☰
Explorar
Assinar em
Inscrever-se
Envio
×
Baixar
Sem categoria
taxocenose de serpentes da ilha de marajó, estado do pará
Belém, 26 de novembro de 2015. LISTA DOS CANDIDATOS PRÉ
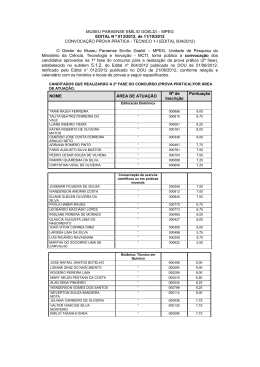
MUSEU PARAENSE EMÍLIO GOELDI – MPEG EDITAL N º 013/2012
Orientação para o seminário.
MPEG - DCC
Retificação Mestrado - Museu Paraense Emílio Goeldi
Descarregar - Receptores digitales
PRA Apresentacao Hangar
Slides
Termo de Autorização para inserir Tese e Dissertação na
Chamada 01/2015 - Museu Paraense Emílio Goeldi