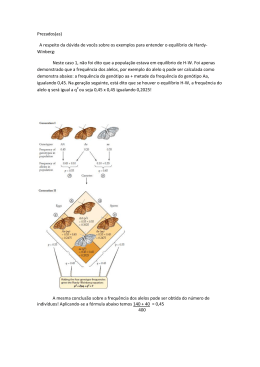
1 Processos Evolutivos Genética de Populações http://dreyfus.ib.usp.br/bio212/ 2 Genética de populações A genética de populações e suas relações com a Evolução Durante os últimos anos temos assistido a uma explosão na quantidade de informações tanto sobre a variabilidade genética das populações como sobre a quantidade de diferenciação genética existente entre as espécies atuais. Os dados obtidos por técnicas de Biologia Molecular têm se avolumado de tal maneira que existe um consenso entre os especialistas sobre a necessidade de métodos de análise mais poderosos ou até mesmo de suporte teórico novo para se poder lidar com tamanha massa de dados. Por outro lado, a preocupação crescente com a diminuição da biodiversidade tem levantado questões sobre tópicos como a fragilidade genética de populações pequenas, estratégias genéticas para a conservação de espécies ameaçadas e problemas correlatos, que naturalmente demandam informações sobre a estrutura genética de populações naturais. Estes problemas atuais necessariamente se baseiam numa área específica e recente da Ciência como um todo, mas antiga se considerarmos o desenvolvimento da genética: a genética de populações. A seguir traçaremos um pequeno histórico do seu desenvolvimento. O trabalho mais importante de Charles Robert Darwin (1809-1882), "A origem das espécies", e o trabalho de Alfred Russell Wallace forneceram uma base fenomenológica para o processo de evolução orgânica. "A origem", publicada em 1859, quando o autor contava com 50 anos de idade, provocou, sem dúvida, um grande impacto. A primeira edição da obra, com 1.250 exemplares, esgotou no primeiro dia de publicação, 22 de novembro, e até 1876, somente na Inglaterra, já haviam sido vendidos 16.000 exemplares. Isso não significa que Darwin só tenha recebido aplausos; muito pelo contrário, esta obra encontrou violenta oposição, não exatamente de natureza científica, mas de caráter emocional por parte da Igreja Anglicana (liderada inicialmente pelo bispo Wilberforce) e por parte da sociedade leiga, por não se admitir a idéia da origem do homem a partir de primatas. Essa resistência persiste até hoje em alguns grupos de religiosos fundamentalistas, principalmente os criacionistas, que defendem a interpretação bíblica "ipsis litteris" da criação do mundo e dos seres vivos. A Igreja Católica admitiu, após quase 100 anos de resistência, através da Encíclica Papal "Humanis Genesis" (1951), a ascendência biológica do homem. Em 1997, através de comunicado do papa João Paulo II, a Igreja Católica deixou de considerar a evolução biológica como uma teoria científica e passou a considerá-la como um fato. Na comunidade científica a teoria de Darwin foi aos poucos encontrando abrigo e se tornando cada vez mais uma fonte de inspiração para o desenvolvimento de novas pesquisas. Além disso, a teoria da evolução, com a devida complementação que mais tarde a genética lhe emprestou, permitiu uma visão unificada de toda a Biologia. Os primeiros grandes defensores e divulgadores da teoria da evolução foram Thomas H. Huxley (avô do biólogo Julian Huxley e do escritor Aldous Huxley), na Inglaterra, e Ernst H. Haeckel na Alemanha, onde o darwinismo foi ensinado pela primeira vez em 1860. É importante salientar que o mecanismo da hereditariedade é fundamental para a compreensão da teoria da evolução. Os trabalhos de Mendel (realizados praticamente ao mesmo tempo em que Darwin formulava sua teoria) tornaram-se amplamente conhecidos somente a partir de 1900. Apenas a partir daquele momento foi possível o estabelecimento de ligações adequadas entre a teoria da evolução e a mecânica da hereditariedade. Antes disso, as idéias predominantes sobre a herança biológica eram oriundas do pensamento de Francis Galton, um primo de Darwin que se dedicava ao estudo de caracteres quantitativos através da aplicação de métodos estatísticos para desvendar os princípios da hereditariedade. Em 1897 Galton propôs a lei da ancestralidade que viria a provocar, no futuro, grande resistência à aceitação do 3 mendelismo; independentemente disso Galton foi um cientista muito importante por ter criado uma escola de biometria, com cientistas do porte de Karl Pearson, que desenvolveu métodos estatísticos usados até hoje. A discrepância entre a teoria de Galton e a genética mendeliana só foi resolvida mais tarde, graças principalmente aos trabalhos do dinamarquês Johanssen sobre a herança de caracteres quantitativos (1909) e a um trabalho publicado em 1918 por Ronald Alymer Fisher. Assim, no começo deste século, a teoria da evolução de Darwin sofreu o forte impacto das novas descobertas da genética, iniciadas com a divulgação do trabalho de Mendel. Tornou-se claro que a matéria-prima da evolução são os genes e as suas leis de transmissão de uma geração a outra, ao nível populacional. A união das idéias de Darwin com as noções exatas da mecânica da transmissão do material hereditário originou a teoria moderna da evolução, também conhecida pelo nome de neodarwinismo. As grandes sínteses empíricas desta visão renovada da teoria de evolução foram feitas nas décadas de 30 e de 40 por três importantes nomes da Ciência: Theodosius Dobzhansky, na sua obra clássica "Genetics and the Origin of Species", cuja primeira edição apareceu em 1937, seguida de várias reimpressões e edições revistas e ampliadas; Julian Huxley em "Evolution, the Modern Synthesis", publicada em 1942, e Ernst Mayr em "Systematics and the Origin of Species", de 1942. A ausência de um bom conhecimento da genética no tempo de Darwin não o impediu de elaborar a sua teoria sobre a origem das espécies, mas deixou um vazio que já foi preenchido nas três primeiras décadas do século 20, com os conhecimentos que viriam a se constituir naquilo que é chamado hoje de "genética de populações". A genética de populações estuda as manifestações da herança no nível populacional. Ela trabalha com modelos, ou seja, representações simplificadas da realidade, usando para isso os elementos que participam do fenômeno (genes, genótipos, fenótipos, gametas, etc.), representados simbolicamente e regras operacionais capazes de traduzir os fenômenos que estão sendo estudados. Estas regras operacionais, em geral, estão sujeitas a princípios matemáticos e estatísticos, de modo que os modelos são chamados de modelos matemáticos. A grande importância desses modelos é que partem de informações obtidas por biólogos através de observação e experimentação. Os modelos fornecem meios de estimar parâmetros corretamente e permitem fazer previsões que podem ser testadas experimentalmente. Se os testes experimentais não estiverem de acordo com os modelos, estes serão rejeitados ou modificados e outros modelos mais adequados serão formulados. A cada nova informação, novos modelos podem ser estabelecidos. Os modelos permitem, portanto, um tratamento quantitativo dos fenômenos, o estabelecimento de previsões e uma maneira de testar hipóteses. O conjunto de modelos acumulados desde os primeiros trabalhos feitos nesse campo e sua maneira integrada de tratar os processos evolutivos permitem afirmar que a genética de populações é hoje uma ciência à parte. A primeira publicação relativa à genética de populações foi uma simples nota publicada na revista Science, em 10 de julho de 1908, por Godfrey Harold Hardy, o mais importante matemático inglês deste século. Esse trabalho se deve às questões levantadas por um famoso estatístico, Yule, numa conferência pronunciada pelo geneticista Punnett, na Royal Society of Medicine. Yule declarava que se um alelo dominante fosse introduzido numa população, sua freqüência deveria aumentar até atingir o valor 0,5, fazendo com que a relação entre os fenótipos dominantes e recessivos fosse de 3:1. Punnett, não concordando com aquela afirmação, levou o problema para Hardy, que analisou a questão e demonstrou que na ausência de qualquer fator perturbador as freqüências gênicas devem permanecer constantes e a distribuição das freqüências dos genótipos AA, Aa e aa dependerá das freqüências gênicas de A e a, assumindo valores conforme a distribuição binomial. Esta distribuição de freqüências de uma população em equilíbrio se tornou o ponto fundamental de todo o desenvolvimento da genética de populações. Mais tarde, verificou-se que o mesmo resultado já havia sido publicado em 13 de janeiro de 1908 por um médico alemão, Wilhelm Weinberg, num estudo sobre a herança da gemelaridade. 4 Assim, esse equilíbrio é hoje conhecido na literatura como "equilíbrio de HardyWeinberg". Um outro aspecto que preocupou os geneticistas da época, em termos populacionais, foi o efeito do endocruzamento na distribuição das freqüências genotípicas. Este aspecto foi tratado independentemente por H. S. Jennings e por R. Pearl em uma série de trabalhos publicados entre 1912 e 1916. Algumas dúvidas sobre a veracidade das proposições de Pearl levaram também o então jovem geneticista americano Sewall Wright (1889-1988) a se envolver no problema de endocruzamento e sistemas de cruzamento de um modo geral, o que culminou, em 1921, com a publicação de uma série de trabalhos com o título "Systems of mating". Wright tornou-se um dos mais importantes teóricos da genética de populações, juntamente com Ronald Alymer Fisher (1890-1962) e John Burdon Sanderson Haldane (1892-1965). Fisher, além de ter sido um pioneiro da genética de populações, também fez inúmeras contribuições extremamente importantes à Estatística. Haldane, que era professor de Bioquímica em Cambridge, desde cedo manteve interesse por problemas de genética e a partir de 1924 iniciou uma série de publicações sobre genética de populações, centradas no estudo da seleção natural. O estudo da genética vinha apresentando grandes progressos e cada vez mais os problemas evolutivos eram analisados sob o ponto de vista genético, ficando claro que a sede das mudanças evolutivas era o próprio material genético. Cada caráter usado para definir uma população, raça ou espécie é, portanto, um caráter hereditário; estudar as mudanças evolutivas a que estes caracteres estão sujeitos é estudar as mudanças que ocorrem no próprio material genético. Trabalhando com genes é possível estabelecer modelos matemáticos para estimar as freqüências dos mesmos e prever as mudanças que podem ocorrer quando submetidos à ação dos agentes evolutivos. Assim, a essência do processo evolutivo é retratada pela genética de populações que, por volta da década de 30, já tinha suas bases completamente estabelecidas. Em 1930, Fisher havia publicado seu livro "The genetical theory of natural selection"; Wright, em 1931, publicava um longo artigo na revista Genetics, intitulado "Evolution in Mendelian populations" e Haldane, em 1932, publicava o livro "The causes of evolution". Outro grande impacto teórico experimentado pela genética de populações foi sofrido no fim dos anos 60 e começo dos anos 70, devido aos avanços da genética bioquímica. O problema apareceu com a descoberta de uma quantidade inesperadamente alta de polimorfismos proteicos e a resposta, proposta principalmente pelo biólogo japonês Motoo Kimura, foi a teoria neutralista da evolução molecular, segundo a qual estes polimorfismos seriam uma espécie de ruído de fundo do processo evolutivo. Esta questão teórica ainda aguarda solução. A disponibilidade recente de informações sobre filogenias gênicas tem aberto novos horizontes, especialmente com o desenvolvimento da teoria da coalescência. Em todos estes avanços teóricos a linguagem usada, principalmente nas demonstrações matemáticas e nas aplicações de estatística, não era acessível para quem não tivesse algum tipo de preparo nestas áreas. O problema foi facilitado pelo aparecimento de livros-texto, como os que citamos a seguir. O primeiro livro-texto didático de genética de populações foi publicado por Hogben, em 1946 e levava o título “An introduction to mathematical genetics”. Em 1948, seguiu-se “Population Genetics”, de autoria de Ching Chung Li, cuja segunda edição, bastante ampliada, foi publicada em 1976 sob o título "First course in population genetics". Li foi um grande divulgador da genética de populações, além de contribuir também com vários trabalhos originais. Além dos livros de C. C. Li, são conhecidas obras dos seguintes autores: Oscar Kempthorne publicou em 1957 "An introduction to genetics statistics", um livro relativamente complexo, exigindo conhecimentos de estatística; D. S. Falconer, em 1960, publicou "Introduction to quantitative genetics", que é um clássico da genética quantitativa; P. A. P. Moran, publicou, em 1962, "The statistical processes of evolutionary theory"; James F. Crow e Motoo Kimura, publicaram "An introduction to population genetics theory" em 1970 e Regina C. Elandt-Johnson, "Probability models and statistical methods in genetics", em 1971. Desta data para cá apareceram inúmeros outros trabalhos, muitos dos quais apresentam o assunto com o mínimo de formalismo matemático. Entre os 5 livros que também abordam o impacto recente dos resultados de Biologia molecular temos o de Masatoshi Nei, "Molecular evolutionary genetics", de 1987, de Daniel Hartl e Andy G. Clark, "Principles of population genetics" (1989) e o de John Maynard Smith, "Evolutionary genetics" (1989). Embora essas obras abordem o assunto de modo mais profundo que o necessário para um curso básico como o nosso, o registro das mesmas fica pela apresentação dos autores e para aqueles que porventura se interessem pelo assunto. EQUILÍBRIO DE HARDY-WEINBERG Um dos aspectos importantes do estudo da Evolução é a análise da variabilidade genética das populações e do seu comportamento ao longo das gerações. Esses aspectos constituem a preocupação fundamental da Genética de populações, que procura descrever a composição genética das populações bem como sua resposta frente à atuação de fatores tais como o tipo de cruzamento, o tamanho da população, a mutação, a migração e os vários tipos de seleção. A Genética de populações, por quantificar os fenômenos evolutivos, fornece parâmetros para a análise da variabilidade genética das populações, sua origem e manutenção. Vamos, portanto, mostrar como essa variabilidade é caracterizada para fins do estudo da Genética de populações. Freqüências gênicas A fim de conceituar freqüência gênica, vamos considerar inicialmente o que ocorre com um par de genes autossômicos em organismos diplóides. Supondo que não haja dominância, poderemos distinguir os três genótipos possíveis, representados por AA, Aa e aa. Esses três genótipos corresponderão a três classes fenotípicas diferentes. Assim, em uma população constituída de N indivíduos poderemos contar D indivíduos AA, H indivíduos Aa e R indivíduos aa. Os valores D, H e R são chamados de freqüências absolutas (note as letras maiúsculas) enquanto que esses valores, divididos pelo total de indivíduos da população (N) nos dão as freqüências relativas (representadas pelas mesmas letras, só que minúsculas). AA d = Aa D N h= H N aa r= R N A soma das freqüências relativas é sempre 1. D H R D+ H + R N + + = = =1 N N N N N As freqüências relativas podem ser interpretadas, no caso de amostragens muito grandes, como probabilidades, ou seja, d é a probabilidade de se tomar "ao acaso" um indivíduo AA desta população. Podemos agora procurar saber quais as freqüências gênicas nessa população. Neste caso, o método direto para estimar as freqüências gênicas é o da contagem simples. Dada a população: 6 AA Aa aa N D H R contamos o número de alelos A e a e estimamos as freqüências gênicas. Por se tratar de uma população diplóide vamos atribuir a cada indivíduo dois genes. A população toda terá, pois, 2N genes. Os indivíduos AA terão 2D genes A e os indivíduos Aa terão H genes A, perfazendo um total de 2D + H genes A em uma população com um total de 2N genes; logo, a freqüência do alelo A será: f (A) = 2D + H 2D H h = + =d+ = p 2N 2N 2N 2 Com o mesmo raciocínio veremos que a freqüência do alelo a será: f (a) = 2R + H 2R H h = + = r+ = q 2N 2N 2N 2 pode-se verificar que: p+q = d+ h h + +r = 1 2 2 Uma vez calculada a freqüência de um alelo, a freqüência do outro pode ser obtida pela diferença em relação à unidade, uma vez que p+q =1 p =1−q e q =1− p Exercícios: 1. Determine nos casos que se seguem fenotípicas e as freqüências gênicas: as freqüências relativas das classes 7 a) AA Aa aa 25 60 15 N = AA Aa aa 320 0 80 d= h= r= N = d= h= f(A) = p = c) b) r= f(a) = q = AA Aa aa 0 120 80 N = f(A) = p = d) f(a) = q = AA Aa aa 10 180 810 d= h= r= N = d= h= f(A) = p = r= f(a) = q = f(A) = p = f(a) = q = 2. Determine, nos casos que se seguem, as freqüências absolutas das classes genotípicas e as freqüências gênicas (para uma população de 1000 indivíduos): a) c) AA D= 0,30 p= Aa H= 0,60 AA D= 0,20 p= Aa H= 0,80 aa R= 0,10 b) q= aa R= 0,00 q= d) AA D= 0,36 p= Aa H= 0,48 AA D= 0,58 p= Aa H= 0,04 aa R= 0,16 q= aa R= 0,38 q= O que você pode concluir a partir dos resultados do exercício 2? 8 EQUILÍBRIO (OU LEI) DE HARDY-WEINBERG O que fizemos até o momento foi representar um par de genes autossômicos, sem dominância, em uma população diplóide e estimar as freqüências dos alelos. Agora verificaremos o que acontecerá com uma população desse tipo na geração seguinte. Por isso consideramos uma população com reprodução sexuada, que se reproduza por fecundação cruzada. Vamos considerar, para simplificar o problema, um modelo com "gerações discretas", ou seja, uma população na qual não haja cruzamentos entre indivíduos pertencentes a duas ou mais gerações diferentes. Assim, temos uma população de N indivíduos adultos: AA Aa aa N D H R Vamos supor que os cruzamentos nesta população ocorram ao acaso. Tal fenômeno é conhecido como pan-mixia e diz-se que a população que se reproduz assim é panmítica. Pan-mixia significa que a probabilidade de um indivíduo de qualquer genótipo cruzar com outro pertencente a qualquer genótipo depende apenas das freqüências genotípicas. Isso é o mesmo que dizer que não há preferências, seja ela por genótipos iguais ou diferentes, na escolha de parceiros. Considerando as freqüências relativas dos genótipos como probabilidades, podemos dizer que a probabilidade dos indivíduos AA, com freqüência relativa d, 2 cruzar com outro indivíduo de mesmo genótipo é simplesmente dXd = d . Assim, podemos construir um quadro com as probabilidades, ou freqüências, dos cruzamentos "ao acaso". QUADRO 1 - Freqüências de cruzamentos "ao acaso". machos AA Aa aa d h r AA freqüências genotípicas d d 2 dh dr Aa h hd h 2 hr aa r rd rh r fêmeas 2 Lembre-se que a soma dessas probabilidades ou freqüências de cruzamentos será sempre igual a 1, pois (d + h + r ) × (d + h + r) = 1× 1 = 1 Agora vamos verificar qual a descendência deixada por cada um desses cruzamentos. Novamente teremos de fazer algumas considerações a respeito de como calcular o número de descendentes. O número de descendentes por casal é variável, mas podemos admitir que este número não dependa dos genótipos dos indivíduos que formam o casal, sendo, em média, o mesmo. 9 Assim, podemos apresentar a freqüência de descendentes de cada classe de casal pela própria freqüência dos cruzamentos. Podemos assim determinar os genótipos dos descendentes de cada casal e suas respectivas freqüências. Exercício: Complete o quadro abaixo, determinando os genótipos dos descendentes de cada tipo de cruzamento e suas respectivas freqüências. machos AA Aa aa d h r 2 d AA dh/2 AA dh/2 Aa fêmeas AA d Aa h aa r Para determinar as freqüências dos diferentes genótipos na nova geração basta somar as freqüências das 3 classes genotípicas dos descendentes. tipo de cruzamento freqüência de cruzamento Descendentes AA AA X AA d2 d2 AA X Aa 2dh dh AA X aa 2dr Aa X Aa h2 Aa X aa 2hr aa X aa r2 Total 1 Aa aa dh 2dr h 2 /4 h 2 /2 h 2 /4 hr hr r2 (d+h/2)2 2(d+h/2)(r+h/2) (r+h/2)2 Lembrando que (d + h/2) = p e (h/2 + r) = q verificamos imediatamente que a distribuição das freqüências genotípicas poderá ser expressa assim: AA p p 2 2 (1-q) 2 Aa aa 2pq q 2p(1-p) (1-p) 2q(1-q) q 2 2 ou 2 ou 10 As duas últimas representações têm a vantagem de ter apenas uma variável de freqüência gênica. As freqüências gênicas não mudam, pois: p1 = d1 + h1 = p 2 + pq = p 2 + p − p2 = p 2 Note que uma série de condições foi imposta na elaboração do modelo, o que levou a população ao equilíbrio. Estas condições são: - população de tamanho infinito; reprodução sexuada, por fecundação cruzada; pan-mixia; ausência de mutação; ausência de migração diferencial; ausência de seleção. Nestas condições, uma população não sofre gênicas, ao longo das gerações, nas proporções: p2 alterações em suas freqüências q2 2pq Estas proporções serão atingidas em uma única geração. Alelos Múltiplos O princípio visto acima pode ser estendido para qualquer número de alelos. Sejam os alelos: A1 p1 A2 p2 A3 p3 ... AN, com as freqüências gênicas: ... pN No equilíbrio, as freqüências dos genótipos homozigotos f ( A1 A1 ) = p 2 1 ; f (A2 A2 ) = p 2 2 ; f (A3 A3 ) = p 2 3 ... serão: f (AN AN ) = p2N e as freqüências dos genótipos heterozigotos serão: f (A1 A2 ) = 2p1 p2 ; f (A1 A3 ) = 2 p1 p3 ; f (A2 A3 ) = 2 p2 p3 ... f (AN−1 AN ) = 2 pN −1 pN Caso de um par de genes autossômicos com dominância Estimativa de freqüências gênicas Muitos caracteres hereditários são determinados por genes alélicos que exibem uma relação de dominância, ou seja, não é possível distinguir fenotipicamente os homozigotos (AA) dos heterozigotos (Aa). Assim, a população seria representada por: A _ D aa R Como já visto anteriormente no caso de herança sem dominância, se a população 2 estiver em equilíbrio, os valores d e r corresponderão, respectivamente, a (p + 11 2 2pq) e q . Dada a impossibilidade de distinguir os indivíduos AA de Aa, não podemos estimar as freqüências gênicas por contagem direta dos genes nos homozigotos e heterozigotos. Entretanto, no caso de a população estar em equilíbrio, pode-se estimar a freqüência do alelo a com base na relação f (aa ) = R = r = q2 N logo, R = r = q = 1 − p. N Exercício: Estimar as freqüências gênicas para as seguintes populações: a) p = A_ 150 aa 50 N = q = b) p = A _ aa 220 80 N = q = Algumas considerações sobre o equilíbrio de H.W. O equilíbrio de H.W. pressupõe uma série de condições que raramente são observadas na prática. Dificilmente será encontrada uma população natural em que todas as condições estejam vigorando simultaneamente. A mutação, por exemplo, é um fenômeno espontâneo, de ocorrência constante. Porém, como as taxas de mutação são, -4 -5 em geral, muito baixas, da ordem de um para dez mil ou cem mil (10 ou 10 ), seu efeito sobre o equilíbrio, no instante em que a amostra for colhida, será desprezível. Quando se observa um polimorfismo, os diversos fenótipos poderão ter valores adaptativos diferentes, significando que a seleção natural está ocorrendo. Além disso, em muitas populações podem estar ocorrendo migrações e muitas populações nem sempre terão um tamanho que permita considerá-las infinitamente grandes. Ainda, nem sempre as populações são pan-míticas. Apesar dessas observações, o equilíbrio de H.W. é extremamente importante e extremamente útil em genética de populações. Ele se constitui de um modelo básico muito simples, porque elimina todos os fatores que redundam em complicações. As condições que são impostas são exatamente aquelas que poderiam promover mudanças nas freqüências gênicas ou genotípicas e que implicariam em evolução. Por isso podemos dizer que o equilíbrio de H.W. é conservador, pois ele descreve uma situação de uma população que não está se modificando. Para estudar as mudanças evolutivas, procura-se analisar o efeito que os fatores isolada ou conjuntamente apresentam sobre o equilíbrio de H.W., chegandose, assim, a retratar as mudanças evolutivas. Teste para verificar se a população está ou não em equilíbrio de H.W. Dada uma amostra populacional com a identificação completa dos genótipos feita, surge o problema: Esta pode ser uma amostra representativa de uma população que está em equilíbrio de Hardy-Weinberg? Vamos estudar primeiramente o caso da ausência de dominância, em um loco com dois alelos: 12 AA Aa aa N D H R onde a freqüência do alelo A é p= 2D + H 2N e a do a é 2R + H 2N q= Note que esta estimativa não requer que a população esteja em equilíbrio. Conhecidos os valores de p e q, podemos então estimar as freqüências das três classes genotípicas, como se a população estivesse em equilíbrio. Essas freqüências absolutas, como já vimos, são: AA Aa aa p 2 .N 2 pq.N q 2 .N , N = D+ H+ R e onde p = 1 − q. Exercício: Calcule as freqüências gênicas e as freqüências (relativas e absolutas) para as seguintes amostras: 1. AA Aa aa 204 494 302 3. N = esperadas Aa aa 220 240 N = p = 2 p = 2 p .N= 2. A 40 genotípicas AA 20 q = 2 q = 2pq= 2pq.N= Aa 50 2 q .N= aa 30 4. N = p = 2 p = 2 p .N= p = 2 p = 2 p .N= AA 80 q = 2 q = 2pq= 2 q .N= 2pq.N= Aa aa 100 120 N = q = 2pq= 2 q = 2pq.N= 2 q .N= p = 2 p = 2 p .N= q = 2 q = 2pq= 2 q .N= 2pq.N= Como temos os valores obtidos e os esperados, podemos ter uma idéia se a população da qual a amostra foi retirada estava em equilíbrio ou não. Como foi visto no exercício acima, aparece uma dificuldade: se os valores forem exatamente iguais (o que, na prática, é pouco provável) concluimos que a população encontra-se em equilíbrio, mas se isto não acontece, temos que lançar mão de um outro método para tomar a decisão de forma objetiva. A estatística nos fornece a ferramenta 2 apropriada para a comparação de proporções: o teste do X (que se lê qui-quadrado). Este teste só pode ser aplicado com os valores absolutos. AA Valores observados (O) Valores esperados (E) D 2 p .N Aa H 2pq.N aa R 2 q .N Total N N 13 2 O valor do X é dado por: (observadoi − esperado i )2 esperado i i=1 k Χ2 = ∑ ( D − p .N ) = 2 ou seja: Χ 2 p 2 .N 2 + ( H − 2pq.N )2 2pq.N ( R − q .N ) + 2 2 q2 .N 2 O valor obtido através dessa soma corresponde ao X . Para saber se a população está ou não em equilíbrio, ou seja, se a amostra foi ou não retirada de uma população em equilíbrio, basta comparar este valor com os valores dados na tabela 2 2 do X . O estabelecimento de um determinado valor de X dependerá, inicialmente, da escolha do "nível de significância", que consiste no risco que se corre, em termos de probabilidade, de rejeitar uma hipótese verdadeira. Esse nível deve ser fixado previamente e, em geral, para os experimentos biológicos usa-se o nível de significância (alfa) de 0,05 (ou 5%). 2 Temos, entretanto, vários valores de X para 5%, cada um correspondendo a um determinado número de graus de liberdade (primeira coluna da tabela). É através 2 desse valor que vamos procurar o X na tabela. O número de graus de liberdade é dado pela quantidade de classes independentes envolvidas na análise. No caso de três genótipos, temos apenas 1 classe independente para o mesmo N e o mesmo p. Por exemplo, se existem 34 indivíduos AA em um total de 100 e a freqüência do alelo A é 0,6, então as outras duas classes estarão já determinadas: 52 Aa e 14 aa. Quaisquer outros valores de H e R alterarão as freqüências gênicas ou o total, o que não é permitido. Então existe uma única classe independente e, portanto, um único grau de liberdade. Na prática, o número de graus de liberdade pode ser calculado assim: g.l.= Número de classes-(número de parâmetros da amostra utilizados no teste para calcular os esperados das classes) No nosso caso, temos 3 classes e usamos dois parâmetros: a freqüência de um alelo e o total. g.l. = 3 - 1 - 1 = 1. Assim, vamos procurar na tabela simplificada, anexa, na linha correspondente 2 ao número de graus de liberdade 1, o valor do X para 5%, que é 3,841. Se o valor obtido for maior que 3,841, rejeitamos a hipótese de equilíbrio; se for menor aceitamos a hipótese. Exercício: 2 Aplique o teste do X para as populações do último exercício, e verifique quais estão em equilíbrio. Caso de dois alelos com dominância 2 Não é possível aplicar o teste do X para saber se a população está ou não em equilíbrio no caso de características monogênicas com dominância. A justificativa é muito simples: o cálculo da freqüência do alelo a, através da expressão q= r , é baseado na condição de que a população esteja em equilíbrio, portanto, não há o que testar. 14 Um outro argumento é que não sobra grau de liberdade para o teste. No caso de dominância temos duas classes (A- e aa). Com esses dados estimamos o parâmetro q = R/N. Logo o número de graus de liberdade será: 2(número de classes) - 1(total) - 1(freqüência) = 0 g.l., o que invalida o teste. TABELA SIMPLIFICADA DE VALORES CRÍTICOS DO QUI-QUADRADO Graus de liberdade 1 2 3 4 5 Alfa = 0,05 3,841 5,991 7,815 9,488 11,070 Alfa = 0,01 6,635 9,210 11,345 13,277 15,086 EQUILÍBRIO PARA GENES LIGADOS AO SEXO O processo de reprodução sexuada geralmente envolve a existência de dois sexos que são determinados por um par de cromossomos. Um dos sexos é homogamético (dois cromossomos sexuais iguais) e o outro é heterogamético (dois cromossomos sexuais diferentes). Vamos utilizar arbitrariamente o sistema de determinação sexual XX XY, onde indivíduos XX são fêmeas e XY são machos e que é o sistema mais comum entre os animais. As conclusões permanecem válidas para sistemas ZZ ZW, bastando apenas inverter os sexos. A genética dos cromossomos sexuais pode então ser de três tipos: No caso de genes que estão situados na região de homologia entre os cromossomos X e Y, continuam valendo as condições de herança autossômica. No caso de genes situados na região não homóloga do cromossomo Y, ficam valendo os princípios da herança em organismos haplóides, para os machos. O caso dos genes exclusivos do cromossomo X merece um tratamento especial devido à haplo-diploidia, como veremos a seguir. Tomemos uma população de fêmeas e machos, respectivamente, na geração 0: AA Aa aa D0 H0 R0 N f e AY aY S0 T0 Nm onde D, H e R são as freqüências absolutas de cada um dos genótipos das fêmeas e S e T as freqüências absolutas de cada um dos genótipos dos machos , Nf e Nm o número de fêmeas e machos, respectivamente. As freqüências alélicas entre as fêmeas são: f ( A) = p f = 2D0 + H0 h = d0 + 0 2N f 2 Entre os machos teremos: e f (A) = pm = f (a) = q f = S0 = s0 Nm e 2R0 + H0 h = r0 + 0 2N f 2 f (a) = qm = T0 = t0 Nm Para verificar o que acontece na geração seguinte, g , deixemos os indivíduos da 1 geração 0 reproduzirem-se através de cruzamentos pan-míticos: Cruzamento tipo freq. d 0s0 AA-AY Machos AY d 0s0 aY - AA d 0s0 Fêmeas Aa aa 15 h0 s0 2 Aa-AY h 0s0 aa-AY r0s0 AA-aY d 0t0 d 0t0 Aa-aY h 0t0 h0 t0 2 aa-aY r0t0 Total 1 h0 s0 2 h0 s0 2 h0 s0 2 r0s0 r0s0 d 0t0 h0 t0 2 h0 t0 2 h0 t0 2 r0t0 (d0+h 0/2) r0t0 (r0+h 0/2) s0(d0+h 0/2) s0(r0+h 0/2)+t 0(d0+h 0/2) As freqüências pm1 e qm1 dos alelos A e a, entre os machos, na t0(r0+h 0/2) geração seguinte, serão: pm1 = f ( A) = d0 + h0 = pf0 2 Isso quer dizer que as freqüências geração g , são idênticas às freqüências 1 geração anterior g . Isto é esperado, 0 exclusivamente das fêmeas. As freqüências pf1 e qf1 dos alelos obtido no quadro 1, serão: e qm1 = f (a) = r0 + h0 = qf 0 2 dos alelos A e a, entre os machos, na destes mesmos alelos entre as fêmeas, na pois os machos recebem o cromossomo A e a entre as fêmeas, de acordo com o h 1 h h p f 1 = f (A) = s0 d0 + 0 + s0 r0 + 0 + t 0 d0 + 0 2 2 2 2 e h 1 h h q f 1 = f (a) = t0 r0 + 0 + s0 r0 + 0 + t0 d0 + 0 2 2 2 2 Substituindo s0 e t0 por pm0 e qm0 , e (d0+h0/2) e (h0/2+r0) por pf0 e qf0 teremos: 1 ( p .q + qmo.p f 0) = 2 m0 f 0 + pm0 .q f 0 + qm 0 .p f 0 = 2 p f 1 = pm 0 .p f 0 + = = = 2 pm0 .p f 0 pm0 .( pf 0 + q f 0 ) + p f 0 .( pm 0 + qm0 ) 2 pm0 + p f 0 2 pois pf0 + qf0 = 1 e pm0 + qm0 = 1. Com o mesmo raciocínio demonstra-se que X qf1 = qm 0 + q f 0 2 = 16 Isso quer dizer que pf1 é a média aritmética das freqüências do alelo A entre os machos (pm0 ) e as fêmeas (pf0 ) da geração anterior g0, o que é intuitivo, pois cada um dos dois cromossomos sexuais vem de um progenitor com sexo diferente. EXERCÍCIO: 1. Determine a composição genética de uma população na geração g1, considerando que a população g0 apresenta as seguintes composições: a) b) | AA | Aa |810 180 | AA | |0,50 Aa 0,40 aa | | Nf=1000 10 | | AY | |900 aa | | 0,10 | | AY | |0,50 aY | | Nm=1000 100 | aY | | 0,50| 2. Determine quais as freqüências gênicas das populações parentais femininas e masculinas que deram origem às seguintes populações: a) b) | AA | |0,35 | AA | |0,06 Aa 0,50 Aa 0,58 aa | | 0,15| | AY | |0,70 aY | | 0,30| aa | | 0,36| | AY | |0,60 aY | | 0,40| Condições de equilíbrio As condições de equilíbrio no caso da herança ligada ao sexo são duas: primeira, que as freqüências gênicas sejam as mesmas, entre fêmeas e machos; segunda, que entre as fêmeas a distribuição das freqüências genotípicas obedeça às 2 2 proporções p :2pq:q . Vamos verificar a seguir como as freqüências alélicas de um gene ligado ao sexo entram em equilíbrio. Para isto, utilizaremos a quantidade d, que mede a diferença entre as freqüências alélicas entre os machos e as fêmeas. d = qf - qm como d1 = qf1 = q f 0 + qm0 q f 0 + qm 0 2 d1 é portanto 2 e − qf 0 = − d0 , 2 qm1 = q f 0 , q f 0 + qm 0 − 2q f 0 2 = qm 0 − q f 0 2 ; que é o mesmo que dizer que a diferença entre as freqüências dos dois sexos cai pela metade em cada geração, em termos absolutos, e que essa diferença muda de sinal, também a cada geração. A diferença tende a 0 e, em equilíbrio, a freqüência gênica será qˆ = qˆ f = qˆm , sendo que em cada geração: 17 qn = q mn + 2q fn 3 , que é a média ponderada pelo número de cromossomos X de cada sexo e representa a freqüência do alelo na população total (machos e fêmeas). EXERCÍCIO: 3. Verifique como a população tende ao ponto de equilíbrio calculando, a partir da constituição inicial dada abaixo, as freqüências gênicas e genotípicas e o valor de d em pelo menos 4 gerações consecutivas. Represente graficamente os valores destas freqüências. Calcule também o valor médio das freqüências gênicas em cada uma das gerações: | AA | |0,10 Aa 0,40 aa | | 0,50| | A | |0,70 a | | 0,30| Estimativa das freqüências gênicas Para estimarmos as freqüências gênicas de um gene ligado ao sexo com a finalidade de verificarmos se a população está ou não em equilíbrio, fazemos uma única estimativa, a da freqüência de um dos alelos na população total: AA Aa aa D H R N f p = f (A) = AY aY N S T m 2D + H + S 2R + H + T ; q = f (a) = 2N f + Nm 2N f + Nm Para o teste de X2, teremos 5 classes, dois totais independentes (um total para machos e outro para fêmeas) e uma freqüência estimada a partir da amostra total; portanto teremos 5 - 2 - 1= 2 graus de liberdade. Genes ligados ao sexo com dominância No caso de haver dominância, podemos estimar as freqüências gênicas através de uma equação que será fornecida sem comentários, pois sua dedução exige conhecimentos mais avançados de matemática: A _ D q= aa N R f AY aY N S T m − S + S2 + 4(2N f + Nm ).(2R + T ). 4N f + 2 Nm (quando há dominância em herança ligada ao sexo) O valor de q estimado pela equação acima somente corresponderá ao valor verdadeiro se a população estiver em equilíbrio. Podemos mesmo assim verificar a condição de equilíbrio pelo emprego do teste de X2, pois teremos 4 classes - 2 totais - 1 freqüência gênica, o que resulta em um grau de liberdade. EXERCÍCIO: 18 O resultado da análise de uma dada geração de uma caixa de populações de drosófila com olhos vermelhos (w+_) e brancos (ww) foi: + + | w w | |280 w + 170 ww | | Nf 50 | 500 + | w y | | 400 wy | | Nm 100| 500 Calcule a freqüência do alelo para olhos brancos (w) entre as fêmeas, entre os machos e na população total. Suponha, agora, que nesta mesma amostra não se saiba quantas fêmeas de olhos vermelhos são homozigotas. Usando o estimador apropriado, estime a freqüência do 2 alelo w para essa população e compare com a estimativa anterior. Faça o teste do X para verificar se esses dados estão de acordo com o esperado em equilíbrio. Suponha que você tenha apenas 300 machos, mas que as freqüências gênicas sejam as mesmas + (w y = 240; wy = 60). Estime de novo a freqüência do alelo w, usando esse dado, em lugar de 400 e 100, e verifique que alterações isso acarreta na estimativa. DESVIOS DA PAN-MIXIA: CRUZAMENTOS PREFERENCIAIS E ENDOCRUZAMENTO Entre as várias condições que são impostas para que o equilíbrio de HardyWeinberg (H.W.) se verifique, uma delas é que a população seja completamente panmítica, isto é, os cruzamentos devem ocorrer totalmente ao acaso. Acontece, porém, que podem haver inúmeras maneiras diferentes dos indivíduos se associarem em acasalamento. Essas diferentes maneiras dependem da própria biologia do organismo, devido a determinadas características morfológicas, fisiológicas ou comportamentais. Podemos lembrar, por exemplo, que existem organismos monóicos, como o caso da ervilha, em que a autofecundação é praticamente obrigatória. Por outro lado, alguns organismos, embora sendo monóicos ou hermafroditas, dispõem de mecanismos que evitam a realização da autofecundação. Em determinadas plantas, como a Nicotiana tabacum, existe um sistema regulado por um loco com vários alelos (S1, S2, S3, ...) que determinam a auto-esterilidade. O pólen S1 é incapaz de produzir o desenvolvimento do tubo polínico no estigma de uma planta cujo genótipo tenha o alelo S1. O mesmo acontece para qualquer outro alelo, de tal forma que a autofecundação nunca deixa descendência e também nunca se formam homozigotos para quaisquer dos alelos do loco. Os sistemas de cruzamentos podem também ser alterados artificialmente pelo homem, que interfere seletivamente na escolha dos cruzamentos de plantas e animais domésticos. Em ambos os casos, teremos alterações na distribuição das freqüências genotípicas das populações. Conseqüentemente, devemos procurar analisar os vários modelos que produzem desvios da pan-mixia e determinar quais as conseqüências que acarretam na estrutura genética da população. Basicamente distinguimos duas categorias de alterações da pan-mixia, uma delas devida a um maior índice de cruzamentos consangüíneos, fenômeno esse chamado de endogamia ou de endocruzamento, e a outra categoria corresponde aos cruzamentos preferenciais, positivos e negativos, com relação a um dado caráter genético. Cruzamento preferencial totalmente negativo Cruzamento preferencial totalmente negativo é aquele em que os cruzamentos só ocorrem entre indivíduos de fenótipos (ou genótipos) diferentes. Considerando fenótipos determinados por mecanismo monogênico, as alterações que esse tipo de cruzamento determina na constituição genética da população serão apenas em relação a esse par de genes. 19 Vamos supor um par de alelos A,a, com dominância, que determina um caráter para o qual só ocorra cruzamento preferencial totalmente negativo, ou seja, indivíduos com o fenótipo dominante só cruzam com os de fenótipo recessivo. Assim, a população: AA Aa e aa passaria logo a: Aa aa já que os indivíduos AA ou Aa só se cruzariam com aa. Este tipo de cruzamento preferencial negativo é o que ocorre, por exemplo, na reprodução sexuada com relação aos cromossomos X e Y. Cruzamento preferencial totalmente positivo Diz-se que há cruzamento preferencial totalmente positivo, em relação a um dado caráter, quando há cruzamentos apenas entre os indivíduos fenotipicamente iguais. Seja, pois, um par de genes A,a, com dominância, responsáveis por um caráter em relação ao qual somente ocorram cruzamentos preferenciais totalmente positivos. Assim, em relação a esse par de genes, temos a população: genótipos: freqüências: AA d Aa h aa r Total 1 Lembrando que f(A) = (d + h/2) e f(a) = (r + h/2) Como os indivíduos com o fenótipo dominante só se cruzam entre si, as freqüências de cruzamentos serão expressas em relação ao total de indivíduos desta classe, que é d + h = 1 - r A Tabela abaixo fornece a descendência dos cruzamentos possíveis. (Observar que d, h e r são freqüências genotípicas quaisquer, independente de equilíbrio de H.W.) Tipo de cruzamento AA X AA AA X Aa Aa X Aa aa X aa Freqüência de cruzamento 2 AA 2 Descendência Aa aa d 1−r 2dh 1−r h2 1−r d 1−r dh 1−r h2 4 1−r __ __ dh 1−r h2 2 1−r __ r2 r __ __ h2 4 1−r r2 r 20 Totais [ ( )] [ d + (h 2)] 2 1 h d + h2 1−r 1− r (h 2) 1−r Nesta tabela, as duas "sub-populações" de fenótipos idênticos estão destacadas por linhas duplas. Note que a transferência entre as subpopulações resulta do cruzamento entre heterozigotos. A freqüência do alelo A, nesta nova geração, será: [ ( )] d + h2 f (A) = 1−r 2 ( ) ( ) h . d+h h 2 = d + 2 .(d + h ) + 2 1−r 1− r Como d + h = 1 - r, temos: f(A) = [d + (h/2)], que é igual à freqüência deste alelo na geração anterior. EXERCÍCIO: 1) Analise o que acontece com as freqüências gênicas, genotípicas e fenotípicas Pop. 1 Pop. 2 AA Aa aa 0,45 0,10 0,35 0,30 0,20 0,60 em duas gerações consecutivas, no caso de cruzamentos preferenciais totalmente positivos. Endocruzamento ou endogamia Considera-se endocruzamento ou cruzamento endogâmico quando os indivíduos que se cruzam apresentam ancestrais comuns próximos. Se admitirmos que a população é pan-mítica, pode-se esperar uma determinada freqüência de cruzamentos endogâmicos. Quando a freqüência de cruzamentos endogâmicos observada for maior do que a freqüência esperada (pela pan-mixia), então se diz que está ocorrendo endogamia. A população será, então, endogâmica. Endogamia é, portanto, o desvio da pan-mixia devido ao excesso de endocruzamentos (ou cruzamentos endogâmicos) na população. O endocruzamento ou endogamia corresponde ao termo inglês "inbreeding"; o produto de um "inbreeding" é chamado "inbred". A conseqüência da ocorrência de endocruzamento numa população será o aumento da freqüência dos indivíduos homozigotos e a redução da freqüência dos heterozigotos na população. Para entender melhor o que acontece quando há cruzamento endogâmico, vamos comparar duas situações diferentes, uma com endocruzamento e outra sem: 1) 2 +r 21 Aa AA AA Aa Aa AA Aa Aa aa 2) Aa AA AA Aa Aa Aa aa No caso 1 podemos verificar que o homozigoto aa é constituído por dois genes a, sendo que ambos são cópias do mesmo gene original existente num dos bisavós. No caso 2, também temos um homozigoto aa, formado, porém, por cópias de dois genes a presentes nos avós, de origens independentes. No primeiro caso, diz-se que os dois alelos são iguais por descendência (i.p.d.) e, no segundo, iguais pela origem (i.p.o.). Cotterman chamou os homozigotos com i.p.d. de autozigotos e os homozigotos com genes i.p.o de alozigotos. O importante será determinar quantitativamente o efeito dos cruzamentos endogâmicos sobre a estrutura da população. É claro que o grau de endogamia pode variar e, conseqüentemente, o efeito que acarreta para a estrutura da população também variará. Para se ter uma idéia mais objetiva da conseqüência do endocruzamento na população vamos inicialmente analisar o que ocorre em uma população que se reproduz por autofecundação. A autofecundação corresponde ao grau máximo de endogamia. Consideremos um par de genes autossômicos. Assim, na geração inicial g a 0 população é : AA Aa aa N D H R 0 0 0 g0 f (A) = em que D0 + H0 2 N =p f ( a) = e R0 + H0 N 2 =q Admitindo, como foi feito anteriormente, que o número médio de descendentes por entidade reprodutora seja o mesmo e representando esse número pelo próprio número de cruzamentos ocorridos, podemos ver que, nas gerações seguintes, a população terá as seguintes constituições: geração 0 1 AA Aa aa d0 h0 h0 2 r0 d0 + h0 4 r0 + h0 4 22 h0 h0 + 4 8 h0 4 ... ... ... 1 1 1 d0 + h0 . + + ...+ 4 8 2.2n h0 n 2 1 1 1 r0 + h0 . + + ...+ 4 8 2.2n 2 d0 + ... n r0 + h0 h0 + 4 8 No equilíbrio, teremos: h dˆ = d0 + 0 2 ; hˆ = 0 e rˆ = r0 + h0 2 , pois 1 =1 i i→ ∞ i =1 2 lim ∑ Portanto, na enésima geração, a população estará em constituída apenas pelas duas classes homozigotas. Em equilíbrio, a composição genética da população será: AA Aa aa p 0 q equilíbrio e será Como se pode ver, não ocorreram mudanças nas freqüências gênicas, mas apenas nas freqüências genotípicas, sendo que o valor da classe heterozigota foi diminuindo e os valores das classes homozigotas, aumentando. A endogamia não produz modificação em freqüências gênicas, mas pode ser importante, por exemplo, na eliminação mais rápida de genes letais ou detrimentais da população, quando associada à seleção natural. Podemos definir um índice de heterozigose da população, h /h , chamado por n 0 Wright de índice de pan-mixia, P: P= hn h0 No exemplo que acabamos de examinar, o índice de heterozigose P, é zero, porque h é zero; isso significa que, nessa população, não há qualquer parcela que n seja pan-mítica, isto é, ela é totalmente formada por indivíduos que se cruzam por autofecundação. O complemento de P, isto é, (1-P), é o coeficiente de endocruzamento F: F =1−P =1− hn h0 − hn = h0 h0 O coeficiente de endocruzamento F pode variar entre 1 (quando hn for igual a zero) e zero (quando hn for igual a h0): h0 − 0 =1 h0 h0 − h0 =0 h0 Podemos interpretar o coeficiente F como sendo um fator de proporcionalidade que divide a população em duas partes, uma fração F, na qual haveria endocruzamento total, e uma fração 1-F, completamente pan-mítica. Assim, poderíamos dizer que uma população AA Aa aa 23 em que existisse um coeficiente de endocruzamento F, constante ao gerações, seria constituída por uma fração F de indivíduos autozigotos AA Aa aa p 0 q longo das e uma fração 1-F constituída por indivíduos alozigotos AA 2 p Aa aa 2 q 2pq Ou então: (1 − F).(p2 + 2 pq + q2 ) + F(p + 0 + q) Se desdobrarmos esses termos e efetuarmos a soma, teremos: p 2 + 2 pq + q 2 − Fp 2 − 2Fpq − Fq 2 + Fp + Fq que, reescrito, dará p 2 − Fp2 + Fp + 2pq − 2Fpq + q 2 − Fq2 + Fq que será igual a: p 2 + Fp(1 − p) + 2pq − 2Fpq + q2 + Fq(1 − q) = = p2 + Fpq + 2 pq − 2Fpq + q 2 + Fpq = p2 + Fpq + 2 pq(1 − F) + q2 + Fpq ou seja: AA Aa aa 2 p +Fpq 2pq(1-F) 2 q +Fpq Essa é a distribuição de freqüências de uma população com coeficiente de endocruzamento F. Essa população está num equilíbrio conhecido pelo nome de equilíbrio de Wright, uma vez que esse tipo de equilíbrio foi demonstrado, pela primeira vez, por esse autor. EXERCÍCIOS: 1)Dadas as populações pan-míticas, com a constituição abaixo, AA Aa aa Pop. 1 356 713 356 Pop. 2 523 698 232 Pop. 3 544 467 100 Pop. 4 40 316 632 quais seriam as suas composições genotípicas se passassem endocruzamento com um coeficiente F = 0,10? e com F=0,25? a 2) Calcule os coeficientes F de endogamia das seguintes populações: um regime de 24 Pop. Pop. Pop. Pop. 1 2 3 4 AA 672 43 243 258 Aa 256 125 298 661 aa 72 182 113 258 Mutações Toda variabilidade genética existente origina-se por mutações, que podem ser induzidas ou espontâneas. Em ambos os casos, podemos considerar que as mutações são recorrentes, ou seja, não são eventos únicos. Seja para sítio de nucleotídeo, aminoácido ou para genes inteiros, a probabilidade de ocorrência de mutação é a chamada taxa. No caso de processos evolutivos, as mutações importantes são aquelas que envolvem a linhagem germinativa, na produção de gametas. No entanto, os modelos vistos a seguir podem ser facilmente modificados para mutações somáticas, tais como aquelas que envolvem a origem de tumores, ou no caso de propagação vegetativa. As taxas de mutação são expressas em termos de proporção de gametas mutantes que aparecem por geração por gene (ou por sítio). O modelo a seguir considera um loco com dois alelos, A e a. Sendo µ a taxa de mutação de A para a, e p0 e q0 as respectivas freqüências gênicas na geração inicial, então podemos escrever: p1 = p0 (1 − µ) p2 = p1 (1 − µ) = p0 (1 − µ)(1− µ) = p0 (1− µ) 2 da mesma forma: p3 = p2 (1− µ) = p0 (1− µ) 2 (1− µ) = p0 (1 − µ)3 pn = p0 (1 − µ)n logo: pn (1 − qn ) = = (1 − µ )n p0 (1 − q0 ) multiplicando µ por n/n e substituindo p por 1-q temos: (1 − q n ) nµ n = (1 − ) (1 − q 0 ) n A função exponencial (ex ) é definida por ex = lim x n 1 + n → ∞ n e se considerarmos um número grande de gerações, (1 − q n ) ≅ e − nµ (1 − q 0 ) tomando os logaritmos naturais (na base e) de ambos os lados da equação: −nµ ≅ ln(1 − qn ) − ln(1− q0 ) Portanto podemos calcular aproximadamente quantas gerações são necessárias para que um determinado gene mude de freqüência com um determinado ∆q, sabendo-se sua freqüência inicial e sua taxa de mutação: 25 n≅ ln(1 − q 0 ) − ln(1 − q n ) µ Mutação reversa É possível que o alelo A mute para a com taxa µ e que o alelo a mute para A com taxa v: µ A →a A ← ν a Assim, em cada geração teremos uma variação nas freqüências dos alelos, que pode ser expressa por: ∆q = µ. p − ν . q Haveria equilíbrio quando ∆q = 0. Neste caso: µp = vq sendo p = 1-q, segue µ(1-q) = νq µ - µq = νq µ = (µ + ν)q Logo, q (em equilíbrio) = q$ = µ µ +ν Note que para este equilíbrio ser alcançado, o número de gerações é muito grande, pois as taxas são muito pequenas. Efeitos das migrações e suas aplicações Abordaremos este assunto primeiramente examinando o modelo proposto por Glass e Li (1953). Sejam: q0 - freqüência original de determinado alelo na população que recebe os migrantes. Q - freqüência do mesmo alelo na população migrante; qn - freqüência do mesmo alelo na população resultante na geração n; m - fração do “pool” gênico que é substituída a cada geração por genes de migrantes, através do inter-cruzamento. Temos, portanto: q1 = (1 − m)q 0 + mQ q 2 = (1 − m) q1 + mQ = (1 − m) 2 q0 + mQ[1 + (1 − m)] q3 = (1 − m)q2 + mQ = (1 − m)3 q0 + mQ[1 + (1 − m) + (1 − m)2 ] ... qn = (1− m) n q0 + mQ[1 + (1− m) + (1 − m)2 ...+ (1− m)n−1 ] Entre colchetes temos uma soma de termos de uma progressão geométrica, onde o n-1 primeiro termo (a1)=1; o último termo (an)=(1-m) e a razão (q)=(1-m) A fórmula geral para a soma dos termos dessa progressão é: 26 n ∑aq i =1 1 i = an × q − a1 q −1 portanto: (1− m) n−1 × (1 − m) − 1 qn = (1− m) n q0 + mQ = (1 − m) − 1 (1− m) n − 1 = (1− m) n q0 + mQ = (1− m) n q0 + Q[1 − (1− m)n ] = − m = (1− m) n q0 + Q − Q(1 − m) n = (1− m)n (q0 − Q) + Q q n − Q = (1 − m) n (q 0 − Q) q n = (1 − m) n (q0 − Q) + Q (1 − m) n = (q n − Q) (q 0 − Q) (q − Q) ln(1− m)n = ln n (q0 − Q) (q − Q) n ln(1 − m) = ln n (q0 − Q) ln e n= (qn − Q) (q0 − Q) ln(1− m) = ln(qn − Q) − ln(q0 − Q) ln(1− m) Exercícios: 1. Uma população tem um alelo em um loco autossômico com freqüência 0,2. Passa a receber uma proporção fixa de 10% de migrantes de outra população cuja freqüência deste mesmo alelo é 0,8. Após 5 gerações, qual será a freqüência deste alelo na população que recebeu os migrantes? Após quantas gerações este alelo terá a freqüência 0,65? E após quantas gerações este valor será 0,78? DERIVA GENÉTICA Seja uma população gerações; sejam ainda p e q 0 0 de tamanho finito N, constante ao longo das as freqüências dos alelos A e a de um loco autossômico na geração 0; como o tamanho da população é constante, a geração 1 é formada da união de 2N gametas ao acaso dentre os indivíduos da geração 0: (p0+q0)2N; q1 pode tomar, portanto, qualquer um dos (2N+1) valores seguintes: 0 1 2 2 N − 2 2N −1 2N ; ; ; ...; ; ; 2N 2N 2 N 2N 2N 2N 27 A probabilidade de que q tome o valor particular qj = j/2N é 2N 2 N − j j 2N .p .q = .(1 − q)2 N − j .q j j j Onde 2N (2N)! = j j!(2N − j)! (combinação de 2N elementos j a j) Seja o seguinte exemplo: N = 2, 2N = 4 genes; p0 = q0 = 1/2 f(A) = p = 1 f(a) = q = 0 estado j = 0 3/4 1/4 1 1/2 1/2 2 1/4 3/4 3 0 1 4 As probabilidades de que a população 1 esteja nos estados j = 0, 1, 2, 3 ou 4 são, respectivamente, 0: 1: 2: 3: 4: (1/2)4 = 1/16 4(1/2)3(1/2) = 1/4 6(1/2)2(1/2)2 = 3/8 4(1/2)(1/2)3 = 1/4 (1/2)4 = 1/16 o que define o vetor da linha Q(1) = (1/16 1/4 3/8 1/4 1/16). Se a população 1 estiver no estado j = 0 (p1 = 1, q1 = 0), o que ocorre com uma probabilidade de 1/16, as probabilidades de que a população 2 esteja nos estados j = 0, 1, 2, 3, 4 são, respectivamente, 0: 1: 2: 3: 4: 1 0 0 0 0. Se a população 1 estiver no estado j=1 (p1 = 3/4, q1 = 1/4), o que ocorre com uma probabilidade de 1/4, as probabilidades de que a população 2 esteja nos estados j = 0, 1, 2, 3, 4 são, respectivamente, 0: 1: 2: 3: 4: (3/4)4 = 81/256 4(3/4)3(1/4) = 27/64 6(3/4)2(1/4)2 = 27/128 4(3/4)(1/4)3 = 3/64 (1/4)4 = 1/256. Se a população 1 estiver no estado j=2 (p1 = q1 = 1/2), o que ocorre com uma probabilidade de 3/8, as probabilidades de que a população 2 esteja nos estados j=0, 1, 2, 3, 4 são respectivamente, 0: 1: 2: 3: 4: (1/2)4 = 1/16 4(1/2)3(1/2) = 1/4 6(1/2)2(1/2)2 = 3/8 4(1/2)(1/2)3 = 1/4 (1/2)4 = 1/16. Se a população 1 estiver no estado j=3 (p1 = 1/4, q1 = 3/4), o que ocorre com uma probabilidade de 1/4, as probabilidades de que a população 2 esteja nos estados j = 0, 1, 2, 3, 4 são, respectivamente, 28 0: 1: 2: 3: 4; (1/4)4 = 1/256 4(1/4)3(3/4) = 3/64 6(1/4)2(3/4)2 = 27/128 4(1/4)(3/4)3 = 27/64 (3/4)4 = 81/256. Se a população 1 estiver no estado j=4 (p1 = 0, q1 = 1), o que ocorre com uma probabilidade de 1/16, as probabilidades de que a população 2 esteja nos estados j=0, 1, 2, 3, 4 são, respectivamente, 0: 1: 2: 3: 4: 0 0 0 0 1. 29 Logo, as probabilidades de que a população 2 esteja nos estados j = 0, 1, 2, 3, 4 são, respectivamente, 0: 1/16 x 1 + 1/4 x 81/256 + 3/8 x 1/16 + 1/4 x 1/256 + 1/16 x 0 = 85/512 = 0,166016 1: 1/16 x 0 + 1/4 x 27/64 + 3/8 x 1/4 + 1/4 x 3/64 + 1/16 x 0 = 27/128 = 0,210938 2: 1/16 x 0 + 1/4 x 27/128 + 3/8 x 3/8 + 1/4 x 27/128 + 1/16 x 0 = 63/256 = 0,246094 3: 1/16 x 0 + 1/4 x 3/64 + 3/8 x 1/4 + 1/4 x 27/64 + 1/16 x 0 = 27/128 = 0,210938 4: 1/16 x 0 + 1/4 x 1/256 + 3/8 x 1/16 + 1/4 x 81/256 + 1/16 x 1 = 85/512 = 0,166016 o que define o vetor de linha Q(2) = (85/512 27/128 63/256 27/128 85/512). Sob forma matricial, as operações podem ser reescritas como 1 16 1 4 3 8 1 4 0 0 0 0 1 81 27 27 3 1 64 128 64 256 1 1256 1 3 1 1 × 4 8 4 16 16 1 16 3 27 27 81 256 64 128 64 256 0 0 0 1 0 [ = 85 512 27 128 63 256 27128 85 512 ] ou, abreviadamente, Q(1).T = Q(2), em que T é uma matriz transicional de probabilidades condicionais (matrizes desse tipo caracterizam-se por suas linhas somarem 1). Generalizando, Q(n).T = Q(n+1). O que foi visto foi a análise de um processo em que, dadas as condições de uma determinada população (tamanho e freqüência), podemos determinar as probabilidades da população estar na mesma condição (freqüências gênicas iguais) ou em condições diferentes. Como a deriva genética é um processo de amostragem casual, não podemos prever o que pode acontecer com a freqüência gênica de uma determinada população pequena. O que pode ser feito, no entanto, é estudar o comportamento de um número muito grande de populações com mesmo tamanho, em que podemos esperar que algumas aumentem as freqüências gênicas, outras diminuam e outras permaneçam com freqüências gênica iguais. A teoria da Estatística nos fornece meios de prever a dispersão das freqüências gênicas em muitas populações. Para isso, usamos a medida da variância, na enésima geração, em um grupo de populações que na geração 0 têm as mesmas freqüências gênicas. A previsão da variância no decorrer das gerações exige conhecimentos avançados de Estatística, mas está representada abaixo apenas para ilustração: Na geração 0, não há variação, todas as freqüências são idênticas, portanto: σ 02 = 0 Na primeira geração (da distribuição binomial): σ 12 = q0 (1 − q0 ) 2N A média das freqüências é igual a esperança: 30 E(q1 ) = q0 = q e a variância: σ 12 = E(q12 ) − q2 = E(q12 ) − q02 q (1 − q0 ) ∴ E(q12 ) = σ 12 + q02 = q02 + 0 2N A proporção de heterozigotos esperada na geração 1 é: h1 = E(2 p1q1 ) = 2E(q1 ) − 2E(q1 2 ) = 2q0 − 2q02 − 2q0 (1 − q0 ) − 2q0 (1 − q0 ) = 2N 2q0 (1− q0 ) 1 = 2q0 (1 − q0 ) 1 − = 2N 2N 1 h0 1 − ∴ 2N como a proporção de heterozigotos de uma geração é a multiplicação da quantidade de heterozigotos da geração seguinte por uma constante, na n-ésima geração: 1 n hn = h0 1 − 2N A variância na n-ésima geração: σ n2 = E(qn2 ) − q02 Colocando o termo E(q2n ) em termos de hn(que já conhecemos) e q0: hn = E(2 pn qn ) = E [2qn (1 − qn )] = 2E(qn ) − 2E(qn2 ) = 2q0 − 2E(qn2 ) E(q2n ) = 2q0 − hn 2 hn 1 n 2 2 σ = q0 − − q0 = q0 − q0 − q0 (1 − q0 ) 1− = 2N 2 2 n 1 n = q0 (1 − q0 ) − q0 (1 − q0 ) 1− = 2n 1 n = q0 (1 − q0 ) 1 − 1− 2N 1 n σ = p0 q0 1− 1 − 2N 2 n O limite de σ 2n quando n tende a infinito é q (1-q ). 0 0 A Tabela abaixo mostra, para um número infinito de populações compostas, cada uma, por N = 2 indivíduos com p = q = 1/2 na geração inicial, os 0 0 31 valores das probabilidades dos estados j = 0, 1, 2, 3, 4 e da variância calculados segundo os métodos mostrados anteriormente. j geração 0 1 2 3 4 5 10 15 20 25 ∞ 0 1 2 3 4 σ 2n 0,000000 0,062500 0,166016 0,248962 0,311670 0,358748 0,466480 0,492046 0,498112 0,499552 0,500000 0,000000 0,250000 0,210938 0,160400 0,120506 0,090399 0,021453 0,005091 0,001208 0,000287 0,000000 1,000000 0,375000 0,246094 0,181274 0,135647 0,101706 0,024124 0,005727 0,001359 0,000323 0,000000 0,000000 0,250000 0,210938 0,160400 0,120506 0,090399 0,021453 0,005091 0,001208 0,000287 0,000000 0,000000 0,062500 0,166016 0,248962 0,311670 0,358748 0,466480 0,492046 0,498112 0,499552 0,500000 0,000000 0,062500 0,109375 0,144531 0,170898 0,190674 0,235922 0,246659 0,249207 0,249812 0,250000 σ 2n , Seleção Natural "I have called this principle, by which each slight variation, if useful, is preserved, by the term Natural Selection, in order to mark its relation to man's power of selection." (Denominei este princípio, pelo qual cada variação diminuta, se útil, é preservada, com o termo Seleção Natural, com a finalidade de salientar sua relação com o poder humano de seleção.) Darwin, The origin of Species, cap III. Quando Darwin estabeleceu o conceito de seleção natural comparou-a com a prática da seleção genética de animais e plantas (domesticação), que vinha sendo realizada com sucesso desde há muito tempo. A seleção artificial era sempre direcionada para o desenvolvimento de características desejáveis pelos seres humanos, chegando a satisfazer até mesmo caprichos bizarros. Em princípio, o que se fazia era escolher os organismos que apresentassem caracteres interessantes para serem os reprodutores. Darwin raciocinou acertadamente que, na natureza, aqueles indivíduos que apresentassem atributos que aumentassem a chance de deixar mais descendentes deixavam mais descendentes. Se estes atributos fossem hereditários e variáveis, os descendentes dos indivíduos "mais aptos" apresentariam, com maior probabilidade, as características de sucesso. Este raciocínio é correto uma vez que as chances de sucesso dependem de fatores extrínsecos aos organismos, os chamados fatores ambientais, que podem ser, inclusive, outros organismos, tais como predadores, parasitas, competidores, etc. A dependência com relação ao ambiente confere significado ao que se conhece como "valor adaptativo". A variação não genética (ou variação ambiental) por não ser herdada, não influencia o valor adaptativo. A potencialidade genética para responder ao ambiente, por ser herdável, também é passível de seleção. O valor adaptativo, além de ser dependente dos fatores ambientais, é de natureza estatística. Um indivíduo com um genótipo que apresenta características vantajosas para uma determinada situação ambiental pode ter insucesso reprodutivo, enquanto outros não geneticamente favorecidos podem deixar proles enormes. Neste caso, diferenças entre valores adaptativos são diferenças entre médias apresentadas pelos diversos indivíduos de cada genótipo. O valor adaptativo, por ser um parâmetro que depende de genótipo, é sempre relativo aos outros genótipos. 32 Cálculo do valor adaptativo. O valor adaptativo é calculado geralmente a partir da divisão dos indivíduos em classes genotípicas com relação a apenas uma fração da variação genética existente, considerando que a variação restante tem efeito igual sobre os genótipos a ser analisados. Exemplificando: se quisermos verificar o efeito sobre o sucesso reprodutivo que a variação em um loco com dois alelos exerce, dividimos os indivíduos em três classes: AA, Aa e aa. Dentro de cada uma destas classes haverá variantes em outros locos (BB, Bb e bb; CC, Cc e cc, etc.), mas como a divisão não considera estes locos, pode-se admitir que eles atuam de forma semelhante sobre o(s) loco(s) cujas classes genotípicas serviram de base para a divisão. Aqui cada classe genotípica pode ser analisada quanto a qualquer componente do valor adaptativo, por exemplo, número de ovos, sementes, taxa de fertilidade, etc., mas a avaliação global do valor adaptativo são as próprias relações entre freqüências de duas gerações consecutivas. Sejam as freqüências genotípicas na geração inicial: AA d0 Aa h0 aa r0 Aa h1 aa r1 e na geração seguinte: AA d1 os valores adaptativos serão: w1(genót. AA)= d1 d0 w2(genót. AA)= h1 h0 . w1(genót. aa)= r1 r0 como são valores relativos, os valores adaptativos podem dividindo-se cada um deles pelo maior, que passará a valer 1. ser normalizados Exemplo: Tomou-se uma população de 1000 indivíduos que foi observada por duas gerações obtendo-se os resultados: AA 250 360 Aa 500 480 aa 250 160 Os valores adaptativos para cada um dos genótipos são, portanto: w1 = 360 =1,44 250 normalizando: w1= 1,44 1,44 = 1,00 w2= 480 500 w2= 0,96 1,44 =0,96 w3= 160 250 = 0,67 = 0,64 w3 = 0,64 1,44 = 0,44 Assim, os indivíduos de genótipo Aa deixam, em média 67% de descendentes com relação aos de genótipo AA e os de genótipo aa deixam apenas 44%, com relação ao mais adaptado (AA). O modelo geral de seleção. 33 O conceito de valor adaptativo nos permite fazer previsões com relação à composição genética de populações naturais, desde que aplicadas a modelos matemáticos adequados. Supondo um sistema de cruzamento ao acaso (pode-se modelar com outros sistemas, sendo que a seqüência de procedimentos é a mesma). Supõe-se também um loco com dois alelos e com valores adaptativos constantes ao longo do tempo. Temos: Genótipos AA Aa aa valores W1 W2 W3 p2 2 pq q p 2 .w1 2 pq.w2 q 2 .w3 Total adaptativos freqüências antes da seleção 2 1 contribuição proporcional freqüências após seleção p 2 .w1 w 2 pq.w2 w q 2 .w3 w w 1 O valor adaptativo médio - w - (Atenção, w NÃO é uma média aritmética simples, é a média ponderada dos valores adaptativos dos genótipos pelas freqüências genotípicas) nos mostra o quanto a população como um todo está adaptada. Por ser uma média ponderada pelas freqüências genotípicas, com valores adaptativos iguais, o valor adaptativo médio pode assumir diversos valores. A quantidade de seleção sofrida por cada um dos genótipos também pode ser expressa através do valor complementar ao do valor adaptativo, o coeficiente de seleção: s1 = 1 - w1 s2 = 1 - w2 s3 = 1 - w3 Casos especiais I. Seleção contra homozigotos recessivos ( w1 = w2 > w3) obs: como só existe um coeficiente de seleção, ele será, neste caso, designado apenas por s. Genótipos AA Aa aa Total valores 1 1 (1 − s) p2 2 pq q2 1 p2 2 pq q 2 (1− s) w = 1 − sq2 adaptativos freqüências antes da seleção contribuição proporcional 34 p2 1 − sq2 freqüências após seleção 2 pq 1 − sq2 q 2 (1− s) 1 − sq2 1 a freqüência do alelo a na próxima geração será (r+h/2): q1 = q 2 − sq 2 + pq q 2 − sq2 + q(1 − q) q2 − sq2 + q − q 2 q − sq 2 = = = 1− sq 2 1− sq2 1− sq2 1 − sq2 A variação da freqüência gênica de a, ∆q será: q − sq2 q − sq 2 − q(1 − sq 2 ) sq2 + sq3 ∆q = q1 − q0 = −q= =− 1 − sq2 1 − sq 2 1 − sq2 sq2 (1 − q) ∆q = − 1 − sq2 (seleção contra homozigotos recessivos) Como s e q são quantidades positivas e menores que 1, q será sempre negativo, ou seja, haverá seleção até a extinção do alelo a. Se a seleção for total (s=1), poderemos prever a freqüência do gene a (q) para a nésima geração: q(1 − q ) q − q2 q q1 = = = 2 1−q (1 + q)(1 − q) 1+ q q2 = q1 , 1+ q1 reaplicando q0 1 + 3q0 e q q 1+q 1+ q q = q = 1 + q + q = 1 + 2q 1+ 1+q 1+ q assim q3 = qn = q0 1+ nq0 (quando há seleção total contra homozigotos recessivos) Se n fica muito grande, q tende a zero, ou seja, a população tende a ficar sem o gene recessivo. 35 II. Seleção favorecendo heterozigotos. (w1<w2>w3) Genótipos AA Aa aa (1 − s1 ) 1 (1 − s3 ) p2 2 pq q2 1 p 2 (1− s1 ) 2 pq q 2 (1− s3 ) w = 1 − s1 p 2 − s3q 2 2 pq w q 2 (1− s3 ) w valores Total adaptativos freqüências antes da seleção contribuição proporcional freqüências após seleção p 2 (1− s1 ) w 1 a freqüência do alelo a na próxima geração será (r+h/2): q 2 − s3 q2 + pq q2 − s3 q2 + q − q2 q − s3q 2 q1 = = = 1− s1 p 2 − s3 q 2 1 − s1 p 2 − s3q 2 1− s1 p2 − s3 q 2 a diferença entre duas gerações consecutivas (∆q) será: q − s3 q2 − q(1 − s1 p2 − s3q 2 ) q − s3 q2 − q + s1 p2 q + s3 q3 ∆q = q1 − q = = = 1 − s1 p2 − s3 q2 1− s1 p2 − s3 q 2 2 s1 p 2 q + s3 q3 − s3 q2 q(s1 p2 + s3 q2 − s3 q) q[ s1 p − qs3 (1 − q)] = = = = 1− s1 p 2 − s3 q 2 1 − s1 p 2 − s3q 2 1 − s1 p2 − s3 q2 = q(s1 p2 − qs3 p) 1− s1 p2 − s3 q 2 ∆q = = pq(s1 p − s3 q) 1− s1 p2 − s3 q 2 (para seleção a favor de heterozigotos) Este valor (∆q) assumirá um valor negativo ou positivo dependendo das freqüências gênicas. Isto significa que existe um valor de equilíbrio onde as freqüência gênicas não mudarão. No equilíbrio, portanto as relações serão constantes: p1 q1 = p q ; substituindo: p − s1 p2 q − s3q 2 = wp wq então: p(1− s1 p) q(1 − s3 q) = wp wq 1 − s1 p = 1 − s3 q ; portanto s1 p = s3 q ; s1 (1− q) = s3 q 36 qˆ = s1 s1 + s3 (somente em equilíbrio, com superioridade do heterozigoto) este é o valor da freqüência genica q, em equilíbrio, significando que esta depende apenas das intensidades dos coeficientes de seleção contra os homozigotos. Equilíbrio entre mutação e seleção Mutações dominantes deletérias são imediatamente eliminadas assim que surgem na formação de heterozigotos. Mas as mutações recessivas se mantêm nos heterozigotos. Para estudarmos o equilíbrio entre mutação recorrente para um gene recessivo deletério, podemos combinar os modelos de seleção contra homozigotos recessivos com o modelo de mutação. A proporção de genes recessivos novos que entra em cada geração por mutação é: µ.(1 − q) A proporção de alelos que é eliminada por seleção contra homozigotos é: sq2 (1 − q) ∆q = − 1 − sq2 Se o coeficiente de seleção for alto, ou seja, se o alelo em homozigose for deletério, este ocorrerá em freqüência baixa, de tal forma que 1-q será um valor próximo de 1. Neste caso, 1-sq 2 será praticamente 1. Podemos escrever então: µ(1 − q) = sq2 (1 − q) µ ) µ = sq 2 ∴ qˆ2 = s No equilíbrio: Podemos estimar, então, no equilíbrio entre mutação e seleção: qˆ = µ s e no caso de s=1 (gene letal): qˆ = µ Estas equações têm sido utilizadas na estimativa indireta de taxas de mutação em organismos diplóides. 37 Exercícios de Genética de Populações 1. Em uma população, as freqüências genotípicas absolutas são as seguintes: AA 100 Aa 300 aa 380 Responda: a)quais são as freqüências gênicas e genotípicas relativas? b)esta população está em equilíbrio de Hardy-Weinberg com relação a este par de alelos? c)qual é a freqüência esperada de cruzamentos AA x aa ? d)dentre os indivíduos aa da próxima geração, que proporção será oriunda apenas de cruzamentos aa x aa? 2. Dada a distribuição de freqüências genotípicas: AA Aa aa d h r para uma população pan-mítica, qual é a freqüência esperada de cruzamentos: a) machos aa x fêmeas AA b) machos aa x fêmeas Aa c) entre indivíduos AA e Aa indique, em cada caso, a freqüência de descendentes machos de genótipo Aa (lembrese que há descendentes machos e fêmeas). 3. Dada a população: AA Aa aa 200 300 500 qual será a distribuição de freqüências genotípicas na geração seguinte, admitindose que os cruzamentos ocorrerão totalmente ao acaso? 4. Em uma população em equilíbrio de Hardy-Weinberg, existem 2 vezes mais homozigotos de um dos tipos que heterozigotos, para um loco autossômico com dois alelos. Quais são as freqüências dos alelos? 5. Temos, em um laboratório, duas populações de uma mesma espécie, com as seguintes composições de um caráter codominante: população I população II AA 36 40 Aa 48 120 aa 16 97 Juntando-se estas duas populações, que deverão se comportar como uma única população pan-mítica, pergunta-se: a)quais são as freqüências gênicas e genotípicas das duas populações quando separadas e quando juntas? b)quais serão as freqüências genotípicas da geração seguinte na população juntada. 6. Das populações abaixo, qual delas não se encontra evidentemente em equilíbrio de Hardy-Weinberg? Por quê? pop. 1 pop. 2 AA 143 340 Aa 632 483 aa 225 177 N 1000 1000 7. De uma população de uma espécie de inseto, foram analisados 1000 indivíduos quanto à pigmentação do tórax. Destes, 40 eram granulados, 640 estriados e 320 uniformes. Considerando equilíbrio de H.W., aponte o genótipo heterozigoto, supondo herança codominante e um par de alelos. 8. Construa 4 populações com 1000 indivíduos com genótipos formados pelos alelos 'A' (freqüência=0,6) e 'a' (freqüência=0,4), que NÃO estejam em equilíbrio de H.W. Demonstre isto com o emprego do teste de qui-quadrado. 9. Demonstre, gráfica ou algebricamente, que uma população com dois alelos não poderá ter freqüência de heterozigotos superior a 0,5, se em equilíbrio de H.W. 38 10. Um sistema genético é constituido por 4 alelos autossômicos codominantes, designados pelas letras A,B,C e D. Se uma população é pan-mítica e as freqüências dos alelos são respectivamente 0,1; 0,2; 0,3 e 0,4 quais são as freqüências genotípicas esperadas? 11. Em uma população de Prochilodus scrofa (curimbatá), Galhardo (1989), estudando um polimorfismo de transferrinas do plasma sanguíneo, encontrou 5 alelos (TfA, TfB, TfC, TfD e TfE), assim distribuídos nos genótipos: genótipo Tf N. indivíduos AA AB AC 22 19 17 AD AE BB 13 12 29 BC BD BE 25 29 12 CC CD CE 34 22 25 DD DE EE 29 23 28 Calcule: a)as freqüências genotípicas relativas de cada genótipo b)as freqüências absolutas esperadas c)o número de graus de liberdade para um teste de qui-quadrado 12. Com n alelos em um loco autossômico, quantos genótipos homozigotos são possíveis? E heterozigotos? 13. Que proporção de crianças MN tem mães MN? 14. Que proporção de pessoas MM tem ambos os progenitores MM? 15. Numa população em equilíbrio de H.W. existem 10 vezes mais genótipos MN que NN. Qual é a freqüência do alelo N? 16. Um indivíduo é falsamente acusado de haver cometido um roubo. O verdadeiro ladrão, ao arrombar o cofre, machucou a mão. Isso permitiu determinar que o ladrão pertencia aos grupos sanguíneos M do sistema MN e O (genótipo ii) do sistema ABO. Qual é a chance de que, uma vez determinados os grupos sanguíneos MN e ABO do indivíduo falsamente acusado, ele seja excluído dessa acusação? Sabe-se que a freqüência do gene M é 0,55 e a freqüência do gene i é 0,65. 17. Qual a probabilidade de uma mãe Rh- ter um filho Rh+? 18. Em algumas variedades de carneiros, a presença de chifres é determinada por um alelo que é dominante nos machos e recessivo nas fêmeas. Se 96% dos machos têm chifres, qual é a proporção de fêmeas que os apresentam? 19. Usando o programa SUPERPOP (exercícios 19-23), na opção de genes ligados ao sexo, inicie algumas populações com freqüências gênicas iguais e diferentes para os dois sexos - pf igual a pm ,e pf diferente de pm , onde pf é a freqüência do gene A nas fêmeas e pm é a freqüência do mesmo alelo nos machos. Simule pelo menos 2 situações para freqüências iguais e 3 para diferentes. Tente obter também uma situação na qual as freqüências iniciem com valores diferentes para machos e para fêmeas e que se obtenha no final pf =pm =0,4. Tente novamente, só que com freqüência final pf =pm =0,8. Em seguida, procure responder as seguintes questões: 20. Considerando a geração genérica t (em relação à geração t-1), qual é a freqüência pf (das fêmeas)? 21. Qual é a freqüência pm (dos machos)? 22. Este modelo é válido se a razão sexual for diferente de 1:1? Por quê? 23. Para as freqüências gênicas iniciais pf0 =0,80 e pm0 =0,20, calcule as freqüências para machos e fêmeas durante 6 gerações de cruzamentos ao acaso. 39 24. Na Drosophila melanogaster, a característica Bar (olho com um número reduzido de omatídeos) é condicionada por um gene b (na verdade uma duplicação gênica), situado no cromossomo X. As fêmeas heterozigotas Bb apresentam o fenótipo “reniforme”. Uma população é constituída por: 320 fêmeas com olho normal 236 fêmeas com olho reniforme 44 fêmeas com olho Bar 174 machos com olho normal 126 machos com olho Bar Calcule, admitindo pan-mixia, nesta geração e na geração seguinte as freqüências gênicas: a) entre machos e fêmeas separadamente b) na população total Verifique se a população encontra-se em equilíbrio para genes ligados ao sexo. 25. Numa amostragem de 150 homens e 300 mulheres, Tönz e Rossi (1964) verificaram a seguinte distribuição de genótipos quanto à deficiência de G6PD (enzima codificada por um gene ligado ao cromossomo X): Genótipo N A X Y 137 a X Y 13 A A X X 247 A a X X 50 a a X X 3 a Estime a freqüência do alelo X na população masculina, na população feminina e na população total. Verifique se a distribuição dos genótipos está em equilíbrio para genes ligados ao sexo. 26. Usando o programa SUPERPOP, verifique os casos 1,2 e 3 de cruzamentos preferenciais, usando valores diferentes de freqüências genotípicas iniciais (cruzamento preferencial totalmente positivo, com dominância - caso 1 - cruzamento preferencial totalmente positivo sem dominância - caso 2 - e cruzamento preferencial totalmente negativo sem dominância - caso 3). Para cada um deles, responda: a. Existe mudança nas freqüências gênicas? b. Quais são os valores de equilíbrio (para freqüências genotípicas) c. Em que os valores de freqüências genotípicas iniciais influem nas freqüências genotípicas de equilíbrio? 27. Baseado nos resultados obtidos no exercício anterior, com relação aos resultados do caso 3, responda: É estável um sistema de três sexos, cada um determinado por um dos genótipos possíveis de um loco com dois alelos, onde cada sexo só pode se cruzar com qualquer dos outros dois? 28. Verifique o que acontece com as freqüências genotípicas na população seguinte durante 4 gerações de autofertilização: AA Aa aa 0,1225 0,455 0,4255 29. Qual seria o coeficiente F de endogamia se ele fosse estimado após a segunda geração de autofertilização da população acima? 30. Uma população tem dois alelos segregando em um loco, A e a. A freqüência do alelo A é 0,46. O coeficiente de endocruzamento é 0,3. Calcule, para 1200 indivíduos, as freqüências genotípicas absolutas esperadas, no equilíbrio de Wright. Faça o teste de equilíbrio de Hardy-Weinberg. 31. Quantas gerações são necessárias para que a freqüência de um genótipo recessivo passe de 1/10.000 para 1/1.000 se a taxa de mutação para a formação deste gene recessivo é de 10-6 ? (não considerar mutação reversa) 40 32. Um biólogo analisa três populações de uma espécie de peixe ao longo de um rio, separadas por barragens. A população A está rio acima e não recebe migrantes. A população B é intermediária e recebe migrantes da população C que está rio abaixo mas não da população A. A população C apenas fornece migrantes para a população B. A freqüência de um alelo é 0,32 na população A, 0,65 na população B e 0,80 na população C. As populações A e B formavam uma única população antes da construção das barragens e a população C passou a fornecer migrantes para a população B depois da construção das barragens, em 1960. Este peixe tem uma única geração por ano. Imaginando que a única fonte de alteração de freqüência deste gene se deve à migração, responda: a) Qual é a taxa de migração anual que a população B recebe? b) Sabendo-se que os tamanhos das populações são constantes e que a população B recebe 14 migrantes por ano, qual é o tamanho da população B? c) Se a taxa de migração que a população B recebe fosse de 4% ao ano, qual seria a freqüência gênica atual? 33. Uma população é composta por dois indivíduos heterozigotos (Aa). A cada geração, a população é substituída por uma progênie também de dois indivíduos, um macho e uma fêmea. Pergunta-se: a) Qual é a freqüência gênica do alelo A na população inicial? b) Quais são as probabilidades de, na geração seguinte, a população possuir freqüências do alelo A igual a 1, 3/4, 1/2, 1/4 e 0? c) Qual é a chance da população na geração 1 ter freqüência gênica exatamente igual à da geração 0? d) Qual é a chance da população, na geração 1 alterar a freqüência gênica em relação à existente na geração 0? e) Se a população na geração 0 fosse composta por 8 indivíduos heterozigotos Aa e o tamanho da população fosse constante ao longo das gerações, qual é a chance de ocorrer alteração na freqüência gênica na geração 1? 34. Spiess e colaboradores estudaram genótipos de Drosophila persimilis e verificaram a sobrevivência relativa de cada um deles, obtendo os seguintes resultados: Genótipo Sobrevivência Em baixa densidade Em alta densidade 37,3% 41,0% 48,3% 47,8% 50,0% 38,4% WT/WT WT/KL KL/KL Baseado nisto, responda: a) quais são os valores adaptativos e os coeficientes de seleção nas duas condições ambientais? b) Quais serão as freqüências dos alelos na condição de equilíbrio nas duas condições ambientais? 35. Uma população, na geração 0, ao nascimento, tem a seguinte composição genotípica: AA 0,16 Aa 0,48 aa 0,36 Para cada uma das combinações de valores adaptativos abaixo, calcule as freqüências genotípicas ao nascimento (ou seja, após a população sofrer seleção e se reproduzir, com cruzamentos ao acaso), na geração seguinte e após um número muito grande de gerações (tendendo ao infinito). caso 1 caso 2 caso 3 AA 1 1 0,3 Aa 1 0,8 1 aa 0,5 0,5 0,8 36. Uma população possui um alelo recessivo que é letal em homozigose. A freqüência ao nascimento dos genótipos homozigotos recessivos é 1/160.000. Supondo-se equilíbrio entre mutação e seleção, qual é a taxa de mutação para o alelo recessivo? Consegue-se uma droga que cura parcialmente a doença provocada pelo genótipo homozigoto. Com esta droga, a letalidade passa a ser de 50%. No equilíbrio, quais serão as novas freqüências gênicas e genotípicas? 41 Projeto 1. Simulação de deriva genética. Utilizando o programa SUPERPOP, seu grupo deverá simular o processo de deriva genética. Como se trata de um fenômeno em que processos meramente casuais atuam em populações biológicas, não é possível fazer previsões a respeito do resultado em termos de direção (por exemplo, não é possível saber se uma freqüência gênica aumentará ou diminuirá). Entretanto, a magnitude da variação pode ser prevista a partir da variância teórica esperada: σe2 t 1 = p0 q0 1 − 1 − 2N p 2 onde δ e é a variância esperada, p0 e q0 são as freqüências gênicas iniciais, Np é o tamanho da população e t é o número de gerações. 2 Temos também a equação de estimativa da variância (δ o , variância observada ou experimental) a partir de dados observados que é: N ∑( p − p ) 2 i σ o2 = i =1 Na esta fórmula é algebricamente equivalente a: σ o2 Na pi Na ∑ 2 i=1 pi − ∑ Na = i =1 Na 2 , que é mais fácil de usar com uma calculadora, onde pi é a i-ésima freqüência após t gerações, p é a média das freqüências observadas após t gerações e Na é o número de freqüências abservadas após t gerações. O objetivo deste projeto é comparar as variâncias observadas e as variâncias teóricas após 15 gerações em populações com tamanhos diferentes: 20, 50, 100, 200 e 500 indivíduos. O programa ilustra graficamente a variação da freqüência de um dos alelos durante todas as gerações, mas os dados a serem utilizados serão apenas os da geração 15. Utilize sempre a mesma freqüência p inicial para todas as simulações (entre 0,33 e 0,66). A comparação entre as variâncias experimentais e as variâncias teóricas pode ser feita com o teste da razão, onde se estima o parâmetro F que é comparado com o valor crítico: F= σo σ 2e 2 O F crítico é obtido de uma tabela de valores F críticos, com nível de significância (alfa) de 0,5; bicaudal, com 10 graus de liberdade no numerador e infinitos graus de liberdade no denominador: Fcritico0,05( 2)10,× = 2,05 42 Tamanho N=20 F= N=50 F= N=100 F= N=200 F= N=500 F= freqüências obtidas p inical= p= p1= p6= p2= p7= p3= p8= σ 2e = p4= p9= F = p5= p10 = σ 2o = p1= p6= p= p2= p7= p3= p8= σ 2e = p4= p9= F = p5= p10 = σ 2o = p1= p6= p= p2= p7= p3= p8= σ 2e = p4= p9= F = p5= p10 = σ 2o = p1= p6= p= p2= p7= p3= p8= σ 2e = p4= p9= F = p5= p10 = σ 2o = p1= p6= p= p2= p7= p3= p8= σ 2e = p4= p9= F = p5= p10 = σ 2o = Com os dados obtidos com o emprego do programa SUPERPOP, preencha a tabela acima e responda: 1. A evolução observada através do processo de deriva genética é também chamada de evolução não darwiniana. Por quê? 2. A variabilidade genética é uma medida que reflete a quantidade de variação genética em um determinado grupo de organismos. Se neste grupo existem muitos alelos com freqüência alta, há muita variabilidade genética. Numa população grande que se fragmenta em muitas sub-populações pequenas, o que ocorre, depois de muito tempo na variabilidade genética: a) em cada uma das subpopulações. b) na população como um todo c) justifique. 43 44 Projeto 2. Simulação de seleção natural. A seleção natural é considerada como o único processo modulador da evolução. Ela atua sempre no sentido de levar as populações a um estado de maior adaptabilidade. Dentro dos processos conhecidos que atuam na evolução dos organismos, a seleção natural pode ter tanto a sua intensidade como sua direção estabelecidas. A seleção natural é um processo estatístico, ou seja, ela atua quando existem diferenças de probabilidades de que determinados genótipos ou fenótipos deixem descendência viável. É importante salientar que a intensidade da atuação da seleção natural é relativa, ou seja, um determinado genótipo pode ser bem adaptado com relação a outros genótipos, mas pode não sê-lo se comparado a um genótipo até então não existente. Um modelo de seleção que pode ser utilizado para uma introdução ao assunto é o elaborado por Sewall Wright e que analisa os efeitos de valores adaptativos (ou coeficientes de seleção) para cada um dos três genótipos de um loco autossômico com dois alelos. O programa SUPERPOP para micros simulará a mudança nas freqüências gênicas em vários casos deste modelo. PROCEDIMENTOS: O programa precisa dos seguintes dados: a) a freqüência inicial do alelo A. Comece sempre com um valor alto (ex. 0.9). Isto simularia uma situação na qual este alelo estaria quase monomórfico e o alelo a surgisse por migração. Use também, para cada caso, valores iniciais diferentes. b) o coeficiente de seleção para cada um dos três genótipos. obs: como se trata de um valor relativo, um dos genótipos deverá apresentar o coeficiente de seleção igual a zero. c) o número de gerações. situação As seguintes situações deverão ser simuladas: valores adaptativos coeficientes de classificação seleção (complete) 1 wAA = WAa > Waa sAA = sAa < saa wAA = WAa < Waa sAA = sAa > saa wAA < WAa = Waa sAA > sAa = saa wAA > WAa = Waa sAA < sAa = saa seleção contra homozigotos recessivos 2 3 4 5 wAA > WAa > Waa sAA < sAa < saa wAA < WAa < Waa sAA > sAa > saa seleção contra homozigotos e heterozigotos 6 7 wAA < WAa > Waa sAA > sAa < saa wAA > WAa < Waa sAA < sAa > saa 8 seleção favorecendo heterozigotos 45 As situações de número 7 e 8 deverão ser repetidas várias vezes com os mesmos valores, modificando-se apenas a freqüência gênica inicial. Questões: 1. Qual é o comportamento analisadas? Por quê? do valor adaptativo médio em todas as situações 2. Compare todas as situações salientando as diferenças e semelhanças em: a) Há tendência de extinção de um alelo? Qual? b) O processo é mais rápido ou mais lento? Atenção: para poder responder esta questão, é importante que se utilize coeficientes de seleção de mesma ordem de magnitude. c) Há apenas equilíbrio trivial (quando há extinção de um alelo) ou há algum tipo de equilíbrio não trivial (quando os dois alelo se mantêm em certas freqüências)? 3. Utilize o programa SUPERPOP para responder as questões 34 e 35 da lista de exercícios.
Baixar