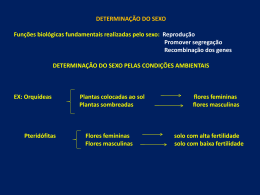
i UNIVERSIDADE FEDERAL DA PARAÍBA CENTRO DE CIÊNCIAS AGRÁRIAS PROGRAMA DE PÓS-GRADUAÇÃO EM ZOOTECNIA COMPORTAMENTO SEXUAL DO MACHO E MATERNO-FILIAL DE OVINOS DA RAÇA MORADA NOVA JAQUELINE DA SILVA TRAJANO Zootecnista AREIA – PB FEVEREIRO/2011 ii COMPORTAMENTO SEXUAL DO MACHO E MATERNO-FILIAL DE OVINOS DA RAÇA MORADA NOVA iii JAQUELINE DA SILVA TRAJANO COMPORTAMENTO SEXUAL DO MACHO E MATERNO-FILIAL DE OVINOS DA RAÇA MORADA NOVA Dissertação apresentada ao Programa de PósGraduação em Zootecnia da Universidade Federal da Paraíba, Centro de Ciências Agrárias Areia, em cumprimento as exigências ao titulo de Mestre em Zootecnia, Área de concentração: Produção Animal. Comitê de Orientação: Prof. Dr. Edgard Cavalcanti Pimenta Filho - Orientador Principal (CCA/UFPB) Prof. Dr. Ariosvaldo Nunes de Medeiros – Co - Orientador (CCA/UFPB) Prof. Dr. Edilson Paes Saraiva – Co - Orientador (CCA/UFPB) AREIA-PB FEVEREIRO 2011 iv Departamento de Zootecnia/CCA Presidente v Se tu o desejas, podes voar, só tens de confiar muito em ti. Mensagem vi Aos meus Pais Risonete da Silva Trajano e Geraldo da Silva Trajano Ao meu irmão Geraldo da Silva Trajano Júnior E em especial ao meu filho, razão da minha vida Mateus Trajano Muniz Dedico vii AGRADECIMENTOS A Deus por todos as portas que ele tem aberto em minha vida e por ter proporcionado mais essa conquista. Aos meus pais pelo amor e apoio em todas as horas que precisei. Ao amor incondicional do meu pequeno príncipe (meu filho) que soube proporcionar muitas alegrias em minha vida nos momentos mais difíceis. A Universidade Federal da Paraíba e ao Programa de Pós-Graduação em Zootecnia pela oportunidade de realização deste mestrado. A Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES, pela concessão da bolsa de estudos. Ao meu orientador Prof. Edgard Cavalcanti Pimenta Filho pela grande oportunidade a mim concedida de ter feito parte da minha via profissional e pessoal, agradeço pela compreensão e apoio nos momentos difíceis durante o experimento. Nem todas as palavras que possa expressar aqui serão suficientes para agradecer a pessoa maravilhosa que o senhor foi durante minha formação enquanto Zootecnista. Ao meu segundo orientador e amigo, Prof. Edilson Paes Saraiva agradeço todo o apoio, conselhos e extrema ajuda em todos os momentos dessa fase tão importante. Agradeço ao Corpo Docente do Programa de Pós Graduação em Zootecnia, PPGZ/CCA. Em especial aos professores Dr. Severino Gonzaga Neto, Dr. Walter Esfrain Pereira, Dra. Carla Saraiva, Profª. Dra. Juliana Silva de Oliveira, As funcionárias do PPGZ e da estação Experimental de São João o Cariri: Seu Son, Inacinho, Seu Jaime, por todos os momentos vividos e pela disposição em ajudarmos sempre que precisamos. Aos meus grandes amigos e companheiros que foram essenciais na execução do trabalho: Vinicius, Mucureba, José Helder, Adriano, Cintia, Ana Jaqueline, Serjão, Luana, Lígia, Rinaldo, Vanúbia, Rosa, Giu, Uriel, e todos que não foram citados, mas que também auxiliaram de alguma forma na execução desse trabalho. viii BIOGRAFIA DA AUTORA JAQUELINE DA SILVA TRAJANO - Nascida em 20 de novembro de 1985, na cidade de Santa Rita - PB e residente no município de Santa Rita - PB, onde estudou na Escola Estadual Maria de Lourdes Araújo durante o ensino fundamental, tendo concluído em 2000. Em 2001, ingressou na Escola Estadual de Ensino Fundamental e Médio Luiz de Azevedo Soares - PB, concluindo o ensino médio em 2003. Em abril de 2009, formou-se em Zootecnia, pelo Centro de Ciências Agrárias da Universidade Federal da Paraíba. Quando acadêmica do Curso de Zootecnia, desenvolveu pesquisa na área de produção de ruminantes, sendo bolsista no 5ª semestre, ano 2006, do Programa de Iniciação Cientifica (Conselho Nacional de Desenvolvimento Cientifico e Tecnológico – CNPq), durante 30 meses. Em Março de 2009, ingressou no Curso de Mestrado em Zootecnia da Universidade Federal da Paraíba (UFPB), sob a orientação do professor Dr. Edgard Cavalcanti Pimenta Filho, no qual foi bolsista da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES, desenvolvendo sua pesquisa na área de Comportamento Animal. ix SUMÁRIO Página Lista de Tabelas........................................................................................................ x Resumo Geral............................................................................................................ xii Abstract..................................................................................................................... xiii Capítulo 1. Referencial Teórico............................................................................... 01 Referências Bibliográficas........................................................................................ 09 Capítulo 2. Comportamento Sexual de carneiros da Raça Morada Nova................ 13 Resumo...................................................................................................................... 14 Abstract..................................................................................................................... 15 Introdução................................................................................................................. 16 Material e Métodos................................................................................................... 17 Resultados e Discussão............................................................................................ 21 Conclusões................................................................................................................ 31 Referências Bibliográficas........................................................................................ 32 Anexos...................................................................................................................... 34 Capítulo 3. Caracterização do Comportamento Materno-Filial no Pós-Parto de ovelhas Morada Nova............................................................................................... 38 Resumo...................................................................................................................... 39 Abstract..................................................................................................................... 40 Introdução................................................................................................................. 41 Material e Métodos................................................................................................... 43 Resultados e Discussão............................................................................................ 47 Conclusões................................................................................................................ 56 Referências Bibliográficas........................................................................................ 57 Implicações e Sugestões............................................................................................ 59 x LISTA DE TABELAS E QUADROS Capítulo 2 Página Quadro 1. Média de temperatura máxima, mínima e umidade relativa dos meses 18 experimentais na fase de acasalamento..................................................................... Tabela 1. Percentagem de prenhez entre estações de monta em ovinos Morada Nova.......................................................................................................................... 21 Tabela 2. Número de fêmeas expostas, com repetição de cio, com prenhez positiva por reprodutor e por estação de monta........................................................ 23 Tabela 3. Média do Tempo de Reação (TR) em segundos e Freqüência do Número de Saltos por tentativa (NST) de ovinos da raça Morada Nova.......................................................................................................................... 24 Tabela 4. Médias das frequências dos padrões de Comportamentos sexuais: cortejo da fêmea (CF); cortejo com a pata (CP); vocalização (VOC); exposição de pênis (EXP); cheirar a região urogenital da fêmea (CRUF) e reflexo de flehmen (RF) apresentados por machos ovinos da raça Morada Nova..................... 25 Tabela 5. Média do Tempo de Reação (TR), em segundos, e média da Freqüência do Número de Saltos por Tentativa (NST) dos reprodutores em relação ao turno de acasalamento.............................................................................. 28 Tabela 6. Médias das freqüências dos padrões de Comportamentos sexuais: Cortejo com a Pata (CP); Vocalização (VOC); Exposição da Língua (EXL); Exposição de Pênis (EXP); Cheirar a Região Urogenital da Fêmea (CRUF) e Reflexo de Flehmen (RF) apresentados em ovinos da raça Morada Nova em relação ao turno de acasalamento.............................................................................. 29 xi Capítulo 3 Página Tabela 1. Frequência das latências para tocar a cria em segundos, ficar em pé e realiza a primeira mamada em minutos, relacionados com o tipo de parto.......................................................................................................................... 48 Tabela 2. Média das Frequências de Posturas e Atividades das ovelhas em relação ao tipo de parto segundo o lote de parição................................................... 49 Tabela 3. Frequências das Posturas e Atividades das crias em relação ao tipo de parto em dos lotes de parição.................................................................................... 52 Tabela 4. Correlação dos comportamentos entre as mães dos dois lotes................. 54 Tabela 5. Correlação dos comportamentos entre as crias dos dois lotes................. 55 xii Comportamento Sexual do macho e Materno-Filial de ovinos Morada Nova Resumo geral: A raça ovina Morada Nova tem sido considerada alvo de esforços integrados por vários grupos de pesquisa de diferentes instituições intuito de conhecermos melhor como explorarmos de forma eficiente. Objetivou-se caracterizar o comportamento sexual e materno-filial de ovinos Morada Nova nas condições de semiárido. Foram registrados todos os eventos relativos à corte do macho perante uma fêmea em cio, registrados em planilhas específicas durante o acasalamento. Os resultados das variáveis referentes aos padrões de comportamento sexual dos machos foram influenciados por características individuais. Isso provavelmente pode ser atribuído a pouca experiência sexual do macho, o que leva a supor que o maior tempo despendido no cortejo poderia ser atribuído a questão da experiência do macho. Após o parto foram observados os comportamentos entre mãe e cria, num período de 2 horas, sendo registrados todos os comportamentos expressados pelas fêmeas. Os resultados obtidos demonstraram que as ovelhas da raça Morada Nova expressam todos os comportamentos iniciais necessários para garantir a sobrevivência da sua cria. Além disso, os resultados observados mostraram que as crias se apresentaram mais ágeis quanto ao tempo de ficar em pé e realizar a primeira mamada independente do lote. Concluiu-se que os ovinos desta raça apresentam a maioria dos padrões do comportamento sexual, durante as fases de cortejo e cópula da fêmea em estro com as diferenças comportamentais observadas atribuídas a características individuais. Além das fêmeas apresentarem uma excelente habilidade materna para as condições do semiárido, sendo fundamental na preservação da espécie, agilidade e sobrevivência da cria até o desmame. Palavras-Chave: etologia, fêmeas, relação mãe-cria, reprodução, sobrevivência. xiii Male and Sexual Behavior Branch of the Maternal and Morada Nova sheep ABSTRACT: The Morada Nova sheep breed has been considered the target of integrated efforts by several research groups from different institutions order to know better how to exploit efficiently. This study aimed to characterize the sexual behavior and mother-infant Morada Nova sheep in semiarid conditions. We recorded all events relating to the cutting of the male before a female in heat, registered in specific worksheets during mating. The results of the variables related to sexual behavior patterns of males were influenced by individual characteristics. This can probably be attributed to inexperienced male sexual, which leads to the supposition that the more time spent in courtship could be attributed to the issue of male experience. After delivery behaviors were observed between mother and soon, within 2 hours, and recorded all behaviors expressed by females. The results showed that sheep Morada Nova express all initial behaviors needed to ensure the survival of their offspring. Moreover, the observed results showed that the pups were more agile as the time to stand and deliver the first batch of independent feeding. It was concluded that the sheep of this breed have most patterns of sexual behavior during the stages of courtship and copulation the female in estrus with the observed behavioral differences attributed to individual characteristics. Besides the females present an excellent maternal ability to semiarid conditions, being instrumental in the preservation of the species, agility and survivability of the offspring to weaning. Keywords: ethology, females, mother-offspring relationship, reproduction, survival. 1 Capítulo 1 Referencial Teórico COMPORTAMENTO SEXUAL DO MACHO E MATERNO-FILIAL DE OVINOS DA RAÇA MORADA NOVA 2 1. Referencial teórico No Nordeste, em decorrência do sistema extensivo de criação e em associação às condições do semiárido brasileiro, os ovinos sofreram uma seleção natural ao longo dos séculos. Esta seleção levou ao desenvolvimento de animais cujas principais características são rusticidade, boa capacidade de reprodução e pele de ótima qualidade (Azevêdo et al., 2003). O desempenho reprodutivo, especialmente a habilidade materna e a taxa de crescimento de seus cordeiros, estão entre os componentes responsáveis pelo sucesso da produção (Macedo et al., 2003). Os ovinos representam para a região Nordeste uma fonte de alimento proteico a baixo custo, apresentando uma inconteste importância socioeconômica para a população da zona rural. A exploração de raças nativas no Nordeste, desde muitos anos, tem trazido uma grande vantagem, principalmente no uso de áreas de Caatinga. Animais, como os da raça Morada Nova, tendem a contribuir com uma produção eficiente por serem bem adaptados às condições do semiárido (Fernandes et al., 2001), com características específicas tais como excelente habilidade materna, altos índices de fertilidade e prolificidade. A raça Morada Nova constitui-se em uma das principais raças nativas de ovinos deslanados do Nordeste Brasileiro, explorados para carne e pele, sendo esta altamente apreciada no mercado internacional. Nas últimas décadas o estudo do comportamento animal vem sendo utilizado no intuito de auxiliar os produtores para que eles possam maximizar a produção animal e o estudo desta raça pelo pode contribuir para melhorias na exploração de ovinos no semiárido. Del-Claro (2004) vai um pouco além e define “estudar o comportamento” como sendo avaliar e mensurar todo o ato executado por um animal, perceptível ou não, ao universo sensorial humano. Cada vez mais o comportamento tem se tornado uma ciência multidisciplinar, envolvendo técnicas de outras subdisciplinas da biologia como a bioquímica, genética, botânica, ecologia, além de manipulações experimentais. Mesmo quando um animal aparentemente não está fazendo nada, esse "não fazer nada", também representa um tipo de comportamento e tem sua função. Animais sociais como os ovinos podem comer mais e digerir melhor os alimentos quando estão em grupo. Da mesma forma, rotinas de criação aplicadas de 3 forma biologicamente inadequada podem diminuir a taxa de produção dos animais (Jensen, 2002). A etologia é uma ciência que pode oferecer muitos tipos de aplicações em situações em que os seres humanos utilizam animais para vários fins (Ferraz, 2011). A intensificação dos estudos de comportamento na área de produção visa uma exploração mais eficiente e obtenção de produtos de maior qualidade sem comprometimento dos animais. Segundo Snowdon (1999), a conservação de uma espécie requer uma compreensão do seu comportamento em seu habitat natural como: padrões migratórios, tamanho de território, interações com outros grupos, demanda de forrageio, comportamento reprodutivo e comunicação. O comportamento animal vem sendo estudado desde a antiguidade. Os homens pré-históricos estudavam o comportamento dos animais à sua volta, seja para se alimentar, se defender, domesticá-los ou apenas conhecê-los. Na atualidade, o estudo do comportamento vem se destacando em uma maior proporção devido à grande contribuição no manejo, nas adequações das instalações de acordo com a espécie, sexo, idade ou fase fisiologia e até no próprio bem-estar dos animais (Ferraz, 2011). Com isso, a caracterização e a compreensão do comportamento natural representam um aspecto central da etologia (Jensen, 2002). Entre esses aspectos, principalmente quando se trata de raças nativas, se mostra de grande relevância o estudo do comportamento sexual e materno-filial, pois existem alguns fatores que podem afetar a conduta sexual dos animais, como fatores genéticos, fisiológicos, estação do ano, raça, níveis hormonais (andrógenos), sanidade, manejo, dominância, idade, ambiente, interação sócio-sexual experiência prévia do indivíduo e ordem de dominância social (Santos et al., 2006) que necessitam serem avaliados com intuito de maximizar a produção desses animais em áreas de caatinga. Na maioria das espécies, a expressão do comportamento sexual depende tanto de fatores internos como os níveis de hormônios esteróides, estado nutricional, incluindo o ambiente físico e social além do fotoperíodo ou estrutura social do grupo (Fabre-Nys, 2000). O comportamento sexual é uma resposta aos estímulos externos, que normalmente são ferormônios, substâncias químicas secretadas pelo animal na urina, fezes ou glândulas, que causam respostas específicas no indivíduo da mesma espécie (Rekwot et al., 2001) 4 A avaliação do comportamento reprodutivo permite a elaboração de um padrão que norteie a identificação de carneiros com eficiência de desempenho sexual, expresso pela demonstração de capacidade de reconhecer fêmeas em cio e efetuar a cobrição. Carneiros classificados como de alto desempenho reprodutivo deixam maior número de descendentes (Stellflug et al., 2006). A identificação e seleção de machos para serem utilizados como reprodutores é uma ferramenta extremamente importante, pois carneiros com melhor desempenho sexual provavelmente servirão um maior número de ovelhas em um curto período de tempo, aumentando a pressão de seleção e a disseminação do material genético desejável. As principais características a serem avaliadas na escolha de potenciais reprodutores incluem a qualidade seminal, as características testiculares e o comportamento sexual (Pacheco et al., 2008). Mellado et al. (2000) citam que nas condições de criação em regiões áridas e semiáridas, os machos percorrem grandes distâncias tanto à procura de alimentos como de fêmeas no cio, o que leva a um significante estresse durante este período, especialmente nos casos de alta relação macho e fêmea, altas temperaturas ambientais e vegetação escassa. Considera-se que os resultados advindos de estudos do comportamento sexual envolvendo a capacidade de monta ao longo do dia, a frequência ejaculatória, a libido, variações no peso corporal, constituem importantes bases de informações para o estabelecimento de programas reprodutivos em condições de monta a campo. Macedo e Alvarado (2005), trabalhando com ovelhas em diferentes sistemas de produção e épocas de monta, observaram que a produtividade foi maior quando houve maior intensificação dos sistemas, mostrando que a eficiência de uma propriedade também está relacionada com a tecnologia ou manejo adequado que esta utiliza. O comportamento sexual, tanto em machos como em fêmeas, pode ser dividido em duas fases. A primeira, conhecida como pré-copulatória, é chamada fase de "excitação" e depende, essencialmente, da motivação dos parceiros sexuais. Durante esta fase, o indivíduo procura manter contato com o companheiro e o incentiva numa interação sexual (Fabre-Nys, 2000). De acordo com Rutter (2002), os machos utilizam estímulos olfativos e gustativos para detectar fêmeas no cio. Conhecido como comportamento de Flehmen, o macho cheira a urina da fêmea e, em seguida, fica parado, com a cabeça levantada e os lábios curvados, sendo esta investigação olfatória controlada por ação da 5 testosterona (Ladewig et al., 1980) e que dura normalmente entre 10 e 30 segundos nos machos caprinos e ovinos. Estes autores citam que ao ser desencadeado o efeito Flehmen nos machos, estes já teriam previamente identificado as fêmeas no cio, sugerindo-se que este ato seja uma confirmação do diagnóstico e um estímulo para excitação sexual. A fêmea também cheira o corpo e genitais do macho, nesta fase de acasalamento. Este comportamento de abordagem genital de ambos os sexos os levam a circular o outro. O macho faz grunhidos de corte e lambe a genitália da fêmea durante a abordagem sexual, depois desloca a fêmea, e ela vira a cabeça para trás sendo o inicio da imobilização que antecede a cópula (Rutter, 2002). Na segunda fase, chamada de fase de "consumação", ocorre o acasalamento propriamente dito. Um dos principais fatores de variação no comportamento sexual nos machos é o nível de testosterona. A primeira indicação do papel da testosterona é a observada no comportamento sexual do machos (Fabre-Nys, 2000). Após a corte, o macho desloca a fêmea e ela vira a cabeça para trás. Para o macho, o acasalamento termina quando a fêmea torna-se imóvel e adota uma postura que permite que o macho a monte. Se a fêmea é receptiva, a cópula pode ocorrer rapidamente, e é muito breve, o macho move a cabeça rapidamente para trás, e ocorre a ejaculação com o comportamento característico dos ruminantes chamado de arranque. Após o acasalamento, os carneiros esticam sua cabeça e pescoço (Rutter, 2002). Após o acasalamento, inicia-se outra fase na vida da fêmea: a fase gestacional, no caso das ovelhas que ficaram prenhez. Nesse período tanto a fisiologia quanto o comportamento da fêmea são modificados devido às transformações ocorridas. O parto é um momento muito importante no ciclo reprodutivo da ovelha que tem seu comportamento profundamente alterado entre algumas horas e alguns dias anteriores ao parto. A ovelha transforma-se de um animal altamente gregário para um animal isolado do rebanho (Silva et al., 2010a). Para a formação do laço materno-filial é necessário que haja um conjunto de comportamentos de atenção, incluindo a vocalização, o ato de cheirar, lamber e proteger o neonato. Estes comportamentos são direcionados à formação de um vínculo olfatório exclusivo entre a ovelha e o cordeiro Por muito tempo, acreditou-se que a ocitocina seria responsável apenas pelas contrações uterinas e ejeção de leite. No entanto, esse hormônio parece ser, também, a chave que estimula o comportamento materno (Lévy et al., 2004). 6 O comportamento materno envolve uma variedade de hormônios, a experiência como mãe, tendências hereditárias e o estímulo do neonato (Reece, 2006). Nas primeiras horas após o parto as principais funções dos cuidados maternos em mamíferos são fornecer colostro, para satisfazer as necessidades metabólicas do recém-nascido, e protegê-lo de agressão por parte do ambiente externo (Nowak e Poindron, 2006). Segundo Dwyer et al. (1998), os ovinos têm uma vocalização específica ao parto: um balido de tom baixo ou ronco, que é feito exclusivamente para o cordeiro, e de grande importância no estabelecimento do vínculo ovelha e cordeiro. Em seu experimento, constataram que esse balido foi menor em ovelhas primíparas do que em multíparas, concordando com a questão do comportamento materno de ovelhas de primeira cria, que desenvolvem um vínculo com seu filhote muito mais lentamente. De acordo com Paranhos da Costa e Cromberg (1998), com a proximidade do parto ocorrem algumas alterações comportamentais que podem ter início desde alguns dias até poucas horas antes do nascimento, demonstrando a possibilidade de se selecionar animais através de suas habilidades para cuidar da prole, associando o temperamento materno ao vigor da cria. Com isso, se podem tomar medidas mais precisas do efeito materno nos programas de melhoramento genético animal. As ovelhas recém paridas despendem seu tempo, principalmente, com comportamentos voltados à formação de um vínculo exclusivo com a cria e com cuidados importantes para a sobrevivência do cordeiro, como a limpeza corporal do neonato, enquanto que a cria após se levantar e locomover, procura ingerir o colostro (Rainere, 2008). O período no qual a ovelha forma uma ligação com seu cordeiro é de cerca de 24 horas, sendo necessária a exposição do cordeiro por apenas alguns minutos que ocasionam alterações hormonais desencadeando o comportamento maternal (Reece, 2006). Poucas horas depois do nascimento a ingestão de colostro deve ser feita para aquisição de imunidade e energia, conferindo resistência a doenças e aumentando as chances de sobrevivência da cria. Para que isto ocorra, é desejável que a mãe inicie o quanto antes o processo de limpeza da cria, com consequente formação de vínculo afetivo e que permita ao recém-nascido acesso ao alimento, ou seja, não dificulte a mamada. Neste aspecto, são relevantes duas características: o comportamento materno (de aceitar a aproximação e manipulação do úbere ou tetos pela cria) e a conformação do aparelho mamário (Schmidek, 2004). 7 A mãe é responsável pela sobrevivência imediata do recém-nascido, que nas diferentes espécies não nasce com o mesmo grau de maturidade motora, sensorial ou de termorregulação e, consequentemente, exige um comportamento perfeitamente adaptado da mãe à sua necessidade. Nas espécies do grupo dos ungulados, como bubalinos, ovinos, bovinos e caprinos, os recém-nascidos são classificados como precoces por seguirem as mães pouco tempo após o nascimento (Poindron, 2005). Para Dwyer (2008), essas características do comportamento materno são expressas após o nascimento associados à sobrevivência das crias e são divididas em duas categorias. A primeira é de cuidados com o cordeiro que facilitam a transição da vida pré-natal para a pós-natal; esses comportamentos incluem balidos de baixa intensidade (uma vocalização específica para a comunicação com a cria, lambidas ou grooming). A segunda categoria é a da formação de uma memória que permita o reconhecimento do cordeiro, dirigindo os cuidados maternos apenas para a sua própria cria. Grandinson (2005) aponta como componentes principais do comportamento materno o estabelecimento de vínculo entre mãe e filho, o comportamento de cuidado, a receptividade, a atenção para com a prole e proteção contra intrusos. Segundo Rech et al. (2008), o comportamento materno se modifica de acordo com a idade e experiência da ovelha, relatando que quanto maior o tempo que os cordeiros permanecem com a mãe aumenta a probabilidade de sua sobrevivência. O comportamento materno tem grande influência sobre a sobrevivência da prole desde o aleitamento ou até o desmame. A manutenção do comportamento materno é promovida pela ação hormonal (estradiol, prolactina, ocitocina e opióides) e sensorial (audição, olfato e visão). O olfato é extensivamente usado em muitos aspectos maternais em mamíferos, assegurando a coordenação das interações materno-filiais e, por conseguinte, o desenvolvimento normal da descendência. Fora do período de parição e lactação, componentes olfatórios exercem um papel de inibição da responsabilidade materna, fazendo com que as fêmeas não prenhes, ou em estádio inicial de gestação, considerem aversivo o odor de um jovem (Levy et al., 2004). A relação mantida entre mãe e neonato durante o período colostral depende muito do que ocorre no chamado período sensível, que é um período sensório específico, dependente de fatores internos da mãe relacionados com o parto, que ativam o comportamento materno em um primeiro momento. Este período é 8 caracterizado pela receptividade materna a algumas pistas táteis, olfativas e ou auditivas fornecidas pelo neonato e é nele que se estabelecem os laços que garantirão o empenho materno na sobrevivência do recém-nascido até à desmama (Poindron, 2005). Apesar dos estímulos visuais serem, sem dúvida, importantes no reconhecimento individual, o fato de ovinos gastarem muito tempo se cheirando, especialmente quando um animal estranho é introduzido no grupo, implica que o olfato também é importante a este respeito. À medida que o cordeiro cresce e melhora o reconhecimento mútuo, o cordeiro e a ovelha ficam mais distantes um do outro. A ovelha pode chamar o cordeiro para mamar, mas ela também mostra uma tendência crescente para encerrar o aleitamento quando o cordeiro cresce. Ovelhas com duas crias só permitirá a mamada quando ambos os cordeiros estiverem presentes, indicando que ela reconhece que tem dois filhos (Rutter, 2002). A habilidade da matriz, a condição de escore corporal, a idade e o estado imunológico da fêmea influenciam no comportamento materno-filial e no desenvolvimento dos cordeiros, além disso, podem contribuir para um maior ou menor cuidado maternal as alterações fisiológicas, o clima, o grupo genético e o estresse. Desta forma, para diminuir o índice de perdas nesse período é fundamental o conhecimento da etologia, pois, desta maneira, pode-se utilizar o comportamento como ferramenta seletiva de matrizes, avaliando-se a habilidade, possíveis correlações dos cuidados maternos com o peso das crias ao desmame e com o índice de sobrevivência (Silva et al., 2010b). 9 2. REFERENCIAS BIBLIOGRÁFICAS AZEVÊDO, D.M.R.; et al. A Ovinocultura no Mundo e no Brasil: Uma Realidade. Ciência Rural, v.33, n.2, p.357-361, 2003. DEL-CLARO, K. Comportamento Animal: uma introdução a ecologia comportamental. Jundiaí: Livraria Conceito, v.1, 2004, 132p. DWYER, C.M.; MCLEAN, K.A.; DEANS, L.A. et al. Vocalisations between mother and young in sheeps: effects of breed and maternal experience. Applied Animal Behaviour Science, v.58, n.1-2, p.105-119, 1998. DWYER, C. M. Genetic and physiological determinants of maternal behavior and lamb survival: Implications for low input sheep management. Journal Animal Science, v.86, p.246-258, 2008. FABRE-NYS, C. Le Comportement sexuel des caprins: contrôle hormonal et facteurs sociaux. Inra Productions Animales, v.13, n.1, p.11-23, 2000. FERNANDES, A.A.O.; et al. Avaliação dos Fatores Ambientais no Desenvolvimento Corporal de Cordeiros Deslanados da Raça Morada Nova. Revista Brasileira de Zootecnia, v.30, n.5, p.1460-1465, 2001. FERRAZ, M.R. Manual do comportamento animal. Rio de janeiro: Rubio, 2011. 209p. GRANDINSON, K. Genetic background of maternal behaviour and its relation to offspring survival. Livestock Production Science, v.93, p.43–50, 2005. JENSEN, P. The Study of Animal Behaviour and its Applications. In: JENSEN, P. The Ethology of Domestic Animals. 1.ed. CAB International: Skara, 2002. p.3-11. LADEWIG, J.; PRICE, E. O; HART, B. L. Flehmen in male goats: Role in sexual Behaviour. Behavioral and nasal Biology, v. 30, p. 312-322, 1980. 10 LÉVY, F.; KELLER, M.; POINDRON, P. Olfactory regulation of maternal behavior in mammals. Hormones and Behavior, v.46, n.3, p.284-302, 2004. MACEDO, F.A.F.; ZUNDT, M.; MEXIA, A. A. Parâmetros reprodutivos de matrizes ovinas, rebanho base para produção de cordeiros para abate. Revista Brasileira de Reprodução Animal, v. 27, n. 2, p. 127-133, 2003. MACEDO, R.; ALVARADO, A. Efecto de la Época de Monta sobre la Productividad de Ovejas Pelibuey Bajo dos Sistemas de Alimentación en Colima, México. Archivos de Zootecnia, v. 54, p. 51-62, 2005. MELLADO, M., CARDENAS, C. RUIZ, F. Mating behavior of bucks and dors in goat operations undes range conditions. Applied Animal Behavious Science, v.67, p. 8996, 2000. NOWAK, R.; POINDRON, P. From birth colostum: early steps leading to lamb s urvival. Reprod. Nutr. Dev., v.46, p. 431-446, 2006. PACHECO, A.; QUIRINO, C.R.; OLIVEIRA, A. DE F.M. Avaliação do comportamento sexual de ovinos jovens da raça Santa Inês, com e sem experiência prévia com fêmeas. Archivos Latinoamericanos de Producción Animal, v.17, n.1, p.22-30, 2008. PARANHOS, M.J.R.; CROMBERG, V.U. Relações materno-filiais em bovinos de corte nas primeiras horas após o parto. In: PARANHOS, M.J.R.; CROMBERG, V.U. Comportamento materno em mamíferos: bases teóricas e aplicações aos ruminantes domésticos. São Paulo: sociedade brasileira de etologia, 1998, p.215-236. POINDRON, P. Mechanisms of activation of maternal behaviour in mammals. Reprod. Nutr. Dev., v.45, p. 341–351, 2005. RAINERE, C. Perfil do Comportamento materno-filial de ovinos da raça Santa Inês e sua influência no desempenho dos cordeiros ao desmame. 2008. 61f. Dissertação (Mestrado em Zootecnia) – Faculdade de Zootecnia e Engenharia de Alimentos, Universidade de São Paulo, Pirassununga. 11 RECH, C.L.S.; RECH, J.L.; FISCHER, V. et al. Temperamento e comportamento materno-filial de ovinos das raças Corriedale e Ideal e sua relação com a sobrevivência dos cordeiros. Ciência Rural, v.38, n.5, p.1388-1393, 2008. REECE, W. O. Dukes, Fisiologia dos animais domésticos. 10.ed. Guanabara Koogan: Rio de Janeiro, 2006. 909p. REKWOT, P.I., D. OGWU, E.O. OYEDIPE, AND V.O. SEKONI. The role of pheromones and biostimulation in animal reproduction. Animal Reproduction Science, v.65, p.157-170, 2001. RUTTER, S.M. Behaviour of Sheep and Goats In: Jensen, P. The Ethology of Domestic Animals. 1.ed. CAB International: Skara, 2002. p.145-158. SANTOS, A.D.F.; TORRES, C.A.A.; FONSECA, J.F. et al. Parâmetros reprodutivos de bodes submetidos ao manejo de fotoperíodo artificial. Revista Brasileira de Zootecnia, v.35, n.5, p.1926-1933, 2006. SCHMIDEK, A. Habilidade Materna e Aspectos Relacionados à Sobrevivência de Bezerros: Valores Ótimos nem Sempre são Valores Extremos. ABCZ, Uberaba, n.21, p.72-75, 2004. SILVA, M. G. B. da.; COSTA, C.; MEIRELLES, P. R. de L. et al. Comportamento materno-filial de ovinos. In: ENCONTRO DE ZOOTECNIA, 7., 2010a, Dracena. Anais... Dracena: Simpósio de Ciências da Unesp, 2010a. p.3. SILVA, R.S.M.; SOUSA, D.L.; HAGAS, G.R. et al. Avaliação do comportamento materno-filial de ovinos deslanados Morada Nova durante o parto. In: CONGRESSO NORDESTINO DE PRODUÇÃO ANIMAL., 6., 2010b, Mosoró. Anais…Mosoró: Sociedade Nordestina de Produção Animal, 2010b. p.5. SNOWDON, C.T. O significado da pesquisa em comportamento animal. Estudos de Psicologia, v.4, n.2, p. 365-373, 1999. 12 STELLFLUG, J.N.; COCKETT, N.E.; LEWIS, G.S. The relationship between sexual behavior classifications of rams and lambs sired in a competitive breeding environment. Journal Animal Science, v.84, p.463–468, 2006. 13 Capítulo 2 COMPORTAMENTO SEXUAL DE CARNEIROS DA RAÇA MORADA NOVA 14 COMPORTAMENTO SEXUAL DE CARNEIROS DA RAÇA MORADA NOVA Resumo: Objetivou-se caracterizar o comportamento sexual de reprodutores Morada Nova nas condições de semiárido. Foram registrados todos os eventos relativos à corte, registrados em planilhas específicas durante o acasalamento como: Tempo de Reação (TR), Número de saltos por Tentativa (NST), Cortejo da Fêmea (CF), Cortejo com a Pata (CP), Vocalização (VOC), Exposição do Pênis (EXP), Cheirar a região Urogenital da Fêmea (CRUF) e Reflexo de Flehmen (RF). O delineamento experimental utilizado foi um DIC, adotando o qui-quadrado para a análise dos dados, através do procedimento GLM do SAS (1999). Os resultados das variáveis referentes aos padrões de comportamento sexual dos machos foram influenciados por características individuais. As variáveis CP, VOC, EXL variaram significativamente (P<0,05) entre os reprodutores, porém as variáveis CF, EXP, CRUF e RF não diferiram estatisticamente entre os carneiros. Isso provavelmente pode ser atribuído a pouca experiência sexual do macho, o que leva a supor que o maior tempo despendido no cortejo poderia ser atribuído a questão da experiência do macho. Portanto, concluiu-se que a maioria dos padrões do comportamento sexual de ovinos da raça Morada Nova foi apresentada durante as fases de cortejo e cópula da fêmea em estro, com as diferenças comportamentais observadas atribuídas a características individuais, não sendo observada influência do turno de monta o que pode evidenciar o valor adaptativo de ovinos da raça Morada Nova. Palavras-Chave: ovinos nativos, etologia, cortejo sexual. 15 SEXUAL BEHAVIOR OF SHEEP BREED MORADA NOVA Abstract: This study aimed to characterize the sexual behavior of breeding Morada Nova semiarid conditions. We recorded all events relating to the court, registered in specific worksheets during mating as: Reaction Time (RT), number of jumps per Attempt (NST), Procession of Female (CF), Procession with Pata (CP), Vocalization (VOC), Dick Exposure (EXP), Smell of Female Urogenital region (CRUF) and Reflex Flehmen (RF). The experimental design was a DIC, adopting the chi-square test for the analysis of data through the GLM procedure of SAS (1999). The results of the variables related to sexual behavior patterns of males were influenced by individual characteristics. The variables CP, VOC, EXL varied significantly (P <0.05) among the players, but the variables CF, EXP, CRUF and RF did not differ statistically among the sheep. This can probably be attributed to inexperienced male sexual, which leads to the supposition that the more time spent in courtship could be attributed to the issue of male experience. Therefore, it was concluded that most of the sexual behavior patterns of Morada Nova sheep was presented during the phases of courtship and copulation the female in estrus, with the observed behavioral differences attributed to individual characteristics, it did not influence the shift rides can show that the adaptive value of Morada Nova sheep. Keywords: mating, sheep, procession, ethology, experience 16 1. INTRODUÇÃO O estudo do comportamento animal vem sendo valorizado, nos últimos anos, com intuito de adequar o manejo dos rebanhos e como forma de auxiliar no aumento dos índices produtivos e reprodutivos. Em carneiros, a atividade reprodutiva pode ser afetada por vários fatores externos, incluindo sócio-sexuais, temperatura, fotoperíodo e nutrição (Maurya et al., 2010). Em regiões áridas e semiáridas para os animais nativos a reprodução é uma forma de prolongar a sobrevivência da espécie através de novos descendentes. Consequentemente, se o macho não estiver em boa condição ou até mesmo com baixo a libido, prejudicaria a reprodução da espécie. Para que haja sucesso na reprodução dos animais em condições de pastejo, os recursos existentes devem ser geridos de forma eficiente para atingir o nível ideal. Isto requer um entendimento claro do requisito tanto nutricional quanto dos vários sistemas fisiológicos concorrentes para que os animais possam externar todo seu potencial reprodutivo. Durante a estação de monta o reprodutor tem uma contribuição importante para a taxa reprodutiva e o melhoramento genético de rebanho. Por isso, é importante prestar atenção aos fatores que podem influenciar a fertilidade do macho, como a libido, capacidade de acasalamento e qualidade do sêmen (Maurya et al., 2010). A compreensão do comportamento sexual dos machos fornece informações de grande importância, pois no acasalamento o macho desencadeia comportamentos específicos nesse período, desde o cortejo da fêmea até o ato final da cobrição, acompanhada da efetiva concepção. Estes comportamentos podem ser influenciados pela idade, estado fisiológico, e a libido do macho. Durante este momento a eficiência do reprodutor se mostra como uma ferramenta importantíssima para o sucesso reprodutivo do rebanho. A seleção de machos para serem utilizados como reprodutores é extremamente importante, pois carneiros com alto desempenho sexual servirão um maior número de ovelhas em um curto período de tempo, aumentando a pressão de seleção e a disseminação do material genético desejável (Pacheco e Quirino, 2006). Objetivou-se com este trabalho caracterizar o comportamento sexual de machos da raça Morada Nova em regime semi-intensivo em condições do semiárido. 17 2. MATERIAL E MÉTODOS 2.1. Local do experimento O trabalho foi desenvolvido na Unidade de Pesquisa em Pequenos Ruminantes, da Universidade Federal da Paraíba, localizada na Microrregião do Cariri Oriental Paraibano. A região é caracterizada por apresentar uma vegetação típica classificada como caatinga hiperxerófila, clima quente e seco com distribuição irregular de chuvas em curtos períodos, podendo apresentar uma estação seca prolongada, com precipitação média de 450 mm anuais e temperaturas médias anuais em torno de 25º C (Bacia Escola UFCG). 2.2. Animais Foram utilizados quatro reprodutores da raça Morada Nova, numerados de um a quatro, com média de escore corporal entre 3,5 e 4,0, sendo todos jovens, com idade em torno de dois anos. Foram utilizadas sessenta e nove fêmeas da mesma raça, com idades que variam entre quatro e treze anos, todas multíparas. 2.3 Condições climáticas Os acasalamentos foram realizados em duas estações de monta, nos turnos da manhã e tarde durante os meses de janeiro, fevereiro, março e abril, período seco, apresentando um índice pluviométrico médio de 75,6 mm (Bacia Escola UFCG). No Quadro 1 estão apresentadas as médias de temperatura máxima (Tmáx), temperatura mínima (Tmín) e umidade relativa (UR) nos meses em que se realizaram as montas. As fêmeas foram suplementadas num período de cinco semanas (duas semanas antes do início dos acasalamentos e três semanas após) e os machos foram mantidos confinados em baias individuais durante todo o período experimental recebendo feno de sorgo à vontade no cocho e uma suplementação concentrada. As ovelhas receberam suplementação formulada de acordo o NRC (1985), apenas à tarde após retornarem do 18 pastejo na caatinga. Durante esse período foi utilizado também o efeito macho através do contato dos carneiros com as fêmeas duas vezes ao dia, ou seja, antes e depois delas retornarem do pastejo (6h30 h e 17h). Quadro 1. Média de temperatura máxima, mínima e umidade relativa dos meses experimentais na fase de acasalamento Meses Manhã Tarde UR Tmáx Tmín Tmáx Tmín Janeiro 27,82 21,54 31,71 23,60 74,01 Fevereiro 26,81 21,54 31,82 25,57 75,01 Março 26,77 21,84 34,38 25,42 64,77 Abril 27,38 21,06 32,11 27,55 S/L Média 27,19 21,49 32,50 25,53 71,26 S/L= sem leitura 2.4 Estação de Monta Foram realizadas duas estações de monta em períodos distintos, com duração de 45 dias cada e com um intervalo de quinze dias entre as duas estações. Determinouse um escore corporal mínimo entre 2,5 e 3,0 para entrada na estação de monta. Quarenta ovelhas foram destinadas para a primeira fase, no período janeiro-fevereiro, denominada Estação A. As demais vinte e nove fêmeas foram direcionadas a um tratamento alimentar para alcançar o escore corporal mínimo, compondo a Estação B, realizada no período de março a abril. As treze fêmeas que repetiram o cio na Estação A foram incorporadas na Estação B. O lote de fêmeas foi dividido em grupos ao acaso para facilitar o trabalho do rufião que permanecia durante um período de 5 minutos para cada grupo com intervalo de 2 minutos entre os grupos. À medida que as fêmeas eram identificadas pelo rufião, as mesmas eram expostas aos reprodutores de acordo com a ordem de identificação no grupo. Após a detecção do estro, as fêmeas eram conduzidas aos reprodutores, de uma em uma, para cada carneiro, sequencialmente do 1 ao 4. Os animais eram identificados pela numeração dos colares e tatuagem na região dorso lombar. Foram utilizados colares com cores diferentes para identificação 19 da condição reprodutiva das fêmeas. Quando apresentava cio era colocado um colar verde e quando cobertas e não aceitavam mais a monta pelo macho era colocado um colar branco. O rebanho foi devidamente vacinado contra clostridiose e desverminados de acordo com o calendário Zootécnico da estação experimental. Os reprodutores também foram pesados e avaliados o escore, quinzenalmente para acompanhamento do peso e da condição corporal. 2.5 Comportamento Sexual Para a caracterização do comportamento sexual foi utilizado o método de observação direta, com amostragem tipo animal focal e avaliação em forma de frequência. Os padrões comportamentais dos reprodutores foram observados e anotados e as avaliações realizadas durante o turno da manhã e da tarde após a identificação do cio pelo rufião. As anotações foram realizadas por observadores treinados e posicionados em locais estratégicos no curral de acasalamento. Foi avaliado o tempo de reação (TR), período de tempo de liberação do carneiro para a monta até a primeira ejaculação, e o número de saltos por tentativa (NST) durante o tempo de reação, sendo registrado em cronômetro. Além disso, foram observados os padrões de comportamento como: cortejo da fêmea (CF); cortejo com a pata (CP); vocalização (VOC); exposição de língua (EXL); cheirar a região urogenital da fêmea (CRUF); exposição do pênis (EXP) e reflexo de flehmen (RF) segundo a metodologia de Santos et al. (2006). Os reprodutores realizaram uma monta a cada 12 horas, sendo estabelecido um intervalo de 40 minutos entre duas montas consecutivas num mesmo turno, garantindo um período de restabelecimento do macho sem prejudicar o processo da espermatogênese. Todos os comportamentos foram registrados em planilhas, seguindo um etograma de trabalho específico para avaliação do comportamento sexual dos reprodutores. 20 2.5.1 Etograma de trabalho para as avaliações comportamentais 1- Tempo de Reação: Período de tempo de liberação do carneiro para a monta até a primeira ejaculação; 2- Números de saltos por tentativa: Número de vezes que o animal monta na fêmea até executar a monta completa com ejaculado; 3- Cortejo da fêmea: Perseguir a fêmea insistentemente durante o cortejo; 4- Cortejo com a pata: cortejar a fêmea com os membros anteriores; 5- Exposição da Língua: Expor a língua durante a corte da fêmea; 6- Exposição do pênis: Expor o pênis como forma de excitação antecedendo a cópula; 7- Vocalização: emissão de sons em tons graves durante o cortejo; 8- Cheirar a região urogenital da fêmea: Cheirar e/ou lamber a região urogenital reconhecendo os ferormônios que a fêmea libera em estro; 9- Reflexo de Flehmen: O macho cheira a urina da fêmea e então levanta a cabeça, com os lábios torcidos. 2.6 Análises Estatísticas O delineamento utilizado foi o inteiramente casualizado (DIC) e a análise estatística adotada foi o teste do Qui-quadrado. Os índices reprodutivos foram expressos de forma descritiva, através de porcentagem. Os dados de frequência das avaliações comportamentais foram analisados utilizando modelos lineares generalizados considerando uma distribuição binomial negativa a 5% de probabilidade, através do procedimento GLM do programa estatístico SAS (1999). 21 3. RESULTADOS E DISCUSSÃO 3.1 Indicadores reprodutivos Os resultados dos serviços realizados pelos reprodutores nas duas estações, com respectivas taxas de prenhez total e número de fêmeas expostas estão apresentados na Tabela 1. Das quarenta fêmeas expostas na estação A, vinte e cinco emprenharam, uma abortou, duas não apresentaram cio e treze não emprenharam. Com estas últimas sendo expostas na estação seguinte. Durante essa primeira estações de monta foram observados problemas reprodutivos como hidrometria e cistos no útero detectados por meio da ultrassonografia transretal nas ovelhas que obtiveram diagnóstico negativo de prenhez. Na estação B, foram expostas quarenta e três fêmeas, das quais treze foram remanejadas da estação A, porque repetiram o cio. Apenas trinta e três ovelhas ficaram prenhas. Se observados apenas o número de fêmeas pertencentes à estação B, contabilizaríamos vinte e nove ovelhas, das quais vinte e sete emprenharam, evidenciando uma taxa de prenhez de 93,1% para esse grupo. As ovelhas expostas na Estação B apresentaram uma maior taxa de prenhez em relação aos da Estação A. Este grupo era composto em maior parte por fêmeas que se encontravam com uma condição corporal abaixo do recomendado para início da atividade reprodutiva, com vinte e sete das fêmeas prenhas, representando um percentual de 93,1%. Quando se incorporou as ovelhas da Estação A esse percentual diminui para 76,74% (Tabela 1), mas mesmo assim representa um valor considerável para fêmeas em regime de caantiga. Tabela 1. Percentagem de prenhez entre estações de monta em ovinos Morada Nova Fêmeas Expostas Prenhez total % Total de prenhez Estação A 40 25 62,50 Estação B 43 33 76,74 Resultados obtidos por análise descritiva. A maior eficiência das ovelhas da estação B poderia ser explicada por uma melhor resposta à suplementação alimentar, com consequente aumento na taxa de 22 ovulação. Esse raciocínio tem como premissa o fato de que as referências para classificação do escore corporal são baseadas em trabalhos com raças especializadas. Para alguns autores, as ovelhas no momento da cobertura devem estar com escore corporal acima de 2 para alcançar uma ótima eficiência reprodutiva resultando na prenhez (Cezar e Sousa, 2006). Nesse caso, é possível que o verdadeiro padrão para raças nativas, com a Morada Nova, esteja naturalmente abaixo desse escore. Assim, as ovelhas da Estação A estariam, na realidade, acima da condição corporal desejável, ou seja, com deposição de gordura no aparelho reprodutor acarretando em uma menor eficiência reprodutiva. Na primeira estação de monta algumas fêmeas apresentaram retorno ao estro em maior número em relação à segunda estação. Isso pode ser atribuído à condição corporal das ovelhas, pois fêmeas mais gordas tendem a depositar gordura entre as gônadas (ovários), podendo apresentar problemas reprodutivos e consequentemente prejudicar a liberação do óvulo para posterior fertilização. Para Cezar & Sousa (2006) a eficiência reprodutiva à monta responde ao consumo de energia ou às mudanças de escore corporal, mostrando que se o animal chegar à monta com um escore abaixo ou acima a perda ou o ganho não vai piorar nem melhorar a baixa eficiência reprodutiva das matrizes, porém, a melhoria do escore resulta em incremento da eficiência, enquanto a perda diminuirá o desempenho reprodutivo das matrizes. A condição corporal alcançada pela ovelha durante a monta tem uma ação reguladora sobre a eficiência reprodutiva, afetando a taxa de fertilidade e, principalmente, o índice de prolificidade das matrizes. Selaive-Vilarroel & Fernandes (1994) observaram em ovelhas Morada Nova com escore corporal entre 1 e 3, taxa de parição de 75 e 91% , respectivamente. A tabela 2 apresenta os valores referentes às fêmeas expostas por reprodutor e seus respectivos percentuais de prenhez em cada estação. Na Estação A foram apresentadas menores taxas do que na B, exceto para o macho 3. Esse reprodutor apresentou uma maior taxa do que os demais reprodutores, que obtiveram índices similares. As taxas de prenhez segundo o reprodutor mostraram que o valor individual prevaleceu entre os animais, pois todos os machos receberam manejo nutricional, sanitário e números de fêmeas expostas semelhantes. 23 Tabela 2. Número de fêmeas expostas, com repetição de cio, com prenhez positiva por reprodutor e por estação de monta. Reprodutor Estação 1 2 3 4 Fêmeas Repetição Expostas de cio Prenhez % Positiva Prenhez A 12 5 6 50,00 B 12 4 8 66,66 A 12 4 6 50,00 B 13 6 7 53,84 A 10 2 8 80,00 B 13 3 10 76,92 A 10 4 5 50,00 B 13 5 8 61,54 % média de prenhez/macho 58,33 51,92 78,46 55,77 Resultados obtidos por análise descritiva. O maior percentual médio de prenhez apresentado pelo carneiro 3 pode estar associado a um maior vigor espermático somado a uma mais rápida recuperação, que pode ter refletido num melhor desempenho. Estes resultados demonstram a importância do valor individual do reprodutor, o que pode ser um dos pontos de grande relevância na escolha de um macho, visando uma alta eficiência reprodutiva. Em virtude da impossibilidade da realização do exame andrológico, estes resultados não puderam, no entanto, ser associados ao vigor espermático. 3.2 Variáveis comportamentais dos reprodutores Morada Nova Os resultados apresentados para a variável Tempo de Reação (TR) diferiram de forma significativa (P<0,05) entre os reprodutores. Os carneiros 2 e 3 apresentaram maior TR em relação aos machos 1 e 4, resultados possivelmente atribuídos às diferenças individuais. O maior tempo que o reprodutor 3 levou para realizar a monta pode ter sido um dos fatores responsáveis pela maior taxa de prenhez. Novamente, essa diferença pode ser conferida ao comportamento individual relativa à capacidade de aprendizagem de cada macho o que influenciará na eficiência durante o acasalamento, tornando-se um fator de escolha de reprodutores que se mostrem eficientes no rebanho (Tabela 3). 24 Tabela 3. Média do Tempo de Reação (TR) em segundos e Freqüência do Número de Saltos por tentativa (NST) de ovinos da raça Morada Nova Reprodutores Tempo de Reação N° Saltos/Tentativa N° observações 1 26,95 b 1,48 b 45 2 39,73 a b 38 3 39,07 a 2,95 a 40 4 22,15 b 1,64 b 39 1,92 Médias seguidas de letras diferentes na coluna diferem estatisticamente, ao nível de 5% de probabilidade, pelo teste do Qui-quadrado Diferentemente do que se espera de um animal jovem que, pela sua exacerbada libido, realiza a cópula de forma mais rápida, os reprodutores 2 e 3 levaram mais tempo para realizar a monta. Isso pode ter estimulado mais as fêmeas com aumento na liberação de LH, refletindo diretamente nas taxas de ovulação o que repercutiu num maior índices de prenhez apresentado pelo carneiro 3 em comparação aos outros carneiros. O fato do reprodutor 1 ter tido uma experiência sexual anterior em relação aos outros, não se mostrou como um fator de diferenciação em relação aos machos inexperientes. Se essa diferença no TR fosse atribuída à experiência, apenas o macho 1 apresentaria menor TR, mas o que não se mostrou relevante para essa explicação entre os machos, pois percebe-se que o fator mais importante nesse padrão será o nível de estimulação ocasionado na fêmea, que influenciará na taxa de prenhez do rebanho. Pacheco et al. (2008) trabalhando com carneiros jovens com contato prévio e sem contato prévio, observaram que o tempo de reação entre os animais não variaram, apresentando um TR médio de 26,5 segundos, sendo semelhante aos nossos resultados, apenas que neste estudo o autor mensurou o TR a partir do tempo que cada macho levava para manifestar qualquer tipo de evento sexual. O macho 3 apresentou um NST superior aos demais (P<0,05). A explicação para esse resultado pode ser atribuída ao alto TR e às diferenças individuais. Deve ser destacado também que o macho 3 apresentou um desempenho superior no que se refere à taxa de prenhez das fêmeas por ele cobertas em ambas as estações. Como este reprodutor levou mais tempo em realizar a monta, isso pode ter promovido um maior estimulo no cortejo dando respaldo aos índices reprodutivos. 25 A característica individual no comportamento sexual influencia a eficiência do macho e pode proporcionar diferenças no número de descendentes por reprodutor. No caso do macho 3, a resposta ao aumento no número de saltos pode ter refletido numa possível indução à liberação de hormônios estrais promovidos possivelmente por esse maior contato ou cortejo no acasalamento. Pimentel Gómez et al. (2005) relataram, em seus trabalhos com diferentes genótipos de ovinos, que a influência da idade e as características como número de serviços e número de montas tendem a melhorar com o aumento da idade dos reprodutores. Os comportamentos apresentados durante a fase preparatória da cópula como cortejar a fêmea com a pata (CP), vocalização (VOC), exposição de língua (EXL) variaram de forma significativa (P<0,05) entre os reprodutores, enquanto que comportamentos como cortejo da fêmea (CF), exposição de pênis (EXP), cheirar a região urogenital da fêmea (CRUF) e reflexo de flehmen (RF), denominados eventos pré-copulatórios, não diferiram estatisticamente (Tabela 4). Tabela 4. Médias das frequências dos padrões de Comportamentos sexuais: cortejo da fêmea (CF); cortejo com a pata (CP); vocalização (VOC); exposição de pênis (EXP); cheirar a região urogenital da fêmea (CRUF) e reflexo de flehmen (RF) apresentados por machos ovinos da raça Morada Nova Reprodutores Padrões Comportamentais CF CP VOC EXL EXP CRUF RF 1 1,00 1,23 b 1,71 b 1,60 b 1,80 1,48 1,09 2 1,50 1,19 b 2,85 a 2,27 ab 1,71 1,75 1,20 3 1,40 2,89 a 3,16 a 2,75 a 1,60 1,41 1,25 1,50 b b b 1,28 1,29 1,00 4 1,60 1,60 1,60 Médias seguidas de letras diferentes na coluna diferem estatisticamente, ao nível de 5% de probabilidade, pelo teste do Qui-quadrado do valor da média estimada. O macho 3 apresentou 2,89 vezes mais o CP do que os outros animais. Este resultado pode ter ocorrido em função do maior TR, o que mostra que os comportamentos iniciais do cortejo da fêmea em estro podem ser importantes na estimulação. Possivelmente, o comportamento apresentado pelo macho 3 respalda a maior taxa de prenhez das fêmeas servidas por ele em ambas estações de monta. O 26 reprodutor 2 mesmo com um maior TR, apresentou uma menor frequência deste comportamento, o que possa ser uma das explicações da menor taxa de prenhez. Kridli e Al-Yacoub (2006) não observaram efeito significativo quanto aos comportamentos cortejo com a pata, cheirar a região urogenital da fêmea, tentativa de monta e monta quando o grupo era constituído apenas de machos. Porém quando os machos foram inseridos a grupos misturados com fêmeas mostraram mais padrões de comportamentos durante o acasalamento. Os reprodutores 2 e 3 tenderam a vocalizar mais do que os outros machos, fato também associado ao maior TR. Segundo Pacheco et al. (2008), avaliando o comportamento sexual de ovinos Santa Inês, a vocalização excessiva pode ser uma atitude provavelmente relacionada com a ansiedade gerada pela separação dos companheiros pertencentes ao seu grupo de convívio. Mas, neste estudo, este padrão de comportamento possivelmente estaria relacionado com a comunicação entre os animais durante o cortejo. Ao observar a EXL entre os reprodutores, os resultados mostram que o macho 3 apresentou em maior número este padrão de comportamento da fase pré-copulatória, mas que não diferiu do macho 2 e este não diferiu dos demais. Este tipo de comportamento está associado com a captura dos ferormônios liberados pelas fêmeas em estro o que pode também respaldar a taxa de prenhez apresentada pelo carneiro 3. Os outros padrões envolvidos na fase preparatória ou pré-copulatória como CF, EXP, CRUF e RF não apresentaram diferença significativa, possivelmente por se tratarem de animais jovens e estes tipos de comportamentos serem apresentados com maior frequência em animais adultos. Madella-Oliveira et al. (2009) observaram que entre os padrões comportamentais realizados pelos carneiros os que apresentaram maiores frequências foram: perseguir (16,1%), lamber (18,5%) e cheirar a região genital das fêmeas em estro (11,6%). Essas altas frequências evidenciam que mesmo sem experiência prévia, o comportamento sexual dos carneiros durante a fase de monta pode ser expresso de forma satisfatória. Para Hafez (2004), o nível de experiência sexual não altera os padrões motores de conduta do cortejo, mas age principalmente sobre a eficiência do acasalamento. De acordo com Bench et al. (2001), o contato prévio dos carneiros com as fêmeas pode fornecer prática na execução da cópula podendo auxiliar na desinibição de machos inexperientes. Entretanto, o início do desenvolvimento da orientação sexual 27 também pode surgir pelo convívio com outros machos. Para Kridli e Al-Yacoub (2006), a criação de machos separadamente ou junto das fêmeas, antes da entrada na puberdade, parece não influenciar no desempenho sexual. Os carneiros 2 e 3, sem experiência prévia, apresentaram padrões de comportamentos semelhantes, enquanto que o macho 1, com experiência, e o 4, sem experiência prévia, demonstraram resultados similares para a maioria dos comportamentos avaliados. Stellflug e Lewis (2007) relataram que a frequência e duração de cada comportamento dependem de cada reprodutor e de sua experiência sexual prévia, podendo servir como critério de avaliação, classificação e seleção de bons reprodutores. Os resultados mostraram que o fator experiência não se apresentou como fator de diferenciação entre os machos para expressar os comportamentos necessários de estímulo das fêmeas em estro. Godfrey et al. (1998) avaliaram o comportamento sexual de carneiros que não tiveram contato físico nem visual com fêmeas desde o desmame e concluíram que a falta de contato com fêmeas parece não influenciar no desencadeamento dos comportamentos de cortejo. Apenas com um contato prévio com fêmeas em cio os machos são estimulados e adquirem experiência para a realização do comportamento e reconhecimento do estado fisiológico da fêmea (Pacheco et al., 2008). Os comportamentos CF, EXP, CRUF e RF podem ser apresentados durante o acasalamento, porém em menor frequência do que outros, mas que se mostram muito importantes durante a fase de cortejo, podendo desencadear processos como liberação de hormônios sexuais envolvidos na cópula. Stellflug e Lewis (2007) observaram que os cortejos aumentaram siginificativamente (P <0,01) para machos em contato com fêmeas desde cordeiros, comparado com os que não tinham contato, sendo um importante indicativo para realizações de teste de libido. Na Tabela 5 estão apresentados os resultados relacionados com o turno. Não houve diferença estatística entre TR e NST nos dois turnos, demonstrando que para ovinos desta raça durante a época chuvosa do ano o turno de acasalamento não influenciou na eficiência da monta. Esses resultados diferem dos apresentados por Alves et al. (2006), que relatam diferença significativa entre os períodos de monta (manhã e tarde) observando que os animais, tanto jovens como adultos, tiveram TR menor pela manhã (das 6 às 12 h) do que à tarde (das 12 às 18 h). Atribuíram os resultados, primeiramente, ao cansaço 28 natural advindo da exposição sequencial dos reprodutores à monta e, em segundo lugar, ao fator climático, uma vez que durante o período experimental as temperaturas no turno da tarde foram superiores àquelas da manhã. Tabela 5. Média do Tempo de Reação (TR), em segundos, e média da Freqüência do Número de Saltos por Tentativa (NST) dos reprodutores em relação ao turno de acasalamento. Qui-Quadrado Turno N° observações TR NST Manhã 31,27 1,85 80 Tarde 32,41 2,12 82 Efeito não significativo (P>0,05) pelo Qui-Quadrado Nos períodos de acasalamento as temperaturas médias máximas (Tmáx) e mínimas (Tmín) no turno da manhã (das 6 as 7:30 h) foram 27,19°C e 21,49°C, respectivamente, e no turno da tarde (das 16:30 as 17 h) foram 32,50°C e 25,53°C apresentaram dentro da zona de conforto térmico que varia de 15 a 30°C de ovinos adultos que conforme Baêta e Souza (1997), sendo a zona crítica acima de 35°C. Por ter sido realizada na época considerada chuvosa os dados relacionados com a temperatura se encontram relativamente adequadas para a espécie, além de que a raça apresenta um excelente valor adaptativo o que não afetou o seu desempenho durante a monta em dias com temperaturas mais altas. Nas condições de temperatura elevada os carneiros devem expressem níveis adequados de libido e fertilidade para o sucesso reprodutivo de um grande número de fêmeas (Godfrey et al., 1998). Não houve diferenças significativas (P<0,05) entre os turnos com relação às variáveis comportamentais CF, CP, VOC, EXL, EXP, CRUF e RF (Tabela 6). Os padrões de comportamentos apresentados foram todos avaliados dentro de um período que compreendeu, da soltura dos machos até ele realizar a monta completa com ejaculado (TR). Os resultados apresentados mostraram que os ovinos da raça Morada Nova, em regime de monta natural controlada, apresentam todos os comportamentos necessários para estímulo da fêmea em estro. Os reprodutores nos dois turnos realizaram no máximo duas montas por turno, para não haver o desgaste físico dos mesmos, com 29 isso favorecendo uma renovação na produção dos espermatozoides. Também foi feito uma distribuição das fêmeas semelhante como forma de fornecer as mesmas condições dos machos demonstrarem seus padrões comportamentais diante das ovelhas em estro. Tabela 6. Médias das freqüências dos padrões de Comportamentos sexuais: Cortejo com a Pata (CP); Vocalização (VOC); Exposição da Língua (EXL); Exposição de Pênis (EXP); Cheirar a Região Urogenital da Fêmea (CRUF) e Reflexo de Flehmen (RF) apresentados em ovinos da raça Morada Nova em relação ao turno de acasalamento Padrões Comportamentais Turno CF CP VOC EXL EXP CRUF RF Manhã 1,42 1,74 2,32 2,12 1,54 1,47 1,06 Tarde 1,33 2,20 2,47 2,16 1,67 1,52 1,20 Efeito não significativo (P>0,05) pelo Qui-Quadrado O comportamento CF foi pouco observado nos machos isso possa ser explicado pelo motivo das ovelhas quando expostas aos reprodutores já se encontravam em estro, o que não necessitaria de um investimento do macho na insistência em perseguir lá. Com isso, refletindo num menor desgaste físico dos reprodutores, consequentemente auxiliando numa monta mais eficiente. Enquanto que o CP que também é considerado como um dos comportamentos iniciais da fase pré-copulatória, não foi influenciado pelo turno de monta, estando mais associada à característica individual dos machos do que efeito dentro do turno de acasalamento. A VOC em relação aos outros padrões de comportamento se apresentou com uma média de frequência um pouco maior, isso possa está relacionada à idade dos animais e possivelmente a algum tipo de comunicação entre eles durante o acasalamento. O que demonstra que mesmo se tratando de animais jovens, eles também desencadeiam estímulos auditivos no momento da cópula. Para a EXL junto com a VOC também apresentou com uma média de frequência relativamente mais alta do que os outros comportamentos, associada principalmente a detecção do estro através dos feromonios liberados pelas fêmeas, sendo este comportamento associado com a confirmação do cio. Na EXP não apresentou diferença estatística, obtendo uma frequência semelhante entre os dois turnos, mostrado principalmente na fase de excitação diante 30 uma fêmea em cio, antecedendo a ereção na preparação a monta. De acordo com Santos et al. (2006), existe uma variação na frequência das manifestações das características de cortejo (exposição da língua e exposição do pênis) entre animais jovens e adultos, tendo maior intensidade nos animais mais velhos, devido à maior experiência sexual. Neste trabalho os resultados para os reprodutores foram semelhantes por se tratarem de animais de mesma faixa etária com relação aos jovens, com apenas o carneiro 1 apresentando uma experiência prévia de monta o que não se apresentou como fator de diferença na demonstração dos padrões de comportamento sexual na fase de acasalamento. O ato de cheirar a região urogenital da fêmea (CRUF) se mostra como um dos padrões mais importantes que antecede a monta, aumentando a excitação do macho na detecção da fêmea em cio ao cheirar a urina fresca da ovelha, odores dos feromonios liberados pelos hormônios envolvidos no estro como FSH, LH e estrógeno. Com relação ao reflexo de Flehmen, verificou-se uma frequência muito baixa, independente do turno. Após cheirar e lamber a urina, o carneiro realiza um movimento bem característico dos ruminantes e equídeos, chamado de reflexo de Flehmen, levantando a cabeça e curvando o lábio superior, estando associado como uma investigação olfatória do estro da fêmea, sendo controlada por ação da testosterona. Alves et al. (2006) atribuem esta característica aos animais mais velhos e experientes que a utilizam como forma de estímulo sexual antes da cópula. Os resultados apresentados mostram que, independente do turno, todos os padrões de comportamentos foram apresentados, o que pode caracterizar a adaptação desta raça na região semiárida nordestina. 31 4. CONCLUSÃO Concluiu-se que a maioria dos padrões do comportamento sexual de ovinos da raça Morada Nova foi apresentada durante as fases de cortejo e cópula da fêmea em estro, com as diferenças comportamentais observadas atribuídas a características individuais. 32 5. Referências Bibliográficas ALVES, D.; PEÑA-ALFARO, C.E.; LEITE, S.V.F. et al. Comportamento sexual de caprinos da raça bôer submetidos a regime intensivo de coleta de sêmen. Agropecuária Científica no Semiárido, Patos, v.2, n.1, p. 55-61, 2006. BAÊTA, F.C.; SOUZA, C. Ambiência em edificações rurais: conforto animal. Viçosa, Editora: UFV, 246 p, 1997. BENCH, C.J; PRICE, E.O; DALLY, M.R; BORGWARDT, R. E. Artificial selection of rams for sexual performance and its effect on the sexual behavior and fecundity of male and female progeny. Applied Animal Behaviour Science, v.72, p.41-50, 2001. CEZAR, M. F.; SOUSA, W. H. DE. Avaliação e utilização da condição corporal como ferramenta de melhoria da reprodução e produção de ovinos e caprinos de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 43, 2006, João Pessoa, PB. Anais... João Pessoa: SBZ, 2006, p. 649-678. Godfrey, R. W., J. R. Collins, and M. L. Gray. Evaluation of sexual behavior of hair sheep rams in a tropical environment. Journal Animal Science, v.76, p.714717,1998. HAFEZ, B.; HAFEZ, E.S.E.; Reprodução Animal. 7ª ed. Barueri, SP Editora: Manole, Cap.19, p.293-300, 2004 KRIDLI, R.T; AL-YACOUB, A.N. Sexual performance of Awassi ram lambs reared in different sex composition groups. Applied Animal Behaviour Science, v.96, p.261-267, 2006. MADELLA-OLIVEIRA, A. F.; MARCIAL, M. P. S.; JESUS, T. G.; BARROS, C. J. M.; NEVES, A. C. Comportamento Social E Sexual De Carneiros Jovens Em Sistema Intensivo. In: CONGRESSO NACIONAL DE ZOOTECNIA, 19, 2009, Águas de Lindoia, SP. Anais... Águas de Lindoia, ZOOTEC, 2009, p.4. 33 MAURYA, V.P; SEJIAN, V.; KUMAR, D.; NAQVI, S. M. K. Effect of induced body condition score differences on sexual behavior, scrotal measurements, semen attributes and endocrine responses in Malpura rams under hot semi-arid environment. Journal of Animal Physiology and Animal Nutrition, v.10, n., p.110, 2010. NATIONAL RESEACH COUNCIL. Nutrients Requirements of sheep. 6 ed. Washington, DC, USA, 1985. 99p. PACHECO, A.; QUIRINO, C.R.; OLIVEIRA, A. DE F.M. Avaliação do comportamento sexual de ovinos jovens da raça Santa Inês, com e sem experiência prévia com fêmeas. Archivos Latinoamericanos de Producción Animal, v.17, n.1, p.22-30, 2008. PIMENTEL GÓMEZ, J.; GARZA, R.P.; MARTÍNEZ, L.Z. et al. Caracterización Reproductiva integral Del Morueco en El ganado lanar de Chiapas. Archivos de Zootecnia, v.54, n.206-207, p.557-564, 2005. SANTOS, A.D.F.; TORRES, C.A.A.; FONSECA, J.F. et al. Parâmetros reprodutivos de bodes submetidos ao manejo de fotoperíodo artificial. Revista Brasileira de Zootecnia, v. 35, n.5, p. 1926-1933, 2006. SAS, Statistical Analysis Systems Institute, User`s guide, version 8. SAS institute Inc., North Carolina, 1999, 295p. SELAIVE-VILARROEL, A.B.; FERNANDES, A.A.O. Avaliação da condição corporal ao acasalamento e sua influência no desempenho reprodutivo de ovelhas Morada Nova no Semi-árido do Ceará. Revista Científica de Produção Animal, v. 4, n.1, p.914, 1994. STELLFLUG, J.N; LEWIS, G.S. Effect of early and late exposure to estrual ewes on ram sexual performance classifications. Animal Reproductive Science. v.97, p.295-302, 2007. 34 ANEXOS 35 36 Fêmea no curral de monta Farejando os ferormonios 37 Fêmea e macho durante o corte Imobilização da Fêmea Monta Completa Macho cheirando a CRUF da fêmea Salto sobre a fêmea Monta completa evidenciando o arranque Curral onde foram realizados os acasalamentos 38 Capítulo 3 CARACTERIZAÇÃO DO COMPORTAMENTO MATERNO-FILIAL NO PÓS-PARTO DE OVELHAS MORADA NOVA 39 CARACTERIZAÇÃO DO COMPORTAMENTO MATERNO-FILIAL NO PÓS-PARTO DE OVELHAS MORADA NOVA Resumo: O comportamento materno-filial em ovinos tem uma importância fundamental na preservação da espécie, agilidade e sobrevivência da cria até o desmame, tendo reflexo direto na eficiência econômica do sistema de produção. Objetivou-se caracterizar a relação materno-filial de ovelhas da raça Morada Nova após o parto. Para tais observações comportamentais foram acompanhadas 39 fêmeas adultas, multíparas, distribuídas em duas estações de monta consecutivas, com intervalo de quinze dias entre elas. As observações das variáveis foram feitas iniciando-se a partir da localização de uma fêmea com características de pré-parto. Foram registrados todos os eventos comportamentais realizados entre mãe e cria, num período de 2 horas. A análise estatística adotada foi o teste do Qui-quadrado considerando uma distribuição binomial negativa a 5% de probabilidade. Os resultados obtidos demonstraram que as ovelhas da raça Morada Nova expressam todos os comportamentos iniciais necessários para garantir a sobrevivência da sua cria. Além disso, os resultados observados mostraram que as crias se apresentaram mais ágeis quanto ao tempo de ficar em pé (TEP) e realizar a primeira mamada (TPM) independente do lote. Pode-se obsevar que para as outras variáveis comportamentos relacionadas com a relação materno-filial como: limpeza do cordeiro (LCO), ingerindo membranas (IME), facilitando a mamada (FMA) e dificultando a mamada (DMA). E nas crias os padrões observados foram: procurar úbere (PUB), tentar mamar (TMA) e mamando (MAM) mostram que independente da estação de parição ou condição corporal da fêmea antes do parto não prejudica na apresentação dos estímulos iniciais da mãe com sua cria. Concluiu-se que os ovinos desta raça apresentam uma excelente habilidade materna para as condições do semiárido brasileiro, ambiente em que foi conduzido o trabalho. Palavras–chave: mãe-cria, prolificidade, sobrevivência. 40 CHARACTERIZATION OF CONDUCT BRANCH IN MOTHER-OF SHEEP MORADA NOVA POSTPARTUM Abstract: The maternal-filial behavior in sheep is of fundamental importance in the preservation of the species, agility and survivability of the offspring to weaning, having a direct impact on economic efficiency of the production system. This study aimed to characterize the relationship of mother-infant Morada Nova sheep after delivery. For such behavioral observations were accompanied by 39 adult females, multiparous, distributed in two consecutive breeding seasons, with two week interval between them. The electrocardiographic changes were made starting from the location of a female with characteristics of pre-birth. All were recorded behavioral events performed between mother and creates, over a period of 2 hours. The statistical analysis used was the chi-square test assuming a negative binomial distribution at 5% probability. The results showed that sheep Morada Nova express all initial behaviors needed to ensure the survival of their offspring. Moreover, the observed results showed that the pups were more agile in terms of time of standing (TEP) and perform the first feeding (TPM) independent of the lot. You can obsevar that behavior for other variables related to maternal-filial relationship as: cleaning Lamb (LCO), ingesting membranes (IME), facilitating breastfeeding (FMA) and hindering breastfeeding (DMA). And the patterns observed in the offspring were looking udder (PUB), try to breastfeed (TMA) and feeding (MAM) show that regardless of the calving season or female body condition before calving did not affect the presentation of the stimuli with his mother's initials creates. It was concluded that this breed sheep have excellent maternal ability to Brazilian semiarid conditions, the environment in which the work was conducted. Keywords: mother-offspring, prolificacy, survival. 41 1. INTRODUÇÃO As raças nativas do semiárido brasileiro, face ao desconhecimento de seu potencial produtivo mesmo no seu ambiente de origem, vêm sendo, gradativamente, substituídas por raças de origem europeia ou sul-africana. A caracterização fenotípica, fundamental para o programa de conservação, inclui, também, as características de comportamento. Dentro do estudo etológico, destaca-se a relação materno-filial, responsável pelo padrão de desenvolvimento da cria no início de sua vida e, por consequência, da taxa de mortalidade. Além disso, o conhecimento do comportamento animal, em todas as fases, tem auxiliado não somente no manejo dos animais, mas tem também contribuído para reduzir o estresse advindo da intensa exploração produtiva dos rebanhos. Na espécie ovina, como na maioria dos mamíferos, a relação materno-filial inicia-se nos primeiros dias após a concepção por meio de mecanismos hormonais que irão estabelecer o contato entre a mãe e o feto denominado de reconhecimento materno da gestação (Paranhos da Costa et al., 2007). Nas primeiras horas de vida os cordeiros nascem bastante frágeis, com pouco tecido de reserva e muito dependentes da nutrição e proteção materna. Por isso, a adequação do manejo pós-parto se mostra de suma importância, pois pode acarretar em sérios problemas de rejeição da cria o que pode ocasionar um aumento na taxa de mortalidade na propriedade. Um manejo inadequado para um determinado genótipo pode influenciar a mortalidade das crias. A sobrevivência das crias deve ser encarada principalmente como uma parceria bem sucedida entre mãe e cria durante todas as fases criticas da vida do recém-nascido. Para tanto, pesquisas vêm sendo realizadas com a raça Morada Nova no intuito de conhecer melhor seus hábitos, formas de interação e relação entre mãe-cria. Estudos anteriores já mostraram que ovelhas da raça Morada Nova mesmo recebendo níveis baixos de energia na dieta não deixaram de expressar seus cuidados, nem tampouco deixaram de rejeitar suas crias. Esse resultado pode estar amparado pela pressão de seleção natural imposta à raça, que permitiu evidenciar uma ótima habilidade materna, mesmo em condições de restrição energética (Mariz et al., 2007). Em programas da própria Embrapa Caprinos e Ovinos esta raça é considerada como raça mãe tendo como base a sua habilidade materna. No entanto, informações 42 advindas de estudos planejados sobre a relação materno-filial nessa raça ainda são limitadas, demandando uma caracterização melhor definida. Neste trabalho, objetivou-se caracterizar a relação materno-filial de ovelhas da raça Morada Nova após o parto em regime de criação semi-intensivo no semiárido. 43 2. MATERIAL E MÉTODOS 2.1. Fase pré-experimental e Experimental A pesquisa foi desenvolvida na Unidade em Pequenos Ruminantes, pertencente ao CCA/UFPB, Campus II, localizada no município de São João do Cariri-PB, na Microrregião do Cariri Oriental Paraibano, apresentando uma média anual de chuvas em torno de 450 mm anuais e temperaturas médias de 25º C (Bacia Escola, UFCG). Para a avaliação do comportamento materno-filial foram utilizados dois currais, sendo um para a maternidade com as dimensões 9,00m x 10,30m e outro curral de apoio para as fêmeas e borregos no pós- parto. Foram utilizadas 39 fêmeas prenhes da raça Morada Nova, com idades entre quatro e treze anos, todas multíparas, provenientes de duas estações de monta realizadas nos meses de janeiro a fevereiro, estação A, e de março a abril, denominada de B. Na estação A, foram utilizadas 19 fêmeas e na estação B foram utilizadas 20 fêmeas. Para o acasalamento foram utilizados quatro carneiros e um rufião para detectar as fêmeas em estro. Durante a fase pré-experimental as ovelhas foram pesadas com uso de uma balança digital e avaliadas quanto ao escore corporal (ECC) a cada 28 dias. A avaliação do escore foi feito pelo método de apalpação das vértebras lombares na região dos processos transversais e processos espinhais, atribuindo um valor de 1 a 5 (sendo 1 os animais muito magros e 5 os animais obesos), com um único avaliador. Depois de 30 dias após o acasalamento realizou-se o diagnóstico de gestação por ultrassonografia trans retal para cada grupo de fêmeas e consequentemente foi estimado o período de parição de todas as ovelhas dentro de cada lote. A previsão do parto foi contabilizada a partir da primeira cópula da fêmea com o macho e somado a cento e cinquenta dias de gestação. A Fase experimental iniciou-se nos meses de junho, julho e agosto, onde se concentraram os partos, sendo este período considerado de transição entre o chuvoso e início do período seco, apresentando uma média pluviométrica de 78,6 mm e temperatura variando de 21,9 a 28,2 ºC de acordo com dados provenientes da Bacia Escola – UFCG. 44 No período experimental o manejo nutricional adotado seguiu as exigências estabelecidas pelo NRC (1985) de acordo com a fase gestacional dos animais, sendo fornecido 600g de ração concentrada/animal/dia. O rebanho foi vacinado de acordo com o calendário Zootécnico e as ovelhas foram everminadas logo após o parto e as crias 15 dias após, segundo um protocolo específico de acordo com a infestação parasitária. No período que antecedia o parto, cerca de sete dias antes da data prevista de parição as ovelhas foram transferidas para uma área reservada, num curral maternidade e permaneceram nessas instalações até o parto. Para as avaliações da relação materno-filial foram realizadas observações comportamentais de forma direta, com registro instantâneo e amostragem focal. As observações do comportamento foram iniciadas a partir da localização de uma fêmea com características de pré-parto. Logo após a expulsão completa do feto, era anotada a hora do parto e iniciados os registros dos comportamentos da fêmea em relação à cria e da cria em relação à mãe por um período de 2 horas. Os registros eram feitos durante dez minutos com um intervalo de cinco minutos entre as amostragens, seguindo a metodologia apresentada por Rainere (2008); Paranhos da Costa e Cromberg (1998), adotando-se períodos de observação de 24 horas contínuos. A avaliação da relação mãe-cria foi feita seguindo uma padronização, onde apenas a primeira cria independente do tipo de parto foi avaliada, para não gerar tendências nos resultados. Foram também caracterizados a duração e o tipo de parto (simples, duplos ou triplos) referente ao número de cordeiros nascidos por parto. O comportamento materno-filial foi caracterizado através do registro das variáveis atividade (ATM) e postura (PTM) da ovelha e do borrego. Os horários do primeiro toque da mãe no cordeiro (TTC), momento em que o cordeiro ficou em pé (TEP) e mamou pela primeira vez (TPM) foram registrados independentemente dos intervalos de amostragem. Todos os dados foram colhidos por observadores treinados e registrados em planilhas específicas, seguindo um etograma de trabalho pré-estabelecido. Nas fêmeas foram observados os padrões comportamentais: contato com o cordeiro (CCO), limpeza do cordeiro (LCO), ingerindo membranas (IME), facilitando a mamada (FMA), dificultando a mamada (DMA). Para as crias os padrões observados foram: tentando levantar (TLE), levantar (LEV), procurar úbere (PUB), tentar mamar (TMA), mamando (MAM) e os parâmetros como: sem atividade aparente (SAA) e 45 outras atividades (OAT) foram utilizados tanto para as mães como para as crias, como mostrado abaixo. Depois de transcorrido o período de duas horas de observação às fêmeas e suas crias foram levadas a um curral de apoio, onde permaneciam com suas crias até completarem um período de 15 dias para daí então serem liberadas para o pastejo. Os nascimentos foram concentrados durante o período de 13 de junho a 07 de julho de 2010, referente à estação A e no período de 30 de julho a 17 de agosto de 2010, referente à estação B. 2.2 Descrição das atividades observadas nas ovelhas 1- Contato ou toque: a ovelha cheira ou toca o cordeiro com o focinho; 2- Limpeza do cordeiro: a ovelha lambe o cordeiro; 3- Ingerindo membranas: a ovelha ingere membranas do solo, cordeiro ou do seu corpo; 4- Facilitando a mamada: a ovelha expõe o úbere para o cordeiro ou direciona o neonato; 5- Dificultando a mamada: movimentos para frente, para trás ou em círculos quando o cordeiro aproxima-se do úbere; 6- Afastar-se da cria: caminhar para outra direção, interessada por outro estímulo; 7- Agressão contra a cria: a ovelha afasta ou agride a cria com movimentos da cabeça; 8- Sem atividade aparente: a ovelha não demonstra nenhuma atividade; 9- Estimulando a cria: a ovelha toca o neonato com o focinho ou patas para fazê-lo levantar; 10- Outras atividades: a ovelha realiza atividades diferentes das avaliadas; 11- Vocalização: emissão de sons em tons graves; 12- Rejeição da cria: quando a fêmea rejeita sua cria dando cabeças ou afastandose. 46 2.3 Descrição das atividades observadas nos cordeiros 1- Tentar levantar: cordeiro mantém pelo menos uma parte do corpo fora do solo; 2- Levantar: cordeiro sobre as quatro patas por no mínimo 5 segundos; 3- Procurar úbere: cordeiro explorando o corpo da mãe; 4- Tentar mamar: cordeiro explorando o úbere da mãe por no mínimo 5 segundos; 5- Sem atividade aparente: o cordeiro não demonstra nenhuma atividade; 6- Outras atividades: o cordeiro realiza atividades diferentes das avaliadas. 2.4 Análises Estatísticas O delineamento utilizado foi o inteiramente casualizado (DIC) e a análise adotada paras os padrões de comportamentos foi o teste do Qui-quadrado. Os dados de frequência foram analisados utilizando modelos lineares generalizados considerando uma distribuição binomial negativa a 5% de probabilidade, utilizando-se do procedimento GLIMMIX do programa estatístico SAS (1999). 47 3. RESULTADOS E DISCUSSÃO 3.1 Padrões comportamentais de fêmeas e crias durante as primeiras horas pósparto Os dados observados para as médias das frequências para latência de tocar a cria, tempo de ficar em pé e realizar a primeira mamada não diferiram entre os tipos de parto simples e duplo dentro de cada lote (Tabela 1). Os resultados mostraram que as ovelhas Morada Nova apresentam uma habilidade materna satisfatória no que se refere ao tempo de contato com sua cria dando estímulos que proporcionam maior agilidade as suas crias através dos comportamentos necessários para sobrevivência nas primeiras horas de vida. O TTC em ambos os lotes foram semelhantes, demonstrando que mesmo uma fêmea que pare dois borregos consegue imprimir os primeiros cuidados maternais necessários para manter uma relação de contato com sua prole. Isso implica em afirmar que a ovelha da raça Morada Nova possui uma boa habilidade materna desde os primeiros momentos após o parto independente do número de crias paridas. Para as variáveis TTC e TEP os resultados apresentados neste trabalho foram similares aos encontrados por Rainere (2008) que relataram TTC de 1,8 segundos e TEP 20,87 minutos para ovinos da raça Santa Inês. Mariz et al. (2007) Trabalhando com ovinos Morada Nova em confinamento, apresentaram um valor para TPM de 44,09 minutos. Esse tempo foi superior aos resultados encontrados no presente trabalho. Essa diferença pode ser atribuída ao fato das fêmeas, naquele estudo, estarem em restrição alimentar o que possa ter apresentado como uma condição menos apropriada para o manejo de animais dessa raça. A característica tempo para ficar em pé é uma das mais importantes na determinação da capacidade de sobrevivência. Quanto menor é esse tempo maior é a capacidade da cria de buscar a teta para a mamada. Em situação de manejo extensivo essa capacidade é imprescindível para que a taxa de mortalidade não se eleve muito. Em trabalho realizado com ovino da raça Ideal, Rech et al. (2008) observaram um tempo de 27,83 minutos para as crias ficarem em pé e de 78,87 minutos para realizar a primeira mamada, tempo superior aos registrados no presente trabalho. Quanto mais o cordeiro demorar em levantar, mais tempo ele desprenderá para ingerir o colostro e, possivelmente, serão menos precoces (Kippert et al., 2006). 48 Tabela 1. Frequência das latências para tocar a cria em segundos, ficar em pé e realiza a primeira mamada em minutos, relacionados com o tipo de parto Tipo de Parto Variáveis Lote A Lote B Simples Duplo Simples Duplo TTC 3 3 2 2 TEP 3,88 8,62 5,34 6,27 TPM 30,83 32,32 24,22 24,22 TTC= Tempo de tocar a cria (segundos); TEP= Tempo para ficar em pé (minutos); TPM= Tempo para mamada (minutos) As variáveis comportamentais TEP e TPM mostraram-se relativamente maiores para as ovelhas de parto duplo em ambos os lotes de parição, possivelmente pelo fato dessas fêmeas ainda estarem em trabalho de parto. Observou-se que o cuidado desprendido com o primeiro borrego é diminuído, já que uma segunda cria prestes a nascer necessita de maior atenção da fêmea. Outro fator importante para garantir a sobrevivência relaciona-se com o tempo para realizar a primeira mamada. Neste trabalho, os baixos valores de TPP, associados à agilidade da cria Morada Nova em procurar o úbere e mamar, independente do tipo de parto, sugere possibilidade de maiores taxas de sobrevivência das crias nas primeiras horas após o parto. Para Toledo et al. (2007), as variáveis latência de ficar em pé e realizar a primeira mamada podem ser utilizadas como medida de vigor ou agilidade em ingerir o colostro logo após o parto. Na Tabela 2 são apresentadas as variáveis comportamentais demonstradas pelas ovelhas em relação às crias, logo após o parto, nos dois lotes. No Lote B, as ovelhas de parto duplo apresentaram menor frequência da postura em pé do que as de parto simples. Isso pode ser atribuído ao maior desgaste da ovelha após trabalho de parto, ocasionando menor permanência destes animais em pé pelo maior esforço físico gerado. Também no Lote B, as ovelhas de parto simples dedicaram mais tempo à atividade de limpeza da cria. A princípio, com a massagem que o ato de limpeza proporciona era de se esperar uma diferença no tempo para ficar em pé, resultante desse estímulo. 49 O contato com o cordeiro (CCO) é a primeira estimulação de reconhecimento do laço maternal. Os padrões comportamentais apresentados pelas ovelhas são decisivos para a sobrevivência do cordeiro, favorecendo o desenvolvimento de laços de exclusividade entre mãe e cria. Os sistemas de cuidados maternos entre os mamíferos são bem variados, pois existem cuidados que envolvem o contato mãefilhote mais continuado. Tabela 2. Média das Frequências de Posturas e Atividades das ovelhas em relação ao tipo de parto segundo o lote de parição Lote A Tipo de Parto Lote B Simples Duplo Simples Duplo Deitada 0,16 1,83 1,42 2,09 Em pé 17,00 16,58 26,14 a 13,60 b Deslocamento 9,16 19,25 23,71 19,27 Contato com a cria 8,33 8,50 18,28 15,72 Ingerindo membranas 8,33 13,08 4,71 11,18 Limpeza da cria 58,50 47,50 116,57 a 54,00 b Estimulando a cria 8,66 5,83 A 0,42 1,00 B Facilitando a mamada 13,16 20,75 15,85 16,00 Dificultando a mamada 0,16 2,16 0,57 0,18 293,83 349,00 190,71 233,18 Sem atividade aparente 9,50 13,33 A 7,28 4,00 B Outras atividades 7,83 1,58 B 9,00 13,00 A Postura Atividade Vocalizando Médias seguidas de letras minúsculas diferentes na linha dentro do lote diferem estatisticamente, e médias seguidas de letras maiúsculas de acordo com o tipo de parto entre os lotes diferem entre si ao nível de 5% de probabilidade, pelo teste do Qui-quadrado. As frequências de limpeza da cria se mostraram significativas apenas no Lote B, nas quais as ovelhas de parto simples apresentaram maior frequência deste comportamento em relação às fêmeas de parto duplo. De fato, a limpeza já se configura como um ato de estímulo afetivo, acarretando em um mecanismo eficaz para o enfrentamento da cria. A menor frequência de limpeza apresentada pelas ovelhas de 50 parto duplo pode ser explicada pelo fato de que apenas a primeira cria nascida era avaliada e isso possa ter interferido nos resultados relativos a este comportamento. A limpeza da cria proporciona o reconhecimento, desobstrução das vias aéreas, aumento da circulação sanguínea, regulação da homeotermia e secagem dos borregos. Segundo Dias et al. (2008), o olfato é utilizado em muitos aspectos do comportamento materno-filial para garantir a sobrevivência da prole. No momento do parto, mecanismos neuroendócrinos modificam a resposta da fêmea frente aos recémnascidos, sendo estabelecido o vínculo materno-filial através de informações olfatórias. Outro padrão relacionado com a habilidade materna da ovelha, que é a facilitação da mamada (FMA), é um dos pontos mais importantes no que diz respeito à sobrevivência do cordeiro, uma vez que é através do colostro que o animal adquire imunidade. Em ambos os lotes, este tipo de comportamento demonstrou frequências semelhantes o que mostra relação positiva com o sucesso da primeira mamada nessa raça. A vocalização se apresentou com elevada frequência nas ovelhas, independente do tipo de parto e da estação de parto. Pode-se concluir que a vocalização representa, para a raça ovina Morada Nova, em um mecanismo de destaque no estabelecimento de vínculo entre a mãe e a cria. A identificação da mãe pela cria e da cria pela mãe por meio dos sons, além dos parâmetros visuais, concorrem para facilitar o acompanhamento quando a visualização está prejudicada. Isso é essencialmente importante nas condições da caatinga. Apenas as variáveis observadas nas ovelhas de parto duplo estímulo da cria, sem atividade aparente e outras atividades apresentaram diferença estatística (P>0,05) entre os lotes de parição (Tabela 2). No entanto, ressalta-se que dessas variáveis a que sugere maior importância na sobrevivência da cria é aquela ligada ao seu estímulo. As fêmeas de parto duplo do lote A demonstraram estimular mais seus filhotes comparadas às fêmeas de parto duplo do lote B, o que possivelmente sugere a maior agilidade das crias em ficarem de pé, num menor tempo comparado com o do Lote B, mas que não apresentou efeito significativo entre os lotes com relação às variáveis TEP e TPM. As fêmeas de parto duplo do lote A ficaram sem realizar nenhum tipo de atividade com maior frequência comparada com as de parto duplo do lote B. Isto possa ser atribuído ao fato das fêmeas do lote B terem gastado mais tempo lambendo e limpando suas crias. 51 Durante o período de observação as fêmeas apresentaram algumas atividades diferentes das constantes no etograma. Isso aconteceu principalmente aquelas de parto duplo. Essas atividades foram inquietação, batendo a pata no chão e se sacudindo, relacionados mais ao parto da segunda cria. Tanto com relação às características de postura quanto de atividades, não houve evidências de diferenças quando se compararam ovelhas de partos simples e duplos. Novamente, esses resultados podem estar sugerindo uma suficiente habilidade materna uma vez que poderia haver menor atividade da ovelha dedicada ao primeiro filho visto haver um segundo borrego para cuidar. No entanto, o que se verificou foi uma semelhança de comportamento em relação às ovelhas que pariram de apenas um borrego. Comportamentos como afastar-se e apresentar agressão contra a cria não foram observadas. Esse registro é uma comprovação de observações realizadas por criadores que atestam a acentuada habilidade de cuidar das crias apresentada por esta raça. Neste trabalho a frequência de comportamentos agonísticos foi praticamente nula. Possivelmente esses comportamentos não façam parte da herança genética desses animais que apresentam uma excelente habilidade em cuidar de suas crias. Observa-se que crias desta raça se mostram ágeis em permanecer em pé e realizar a mamada logos nos primeiros minutos, comparados com ovinos de outras raças. Isto pode estar vinculado com a herança genética advinda das raças que participaram da formação da raça Morada Nova. Na verdade, uma questão crítica em este estabelecimento é se o recém-nascido aprende a reconhecer algumas características físicas individuais da mãe ou responde a alguns sinais de aceitação (como por exemplo, acesso à lambida, permitindo então que a mãe exponha o úbere para realização da mamada ou a produção de um determinado tipo de vocalização) ou comportamentos de rejeição (por exemplo, agressão e agitação), mostrado pela mãe ao interagir com os animais jovens (Sèbe et al. 2010). Na Tabela 3 os padrões de comportamento das crias nas primeiras horas pósparto não variaram em função do tipo de parto dentro de cada lote. Quando se comparou os lotes de parição observou-se diferença significativa (P>0,05) com relação às variáveis em pé, deslocamento e procurando o úbere entre os tipos de parto duplo. As crias do lote B apresentaram menores frequências dessas variáveis quando comparadas com as do Lote A. Provavelmente esses resultados possam ser explicados pelas TEP e TPM que para as crias do lote B os animais se mostraram mais ágeis. 52 Tabela 3. Frequências das Posturas e Atividades das crias em relação ao tipo de parto em dos lotes de parição VARIÁVEIS COMPORTAMENTAIS Lote A Lote B Postura Simples Duplo Simples Duplo Deitada 9,33 7,75 9,00 8,72 Em pé 11,00 18,16 A 15,42 10,54 B Deslocamento 23,26 49,67 A 39,71 26,18 B Tentando levantar 17,33 8,25 8,28 10,54 Levantando 2,33 7,16 3,85 2,18 Procurando o úbere 24,33 34,25 A 17,85 15,81 B Tentando mamar 18,67 25,16 19,00 21,00 Mamando 13,83 10,75 14,85 11,18 Vocalizando 70,67 37,41 52,28 50,90 Sem atividade aparente 14,16 13,41 11,71 8,90 Outras atividades 4,00 6,00 11,28 4,81 Atividades Médias seguidas de letras maiúsculas na linha de acordo com o tipo de parto entre os lotes diferem entre si ao nível de 5% de probabilidade, pelo teste do Qui-quadrado. Os resultados apresentados para as atividades e posturas das crias dentro de cada lote não demonstram diferença estatística independente do tipo de parto, que as fêmeas promovem todos os estímulos necessários às crias. Observando os lotes, referente a outras atividades apresentadas pelas crias, os animais do Lote B demonstraram apresentar mais relacionadas à interação com outras fêmeas, atividades relacionadas com comportamentos lúdicos e tentar mamar em outras ovelhas, comparados com os do Lote A. Estes comportamentos foram observados durante as avaliações no período experimental. Paranhos e Cromberg (1998) ressaltam que o sucesso da mamada parece sugerir uma sequência definida, onde se identifica os comportamentos ficar em pé, procurar os tetos e finalmente mamar. 53 No entanto, apesar da mãe desempenhar um papel importante no investimento parental, o animal jovem pode também contribuir para o sucesso do reconhecimento maternal através do desenvolvimento de um relacionamento com sua mãe. Com efeito, o reconhecimento é crucial para a sobrevivência da cria, ao passo que um erro no reconhecimento representa um maior custo para fêmeas adultas (Sèbe et al., 2010). Foram realizadas as correlações entre os padrões de comportamento estudados nas mães e crias, com isso foram observadas algumas correlações positivas entre as variáveis comportamentais encontradas nas Tabelas 4 e 5. Há relatos de correlação negativa entre a latência para ficar de pé e a intensidade e quantidade de contato entre a mãe e o filho durante a primeira hora de vida (Cromberg et al., 1997). Para a variável IME nas fêmeas observou-se correlação positiva com as variáveis FMA e VOC, e estas correlacionaram positivamente com a IME. Enquanto que nas crias observamos correlação entre DES e PUB e TLE com VOC e vice-versa, podem ser indícios da adaptação comportamental dessa raça em condições semiextensivas. 54 Tabela 4. Correlação dos comportamentos entre as mães dos dois lotes D EP DES CCO LCO IME FMA DMA SAA ESC VOC OAT D 1.00000 -0.15561 0.3648 0.02134 0.9017 0.20116 0.2394 0.07136 0.6792 -0.05330 0.7575 -0.10923 0.5260 0.28322 0.0942 -0.06054 0.7258 0.22364 0.1898 -0.25005 0.1413 0.22364 0.1898 EP -0.15561 0.3648 1.00000 DES 0.02134 0.9017 0.22493 0.1872 1.00000 CCO 0.20116 0.2394 0.21316 0.2119 0.30585 0.0697 1.00000 0.22493 0.1872 0.21316 0.2119 0.12916 0.4528 -0.37936 0.0225 -0.06980 0.6858 -0.06260 0.7168 0.13050 0.4481 0.05300 0.7589 -0.14109 0.4118 0.30585 0.0697 -0.04856 0.7785 0.15746 0.3591 0.15033 0.3815 0.15995 0.3514 -0.02076 0.9043 0.06510 0.7060 -0.10042 0.5601 0.26651 0.1161 -0.22899 0.1791 -0.03646 0.8328 -0.06351 0.7129 -0.08528 0.6209 0.04675 0.7866 -0.29109 0.0850 0.21357 0.2111 0.03738 0.8286 0.15008 0.3823 LCO 0.07136 0.6792 0.12916 0.4528 -0.04856 0.7785 0.26651 0.1161 1.00000 -0.33234 0.0477 -0.19327 0.2587 0.11176 0.5164 -0.08118 0.6379 -0.01918 0.9116 -0.01029 0.9525 0.23827 0.1617 IME FMA -0.05330 -0.10923 0.7575 0.5260 -0.37936 -0.06980 0.0225 0.6858 0.15746 0.15033 0.3591 0.3815 -0.22899 -0.03646 0.1791 0.8328 -0.33234 -0.19327 0.0477 0.2587 1.00000 0.53542* 0.0008 0.53542* 1.00000 0.0008 -0.09141 -0.05416 0.5959 0.7537 -0.10198 0.10182 0.5540 0.5546 -0.27625 -0.13233 0.1029 0.4417 0.53814* 0.47848 0.0007 0.0032 DMA 0.28322 0.0942 -0.06260 0.7168 0.15995 0.3514 -0.06351 0.7129 0.11176 0.5164 -0.09141 0.5959 -0.05416 0.7537 1.00000 -0.21014 0.2186 -0.17575 0.3052 -0.46876 0.0039 -0.05859 0.7343 0.24567 0.1487 0.18862 0.2706 SAA -0.06054 0.7258 0.13050 0.4481 -0.02076 0.9043 -0.08528 0.6209 -0.08118 0.6379 -0.10198 0.5540 0.10182 0.5546 -0.05859 0.7343 1.00000 ESC 0.22364 0.1898 0.05300 0.7589 0.06510 0.7060 0.04675 0.7866 -0.01918 0.9116 -0.27625 0.1029 -0.13233 0.4417 0.24567 0.1487 0.41585 0.0117 1.00000 0.41585 0.0117 -0.00009 0.07804 0.9996 0.6510 VOC -0.25005 0.1413 -0.14109 0.4118 -0.10042 0.5601 -0.29109 0.0850 -0.01029 0.9525 0.53814* 0.0007 0.47848 0.0032 0.18862 0.2706 -0.00009 0.9996 0.07804 0.6510 1.00000 1.00000 OAT 0.02275 0.8952 0.21357 0.2111 -0.03738 0.8286 0.15008 0.3823 0.23827 0.1617 -0.21014 0.2186 -0.46876 0.0039 -0.17575 0.3052 -0.10177 0.5548 -0.13181 0.4435 -0.31535 0.0610 -0.10177 0.5548 -0.13181 0.4435 -0.31535 0.0610 1.00000 54 55 Tabela 5. Correlação dos comportamentos entre as crias dos dois lotes D D 1.00000 LEV 0.32808 0.0508 0.21941 0.1985 0.11055 0.5210 -0.03728 PUB 0.8291 0.08771 EP DES TLE VOC TMA MAM SAA OAT 0.31017 0.0656 0.05350 0.7566 0.20823 0.2230 0.00637 0.9706 0.09769 0.5766 EP 0.32808 DES 0.21941 TLE 0.11055 LEV PUB -0.03728 0.08771 VOC 0.31017 TMA 0.05350 MAM 0.20823 SAA 0.00637 OAT 0.09769 0.0508 1.00000 0.1985 0.46364 0.0044 1.00000 0.5210 -0.09997 0.5618 -0.12312 0.4744 1.00000 0.6110 0.35119 0.0357 0.66039* <.0001 0.18739 0.2738 0.10433 0.0656 0.16209 0.3449 0.14498 0.3989 0.54855* 0.0005 -0.02672 0.7566 0.10142 0.5562 0.24802 0.1447 -0.06514 0.7058 -0.03551 0.2230 0.29341 0.0824 0.11137 0.5178 0.01474 0.9320 -0.01393 0.9706 0.22283 0.1915 -0.08355 0.6280 0.17481 0.3079 -0.15468 0.5766 0.23096 0.1819 0.51333 0.0016 -0.10053 0.5656 0.05764 0.5448 1.00000 0.8770 0.11966 0.4870 1.00000 0.8371 0.49510 0.0021 0.13081 0.4470 1.00000 0.9357 0.34643 0.0385 -0.02410 0.8890 0.50385 0.0017 1.00000 0.3677 0.17073 0.3195 0.17789 0.2993 0.05052 0.7698 0.05537 0.7484 1.00000 0.7423 0.42652 0.0106 0.15821 0.3640 0.26580 0.1228 0.36345 0.0319 0.27669 0.1076 1.00000 0.46364 0.0044 -0.09997 0.5618 -0.01751 -0.12312 0.4744 0.10722 -0.16316 0.8291 -0.01751 0.9193 0.10722 0.5337 -0.16316 0.3417 1.00000 0.9193 0.35119 0.0357 0.16209 0.3449 0.10142 0.5562 0.29341 0.0824 0.22283 0.1915 0.23096 0.1819 0.5337 0.3417 0.66039* 0.18739 <.0001 0.2738 0.14498 0.54855* 0.3989 0.0005 0.24802 -0.06514 0.1447 0.7058 0.11137 0.01474 0.5178 0.9320 -0.08355 0.17481 0.6280 0.3079 0.51333 -0.10053 0.0016 0.5656 0.10433 0.5448 -0.02672 0.8770 -0.03551 0.8371 -0.01393 0.9357 -0.15468 0.3677 0.05764 0.7423 0.11966 0.4870 0.49510 0.0021 0.34643 0.0385 0.17073 0.3195 0.42652 0.0106 0.13081 0.4470 -0.02410 0.8890 0.17789 0.2993 0.15821 0.3640 0.50385 0.0017 0.05052 0.7698 0.26580 0.1228 0.05537 0.7484 0.36345 0.0319 0.27669 0.1076 55 56 4. CONCLUSÃO Concluiu-se que ovelhas da raça Morada Nova, independente do tipo de parto, demonstram os padrões de comportamentos maternais necessários à sobrevivência da cria, destacando-se a rapidez como praticam os cuidados alo corporais com a cria. 57 5. Referências Bibliográficas CROMBERG, V. U.; PARANHOS da COSTA, M. J. R. Mamando logo, para fazer crescer a receita. ANUALPEC: Anuário da Pecuária Brasileira. São Paulo, FNP, 1997, p.215-217. 54 DIAS, C.G.A.; SILVA, L.D.M. da.; MATTOS, M.R.F. Papel do olfato no estabelecimento do vínculo materno-filial em mamíferos. Ciência Animal, v.18, n.2, p.57-66, 2008. KIPPERT, C.J.; RORATO, P.R.N.; CAMPOS, L.T. et al. Efeito de fatores ambientais sobre escores de avaliação visual à desmama e estimativa de parâmetros genéticos, para bezerros da raça Charolês. Ciência Rural, v.36, n.2, p.579-585, 2006. MARIZ, T.M. A.; PIMENTA FILHO, E.C.; MEDEIROS, A.N. et al. Relação maternofilial da raça Morada Nova recebendo dietas com três níveis de energia, ao final da gestação. Revista Brasileira de Zootecnia, v.36, n.6, p.1889-1893, 2007. NATIONAL RESEACH COUNCIL. Nutrients Requirements of sheep. 6 ed. Washington, DC, USA, 1985. 99 p. PARANHOS DA COSTA, M. J. R; SCHMIDEK, A.; TOLEDO, L. M. Relações materno-filiais em bovinos de corte do nascimento à desmama. Revista Brasileira Reprodução Animal, v.31, n.2, p.183-189, 2007. PARANHOS da COSTA, M. J. R.; e CROMBERG, V. U. Relações materno-filiais em bovinos de corte nas primeiras horas após o parto. In: Comportamento Materno em Mamíferos: bases teóricas e aplicações aos ruminantes domésticos, São Paulo: ed. Sociedade Brasileira de Etologia, p.215-235, 1998. RAINERE, C. Perfil do Comportamento materno-filial de ovinos da raça Santa Inês e sua influência no desempenho dos cordeiros ao desmame. 2008. 61f. 58 Dissertação (Mestrado em Zootecnia) – Faculdade de Zootecnia e Engenharia de Alimentos, Universidade de São Paulo, Pirassununga. RECH, C. L. S.; RECH, J. L.; FISCHER, V. et al. Temperamento e comportamento materno-filial de ovinos das raças Corriedale e Ideal e sua relação com a sobrevivência dos cordeiros. Ciência Rural, v.38, n.5, p.1388-1393, 2008 SAS, 1999. Statistical Analysis Systems Institute, User`s guide,version 8. SAS institute Inc., North Carolina 295 pp. SÈBE, F.; DUBOSCQ, J.; AUBIN, T.; LIGOUT, S.; POINDRON, P. Early vocal recognition of mother by lambs: contribution of low- and high-frequency vocalizations. Animal Behaviour, n.79, p.1055-1066, 2010. SÈBE, F.; AUBIN, T.; BOUÉ, A.; POINDRON, P. Mothereyoung vocal communication and acoustic recognition promote preferential nursing in sheep. Journal of Experimental Biology, n.211, p.3554-3562, 2008. TOLEDO; L. M.; PARANHOS DA COSTA, M. J. R.; TITTO, E. A. L.; FIGUEIREDO, L. A.; ABLAS, D. S. Impactos de variáveis climáticas na agilidade de bezerros Nelore neonatos. Ciência Rural, v.37, n.5, p.1399-1404, 2007. 59 6. Implicações e Sugestões Mais estudos devem ser desenvolvidos na intenção de melhorar a forma de avaliação de animais nativos nas condições do semiárido; Algumas metodologias simples podem ser adaptadas para realizarem as avaliações a pasto; Provavelmente as fêmeas dessa raça necessitem de uma avaliação mais acurada, quanto ao escore corporal ideal para iniciarem uma estação de monta; Sugeri-se que durante as avaliações de comportamento materno-filial, possa se disponibilizar uma área maior para o curral maternidade, para que possa evitar a concorrência por espaço e o roubo de crias por outras fêmeas.
Baixar