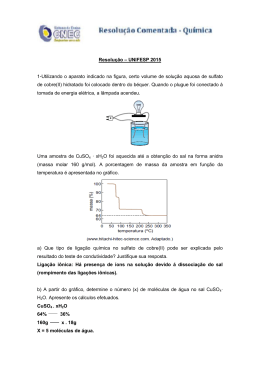
67 REVISÃO PARA QUE SERVE A CAFEÍNA EM PLANTAS? 1 Paulo Mazzafera 2,3 , Dirce Mithico Yamaoka-Yano 2,4 e Angela Pierre Vitória 2,4 Departamento de Fisiologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, CP 6109, Campinas, SP, 13081-970, Brasil. RESUMO- Cafeína é encontrada em mais de sessenta espécies vegetais e, assim como para muitos alcalóides, seu papel fisiológico ainda é desconhecido. Os poucos dados sobre o assunto foram obtidos com chá (Camellia sinensis) e café (Coffea spp.) e, em sua maioria,indicaram cafeína como um agente alelopático e anti-herbivoria. Por confrontação com outros trabalhos não relacionados especificamente com o papel fisiológico de cafeína, esses dados são discutidos e contestados. Também se discute cafeína como uma molécula armazenadora de nitrogênio e seu possível envolvimento com a resistência de doenças. Em adição, são apresentados dados indicando o não relacionamento de cafeína com a metilação do DNA em folhas de espécies de café com diferentes teores do alcalóide. Termos adicionais para indexação: alelopatia, Camellia sinensis, Coffea, metilação de DNA, resistência a doenças, resistência a insetos. DOES CAFFEINE PLAY ANY ROLE IN PLANTS ? ABSTRACT- Caffeine is found in more than sixty plant species and as for many other alkaloids its physiological role is unknown. Scanty data from the literature on the subject refer to tea (Camellia sinensis) and coffee (Coffea spp), and in most cases suggest caffeine as being either an allelopathic agent or an anti-herbivory compound. Through comparing such data with other information not specifically related to the physiological role of caffeine, we dismiss the role of caffeine as an allelopathic or 1 Recebido em 08/09/1995 e aceito em 26/01/1996. 2 Bolsistas do CNPq (P.M., pesquisa; D.M.Y.Y, pós doutorado) e CAPES (A.P.V., doutorado). 3 Prof. Livre Docente, autor para correspondência, fax 0192 393124, e-mail [email protected] 4 D.M.Y.Y. - Bióloga, Pós doutorando; A.PV. - Bióloga, doutorando. anti-herbivory compound. The role of caffeine as a nitrogen-storage compound and its involvement in resistance against diseases is also discussed. In addition, new data are presented showing the lack of a relationship between caffeine and DNA methylation in young coffee leaves. Additional index terms: allelopathy, Camellia sinensis, Coffea, DNA methylation, disease resistance, insect resistance. INTRODUÇÃO Várias revisões mostram a dificuldade em se estabelecer o papel dos alcalóides em plantas (Robinson, 1968,1974; Waller & Nowacki, 1978). Entre as várias propostas apresentadas, duas têm merecido maior atenção: que alcalóides têm efeito alelopático e que atuam como agentes protetores contra a predação por herbívoros. Apesar dos muitos trabalhos mostrando algum efeito alelopático ou anti-herbivoria de um dado alcalóide, tais funções não podem ser ainda aplicadas para toda a classe. Além disso, alguns trabalhos mostram que um determinado alcalóide pode ter uma dessas funções em determinadas situações, mas não em outras (Waller & Nowacki, 1978). Dessa forma, cada caso deve ser analisado separadamente. Outra função apontada é a de que alcalóides poderiam atuar como moléculas armazenadoras de nitrogênio (Waller & Nowacki, 1978), tal como asparagina, glutamina e os ureídeos, ácido alantóico e alantoína. Cafeína (1,3,7-trimetilxantina) é um alcalóide presente em um número não muito grande de espécies, totalizando um pouco mais de sessenta (Suzuki & Waller, 1988). Plantas contendo cafeína têm distribuição ampla no globo terrestre, podendo ser encontradas desde regiões de clima frio ( Erodium Escandinávia; chá-mate Ilex cícutarum, paraguariensis, Argentina) até tropical (cacaueiro - R. Bras. Fisiol. Veg., 8(1):67-74, 1996 68 Theobroma cacao, Amazônia; guaraná - Paullinia cupana, Amazônia; café - Coffea spp, África). tecidos analisados tivessem sido expostos ao ataque de algum inseto. Dos poucos trabalhos em que algum papel fisiológico foi atribuído à cafeína, a maioria foi feita com chá e café. Nesse artigo, estas informações serão discutidas com principal atenção ao cafeeiro, e dados de diferentes trabalhos serão confrontados para dar suporte a algumas conclusões. Dados inéditos também serão apresentados. Apesar de já ter sido comunicado na literatura, o ataque de cafeeiros por formigas cortadeiras do gênero Atta não é de ocorrência frequente (Oliveira Filho, 1934). Porém, Mazzafera (1991) observou que em certos cafeeiros frequentemente atacados por saúvas, entre vários compostos analisados nas folhas que poderiam ter características repelentes ou atrativas, somente o baixo teor de cafeína diferenciava essas plantas de outras não atacadas. Cafeina como um inseticida natural Poucos trabalhos investigaram cafeína como composto protetor contra o ataque de insetos. Em Callosobruchus chínensis, uma broca de grãos, cafeína causou l00% de esterilidade quando adicionada à dieta artificial, na concentração de 1,5% (Rizvi et al., 1980b). Nathanson (1984) usou plantas de tomate para estudar o efeito pesticida de cafeína contra a lagarta Manduca sexta. Folhas de tomateiros aspergidas com soluções de cafeína foram usadas para alimentar larvas de Manduca sexta. À concentração de 0,3%, houve 60% de redução da área foliar comida; à 1%, mais de 80% de redução ocorria. Adição de pó de café ou cafeína pura à dieta artificial levaram à diminuição do crescimento das lagartas. Analisando gemas apicais e folhas emergentes de café (Coffea arabica), Frischknecht et al. (1986) detectaram que cafeína representava até 4% da massa seca. Com o crescimento ocorria decréscimo no nível do alcalóide. Estimou-se que nesses tecidos, em termos de alocação de carbono, a cafeína representava 15% do carbono usado na respiração. Os autores argumentaram que, nesse estádio de desenvolvimento, o tecido foliar é bastante tenro, não oferecendo nenhum tipo de resistência mecânica ao ataque de insetos e, em contrapartida, cafeína atuaria como uma defesa química, justificando o gasto metabólico. No entanto, um forte argumento contra a interpretação dada por aqueles autores é que tecidos jovens têm grande atividade metabólica de síntese, ou seja, alta taxa de transcrição, apresentando um "pool" maior de nucleotídeos e com alta taxa de reciclagem. Uma vez que cafeína é sintetizada a partir de adenina e guanina, constituintes desse "pool", seria esperado que tecidos jovens de café tivessem alto conteúdo desse alcalóide (Suzuki & Waller, 1984b). De fato, os maiores teores de cafeína são encontrados em tecidos jovens (Herndlhofer, 1933; Raju & Gopal, 1979; Suzuki & Waller, 1984b). Assim, a hipótese apresentada por Frischknecht et al. (1986) somente poderia ser validada se os Apesar disso, o autor avalia com cautela os resultados, pois ataques de saúvas a cafeeiros com alto teor de cafeína nas folhas já haviam sido constatados (P. Mazzafera, dados não publicados). Em nenhum dos quatro trabalhos citados avaliou-se o efeito da cafeína, considerando-se o cafeeiro e uma praga específica dessa planta. Levando-se em conta que existe variabilidade dentro do gênero Coffea para o teor de cafeína, se ela repele insetos, deveria haver, então, variação do ataque por uma determinada praga. Assim, pode-se considerar que os dados citados não são fortes indicadores de que a cafeína atue como repelente ou que tenha característica de inseticida natural, como sugerido por Nathanson (1984). TABELA 1- Conteúdo de cafeína em folhas de espécies de café apresentando diferentes níveis de resistência ao bicho mineiro. Espécies de café Cafeína (%) Resistência C. stenophylla 0.03 muito resistente C. brevipes ND muito resistente C. liberica 0.02 muito resistente C. salvatrix 0.O1 muito resistente C. racemosa 0.03 resistente C. kapakata ND resistente C. dewevrei 0.02 resistente C. eugenioides ND resistente C. congensis 0.32 suscetível C. canephora 0.45 suscetível C. arabica 0.98 muito suscetível ND = não detectada Duas são as principais pragas atacando o cafeeiro no Brasil. Hypothenemus hampei , conhecida como a broca do café, ataca os frutos do café, comendo a semente; Perileucoptera coffeela, R. Bras. Fisiol. Veg., 8(1):67-74, 1996 69 conhecido como bicho mineiro, é um minador de folhas, comendo o mesófilo foliar. Com base nos dados de Guerreiro Filho et al. (1991), que avaliaram a resistência de espécies de café ao bicho mineiro, com os de Mazzafera e Magalhães (1991), que determinaram o conteúdo de cafeína em folhas das mesmas plantas dessas espécies, pode-se construir a tabela 1. Aparentemente, a primeira conclusão que se tem é de que cafeína não determinaria a resistência, uma vez que a relação entre elas é inversa. TABELA 2- Teor de cafeína nas sementes de espécies de café com diferentes níveis de resistência à broca do café. Espécies de café Cafeína (%) Resistência Referências em Montalvo & Botero (1987) C. canephora 1.710 24 C. liberica 0. 021 C. canephora menos atacada que C. liberica C. excelsa 1.205 C. liberica 0.021 272 C. abeokutae 1.322 C . excelsa, C. liberica, C. abeokutae menos atacadas que C. canephora C. canephora 1.710 C. canephora 1.710 126 C. liberica 0.021 C. canephora menos atacada que C. liberica No entanto, alguma evidência do efeito de cafeína na infestação do cafeeiro pelo bicho mineiro foi obtida por Guerreiro Filho (1994), que avaliou o teor de cafeína em folhas de cafeeiros resistentes e suscetíveis de uma população de híbridos entre C. arabica (suscetível) e C. racemosa (resistente). Uma das folhas de cada par de folhas foi exposta à infestação, sendo a outra protegida contra a oviposição pelos insetos adultos. Nas plantas resistentes a concentração do alcalóide foi superior nas folhas infestadas. 0 autor concluiu que cafeína poderia estar envolvida de alguma forma na resistência do cafeeiro ao bicho mineiro. A outra praga de importância econômica atacando o cafeeiro é a broca dos grãos. Os dados sobre o conteúdo de cafeína em sementes de várias espécies de café obtidos por Mazzafera e Magalhães (1991) e de outros autores (ver referências em Montalvo & Botero, 1987) que estudaram o ataque da broca do café em espécies de café originaram a tabela 2. Em nenhum caso examinado foi detectada imunidade ao ataque da broca, e em todos eles a resistência de uma dada espécie era relativa às outras espécies estudadas. Também aqui não se chega a alguma conclusão sobre a relação entre cafeína e resistência do cafeeiro ao ataque da broca. Cafeína como reserva de nitrogênio A cafeína possui 4 átomos de N, quase 29% da molécula, e uma relação C/N = 2. Com isto, ela poderia ter a função de armazenar nitrogênio, a exemplo do que ocorre com glutamina e asparagina (C/N = 2), e os ureídeos alantoína e ácido alantóico (C/N = 1) em leguminosas tropicais, compostos orgânicos contendo nitrogênio que são predominantemente translocados na planta. Weevers (1930) observou que folhas velhas do cafeeiro apresentavam teor reduzido de cafeína, sugerindo que, ao ser degradada, o nitrogênio da molécula poderia ser reaproveitado. Esta hipótese recebeu maior crédito quando Hamidi & Wanner (1964) não detectaram cafeína em folhas velhas de café, contradizendo, porém, os dados de Herhdlhofer (1933), que detectou o alcalóide em todas as partes do cafeeiro. As diferenças entre os dados desses autores provavelmente seriam metodológicas, pois com métodos de análise mais modernos (cromatografia líquida de alta eficiência), pode-se confirmar a presença de cafeína em folhas com mais de um ano de idade (P. Mazzafera, dados não publicados). Em chá foi observado que durante o ano o teor de cafeína variava em função do nível de nitrogênio na planta, refletindo a dependência de sua síntese em função desse elemento (Cloughley,1982; Suzuki & waller, 1986). Tais variações eram acompanhadas pelas atividades das enzimas envolvidas na sua via de síntese (Fujimori et al., 1991). Em café, a relação cafeína-nitrogênio também foi observada (Beaudin-Dufour & Müller, 1971). Atuando como molécula armazenadora de N, a cafeína poderia ser degradada localmente ou translocada para outras partes da planta, liberando posteriormente o nitrogênio contido nela. Experimentos em que cafeína com dupla marcação por isótopos (14C e 15N) foi fornecida a folhas de café em crescimento, não se observou translocação para frutos posicionados no mesmo nó foliar e muito pouco para a folha do mesmo nó foliar (Baumann & Wanner, 1972). Na célula, cafeína se encontra complexada com potássio e ácido clorogênico (Horman & Viani, 1972; Martin et al., 1987). Porém, cafeína também é encontrada no espaço intercelular (Pfrunder et al., 1980), e folhas do quinto e sexto nós de ramos plagiotrópicos apresentam mais cafeína intercelular do que folhas do primeiro ou segundo par (A.P. Vitória e P. Mazzafera, dados não publicados). R. Bras. Fisiol. Veg., 8(1):67-74, 1996 70 Experimentos com calos de café crescidos em meio de cultura e mantidos no escuro mostraram que a cafeína era liberada no meio de cultura em grande quantidade (Baumann & Gabriel, 1984; Kurata & Furusaki, 1993). Estes resultados foram explicados com base na dependência de luz da enzima fenilalanina amônia-liase. Uma vez inibida sua atividade, não há síntese de ácido clorogênico, e como conseqüência, diminui sua ligação com cafeína. Acredita-se que a complexação de cafeína com o ácido clorogênico seja o artifício responsável pela retenção do alcalóide no interior da célula. Dessa forma, a maior quantidade de cafeína intercelular em folhas mais velhas poderia ser reflexo da degradação de ácido clorogênico. Gabriel, 1984; Friedman & Waller, 1983a; Suzuki & Waller, 1987). Desde que quase não ocorre o transporte de cafeína entre orgãos de uma mesma planta, a cafeína intra e intercelular poderia ser metabolizada localmente para liberação do nitrogênio contido nela. Em frutos e folhas de café e chá, a cafeína pode ser degradada completamente até amônia (Suzuki & Takahashi, 1975; Suzuki & Waller, 1984a, 1984b). 0 fornecimento de cafeína marcada com radioisótopos mostrou, porém, que em várias espécies de café a taxa de degradação de cafeína é praticamente nula, em períodos relativamente curtos (Kalberer, 1965; Mazzafera, 1993; Mazzafera et al., 1991; Mazzafera et al., 1994). Waller et al. (1986) observaram que a adição de cafeína a solo coletado sob a saia do cafeeiro era facilmente recuperada. Por outro lado, em solo coletado a dois metros de distância da planta a recuperação era bastante baixa, mesmo utilizando condições drásticas na extração. A conclusão dos autores foi de que, sendo a cafeína fortemente retida pelas argilas do solo (Lailach et al., 1968), no solo coletado sob a planta haveria saturação das argilas pelo alcalóide, o contrário ocorrendo naquele coletado a dois metros. Isto explicaria a fácil recuperação no primeiro solo. Apesar de sementes de café e de chá conterem considerável quantidade de cafeína, vários trabalhos mostraram que durante a germinação este alcalóide não atuaria como fator nutritivo (Baumann & Gabriel, 1984; Mazzafera, 1990; Suzuki & Waller, 1987). Cafeína como agente alelopático Os primeiros resultados indicando o efeito alelopático de cafeína foram obtidos colocando-se sementes de Amaranthus spinosus para germinar em extratos obtidos a partir de folhas de quarenta e nove plantas (Rizvi et al., 1980a). Após fracionamento com vários solventes orgânicos, a fração clorofórmio de folhas de café (Coffea arabica), que havia sido seca e solubilizada em água, foi a que causou maior inibição na germinação. Identificou-se cafeína como o principal componente dessa fração (Rizvi et al., 1981). Posteriormente, vários trabalhos confirmaram o efeito alelopático de cafeína (Chou & Waller, 1980; Shettel & Balke, 1983; Smyth, 1992; Waller et al., 1986). Outros estudos realizados em laboratório demonstraram que sementes de chá e de café liberam cafeína durante a germinação, podendo, inclusive, causar autoinibição da germinação (Baumann & Acredita-se que, em café, a primeira divisão mitótica da radícula aconteça somente após sua protusão do endosperma, constituindo-se como um mecanismo para evitar o contato com o alcalóide (Friedman & Waller, 1983a) . Sementes de café que caem da planta e permanecem no chão poderiam ser uma das fontes de cafeína no solo (Friedman & Waller, 1983b). Outras fontes seriam a cafeína proveniente da lavagem das folhas pela água das chuvas e aquela liberada pela deterioração de folhas caídas e outras partes da planta de café (Waller et al., 1986) . Waller et al. (1986) argumentaram também que, na projeção da copa do cafeeiro, a cafeína ficaria livre e poderia levar a autointoxicação da planta, explicando-se, portanto, a depauperação de plantas entre 10 e 25 anos de idade. No solo, os compostos liberados por plantas podem ser modificados devido a reações químicas como também a sua transformação pelo ataque de microrganismos (Rice, 1984). No entanto, a hipótese de que cafeína poderia ser degradada no solo foi considerada pouco provável por Waller et al. (1986). Segundo os autores, a cafeína poderia sim, inibir o desenvolvimento de microrganismos, acumulando-se mais ainda no solo. Bactérias capazes de degradar cafeína já foram isoladas de solos que nunca reberam cafeína (Blecher & Lingens, 1977; Woolfolk, 1975), utilizando-se uma estratégia metodológica denominada enriquecimento. Nela, cultiva-se a bactéria em meio rico em nutrientes, mas incrementado com baixa concentração de cafeína. Após várias transferências para o mesmo meio, mutantes com capacidade de degradar cafeína podem ser isolados. Recentemente, bactérias capazes de degradar cafeína foram isoladas de solos cultivados com café, sem a necessidade de enriquecimento. Amostras desses solo foram incubadas por 24 horas em meio mineral líquido contendo cafeína como única fonte de R. Bras. Fisiol. Veg., 8(1):67-74, 1996 71 FIGURA 1- Bandeamento de DNA de folhas jovens de C. arabica e C. dewevrei digeridos com as seguintes combinações de enzimas de restrição: 1 - ECO RI/ALU I; 2 - ECO RI/Hae III; 3 - ECO RI/Hpa II; 4 - Bam HI/Alu I; 5 - Bam HI/Hae III; 6 - Bam HI/Hpa II. Da esquerda para a direita, a primeira coluna mostra o padrão de DNA (fragmentos de 12.200 a 140 pares de bases), a segunda o DNA de C. arabica e a terceira o de C. dewevrei. carbono e nitrogênio (Yamaoka-Yano & Mazzafera, 1996), ou simplesmente em água (Mazzafera et al., 1996), e depois semeadas em meio mineral sólido com cafeína. Além de Serratia marcescens (Mazzafera et al., 1996), outros dezenove isolados foram obtidos (Yamaoka-Yano & Mazzafera, 1996), sendo oito deles identificados como Pseudomona putida. Pseudomonas é o gênero de bactérias mais comumente encontrado no solo, e isto se deve a sua versátil capacidade de degradar mais de 80 substâncias, garantindo-lhe o crescimento nos mais variados tipos de substrato (Clarke & Slater, 1986). Porém, nem todos os genes envolvidos na degradação desses compostos são expressos ao mesmo tempo, o que se constituiria num gasto energético muito grande. Eles permaneceriam silenciados até serem transcritos devido a um estímulo ocasionado pelo substrato (Bennet & Richmond, 1978; Clarke & Slater, 1986; Unger et al., 1986). Este estímulo também parece ocorrer no caso de cafeína e Pseudomonas (Blecher & Lingens, 1977). Quando a semeadura em meio com alcalóide era feita com inóculo crescido em meio nutritivo rico sem cafeína, o crescimento inicial era mais lento do que quando usado inóculo produzido em meio com cafeína. Waller et al. (1986) não conseguiram extrair cafeína "endógena" do solo coletado sob a copa do cafeeiro. A confirmação do papel de bactérias na degradação de cafeína foi obtida por Yamaoka-Yano & Mazzafera (1996), que adicionaram quantidades conhecidas do alcalóide em amostras de solo. Duas duplicatas foram feitas, uma esterilizada em autoclave e outra não. Após uma semana de incubação a quantidade de cafeína tinha caído drasticamente na duplicata não estéril. Outras investígações sobre o papel fisiológico da cafeina Quando infectadas, plantas de café suscetíveis ao fungo Hemileia vastatrix têm aumentada a concentração de cafeína nas folhas (Medeiros et al., 1989). Semelhantemente, o mesmo ocorreu no caso de cafeeiros suscetíveis infectados por Meloidogyne incognita (Gonçalves et al., 1995). Cafeína não poderia estar envolvida na resistência, uma vez que em ambos os casos ocorreu o estabelecimento do patógeno na planta. Muito provavelmente, a explicação para esse fato é similar à justificativa dada para o maior teor de cafeína em tecidos novos no cafeeiro, ou seja, ela seria decorrente do aumento do metabolismo geral da planta em resposta à infecção, aumentando, portanto, o "pool" de nucleotídeos derivados da reciclagem de m RNA. É conhecido que no DNA de plantas, a metilação de citosina, formando 5-metilcitosina, leva a repressão de genes (Hepburn et al., 1987; Quemada et al., 1987). Tendo a cafeína três radicais metil e considerando-se que em tecidos novos sua degradação é mais intensa (Mazzafera et al., 1994; Suzuki & Waller, 1984a), investigou-se a possibilidade R. Bras. Fisiol. Veg., 8(1):67-74, 1996 Portanto, existe grande possibilidade de que os isolados de Pseudomonas obtidos por Yamaoka-Yano & Mazzafera (1996) já estivessem expressando os genes necessários para a degradação de cafeína. A degradação por microrganismos explicaria porque 72 de haver alguma relação entre o teor do alcalóide e a CLARKE, P.H. & SLATER, J.H. Evolution of enzymes structure and function in Pseudomonas . In: metilação do DNA. Folhas do primeiro par de folhas de SOKATCH, J.R.E ed. The Bacteria: A treatise on cafeeiros C. arabica e C. dewevrei, com 0,9%e 0,2% structure and function. London, Academic Press, de cafeína, respectivamente (Mazzafera et al., 1991), 1986. v. 10, p.71-144. tiveram o DNA extraído e digerido com enzimas de restrição, em combinação duas a duas, que CLOUGHLEY, J.B. Factors influencing the caffeine content of black tea: par t 1 - the effect of field reconhecem pontos de metilação no DNA. Os variables. Food Chemistry, 9:269-276, 1982 . fragmentos originados foram submetidos a eletroforese em gel de agarose. Em todas as FRIEDMAN, J. & WALLER, G.R. Caffeine hazards and combinações de enzimas, o padrão eletroforético foi o their prevention in germinating seeds of coffee (Coffea arabica L.). Journal of Chemical Ecology, mesmo para as duas plantas (Fig. 1). 9:1099-1106, 1983a. 0 não envolvimento de cafeína com a metilação do FRIEDMAN, J. & WALLER, G.R. Seeds as allelopathic DNA foi confirmado crescendo-se calos de folhas de agents. Journal of Chemical Ecology, C. arabica em meio MS estéril (Söndahl et al., 1984), 9:1107-1117, 1983b. contendo cafeína (2mM), 5-azacitosina (5µM), ou os dois juntos. Acima daquela concentração, cafeína FRISCHKNECHT, P.M.; BAUMANN, T.W. & WANNER, H. Tissue culture of Coffea arabica. Growth and caffeine passa a inibir o crescimento de calos de café formation. Planta Medica, 31:344-350, 1977. (Frischknecht et al., 1977). 5-Azacitosina é um análogo de citosina e incorpora-se ao DNA, inibindo a FRISCHKNECHT, P.M.; ULMER-DUFEK, J. & BAUMANN, T.W. Purine alkaloid formation in buds ação da DNA-metiltransferase (Hepburn et al., 1987). and developing leaflets of Coffea arabica : Como tratamento controle, não adicionou-se cafeína Expression of an optimal defence strategy? ou 5-azacitosina no meio de cultura. 0 DNA foi Phytochemistry, 25:613-616, 1986. extraído desses calos, digerido com ácido fórmico e as quantidades de citosina e 5-azacitosina FUJIMORI, N.; SUZUKI, T. & ASHIHARA, H. Seasonal variations in biosynthetic capacity for the synthesis determinadas (Klaas et al., 1989). As proporções of caffeine in tea leaves. Phytochemistry, citosina/5-metilcitosina observadas foram: tratamento 30:2245-2248, 1991. controle - 6,2+0,28, tratamento com cafeína 7,14+0,14, tratamento com 5-azacitosina - 9,7+0,14, GONÇALVES, W.; MAZZAFERA, P.; FERRAZ, L.C.C.B.; SILVAROLLA, M.B. & LIMA, M.M.A. Biochemical tratamento com cafeína e 5-azacitosina = 10,62+2,38. basis of coffee tree resistance to Meloidogyne Portanto, houve inibição da metilação do DNA por incognita. Plantation Recherche Développement, 5-azacitosina, mas a cafeína não reverteu este efeito. 2:54-60, 1995. GUERREIRO FILHO, 0. Identification de genes de REFERÊNCIAS résistance à Perileucoptera coffeella en vue de BAUMANN, T.W. & GABRIEL, H. Metabolism and 1'amélioration de Coffea arabica: Potentiel excretion of caffeine during germination of Coffea d'espèces diploides du genre Coffea ; genes de L. Plant and Cell Physiology, arabica Bacillus thuringiensis . Montpellier, ENSAM, 1994. 25:1431-1436, 1984. 185p. Tese de Doutorado. BAUMANN, T.W. & WANNER, H. Untersuchungen uber GUERREIRO FILHO, O.; MEDINA FILHO, H.P. & den transport von kaffein in der kaffeepflanze (Coffea CARVALHO, A. Fontes de resistência ao bicho arabica). Planta, 108:11-19, 1972 . mineiro, Perileucoptera coffeela, em Coffea spp . Bragantia, 50:45-55, 1991. BEAUDIN-DUFOUR, D. & MÌÜLLER, L.E. Effet de la radiation solaire et de 1'âge sur le contenu en HAMIDI, A. & WANNER, H. The distribution pattern of caféine et en azote des feuilles et des fruits de trois chlorogenic acid and caffeine in Coffea arabica. espèces de caféiers. Turrialba, 21:387-392, 1971. Planta, 61:90-96, 1964. BENNET, P.M. & RICHMOND, M.H. Plasmids and their HEPBURN, A.G.; BELANGER, F.C. & MATTHEIS, J.R. possible influence on bacterial evolution. In: DNA methylation in plants. Developmental ORNSTON, L.N. & SOKATCH, J.R.E ed. The Genetics, 8:475-493, 1987. bacteria: A treatise on structure and function, HERNDLHOFER, E. A distribuição das proteínas, da London, Academic Pres, 1978. v. 6, p.1-69. cafeína dos mono-amino-ácidos e dos BLECHER, R. & LINGENS, F. The metabolism of caffeine di-amino-ácidos no cafeeiro e as variações da by a Pseudomonas putida strain. Hoppe-Seyler's porcentagem destas substâncias no percurso de um Zeitschrift für Physiologische Chemmie, ano. Boletim de Agricultura de São Paulo, 358:807-817, 1977. 34:163-251, 1933. CHOU, C.-H. & WALLER, G.R. Possible allelopathic HORMAN, I. & VIANI, R. The nature and confirmation of constituents of Coffea arabica. Journal of Chemical the caffeine-chlorogenate complex of coffee. Journal Ecology, 6:643-654, 1980. of Food Science, 37:925-927, 1972. R. Bras. Fisiol. Veg., 8(1):67-74, 1996 73 KALBERER, P. Breakdown of caffeine in the leaves of Coffea arabica L. Nature , 205:597-598, 1965. KLAAS, M.; JOHN, M.C.; CROWELL, D.N. & AMASINO, R.M. Rapid induction of genomic demethylation and T-DNA gene expression in plant cells by 5-azacytosine derivatives. Plant Molecular Biology, 14:413-423, 1989. KURATA, H. & FURUSAKI, S. Immobilized Coffea arabica cell culture using a bubble-column reactor with controlled light intensity. Biotechnology and Bioengeneering, 42:494-502, 1993. LAILACH, G.E.; THOMPSON, T.D. & BRINDLEY, G.W. Absorption of pyrimidines, purines, and nucleosides by Li-, Na-, Mg-, and Ca-Montmorillonite (clay-organic studies XII). Clays and Clay Minerals, 16:286-293, 1968. MARTIN, R.; LILLEY, T.H.; FALSHAW, C.P.; HASLAM, E., BEGLEY, M.J. & MAGNOLATO, D. The caffeine-potassium chlorogenate molecular complex. Phytochemistry, 26:273-279, 1987. MAZZAFERA, P. Estudo sobre o papel da cafeína em plântulas de café (Coffea arabica L.). Revista Brasileira de Botânica, 13:97-102, 1990. MAZZAFERA, P. Análises químicas em folhas de cafeeiros atacados por Atta spp . Revista de Agricultura, 66:33-45, 1991. MAZZAFERA, P. 7-methylxanthine is not involved in caffeine catabolism in Coffea dewevrei. Journal of Agricultural and Food Chemistry, 41:1541- 1543, 1993. MAZZAFERA, P. & MAGALHÃES, A.C.N. Cafeína em folhas e sementes de Coffea e Paracoffea . Revista Brasileira de Botânica, 14:157-160, 1991. MAZZAFERA, P.; CROZIER, A. & MAGALHÃES, A.C. Caffeine metabolism in Coffea arabica and other species of coffee. Phytochemistry, 30:3913-3916, 1991. MAZZAFERA, P.; CROZIER, A. & SANDBERG, G. Studies on the metabolic control of caffeine turnover in developing endosperms and leaves of Coffea and Coffea dewevrei . Journal of arabica Agricultural and Food Chemistry, 42:1423, 1994. MAZZAFERA, P.; OLSSON, 0. & SANDBERG, G. Degradation of caffeine and related methylxanthines by Serratia marcescens isolated from soil under coffee cultivation. Microbial Ecology, 31:199-207, 1996. MEDEIROS, M.A.P.X.L.; GUEDES, M.E.M. & SOUSA, M.L.B. Has caffeine a role in the resistance of coffee to orange rust? In: COLLOQUE INTERNATIONAL SUR LA CHIMIE DES CAFÉS VERTS, TORRÉFIÉS ET LEURS DÉRIVÉS, 13, 1989, Paipa, Colômbia, Association Scientifique International du Café, 1989. p. 733-744. MONTALVO, L.A.M. & BOTERO, M.P.M. La Broca de la Cereza del Cafeto ( Hypothenemus hampei Ferrari 1867): Resúmenes Analiticos. Chinchiná, Federacion Nacional de Cafeteros de Colombia -Cenicafé, 1987. NATHANSON, J. A. Caffeine and related methylxanthines: possible naturally occurring pesticides. Science, 226:184-187, 1984. OLIVEIRA FILHO, M.L. 0 problema da sauva. Revista do Instituto do Café, 9:110-113, 1934. PFRUNDER, R.; WANNER, H.; FRISCHKNECHT, P. M. & BAUMANN, T. W. An attempt to localize caffeine in the cell by its washout kinetics. In: COLLOQUE INTERNATIONAL SUR LA CHIMIE DES CAFÉS VERTS, TORRÉFIÉS ET LEURS DÉRIVÉS, 9, 1980, London. Association Scientifique International du Café, 1980. p. 169-175. QUEMADA, H.; ROTH, E.J. & LARK, K.G. Changes in methylation of tissue cultured soybean cells detected by digestion with the restriction enzymes Hpa II and Msp I. Plant Cell Reports, 6:63-66, 1987. RAJU, K.I. & GOPAL, N.H. Distribution of caffeine in arabica and robusta coffee plants. Journal of Coffee Research, 9:83-90,1979. RICE, E.L. Alellopathy, 2 ed. Orlando, Academic Press, 1984. p. 422 . RIZVI, S.J.H.; MUKERJI, D. & MATHUR, S.N. A new repor t on a possible source of natural herbicide. Indian Journal of Experimental Biology, 18:777-778, 1980a. RIZVI, S.J.H.; PANDEY, S.K.; MUKERJI, D. & MATHUR, S.N. 1,3,7-Trimethylxanthine, a new chemosterilant for stored grain pest - Callosobruchus chinensis. Zeitschrift für Angewandte Entomologie, 90: 777-778, 1980b. RIZVI, S.J.H.; MUKERJI, D. & MATHUR, S.N. Selective phyto-toxicity of 1,3,7-trimethylxanthine between Phaseolus mungo and some weeds. Agricultural and Biological Chemistry, 45:1255-1256, 1981. ROBINSON, T. The biochemistry of alkaloids. Berlin, Springer-Verlag, 1968. p.149. ROBINSON, T. Metabolism and function of alkaloids in plants. Science, 184:430-435, 1974. SHETTEL, N.L. & BALKE, N.E. Plant growth response to several allelopathic chemicals. Weed Science, 31:293-298, 1983. SMYTH, D.A. Effect of methylxanthine treatment on rice seedling growth. Journal of Plant Growth Regulation, 11:125-128, 1992. SÖNDAHL, M.R.; NAKAMIJRA, T.; FILHO, H.P.M.; CARVALHO, R.; FAZUOLI, L.C. & COSTA, W.M. Coffee. In: AMMIRATO, P.V.; EVANS, D.A.; SHARP, W.R. & YAMADA, Y.E. ed. Handbook of Plant Cell Culture. Crop Sciences. New York, Macmillan, 1984. v.3, p.564-590. SUZUKI, T. & TAKAHASHI, E. Metabolism of xanthine and hypoxanthine in the tea plant ( Thea sinensis L.). Biochemical Journal, 146:79-85, 1975. R. Bras. Fisiol. Veg., 8(1):67-74, 1996 74 SUZUKI, T. & WALLER, G.R. Biodegradation of caffeine: formation of theophylline and theobromine from caffeine in mature Coffea arabica fruits. Journal of the Science of Food and Agriculture, 35:66-70, 1984a. SUZUKI, T. & WALLER, G.R. Biosynthesis and biodegradation of caffeine, theobromine, and theophylline in Coffea arabica L. fruits. Journal of Agricultural and Food Chemistry, 32:845-848, 1984b. SUZUKI, T. & WALLER, G.R. Total nitrogen and purine allaoids in the tea plant throughout the year. Journal of the Science of Food and Agriculture, 37:862-866, 1986. SUZUKI, T. & WALLER, G.R. Allelopathy due to purine alkaloids in tea seeds during germination. Plant Soil, 98:131-136, 1987. SUZUKI, T. & WALLER, G.R. Metabolism and analysis of caffeine and other methylxanthines in coffee, tea, cola, guarana and cacao. In: LINSKENS, H.F. & JACKSON, J.F.E. ed. Analysis of Nonalcoholic Beverages. Berlin, Springer-Verlag, 1988. p.184-220. UNGER, B.P.; SLIGAR, S.G. & GUNSALUS, I.C. Pseudomonas cytochromes P-450. In: SOKATCH, J.R.E. ed. The Bacteria: A treatise on structure and function, London, Academic Pres, 1986. v. 10, p.557-589. WALLER, G.R.; KUMARI, D.; FRIEDMAN, J.; FRIEDMAN, N. & CHOU, C.H. Caffeine autotoxicity in Coffea arabica L. In: PUTNAM, A.R. & TANG, C.-S.E. ed. The Science of Allelopathy. New York, John Wiley, 1986. p.243-269. WALLER, G.R. & NOWACKI, E.K. Alkaloid Biology and Metabolism in Plants. New York, Plenum Press, 1978, 293p. WEEVERS, T. Die Funktion der Xanthinderivative im Pflanzenstoffwechsel. Archives Néerlandaises des Sciences Exactes et Naturelles, Ser.3 B 5:111-195, 1930. WOOLFOLK, C.A. Metabolism of N-methylpurines by a Pseudomonas putida strain isolated by enrichment on the caffeine as sole source of carbon and nitrogen. Journal of Bacteriology, 123:1088-1106, 1975. YAMAOKA-YANO, D.M. & MAZZAFERA, P. Degradation of caffeine by Pseudomonas and other bacteria isolated from soil. Microbial Ecology, 1996. Manuscrito (submetido para publicação). R. Bras. Fisiol. Veg., 8(1):67-74, 1996
Download