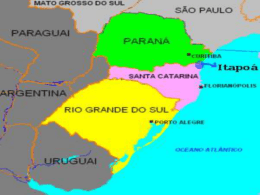
UNIVERSIDADE ESTADUAL DO OESTE DO PARANÁ CENTRO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE PROGRAMA DE PÓS-GRADUAÇÃO STRICTO SENSU EM CONSERVAÇÃO E MANEJO DE RECURSOS NATURAIS – NÍVEL MESTRADO LUCIANE LUVIZON CAMPAGNOLO COMPOSIÇÃO E ESTRUTURA DA ICTIOFAUNA EM UMA ÁREA AQUÍCOLA DO RESERVATÓRIO DE SALTO CAXIAS, RIO IGUAÇU/PR CASCAVEL-PR SETEMBRO/2012 LUCIANE LUVIZON CAMPAGNOLO COMPOSIÇÃO E ESTRUTURA DA ICTIOFAUNA EM UMA ÁREA AQUÍCOLA DO RESERVATÓRIO DE SALTO CAXIAS, RIO IGUAÇU/PR Dissertação apresentada ao Programa de Pósgraduação Stricto Sensu em Conservação e Manejo de Recursos Naturais – Nível Mestrado, do Centro de Ciências Biológicas e da Saúde, da Universidade estadual do Oeste do Paraná, como requisito parcial para a obtenção do título de Mestre em Conservação e Manejo de Recursos Naturais Área de Concentração: Conservação e Manejo de Recursos Naturais CASCAVEL-PR SETEMBRO/2012 Ficha Catalográfica Elemento obrigatório* (APÓS CORREÇÕES-CÓPIA FINAL) *Elaborado somente pela Biblioteca da UNIOESTE FOLHA DE APROVAÇÃO LUCIANE LUVIZON CAMPAGNOLO COMPOSIÇÃO E ESTRUTURA DA ICTIOFAUNA EM UMA ÁREA AQUÍCOLA DO RESERVATÓRIO DE SALTO CAXIAS, RIO IGUAÇU/PR Dissertação apresentada ao Programa de Pós-Graduação Stricto Sensu em Conservação e Manejo de Recursos Naturais - Nível de Mestrado, do Centro de Ciências Biológicas e da Saúde, da Universidade Estadual do Oeste do Paraná, como requisito parcial para a obtenção do título de Mestre em Conservação e Manejo de Recursos Naturais, pela comissão Examinadora composta pelos membros: Profa. Dra. Maristela Cavicchioli Makrakis Universidade Estadual do Oeste do Paraná (Presidente) ____________________________________ Profa. Dra. Rosilene Luciana Delariva Universidade Estadual do Oeste do Paraná ____________________________________ Profa. Dra. Valéria Flávia Batista da Silva Universidade Estadual do Mato Grosso do Sul Aprovada em 14 de setembro de 2012 Local de Defesa: Unioeste, Prédio de Salas de Aula, Sala 56, Cascavel-PR. Aos meus pais Ione Teresinha Presotto e Leodir João Luvizon, pelo exemplo de sabedoria, humanidade e amor! Dedico “A imensa satisfação intelectual, o imenso prazer de poder de fato entender por que as coisas são como são na natureza, nos abrem as portas para um eterno, inesgotável caso de amor com o mundo natural”. (Fernando Fernandez em sua obra O Poema Imperfeito: Crônicas de Biologia, Conservação da Natureza e seus Heróis). AGRADECIMENTOS Agradeço a Deus! Por ser meu rumo, minha fonte de fé e esperança! Ao meu esposo, Marlon Robson Campagnolo, pelo apoio, paciência, dedicação, cuidado e amor! Sem dúvidas, o companheiro da minha vida inteira! Ao meu amado irmão Jariel Guilherme Luvizon, por todos os dias de ajuda, pela paciência nos momentos de nervosismo, pelo carinho, por acreditar em mim e por sua serenidade, que fez meus dias serem menos difíceis! As minhas amigas do mestrado, Paula, Salete, Marinêz, Renata, Liege e Eliana, pelas brincadeiras, pelos almoços, por cederem um lugar em suas casas durante as disciplinas, por segurarem a “barra” durante minha recuperação do acidente, e principalmente, pela amizade desinteressada! A minha orientadora Dra. Maristela Cavicchioli Makrakis e Co-orientadora Dra. Elaine Antoniassi Luiz Kashiwaqui por todos os momentos de compreensão, de ensino, de conversas, pelos conselhos, lições e pelo carinho. Vocês acrescentaram muito à minha formação, profissional e pessoal! Ao programa Nacional de Pós-Doutorado (PNPD)/Coordenação de Aperfeiçoamento (CAPES) pelo apoio financeiro a pesquisa; Aos integrantes do grupo de pesquisa GETECH, muito obrigada por todos os ensinamentos e pela ajuda durante as coletas. Ao grupo de pesquisa GEMAQ, pela disponibilização de informações. Aos meus amigos de toda a vida, que de uma forma ou de outra, contribuíram para tornar possível essa conquista! E principalmente a toda a minha família, meus pais, meus irmãos e meu marido, pois sempre virão em primeiro lugar na minha vida, em quaisquer circunstâncias! SUMÁRIO CAPÍTULO 1: COMPOSIÇÃO E ESTRUTURA DA ICTIOFAUNA EM UMA ÁREA AQUÍCOLA DO RESERVATÓRIO DE SALTO CAXIAS, RIO IGUAÇU/PR.................8 Resumo .....................................................................................................................8 Abstract ....................................................................................................................9 Introdução............................................................................................................... 10 Material e Métodos ................................................................................................. 13 Resultados .............................................................................................................. 18 Discussão ................................................................................................................ 29 Agradecimentos ...................................................................................................... 33 Referências ............................................................................................................. 33 Anexos ................................................................................................................... 44 8 COMPOSIÇÃO E ESTRUTURA DA ICTIOFAUNA EM UMA ÁREA AQUÍCOLA DO RESERVATÓRIO DE SALTO CAXIAS, RIO IGUAÇU/PR Luciane Luvizon Campagnolo1, Maristela Cavicchioli Makrakis2, Elaine Antoniassi Luiz Kashiwaqui3 1 Universidade Estadual do Oeste do Paraná, Programa de Pós-Graduação em Conservação e Manejo de Recursos Naturais, Rua Universitária, 2019, Jardim Universitário, 85819110, Cascavel, Paraná, Brasil. [email protected] 2 Universidade Estadual do Oeste do Paraná - UNIOESTE, GETECH. Programa de Pós- Graduação em Recursos Pesqueiros e Engenharia de Pesca. Rua da Faculdade, 645. Jardim Santa Maria. 85903-000. Toledo, PR, Brazil. [email protected]. 3 Universidade Estadual de Mato Grosso do Sul - Grupo de Estudos em Ciências Ambientais e Educação (GEAMBE), BR 163 km 20,2; Mundo Novo, Mato Grosso do Sul. [email protected] Resumo Este trabalho avaliou os padrões espaciais na composição e estrutura da ictiofauna, bem como a relação desses padrões com as características físicas e químicas de dois pontos de uma área aquícola no reservatório de Salto Caxias, rio Iguaçu/PR, sendo um com influência direta do cultivo de peixes em tanques-rede e outro sem cultivo (controle). Para caracterizar a estrutura da ictiofauna, foram avaliados os atributos ecológicos como diversidade, equitabilidade, riqueza e dominância, assim como, avaliadas as condições abióticas dos locais de estudo. As coletas foram trimestrais, no período de julho de 2010 a 9 abril de 2011, utilizando-se redes de espera de diferentes malhagens, nos dois pontos do reservatório: na área aquícola (denominado tanques-rede), e no local com influência de um ambiente lótico, o rio Jacutinga (denominado Jacutinga). Durante o período de estudo, foram capturados 1060 peixes, pertencentes a quatro Ordens, 11 famílias e 31 espécies, com a maior abundância observada na área aquícola, correspondente a 629 peixes (59,3%). Os atributos analisados mostraram que o ponto tanques-rede apresentou maior diversidade, riqueza e dominância de espécies, enquanto que a equitabilidade foi maior no ponto Jacutinga, embora não houve diferenças significativas entre os dois ambientes, para estes atributos. Os fatores abióticos que mais influenciaram a posição das amostras evidenciadas pela Análise de Componentes Principais (ACP) foram a transparência, DBO, ortofosfato, turbidez, fósforo total, clorofila-α, pH, NO2 e condutividade elétrica. Os fatores abióticos analisados não apresentaram diferenças significativas entre os pontos de coleta. Nossos resultados sugerem que a atividade de aquicultura durante o período analisado não influenciou de forma significativa a abundância, os atributos ecológicos da assembleia de peixes e os fatores abióticos, da região aquícola, a ponto de interferir na estrutura da ictiofauna analisada. Abstract This study evaluated the spatial patterns in the composition and structure of fish fauna, as well as investigated the relationship between these patterns with the physical and chemical characteristics of two environments in an aquaculture area of the Salto Caxias reservoir, Iguaçu River/PR, one with direct influence of the fish cages and the other without influence (control). To characterize the structure of the ichthyofauna, we assessed the ecological attributes such as diversity, evenness, richness and dominance, as well as the 10 abiotic conditions of the study sites. Samples were collected quarterly from July 2010 to April 2011, using gill nets of different mesh sizes, in both sites of the reservoir: in the aquaculture area (named Cages), and in local with influence of a lotic environment, the Jacutinga River (named Jacutinga). We captured 1060 fish belonging to four orders, 11 families and 31 species, with the highest abundance observed in the aquaculture area, corresponding to 629 fishes (59.3%). The attributes analyzed showed that the Cages site showed greater diversity, richness and dominance of species, while evenness was higher in Jacutinga site, although no significant differences was observed between the two environments. The abiotic variables that influenced the position of the sample (PCA) were water transparency, DBO, orthophosphate, turbidity, total phosphorus, chlorophyll a, pH, electrical conductivity and NO2. The abiotic factors analyzed showed no significant differences between sampling sites, only differences among periods (trials). Our results suggest that the activity of aquaculture during the study period did not influence significantly the abundance, the ecological attributes of fish assembly and abiotic factors in the aquaculture area to interfere in the structure of the fishes examined. Keywords: Fish cages, water quality, ecological attributes, abundance. Introdução A bacia hidrográfica do rio Iguaçu possui uma área de aproximadamente 72.000 km2, da qual 79% pertencem ao Estado do Paraná, 19% à Santa Catarina e 2% à Argentina (Agostinho & Gomes 1997), percorrendo 1060 km, desde sua nascente na vertente ocidental da serra do Mar, proximidades de Curitiba, até a foz, no rio Paraná (Júlio Jr. et al., 1997). 11 O elevado desnível da bacia do rio Iguaçu no terceiro planalto se constitui em grande atrativo para o aproveitamento hidrelétrico, resultando na construção de barragens, formando grandes reservatórios de água. Cerca de cinco reservatórios hidrelétricos foram formados no leito principal do rio Iguaçu, que é um dos principais tributários do rio Paraná (Agostinho et al., 2007), sendo estes das Usinas: Foz do Areia, no município de Pinhão; Segredo, município de Mangueirinha; Salto Caxias, município de Capitão Leônidas Marques; Salto Santiago, município de Saudade do Iguaçu e Salto Osório, no município de Quedas do Iguaçu (COPEL, 2002). No que se refere à ictiofauna da bacia do rio Iguaçu, um levantamento realizado por Garavello et al. (1997), no reservatório de Segredo, totalizou 52 espécies, dentre essas um gênero e 14 espécies não descritas, provavelmente endêmicas, e outras três introduzidas de outras bacias, conforme definição dos autores. Baumgartner et. al. (2012) ampliaram esse número para 106 espécies de peixes para o baixo rio Iguaçu, em seu inventário e redescrição de espécies, incluindo espécies não indígenas. A bacia é caracterizada por apresentar um elevado grau de endemismo em razão do isolamento provocado pelo surgimento das Cataratas do Iguaçu (Garavello et al., 1997). Na década de 90, o endemismo ictiofaunístico da bacia foi estimado em 75% por Zawadzki et al. (1999), no entanto, mais recentemente, um estudo realizado por Baumgartner et al. (2012) diminuiu o percentual para 69,7%. Os reservatórios artificiais têm sido utilizados para múltiplas finalidades, dentre elas a produção de alimento por meio da piscicultura (Tundisi, 2005), tornando-os ambientes de grande importância em termos sociais e econômicos (Mallasen, et al., 2008). As estatísticas do Ministério da Pesca e Aquicultura revelaram que a produção de pescado no Brasil teve um crescimento de 25% nos últimos oito anos, sendo 15,7% somente em 2008 e 2009 (MPA, 2010). 12 Nesse sentido, entre as alternativas da piscicultura, destaca-se o cultivo de peixes em tanques-rede. Essa modalidade de cultivo pode incrementar consideravelmente a produção aquícola, criar condições para atrair novos investidores e tornar-se uma excelente alternativa de geração de emprego e renda, além de diminuir a pressão sobre os estoques pesqueiros naturais (Ayroza et al., 2005). Borghetti & Silva (2008) estimam custos operacionais entre 30 e 40% daquele para produzir uma tonelada de peixes com sistemas tradicionais. As principais vantagens do sistema de produção de peixes em tanques-rede, segundo Furlaneto et al. (2006), são: menor variação dos parâmetros físicos e químicos da água durante a criação, maior facilidade de retirada dos peixes para venda, menor investimento inicial, facilidade de movimentação e realocação dos peixes, intensificação da produção, facilidade de observação dos peixes, redução do manuseio dos peixes e, diminuição dos custos com tratamentos de doenças. Todavia, a disponibilidade de alimento artificial aliado à presença física da estrutura dos tanques-rede, além de atrair e agregar as assembleias de peixes silvestres nas proximidades dos tanques-rede atrai piscívoros e outros predadores como mamíferos, aves e répteis, que podem alterar a estrutura trófica local, danificar os tanques, promovendo escapes e permitir que ciclos de parasitas sejam fechados, com aumento das chances de troca de patógenos entre peixes silvestres e cultivados (Beveridge, 2004; Agostinho et al., 2007). Nesse sentido, Zaniboni-Filho et al. (2005), destacam a origem dos impactos decorrentes do cultivo intensivo em tanques-rede como sendo a matéria orgânica depositada no ambiente, tanto como resultado do metabolismo dos peixes como pelo excedente de ração não ingerida. Essa matéria causa mudanças nas características físicas, químicas e biológicas do local onde estão instalados os tanques-rede, sendo que a grande dificuldade no cultivo em tanques-rede, considerando a estrutura utilizada atualmente, é a 13 impossibilidade do tratamento do efluente produzido (Zaniboni-Filho et al., 2005; Araripe et al., 2006). Alguns estudos revelaram os diversos impactos causados por essa modalidade de cultivo (Tundisi & Henry 1986; Damato 2001; Guo & Li, 2003; Agostinho et al., 2007; Demetrio, 2010 e Loureiro et al., 2012). No entanto, ainda são escassos, principalmente no que se refere ao impacto na ictiofauna, justificando a necessidade da realização de estudos dessa natureza. Segundo Tundisi et al. (2008), esse conhecimento deve ser incluído entre os “desafios” a serem superados para possibilitar a racionalização do uso dos reservatórios. Sendo assim, considerando a presença dos tanques-rede e as premissas de que uma área aquícola promove o aporte de nutrientes (Araripe et al., 2006), a atração de indivíduos (Zanatta, 2011) e mudanças nas condições físicas e químicas da água (Bueno et al., 2008), os objetivos deste estudo foram (I) avaliar os padrões espaciais na composição, estrutura e distribuição da ictiofauna residente e (II) relacionar esses padrões com as características físicas e químicas de dois pontos no reservatório de Salto Caxias, rio Iguaçu/PR, sendo um na área aquícola, com influência direta do cultivo de peixes em tanques-rede e outro sem cultivo. Desta forma, baseado na hipótese de que ambientes modificados alteram a estrutura de suas comunidades. Esse estudo busca testar a seguinte predição: de que um ambiente aquático sob efeito do cultivo em tanques-redes possui a estrutura e distribuição das assembleias de peixes diferenciadas, em relação a um ambiente sem essa influência. Material e Métodos Área de estudo O reservatório de Salto Caxias localiza-se na região Sudoeste do estado do Paraná (25º 32’S e 53º 30’W; 25º 35’S e 53º 06’W), à aproximadamente 180 km a montante das 14 Cataratas do Iguaçu e a 100 km a jusante da barragem do reservatório de Salto Osório (Novakowski et al., 2007), sendo o último de uma cascata de cinco reservatórios instalados a montante do médio rio Iguaçu (Delariva, 2002). O reservatório foi fechado em 1998, inundando uma área de 124 km2, operado no sistema fio d’água (ONS, 2012) e com tempo de residência de 31 dias (IAP, 2009). O Instituto Ambiental do Paraná caracterizou o reservatório de Salto Caxias como pouco degradado (classe II) e destaca ainda que o padrão de estratificação é de um lago monomítico quente, com isotermia no inverno e depleção acentuada de oxigênio nos meses mais quentes, com hipoxia na camada próxima ao fundo (IAP, 2009). A área aquícola na qual foi desenvolvida a pesquisa é o resultado de um Convênio firmado em 2005, entre Conselho Pró Desenvolvimento dos municípios Lindeiros ao Lago de Salto Caxias, Universidade Estadual do Oeste do Paraná e Secretaria do Estado da Ciência, Tecnologia e Ensino Superior – SETI, com o objetivo de desenvolver economicamente a região. Cabe ressaltar que este é o único Centro de Difusão de Tecnologia (CDT) existente no rio Iguaçu. Contém cerca de vinte tanques-rede experimentais instalados em 2007, com área de 4,05 m3 cada, com matrizes de Rhamdia voulezi e Steindachneridion melanodermatum (Feiden et al., 2010; Bittencourt et al., 2009), entre outras espécies endêmicas da bacia. Os pontos de coleta estão localizados em duas áreas de remanso (tributários alagados do reservatório) (Fig. 1), assim identificados: - Ponto 1 (denominado Jacutinga – sem cultivo): braço do reservatório que possui influência de um afluente, o rio Jacutinga. Considerado como área a montante dos tanquesrede (25º 26’30,42” S e 52º 26’ 28,99” W); - Ponto 2 (denominado Tanques-rede) : braço no qual está situada a área aquícola, com os tanques-rede desde 2007 (25º 26’ 55,00” S e 53º 26’ 41,81”W). 15 Durante o período das coletas, entre julho de 2010 e abril de 2011, as espécies endêmicas da bacia como o jundiá (Rhamdia branneri, Rhamdia voulezi), mandi pintado (Pimelodus britskii), surubim do Iguaçu (Steindachneridion melanodermatum) e lambaris (Astyanax spp.), estavam sendo cultivadas nos tanques-rede. A biomassa de matrizes cultivada correspondia a 507,7 kg, para as respectivas espécies, alimentadas diariamente com 13 a 15 kg de ração comercial contendo 28% de proteína bruta e também algumas rações experimentais. Fig. 1 Localização dos pontos de coleta no reservatório de Salto Caxias, rio Iguaçu/PR (P1= Jacutinga e P2= Tanques-rede). Coletas de dados As amostragens da ictiofauna foram trimestrais entre julho/2010 e abril/2011, compreendendo as estações do ano, totalizando quatro coletas para cada ponto. Para isso, 16 foram utilizadas redes de espera expostas por 24 horas, sendo 12 do tipo simples (malhas entre 2,5 e 12 cm) e três do tipo tresmalhos (“feiticeira”, malhas de 6,0; 7,0 e 8,0 cm). As despescas aconteceram a cada oito horas (manhã, tarde, noite). Os indivíduos capturados foram levados ao Centro de Difusão e Desenvolvimento Tecnológico para o rio Iguaçu (CDT-Iguaçu), localizado próximo ao local de coleta, onde foram anestesiados com formaldeído 10% e imolados para posterior identificação, de acordo com Garavello et al. (1997), Reis et al.(2003) e Baumgartner et al. (2012). Exemplares testemunhos estão depositados no Laboratório de Ictiologia do Curso de Engenharia de Pesca-Unioeste-Toledo/PR. Os fatores abióticos foram obtidos concomitante às coletas de peixes, sendo a temperatura (oC), oxigênio dissolvido (mg/L-1 e %), pH, turbidez (NTU) e condutividade elétrica (µS.cm-1) analisados com o multiparâmetro digital YSI, modelo Profissional Plus, e a transparência da água por meio do Disco de Secchi. As análises de clorofila (µg/L-1) e ortofosfato (µg/L-1), foram realizadas como sugerido por Tavares & Moreno (1994) e Mackereth et al. (1978), respectivamente. A determinação da amônia (µg/L-1) foi feita com base na metodologia proposta por Koroleff et al. (1976) e o nitrito (µg/L-1) por Strickland et al. (1972). A Demanda bioquímica de Oxigênio DBO (mg/L-1) foi determinada pelo “Standard Methods for the examination of water and wastewater”, segundo Apha (1998). Os dados pluviométricos do período das coletas foram obtidos junto ao Instituto Tecnológico SIMEPAR, da estação meteorológica Salto Caxias, localizada no município de Capitão Leônidas Marques-PR (latitude -25.5166 e longitude -53.4833). Os fatores abióticos foram utilizados para caracterizar as condições abióticas do local de estudo e foram comparados com a Resolução CONAMA 357/05, a qual estabelece os padrões de lançamento de efluentes para os corpos d’água Classe II. 17 Análise dos dados A comparação da estrutura das assembleias de peixes entre os pontos amostrados foi realizada utilizando-se como atributos ecológicos: composição, abundância, diversidade, riqueza, dominância, equitabilidade e constância. A abundância foi determinada em número de indivíduos e biomassa e expressa em captura por unidade de esforço (CPUE), dada em número de indivíduos por 1000 m2 de rede em 24 horas. A diversidade da assembleia foi calculada pelo índice de Shannon – Wiener (H’) (Magurran, 1988), calculado pela fórmula H’ = - ∑ (n/N) log (n/N), onde n = número de indivíduos de cada espécie e N = número total de indivíduos. A riqueza foi definida como o número de espécies encontradas nas comunidades coletadas. A dominância foi calculada pelo índice de Simpson (Magurran, 1988), baseando-se na fórmula (y1 + y2)/Y, onde y1= abundância da 1ª espécie mais abundante, y2=abundância da 2ª espécie mais abundante e Y= abundância total de todas as espécies. A equitabilidade da comunidade de peixes foi conhecida pelo índice de Pielou (1984). A base para o cálculo do índice de equitabilidade se deu pela equação J = H’/ln S, onde: S = número de espécies por coleta e H’= Índice de Shannon. A partir das coletas, calculou-se também a constância de ocorrência de Dajoz (1983), estimada pela fórmula C= (ni / N) x 100, em que: C= constância de ocorrência, n= número de coletas onde a espécie i ocorreu e N = número total de coletas. As espécies foram classificadas em: constantes (C>50%), acessórias (50≥C≥25%) e acidentais (C<25%). Fezse também a estimativa de proporção de captura (PC) das espécies, calculada pelo número total de indivíduos de uma dada espécie em relação ao número total de peixes capturados, expressa em percentagem, para cada local de coleta. 18 Foram utilizadas técnicas multivariadas visando estabelecer a relação entre a composição das assembleias, atributos ecológicos e fatores abióticos com os locais de coleta. Para sumarizar a matriz de dados de composição da assembleia, abundância e locais de coleta foi utilizada a análise de correspondência com remoção do efeito do arco (DCA; Gauch Jr., 1986; Jongman et al., 1995). Essa análise foi realizada com a CPUE das espécies coletadas nos diferentes meses. No que se refere às variáveis abióticas, estas foram resumidas pela análise de componentes principais (ACP; Gauch Jr. 1986), e os eixos retidos para interpretação foram aqueles que apresentaram autovalores maiores que os obtidos pelo modelo aleatório de “Broken-stick” (Jackson 1993). Para verificar as possíveis correlações entre as assembleias de peixes (abundância numérica - CPUE) com os fatores abióticos, aplicou-se uma análise de correspondência canônica (ACC). Cabe ressaltar que os dados de CPUE foram previamente transformados em raiz quadrada. Para verificar possíveis diferenças nas médias dos atributos e dos escores dos eixos da ACP, DCA e ACC nos diferentes locais de coletas (P1 e P2) foram empregadas análises de variância unifatoriais (ANOVA unifatorial), respeitando-se os pressupostos de normalidade e homocedasticidade dos dados (Zar, 1999), no programa Statistica 7.0 (Statsoft, 2005). A ACC e o cálculo dos atributos foram realizados no programa PAST (Hammer et al., 2003). Para a realização da ACP e DCA utilizou-se o programa PC-ORD (McCune & Mefford, 1997). Resultados O levantamento ictiofaunístico, independente do ponto amostrado, resultou na captura de 1060 indivíduos, pertencentes a quatro ordens (Characiformes, Siluriformes, Perciformes e 19 Gymnotiformes), 11 famílias e 31 espécies (Tabela 1). Do total de espécies, 18 (58,06%) são endêmicas da bacia do rio Iguaçu, 10 (32,25%) são nativas e três (9,6%) são introduzidas de outras bacias. No que se refere à distribuição dessas entre os pontos, 20 foram comuns aos dois pontos amostrados, quatro foram exclusivas do ponto Jacutinga: Bryconamericus sp. C, Crenicichla lepidota, Crenicichla sp. 2 e Gymnogeophagus setequedas e sete foram exclusivas do ponto Tanques-rede: Astyanax gymnogenys, Clarias gariepinus, Hypostomus myersi, Psalidodon gymnodontus, Rhamdia branneri, Steindachneridion melanodermatum e Tilapia rendalli. O resultado da constância de ocorrência das espécies mostrou que, para o ponto Jacutinga, o número de espécies constantes e acessórias correspondeu a 12 espécies (50% cada). Para as constantes, 9 apareceram em todas as coletas e 3 espécies em três coletas, e as acessórias com 9 de ocorrência em apenas uma coleta. No ponto Tanques-rede, o número de espécies constantes correspondeu a 13 espécies (48,1%), sendo 10 em todas as coletas e 3 em três coletas, e as acessórias representaram 14 espécies, das quais 3 em três coletas e 11 em uma coleta (Tabela 1). Tabela 1 Lista de espécies capturadas no reservatório de Salto Caxias – rio Iguaçu – Paraná, de julho de 2010 a abril de 2011. ♦ Espécie endêmica ◊ Espécie nativa ■ Espécie Introduzida. Família Parodontidae Curimatidae Erythrinidae Characidae Espécie Characiformes Apareiodon vittatus Cyphocharax cf. santacatarinae Hoplias aff. malabaricus Astyanax altiparanae Astyanax gymnogenys Astyanax gymnodontus Astyanax bifasciatus Astyanax minor Astyanax dissimilis Oligosarcus longirostris Sigla Distribuição Classificação J Avit Csan Hama Aalt Agyg Agyd Astb Astm Astd Olon ♦ ◊ ◊ ◊ ♦ ♦ ♦ ♦ ♦ ♦ Co Co Co Co Co Co Co Ace Co T Co Ace Co Co Ace Co Co Co Co Co 20 Bryconamericus ikaa Bryconamericus sp. C Psalidodon gymnodontus Bika BspC Pgym ♦ ♦ ♦ Ace Ace - Co Ace Cgar Cpal Hder Hmye Hysp Pbri Port Smel Rbra ■ ◊ ◊ ◊ ◊ ♦ ♦ ♦ ♦ Ace Co Ace Co Ace - Ace Gsyl ■ Ace Ace Gbra Tren Cigu Clep Crsp Csp1 Csp2 Gset ◊ ■ ♦ ◊ ♦ ♦ ♦ ◊ Ace Co Ace Co Ace Ace Ace Ace Ace Co Co Ace - Siluriformes Heptapteridae Clarias gariepinus Corydoras aff. paleatus Hypostomus derbyi Hypostomus myersi Hypostomus sp. Pimelodus britskii Pimelodus ortmanni Steindachneridion melanodermatum Rhamdia branneri Gymnotidae Gymnotus sylvius Cichlidae Geophagus brasiliensis Tilapia rendalli Crenicichla iguassuensis Crenicichla lepidota Crenicichla sp. Crenicichla sp. 1 Crenicichla sp. 2 Gymnogeophagus setequedas Clariidae Callichthyidae Loricariidae Pimelodidae Ace Co Ace Ace Co Ace Ace Ace Gymnotiformes Perciformes Abundância Do total de indivíduos coletados, 431 (40,7%) foram capturados no ponto Jacutinga e 629 (59,3%) no ponto Tanques-rede. A abundância numérica das capturas totais expressa em CPUE relevou que as espécies mais abundantes foram A. bifasciatus, A. altiparanae, A. vittatus e A. minor. No que se refere à abundância, considerando os locais de coleta, a maior abundância foi observada para as espécies A. bifasciatus e A. altiparanae tanto no ponto Jacutinga como no ponto Tanques-rede. No entanto, a terceira espécie mais abundante no ponto Jacutinga foi A. vittatus, enquanto que no ponto Tanques-rede foi O. longirostris. No que se refere à CPUE expressa em biomassa, os maiores índices foram observados no ponto Jacutinga para P. britskii, H. aff. malabaricus, O. longirostris, e no ponto Tanques- 21 rede para H. derbyi, H. aff. malabaricus e O. longirostris. No entanto, destaca-se que o ponto Tanques-rede obteve uma biomassa (kg) em CPUE mais elevada que o ponto Jacutinga (Figura 2). Jacutinga Tanques-rede Abif Aalt Olon Crsp Amin Agyd Cigu Cpal Adis Pbri Avit Bikaa Hama Hder Gbra Csp1 Agyg Csan Port Pgyd Cgar Gsyl Hmye Hysp Rbra Smel Tren Abif Aalt Avit Amin Olon Crsp Pbri Csan Cpal Cigu Agyd Adis Hama Hder Gbra Bika Csp1 BspC Clepi Csp2 Gset Gsyl Hysp Port 400 300 200 100 0 CPUE (número) 5 10 kg CPUE (biomassa) 15 20 600 450 300 150 0 5 10 15 20 kg CPUE (número) CPUE (biomassa) Fig. 2. Captura por unidade de esforço (CPUE) expressa em número de indivíduos e biomassa (kg) para as espécies capturadas nos pontos de coleta na área aquícola do reservatório de Salto Caxias, rio Iguaçu-PR. As siglas das espécies estão relacionadas na Tabela 1. Estrutura da ictiofauna Por meio da análise dos atributos ecológicos, identificou-se que houve pouca variação dos mesmos entre locais analisados (Tabela 2). A análise de variância unifatorial (ANOVA) realizada mostrou que as diferenças não foram significativas para nenhum dos atributos analisados (Figuras 3 e 4). 22 Tabela 2 Riqueza de espécies de peixes (S), equitabilidade (E), índice de diversidade de Shannon-Wiener (H`) e dominância (D`) obtidos para os pontos de coleta e períodos amostrados na área aquícola do reservatório de Salto Caxias, rio Iguaçu/PR. Local Mês Jacutinga Jacutinga Jacutinga Jacutinga Tanques-rede Tanques-rede Tanques-rede Tanques-rede Julho Outubro Janeiro Abril Julho Outubro Janeiro Abril S 16 20 14 10 16 21 15 14 E 0,768 0,793 0,855 0,869 0,79 0,79 0,836 0,837 H` 2,131 2,375 2,257 2,002 2,191 2,404 2,265 2,21 D` 0,8342 0,8599 0,8718 0,8399 0,8326 0,8752 0,8526 0,8555 0,85 19 Média 18 Média 0,84 Equitabilidade (E) Riqueza (S) 17 16 15 14 0,83 0,82 0,81 0,80 13 S: 12 Média±EP Média±EP Jacutinga F(1;6) = 0,3333; p = 0,5847 Tanques rede E: F(1;6) = 0,0833; p = 0,7825 0,79 Jacutinga Tanques rede Fig. 3 Valores médios (± erro padrão) dos atributos ecológicos riqueza e equitabilidade, das espécies de peixes coletadas nos pontos Jacutinga e tanques-rede na área aquícola de Salto Caxias, rio Iguaçu/PR. 23 0,864 0,862 Mean Mean±EP 0,860 0,858 Média Média±EP Dominância (D') Índice de Diversidade (H') 2,32 2,28 2,24 2,20 0,856 0,854 0,852 0,850 0,848 0,846 2,16 0,844 2,12 0,842 H: 2,08 D`: F(1;6) = 0,0419; p = 0,8447 F(1;6) = 0,6624; p = 0,4468 0,840 Jacutinga Tanques rede Jacutinga Tanques-rede Fig. 4 Valores médios (± erro padrão) dos atributos ecológicos diversidade e dominância das espécies de peixes coletadas nos pontos Jacutinga e tanques-rede na área aquícola de Salto Caxias, rio Iguaçu/PR. Os padrões espaciais de variações na composição e abundância foram analisados pela DCA, sendo que o eixo 1 apresentou autovalor de 0,24, enquanto o eixo 2 apresentou autovalor de 0,047, desta forma, somente os escores do eixo 1 foram retidos para interpretação (Matthews, 1998). Mesmo assim, os eixos 1 e 2 foram separados para a apresentação gráfica. As espécies mais correlacionadas com o primeiro eixo da DCA (Figura 5a), ou seja, as que mais influenciaram na ordenação, foram, positivamente, C. gariepinus, H. myersi, R. branneri e S. melanodermatum; e negativamente, Bryconamericus sp. C, T. rendalli, P. gymnodontus e C. lepidota. 24 120 80 abr11 out10 jan11 60 DCA2 Jacutinga Tanques rede jan11 100 jul10 abr11 out10 40 20 jul10 0 -20 -20 b 0 20 40 60 80 100 120 140 160 DCA1 Fig. 5 Ordenação resultante da Análise de Correspondência Destendênciada (DCA) com remoção do efeito do arco (a e b), com os dados da CPUE (ind/1000m2 de rede por 24 h) das espécies de peixes da área aquícola do reservatório de Salto Caxias, rio Iguaçu/PR. Com relação à ordenação dos meses e pontos de coleta, verificou-se que, outubro para os dois pontos de coleta permaneceu no espaço positivo e separado dos demais meses, assim como, julho para o ponto tanques-rede e janeiro para o ponto Jacutinga (Figura 5b). A aplicação da ANOVA unifatorial não revelou diferenças significativas nas médias desses escores entre os locais de coleta. 25 Fatores abióticos Com base na análise da Tabela 3, verifica-se que a temperatura da água (oC) obteve os mesmos valores para os dois pontos de coleta; a temperatura do ar (oC) foi mais elevada para o ponto tanques-rede nos meses de julho e janeiro; para a variável oxigênio dissolvido (mg/L e %) os menores valores foram obtidos em abril para os dois pontos. Já a turbidez se mostrou bastante elevada no mês de outubro para ambos os pontos de coleta, no entanto, ainda maior para o ponto Jacutinga; o mês com menor transparência da água foi o de outubro, no ponto tanques-rede, e a maior transparência foi identificada no mês de janeiro para ambos os pontos de coleta. O pH não apresentou grande variação entre os períodos analisados; o parâmetro condutividade elétrica (µS/Cm) foi maior no mês de janeiro e obteve os menores valores no mês de abril tanto para o ponto Jacutinga como no ponto tanques-rede. Com relação à DBO, essa se mostrou maior nos dois pontos no mês de outubro, e no mês de abril essa variável apresentou os menores valores no ponto tanques-rede. O fósforo total teve o menor valor no ponto Jacutinga no mês de outubro, já no ponto tanques-rede no mês de abril se obteve as maiores concentrações. Para o ortofosfato, registraram-se valores bem acima dos outros meses no mês de outubro, para os dois pontos de coleta. A concentração de nitrito (NO2) apresentou alta variação nos dois pontos no mês de janeiro, mantendo-se estável nos outros meses. Para a amônia (NH3), o ponto Jacutinga apresentou valores superiores ao ponto tanques-rede em todos os meses, porém, ambos tiveram valores mais elevados no mês de outubro. Já a clorofila-ɑ apresentou um valor bem elevado no mês de abril no ponto tanques-rede. 26 Tabela 3 Resultado dos fatores abióticos obtidos nos pontos de coleta na área aquícola do reservatório de Salto Caxias, rio Iguaçu/PR, de julho de 2010 a abril de 2011. Legenda: Jacu = Jacutinga; Tan = Tanques; jul = Julho; out = Outubro; jan = Janeiro e abr = Abril. Local T. ar T. OD Cond. T Secchi DBO PT ORTO NO2 NH3 Clor. a pH OD% o C águaoC (mg/L) Elet. (NTU) (m) mg/L µg/L µg/L µg/L µg/L µg/L Jacujul10 11,7 18,5 9,0 7,0 94,2 56,3 3,04 1,85 20,9 23,7 5,41 0,2 21,3 4,8 Jacuout10 18,3 21,6 8,2 7,2 95,6 42,8 14,1 1,7 27,0 13,1 23,7 1,1 70,2 2,4 Jacujan11 23,5 29,3 7,5 8,0 99,7 67,8 1,7 2,3 8,3 18,4 4,6 5,7 46,0 1,7 Jacuabr11 20,6 25 5,3 7,2 63,7 30,8 2,8 2,2 7,9 21,5 0,5 0,5 33,1 1,7 Tanqjul10 12,7 18,5 8,8 7,1 94,2 47,6 3,6 1,7 4,3 19,3 3,4 0,0 11,1 2,5 Tanqout10 18,3 21,6 8,4 7,2 95,7 40,0 10,2 1,1 22,0 19,3 24,6 0,4 38,0 1,2 Tanqjan11 24,2 28,9 7,5 8,0 95,5 66,7 2,5 2,4 9,4 19,5 4,2 6,4 23,5 3,9 Tanqabr11 20,6 24,9 5,3 7,1 65,0 31,5 2,9 2,2 4,8 33,7 0,9 0,67 14,0 21,9 Com a realização da análise de componentes principais (ACP), que sumarizou os dados abióticos, somente os CP1 e CP2 apresentaram autovalores (5,76 e 4,53) maiores que aqueles obtidos pelo modelo “broken stick” (3,25 e 2,25) e, portanto, foram retidos para interpretação. Estes dois eixos explicaram juntos, 73,5% da variabilidade no gradiente apresentado para os dados abióticos (41,1% e 32,4%, para o eixo um e dois, respectivamente). As variáveis que mais influenciaram a posição das amostras na ordenação foram para o eixo um, positivamente, a transparência e, negativamente, a DBO, o ortofosfato e a turbidez. Já para o eixo dois, fósforo total e clorofila influenciaram positivamente, enquanto que o pH, NO2 e condutividade elétrica negativamente (Figura 6). 27 PC1 PC2 0.4 0.2 0 -0.2 -0.4 -0.6 i a cch gu Se T. a PT T. ar Clor. NO2 pH d. El NH3 D% g/L TU) BO RTO D O O (m (N n T Co OD Fig. 6 Autovetores (correlações) das variáveis utilizadas na análise de componentes principais (ACP) dos eixos retidos para interpretação. Com a ordenação dos pontos e meses, em relação às variáveis abióticas podemos afirmar que não houve diferenças significativas nas condições abióticas entre o ponto Jacutinga (Ponto 1) e tanques-rede (Ponto 2), (Eixo 1 F= 8,82; P = 0,59 e Eixo 2 F = 4,84; P = 0,11) nos dois eixos retidos da ACP. Os resultados demonstram que o primeiro eixo ordenou os dados de acordo com gradiente de radiação subaquática, disponibilidade de oxigênio no meio e nutrientes, enquanto o segundo ordenou as variáveis de acordo com gradiente de trofia, disponibilidade de carbono inorgânico e variabilidade de íons H+ e OH-. Para a variável turbidez, essa separou outubro dos demais meses, a qual se mostrou bastante elevada naquela coleta. Isso pode ser explicado pela elevada precipitação que ocorreu naquele período (Figura 7), ocasionando o carreamento de partículas sólidas suspensas da área de entorno para o reservatório, em consequência, aumentando sua turbidez. 28 Precipitação (mm) Precipitação Estação Meteorológica Salto Caxias/PR 16 14 12 10 8 6 4 2 0 jul/10 out/10 jan/11 abr/11 Fig. 7 Precipitação (mm) registrada nos períodos de coleta na estação meteorológica Salto Caxias, rio Iguaçu/PR. Fonte: Dados do Instituto Tecnológico SIMEPAR Quanto ao atendimento à Resolução CONAMA 357/05, todos os parâmetros atenderam os padrões de lançamento de efluentes nos corpos d’água para a Classe II, com exceção do parâmetro DBO (mg/L), que no ponto Jacutinga em todas as coletas se apresentou superior aos limites tolerados e no ponto tanques-rede nas coleta do mês de outubro. A análise de correspondência canônica (ACC), na Figura 7, apresenta a relação das variáveis abióticas com a abundância dos peixes amostrados. Na análise, as duas primeiras variáveis canônicas explicaram 53,19% da variabilidade dos dados, sendo 34,66% e 18,53% para o 1º e 2º eixo, respectivamente. Na primeira variável, Bryconamericus sp. C, C. lepidota e C. santacatarinae se associaram positivamente com a condutividade elétrica, nitrito e transparência da água e negativamente, A. gymnogenys, A. vittatus, A. minor, H. derbyi, O. longirostris, C. iguassuensis e A. bifasciatus, se associaram aos parâmetros clorofila-ɑ, fósforo total e temperatura da água e do ar. Para a segunda variável, positivamente, as espécies Crenicichla sp. 2, Hypostomus sp, G. setequedas, G. brasiliensis, G. sylvius e Crenicichla sp. 1, se associaram à amônia, DBO, OD (mg/L e %), ortofosfato e à turbidez. 29 Fig. 8 Triplot gerado com os resultados da Análise Correspondência Canônica (ACC) entre a abundância numérica (raiz quadrada da CPUE) e as variáveis abióticas nos pontos de coleta na área aquícola de Salto Caxias, rio Iguaçu/PR. Discussão Com base na análise da composição específica e riqueza da ictiofauna coletada na área de estudo, o menor número de espécies amostradas, comparada com os outros levantamentos realizados na bacia, (Garavello et al., 1997; Baumgartner et al., 2006; Novakowski et al., 2007), pode ser explicado pelas características de distribuição da ictiofauna e também pela menor área de amostragem, considerando a bacia do rio Iguaçu. Garavello et al. (1997), ressaltam que a distribuição longitudinal da ictiofauna do rio Iguaçu acima das Cataratas, ao longo de seu curso, não é uniforme. Algumas espécies são encontradas em regiões de maior altitude e outras em regiões mais baixas. A substituição de espécies e a variação no grau de dominância entre elas podem ser notadas ao longo da bacia do rio Iguaçu. 30 No que se refere à abundância numérica, A. bifasciatus foi a espécie mais capturada nas coletas realizadas. Essa espécie foi a mais abundante em um estudo realizado em 13 reservatórios do rio Iguaçu (Luiz, et al., 2005) e é uma das espécies mais abundantes no baixo rio Iguaçu, segundo evidenciado por Bailly et al. (2003). A maior intensidade reprodutiva dessa espécie ocorre na primavera e início do verão (Suzuki, 1999), sendo que as maiores capturas foram identificadas para o ponto Jacutinga em outubro (primavera) e no ponto tanques-rede em abril (outono). A segunda espécie com maior abundância nas capturas realizadas, A. altiparanae, pode ser atribuída à grande flexibilidade de hábitos alimentares e capacidade de reprodução em diversos habitats (Bennemann et al., 1995). Conforme relatado por Hoffmann et al. (2005) em seu estudo no reservatório de Capivara, rio Paranapanema, na Bacia do Alto rio Paraná, estes fatores devem ter permitido a expressiva abundância dessa espécie. A captura expressiva da espécie também foi evidenciada por Bailly et al. (2003) no reservatório de Salto Caxias. Entre os characiformes há um predomínio de espécies de pequeno porte e/ou capazes de concluir seu ciclo de vida em ambientes lênticos (Hoffmann et al., 2005), o que justifica a elevada abundância de espécies dessa ordem. O elevado número de espécies do gênero Astyanax nesse estudo, estando entre as mais abundantes, pode ser explicado pelo fato de apresentarem ovócitos pequenos, elevada fecundidade relativa, rápido desenvolvimento, fertilização externa e ausência de cuidado parental, fatos que possibilitam a essas espécies um amplo predomínio sobre as demais (Agostinho et al. 1999). Muitos estudos sugerem que na maioria das comunidades animais há poucas espécies abundantes e muitas espécies representadas por poucos indivíduos (Matthews, 1998). Esta afirmação foi confirmada, visto que foram 14 espécies (45,16%) com menos de quatro indivíduos capturados em todas as coletas. 31 A participação das espécies H. aff. malabaricus, P. britskii e H. derbyi se deu em função do maior porte dos indivíduos. Esse aspecto também foi verificado por Oliveira et al. (2008), na caracterização da assembleia de peixes no reservatório do Iraí, região do alto rio Iguaçu/PR. A despeito da captura de um indivíduo da espécie S. melanodermatum, salienta-se que a mesma ocorreu no ponto tanques-rede, onde ocorre o cultivo de matrizes dessa espécie, e muito possivelmente, este indivíduo tenha origem do escape de algum dos tanques-rede. A medida de constância de uma dada espécie em um ambiente indica o estabelecimento ou a temporariedade na utilização daquele biótopo pela espécie (Delariva et al., 2009). Nesse sentido, os resultados mostraram que as espécies constantes foram também as mais abundantes nos dois locais de coleta. Obtiveram resultados similares os estudos realizados por Dufech & Fialho (2009) no Parque Estadual de Itapuã, Sul do Brasil, e Lucena et al. (1994) na praia de Itapuã, no lago Guaíba, RS. Analisando os dados de abundância e riqueza, Zanatta (2011) em um estudo no reservatório de Chavantes (médio rio Paranapanema SP/PR) em um sistema particular de tanques-rede e uma área controle observou que essas foram maiores na área controle, porém, corroboram com os resultados obtidos neste estudo o que diz respeito à dominância, a qual também foi maior na área dos tanques-rede. No que se refere à abundância na área dos tanques-rede, diversos são os estudos que mostram os sistemas de piscicultura em tanques-rede como mecanismos de agregação de peixes (Valle et al., 2007; Dempster et al., 2002). Esses estudos demonstram grandes diferenças na abundância e biomassa de peixes quando se comparam trechos controle e regiões com áreas de cultivo em tanques-rede. Os autores atribuem às diferenças ao fato de que: i) essas estruturas podem servir de abrigo a peixes de pequeno porte que assim atraem os predadores de maior porte, ii) que o aporte de nutrientes derivado da sobra da ração e do 32 metabolismo dos peixes confinados servem de alimento para os peixes atraídos ou incrementa a produção primária, via ciclagem de nutrientes, e ainda, iii) ao fato de que a ocupação de áreas para a produção estaria reduzindo a pressão pesqueira nessas regiões (Zaniboni et al., 2005). Entretanto, este estudo apresentou resultados distintos dos mencionados acima, pois quando comparado a abundância numérica (CPUE) por meio Anova, essa não apresentou diferença significativa entre os pontos analisados. Apesar dos fatores abióticos analisados não apresentarem diferenças significativas entre os pontos analisados, a DBO não atendeu aos padrões estabelecidos pela Resolução CONAMA 357/05. Cabe ressaltar que no ponto Jacutinga o mês com maior concentração de DBO foi o mês de outubro, mês de maior precipitação registrada durante as coletas. Com base nas outras variáveis que também sofreram alterações nesse mês de coleta, indicam que as alterações físicas e químicas da água não são somente oriundas da área do cultivo em tanques-rede, visto que os maiores índices foram observados em períodos de entrada de material alóctone no reservatório. A região do entorno do reservatório de Salto Caxias possui uma extensa área de pastagens (54,4%) e agricultura (27,8%) (COPEL, 2002), assim como estradas não adequadas, e está entre os reservatórios da Copel que precisam de maior quantidade de vegetação a ser recomposta (67%) (Andrade et al., 2005), o que deixa suas margens vulneráveis à degradação. Outro fato que demonstra isso são os índices de DBO obtidos no ponto Jacutinga (controle), maiores que os do ponto tanques-rede, reforçando à entrada de material orgânico das áreas a montante que podem ter sido carreadas pelo próprio rio Jacutinga. Cabe destacar que, o efeito do cultivo em tanques-rede sobre o ambiente depende, primariamente, da produção anual de peixes (Zaniboni et al., 2005) e que o cultivo instalado na área aquícola de Salto Caxias correspondeu, no período das coletas, à uma 33 biomassa de 507,7 kg, para os quais não foi verificado diferença significativa na composição e estrutura da comunidade entre os dois ambientes, com a biomassa (kg) e número de tanques-rede instalados no local. No que concerne à análise dos atributos ecológicos, a predição de que áreas com a presença de cultivo em tanques-rede possuem a estrutura e distribuição das assembleias de peixes diferenciadas, em relação a um ambiente sem essa influência, não foi comprovada. Com esses resultados, percebeu-se que, o ponto Tanques-rede apresentou uma maior abundância (atração de indivíduos), diversidade, riqueza e dominância de espécies, e essas alterações, embora não significativas, permitem concluir que este sistema apresenta fontes de impactos pontuais do ponto de vista ecológico e ambiental, efeito este que poderá ser agravado caso a biomassa (kg) e área aquícola seja ampliada, sendo necessário um acompanhando periódico desse sistema. Agradecimentos Os autores são gratos a CAPES pelo auxílio financeiro a pesquisa (Projeto PNPD/2009), à Elaine Bartozek Rodrigues pelo fornecimento de dados de alguns fatores abióticos, aos grupos de pesquisa GETECH e GEMAQ da Unioeste – Campus Toledo, pelo apoio logístico na realização das coletas e ao Mestrado em Conservação e Manejo de Recursos Naturais – Unioeste – Cascavel/PR. Referências Agostinho, A. A., L. C. Gomes & F. M. Pelicice, 2007. Ecologia e Manejo de recursos pesqueiros em reservatórios do Brasil. Maringá, EDUEM, 501p. 34 Agostinho, A. A, L. E Miranda, L. M. Bini, L. C. Gomes, S. M. Thomaz. & H. I. Susuki. 1999. Patterns of colonization in neotropical reservoirs, and prognosis on aging. In: Bailly, D., A. A. Agostinho, H. I. Suzuki & E. A. Luiz. 2003. Características reprodutivas relacionadas ao sucesso na colonização de espécies do gênero Astyanax em reservatórios da bacia do rio Iguaçu – PR. Bio Ecologia de Peixes, Maringá, p. 259-265. Agostinho, AA. & LC Gomes. 1997. Reservatório de Segredo: bases ecológicas para o manejo. Maringá, EDUEM, 387p. Andrade, J de, CR. Sanquetta. & C. Ugaya. 2005. Identificação de áreas prioritárias para recuperação da mata ciliar na UHE Salto Caxias. Espaço Energia, 3ª Edição. APHA, AWWA, WEF. 1998. Standard Methods for the Examination of Water and Wastewater. Washington, American Puclic Health Association, 1157p. Araripe, MNBA, Segungo, LFF. Lopes, JB. Araripe, HGA. 2006. Efeito do Cultivo de Peixes em Tanques Rede sobre o Aporte de Fósforo para o Ambiente. Revista Científica de Produção Animal, 8 (2). Ayroza, LMS, FPB. Furlaneto, DMMR. Ayroza & FR. Sussel. 2005. Piscicultura no Médio Paranapanema: situação e perspectivas. Aqüicultura e Pesca, São Paulo, 12: 27-32. 35 Bailly, D, AA. Agostinho, HI. Suzuki & EA. Luiz. 2003. Características reprodutivas relacionadas ao sucesso na colonização de espécies do gênero Astyanax em reservatórios da bacia do rio Iguaçu – PR. Bio Ecologia de Peixes, Maringá, p. 259-265. Baumgartner, G, CS Pavanelli, D Baumgartner, AG Bifi, T Debona & VA Frana. 2012. Peixes do baixo rio Iguaçu. Maringá, Eduem, 203p. Baumgartner, D, G. Baumgartner, CS. Pavanelli, PRL. da Silva, VA. Frana, LC. de Oliveira & MR. Michelon. 2006. Fish, Salto Osório Reservoir, Iguaçu River basin, Paraná State, Brazil. Check List (UNESP), São Paulo, 2 (1): 1-4. Bennemann, ST, ATE. Silva-Souza & GRA. Rocha. 1995. Composicion ictiofaunistica en cinco localidades de la cuenca del Rio Tibagi, PR – Brasil. Interciencia 20 (1): 7-13. Beveridge, M. 2004. Cage Aquaculture. Brackwell publishing. Scotlan. In: Demetrio, JA. Atração de peixes silvestres em áreas de cultivo de tilápias em tanques-rede no rio do Corvo – Reservatório de Rosana – Brasil. Tese de Doutorado. Universidade Estadual de Maringá, Maringá, 28p. Bittencourt, F, A. Feiden, AA. Signor, WR. Boscolo, A. Coldebella & C. Manske. 2009. Densidade de estocagem do Surubim do Iguaçu (Steindachneridion melanodermatum) cultivado em tanque-rede no reservatório José Richa. Congresso Brasileiro de Zootecnia, Águas de Lindóia, São Paulo. 36 Borghetti, JR. & UAT. Silva. 2008. Sistemas de cultivo empregados na aqüicultura continental. Pp. 73-95. In: Ostrensky, A, JR. Borghetti & D. Soto (Eds.). Aquicultura no Brasil: o desafio é crescer. Brasília, FAO, 276p. Bueno, G. W. Marengoni, NG. Gonçalves Júnior, AC. Boscolo, WR. Teixeira, RA. 2008. Estado trófico e bioacumulação do fósforo total no cultivo de peixes em tanques-rede na área aquícola do reservatório de Itaipu. Acta Scientiarum. Biological Sciences. Maringá, 30 (3): 237-243. Companhia Paranaense de Energia - COPEL. 2002. Plano Ambiental de Conservação e uso do reservatório e entorno da usina hidrelétrica Salto Caxias. Curitiba. In: RIBEIRO, LHL, AL. BRANDIMARTE, & RT. KISHI. 2005. Formation of the Salto Caxias Reservoir (PR) – an approach on the eutrophication process. Acta Limnológica Brasiliensia. 17 (2): 155165. Dajoz, R. 1983. Ecologia Geral. Petrópolis, Vozes, 472p. Damato, M. 2001. O emprego de indicadores biológicos na determinação de poluentes orgânicos perigosos. Pp. 229-236. In: Bollmann, HA, NB. Maia, W. Barrela (Eds.). Indicadores ambientais: conceitos e aplicações. São Paulo, EDUC/COMPED/INEP, 285p. Delariva, RL, FC. Canteri, PV. Sanches & G. Baumgartner. 2009. Composição e estrutura da ictiofauna de área marginal da lagoa Xambrê, Parque Nacional de Ilha Grande, PR, Brasil. Revista em Agronegócios e Meio Ambiente, 2 (1): 141-153. 37 Delariva, RL. 2002. Ecologia trófica do rio Iguaçu – PR sob efeitos do represamento de Salto Caxias. Tese de Doutorado, Universidade Estadual de Maringá, Maringá. 65p. Demetrio, JA. 2010. Atração de peixes silvestres em áreas de cultivo de tilápias em tanques-rede no rio do Corvo – Reservatório de Rosana – Brasil. Tese de Doutorado. Universidade Estadual de Maringá, Maringá. 29p. Dempster, T, P. Sanchez-Jerez, JT. Bayle-Sempere, F. Giménez-Casalduero & C. Valle. 2002. Attraction of wild fish to sea-cage fish farms in the south-western Mediterranean Sea: spatial and short-term temporal variability. Marina Ecology Progress Series. (242): 237-257. In: Zanatta, AS. 2011. As interferências das pisciculturas em tanques-rede sobre as assembleias de peixes em grandes reservatórios do rio Paranapanema (bacia do Alto Paraná). Tese de Doutorado, Universidade Estadual Paulista, Botucatu. 73p. Dufech, APS & CB. Fialho. 2009. Estudo comparado da taxocenose de peixes em dois ambientes aquáticos do Parque Estadual de Itapuã, sul do Brasil. Iheringia Série Zoologia, Porto Alegre, 99 (2): 177-188. Feiden, A, AA. Signor, O. Diemer, C. Sary, WR. Boscolo & DH. Neu. 2010. Desempenho de juvenis de jundiás (Rhamdia voulezi) submetidos à alimentação com ração orgânica certificada e comercial. Revista Acadêmica Ciências Agrárias e Ambientais, Curitiba, 8 (4): 381-387. 38 Furlaneto, FPB, DMMR. Ayroza, LMS. Ayroza. 2006. Custo e Rentabilidade da Produção de Tilápia (Oreochromis spp.) em Tanque-rede no Médio Paranapanema, Estado de São Paulo, Safra 2004/05. Informações Econômicas, São Paulo, 36 (3): 63-69. Garavello, JC, CS. Pavanelli & HI. Suzuki. 1997. Caracterização da ictiofauna do rio Iguaçu. Pp. 61-84. In: AGOSTINHO, AA. & LC. GOMES. 1997. Reservatório de Segredo: bases ecológicas para o manejo. Maringá, EDUEM, 387p. Gauch Jr, HG. 1986. Multivariate analysis in community ecology. Cambridge University Press, 1982 (reprinted 1986), Cambridge. Guo, L & Z. Li. 2003. Effects of nitrogen and phosphorus from fish cage-culture on the communities of a shallow lake in middle Yangtze river basin of China. Aquaculture, 226: 201-212. Hammer, Ø, DAT. Harper & PD. Ryan. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 4 (1): 9p. Hoffmann, AC, ML. Orsi & AO. Shibatta. 2005. Diversidade de peixes do reservatório da UHE Escola Engenharia Mackenzie (Capivara), Rio Paranapanema, bacia do alto rio Paraná, Brasil, e a importância dos grandes tributários na sua manutenção. Iheringia Série Zoologia, Porto Alegre, 95 (3): 319-325. Instituto Ambiental do Paraná – IAP, 2009. Monitoramento da Qualidade da Água dos Reservatórios do Paraná no período de 2005 a 2008. 120p + anexos. 39 Jackson, DA. 1993. Stopping rules in principal component analyses: a comparison of heuristical and statistical approaches. Ecology, Washington, DC, 74: 2204-2214. Jongman, RHG, CJF. TER Braak & OFR. Van Tongeren. 1995. Data analysis in community and landscape ecology. Cambridge University Press, Cambridge. Júlio-Júnior, HF, CC. Bonecker & AA. Agostinho. 1997. Reservatório de Segredo e sua inserção na bacia do rio Iguaçu. Pp. 1-17. In: Agostinho, AA, & LC Gomes, (Eds.). Reservatório de Segredo: bases ecológicas para o manejo. Maringá, EDUEM, 387p. Koroleff, F & K. Grasshoff. 1976. Methods of Seawater analysis – verlos chemie, Wheinheinv, New York, p. 117-181. Loureiro, BR, CWC. Branco & E. Zaniboni Filho. 2012. Influence of net-cage fish farming on zooplankton biomass in the Itá reservoir, SC, Brazil. Acta Limnologica Brasiliensia. Lucena, CAS, AS. Jardim & ES. Vidal. 1994. Ocorrência, distribuição e abundância da fauna de peixes da praia de Itapuã, Viamão, Rio Grande do Sul, Brasil. Comunicações do Museu de Ciências e Tecnologia - PUCRS, Série Zoologia, 7: 3-27. Luiz, EA, AC. Petry, CS. Pavanelli, HF. Júlio-Júnior, JD. Latini & VM. Domingues. 2005. As assembleias de peixes de reservatórios hidrelétricos do Estado do Paraná e bacias limítrofes. Pp. 169-184. In: Rodrigues, L, SM. Thomaz, AA. Agostinho & LC. Gomes 40 (Eds.). Biocenoses em reservatórios: padrões espaciais e temporais. São Carlos RiMa, 333p. Mackereth, FJH, J Heron & JF Talling. 1978. Water analysis: some revised methods for limnologist. Kendal: Titus Wilson and Sons LTDA. Freshwater Biological Association Scientific Publication, 121p. Magurran, AE. 1988. Ecological diversity and its measurement. Princeton University Press, New Jersey. Mallasen, M, HP. de Barros & EY. Yamashita. 2008. Produção de peixes em tanques-rede e a qualidade da água. Revista Tecnologia e Inovação Agropecuária, Apta, p.47-51. Matthews, WJ. 1998. Patterns in Freshwater Fish Ecology. Chapman and Hall, Kluwer Academic Press, New York, NY, 757p. McCune, B & MJ Mefford. 1997. PC-ORD: Multivariate analysis of ecological data. Version 3.0. Oregon, USA: MjM Software Design. Ministério da Pesca e Aquicultura – MPA. 2010. Produção Pesqueira e Aquícola: Estatística 2008 e 2009. Disponível em http://www.mpa.gov.br/info-estatistica/estatisticada-pesca-e-aquicultura. Acesso dia 06 de fevereiro de 2012. 41 Navakowski, GC, NS. Hahn & R. Fugi. 2007. Alimentação de peixes piscívoros antes e após a formação do reservatório de Salto Caxias, Paraná, Brasil. Biota Neotrop. 7(2):149154. Oliveira, EC. de, LE. de Santos, LF. Fávaro & V. Abilhoa. 2008. Caracterização da assembleia de peixes em um reservatório recém-formado no Sul do Brasil. Revista Estudos de Biologia, 30: 125-32. ONS – Operador Nacional do Sistema Elétrico. Disponível em: <http://www.ons.gov.br>, Acesso em 31 de junho de 2012. Pielou, EC. 1984. The interpretation of ecological data: a primer on classification and ordination. New York, Wiley. Reis, RE, SO Kullander & Jr Ferraris. 2003. Check list of the freshwater fishes of South and Central America. Porto Alegre, EDIPUCRS, 279p. Strickland, JDH. & TR Parsons. 1972. A pratical handbook of sea water analysis. Fish Res. Board of Canada, Ottawa, 310p. Suzuki, HI. 1999. Estratégias reprodutivas de peixes relacionadas ao sucesso na colonização em dois reservatórios do rio Iguaçu, Brasil. Tese de Doutorado. Universidade Federal de São Carlos, São Carlos, 97p. 42 Tundisi, JG. 2005. Gerenciamento integrado de bacias hidrográficas e reservatórios: estudos de caso e perspectivas. In: Mallasen, M, HP. de Barros & EY. Yamashita. 2008. Produção de Peixes em tanques-rede e a qualidade da água. Revista Tecnologia e Inovação Agropecuária, Apta. 47-51. Tavares, LHS. & SQ. Moreno. 1994. Variação dos parâmetros limnológicos em um viveiro de piscicultura nos períodos de seca e chuva. Revista Unimar, 16: 229-242. Tundisi, JG, T. Matsumura-Tundisi & JEM. Tundisi. 2008. Reservoirs and human well being: new challenges for evaluating impacts and benefits in the neotropics. Brazilian Journal of Biology. 68 (4): 1133-1135. Tundisi, JG & R. Henry. 1986. Effects of enrichment on the summer surface phytoplanktonic community in a stratified tropical lake. Revista Brasileira de Biologia, 46 (1): 231-237. Valle, C, JT. Bayle-Sempere, T. Dempster, P. Sanchez-Jerez & F. Giménez-Casalduero. 2007. Temporal variability of wild fish assemblages associated with a sea-cage fish farm in the south-western Mediterranean Sea. Estuarine, Coastal and Shelf Science, 72: 299-307. Zanatta, AS. 2011. As interferências das pisciculturas em tanques-rede sobre as assembleias de peixes em grandes reservatórios do rio Paranapanema (bacia do Alto Paraná). Tese de Doutorado, Universidade Estadual Paulista, Botucatu. 73p. 43 Zaniboni Filho, E, AP. de Oliveira Nuñer, RM. Guereschi, S. Hermes-Silva. 2005. Cultivo de peixes em tanques-rede e impactos ambientais. Pp. 57-78. In: Cultivo de peixes em tanques-rede: desafios e oportunidades para um desenvolvimento sustentável. Belo Horizonte, EPMIG, 104p. Zar, JH. 1999. Biostatistical analysis. New Jersey, Prentice Hall, 662p. Zawadzki, CH, E. Renesto & LM. Bini. 1999. Genetic and morphometric analysis of three species of the genus Hypostomus Lacépède, 1803 (Osteichthyes: Loricariidae) from the rio Iguaçu (Brazil). Revue Suisse de Zoologie, 106: 91-105. 44 Anexos (Normas da Revista Neotropical Ichthyology) INSTRUCTIONS TO AUTHORS ISSN 1679-6225 printed version ISSN 1982-0224 online version Scope and policy Form and preparation of manuscripts Neotropical Ichthyology taxonomic contribution style sheet Submission of manuscripts Steps followed during manuscript submission Further information Scope and policy Scope and policy The manuscript must contain results of original research on marine and freshwater Neotropical fishes, in the areas of Biology, Ecology, Etology, Physiology, Genetics and Molecular Biology, and Systematics. Scientific notes on morphological or chromatic aberrations, or new records of geographical distribution are not accepted. The Editor and Area Editors will perform a previous analysis of the submitted manuscript to analyze if its contents are appropriate to publish inNeotropical Ichthyology. The Journal is open for submissions to all researchers on Neotropical fish fauna. Payment of publishing costs may apply if none of the author is a member of the Brazilian Society of Ichthyology. Manuscripts that are not formatted according to instructions will not be accepted. Form and preparation of manuscripts Manuscripts Manuscripts must be submitted as digital files. Text must be in Word for Windows or rtf files. Photos and figures must be sent as separate and individual tif or jpg files. Format In taxonomic papers check also: Neotropical Ichthyology taxonomic contribution style sheet Text must be submitted in English. Manuscript must contain the following, in the cited 45 order: - Title - authors names (*) - addresses (do not use a footnote) - Abstract - Resumo - key words (up to five, not repeating title information) - Introduction - Material and Methods - Results - Discussion - Acknowledgments - Literature Cited - Tables - Figure Legend. Manuscripts should not exceed 60 pages, including Figures and Tables. The Editorial Board will decide on possible exceptions. Scientific Notes must contain the following, in the cited order: - Title - authors names (*) - addresses (do not use a footnote) - Abstract - Resumo - key words (up to five, not repeating title information) - text without subtitles, including Introduction, Material and Methods, Results and Discussion - Acknowledgments - Literature Cited - Tables - Figure Legend Scientific Notes will be accepted only if they contain information that justifies immediate publication. Text Text pages cannot include headers, footers, or footnotes (except page number), or any paragraph format. Never use "Tab" or "space" to format references. Text must be aligned to the left, not fully justified. Species, genera and Latin terms (et al., cf., aff., in vitro, in vivo) must be in italics. Do not underline words. Only the following titles should de bold formatted: Abstract, Introduction, Material and Methods, Results, Discussion, Acknowledgements, Literature Cited. 46 List abbreviations used in the text under Material and Methods, except for those in common use (e.g., min, km, mm, kg, m, sec, h, ml, L, g). Measurements must use the metric system. Manuscripts must contain the institutional acronyms and catalog numbers for voucher specimens. Acknowledgments must be concise and include both first and last names. Figures and Tables must be sequentially numbered according to their citation in the text, using the following formats: Fig. 1, Figs. 1-2, Fig. 1a, Figs. 1ab, Table 1, Tables 1-2. The words Table and Fig. must be bold in legends. A list of figure legends must be included at the end of the manuscript. Tables must be constructed using lines and columns, but not "Tab" or "space". Tables can not contain vertical lines or footnotes. Digital files of tables must be formatted in cells. Digital files of tables with columns separated by "Tab" or "space" will not be accepted. Tables and legends must be included at the end of the manuscript, in the following format: Table 1. Monthly variation of the gonadosomatic index in Diapoma speculiferum.... Approximate locations where tables and figures should be inserted must be indicated along the margin of the text. Nomenclature Scientific names should be cited according to the ICZN (1999). Authorship is required only in taxonomic papers and at the first reference of a species or genus. Do not include authorship in the abstract or resumo. Figures Figures must be of high quality and definition. Text included in graphs and pictures must be of a font size compatible with reductions to page width (175 mm) or column width (85 mm). Graphs will be preferably printed as one column (85 mm). Color photos will be accepted only if necessary for the paper and authors will be charged for the cost of printing the color photo. Composed figures must be identified with lower case letters (a, b, ..) in the left lower of each illustration. Composed figures must be prepared so as to fit either the page (175 mm) or column width (85 mm). Illustrations must include either a scale or reference to the size of the item in the figure legend. 47 Literature Cited Use the following formats in the text : Eigenmann (1915, 1921) or (Eigenmann, 1915, 1921; Fowler, 1945, 1948) or Eigenmann & Norris (1918) or Eigenmann et al. (1910a, 1910b). Do not include abstracts or technical reports in Literature Cited. Avoid unnecessary references to thesis or dissertations. Literature Cited must be in alphabetical order, using the following formats: Books: Campos-da-Paz, R. & J. S. Albert. 1998. The gymnotiform "eels" of Tropical America: a history of classification and phylogeny of the South American electric knifefishes (Teleostei: Ostariophysi: Siluriphysi). Pp. 419-446. In: Malabarba, L. R., R. E. Reis, R. P. Vari, Z. M. S. Lucena & C. A. S. Lucena (Eds.). Phylogeny and Classification of Neotropical Fishes. Porto Alegre, Edipucrs, 603p. Thesis/Dissertations: Langeani, F. 1996. Estudo filogenético e revisão taxonômica da família Hemiodontidae Boulenger, 1904 (sensu Roberts, 1974) (Ostariophysi, Characiformes). Unpublished Ph.D. Dissertation, Universidade de São Paulo, São Paulo. 171p. Articles (list full periodic names): Lundberg, J. G., F. Mago-Leccia & P. Nass. 1991. Exallodontus aguanai, a new genus and species of Pimelodidae (Teleostei: Siluriformes) from deep river channels of South America and delimitation of the subfamily Pimelodinae. Proceedings of the Biological Society of Washington, 104(4): 840-869. Articles in press: Burns, J. R., A. D. Meisner, S. H. Weitzman & L. R. Malabarba. (in press). Sperm and spermatozeugma ultrastructure in the inseminating catfish, Trachelyopterus lucenai (Ostariophysi: Siluriformes: Auchenipteridae). Copeia, 2002: 173-179. Internet resources Author. 2002. Title of website, database or other resources, Publisher name and location (if indicated), number of pages (if known). Available from: http://xxx.xxx.xxx/ (Date of access). Neotropical Ichthyology taxonomic contribution style sheet This summary is intended to provide specific information for taxonomic manuscripts only. 48 Generic accounts Order of presentation: Name Author, Year (or new genus [Do not abbreviate.]) [Synonymy] Type species. Diagnosis. (see below the CONSIDERATIONS ON HOW TO PREPARE DIAGNOSES) Etymology. (for new species only) [Remarks.] Key to species. Comments on above: Type species: For newly proposed genera, the original Name of the proposed type species, followed by Author and Year of publication (or new species) is sufficient. For previously proposed generic names, the following additional information is required (in this order): Nature of type designation (e. g., original designation, monotypy, absolute tautonymy. etc). If the type species was not designated in the original publication, the author, year and page of the designation should be cited (e. g., Type by subsequent designation by Jordan, 1919: 45). Diagnosis: diagnoses should NOT be written in telegraphic style (for clarity purposes). Generic diagnoses preferably should list the unique synapomorphies of the genus, followed by homoplastic derived characters and/or other useful distinguishing characteristics. Etymology: For new names, state the gender, even though it may be obvious from the construction. Do not give an etymology for preexisting names. If it is necessary to discuss the etymology of an old name (for example, to justify an interpretation of its gender), put that in the Remarks section. Species accounts Order of presentation: Name, Author, Year (or "new species" [Do not abbreviate.]) [Synonymy] Holotype. [for new species only - include full collection data (see details, below)] [Paratype(s).] [for new species only - include full collection data (see details, below)] [Non-types .] [for new species only - include reduced collection data (see details, below) (Justification for separating non types should be provided in Remarks)] 49 Diagnosis. (see below the CONSIDERATIONS ON HOW TO PREPARE SPECIES DIAGNOSES) Description. Coloration. Size. [Sexual dimorphism.] Distribution. [Ecological notes.] Etymology. [for new species only] [Remarks.] [Material examined.] (for accounts of previously named species) Some comments on the above listed categories: Types: Should be listed separately from other material examined only for new species. Should include full collection data, in the following order: Catalog number, # specimens (except for holotype), size range, number and size range of measured specimens, if different - in parentheses, locality, date of collection [in Day, Month (3 letter abbreviation only) and Year format], and collector(s) (e.g., LIRP 5640, 25, 38.5-90.3 mm SL (12, 75.0-90.3 mm SL), Brazil, São Paulo, Município de Marapoama, rio Tietê basin, ribeirão Cubatão at road between Marapoama and Elisiário, 21º11'35"S 49º07'22"W, 10 Feb 2003, A. L. A. Melo). Diagnosis: diagnoses should NOT be written in telegraphic style (for clarity purposes). Description: In telegraphic style (i. e., no verbs or articles) Coloration: In telegraphic style (i. e., no verbs or articles), may be divided in Color in alcohol and Color in life. Etymology: For new names, state the usage (adjective, noun, patronym, etc.), even though it may be obvious from the construction. Do not provide an etymology for preexisting names, unless the etymology is necessary to justify the spelling. In such cases, this information belongs in the Remarks and not as a separate heading. Material examined: Provide only locality, catalog number, number of specimens and size range. In addition, indicate any types by: (Holo- Syn-, etc.) type of Xus yus Author, date. For Lectotype or Neotype, also provide citation for source of designation (e.g., USNM 123456, 75 mm SL, Amazon River near Manaus, lectotype of Xus yus, Author, date, designated by Isbrücker (1971: 85) [or designated herein]). Specimen lots should be arranged by Country, then by State or Territory, then by river basin, if relevant. Country should be written in Bold font and should not be 50 repeated after the first usage in a species account. Specific issues: Scientific names must always include the generic name, or at least an abbreviation for the generic name. This applies to tables and figure captions, as well as the text of the manuscript. Typically, the whole generic name should be spelled out in full at the first usage in each paragraph. Thereafter, an abbreviation can be used provided that there is no possibility of confusion with another generic name. Bilaterally paired structures must be treated in the singular (e. g., pelvic fin short, not pelvic fins short) Compound adjectives that include a noun should be connected by a hyphen (e. g., pectoral-fin spine, NOT pectoral fin spine). Fin-ray formulae should be reported with unbranched rays in lower case Roman numerals, spines in upper case Roman, and branched rays in Arabic numerals. Transitions between different types of rays should be indicated by a comma (,) and not a plus sign (+), or dash (-) (e. g, iii,7 or II,9. Not iii-7 or iii+7; no spaces should be inserted after the comma). We treat the catfish spinelet as a spine, so dorsal fin counts that include a spinelet should be reported as II,6 (or whatever the branched ray count is). Latitude and Longitude: No spaces between numerals and symbols. For degree sign, use Control +@, space (in MS Word) and not superscript O; for seconds, do not use the single quote mark twice, use the double quote mark (Shift quote). Percents: no space between numeral and % (e. g. 25%). Revisions and reviews Species accounts should be in alphabetical order. CONSIDERATIONS ON HOW TO PREPARE SPECIES DIAGNOSES A species diagnosis is typically a paragraph constructed of full sentences which list the most important traits that allow the reader to unequivocally identify the species. Ideally, the diagnosis includes one or more features that are unique to the species, preferably autapomorphic characters. If unique features were not discovered, the next best option is a differential diagnosis, within which a series of direct comparisons are made among species and the alternative character states specified by contrasts are stated explicitly (using "vs." followed by the condition found in the species, 51 or group of species, being compared, for each diagnostic feature). Diagnoses that consist only of a combination of characters (i.e., traits listed sequentially which, when considered together, distinguish the species from congeners) in many cases fail to make a convincing case that the species warrants recognition, mostly because too little information is offered in the way of direct comparisons with congeners. For that reason, this form of diagnosis should be avoided. Submission of manuscripts Send manuscripts electronically to the Editor through the journal website (http://www6.ufrgs.br/seerni/ojs/index.php) or through e-mail at [email protected]. Send a statement that it constitutes original research and is not being submitted to other journals. In multi-authored papers, indicate the name and address of the author responsible by all contacts and the e-mail addresses of co-authors. Indicate the area of Ichthyology (Biochemistry and Physiology, Biology, Ecology, Etology, Genetics and Molecular Biology, Systematics) to which the manuscript is referable. List three possible referees (name, institution, country and e-mail). Steps followed during manuscript submission 1 - Manuscripts and additional documents are sent to the Scientific Editor. 2 - The Scientific Editor analyses the contents, format and enclosed documents. 3 - If the manuscript fills all requirements, the Scientific Editor forward all documents to the Editor of the area related to the manuscript. 4 - The Area Editor analyses the contents of the submitted manuscript. 5 - If its contents are considered appropriate to the journal, the Area Editor send the manuscript, along with an evaluation form, to two independent reviewers (Referees). 6 - The Referees make a critical analysis of the contents of the manuscript, making the necessary suggestions for their improvement, and giving their expert opinion regarding its acceptance or refuse for publication. 52 7 - The Area Editor receives referees review and evaluation of the manuscript. - If recommended without corrections the Area Editor forwards the manuscript to the Scientific Editor; - If recommended with corrections the Area Editor returns the manuscript to Author, with the suggestions of the Referees; - If not recommended, the manuscript is returned to Author. 8 - When receiving the manuscript with referees comments and editorial notes, author must make corrections immediately (maximum four weeks) and return to the area editor. If Author does not agree with referees suggestions, must justify its no adoption. 9 - The Area Editor receives the final version of the manuscript, check if necessary changes/improvements were made accordingly, and forward all documents (final version of the manuscript and illustrations) to the Scientific Editor. 10 - The Scientific Editor receives the documents, makes a final review of format and forward to printer. 11 - Proofs are reviewed by the Scientific Editor and forward to Author for review. Further information Luiz R. Malabarba Editor Laboratório de Ictiologia Departamento de Zoologia Universidade Federal do Rio Grande do Sul Av. Bento Gonçalves, 9500 - bloco IV - Prédio 43435 91501-970 - Porto Alegre, RS - Brazil Fone: 55 51 33087719 Fax: 55 51 33087696 e-mail: [email protected]; [email protected] www.ufrgs.br/ni [Home] [About the journal] [Editorial board] [Subscriptions] All the content of the journal, except where otherwise noted, is licensed under a Creative Commons License Departamento de Zoologia - IB Universidade federal do Rio Grande do Sul Av. Bento Gon�alves, 9500 - bloco IV - Pr�dio 43435 91501-970 - Porto Alegre, RS - Brasil 53 Tel.: 55-21-2568-8262 [email protected]
Download