se numa parte externa, que produz
cortisol e uma parte interna (zona supra-renal fetal) que segrega DHEA-S. Quando o DHEA-S é
fornecido à placenta é convertido em estrogénios (sobretudo estriol) que estimulam, no útero, a produção
de receptores para a oxitocina, receptores para a prostaglandinas e de gap-junctions entre as células
miometriais. A síntese de receptores para as prostaglandinas e oxitocina torna o miométrio mais sensível
a estes agentes. As gap junctions (que funcionam como sinapses eléctricas) sincronizam e coordenam as
contracções uterinas.
Módulo reprodução e manutenção da espécie
Página
38
Universidade do Minho
SOF-Fisiologia
Todos estes acontecimentos podem ser desencadeados, segundo uma hipótese recente, pela placenta,
através da secreção da hormona libertadora da corticotrofina (CRH). A CRH estimula a adenohipófise,
que produz hormona adrenocorticotrófica (ACTH) que vai estimular a supra-renal fetal. A CRH pode
também actuar directamente em receptores para a CRH, localizados na supra-renal fetal. A estimulação,
directa ou indirecta, do córtex da supra-renal fetal, resulta na secreção de cortisol e DHEA-S.
O cortisol estimula a maturação pulmonar fetal, aumenta as reservas de glicogénio, induz enzimas
digestivas e sistemas de transporte intestinais e promove o encerramento do canal arterial. Também
participa num mecanismo de feedback positivo, estimulando a síntese placentar de CRH, aumentando,
assim, a produção de DHEA-S e do próprio cortisol. O DHEA-S é convertido em estriol que aumenta,
como já se referiu, a sensibilidade uterina às prostaglandinas e oxitocina.
As prostaglandinas promovem a entrada de cálcio nas células miometriais, desencadeando as contracções
uterinas. Relativamente à oxitocina, embora os seus níveis plasmáticos possam não aumentar, a sua
síntese local (pela decídua e membranas fetais) e os seus receptores aumentam muito no termo da
gravidez. A oxitocina reforça as contracções do parto e maximiza, particularmente, as contracções que se
seguem à expulsão do feto, minimizando as perdas hemorrágicas maternas.
As contracções uterinas também podem ser moduladas pelas catecolaminas: a estimulação α-adrenérgica
promove as contracções uterinas e a estimulação β-adrenérgica (e os agonistas β, como a ritodrina,
terbutalina e salbutamol) inibe as contracções.
Em termos hormonais, assim que se dá a expulsão da placenta todos os seus produtos desaparecem do
plasma, de acordo com as respectivas semi-vidas. Em geral, 48 a 72 horas após o nascimento, as
hormonas esteróides e os peptídeos hormonais regressam aos níveis anteriores à gravidez.
LACTAÇÃO
Desenvolvimento da mama
Cada glândula mamária é composta por 15-20 lobos, envolvidos por tecido adiposo. A quantidade de
tecido adiposo determina o tamanho e forma da mama, mas é independente da capacidade de
amamentação. Cada lobo subdivide-se em lóbulos, que contêm os alvéolos (glândulas) que segregam o
leite para os túbulos secundários. Estes convergem nos ductos mamários que, por sua vez, confluem no
canal galactóforo que se abre no mamilo.
Durante toda a gravidez, as enormes quantidade de estrogénios produzidos pela placenta estimulam o
crescimento e ramificação do sistema ductal. A insulina, o cortisol, a tiroxina, hormonas moduladoras do
metabolismo proteico, são também importantes no crescimento do sistema de ductos (figura 27).
Módulo reprodução e manutenção da espécie
Página
39
Universidade do Minho
SOF-Fisiologia
Figura 27. Regulação do desenvolvimento mamário.
Depois do desenvolvimento do sistema de ductos, a progesterona, actuando sinergicamnte com os
estrogénios (e também com todas as outras hormonas já referidas) estimula o crescimento dos alvéolos e
o desenvolvimento das características secretoras.
A produção de leite. Prolactina e sua regulação.
Apesar de os estrogénios e a progesterona promoverem o desenvolvimento físico das mamas durante a
gestação, inibem a secreção de leite. A secreção das proteínas do leite, caseína e lactalbumina, é
estimulada por outra hormona, a prolactina, segregada pela hipófise anterior. A concentração de
prolactina sobe desde a 5ª semana até ao final da gravidez, quando atinge níveis 10-20 vezes superiores
ao basal. Devido aos efeitos supressivos dos estrogénios e progesterona são segregados, até ao
nascimento, apenas alguns mililitros de líquido. O líquido segregado nos últimos dias antes do parto é o
colostro, que contém concentrações de proteínas e glicose semelhantes às do leite, mas muito pouca
gordura.
Imediatamente após o nascimento da criança, perde-se o efeito inibitório dos estrogénios e progesterona,
e o efeito lactogéneo da prolactina exerce-se em toda a sua amplitude. Porém, após o nascimento, a
secreção basal de prolactina retoma, em poucas semanas, os níveis anteriores à gravidez, ocorrendo,
apenas, picos de secreção de cada vez que a mãe amamenta.
O hipotálamo regula a secreção de prolactina, tal como faz relativamente às outras hormonas do lobo
anterior da hipófise. Todavia, a regulação da prolactina faz-se por inibição e não por estimulação, como
Módulo reprodução e manutenção da espécie
Página
40
Universidade do Minho
SOF-Fisiologia
acontece em relação às outras hormonas. O factor inibidor da secreção de prolactina é designado por
hormona inibidora da prolactina (PIH) e crê-se que seja a dopamina.
Durante os primeiros 7 a 10 dias do pós-parto, os níveis plasmáticos de FSH e LH permanecem baixos.
Depois, enquanto a secreção de LH continua inibida, a FSH sobe para níveis um pouco superiores aos da
fase folicular normal. Porém, a prolactina inibe as acções da FSH sobre o ovário e pensa-se que também
iniba a secreção de LH pelo lobo anterior da hipófise. A função reprodutora feminina é, pois, inibida pela
lactação. Com o fim da amamentação, quando baixam os níveis de prolactina (ou por administração de
um agonista dopaminérgico, como a bromocriptina), é desencadeada a libertação de LH e o consequente
início de um ciclo menstrual normal.
A ejecção do leite
O leite é segregado continuamente para dentro dos alvéolos, mas não flui livremente para o sistema ductal
e, logo, não chega ao mamilo. Tem, para tal, de ser ejectado.
A ejecção do leite é causada por um reflexo neuroendócrino, desencadeado pelo estímulo da sucção
mamilar que, conduzido pelos nervos somáticos e medula espinal, chega ao hipotálamo (figura 28). Aí, é
inibida a produção de PIH e, possivelmente, produzida uma hormona libertadora de prolactina, resultando
num pico de secreção de prolactina que estimula a secreção alveolar de leite. Por outro lado, o reflexo
condicioa a secreção de oxitocina pela hipófise posterior. A oxitocina estimula a contracção das células
mioepiteliais que circundam os alvéolos mamários e a contracção das paredes do canal galactóforo
permitindo a chegada do leite ao mamilo.
Figura 28. Reflexo neuroendócrino de ejecção do leite.
Módulo reprodução e manutenção da espécie
Página
41
Universidade do Minho
SOF-Fisiologia
A ejecção do leite pode, ainda, ser estimulada por impulsos visuais, auditivos e tácteis (p. ex., o choro do
bébé, o contacto físico). A estimulação adrenérgica (p. ex. devida a situações de ansiedade) pode inibir a
secreção de oxitocina e impedir a ejecção do leite.
Módulo reprodução e manutenção da espécie
Página
42
Universidade do Minho
SOF-Fisiologia
BIBLIOGRAFIA
1. Female reproductive system. In: Stuart Ira Fox. Human Physiology. Mc Graw Hill. 7th
editon, 2002: 656-681.
2. The reproductive glands. In: Berne RM, Levy MN, editors. Physiology. Mosby. 4th
edition, 1998: 987-1013.
3. The female reproductive system. In: Ganong, WF. Review of medical physiology.
Appleton & Lange, 18 th edition, 1997: 407-424.
4. Guyton AG, Hall JE, editors. Textbook of Medical Physiology. Philadelphia: Saunders,
10 th edition, 2000: 1017-1046.
5. Mendes da Graça, L. Medicina Materno Fetal 1 (2ª ed.). Lidel: 2000.
6. Mastroianni L, Coutifaris C. Fisiologia Reprodutiva. Vol. 1 F.I.G.O., 1988.
7. Gougeon A Regulation of ovarian follicular development in primates: facts and
hypotheses. Endocrine Reviews 17 (2): 121-155. 1996.
8. Simoni M et al. The FSH receptor: biochemistry, molecular biology, physiology and
pathophysiology. Endocrine Reviews 18(6): 739-773. 1997.
9. Nadal A et al. The estrogen trinity: membrane, cytosolic and nuclear effects. News
Physiol Sci. 16: 251-255. 2001.
10. McCann, S.M. et al. Nitric oxide controls the hipotalamic-pituitary response to
citokines. Neuroimmunomodulation. 4: 98-106. 1997.
11. Richards JS et al. Ovulation: new dimensions and new regulators of the inflammatorylike response. Annual Review of Physiology. 2002: 69-89.
Módulo reprodução e manutenção da espécie
Página
43
Universidade do Minho
Módulo reprodução e manutenção da espécie
SOF-Fisiologia
Página
44
Download