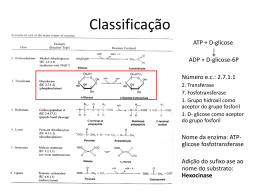
UNIVERSIDADE ESTADUAL DE FEIRA DE i e m q u a l q DEPARTAMENTO DE CIÊNCIAS BIOLÓGICAS u PROGRAMA DE PÓS-GRADUAÇÃO EM RECURSOS GENÉTICOSe r l VEGETAIS u g a r d o d o c u ALINE SILVA COSTA m e n t o . U DETERMINAÇÃO DA ATIVIDADE ENZIMÁTICA E s ANTIOXIDANTE DE VARIEDADES COMERCIAIS DE Ananas e a comosus E AVALIAÇÃO DO EFEITO DE TRATAMENTO TÉRMICOg u E DO USO DE ADITIVOS QUÍMICOS i a F e r r a m e n t a s d e FEIRA DE SANTANA-BA C a 2011 i x a d e T SANTANA ALINE SILVA COSTA DETERMINAÇÃO DA ATIVIDADE ENZIMÁTICA E ANTIOXIDANTE DE VARIEDADES COMERCIAIS DE Ananas comosus E AVALIAÇÃO DO EFEITO DE TRATAMENTO TÉRMICO E DO USO DE ADITIVOS QUÍMICOS FEIRA DE SANTANA-BA 2011 ii e m q u a l q u e r l u g a r d o d o c u m e n t o . U s e a g u i a F e r r a m e n t a s d e C a i x a d e T ALINE SILVA COSTA iii e m q u a l q u e r l u g DETERMINAÇÃO DA ATIVIDADE ENZIMÁTICA E a r ANTIOXIDANTE DE VARIEDADES COMERCIAIS DE d o d Ananas comosus E AVALIAÇÃO DO EFEITO DE o TRATAMENTO TÉRMICO E DO USO DE ADITIVOS c u m QUÍMICOS e n t o . U Dissertação apresentada ao Programa de Pós-Graduação em Recursos s Genéticos Vegetais, da Universidade Estadual de Feira de Santana, como requisito parcial para a obtenção do título de Mestre eem Recursos Genéticos Vegetais. a g u Orientador(a): Prof.ª Dr.ª Maria Gabriela Bello Koblitz i Co-orientador: Prof.º Dr.º Hugo Neves Brandão a Co-orientador(a): Prof.ª Dr.ª Sandra Aparecida de Assis F e r r a m e n t a s d e FEIRA DE SANTANA-BA C a 2011 i x a d e T iv Ficha Catalográfica – Biblioteca Central Julieta Carteado C87d Costa, Aline Silva Determinação da atividade enzimática e antioxidante de variedades comerciais de Ananas comosus e avaliação do efeito de tratamento térmico e do uso de aditivos químicos/ Aline Silva Costa. – Feira de Santana, 2011. 85 f : il. Orientadora: Maria Gabriela Bello Koblitz Co-Orientadores: Hugo Neves Brandão Sandra Aparecida de Assis Dissertação (mestrado) – Universidade Estadual de Feira de Santana. Programa de Pós-Graduação em Recursos Genéticos Vegetais., 2011. 1.Abacaxi – Característica bioquímica. 2.Poligalacturonases (PG). 3.Pectinametilesterases (PME). 4.Polifenol-oxidases (PFO) 5.Peroxidases (PER) 6.Compostos fenólicos. I.Koblitz, Maria Gabriela Bello. II.Brandão, Hugo Neves. III.Assis, Sandra Aparecida de. IV.Universidade Estadual de Feira de Santana. V. Título. CDU: 634.774 v BANCA EXAMINADORA __________________________________________ Prof. Dr. __________________________________________ Prof. Dr. __________________________________________ Prof.ª Dr.ª Maria Gabriela Bello Koblitz Orientador(a) e Presidente da Banca Feira de Santana – BA 2011 vi AGRADECIMENTOS Á Deus, pela oportunidade cedida a mim, conduzindo todos os meus passos e permitindo vivenciar experiências inesquecíveis para meu crescimento. Aos meus pais, Elias e Lene, que sempre me apoiaram e acreditaram na minha capacidade... Meu eterno muito obrigada, por tudo que vocês são e pelo que eu sou. À Prof.ª Dr.ª Maria Gabriela Bello Koblitz, pela orientação, compreensão e incentivo e mesmo distante se fez tão presente. Ao Prof. Dr. Hugo Neves Brandão, pela ajuda, paciência e colaboração. À professora Dr.ª Sandra Aparecida de Assis, pelas contribuições neste trabalho. À Prof.ª Marília Lordêlo Cardoso Silva, pela participação neste trabalho com prestimosas sugestões, por ter disponibilizado o laboratório de analise físico-química de alimentos e pela amizade. À Prof.ª Taís Oliveira Brandão, pela ajuda no teste de compostos fenólicos. Aos professores do ALI por ter disponibilizado os laboratórios e equipamentos. À Cintia Reis, pela disponibilidade em ajudar e pela amizade. À Danielle Figuerêdo, pela ajuda no teste DPPH. Às minhas IC‟s, Isabela e minha querida irmã Larissa, pelo trabalho prestado, companheirismo e amizade. Aos amigos de sempre, Verônica Santos e Andson Rocha pelos bons momentos de convivência, que serão lembrados com muito carinho, e pelo apoio para seguir sempre com coerência. Aos meus tios Maria e Dore, e a toda minha família, irmãos, cunhada, tios e primos pelo incentivo e torcida. A André Luiz, por ter sido tão paciente e compreensivo nesta etapa final. À FAPESB pela concessão da bolsa e suporte financeiro. À EMBRAPA Mandioca e Fruticultura, a quem direciono meus agradecimentos à Fernanda Vidigal e Eliseth Viana. Enfim, a todas as pessoas não mencionadas, que direta ou indiretamente contribuíram para a realização desse trabalho. Muito Obrigada! vii RESUMO O abacaxizeiro constitui uma das fruteiras tropicais mais cultivadas no país e o seu fruto é consumido ao natural, ou na forma de diversos produtos industrializados. Porém, poucas variedades de abacaxi são destinadas à industrialização por falta de conhecimento das suas características bioquímicas, dentre elas a atividade das enzimas pectinolíticas e oxidativas que estão entre aquelas que reduzem a vida de prateleira de produtos a base de frutas. Além disso, apesar do crescente interesse a respeito da atividade antioxidante em produtos de frutas, pouco se sabe sobre o efeito do processamento sobre essa atividade, especialmente em derivados de abacaxi. Este trabalho teve como intuito estudar a atividade das enzimas poligalacturonases (PG), pectinametilesterases (PME), polifenol-oxidases (PFO) e peroxidases (PER), e formas de inativá-las pela adição de inibidores químicos (ácido ascórbico, metabissulfito de sódio e cloreto de cálcio), e físicos (tempo e temperatura de tratamento) de variedades comerciais de Ananas comosus. Estudou-se também o teor de fenóis totais e atividade antioxidante no abacaxi e a possível alteração desta atividade depois dos tratamentos para inativação enzimática aplicados. Para obtenção da máxima atividade enzimática foi feito um planejamento experimental analisando as variáveis pH e concentração de NaCl no método da extração enzimática. Os resultados mostraram que as maiores atividades obtidas nesse experimento foram: 4752,9 U/g para PER; 81,6 U/g para PFO; 7961,3 U/g para PME e 0,000036 U/g h-1 para PG. Foram ainda determinadas as condições mais indicadas para extração das enzimas a saber: pH=6,0 e concentração de NaCl de 1,0 M para PER; pH inferior a 4,6 para PFO; concentração de NaCl de 1,0 M para PME e concentração de NaCl de 2,0 M para PG. Utilizando estes parâmetros foi determinada a atividade enzimática nas variedades Imperial, Vitória, e Pérola adquiridas em dois locais distintos. Observaram-se maiores atividades das enzimas pectinolíticas nas variedades Imperial e Vitória, no entanto estas variedades apresentaram menor atividades das enzimas oxidativas, característica mais vantajosa para a indústria de alimentos, pois a PER é a enzima mais termorresistente (difícil inativação). Utilizou-se o delineamento composto central e a metodologia de superfície de resposta para estudar o efeito do tempo (1 a 9 min), da temperatura (27 a 86 °C) e da concentração do inibidor químico (0 a 200 mg/L de metabissulfito de sódio, ácido ascórbico e cloreto de cálcio) na inativação das enzimas PER e PFO. O metabissulfito de sódio mostrou-se o melhor inibidor para PER, sendo capaz de garantir 100% de inativação com uma concentração de viii 130 mg/L, a temperatura de 78 °C, por 3 min, evitando danos de altas dosagens do inibidor e altas temperaturas no fruto. Para inativação de PFO o tratamento indicado foi com ácido ascórbico a 190 mg/L por 3 min a 75° C, porém este tratamento não foi eficiente para inativar 100% a enzima PER. Estes tratamentos tiveram efeito positivo na quantidade de compostos fenólicos totais e na atividade antioxidante, possivelmente pelo fato destes tratamentos inativarem as enzimas que degradam os compostos fenólicos (PFO e PER), e foi observada uma correlação de moderada a alta entre o teor de fenóis totais e atividade antioxidante. Palavras-chave: abacaxi, poligalacturonases (PG), pectinametilesterases (PME), polifenol-oxidases (PFO), peroxidases (PER), compostos fenólicos. ix ABSTRACT The pineapple is a tropical fruit orchard in the country most cultivated and its fruit is eaten raw, or in the form of ice cream, candy, popsicles, juices processed, fruit in syrup and so on. However, few varieties are used in industry for lack of knowledge of their biochemical characteristics, among them the pectinolytic enzymes activity and oxidative that are among those that decrease the shelf life of products made from fruits. Moreover, despite the growing concern regarding the antioxidant activity in fruit products, little is known about the effect of processing on this activity, especially in derivatives of pineapple. Therefore, the objective of the study to determine the activity of enzymes polygalacturonase (PG), pectin methylesterase (PME), polyphenol oxidase (PPO) and peroxidase (PER), and ways to inactivate them by the addition of chemical inhibitors (ascorbic acid, metabisulfite sodium and calcium chloride) and physical (temperature and time of treatment) of commercial varieties of Ananas comosus. Furthermore, studied the total phenolic content and antioxidant activity in the pineapple and possible amendment of this activity after the treatments applied to enzymatic inactivation. To obtain maximum enzyme activity was made an experimental design by analyzing the pH and NaCl concentration on enzymatic extraction method. The results showed that the highest activities obtained in this experiment were: 1.0 M NaCl, pH 6.0 for PER (4752.9 U / g), pH < 4.6 for PPO (81.6 U / g)1.0 M NaCl for PME (5.8 U / g) and 2.0 M NaCl for PG (0,000034 U/g.h-1) . Using these parameters was determined the enzymatic activity of varieties Imperial, Vitoria, and Perola acquired from two different places. Observed higher activities of pectic enzymes in Variety Imperial and Victoria, but these varieties showed lower activity of oxidative enzymes characteristic most advantageous to the food industry, because the PER is the most thermoresistant enzyme (inactivation difficult). We used the central composite design and response surface methodology to study the effect of time (1-9 min), temperature (27 to 86 ° C) and concentration of the chemical inhibitor (0 to 200 mg / L of sodium metabisulphite, ascorbic acid and calcium chloride) in the inactivation of the PER and PPO. Sodium metabisulphite was found to be the best inhibitor for PER, 100% could inactivate this enzyme at a concentration of 130 mg / L at a temperature of 78 ° C for 3 min damage from high doses of the inhibitor and high temperatures on fruit . For PPO treatment with ascorbic acid was given at 190 mg / L for 3 min at 75 ° C, although this treatment was not effective to inactivate the enzyme 100% PER. These treatments had a x positive effect on the amount of total phenolics and antioxidant activity, possibly because these treatments inactivate enzymes that degrade the phenolic compounds (PPO and PER), and observed a moderate to high correlation between the total phenols and antioxidant activity. Key-words: pineapple, polygalacturonase (PG), pectin methylesterase (PME), polyphenol oxidase (PPO), peroxidase (PER), phenolic compounds. xi SUMÁRIO 1.0 Introdução ....................................................................................................................1 2.0 Revisão da Literatura ...................................................................................................3 2.1 Abacaxi ........................................................................................................................3 2.1.1 Características gerais .............................................................................................3 2.1.2 Melhoramento Genético do abacaxi .......................................................................5 2.2 Enzimas vegetais de importância na indústria de alimentos ..........................................7 2.2.1 Enzimas Oxidativas ...............................................................................................8 2.2.2 Enzimas pectinolíticas ........................................................................................ 12 2.3 Inativação enzimática ................................................................................................. 15 2.4 Atividade antioxidante e Compostos Fenólicos........................................................... 17 2.5 Otimização de processos e delineamento experimental ............................................... 19 3.0 Material e Métodos .................................................................................................... 20 3.1 Experimento 1. Planejamento experimental e otimização do processo de extração enzimática. ....................................................................................................................... 20 3.1.1 Material vegetal ................................................................................................... 20 3.1.2 Planejamento experimental .................................................................................. 20 3.1.3 Determinação da atividade enzimática ................................................................. 21 3.1.4 Análise Estatística ............................................................................................... 22 3.2 Experimento 2. Determinação da atividade enzimática em diferentes variedades de abacaxi. ............................................................................................................................ 23 3.2.1 Material Vegetal .................................................................................................. 23 3.2.2 Determinação da atividade enzimática ................................................................. 23 3.2.3 Análise estatística ................................................................................................ 23 3.3 Experimento 3. Otimização da inativação das enzimas PER e PFO ............................ 24 3.3.1 Material Vegetal .................................................................................................. 24 3.3.2 Planejamento experimental .................................................................................. 24 3.3.3 Determinação da inativação de PER e PFO .......................................................... 25 3.3.4 Análise estatística ............................................................................................... 25 3.3.5 Confirmação dos resultados ................................................................................. 25 3.4 Experimento 4. Determinação da atividade antioxidante e compostos fenólicos totais 25 3.4.1 Material Vegetal .................................................................................................. 25 3.4.2 Extração dos compostos antioxidantes e fenólicos ............................................... 26 3.4.3 Atividade antioxidante ......................................................................................... 26 3.4.4 Determinação do teor de fenóis totais .................................................................. 27 3.4.5 Análise Estatística ............................................................................................... 27 4.0 Resultados e Discussão .............................................................................................. 27 4.1 Experimento 01. Planejamento experimental e otimização do processo de extração enzimática ........................................................................................................................ 27 4.1.1 Peroxidase (PER) ................................................................................................ 29 4.1.2 Polifenol-oxidase (PFO) ...................................................................................... 31 4.1.3 Pectinametilesterase (PME) ................................................................................. 32 4.1.4 Poligalacturonase (PG) ........................................................................................ 33 4.2 Experimento 2. Determinação da atividade enzimática nas variedades de abacaxis .... 35 4.2.1 Peroxidase (PER) ................................................................................................ 35 4.2.2 Polifenol-oxidase (PFO) ...................................................................................... 36 4.2.3 Pectinametilesterase (PME) ................................................................................. 38 xii 4.2.4 Poligalacturonase (PG) ........................................................................................ 39 4.3 Experimento 3. Otimização da inativação enzimática de PFO e PER .......................... 41 4.3.1 Peroxidase (PER) ............................................................................................... 44 4.3.2 Polifenol-oxidase (PFO) ...................................................................................... 47 4.3.3 Confirmação dos resultados ................................................................................. 51 4.4 Experimento 4. Compostos fenólicos e atividade antioxidante .................................... 52 4.4.1 Compostos fenólicos totais .................................................................................. 52 4.4.2 Atividade Antioxidante ........................................................................................ 53 5.0 Conclusão .................................................................................................................. 55 6.0 Referências ................................................................................................................ 57 7.0 Anexos ....................................................................................................................... 75 1 1.0 Introdução O abacaxi ou ananás, nomes utilizados tanto para a fruta como para a planta, pertence à família Bromeliaceae e gênero Ananas Mill. Esse gênero é vastamente distribuído nas regiões tropicais por intermédio da espécie Ananas comosus (L.) Merril, que abrange todas as cultivares plantadas de abacaxi (GIACOMELLI, 1981). O abacaxi é largamente consumido in natura e utilizado na indústria para obtenção de geléias, doces em calda, produtos minimamente processados e sucos, entre outros e seus nutrientes exercem funções importantes no organismo humano (GRANADA et al., 2004). Com relação ao abacaxi como produto processado, é importante ressaltar que o interesse pelos produtos “prontos para o consumo” a base de frutas vem crescendo (VIDAL & GASPARETO, 2000). Mas apenas algumas variedades de abacaxi apresentam atualmente aproveitamento industrial, principalmente pela falta de conhecimentos das suas características bioquímicas. A conservação de polpas e sucos de frutas depende basicamente de se evitar eficientemente a contaminação microbiana e de se contornar os efeitos deletérios causados pela ação das enzimas endógenas dos frutos. Os grupos de enzimas mais danosos em polpas e sucos de frutas são as enzimas pectinolíticas e as enzimas oxidativas (TOIVONEN & BRUMMELL, 2007; MARTINEZ & WHITAKER, 1995). No primeiro grupo encontram-se a pectinametilesterase (PME) e a poligalacturonase (PG): essas enzimas são capazes de hidrolizar a pectina presente na polpa reduzindo sua viscosidade e retirando a turbidez. Conhecendo a atividade das enzimas pectinolíticas, além de algumas de suas características bioquímicas e sabendo da possibilidade de algum dano à polpa ou suco armazenado, pode-se indicar qual a melhor estratégia para evitar esse dano (LEA, 1995; URLAUB, 2002). Entre as enzimas oxidativas, as mais importantes são as polifenol-oxidases (PFO) e as peroxidases (PER). O controle destas enzimas é de grande importância para a tecnologia de alimentos, uma vez que estas são responsáveis pelo escurecimento em frutas e vegetais e seus produtos processados. Além disso, estas enzimas podem participar de um grande número de reações oxidativas e de biodegradação, tais como mudança de cor, degradação da clorofila ou auxinas, oxidação de fenóis, oxidação do ácido indol acético, biossíntese da lignina, e muitos destes fatores também podem ser associados com sabor, aroma cor, textura e 2 qualidade nutricional dos alimentos (BURNETTE, 1977; CLEMENTE, 1995; VÁMOSVIGYÁZÓ, 1981). Os frutos contêm, além dos nutrientes essenciais e de micronutrientes como minerais, fibras e vitaminas, diversos compostos secundários de natureza fenólica, denominados polifenóis que possuem ação antioxidante (KIM et al., 2007). Diversos estudos têm demonstrado que o consumo de substâncias antioxidantes na dieta diária pode produzir uma ação protetora efetiva contra os processos oxidativos que naturalmente ocorrem no organismo. Os compostos fenólicos exibem grande quantidade de propriedades fisiológicas (como antialergênica, antiaterogênica, antiinflamatória, antimicrobiana, antitrombótica, cardioprotetiva e vasodilatadora), mas o principal efeito dos compostos fenólicos tem sido atribuído à sua ação antioxidante em alimentos (BALASUNDRAM et al., 2006). O consumo cada vez mais frequente de alimentos industrializados desperta o interesse na qualidade desses produtos que estão sendo consumidos e a influência do processamento ainda é pouco estudada. Por isso a importância em verificar a influência do processamento nos compostos benéficos que estão presentes nos alimentos in natura (VEDANA, 2008). Diante do exposto, o presente trabalho teve como objetivos avaliar a atividade das enzimas oxidativas e pectinolíticas, importantes para a indústria de alimentos, em diferentes variedades de abacaxi de modo a subsidiar a escolha das variedades com boas características industriais e funcionais. Além de determinar os parâmetros ideais de inativação das enzimas encontradas minimizando custos e danos ao produto e avaliar a atividade antioxidante nas variedades estudadas, antes e depois do tratamento de inativação enzimática. 3 2.0 Revisão da Literatura 2.1 Abacaxi 2.1.1 Características gerais O abacaxizeiro pertence à família Bromeliaceae (subclasse das monocotiledôneas) gênero Ananas, sendo a espécie Ananas comosus L. Merril a que mais se destaca, por sua importância econômica e em razão de abranger todas as variedades de interesse da fruticultura comercial (PIZA et al., 2003). O seu fruto, o abacaxi, é normalmente cilíndrico constituído por 100 a 200 pequenos frutilhos fundidos entre si sobre o eixo central, em que cada “olho” ou “escama” da sua casca é um fruto verdadeiro, que cresceu a partir de uma flor, e estes se fundem em um grande corpo, chamado infrutescência, no topo do qual se forma a coroa (GIACOMELLI, 1981; SILVA & TASSARA, 2001). A parte comestível do fruto é a polpa suculenta formada pelas paredes das lojas dos frutilhos e pelo tecido parenquimatoso que as ligam entre si, bem como pela porção externa do miolo (MEDINA, 1987). O abacaxi é extensivamente produzido em todos os países tropicais, sendo que o Brasil encontra excelentes condições para seu desenvolvimento, sendo cultivado em quase todos os Estados (FAOSTAT, 2010). O Brasil responde por 8,93% da produção mundial e a área plantada em 2009 foi de 60 mil hectares, nos quais foram colhidos 1,479 milhões de frutos. Na safra de 2010, foram produzidos 1,48 milhões de frutos em uma área colhida de 57.727 hectares, um acréscimo de 1,07% em relação ao ano anterior. Os Estados da Paraíba, Minas Gerais, Pará e Bahia são os principais produtores, o que torna o Nordeste a região de maior produção (Figura 01) (IBGE, 2010). A Bahia responde por 8,23% da produção brasileira, ocupando o quarto lugar no ranking dos estados produtores, com uma produção de 121,127 mil frutos no ano de 2009 (IBGE, 2010). No Recôncavo Baiano a cultura do abacaxi vem se destacando a cada ano, gerando emprego e renda para o meio rural, contribuindo para o desenvolvimento local. 4 Figura 01. Produção brasileira de abacaxi por região fisiográfica. Fonte: IBGE, 2010. O Brasil é considerado um dos principais centros de diversidade genética do abacaxi porque, além de A. comosus, todas as espécies do gênero Ananas são encontradas nas formas silvestres ou cultivadas em várias regiões brasileiras (FERREIRA & CABRAL, 1993). Existem centenas de cultivares desenvolvidas no Brasil e no mundo, sendo que a Singapore Spanish, Queen e Espanhola Roja, juntamente com a Perolera, Smooth Cayenne e Pérola, caracterizam as mais conhecidas no mundo (CABRAL et al., 1999). As principais cultivares brasileiras são a Pérola e a Smooth Cayenne, sendo a Pérola a mais cultivada (80% da produção nacional) que é destinada principalmente para o consumo in natura pela sua alta qualidade organoléptica (CABRAL et al., 1999; PINHEIRO et al., 2000). A indústria tem a preferência pela variedade Smooth Cayenne por apresentar fruto com excelente aspecto, de elevado tamanho e formato cilíndrico (o que facilita o descascamento), pesando de 1,5 a 2,0 kg, polpa firme e amarela, com elevado teor de açúcares e elevada acidez (PINHEIRO et al., 2000; NASCENTE et al., 2005). Já o abacaxi Pérola apresenta forma cônica, polpa branca, sucosa, pouca acidez (o que favorece ao consumo in natura), pesando de 1,0 kg a 1,5 kg (CABRAL et al., 2000). A qualidade dos frutos é atribuída ao seu tamanho, forma e cor da casca, cujos fatores associados à composição físico-química da polpa, oferecem aos frutos e aos produtos deles obtidos a qualidade sensorial e nutricional, responsáveis pela aceitação definitiva desses no mercado (SCALON et al., 2004). O abacaxi apresenta ampla variação em sua composição química. Diferentes estudos apresentam amplas faixas para os valores de pH, acidez titulável, açúcares totais e sólidos solúveis dependendo da variedade 5 cultivada, do estádio de maturação, do clima e da época do ano em que o fruto foi produzido, do solo, dos tratos culturais, entre outros fatores (CESAR, 2005). Ele possui em sua composição: ferro, potássio, cálcio, as vitaminas A1, B1, B2 e C, além das vitaminas tiamina, riboflavina e niacina, fibras e outros compostos orgânicos (MEDINA et al., 1978; FRANCO, 1989). As formas mais frequentes de comercialização do abacaxi são in natura, em conserva, suco natural e suco concentrado, polpa em calda, além de ser utilizado também na fabricação de doces, sorvetes, cremes, balas, bolos, licor, vinho, vinagre e aguardente. No mercado externo tem-se observado um particular interesse com relação aos sucos de frutas tropicais, despertando as indústrias nacionais de sucos para o imenso potencial a ser explorado junto aos consumidores da Europa, Ásia e Estados Unidos. Segundo dados do Instituto Brasileiro de Fruticultura (IBRAF, 2008), o Brasil exportou mais de 2 milhões de litros de suco de abacaxi no ano de 2007, enquanto que em 2006 essa marca atingiu apenas 450 mil litros. 2.1.2 Melhoramento Genético do abacaxi Apesar da existência de diversas cultivares de abacaxi, os plantios comerciais em todo o mundo são formados predominantemente com a variedade Smooth Cayenne. Esta estreita base genética é desfavorável à sustentabilidade da cultura, fazendo-se necessário uma diversificação de cultivares (CABRAL, 1999). Os programas de melhoramento do abacaxi visam obter cultivares mais produtivas, adaptadas às condições climáticas locais e resistentes a pragas e moléstias. Entre as principais características preconizadas no melhoramento do abacaxizeiro, está a busca de genótipos resistentes à fusariose (Fusarium subglutinans f.sp. ananas,), principal moléstia da cultura do abacaxi no Brasil. Além disso, são almejados genótipos que evidenciem crescimento rápido, folhas com ausência ou poucos espinhos nas extremidades dos seus bordos, rebentão precoce localizado na base da planta, fruto de forma cilíndrica, casca amarela e pouco fibrosa, elevado teor de sólidos solúveis totais, acidez moderada e alto teor de ácido ascórbico e adaptação ao sistema de produção integrada (CUNHA, 2007; CRESTANI et al., 2010). O Centro Nacional de Pesquisa de Mandioca e Fruticultura da Embrapa possui o maior Banco Ativo de Germoplasma (BAG) de abacaxizeiros do mundo, contando com aproximadamente 700 acessos. A caracterização e avaliação deste BAG permitiu a 6 identificação de acessos resistentes à fusariose, com folhas sem espinhos nos bordos, produção precoce de rebentões, sólidos solúveis totais acima de 15° brix e acidez moderada. Esses acessos são usados para o desenvolvimento de novas cultivares, algumas já lançadas no mercado, como as cultivares Imperial e Vitória (CABRAL et al., 2006; MACHADO et al., 2009). A cultivar Imperial (Figura 02) foi lançada pela Embrapa Mandioca e Fruticultura em 2003. Esta cultivar é resultante do cruzamento das variedades Perolera com Smooth Cayenne. Além de ser resistente à fusariose, o fruto é pequeno, cilíndrico, a polpa é amarela, com elevado teor de açúcar, acidez titulável moderada, alto conteúdo em ácido ascórbico e excelente sabor, de acordo com as análises sensoriais realizadas. Essas características contribuem para que os frutos obtidos sejam destinados para o mercado de consumo in natura e para a industrialização (CABRAL & MATOS, 2005). Figura 02. Fruto Abacaxi Imperial Fonte: CABRAL & MATOS 2005. A cultivar Vitória (Figura 03) é resultado dos estudos da Embrapa Mandioca Fruticultura e do Incarper (Instituto Capixaba de Pesquisa, Assistência Técnica e Extensão Rural). Foi utilizado como parental feminino a cultivar Primavera (PRI) e parental masculino a cultivar Smooth Cayenne (SC), dando no ano de 2006 origem à cultivar Vitória. Os frutos têm polpa branca, elevada acidez, formato cilíndrico, pesando em torno de 1,5 kg e excelente sabor segundo as análises químicas e sensoriais, sendo indicado tanto para o consumo in natura quanto para a industrialização (VENTURA et al., 2006). 7 Figura 03. Abacaxi Vitoria. Fonte: INCARPER, 2010. 2.2 Enzimas vegetais de importância na indústria de alimentos Enzimas são, em sua maioria, proteínas que catalisam, com grande eficiência, as reações metabólicas em determinadas condições de pH, temperatura, meio iônico, entre outros (WHITAKER, 1972). As reações enzimáticas são muito importantes em alimentos. Elas são responsáveis pela formação de compostos extremamente desejáveis, mas também podem provocar consequências desfavoráveis e ocorrem não só no alimento natural, mas também durante o seu processamento e armazenamento. Os alimentos de origem vegetal merecem atenção especial, pois os vegetais possuem em sua composição grupos de enzimas que causam alterações nos alimentos. A respeito do comportamento fisiológico de frutos e vegetais, já está estabelecido que grande parte de sua deterioração - mudanças no sabor, na textura, turbidez e no valor nutricional - são ocasionados por enzimas do grupo das oxidoredutases, sobretudo peroxidases (PER) e polifenol-oxidases (PFO); e pelo grupo de enzimas pectinolíticas pectinametilesterases (PME) e poligalacturonases (PG) (LEE et al., 1984; CHITARRA; CHITARRA, 2005). 8 2.2.1 Enzimas Oxidativas Enzimas oxidativas são aquelas que realizam reações de oxirredução. As enzimas PFO e PER são importantes, pois em frutas e vegetais são responsáveis pela mudança de cor, degradação da clorofila, oxidação de fenóis, do ácido indol acético, sendo que muitos desses fatores estão associados com sabor, aroma, cor, textura e qualidades nutricionais dos alimentos (VÁMOS-VIGYÁZÓ, 1981). O problema mais grave dessas enzimas nos frutos é o escurecimento dos tecidos, que ocorre principalmente pela reação de oxidação dos compostos fenólicos (FILHO & VIEIRA, 2002). O escurecimento enzimático de vegetais inicia-se em resposta às injúrias físicas e/ou fisiológicas (impactos, abrasões, injúria pelo frio, excesso de CO2) e é resultado da oxidação de compostos fenólicos por ação, sobretudo, de PFO. De acordo com Tomás-Barberán & Spín (2001), PER e PFO são relevantes na degradação oxidativa dos compostos fenólicos por causarem a produção de polímeros de coloração marrom conhecidos como melaninas. Whitaker & Lee (1995) acreditam que mais de 50% das perdas em frutas ocorrem como resultado do escurecimento enzimático. O controle do escurecimento, portanto, torna-se crítico para a diminuição das perdas comerciais do agricultor e para a indústria de transformação. Os pigmentos escuros formados pela ação de PER e PFO são geralmente acompanhados de mudanças indesejáveis na aparência e nas propriedades sensoriais do produto, resultando na diminuição da sua vida útil e do seu valor de mercado (ARAÚJO, 1999). Dentre os diversos fatores que constituem entraves na qualidade do abacaxi destinados à exportação, está um distúrbio fisiológico denominado “escurecimento interno”, causado pelo transporte e armazenamento dos frutos em temperaturas inferiores à sua faixa de segurança (temperatura mínima de segurança - TMS), que é considerada entre 7 e 12ºC. Entretanto, tem-se observado que mesmo dentro dessa faixa de temperatura pode ocorrer a manifestação de sintomas dessa injúria no abacaxi (ABREU, 1995). Esse distúrbio fisiológico dos frutos também é associado à ação das enzimas PFO e PER e sua ação sobre os compostos fenólicos (TEISSON, 1979). Com a injúria do tecido pelo frio há o rompimento das membranas celulares liberando as enzimas, que entram em contato com seus substratos (fenólicos e O2 ou H2O2), havendo assim uma oxidação excessiva e nãocontrolada ocasionando os compostos escuros (ABREU, 1995). 9 2.2.1.1 Peroxidases PER (EC 1.11.1.7, doador H2O2 óxido-redutase) são enzimas capazes de oxidar diferentes compostos, na presença de peróxidos, gerando radicais livres. Na ausência de peróxidos, essas enzimas podem ainda catalisar a oxidação de alguns substratos com o auxilio de oxigênio molecular e também hidroxilar diferentes compostos aromáticos (tirosina, fenilalanina e outros fenólicos). Por ser não especifica, ela pode catalisar a oxidação de um grande número de fenóis e compostos com anéis aromáticos que ocorrem naturalmente em tecidos de plantas. Normalmente, essas enzimas estão relacionadas a reações de oxidação que resultam na formação de compostos coloridos, mas também podem promover várias reações de biodegradação (BURNETTE, 1977; KOBLITZ, 2008). O estudo da PER é de grande importância, pois ela pode interferir na qualidade dos produtos, tanto para os industrializados como para os de consumo in natura, podendo até mesmo influenciar a deterioração de alimentos quando armazenados, desenvolvendo sabores e odores indesejáveis nos produtos (HAARD, 1977; LEE & HAMMES, 1979). A PER pode participar da destruição de vitamina C, catalisar o branqueamento dos carotenóides na ausência de ácidos graxos insaturados e a descoloração de antocianinas. Catalisa ainda a reação de degradação de ácidos graxos insaturados, produzindo voláteis que alteram o sabor (RICHARDSON & HYSLOP, 1985). As PER estão entre as enzimas mais termoestáveis relatadas em vegetais (BURNETTE, 1977). Podem apresentar diferentes valores de pH ótimo de atividade, dependendo do substrato empregado e, após certas condições de tratamento, podem regenerar sua atividade durante o armazenamento de alimentos processados (BURNETTE, 1977; CLEMENTE, 1995). A Figura 04 representa o mecanismo de escurecimento enzimático da PER, utilizando como doador de hidrogênio o guaiacol, bastante utilizado nos métodos de detecção da atividade dessa enzima (FILHO & VIEIRA, 2002). Na reação enzimática o peróxido de hidrogênio ou outro peróxido orgânico, como o peróxido de metila ou o etil hidrogênio, é reduzido, enquanto que um doador de elétrons (AH2) é oxidado. O doador de elétrons pode ser ascorbato, fenóis, aminas ou outros compostos orgânicos. Em muitos casos o produto da oxidação é colorido e serve como base para a determinação colorimétrica da atividade da peroxidase (BRITO et al., 2005). 10 Figura 04. Mecanismo de escurecimento enzimático a partir da peroxidase (PER). Fonte: DEBIEN, 2010. Ferreira (1983) citado por Brito (2001) purificou e caracterizou PER de abacaxi da cultivar Pérola. O extrato bruto apresentou uma atividade total de 14,6 U/minuto/mL de suco e uma atividade específica de 8,59 U/minuto/mg de proteína. Beaudreau & Yasunobu (1996) encontraram, para o extrato bruto de abacaxi, uma atividade de 24.800 U/mL. Brito et al. (2005), estudaram algumas características de PER de cultivares de abacaxi desenvolvidos pelo Instituto Agronômico de Campinas (IAC). As PER dos cultivares IAC Gomo-de-mel e de seu clone IAC-1 apresentaram temperatura ótima para a atividade nas faixas de 45 °C a 50 °C e de 50 °C a 55 °C, respectivamente, e apresentaram atividade ótima em pH 4,5 a 50 °C, com menor atividade em valores de pH muito ácidos (2,6) ou alcalinos (superiores a 7,0). Brito et al. (2008), estudando as mesmas variedades, verificaram que IAC Gomo-de-mel apresentou menor atividade de PER bruta (3194 U/g) que os cultivares comerciais Smooth Cayenne (5137 U/g) e Pérola (5624 U/g). 2.2.1.2 Polifenol-oxidases (PFO) PFO são enzimas capazes de oxidar compostos fenólicos com o auxílio do oxigênio molecular. O resultado final das reações catalisadas por PFO são quinonas. Essas substâncias são altamente reativas e combinam-se entre si e com outros componentes do meio para gerar produtos de condensação de alta massa molecular e cor escura, chamados de melaninas (KOBLITZ, 2008). São três as reações catalisadas pela PFO na presença do oxigênio molecular: a hidroxilação de monofenóis a o-difenóis (atividade monoxigenase, EC 1.14.18.1), a oxidação de o-difenóis a o-quinonas (atividade difenoloxidase, EC 1.10.3.2), e a oxidação de p-difenóis a p-quinonas (atividade lacase, EC 1.10.3.1) 11 (ZAWISTOWSKI et al., 1991). A Figura 05 mostra o mecanismo, resumidamente, da ação das PFO no escurecimento enzimático. Em tecidos vivos, esta enzima e o substrato fenólico estão separados dentro das células. No momento da extração ou de algum outro processo que danifique a célula, a enzima e o substrato entram em contato, ocorrendo então a reação. Com a formação da oquinonas, acontece a reação com outras o-quinonas formando as melaninas, além de reagirem também com aminoácidos, peptídeos e proteínas causando alterações estruturais e funcionais, acarretando a diminuição do valor nutritivo do produto (ESCRIBANO et al., 1997). Figura 05. Reação de escurecimento enzimático a partir da polifenol-oxidase. Fonte: DABIEN, 2010. Nos tecidos vegetais, PFO estão localizadas nos cloroplastos e sua concentração (atividade) no tecido vegetal depende do local do plantio, período da colheita, espécie e do estado de amadurecimento, sendo menor em frutos ou vegetais não-amadurecidos (FILHO & VIEIRA, 2002). Em plantas, as PFO podem estar solúveis ou ionicamente ligadas à membrana. Estudos histoquímicos revelaram que essas enzimas estão localizadas predominantemente em membranas tilacóides de cloroplastos e a sua distribuição nas diferentes partes da planta também é variável (MARTINEZ & WHITAKER, 1995; VÁMOS-VIGYÁZÓ, 1981). Thé et al. (2001) estudaram atividade de PFO em diferentes estádios de maturação e formas de armazenamento em polpa de abacaxi e encontraram diferença significativa para a atividade entre todos os fatores estudados, assim como para a interação entre os mesmos. A atividade encontrada variou entre 25,81 U/g a 47,98 U/g. Nos frutos em estádio de maturação 3, armazenados sob refrigeração, a atividade foi mais elevada, o que reforça a 12 teoria da ocorrência de injúria pelo frio (escurecimento interno) e que em frutos maduros a atividade desta enzima é maior. Zhou et al. (2003) estudaram a relação entre a atividade de PER e PFO com o escurecimento interno dos frutos de abacaxi submetidos à refrigeração. Verificaram que a atividade de PFO era muito baixa na colheita dos frutos e manteve-se baixa durante o armazenamento pós-colheita numa temperatura de 25 °C durante 4 semanas. Entretanto, a atividade de PFO aumentou consideravelmente nos frutos após armazenagem em temperatura mais baixa. O aumento da atividade de PFO foi diretamente relacionado à incidência de sintomas do escurecimento interno. 2.2.2 Enzimas pectinolíticas As pectinases formam um grupo de enzimas que degradam substâncias pécticas, hidrolisando ligações glicosídicas ao longo da cadeia carbônica (UENOJO & PASTORE, 2006). As substâncias pécticas são definidas como um grupo complexo de polissacarídeos em forma coloidal, contendo uma grande proporção de unidades de ácido anidro galacturônico em forma de cadeia. Os grupos carboxílicos do ácido poligalacturônico podem estar parcialmente esterificados por grupos metílicos e parcial ou totalmente neutralizados pelo sódio, potássio ou íons de amônio. Incluindo as galactanas e arabinanas (GAVA, 1998; KRATCHANOV et al., 2004). A degradação de polissacarídeos pécticos é uma das principais causas do processo de amaciamento dos frutos. Estão envolvidos na modificação da textura de frutas dois principais processos enzimáticos, cuja ação é devida à atividade das enzimas pectinolíticas pectinametilesterase - PME e da poligalacturonase - PG (ANTHON et al., 2002). Quando os grupos carboxílicos livres das substâncias pécticas encontram-se ligados ao cálcio, formam o pectato de cálcio, que é insolúvel e também designado como protopectina, predominante em frutos imaturos. Com o amadurecimento, há liberação de cálcio e solubilização de protopectina das paredes celulares, possivelmente por ação enzimática. Há então modificação da textura, que se torna gradualmente macia. Essas transformações ocorrem não só durante o amadurecimento, como também no armazenamento de frutas e algumas hortaliças (CHITARRA & CHITARRA, 2005). Em decorrência das modificações das pectinas durante o amadurecimento, a estrutura das paredes celulares torna-se 13 progressivamente mais hidratada. Dessa forma, a textura do fruto é afetada pela facilidade com que as células rompem ou se separam umas das outras (BRUMMELL et al., 2004). PME são enzimas desmetoxilantes que hidrolisam a ligação éster removendo a metoxila de ácidos galacturônicos esterificados com metanol, resultando em pectinas com baixo teor de metoxilação. PG são enzimas despolimerizantes que hidrolisam as ligações glicosídicas entre ácidos galacturônicos (em pectinas com baixo teor de metoxilação ou ácido péctico/poligalacturônico) e sua atividade decresce com o aumento no grau de metoxilação dos substratos (KOBLITZ, 2008). A ação de PME deve preceder a atividade de PG, no sentido de facilitar a atividade desta última, pela desmetilação dos poliuronídeos, uma vez que PG apresentam afinidade pelo substrato linear desmetilado, após a atuação da PME (ANTHON et al., 2002). 2.2.2.1 Pectinametilesterases (PME) Durante o crescimento celular, os homogalacturonanos são secretados nas paredes celulares com um elevado grau de metilesterificação e, progressivamente, são desmetilesterificados. Concomitantemente, ocorre aumento da atividade de PME (EC 3.1.1.11), enzimas que hidrolisam os grupos metiléster da posição C-6 dos homogalacturonanos, formando pectinas com menor grau de metilação e metanol. A atividade de PME é detectada durante todas as fases do desenvolvimento dos frutos, sendo que picos de atividade são observados quando os frutos são ainda pequenos e imaturos e logo no início do processo de amadurecimento (BRUMMELL & HARPSTER, 2001). Em sucos, a diminuição no grau de esterificação e o aumento na densidade de cargas geradas na cadeia péctica causam uma elevação na afinidade pelos íons de cálcio (Ca+2), sendo então geradas ligações cruzadas intermoleculares mediadas por esse íon. Esta auto-associação das regiões não esterificadas da pectina poderá levar à formação de agregados e por consequência sua precipitação (ROMBOUTS et al., 1982; BAKER & CAMERON, 1999; BOAS et al., 2000). O grande problema na indústria de sucos concentrados é a formação desses géis de pectato de cálcio impedindo a reconstituição satisfatória do suco (KOBLITZ, 2008). Por isso o suco é inicialmente submetido a um aquecimento inicial à temperatura de 90 a 95 ºC durante 30 a 40 segundos, visando a inativação enzimática (MAIA et al., 2007). Segundo o estudo elaborado por Thé et al. (2001), a atividade de PME variou de 87,50 U/g a 312,50 U/g em diferentes estádios de maturação do abacaxi. Thé et al. (2006) 14 verificou que a atividade de PME nos frutos de abacaxi analisados no dia da colheita (116,25 U.g-1/min) foi inferior à observada após o armazenamento (variação de 174,0 a 750,00 U.g-1/min). Durante o armazenamento eles utilizaram diversos métodos para prolongar a vida útil do fruto (associação de tratamentos hidrotérmicos, cloreto de cálcio e atmosfera modificada). Os mais efetivos foram aqueles que utilizaram embalagem, resultado também encontrado por Abreu (1995), onde os frutos acondicionados em embalagens de polietileno sem perfuração evidenciaram menor atividade de PME. 2.2.2.2 Poligalacturonases (PG) PG são as enzimas que catalisam a hidrólise das ligações glicosídicas α(1→4) da cadeia de ácido poligalacturônico. As endo-PG (EC 3.2.1.15) hidrolisam as ligações glicosídicas α(1→4) internas de forma randômica, causando a despolimerização da molécula e liberando oligômeros de ácido galacturônico, enquanto as exo-PG (EC 3.2.1.67) removem as moléculas de ácido D-galacturônico pela hidrólise das ligações glicosídicas α(1→4) a partir da extremidade redutora, liberando ácidos mono- ou digalacturônicos (VANDRESEN, 2007). Os homogalacturonanos estão presentes na parede celular numa forma altamente metilesterificada e, para tornarem-se substratos da PG, precisam ser desmetilesterificados (BRUMMELL & HARPSTER, 2001). A hidrólise das ligações glicosídicas existentes nas substâncias pécticas está diretamente ligada ao aumento de pectinas solúveis, à diminuição da rigidez intercelular e à adesividade dos tecidos vegetais (ASSIS et al., 2000; RESENDE et al., 2004). O aumento de pectinas solúveis causados pelas PG afetam diretamente a textura e firmeza dos produtos de frutas, problema encontrado principalmente nos abacaxis minimamente processados. Thé et al. (2009) estudaram a influência da associação de tratamento hidrotérmico, cloreto de cálcio e atmosfera modificada sobre fatores relacionados à textura de abacaxi, cultivar Smooth Cayenne. Os autores relataram que os efeitos do cálcio nos frutos têm recebido muita atenção, visto que a aplicação desse cátion produz efeitos positivos no adiamento da maturação e da senescência. São citados Siddiqui & Bangerth (1996), que afirmam que o acúmulo dos cátions Ca2+ pode facilitar a ligação entre os polímeros de pectina, particularmente na lamela média, aumentando sua resistência. Foi encontrada, no trabalho de Thé et al. (2009), atividade de PG em frutos de abacaxi variando entre 929,38 nmol.g-1 a 526,85 nmol.g-1. Thé et al. (2001), não observou 15 diferenças significativas entre a atividade de PG dos abacaxis Smooth Cayenne em diferentes estádios de maturação, porém a atividade desta enzima foi influenciada pelo armazenamento (com refrigeração: 697,24 U/g e sem refrigeração: 765,53 U/g). Este resultado também foi encontrado por Prado et al. (2003), que obteve o menor valor de atividade para abacaxi Smooth Cayenne minimamente processado sob refrigeração e atmosfera modificada com valores que variaram na faixa de 36,05 a 69,83 U/min/g. 2.3 Inativação enzimática Na indústria de alimentos, a inativação e a ativação enzimáticas são recursos utilizados no processamento de vários produtos. A inativação enzimática objetiva principalmente a conservação do produto, em seu período de elaboração e de armazenamento (EVANGELISTA, 1998). A inibição ou inativação enzimática pode ser realizada através de alguns métodos, como por exemplo: a adição de aditivos inibidores como o sulfito ou dióxido de enxofre, ácido ascórbico; mudanças de pH; exclusão do oxigênio por embalagem a vácuo ou emprego de atmosfera modificada; adição de sacarose para alterar a atividade de água; o congelamento e o tratamento pelo calor, sendo o tratamento térmico e a adição de sulfito ou dióxido de enxofre os mais utilizados. Além da temperatura, outro parâmetro importante no processamento térmico é o tempo. Assim, ao se especificar a temperatura do processamento térmico, é indispensável que se especifique também o tempo de tratamento (CUNHA et al., 2005). Diversos mecanismos têm sido propostos com o intuito de prevenir o escurecimento dos tecidos, os quais visam à diminuição da biossíntese de enzimas envolvidas e cuidados na manutenção da integridade celular do tecido vegetal. O controle do escurecimento torna-se crítico para a diminuição das perdas comerciais do agricultor e para a indústria de transformação (SILVA et al., 2009). O uso de inibidores de escurecimento enzimático em alimentos é restrito pela toxicidade que podem causar dependendo da concentração empregada, e também pelo potencial efeito negativo na textura, aroma, gosto e custos. O ácido ascórbico, ou vitamina C, além de atribuir valor nutricional aos alimentos, também apresenta ação redutora. Juntamente com seus sais neutros compõe um dos principais grupos de antioxidantes empregados em produtos vegetais com o intuito de prevenir o escurecimento e outras reações oxidativas de duas maneiras: (1) agindo diretamente na enzima, complexando o cobre do grupo prostético da PFO ou oxidando os 16 resíduos de histidina que ligam os íons de cobres do sítio ativo, causando sua inibição; (2) reduzindo as quinonas à sua forma anterior de fenóis, impedindo a formação dos pigmentos escuros (SAPERS e MILLER, 1998; KOBLITZ, 2008). O sulfito é um dos inibidores químicos mais comumente utilizados na prevenção do escurecimento enzimático (MARTINEZ et al., 1995). Geralmente é aplicado sob forma de sais (metabissulfito ou bissulfito de sódio ou potássio). São inibidores diretos da PFO atuando sobre seu sítio ativo, além de reagirem com as quinonas formadas, gerando compostos incolores estáveis, o que evita que se polimerizem (KOBLITZ, 2008). Sabe-se que o calor pode reduzir a atividade de enzimas e o cálcio pode contribuir para a manutenção da integridade celular; como conseqüência, a interação entre esses dois fatores também pode contribuir para menor acúmulo de substratos (fenólicos) nos tecidos da polpa do abacaxi reduzindo a interação destes com as enzimas menos ativas, responsáveis pelas reações de escurecimento (GONÇALVES et al., 2000). Resultados obtidos em vários estudos sugerem que a manutenção da integridade celular conferida pela aplicação de CaCl2 nos frutos se reflete numa menor atividade das enzimas oxidativas (PER e PFO) resultando em menor índice de escurecimento, o que pode ser, em parte, atribuído à menor possibilidade de contato das enzimas com substratos e menor disponibilidade de fenólicos (GONÇALVES et al., 2000). O cálcio participa de forma importante da estrutura e da resistência mecânica da parede celular. O acúmulo dos cátions Ca2+ pode facilitar a ligação entre os polímeros de pectina, particularmente na lamela média, aumentando sua resistência. O cálcio, além de conferir insolubilidade ao material péctico, pode inibir a atividade da PG. O pectato formado pela presença do cálcio é resistente à degradação pela PG, que atua preferencialmente na ligação glicosídica adjacente ao grupo carboxílico não-esterificado (THÉ et al., 2006). Diversas alternativas vem sendo estudadas a fim de se fornecer meios para a inativação das enzimas que influenciam o tempo de armazenamento dos frutos e de seus produtos. Silva et al. (2007) estudaram o efeito da radiação ionizante com fonte de Cobalto-60, como um tratamento alternativo para o aumento da vida útil de abacaxi. A atividade de PFO, no entanto, aumentou com aplicação da radiação, apresentando resultado de 32,60 U.g -1/min, enquanto os frutos controle mantiveram atividade de 27,78 U.g –1/min. 17 2.4 Atividade antioxidante e Compostos Fenólicos As frutas, reconhecidas fontes de vitaminas, minerais e fibras, são alimentos nutricionalmente importantes da dieta. No entanto, nos últimos anos, maior atenção tem sido dada a estes alimentos uma vez que evidências epidemiológicas têm demonstrado que o consumo regular de vegetais está associado à redução da mortalidade e morbidade por algumas doenças crônicas não transmissíveis. O efeito protetor exercido por estes alimentos tem sido atribuído à presença de polifenóis com ação antioxidante (MELO et al., 2008). Compostos fenólicos, estruturalmente, compreendem compostos que possuem um anel aromático ligado a um ou mais grupos hidroxila e podem variar de simples moléculas de fenóis a grandes moléculas de compostos polimerizados. Embora haja esta diversidade estrutural, esse grupo de compostos é denominado polifenóis (BALASUNDRAM et al., 2006). Existem cerca de cinco mil fenóis, dentre eles, destacam-se os flavonóides, ácidos fenólicos, fenóis simples, cumarinas, taninos, ligninas e tocoferóis (NACZAK & SHAHIDI, 2004). Principais fontes dietéticas de polifenóis, as frutas, em função de fatores intrínsecos (cultivar, variedade, estádio de maturação) e extrínsecos (condições climáticas e edáficas) apresentam, em termos quantitativos e qualitativos, composição variada desses constituintes. Por sua vez, a eficácia da ação antioxidante depende da estrutura química e da concentração dos polifenóis no alimento (MELO et al., 2008). O excesso de radicais livres no organismo é combatido por antioxidantes produzidos pelo corpo ou absorvidos da dieta. Quando há um desbalanço entre a produção de radicais livres e os mecanismos de defesa antioxidante, ocorre o chamado “estresse oxidativo”. Os radicais livres em excesso podem ser originados por defeitos na respiração mitocondrial, metabolismo do ácido araquidônico, ativação-inibição de sistemas enzimáticos ou por fatores exógenos, como poluição, hábito de fumar ou ingerir álcool, ou ainda, por uma nutrição inadequada (NÚÑEZ-SELLÉS, 2002). Um ponto interessante da atividade farmacológica dos polifenóis naturais é sua eficiência contra o estresse oxidativo. Os polifenóis reduzem o estresse oxidativo, pois neutralizam radicais livres reativos gerando moléculas de baixa reação (MISHRA et al., 2005). De forma geral, denominam-se antioxidantes as substâncias que, presentes em concentrações baixas, comparadas ao substrato oxidável, retardam significativamente ou inibem a oxidação desse substrato. Os radicais formados a partir de antioxidantes não são 18 reativos para propagar a reação em cadeia, sendo neutralizados por reação com outro radical, formando produtos estáveis ou podem ser reciclados por outro antioxidante (OMONI & ALUKO, 2005; BARREIROS et al., 2006). O consumo de substâncias antioxidantes na dieta diária pode produzir uma ação protetora efetiva contra os processos oxidativos do organismo. Foi descoberto que uma série de doenças, entre as quais câncer, aterosclerose, diabetes, artrite, malária, AIDS, doenças do coração, podem estar ligadas aos danos causados por formas de oxigênio extremamente reativas denominadas de “substâncias reativas oxigenadas” ou simplesmente ROS. Estas substâncias também estão ligadas com processos responsáveis pelo envelhecimento do corpo. (BRENNA; PAGLIARINI, 2001; YILDRIM et al., 2001) Um grande número de métodos tem sido desenvolvido para avaliar a atividade antioxidante. Entretanto, a comparação entre esses métodos torna-se muito difícil uma vez que os métodos e os substratos utilizados são muito diferentes. Adicionalmente, formas de acelerar a oxidação dos diversos substratos são empregadas a fim de aumentar a velocidade dos estudos. Desta forma, torna-se impossível expressar a atividade antioxidante como um valor absoluto, sendo muito recomendável a utilização de padrões comparativos independente do método escolhido (KOLEVA et al., 2002). Dentre os métodos mais utilizados para determinação da atividade antioxidante em frutos e hortaliças estão o DPPH (2,2-difenil-1-picrilhidrazil), FRAP (poder antioxidante de redução do ferro), sistema β-caroteno/ácido linoléico e o ABTS (2,2‟ – azino-bis(3etilbenzotiazolina-6-ácido sulfônico) sal diamônio) (WU et al., 2005). Alguns autores recomendam a utilização do método DPPH por ser fácil e preciso para a avaliação da atividade antioxidante de produtos vegetais (RIBEIRO et al., 2002; CARDOSO et al., 2005; JAYAPRAKASHA et al., 2007). O método está baseado na capacidade do DPPH em reagir com doadores de hidrogênio e, na presença de substâncias antioxidantes, receber H+, sendo então reduzido. O DPPH é um radical livre que pode ser obtido diretamente por dissolução do reagente em meio orgânico (RUFINO et al., 2007) e pode ser facilmente detectado por espectroscopia, devido à sua intensa absorção na região visível (BONDET et al., 1997; SANCHEZ-MORENO, 2002), com base na captura do radical DPPH por antioxidantes, produzindo um decréscimo da absorbância a 515 nm (BRAND-WILLIAMS et al., 1995). Um parâmetro cinético importante quando se avalia atividade antioxidante é o EC 50 introduzido por Sánchez-Moreno et al. (1998). O EC50 refere-se à quantidade de antioxidante necessária para reduzir em 50% a quantidade inicial do radical. 19 Melo et al. (2008) estudaram a capacidade antioxidante de diversas frutas tropicais através do método que determina a capacidade de sequestrar o radical DPPH. O abacaxi obteve um significativo resultado com capacidade antioxidante moderada (50-70% de sequestro). 2.5 Otimização de processos e delineamento experimental O planejamento experimental, também denominado delineamento experimental, representa um conjunto de ensaios estabelecido com critérios científicos e estatísticos, com o objetivo de determinar a influência de diversas variáveis nos resultados de um dado sistema ou processo (BUTTON, 2005). A metodologia do planejamento experimental, associada à análise de superfície de respostas, é uma ferramenta fundamentada na teoria estatística, que fornece informações seguras sobre o processo, minimizando o empirismo que envolve técnicas de tentativa e erro (BOX et al., 1978). Com o objetivo da redução do número de pontos experimentais, foi criada a técnica que utiliza os delineamentos compostos. Eles foram desenvolvidos, inicialmente, por Box & Wilson (1951) para estudo de funções polinomiais de resposta na indústria, sendo o erro experimental pequeno, e as condições do experimento mais facilmente controláveis (RAMOS, 2005). A metodologia de superfície de resposta é essencialmente um conjunto de técnicas estatísticas usadas em pesquisas, com a finalidade de determinar as melhores condições e dar maior conhecimento sobre a natureza de certos fenômenos. Sua aplicação permite selecionar a combinação de níveis ótimos na obtenção da melhor resposta para uma dada situação. Então, usando a metodologia, é possível aproximar um modelo empírico a uma relação (inicialmente desconhecida ou conhecida) entre os fatores e as respostas do processo (SILVA, 2005). A modelagem normalmente é feita ajustando-se modelos lineares ou quadráticos a resultados experimentais obtidos a partir de planejamentos experimentais. O modelamento ocorre em busca do caminho de máxima inclinação de um determinado modelo, sendo o caminho onde a resposta varia de forma mais pronunciada (BARROS NETO et al., 2007). A descrição gráfica do modelo chama-se de superfície de resposta, e como segunda alternativa tem-se o gráfico bidimensional de contorno, onde cada contorno corresponde a uma particular altura da superfície de resposta e em cada linha a resposta é constante. A sensibilidade dos parâmetros estudados sobre a resposta de interesse é examinada através do diagrama de Pareto. A utilização dessa ferramenta permite indicar 20 quais parâmetros e interações têm influências significativas sobre cada variável resposta considerada (BARBOSA, 2009). 3.0 Material e Métodos 3.1 Experimento 1. Planejamento experimental e otimização do processo de extração enzimática. 3.1.1 Material vegetal Foi utilizado abacaxi cultivar Pérola adquirido no comércio da cidade de Feira de Santana-Bahia. Os exemplares utilizados apresentaram conteúdo de sólidos solúveis na faixa de 13,11-13,20 °Brix, de modo a padronizar o estádio de maturação dos frutos em estudo. Os abacaxis foram descascados e sua polpa homogeneizada foi armazenada sob congelamento (-10 ºC), até a sua utilização nos experimentos 3.1.2 Planejamento experimental Com o objetivo de verificar a influência das variáveis pH e concentração de NaCl na extração das enzimas, foi elaborado um planejamento composto central com 4 repetições no ponto central, totalizando 12 ensaios. Como variáveis de resposta (dependentes) foram avaliadas a atividade das seguintes enzimas: PER, PFO, PME e PG. Os parâmetros de otimização utilizados encontram-se na Tabela 01. Tabela 01. Parâmetros da otimização pH NaCl (M) -α 4,0 0,0 -1 4,6 0,3 0 +1 +α 6,0 7,4 8,0 1,0 1,7 2,0 Para cada ensaio realizado, 50 g de polpa de abacaxi foram homogeneizados com 50 mL de tampão (0,1 M) -tampão fosfato de sódio para pH 6,0 7,4 e 8,0 e tampão acetato de sódio para pH 4,0 e 4,6- (0,1 M), segundo a Tabela 02, em um liquidificador doméstico, por 3 minutos. Depois, o material obtido foi centrifugado por 15 minutos a 4000 x g na 21 temperatura de 4ºC. O sobrenadante, denominado extrato enzimático, foi aliquotado e armazenado sob congelamento (-10 ºC) até o momento da análise. Tabela 2 - Matriz do planejamento composto central, com quatro repetições no ponto central. Ensaio 1 2 3 4 5 6 7 8 9 (C) 10 (C) 11 (C) 12 (C) pH 4,6 4,6 7,4 7,4 4,0 8,0 6,0 6,0 6,0 6,0 6,0 6,0 NaCl (M) 0,3 1,7 0,3 1,7 1,0 1,0 0,0 2,0 1,0 1,0 1,0 1,0 3.1.3 Determinação da atividade enzimática 3.1.3.1 Atividade de PER A atividade da enzima PER foi determinada em temperatura ambiente, usando-se cubetas de quartzo de 1 cm de caminho óptico. A reação foi lida a 470 nm, em espectrofotômetro UV/VIS, pela formação de tetraguaicol em 3,5 mL de meio reacional contendo: 3,3 mL de guaicol 0,1% (2-metoxi fenol) preparado em tampão fosfato de sódio (0,1 M; pH=6,5) e 0,2 mL de extrato enzimático. A reação foi iniciada pela adição de 0,1 mL de peróxido de hidrogênio 30% e foram realizadas leituras a cada 10 segundos por até 10 minutos. Uma unidade de atividade enzimática foi definida como a quantidade de extrato capaz de aumentar a absorbância em 0,001 unidades por minuto de reação e foi expressa em U/g de polpa (LIU et al., 2008; SHALINI et al., 2008). 3.1.3.2 Atividade de PFO A atividade de PFO foi avaliada segundo a metodologia modificada de Babu et al. (2008), Fang et al. (2007) e Kumar et al. (2008). Utilizou-se 1,5 mL de catecol (pirocatequina) 0,1 M em tampão fosfato pH=6,5 adicionados de 0,7mL de extrato enzimático que foram mantidos em banho a 30°C. A reação foi paralisada por adição de 22 0,8 mL de ácido perclórico (2 M) e acompanhou-se o aumento da absorbância a 420nm, em espectrofotômetro UV/VIS, fazendo leituras a cada 1 minuto por até 10 minutos. Uma unidade de atividade de PFO foi definida como a quantidade de extrato capaz de aumentar a absorbância em 0,001 unidade por minuto e foi expressa em U/g de polpa. 3.1.3.3 Atividade de PME A determinação da atividade da PME foi realizada através do método espectrofotométrico adaptado de Zocca et al. (2007) e Tiwari et al. (2009), utilizando-se 2 mL de solução de pectina cítrica (0,05% , pH=7,5), 0,25 mL de água destilada, 0,15 mL da solução de azul de bromotimol (0,04%) e 0,1 mL de extrato enzimático com pH ajustado para 7,5. Incubou-se em banho a 45 ºC por 24 horas. Usando uma cubeta de quartzo, foi feita a leitura no espectrofotômetro a 620 nm. Uma unidade de atividade enzimática foi definida como a quantidade de extrato capaz de reduzir 0,001 unidades de absorbância por minuto de reação e foi expressa em U/g de polpa. 3.1.3.4 Atividade de PG A atividade de poligalacturonase foi determinada segundo metodologia proposta por Zhou et al. (2000). A atividade de PG foi medida em um viscosímetro, pela mistura de 30 mL do extrato enzimático com 45 mL de ácido poligalacturônico 0,2% em tampão acetato de sódio (0,1 M; pH=4,4). Foi medida a viscosidade inicial e depois de 24 h de incubação a 45 ºC. Uma unidade de atividade de PG foi definida como a quantidade de extrato capaz de causar redução na viscosidade (em segundos) por g de polpa por hora de reação, nas condições do ensaio. 3.1.4 Análise Estatística A influência das condições de extração foi avaliada pela construção de superfície de resposta, utilizando o módulo Experimental Design e pela análise de variância (ANOVA) do software STATISTICA versão 7.0. 23 3.2 Experimento 2. Determinação da atividade enzimática em diferentes variedades de abacaxi. 3.2.1 Material Vegetal Foram analisadas três variedades de abacaxi: 1. Imperial: frutos inteiros adquiridos da Embrapa Mandioca e Fruticultura, 2. Vitória: polpa congelada adquirida da Embrapa Mandioca e Fruticultura, 3. Pérola: provenientes de duas fontes que serão identificadas como: 3.1 Pérola Embrapa: polpa congelada adquirida da Embrapa Mandioca e Fruticultura, 3.2 Pérola Mercado: frutos adquiridos no mercado local, Feira de Santana - Ba. Os frutos foram lavados com água potável e sabão neutro. Em seguida, foram descascados, homogeneizados em liquidificador e mantidos sob temperatura de congelamento até o momento das análises. As polpas oriundas da Embrapa Mandioca e Fruticultura também passaram por esse mesmo processo nos laboratórios da Embrapa. Foram avaliados 3 lotes (cada lote com 5 frutos) de cada variedade estudada e todas as análises foram realizadas em triplicata. 3.2.2 Determinação da atividade enzimática O extrato enzimático foi preparado utilizando uma proporção 1:2 de polpa e solução extratora otimizada no item 3.1.2. homogeneizados em liquidificador durante 3 minutos. O homogeneizado foi centrifugado a 4.000 x g em centrífuga refrigerada a 4ºC durante 15 minutos. As atividades de PER, PFO, PME e PG foram calculadas conforme descrito no item 3.1.3. 3.2.3 Análise estatística Foi realizada análise de variância (ANOVA) e teste de médias de Tukey utilizandose o programa SISVAR. Foram avaliadas as diferenças significativas (p<0,05) para as enzimas analisadas nas diferentes variedades de abacaxi. 24 3.3 Experimento 3. Otimização da inativação das enzimas PER e PFO 3.3.1 Material Vegetal O trabalho foi realizado com abacaxi cultivar Pérola adquiridos no comércio da cidade de Feira de Santana, Bahia. Os abacaxis foram descascados e sua polpa homogeneizada em liquidificador. O material obtido foi centrifugado por 15 minutos a 4000 x g a 4 ºC e armazenado sob congelamento (-10 ºC) até a sua utilização nos experimentos. 3.3.2 Planejamento experimental Foram usados 3 inibidores químicos (metabissulfito de sódio, ácido ascórbico e cloreto de cálcio). Dessa forma, foi elaborado um planejamento composto central (para cada inibidor), utilizando 3 variáveis independentes (tempo – 1, 3, 5, 7 e 9 min, temperatura – 27, 43, 57, 74 e 86 °C, e concentração do inibidor químico – 0, 40, 100, 160 e 200 mg/L) com 4 repetições no ponto central, totalizando 18 ensaios (Tabela 03) (RODRIGUES & IEMMA, 2005). Como variáveis de resposta (dependente) foram avaliadas a inativação das enzimas PER e PFO, em termos de porcentagem de inativação. Tabela 3. Matriz do planejamento composto central, com quatro repetições no ponto central. Ensaio Concentração(mg/L) Temperatura(°C) 1 2 3 4 5 6 7 8 9 10 11 12 13 14 40 40 40 40 160 160 160 160 0 200 100 100 100 100 43 43 74 74 43 43 74 74 57 57 27 86 57 57 Tempo (min) 3 7 3 7 3 7 3 7 5 5 5 5 1 9 25 15 (C) 16(C) 17(C) 18(C) 100 100 100 100 57 57 57 57 5 5 5 5 3.3.3 Determinação da inativação de PER e PFO A atividade das enzimas PER e PFO foi determinada segundo o item 3.1.3. O percentual de inativação enzimática foi calculado baseado na atividade da enzima sem tratamento aplicado. 3.3.4 Análise estatística A influência dos parâmetros na inativação das enzimas PER e PFO foram avaliadas pela construção de superfície de resposta, utilizando o módulo Experimental Design e pela análise de variância (ANOVA) no software STATISTICA versão 7.0. 3.3.5 Confirmação dos resultados Após a análise dos resultados e da escolha dos parâmetros para inativar PER e PFO, foi aplicado o tratamento escolhido como ideal nas variedades Imperial, Vitória e Pérola oriunda da Embrapa do item 3.2.1. Depois foi realizada a determinação da atividade das enzimas PER, PFO, PME e PG segundo o item 3.1.3. 3.4 Experimento 4. Determinação da atividade antioxidante e compostos fenólicos totais 3.4.1 Material Vegetal Foram utilizadas neste experimento as amostras descritas no item 3.2.1. As mesmas amostras passaram pelos tratamentos escolhidos como ideais para inativação de PER e PFO, a fim de avaliar a influencia do tratamento nos compostos fenólicos totais e na atividade antioxidante. 26 3.4.2 Extração dos compostos antioxidantes e fenólicos O procedimento de extração de compostos fenólicos totais e antioxidantes foi baseado em Alothman et al. (2009), com algumas modificações. Foram utilizados 100 g da polpa dos frutos, que foram submetidos à extração com 300mL de etanol a 95% com agitação magnética por 3 horas à temperatura ambiente e no escuro. Os extratos foram centrifugados a 4.000 x g por 15 minutos e os sobrenadantes foram coletados e concentrados, para remoção do solvente, em rota-evaporador a 50 ºC. Com o extrato etanólico obtido foi realizada a determinação de compostos fenólicos totais e a atividade antioxidante. 3.4.3 Atividade antioxidante A atividade antioxidante foi realizada pelo método DPPH (também chamado de capacidade de sequestrar o radical DPPH), utilizando-se o reativo 1,1-diphenil-2picrilhidrazil, usado como radical livre para reagir com os compostos antioxidantes presentes na polpa de abacaxi, segundo Brand-Williams et al., (1995), com modificações. Foram preparadas 5 diluições diferentes (20 a 400 mg/mL) do extrato etanólico, em triplicata. Transferiu-se uma alíquota de 0,05 mL de cada diluição do extrato para tubos de ensaio com 3,0 mL da solução metanólica do radical DPPH (45 µg/mL). Utilizou-se álcool metílico, como branco, para calibrar o espectrofotômetro. Após 15 minutos de incubação à temperatura ambiente, a redução do radical livre DPPH foi mensurada pela leitura da absorbância em 517 nm. O mesmo procedimento foi adotado com um padrão comercial, galato de propila, para efeito comparativo. A capacidade de sequestrar radical livre foi expressa como percentual de inibição de oxidação do radical e calculado conforme a Equação 01. Equação 01 % seqüestro = [(absorbância inicial da amostra – absorbância final da amostra) / (absorbância inicial do padrão – absorbância final do padrão) )]* 100 O valor de EC50 foi calculado através da equação da reta feita com base nas concentrações dos extratos e suas respectivas porcentagem de sequestro do radical DPPH. 27 3.4.4 Determinação do teor de fenóis totais O teor de fenóis totais foi determinado usando o reagente de Folin-Ciocalteau (SINGLETON & ROSSI, 1965; DEGÁSPARI et al., 2004; GEORGÉ et al., 2005), onde foi preparada a reação contendo uma alíquota de 0,5 ml do extrato fenólico (dissolvidos em etanol, a fim de se obter uma concentração de 20 mg de sólidos/mL), 2,5 ml de solução aquosa de Folin-Ciocalteau 10% e 2,0 ml de carbonato de sódio 7,5%. A mistura foi mantida em banho-maria a 50°C durante 5 minutos e então realizada a leitura por espectrofotometria a uma absorbância de 760 nm. O branco que foi utilizado como referência foi preparado substituindo o extrato fenólico por água destilada. Para a determinação dos fenólicos totais o mesmo procedimento foi realizado utilizando soluções padrão de ácido gálico, construindo-se uma curva com o composto em um intervalo de concentração de 1 a 50 mg/L e a quantidade de fenólicos totais foi expressa em em mg de ácido gálico em 100g de polpa. 3.4.5 Análise Estatística Foi realizada análise de variância (ANOVA) e teste de médias de Tukey utilizandose o programa SISVAR. 4.0 Resultados e Discussão 4.1 Experimento 01. Planejamento experimental e otimização do processo de extração enzimática Cada enzima atua sob condições fisiológicas de pH, temperatura e meio iônico. A extração enzimática é um processo no qual a enzima de interesse é transferida a partir da fase aquosa celular para outra fase, seja aquosa ou orgânica. Os reagentes mais utilizados na extração são NaCl e solução tampão (JUNIOR, 2001). Segundo Kamimura (2006), o NaCl aumenta a força iônica da solução tampão e isso possibilita ou melhora a extração das enzimas que estão ligadas ionicamente aos tecidos vegetais. 28 Com o intuito de otimizar o processo de extração das enzimas PER, PFO, PME e PG da polpa de abacaxi utilizou-se a metodologia de superfície de resposta para estudar o efeito do pH (de 4,0 a 8,0) e da concentração de NaCl (de 0,0 a 2,0 M) da solução tampão utilizada na extração das enzimas. A Tabela 04 mostra os resultados experimentais das atividades enzimáticas quando se fez variar os fatores pH e concentração de NaCl. Tabela 4: Resultado experimental em termos de atividade enzimática Ensaios pH 1 2 3 4 5 6 7 8 9 (C) 10 (C) 11 (C) 12 (C) 4,6 4,6 7,4 7,4 4,0 8,0 6,0 6,0 6,0 6,0 6,0 6,0 NaCl (M) 0,3 1,7 0,3 1,7 1,0 1,0 0,0 2,0 1,0 1,0 1,0 1,0 PER (U/g) 2984,60 3117,80 2811,70 3067,40 3039,70 3128,00 2273,30 2827,40 4752,90 2833,90 4511,00 4043,80 PFO (U/g) 49,43 81,86 46,86 66,38 80,71 62,24 33,48 21,76 40,48 36,62 29,95 38,29 PME (U/g) 2672,30 4406,00 1416,70 2441,30 4660,00 3347,30 1113,30 2505,30 7961,30 3109,30 2333,30 3117,30 PG (U/g.h-1) 0,000009 0,000034 0,000003 0,000032 0,000003 0,000036 0,000002 0,000032 0,000048 0,000029 0,000023 0,000023 A atividade máxima encontrada de PER foi de 4752,9 U/g. Brito et al. (2008) encontraram valores superiores de PER tanto para a cultivar Pérola (5624 U/g) quanto para cultivar Smooth Cayenne (5137 U/g). De acordo com Cardello et al. (1998) a atividade enzimática na polpa do fruto pode ser influenciada não apenas por fatores genéticos (variedade comercial de abacaxi), mas também pelo clima e condições meteorológicas, condições de cultura, entre outros. Para PFO as atividades encontradas variaram entre 21,76 e 81,86 U/g. Thé et al. (2001) estudaram atividade de PFO em diferentes estádios de maturação e armazenamento do cultivar Smooth Cayenne e as atividades encontradas variaram entre 25,81 U/g e 47,98 U/g, sendo os valores mais elevados aqueles em que o estádio de maturação estava avançado e sob refrigeração. Estes autores também estudaram a atividade de PME encontrando valores entre 87,50 e 312,50 U/g, valores bem inferiores ao encontrados neste trabalho que foram entre 1113,3 e 7961,3 U/g. Para PG foi encontrada uma atividade entre 0,000009 e 0,000048 U/g h-1. Thé et al. (2009), obtiveram uma atividade de PG variando entre 929,38 nmol.g -1 a 526,85 nmol.g-1 , 29 quando estudaram a influência da associação de tratamento hidrotérmico, cloreto de cálcio e atmosfera modificada sobre fatores relacionados à textura de abacaxi, cultivar Smooth Cayenne. Essas diferenças podem, no entanto, ser creditadas à metodologia de avaliação aplicada, que no caso de Thé et al. (2009) foi baseada na liberação de uronídeos redutores (atividade de exo-PG) e no presente caso foi baseada na redução de viscosidade da solução em 24 h (atividade de endo-PG). A literatura que trata da atividade de PG em abacaxi na sua maioria apresenta os resultados obtidos pela avaliação da liberação de açúcares redutores (ácidos galacturônico) de substâncias pécticas (ácido poligalacturônico) (Prado et al., 2003). Apesar da atividade de endo-PG (hidrólise do polímero de uma forma aleatória diminuindo a viscosidade, mas quase não aumentando o poder redutor da solução) ser detectada por este método, quando a atividade for muito alta, a liberação de açúcares redutores é melhor aplicada para a atvidade de exo-PG (ataca o polímero pelas extremidades, liberando unidades redutoras, mas mostrando pouco efeito sobre a viscosidade). Porém, durante este trabalho, não foi capaz de detectar qualquer atividade de PG, medindo a liberação de açúcares redutores. Observa-se pelos resultados encontrados que houve variação na atividade enzimática em todas as enzimas analisadas, comprovando o quanto a força iônica do meio e o pH influenciam a extração das enzimas. Assim sendo, é fundamental a escolha correta dessas duas variáveis para que se possa obter a quantidade máxima das enzimas na amostra. A influência das condições de extração para cada enzima foi avaliada pela construção de superfícies de resposta, nas quais foi possível determinar as regiões de valores em que se obteve uma maior extração de atividade enzimática. Os modelos propostos foram avaliados por análise de variância (ANOVA). 4.1.1 Peroxidase (PER) A superfície de resposta gerada para a atividade de PER está apresentada a seguir na Figura 06. Pela análise de variância (Tabela 05) conclui-se que o modelo está bem ajustado às observações, a regressão é estatisticamente significativa e serve para fins preditivos, com 99% de confiança. 30 Tabela 05. Análise de variância (ANOVA) para atividade de peroxidase Regressão Resíduo Falta de Ajuste Erro puro Total SQ 5566600 358063 GL 5 5 98192 3 259871 2 5924663 10 MQ Fcalc Ftab(1%) 1113320 15,5464 10,97 71612,6 R² 0,93 Figura 06. Superfície de resposta para atividade de PER. Com a superfície de resposta gerada, foi possível determinar que o processo de extração de PER foi otimizado. As condições mais indicadas para extração dessa enzima são: tampão em pH=6,0 contendo 1,0 M de NaCl. Kamimura (2006), estudando o efeito do pH da solução tampão da extração de PER de yacon, também encontrou como valor ideal para extração de pH 6. 31 4.1.2 Polifenol-oxidase (PFO) A análise de variância e o diagrama de Pareto obtidos para atividade de PFO encontram-se na Tabela 06 e na Figura 07, respectivamente. Tabela 06. Análise de variância (ANOVA) para atividade de polifenol-oxidase Regressão Resíduo Falta de Ajuste Erro puro Total SQ GL 3328,409 5 925,370 6 863,518 3 61,852 3 425,779 11 MQ 665,682 154,228 Fcalc 4,316 Ftab(10%) R² 3,11 0,7824 Pareto Chart of Standardized Effects pH(Q) 11,40277 (1)pH(L) -3,4393 (2)NaCl(L) 1Lby2L NaCl(Q) 2,754473 -1,4216 -,814135 p=,05 Standardized Effect Estimate (Absolute Value) Figura 07. Diagrama de Pareto para atividade da polifenol-oxidase. O coeficiente de determinação alcançado, R², foi de 0,7824, que significa que mais de 78% das variações nos resultados obtidos podem ser explicados pelo modelo ajustado e se devem às diferenças entre os tratamentos, no nível de 10% de probabilidade. No Diagrama de Pareto (Figura 07) é possível visualizar a influência dos fatores estudados sobre a atividade enzimática. No nível de 95% de confiança ficou demonstrado que 32 somente o pH influenciou a extração de PFO. Este resultado é um indicativo de que, na polpa da abacaxi cultivar Pérola, as isoformas de PFO encontram-se majoritariamente em solução e não em formas ionicamente ligadas. A superfície de resposta gerada (Figura 08) mostram que o processo de extração proposto não foi otimizado, porém, de acordo com os resultados obtidos dentre os valores estudados, há uma indicação de que maior atividade de PFO será obtida quando forem utilizados valores de pH inferiores a 4,6. Figura 08. Superfície de resposta para atividade de PFO. 4.1.3 Pectinametilesterase (PME) Na Tabela 07 e na Figura 09 encontram-se a análise de variância e o gráfico bidimensional obtidos para a atividade de PME. O valor do coeficiente de determinação de 0,89 indica que apenas 11% do total da variação das respostas obtidas não é explicado pelo modelo. O modelo é estatisticamente significativo e preditivo e descreve as respostas em função das variáveis analisadas com 95% de confiança. Tabela 07. Análise de variância (ANOVA) para atividade de pectinametilesterase Regressão Resíduo SQ 19,7462 2,4012 GL 5 5 MQ 3,9492 0,4802 Fcalc 8,2241 Ftab(5%) 5,05 R² 0,89 33 Falta de Ajuste Erro puro Total 1,8039 3 0,5973 22,1474 2 10 Figura 09. Superfície de resposta para extração de PME. A análise da superfície de resposta, (Figura 09) mostra que o processo de extração não se encontra otimizado. No entanto, dentro dos valores estudados observa-se que maior extração pode ser alcançada com concentração de NaCl de 1,0M. O valor de pH não foi considerado significativo para a extração de PME de acordo com o diagrama de pareto que se encontra no anexo 1 (Figura 1.e). 4.1.4 Poligalacturonase (PG) A validade do modelo obtido para a atividade de poligalacturonase foi verificada pela analise de variância (ANOVA) apresentada na Tabela 08. O coeficiente de determinação obtido foi de 0,77 e o modelo foi estatisticamente considerado significativo com confiança de 90%, assim foi possível utilizar o modelo gerado (Figura 10) para predizer as melhores condições de extração de PG. De acordo com o Diagrama de Pareto 34 para esse experimento (anexo 1, Figura 1.g), apenas o componente linear da variável concentração de NaCl foi considerado significativo. Assim, o modelo que mais se ajusta aos dados experimentais é o modelo não-quadrático (Figura 10). Tabela 08. Análise de variância (ANOVA) para atividade de poligalacturonase Regressão Resíduo Falta de Ajuste Erro puro Total SQ 2,7505 0,8186 GL 5 5 0,7586 3 0,0600 3,5691 2 10 MQ 0,5501 0,1637 Fcalc 3,4 Ftab(10%) 3,4 R² 0,77 Figura 10. Superfície de resposta para extração de PG. Como pode ser observado pelo modelo (Figura 10), o valor de pH apresenta pouca influência na extração de PG, no entanto, a atividade obtida aumenta com o aumento da concentração de NaCl, indicando que boa parte das isoformas dessa enzima se encontra ionicamente ligada ao tecido vegetal. 35 As condições escolhidas para a extração das enzimas para determinação enzimática em polpa de abacaxi foram: PER - pH=6,0 e concentração de NaCl de 1,0 M ; PFO - pH=4,5 ; PME - Concentração de NaCl de 1,0 M e pH=60 (como é usual a escolha de uma solução tampão para extração de enzimas e o pH não demonstrou influencia na extração de PME, entre os valores analisados, escolheu-se o pH=6,0, pois dessa forma seria feito uma só solução de extração para PME e PER); PG - Concentração de NaCl de 2,0 M e pH=4,6 (por também não sofrer influencia do pH na extração, entre os valores analisados, foi escolhido o pH=4,6 que é o utilizado na solução onde ocorre a reação enzimática na determinação de sua atividade). 4.2 Experimento 2. Determinação da atividade enzimática nas variedades de abacaxis 4.2.1 Peroxidase (PER) Os resultados apresentados na Tabela 09 são referentes à atividade da enzima peroxidase nos três lotes das diferentes amostras de abacaxi analisadas. Tabela 09. Atividade da enzima peroxidase em diferentes variedades de abacaxis. Lotes 1 2 3 Média* DP** PEROXIDASE (U/g) Pérola Pérola EMBRAPA Mercado 2066,67 3165,55 2930 1956,67 3006,67 3100 2013,33 3277,78 2913,3 2012,22 a 3150 c 2981,81 c 55,01 136,22 103,31 Imperial Vitória 2318,43 2287,35 2299,61 2301,80 b 15,65 *Valores assinalados com a mesma letra na linha entre as médias finais, não diferem significativamente (p<0,05), segundo o teste de Tukey. **Estimativa de desvio padrão. Com relação à enzima peroxidase, o abacaxi Pérola se destacou por apresentar maior atividade desta enzima, não havendo diferença significativa entre as duas amostras de Pérola analisadas seguindo da Vitoria e Imperial, (Figura 11). 36 c c b a Figura 11. Atividade de peroxidase em diferentes cultivares de abacaxi. Brito et al. 2001, estudaram a peroxidase no híbrido de abacaxi denominado IAC gomo de mel e seus clones IAC-1, IAC-2 e IAC-3, e encontrou atividades de 3194, 4065, 2751 e 3099 U/g, respectivamente. Antoniolli et al. (2003) determinaram a atividade de peroxidase em abacaxi Pérola minimamente processado, encontrando atividade inicial de 6480 U/g. Todos estes valores são maiores aos da atividade dos cultivares Vitória e Imperial aqui analisado. 4.2.2 Polifenol-oxidase (PFO) Os resultados apresentados na Tabela 10 são referente à atividade da enzima polifenol-oxidase nos três lotes das diferentes amostras de abacaxi analisadas. Tabela 10. Atividade da enzima polifenol-oxidase em diferentes variedades de abacaxis. Lotes 1 2 3 Média* DP** POLIFENOL-OXIDASE (U/g) Imperial Pérola Pérola EMBRAPA Mercado 17,14 29,04 30,00 15,71 32,38 27,14 14,29 30,95 28,57 15,71 a 30,79 c 28,57 c 1,43 1,67 1,43 Vitória 18,98 19,25 20,00 19,41 b 0,53 37 *Valores assinalados com a mesma letra na linha entre as médias finais, não diferem significativamente (p<0,05), segundo o teste de Tukey. **Estimativa de desvio padrão. Os valores mais altos de atividade da enzima polifenol-oxidase foram encontrados nas polpas do abacaxi Pérola Embrapa, diferindo significativamente das cultivares Imperial e Vitória (Figura 12), mas não foi significativamente diferente da outra variedade Pérola analisada oriunda do mercado de Feira de Santana. c c b a Figura 12. Atividade de polifenol-oxidase em diferentes cultivares de abacaxi. Thé et al., (2001) estudaram PFO do abacaxi Smooth Cayenne em diferentes estádio de maturação e encontraram valores que variaram de 25,81 a 28,43 U/g, valores superiores ao das variedades Imperial e Vitória, mas, inferiores ao da variedade Pérola, encontrados neste presente trabalho. Comparando os valores obtidos nesse estudo para a atividade das enzimas polifenol-oxidase e peroxidase nas diferentes cultivares estudadas e os dados descritos na literatura para abacaxi, é possível verificar a existência de diferenças na atividade destas enzimas. Considerando que não houve diferença significativa entre a variedade Pérola oriunda de dois lugares distintos, supõe então que estas diferenças não se devem a outros possíveis fatores de variação da atividade enzimática (condições climáticas, trato cultural, etc.), mas essas diferenças podem ser de origem genética, por se tratarem de cultivares diferentes. 38 Para a indústria de alimentos e até mesmo para o consumo in natura, encontrar variedades que possuam uma menor atividade das enzimas oxidativas é de fundamental importância. Os pigmentos escuros formados pela ação das polifenol-oxidases e peroxidases são geralmente acompanhados de mudanças indesejáveis na aparência e nas propriedades sensoriais do produto, resultando na diminuição da sua vida útil e do seu valor de mercado. Além disso, o problema do escurecimento enzimático sob o aspecto nutricional deve-se à possibilidade das o-quinonas interagirem com grupos amina e tiol, reduzindo a disponibilidade da lisina, metionina, tiamina e outros nutrientes essenciais (ARAÚJO, 1999). Dessa forma as variedades Imperial e Vitória se colocam como uma boa alternativa para a diversificação das variedades utilizadas na indústria de alimentos, onde predomina a Smooth Cayenne. Além da indústria, a exportação brasileira também ganha com essa menor atividade das enzimas oxidativas nessas duas novas variedades, pois diversos estudos (Zhou et al. (2003); Thé et al. (2001); Weerahewa & Adikaram (2005)) vem demonstrando que o principal problema do abacaxi na exportação (injúria pelo frio – escurecimento interno) está ligado diretamente às enzimas oxidativas aqui estudadas. 4.2.3 Pectinametilesterase (PME) A tabela 11 mostra a atividade da enzima pectinolítica PME nas diferentes variedades estudadas. Tabela 11. Atividade da enzima pectinametilesterase em diferentes variedades de abacaxis . Lotes 1 2 3 Média* DP** PECTINAMETILESTERASE (U/g) Imperial Pérola Pérola EMBRAPA Mercado 8660 3753,42 3073,33 9300 3888,89 2973,33 8860 2884,89 2720 8940 a 3509,07 b 2922,22 c 327,41 544,78 182,12 Vitória 9700 9853,33 9986,66 9866,6 a 143,44 *Valores assinalados com a mesma letra na linha entre as médias finais, não diferem significativamente (p<0,05), segundo o teste de Tukey. **Estimativa de desvio padrão. As variedades Imperial e Vitoria não apresentaram diferenças significativas para PME e foram superiores aos da variedade Pérola que apresentaram diferenças 39 significativas entre a variedade oriunda da Embrapa e a do mercado (Figura 13). Valores inferiores foram encontrados por Thé et al. (2006) estudando diferentes formas de armazenamento do abacaxi Smooty Cayenne, as atividades variaram entre 170 a 750 U/g. a a b c Figura 13. Atividade de pectinametilesterase em diferentes cultivares de abacaxi. 4.2.4 Poligalacturonase (PG) A tabela 12 apresenta a atividade de PG encontrada nas diferentes variedades de abacaxis estudadas. Tabela 12. Atividade da enzima poligalacturonase em diferentes variedades de abacaxis. Lotes 1 2 3 Média* DP** POLIGALACTURONASE (U/g.h-1) Imperial Pérola Pérola EMBRAPA Mercado 0,00001898 0,00000494 0,00000297 0,00001898 0,00000440 0,00000304 0,00001574 0,00000417 0,00000289 0,00001790 a 0,00000450b 0,00000297c 0,00000187 0,00000039 0,00000008 Vitória 0,00003657 0,00003472 0,00003287 0,00003472 a 0,00000185 *Valores assinalados com a mesma letra na linha entre as médias finais, não diferem significativamente (p<0,05), segundo o teste de Tukey. **Estimativa de desvio padrão. Não houve diferença significativas entre as novas variedades estudadas, sendo que essas variedades (Imperial e Vitória) apresentaram maior atividade que a variedade Pérola. 40 A metodologia viscosimétrica (utilizada neste experimento) não foi encontrada na literatura para uso em atividade de PG em abacaxi, o que dificulta a comparação da nossa atividade com os outros métodos, pois cada método possui sua unidade específica para a atividade enzimática. Santana (2009) utilizou este método para pêssego Douradão e encontrou valores que variaram de 0,001 a 0,005 U/g. h-1, valores superiores aos encontrados nesse estudo (Figura 14). a a b c Figura 14. Atividade de poligalacturonase em diferentes cultivares de abacaxi. Conforme as Tabelas 11 e 12, houve diferença significativa (p<0,05) dentro da variedade Pérola (Embrapa e Mercado) tanto para PG como PME, possivelmente devido ao fato dessas frutas apresentarem origens distintas considerando, portanto vários fatores, como condições climáticas, condições de cultivo, condições de armazenamento, entre outros. É conhecido que esses fatores influenciam a constituição química das frutas, e por isso devem ser considerados neste tipo de estudo (CARDELLO & CARDELLO, 1998). Não podendo descartar a influencia significativa do estádio de maturação dos frutos que possui relevância nas atividades das enzimas. A ação da pectinametilesterase libera ácido poligalacturônico, substrato para a ação da poligalacturonase, enzima despolimerizante, associada à perda de textura em frutas e vegetais estocados (NIELSEN & CHRISTENSEN, 2002). A interação do ácido pectínico resultante da ação da PME com íons cálcio, com formação de pectato de cálcio insolúvel, causa perda da estabilidade da turbidez, com 41 consequente clarificação do suco (ASSIS et al, 2000), no caso do suco de abacaxi, a turbidez é característica do suco, e esta clarificação causa perda comercial do produto. Assim, a alta atividade de PME nas variedades Imperial e Vitória sugere que estas variedades, ao serem usadas na indústria de alimentos, principalmente de suco concentrado, tenham o processo de inativação destas enzimas bem definidos, de modo a evitar a perda do valor comercial do produto. Apesar das variedades Imperial e Vitória apresentarem uma alta atividade das enzimas PME e PG, elas ainda devem ser recomendadas para a industrialização de abacaxi, pois é mais vantajoso para a indústria ter uma variedade com menor atividade da enzima PER (mais termorresistente e de difícil inativação), do que PME e PG que são enzimas que podem ser mais facilmente inativas través do uso de tratamento térmico. 4.3 Experimento 3. Otimização da inativação enzimática de PFO e PER Na Tabela 13 encontram-se os resultados do planejamento experimental para a inativação enzimática de PER e PFO em polpa de abacaxi Pérola. Tabela 13. Resultados experimentais em termos de porcentagem de inativação enzimática ao adicionar ácido ascórbico, metabissulfito de sódio e cloreto de cálcio. PER PFO % % % % % % Inativação Inativação Inativação inativação Inativação Inativação (ácido (Metabissulfio (cloreto de (ácido (Metabissulfito (cloreto de ascórbico) de sódio) cálcio) ascórbico) de sódio) cálcio) 1 21,38 18,25 22,24 47,54 32,78 36,89 2 21,59 40,46 31,61 56,55 54,91 47,54 3 65,93 70,58 47,39 81,14 77,86 77,05 4 72,72 90,23 54,89 92,62 90,98 86,07 5 31,88 36,02 28,20 63,11 63,93 49,18 6 30,78 31,98 34,19 67,21 80,32 54,92 7 67,35 80,33 59,87 93,44 84,42 82,79 Ensaio 42 8 83,04 85,14 62,42 96,72 95,08 90,16 9 0,00 0,00 32,53 43,44 50,82 52,46 10 39,56 16,16 5,54 69,67 77,04 67,21 11 12,28 36,32 18,47 50 58,19 55,74 12 100,00 100,00 100,00 100,00 100,00 100,00 13 18,36 24,27 29,65 59,01 58,19 59,84 14 33,33 37,47 29,47 68,85 65,57 68,85 15(C) 23,25 31,51 25,14 60,65 62,29 72,95 16(C) 25,30 31,91 27,37 62,29 61,47 72,13 17(C) 23,19 31,54 25,97 61,47 61,47 71,31 18(C) 24,32 32,16 27,46 62,29 60,65 70,49 Observa-se na Tabela 13 que a taxa de porcentagem para a inativação de PFO foi maior do que a de PER, em todos os inibidores utilizados. A grande estabilidade térmica é uma característica comum às peroxidases de várias frutas. Alguns autores atribuem esta grande estabilidade térmica à presença de açúcares na molécula da enzima (POMAR et al, 1997). Também foi relatado que a presença de um grande número de resíduos de cisteína na cadeia polipeptídica contribui para a termoestabilidade das peroxidases (DEEPA, 2002) Em todos os delineamentos só se obteve 100% de inativação quando foi aplicada a temperatura de 86 °C (ensaio 12). Brito et al. (2005) conseguiram inativar peroxidases de abacaxi 'IAC Gomo-de-mel' e „IAC-1‟ com tratamentos a 60 e 120 segundos a 90 ºC. Lourenço & Neves (1997) observaram que a peroxidase de pêssego apresentou rápida inativação em temperaturas acima de 70º C, com perda praticamente total após tratamento a 80º C durante 30 segundos. A influência dos parâmetros de inativação para PER e PFO foi avaliadas pela construção de superfícies de resposta, nas quais foi possível determinar as regiões de valores em que se obteve uma maior inativação enzimática destas enzimas. Os modelos propostos foram avaliados por análise de variância (ANOVA) que se encontram na Tabela 14 e 15. 43 Tabela 14. Análise de variância para porcentagem de inativação de PER. SQ GL MQ Regressão 11672,19 7 1667,45 Resíduo 977,47 10 97,747 Ácido Falta de ajuste 974,46 7 Ascórbico Erro Puro 3,01 3 Total 12649,66 17 Regressão Metabissulfito De Sódio Cloreto de Cálcio F calc F tab (1%) R² 17,05 5,20 0,92 12136,28 9 1348,475 7,010 5,91 0,88 Resíduo 1538,82 8 192,3525 Falta de ajuste 1538,53 5 Erro Puro 0,29 3 Total 13675,10 17 Regressão 6813,526 7 973,36 5,20 0,87 Resíduo 993,36 10 99,336 Falta de ajuste 989,576 7 Erro Puro 3,784 3 Total 7806,886 17 9,7986 Tabela 15. Análise de variância para porcentagem de inativação de PFO. SQ GL MQ Regressão 4700,223 7 671,460 Resíduo 314,26 10 31,426 Ácido Falta de ajuste 312,411 7 Ascórbico Erro Puro 1,849 3 Total 5014,483 17 Regressão 4802,126 9 533,56 F calc F tab (1%) R² 21,366 5,20 0,937 25,24 5,91 0,965 44 Metabissulfito Resíduo 169,059 8 De Falta de ajuste 167,715 5 Sódio Erro Puro 1,344 3 Total 4971,185 17 Regressão 4450,718 7 635,816 Resíduo 118,133 10 11,813 Falta de ajuste 114,774 7 Erro Puro 3,359 3 Total 4568,85 17 Cloreto de Cálcio 21,1323 53,82 5,20 0,987 Observou-se que os modelos empíricos estabelecidos nos planejamentos executados para PER e PFO mostraram boa concordância com os dados experimentais obtidos. Todos obtiveram uma regressão significativa, pois o F calculado foi superior ao F tabelado (1%). A partir das equações de regressão obtiveram-se as superfícies de resposta. 4.3.1 Peroxidase (PER) Todas as superfícies de respostas e os gráficos bidimensionais de contorno para a inativação de PER encontram-se no anexo 2. Na Figura 15, encontra-se a superfície de resposta para o planejamento utilizando ácido ascórbico como inibidor químico de PER. Analisando este gráfico, observa-se que a temperatura mínima para se obter 100% de inativação de PER é 78,41°C, e a concentração de ácido ascórbico dependerá do tempo de tratamento empregado, sendo que quanto menor o tempo maior a concentração requerida para 100% de inativação. Utilizando o modelo proposto chega-se a várias possibilidades para alcançar 100% de inativação, entre elas encontram-se as combinações: 78,41°C, 1 min e 200 mg/L de ácido ascórbico; 78,41°C, 3 min e 174 mg/L de ácido ascórbico; e 78,41 °C, 5 min e 145 mg/L de ácido ascórbico. 45 Figura 15. Superfície de resposta para o efeito temperatura e concentração de ácido ascórbico na inativação enzimática de PER. Na Figura 16, encontra-se o gráfico bidimensional de contorno quando se aplicou metabissulfito de sódio como inibidor químico para PER. Observa-se que a temperatura para se alcançar 100% de inativação dependerá do tempo e da concentração de metabissulfito de sódio empregado. Entre o limite testado, conseguimos 100% de inativação a partir de uma temperatura de 77,52 °C com uma concentração de metabissulfito de 130 mg/L. Figura 16. Gráfico bidimensional de contorno para os efeitos temperatura e concentração de metabissulfito de sódio na inativação enzimática de PER. 46 Na Figura 17, encontra-se a superfície de resposta para a aplicação do cloreto de cálcio na inativação de PER. Analisando o gráfico obtido, observa-se que 100% de inativação é alcançado com uma temperatura de 84°C independente da concentração e do tempo aplicado no tratamento. GONÇALVES et al. (2000) verificaram a influência do tratamento hidrotérmico com a aplicação de cloreto de cálcio em abacaxis Smooth Cayenne. Eles alcançaram a inativação da enzima peroxidase fazendo a imersão dos frutos de abacaxi em uma solução a 1% de cloreto de cálcio em água aquecida (38 ºC ou 40 °C) por 10 ou 20 minutos. Os autores atribuíram a inativação da peroxidase à ação do cálcio na manutenção da integridade celular, reduzindo o contato da enzima com seu substrato. Essa justificativa, entretanto, não deve ser atribuída ao experimento aqui feito, pois utilizou-se polpa homogeneizada, desta forma a enzima já estava em contato com o substrato. Figura 17. Superfície de resposta para os efeitos temperatura e concentração de cloreto de cálcio para inativação enzimática de PER. CLEMENTE (1995) em seu trabalho com isoenzimas de laranja, não conseguiu inativar peroxidase com tratamento de 70ºC e elevando a temperatura para 80 ºC não houve alteração no comportamento da atividade enzimática, mostrando estas temperaturas serem ineficientes para a inativação da peroxidase. Neste trabalho, segundo os modelos propostos, 100% de inativação de peroxidase é alcançada com uma temperatura de 77,52 ºC, isso pode ser atribuído ao efeito benéfico dos 47 agentes químicos empregados nos tratamentos. Quanto menor for o binômio tempo/temperatura a ser aplicado para inativação enzimática, melhor para a indústria de alimentos. Temperaturas e tempos longos podem levar à destruição de vitaminas e à formação de sabor de cozido, com rejeição do consumidor (KOBLITZ, 2008). O metabissulfito de sódio apresentou a melhor resposta de inativação para PER, pois a uma concentração mais baixa e tratamento térmico mais brando, do que quando comparados com os outros inibidores, atingiu-se 100% de inativação enzimática, sem necessitar longo tempo de exposição. Isso representa as vantagens de reduzido emprego de aditivo (diminuindo o uso de altas dosagens que podem provocar gosto e cheiro desagradáveis, além de causarem crises alérgicas em pessoas asmáticas tendo o seu uso controlado pela legislação, no caso do metabissulfito de sódio) e diminuição dos danos causados por tratamento térmicos severo (perdas de vitaminas e sabor de cozido). 4.3.2 Polifenol-oxidase (PFO) Todas as superfícies de respostas e os gráficos bidimensionais de contorno para a inativação de PFO encontram-se no anexo 3. Na Figura 18 encontra-se o gráfico de superfície de resposta para porcentagem de inativação de PFO utilizando ácido ascórbico. Figura 18. Superfície de resposta para porcentagem de inativação de PFO utilizando ácido ascórbico. 48 Observa-se pelo gráfico que à medida que vai aumentado a concentração de ácido ascórbico dimínui-se a temperatura para alcançar 100% de inativação de PFO. Com uma temperatura de 75 °C e concentração de ácido de 200 mg/L é alcançado 100% de inativação. Na Figura 19 encontra-se a superfície de resposta para a inativação de PFO com metabissulfito de sódio. Figura 19. Gráfico Bidimensional de contorno para porcentagem de inativação de PFO utilizando metabissulfito de sódio. O comportamento da inativação de PFO com o metabissulfito de sódio, mostra que são necessárias temperaturas mais altas para se conseguir 100 % de inativação. Todas as variáveis estudadas foram significativas, e a interação entre elas também. Como o principal interesse é conseguir temperaturas mais brandas para a inativação enzimática, com o metabissulfito o modelo indica que isso ocorre com 79 °C, porém com o máximo de metabissulfito de sódio aplicados nos tratamentos (200 mg/L) e tempo de 5 min. Na figura 20, encontra-se a superfície de resposta para a inativação de PFO aplicando cloreto de cálcio. 49 Figura 20. Superfície de resposta para porcentagem de inativação de PFO utilizando cloreto de cálcio. As interações entre as variáveis estudadas aplicando cloreto de cálcio não foram significativas, sendo excluídas do modelo. Observa-se pela superfície de resposta (Figura 20) que a medida que vai aumentado a concentração de cloreto de cálcio diminui a temperatura necessária para se obter 100% de inativação de PFO, porém chega-se um ponto, que aumentando a concentração, aumenta também a temperatura necessária para inativar PFO, demonstrado um efeito negativo de se usar altos valores de cloreto de cálcio para inativar PFO. Alcança-se 100 % de inativação de PFO com aplicação de 100 mg/L de cloreto de cálcio a uma temperatura de 82 °C por 5 min. O cloreto de cálcio não é um inibidor direto da enzima PFO. Mas seu uso é estudado pois sabe-se que é a perda da integridade da parede celular que representa o início das reações de escurecimento enzimático. É o corte, a queda, a ação de pectinases, hemicelulases e celulase que levam à perda da integridade da parede celular dos vegetais, e por consequência, geralmente ao início das reações de escurecimento enzimático. Soluções de sais de cálcio ajudam a manter a parede celular em bom estado: os íons de cálcio ligamse às cadeias de pectina, formando pontes entre elas, aumentando sua força e formando pectato de cálcio, impedindo que o substrato se encontre com a enzima (POOVAIAH, 1986; RENSBURG & ENGELBRECHT, 1986). No presente estudo, foi utilizada polpa 50 homogeneizada, isso faz com que a justificativa da integridade da parede celular conferida pelo cálcio seja descartada. Mas, apesar da inativação com cloreto de cálcio ter apresentado os piores resultados (entre os inibidores testados), a presença do inibidor não pode ser descartada, pois houve um efeito benéfico na inativação. Koblitz (2008) relata a inativação da enzima PFO pela ação de halogenetos, principalmente os sais de cloro, sendo o CaCl2 comercializado, em conjunto com ácido ascórbico, em um preparado comercial inibidor de PFO de excelente eficácia. Fujita et al., 1988, estudou a inativação de PFO em berinjela, a enzima ficou completamente inativada após 30 minutos a 75°C ou 5 minutos a 80°C. Neste presente trabalho, a temperatura mais branda para a inativação de PFO também foi de 75 °C, porém o tempo empregado é de apenas 3 min, confirmando o efeito benéfico de empregar inibidores químicos na inativação de enzimas associado ao binômio temperatura/tempo. O melhor resultado para inativar PFO foi usando ácido ascórbico, pois foi necessária menor temperatura para 100 % de inativação. Na Tabela 16, encontra-se o melhor resultado para cada inibidor químico para as enzimas PER e PFO. Tabela 16. Resumos dos melhores resultados obtidos para inativação das enzimas PER e PFO. Inibidores Ac. ascórbico Metabissulfito de sódio Cloreto de Cálcio T Concen- Tempo T Concen- Tempo T Concen- Tempo (°C) tração (min) (°C) tração (min) (°C) tração (min) (mg/L) (mg/L) (mg/L) PER 79 174 3 78 130 3 84 0 1 PFO 75 190 3 79 200 5 82 100 5 Escolheram-se os 2 melhores tratamentos para inativar PER e PFO, avaliando principalmente o binômio tempo/temperatura, pois é a principal causa de danos ao produto final (sabor de cozido, perdas de nutrientes). Para PER tratamento com 130 mg/L de metabissulfito de sódio a 78º C por 3 min e para PFO tratamento com 190 mg/L de ácido ascórbico a 75° C por 3 min. 51 4.3.3 Confirmação dos resultados O tratamento ideal para PER (130 mg/L de metabissulfito de sódio a 78º C por 3 min ) e para PFO (190 mg/L de ácido ascórbico a 75° C por 3 min) foram aplicados nas variedades citadas no item 3.2.1 e depois foi determinada as atividades das enzimas PG, PME, PFO e PER. Os resultados encontram-se nas tabelas 17 e 18. Tabela 17. Atividade enzimática após aplicação do tratamento para inativação enzimática com 130 mg/L de metabissulfito de sódio a 78 °C por 3 min. Imperial Vitoria Pérola Embrapa Pérola Mercado PFO (U/g) PG (U/g.h-1) PME (U/g) PER (U/g) 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 Tabela 18. Atividade enzimática após aplicação do tratamento para inativação enzimática com 190 mg/L de ácido ascórbico a 75 °C por 3 min. Imperial Vitória Pérola Embrapa Pérola Mercado PFO (U/g) PG (U/g.h-1) PME (U/g) PER (U/g) 0 0 0 312,12 0 0 0 445,35 0 0 0 789,56 0 0 0 659,12 Observa-se que os resultados encontrados pelos modelos propostos foram confirmados, o tratamento do metabissulfito conseguiu inativar peroxidase e também as outras enzimas, e o tratamento com ácido ascórbico conseguiu inativar a PFO e outras enzimas, com exceção da PER. 52 A peroxidase é umas das enzimas mais termoestáveis em vegetais. Considera-se que a inativação da peroxidase é acompanhada pela desnaturação de todas as outras enzimas (BUENETTE, 1977; KHAN & ROBINSON, 1993), fato comprovado neste experimento: o tratamento recomendado de inativação aplicado para PER inativou todas as outras enzimas, já o melhor tratamento pra PFO não foi suficiente para inativar completamente PER. Mello & Clemente (1996), conseguiram inativar PER de abacaxi com uma temperatura de 75ºC, (a mesma utilizada pelo melhor tratamento de PFO), porém eles usaram um tempo de 10 min, bem superior ao utilizado neste experimento, 3 min. A PER parcialmente purificada de tomate mostrou-se menos termorresistente que a de abacaxi, sendo completamente inativada após 10 segundos a 72ºC (ANTHON et al, 2002). 4.4 Experimento 4. Compostos fenólicos e atividade antioxidante 4.4.1 Compostos fenólicos totais Houve diferença significativa do conteúdo de compostos fenólicos totais entre as variedades estudadas e dentro da mesma variedade Pérola (Tabela 19). A variedade Pérola foi a que apresentou maior teor de fenólicos totais, com 5,66 e 5,16 mg de ácido gálico/100g, respectivamente para o Pérola Mercado e Pérola Embrapa. Conteúdos semelhantes de fenólicos totais foram encontrados por Silva (2010), que estudou híbridos de abacaxi sob diferentes tratamentos do solo. Os valores médios dos fenólicos variaram entre 4,34 a 5,80 mg de ácido gálico 100g-1. Porém, Kuskoski et al. (2006) e Melo et al (2008), encontraram valores bem superiores em polpa de abacaxi congelada (21,7 mg/100g) e em abacaxi Pérola (66,3 mg/100g). Essa variação no teor de fenólicos totais é explicada pois o conteúdo de compostos fenólicos é influenciado pelos fatores de pré-colheita, como cultivar, ocorrência de estresses, localização geográfica e outros (ZADERNOWSKI et al., 2005). 53 Tabela 19. Teor de fenóis totais em diferentes variedades de abacaxi. Fenólicos Totais (mg de ácido galico/100 g de polpa) Imperial Vitória Amostra pura 4,90 A a Amostra c/ metabissulfito de sódio Amostra c/ Ac. Ascórbico 4,87 A b Pérola Embrapa 5,16 A c Pérola mercado 5,66 A d 5,4 B 5,37 B 5,98 B 7,01 B 6,19 C 6,73 C 7,18 C 8,97 C Valores assinalados com a mesma letra maiúscula na mesma coluna, não diferem significativamente (p<0,05), segundo o teste de Tukey. Valores assinalados com a mesma letra na primeira linha, não diferem significativamente (p<0,05), segundo o teste de Tukey. Analisando o teor de fenólicos totais depois dos tratamentos aplicados para inativação enzimática, observa-se que houve aumento na quantidade de fenólicos totais havendo diferença significativa nos valores encontrados antes e depois dos tratamentos na mesma variedade. Em todas as variedades, o tratamento com ácido ascórbico foi o que obteve maior conteúdo de fenólicos totais. Vale ressaltar que os tratamentos aplicados obtiveram no experimento de inativação enzimática 100% de inativação de PFO, enzima que tem os compostos fenólicos como substrato o que pode justificar o aumento significativo dos compostos fenólicos dentro da mesma amostra depois que estas passaram pelo processo de inativação enzimática. Essa hipótese é confirmada pelo fato de que, na presença de metabissulfito, o teor de fenólicos foi maior do que na amostra sem tratamento porém inferior ao encontrado na amostra tratada com ácido ascórbico. Verifica-se portanto, que pode haver uma importante influência da presença de enzimas nativas do fruto na extração de compostos fenólicos da polpa. É possível que seja aconselhável o uso de inibidores químicos, prévios à extração, para garantir a manutenção do conteúdo de fenólicos original. 4.4.2 Atividade Antioxidante Os resultados obtidos dos testes frente ao método do radical livre DPPH estão apresentados na Tabela 20 e permitem fazer uma comparação do potencial antioxidante entre as variedades de abacaxi estudadas. 54 Tabela 20. Valores do EC50 dos extratos etanólico de abacaxi. Imperial EC50 (mg/mL) Vitória Pérola embrapa 315,44 b A 263,11 d A Pérola mercado 288,98 c A Amostra pura 326,51 a A Amostra c/ metabissulfito de sódio 301,02 B 289,22 B 249,21 B 187,22 B 257,28 C 191,24 C 160,89 C 138,63 C Amostra c/ Ac. Ascórbico Valores assinalados com a mesma letra maiúscula na mesma coluna, não diferem significativamente (p<0,05), segundo o teste de Tukey. Valores assinalados com a mesma letra na primeira linha, não diferem significativamente (p<0,05), segundo o teste de Tukey. EC50 é definida como a quantidade de amostra suficiente para reduzir a concentração inicial de DPPH para 50%. As substâncias antioxidantes presentes nos extratos reagem com o DPPH, que é um radical estável, e convertem-no em 2,2-difenil-1-picril hidrazina provocando descoloração da solução. O grau de descoloração indica o potencial antioxidante do extrato. Um extrato que apresenta alto potencial em sequestrar radicais livres possui baixo valor de EC50. Desta forma, uma pequena quantidade de extrato é capaz de decrescer a concentração inicial do radical DPPH em 50%, ou seja, inibir a oxidação do radical em 50%. A variedade que apresentou a maior capacidade de sequestrar o radical DPPH foi a Pérola Mercado (EC50=288,98 mg/L) seguida da Pérola Embrapa (EC50=263,11 mg/mL) havendo diferença significativa entre elas. Avaliando o efeito do tratamento de inativação enzimática nas amostras, observa-se que houve aumento na capacidade antioxidante dessas amostras. Este resultado já era esperado para os tratamentos aplicados, diante de que esses aditivos são antioxidantes conhecidos. Porém, o aumento da capacidade antioxidante nessas amostras pode estar relacionada também à inibição enzimática e preservação dos fenólicos quando aplicou-se estes tratamentos, sugerindo a relação entre compostos fenólicos e atividade antioxidante. Desta forma, os estudos realizados indicam a relação entre concentração de fenóis totais e a capacidade de sequestrar radicais livres das variedades de abacaxi, visto que as amostras com maior concentração de fenóis totais são justamente as amostras com maior atividade antioxidante. A correlação entre compostos fenólicos totais e atividade antioxidante entre as diferentes variedades de abacaxi foi de 0,544, valores que corroboram 55 com Vedana (2008) que encontrou uma correlação de 0,540 para fenólicos totais e atividade antioxidante em uvas. A correlação entre fenólicos totais e atividade antioxidante na mesma variedade comparando os tratamentos aplicados foi superior em todas as variedades estudadas, variando de 0,95 a 0,99. Benvenuti et al. (2004) encontrou coeficiente de correlação para DPPH e fenóis totais de 0,992, Roesler (2007) também encontrou relação entre concentração de fenóis totais e a capacidade de sequestrar radicais livres dos extratos de frações das frutas do cerrado. O potencial antioxidante de frutas está intimamente relacionado com a composição específica dos diferentes tipos de compostos fenólicos, uma vez que a atividade antioxidante ótima depende da sua configuração especial e do número e da posição dos grupos hidroxilas presentes na sua estrutura (KUSKOSKI et al., 2006). 5.0 Conclusão O delineamento experimental e a metodologia de superfície de respostas se mostraram métodos eficazes para otimizar o processo de extração enzimática. Observou-se que a concentração de NaCl e o pH da solução tampão de extração influenciam na extração das enzimas. Os resultados mostraram que o pH foi significativo na extração das enzimas PER e PFO, e o NaCl foi significativo na extração de PME e PG. As variedades Imperial e Vitoria são boas alternativas para a industrialização de abacaxi. Apesar de apresentarem alta atividade das enzimas pectiniloticas ( que afetam a textura e turbidez dos alimentos), elas possuem uma menor atividade das enzimas oxidativas (responsáveis pelo escurecimento interno e reações de degradação). Sendo a PER a enzima mais termoresistente, é mais vantajoso ter uma menor atividade desta enzima do que as enzimas pectinolítcas que são mais termolábeis. Estes resultados também servirão de incentivo aos programas de melhoramento genético do abacaxi, que se prendiam em obter cultivares mais produtivas, adaptadas às condições climáticas locais e resistentes a pragas e moléstias, mas não se preocupavam com outros fatores, como características bioquímicas que darão ao fruto possibilidade de uso industrial. 56 Os inibidores químicos utilizados se mostraram eficazes na inativação das enzimas oxidativas, pois reduziram a temperatura necessária para a inativação térmica, comparada com a literatura. O melhor tratamento para inativar PER foi com metabissulfito de sódio, que apresentou a melhor resposta de inativação, pois a uma concentração mais baixa e com tratamento térmico mais brando, do que quando comparados com os outros inibidores, atingiu-se 100% de inativação enzimática, sem necessitar longo tempo de exposição. Isso representa vantagens de reduzido emprego de aditivo e diminuição dos danos causados por tratamentos térmicos severos. Para PFO o tratamento com ácido ascórbico foi o mais indicado para alcançar 100% de inativação. Ao se aplicar estes tratamentos em todas as variedades e estudando todas as enzimas (PER, PFO, PG e PME), observase que o melhor tratamento para PFO não foi eficaz para inativar totalmente a PER, confirmando a termoestabilidade desta enzima. Os tratamentos utilizados para inativação enzimática tiveram influencia positiva no teor dos fenóis totais em todas as variedades de abacaxis estudadas, possivelmente pelo fato da inativação das enzimas oxidativas e permanência dos compostos fenólicos no abacaxi. Houve uma correlação moderada entre os compostos fenólicos e a capacidade de sequestrar o radical DPPH entre as variedades estudadas e uma correlação alta (0,95-0,99) dentro da mesma variedade comparando os tratamentos aplicados. 57 6.0 Referências ABREU, C. M. P. de. Efeito da embalagem de polietileno e da refrigeração no escurecimento interno e composição química durante a maturação do abacaxi cv. Smooth Cayenne. 1995. 94f. Tese (Doutorado em Ciência dos Alimentos) - UFLA, Lavras, Minas Gerais. ALOTHMAN, M.; BHAT R.; KARIM, A. A. Antioxidant capacity and phenolic content of selected tropical fruits from Malaysia, extracted with different solvents. Food Chemistry, v. 115, p. 785–788, 2009. ANTHON, G. E.; SEKINE, Y.; WATANABE, N. Thermal inactivation of pectin methylesterase, polylagacturonase, and perosidase in tomate juice. Journal of Agricultural and Food Chemistry, v. 50, p. 6153-6159, 2002. ANTONIOLLI, L. R.; BENEDETTI, B. C.; SOUZA F. M. de S. M. de. Effect of calcium chloride on quality of freshcut 'Pérola' pineapple. Pesquisa Agropecuária Brasileira, v. 38, n. 9, p. 1105-1110, 2003. AO, C. et al. Evaluation of antioxidant and antibacterial activities of Ficus microcarpa L. fil. Food Control, v. 19, p. 940–948, 2008. ARAÚJO, J. M. A. Química de alimentos: teoria e prática. 2. ed. Viçosa: UFV, 1999. ASSIS, S. A.; LIMA, D. C.; OLIVEIRA, O. M. M. F. Acerola´s pectin methyl esterase: studies of heat inactivation, Food Chemistry, v. 71, p. 465-467, 2000. BABU, B. R.; RASTOGI, N. K.; RAGHAVARAO, K. S. M. S. Liquid–liquid extraction of bromelain and polyphenol oxidase using aqueous two-phase system. Chemical Engineering and Processing, v. 47, p. 83–89, 2008. BALASUNDRAM, N.; SUNDRAM, K.; SAMMAN, S. Phenolic compounds in plants and agriindustrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chemistry, v. 99, p. 191-203, 2006. 58 BAKER, R. A.; CAMERON, R. G. Clouds of citrus juices and drinks. Food technology, v. 53, p. 64-69, 1999. BARBOSA, J. M. D. Influência da areia argilosa na recuperação de petróleo por injeção de vapor. 2009. 143f. Dissertação (Mestrado em Ciência e Engenharia do Petróleo) – Universidade Federal do Rio Grande do Norte, Natal. BARREIROS, A. L.; DAVID, J. M.; DAVID, J. P. Estresse oxidativo: relação entre geração de espécies reativas e defesa do organismo. Quimica Nova, v. 29, p. 113-123, 2006. BARROS NETO, B.; SCARMINIO, I. S.; BRUNS, R. E. Como fazer experimentos: pesquisa e desenvolvimento na ciência e na indústria. 2. ed. Campinas: Editora da UNICAMP, 2002. 401p. BARROS NETO, B; SCARMÍNIO, I. S; BRUNS, R. E. Como fazer experimentos. 3. ed. Campinas: Editora da Unicamp, 2007. 480p. BEAUDREAU, C.; YASUNOBU, K. T. Heme Proteins. VI Crystaline pineapple peroxidase B . Washington DC, v. 5, p. 1404-1412, 1966. BENVENUTI, S. et al. Polyphenols, Anthocyanins, Ascorbic Acid, and Radical Scavenging Activity of Rubus, Ribes and Aronia. Food Chemistry and Toxicology, v. 69, p. 164-169, 2004. BOAS, E. V. B. et al. Modificações Texturais de tomates Heterozigotos no Loco Alcobaça. Pesquisa Agrapecuária Brasileira, v. 35, p. 1447, 2000. BONDET, V.; BRAND-WILLIAMS, W.; BERSET, C. Kinetcs and mechanisms of antioxidant activity using the DPPH free radical method. Food Science Technology-Leb, v. 30, p. 609-615, 1997. BOX, G. E. P.; WILSON, K. B. On the experimental attainment of optimum conditions. Journal Royal Statistics Society, v. 13, p. 38, 1951. 59 BOX, G. E. P.; HUNTER, W. G.; HUNTER, J. S. Statistics for Experimenters. New York: John Wiley & Sons, 1978, 653 p. BRAND-WILLIAMS, W.; CUVELIER, M. E.; BERSET, C. Use of free radical method to evaluate antioxidant activity. Lebensmittel-Wissenschaft und-Technologie, v. 28, p. 2530, 1995. BRENNA, O. V.; PAGLIARINI, E. Multivariate analyses of antioxidant power and polyphenolic composition in red wines. Journal Agricultural Food Chemistry, v. 49, p. 4841-4844, 2001. BRITO, C. A. K. de et al. Características da atividade da peroxidase de abacaxis (Ananas comosus (L.) Merrill) da cultivar IAC Gomo-de-mel e do clone IAC-1. Ciência e Tecnologia de Alimentos, v. 25, p. 244-249, 2005. BRITO, C.A.K.; et al. Caracterização físico-química, enzimática e aceitação sensorial de três cultivares de abacaxi. Revista Brasileira de Tecnologia Agroindustrial, v. 02, p. 0114, 2008. BRITO, C. A. K. Estudo bioquímico das peroxidases brutas de abacaxi Ananas comosus (L.) Merril: cultivar IAC Gomo-de-Mel e clone IAC-1. 2001. 81 f. Dissertação (Mestrado em Ciência de Alimentos) - Universidade Estadual de Campinas, Campinas, São Paulo. BRUEMMER, J. E.; BONGWOO, R.; BOWEN, E. R. Peroxidase reactions and orange juice quality. Journal of Food Science, v. 41, p.186-189, 1976. BRUMMELL, D. A.; HARPSTER, M. H. Cell wall metabolism during in fruit softening and quality and its manipulation in transgenic plants. Plant Molecular Biology, v. 47, p.311-340, 2001. BRUMMELL, D. A. et al. Cell wall metabolism during the development of chilling injury in cold-stored peach fruit association of mealiness with arrested disassembly of cell wall pectins. Journal of Experimental Botany, v. 55, p. 2041-2052, 2004. 60 BUTTON, S. T. Metodologia para planejamento experimental e analise de resultados. Apostila de disciplina. UNICAMP, Faculdade de Engenharia Mecânica. Disponível em: <http://pessoal.utfpr.edu.br/lincolngusmao/arquivos/Planejamento Experimental.pdf>. Acesso em: 10 março 2011. BURNETTE, F. S. Peroxidase and its relationship to food flavour and quality: a review. Journal of Food Science, v. 42, p.1-6, 1977. CABRAL, J. R. S. et al. Variabilidade genética e melhoramento do abacaxi. In: RECURSOS GENÉTICOS E MELHORAMENTO DE PLANTAS PARA O NORDESTE BRASILEIRO, 1999, Petrolina. Anais... Petrolina: Embrapa Semi-Árido, Embrapa Recursos Genéticos e Biotecnologia/Brasília-DF, 1999. V.1, p. 9. Disponível em: <http://www.cpatsa.embrapa.br/catalogo>. Acesso em: 20 novembro 2010. CABRAL, J. R. S.; SOUZA, J. S.; FERREIRA, F. R. Viabilidade genética e melhoramento Genético do Abacaxi. In: QUEIROZ, M. A.; GOEDERT, C. O.; RAMOS, S. R. R. (eds.). Recursos genéticos e melhoramento de plantas para o Nordeste brasileiro. Petrolina, PE: Embrapa Semi-Arido; Brasilia, DF: Embrapa Recursos Genéticos e Biotecnologia, 2006. Disponivel em: <http://www.ceinfo.cnpat.embrapa.br/arquivos/artigo_2562.pdf>. Acesso em: 10 janeiro 2011. CABRAL, J. R. S. Cultivares de abacaxi. Cruz das Almas: Embrapa Mandioca e Fruticultura, 1999. 20 p. (Circular Técnica, 33). CABRAL, J. R. S.; MATOS, A. P. de. Imperial: nova cultivar de abacaxi. Cruz da Almas: Embrapa-CNPMF, 2005. (Embrapa-CNPMF. Comunicado Técnico, 114). CABRAL, J. R. S.; SOUZA, L. S. da S.; REINHARDT, D. H. (org.) Abacaxi: produção (aspectos técnicos). Brasília: Embrapa Comunicação para Transferência de Tecnologia, 2000. 77 p. (Frutas do Brasil, 7). CARDELLO, H. M. A. B.; CARDELLO, L. Teor de vitamina C, atividade de ascorbato oxidase e perfil sensorial de manga (Mangifera indica L.) var. Haden, durante o amadurecimento. Ciência e Tecnologia de Alimentos, v. 18, p. 211-217, 1998. 61 CARDOSO, C. L. et al. New Bioflavonoid and Other Flavonoids from the Leaves of Chimarrhis turbinate and their Antioxidant Activities. Journal of the Brazilian Chemical Society, v.16, p. 1353-1359, 2005. CASTALDO, D. et al. Presence of Residual Pectin Methylesterase Activity in Thermally Stabilized Industrial Fruit Preparations. Lebensmittel-Wissenschaft und-Technologie, v. 30, p. 470-484, 1997. CÉSAR, A. C. W. Análise de viabilidade econômica de um processo de extração e purificação da bromelina do abacaxi. 2005. 98f. Tese (Doutorado em Engenharia Química) – Faculdade de Engenharia Química, Universidade Estadual de Campinas, Campinas. CHITARRA, M. I. F.; CHITARRA, A. B. Pós-colheita de frutos e hortaliças: fisiologia e manuseio. Lavras: ESAL/FAEPE, 2005. 785 p. CHUTINTRASRIA, B.; NOOMHORMB, A. Thermal inactivation of polyphenoloxidase in pineapple puree. LWT - Food Science and Technology, v. 39, p. 492-495, 2006. CLEMENTE, E. Heat stability of soluble and ionically bound peroxidase from Orange. Revista Unimar, v.17, p. 401-408, 1995. CLEMENTE, E.; PASTORE, G. M. Peroxidase and polyphenoloxidase, the importance for food technology. Ciência e Tecnologia de Alimentos, v. 32, p. 167- 171, 1998. CRESTANI, M. et al. Das Américas para o Mundo - origem, domesticação e dispersão do abacaxizeiro. Ciência Rural, v.40, p. 1473-1483, 2010. CUNHA, G. A. P. Abacaxi on-line. Toda Fruta, edição de 17 de dez. de 2003. Disponível em: <http://www.todafruta.com.br/todafruta/mostra_conteudo.asp?conteudo=4859>. Acesso em: 17 outubro 2008. CUNHA, G. A. P. Equipe técnica de abacaxi comemora 30 anos de atividades e realizações. Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, 2007. 20p. 62 (Documentos, 170). Disponível em: <http://www.cnpmf.embrapa.br/ publicacoes/documentos170.pdf>. Acesso em: 10 fevereiro 2011. CUNHA, G. M. A. et al. Estudo da cinética de inativação térmica da peroxidase presente na polpa de goiaba. In: VI Congresso Brasileiro de Engenharia Química em Iniciação Científica, 2005. Disponivel em: < http://www.feq.unicamp.br/~cobeqic/tBT20.pdf>. Acesso em: 15 janeiro 2011. DEEPA, S. S.; ARUMUGHAN, C. Oil palm fruit peroxidase: purification and characterization, Journal of Food Science and Technology, v. 39, p. 8-13, 2002 DEBIEN, I. C. do N. Produção de água de coco (cocus nucifera) utilizando membranas de micro e ultrafiltração de celulose, pes e pvdf. 2010. Dissertação (Mestrado em Engenharia de Alimentos) - Faculdade de Engenharia de Alimentos, Universidade estadual de Campinas, Campinas. DEGÁSPARI, C. H.; WASZCZYNSKYJ, N. Propriedades antioxidantes de compostos fenólicos. Visão Acadêmica, v. 5, p. 33-40, 2004. DEGÁSPARI, C. H.; WASZCZYNSKYJ, N.; SANTOS, R. J. dos. Atividade antioxidante de extrato de fruto de Aroeira (Schinus terebenthifolius Raddi). Visão Acadêmica, v. 5, p. 83-90, 2004. DINIZ F. M.; MARTIN, A. M. Use of response surface methodology to describe the combined effects of pH, temperature and e/s ratio on the hydrolysis of dogfish (Squalus acanthias) muscle. International Journal of Food Science and Technology, v.31, p.19426, 1996. ESCRIBANO, J. et al. Characterization of monophenolase activity of table beet polyphenol oxidase. Determination of cinetic parameters on the tyramine/dopamine pair. Journal of Agricultural and Food Chemistry, v. 45, p. 4209-4214, 1997. EVANGELISTA, Jose. Tecnologia de alimentos. 2. ed. Sao Paulo: Atheneu, 1998. 652p. 63 FAOSTAT - Food and Agriculture Organization of the United Nations Statistical Database. Crops database. Disponível em: <http://faostat.fao.org/site/567/DesktopDefault.aspx?PageID=567#ancor>. Acesso em: 15 junho 2011. FANG, Z. et al. Polyphenol oxidase from bayberry (Myrica rubra Sieb. et Zucc.) and its role in anthocyanin degradation. Food Chemistry, v. 103, p. 268–273. 2007. FERREIRA, J. C. D. Purificação e Caracterização Parcial da Peroxidase do Abacaxi. 1983. 89 f. Dissertação (Mestre em Tecnologia de Alimentos). Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Campinas. FERREIRA, F. R.; CABRAL, J. R. S. Pineapple germplasm in Brazil. Acta Horticulturae, v. 334, p.23-26, 1993. FILHO, O. F.; VIEIRA, I. da C. Uso analítico de tecidos e de extratos brutos vegetais como fonte enzimática. Química Nova, v. 25, p. 455-464, 2002. FRANCO, G. Tabela de composição química dos alimentos. 8. ed. Rio de Janeiro: Livraria Atheneu, 1989. 230 p. FRANK, R. Exportadores brigam por versão superior e supostamente patenteada da fruta. Toda Fruta, edição de 07 de nov. De 2003. Disponível <http://www.todafruta.com.br/todafruta/mostra_conteudo.asp?conteudo=4555> em Acesso em: 12 outubro 2008. FREITAS, A. A. et al. Atividades das enzimas peroxidase (POD) e polifenoloxidase (PPO) nas uvas das cultivares benitaka e rubi e em seus sucos e geléias. Ciência e Tecnologia de Alimentos, v. 28, p. 172-177, 2008. FUJITA, S.; TONO, T. Purification and some properties of polyphenol oxidase in eggplant (Solanum melongena). Journal Science Food and Agriculture, v. 46, p. 115-123, 1988. 64 GAVA, A. J. Princípios da Tecnologia de Alimentos. São Paulo: Ed. Nobel, 1998. GEORGÉ, S. et al. Rapid determination of polyphenois and vitamin C in plant-derived products. Journal of Agricultural and Food Chemistry, v. 53, p.1370-1373, 2005. GIACOMELLI, E. J.; PY, C. Abacaxi no Brasil. Campinas: Fundação Cargill, 1981. 101p. GONÇALVES, N. B.; CARVALHO, V. D.; GONÇALVES, J. R. A. Efeito do cloreto de cálcio e do tratamento hidrotérmico na atividade enzimática e no teor de fenólicos do abacaxi. Pesquisa agropecuária brasileira, v.35, p.2075-2081, 2000. GRANADA, G. G.; ZAMBIAZI, R. Z.; MENDONÇA, C. R. B. Abacaxi: Produção, mercados e produtos. B.CEPPA, v. 22, p. 405-222, 2004. HAMMERSCHMIDT, P. A.; PRATT, D. E. Phenolic antioxidants of dried soybeans. Journal of Food Science, v. 43, p. 556-559, 1978. HAARD, N. F. Physiology cal roles of peroxidase in postharvest fruits and vegetables. In: ORY, R. L.; S.t ANGELO, A. T. (eds.). Enzymes in Food an Beverages Process. Symposium series, p. 143-171, 1977. IBRAF. Instituto Brasileiro de Frutas. Disponível em: <http://www.ibraf.org.br/estatisticas/est_processadas.asp> . Acesso em: 8 outubro 2010. IBGE. Instituto Brasileiro de Geografia e Estatística. Sistema IBGE de Recuperação Automática. SIDRA/LSPA: banco de dados do IBGE. Disponível em: <http://www.sidra.ibge.gov.br >. Acesso em: 07 abril 2010. INCAPER. Assistência Informativo Técnica especial e do Instituto Extensão Capixaba Rural. de Disponível Pesquisa, em: <http://incaper.web407.uni5.net/revista.php?idcap=958>. Acesso em: 04 fevereiro 2011. 65 JAYAPRAKASHA, G. K. et al. Antioxidant and antimutagenic activities of Cinnamomum zeylanicum fruit extracts. Journal of Food Composition and Analysis, v. 20, p. 330-336, 2007. JUNIOR, A. F. Enzimas e suas aplicações: Cinética enzimática. Florianópolis, Universidade Estadual de Santa Catarina, 2001. Disponível em: <http://www.enq.ufsc.br/labs/probio/disc_eng_bioq/lista_exerc/cinetica_enzimatica.pdf>. Acesso em: 13 março 2010. KADER, A . A . Biochemichal and physiological basis for effects of controlled and modified atmospheres on fruits and vegetables. Food Technology, p.99-104, 1986. KUMAR, V. B. A; KISHOR MOHAN, T. C.; MURUGAN, K. Purification and kinetic characterization of polyphenol oxidase from Barbados cherry (Malpighia glabra L.). Food Chemistry, v. 110, p. 328–333, 2008. KAMIMURA, G. K. F. Isolamento, purificação e caracterização da peroxidase de yacon (Smallanthus sonchifolius). 2006. Dissertação (Mestrado em Alimentos e Nutrição, área de Ciências dos Alimentos) - Faculdade de Ciências Farmacêuticas, UNESP. Araraquara. KHAN, A. A.; ROBINSON, D. S. The thermo stability of purified mango isoperoxidases. Food Chemistry, v. 47, p. 53-59, 1993. KE, D.; SALTVEIT, M. E. Wound-induced ethylene production, phenolic metabolism and susceptibility to russet spotting in iceberg lettuce. Physiologia Plantarum, v. 76, p. 412418, 1989. KIM, Y.; GIRAUD, D. W.; DRISKELL, J. A. Journal of Food Composition Analysis, v. 20, p. 458, 2007. KOBLITZ, M. G. B. Bioquímica de alimentos. Teoria e aplicações práticas. Rio de Janeiro: Guanabara Koogan, 2008. 242 p. 66 KOLEVA, I. I. et al. Screening of Plant Extracts for Antioxidant Activity: a Comparative Study of Three Testing Methods. Phytochemical Analysis, v. 13 p. 8-17, 2002. KORNER, B.; ZIMMERMANN, G.; BERK, Z. Orange pectinesterase: purification, properties, and effect on cloud stability. Journal of Food Science., v. 45, p. 1203-1206, 1980. KRATCHANOVA, M.; PAVLOVA E.; PANCHEV, I. The effect of microwave heating o fresh orange peels on the fruit tissue and quality of extracted pectin. Carbohydrate Polymers, v. 56, p. 181-185, 2004. KUSKOSKI, E. M. et al. Frutos tropicais silvestres e polpas de frutas congeladas: atividade antioxidante, polifenóis e antocianinas. Ciência Rural, v.36, p.1283-1287, 2006. LAURILA, E.; KERVINEN, R. Y.; AHVENAINER, R. The inhibition of enzymatic browning in minimally processed vegetables and fruits. Postharvest News and Information, v. 9, p. 53-66, 1998. LEA, A. G. H. Enzymes in the production of beverages and fruit juices. In: TUCKER, G. A.; WOODS, L. F. J. Enzymes in Food Processing, p. 223-249. Blackie Academic & Professional (Chapman & Hall): Londres, 1995. LEE, C. Y.; PENNESI, A. P.; DICKSON, M. H. Characterization of cauliflower peroxidase isoenzyme. Journal of Agricultural And Food Chemistry, v. 32, p. 18-21, 1984. LEE, Y. C.; HAMMES, J. K. Heat inativation of Peroxidase in corn on the cob. Journal of Food Science, v. 44, p. 785-787, 1979. LIU, X. et al. Inactivation of peroxidase and polyphenol oxidase in red beet (Beta vulgaris L.) extract with high pressure carbon dioxide. Innovative Food Science and Emerging Technologies, v. 9, p. 24–31, 2008. LOURENÇO, E. J.; NEVES, V. A. Peroxidase solúvel de pêssego: purificação parcial e propriedades. Ciência e Tecnologia de Alimentos, v.17, p. 42-48, 1997. 67 MACHADO, C. F. et al. Estado da arte do Banco Ativo de Germoplasma de Abacaxi da Embrapa Mandioca e Fruticultura Tropical. In: CONGRESSO BRASILEIRO DE MELHORAMENTO DE PLANTAS, 2009, Guarapari. O melhoramento e os novos cenários da agricultura. Guarapari: SBMP, 2009. MADINEZ, M. V.; WHITAKER, J. R. The biochemistry and control of enzymatic browning. Trends in Food Science & Technology, v. 6, p. 195-200. 1995. MAIA, G. A.; SOUSA, P. H. M.; LIMA, A. S. Processamento de sucos de frutas tropicais. Fortaleza: Edições UFC, 2007. 320p. MAISUTHISAKUL, P.; SUTTAJIT, M.; PONGSAWATMANIT, R. Assessment of phenolic content and free radical-scavenging capacity of some Thai indigenous plants. Food Chemistry, v. 100, p. 1409–1418 2007. MARCO, G. J. A rapid method for evaluation of antioxidants. Journal of the American Oil Chemists‟ Society, v. 45, p. 594-598, 1968. MARTINEZ, M. V.; WHITAKER, J. R. The biochemistry and control of enzymatic browning. Trends in Food Science and Technology, v. 6, p. 195-200, 1995. MEDINA, J. C.; Cultura, In: Abacaxi. 2 ed. Campinas: São Paulo: Ital, 1987. p. 1-133. MELLO, E. T.; CLEMENTE, E. Thermo stability of crude extract of peroxidase from pineapple. Revista Unimar, v. 18, p. 757-763, 1996. MELO, E. A. et al. Capacidade antioxidante de frutas. Revista Brasileira de Ciências Farmacêuticas, v. 44, p. 193-201, 2008. MÉLO, E. A.; LIMA, V. L. A. G; MACIEL, M.I.S. Polyphenol, ascorbic acid and total carotenoid contents in common fruits and vegetables. Brazilian Journal of Food Technology, v. 9, p. 89-94. 2006. 68 MILIAUSKAS, G.; VENSKUTONIS, P. R.; VAN BEEK, T. A. Screening of radical scavenging activity of some medicinal and aromatic plant extracts. Food Chemistry, v. 85, p. 231-237, 2004. MISHRA, B. et al Pulse radiolysis studies of mangiferin: A C-glycosyl xanthone isolated from Mangifera indica. Radiation Physics and Chemistry, v. 75, p. 70-77, 2005. NACZAK, M.; SHAHIDI, F. Extraction and analysis of phenolics in food. Journal of Chromatography A, v. 1054, p. 95-111, 2004. NASCENTE, A. S. et al. Cultivo do abacaxi em Rondônia. Porto Velho, 2005. Disponível em: <http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Abacaxi/CultivodoAbacaxiR O/autores.htm>. Acesso em: 07 fevereiro 2011. NIELSEN, J. E.; CHRISTENSEN, T. M. I. E. Distribution of pectin methyl esterase and acetyl esterase in the genus Citrus visualized by tissue prints and chromatography. Plant Science, v. 162, p. 799-807, 2002 NÚÑEZ-SELLÉS A. J. et al. Isolation and quantitative analysis of phenolic antioxidants, free sugars, and polyols from mango (Mangifera indica L.) stem bark aqueous decoction used in Cuba as nutritional supplement. Journal of Agricultural and Food Chemistry, v. 50, p. 762-766, 2002. OMONI, A.; ALUKO, R. The anticarcinogenic and anti-atherogenic effects of lycopene: a review. Trends in Food Science & Technology, v. 16, p. 344-350, 2005. OMS-OLIU, G. et al. The role of peroxidase on the antioxidant potential of fresh-cut „Piel de Sapo‟ melon packaged under different modified atmospheres. Food Chemistry, v. 106, p. 1085–1092, 2008. PINHEIRO, A. C. M. et al. Influência do CaCl2 sobre a qualidade pós-colheita de abacaxi pérola (Ananas comosus (L.) Merril). In: CONGRESSO BRASILEIRO DE CIÊNCIA E TECNOLOGIA DE ALIMENTOS, 17, 2000, Fortaleza. Anais... Resumos. Fortaleza: SBCTA, 2000. 162p. 69 PIZA, I. M. T.; LIMA, G. P. P.; BRASIL, O. G. Atividade de peroxidase e níveis de proteínas em plantas de abacaxizeiro micropropagadas em meio salino. Revista brasileira de Agrociência, v. 9, p. 361-366, 2003. POMAR, F. et al. Purification, characterization and kinetic properties of pepper fruit acidic peroxidase. Phytochemistry, v. 46, p. 1313-1317, 1997. POOVAIAH, B. W. Role of calcium in prolonging storage life of fruits and vegetables. Food Technology, v. 40, p. 86-89, 1986. PRADO, M. E. T. et al. Armazenamento de abacaxi „Smooth Cayenne‟ minimamente processado sob refrigeração e atmosfera modificada. Revista Brasileira de Fruticultura, v. 25, p. 67-70, 2003. RAMOS, A. W. Delineamento de experimentos. Pro 2712 – controle da qualidade. Escola Politécnica da Universidade de São Paulo, 2005. Disponível em: <http://www.prd.usp.br/>. Acesso em: 12 novembro 2010. RIBEIRO, A. B.; SILVA, D. H. S.; BOLZANI, V. S. Flavonóis glicosilados antioxidantes de Nectandra grandiflora (Lauraceae). Eclética Química, v. 27, p. 35-44, 2002. RENSBURG, E. V; ENGELBRECHT, A. H. P. Effec of calcium salts on susceptibility to browning of avocado fruit. Journal of Food Science, v. 51, p. 1067-1068, 1986. RESENDE, J. M. et al. Atividade de enzimas pectinametilesterase e poligalacturonase durante o amadurecimento de tomates do grupo multilocular. Horticultura Brasileira, v. 22, p. 206-212, 2004. RICHARDSON, T.; HYSLOP, D. B. Enzymes. In: FENNEMA, O. R. 2. ed. New York: Marcel Dekker, 1985, p. 451-452. RODRIGUES, M. I.; IEMMA, A. F. Planejamento e otimização de processos. Uma estratégia sequencial de planejamentos. Ed. Casa do pão. 2005. 326 p. 70 ROESLER, R. Estudo de frutas do cerrado brasileiro para avaliação de propriedade funcional com foco na atividade antioxidante. 2007. 218f. Tese (Doutorado em Ciência de Alimentos) – UNICAMP, Campinas, São Paulo. ROMBOUTS, F. M. et al. Pectinesterases in component parts of citrus fruits related to problems of cloud loss and gelation in citrus products. In Use of Enzymes in food technology. Proceedings Of Iternational Symposium. France: Ed. Versailles, p.483-487, 1982. RUFINO, M. S. M. et al. Determinação da atividade antioxidante total em frutas pela captura do radical livre DPPH. Fortaleza: Embrapa Agroindústria Tropical, 2007. (Comunicado Técnico, 127). SAPERS, G. M.; MILLER, R. L. Browning inhibition in fresh-cut pears. Journal of Food Science, v. 63, p. 342-346, 1998. SANTOS, E. R. Caracterização bioquímica de peroxidase e da polifenoloxidase de açaí (Euterpe oleracea). 2002, 104f. Dissertação (Mestrado em Ciência de Alimentos), Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, Campinas. SÁNCHEZ-MORENO, C.; LARRAURI, J. A.; SAURA-CALIXTO, F. A procedure to measure the antiradical efficiency of polyphenols. Journal Science Food Agriculture, v. 76, p. 270-276, 1998. SÁNCHEZ-MORENO, C. Compuestos polifenólicos: efectos fisiológicos. Actividad antioxidante. Alimentaria, v. 329, p.29-40, 2002. SANTANA, L. R. R. de. Qualidade Pós-colheita de pêssego “Douradão” sob atmosfera controlada e modificada. Campinas. 2009. 176f. Tese (Doutorado em Engenharia Agrícola) - Universidade Estadual de Campinas, Campinas. SCALON, S. de P. Q.; DELL’OLLIO, P.; FORNASIERI, J. L. Temperatura e embalagens na conservação pós-colheita de Eugenia uvalha Cambess – Myrtaceae. Ciência Rural, v. 34, p. 1965-1968, 2004. 71 SIDDIQUI, S.; BANGERTH, F. The effect of calcium infiltration on structural changes in cell walls of stored apples. Journal of Horticultural Science, v. 5, p.703-708, 1996. SILVA, J. M. de; SILVA, J. P.; SPOTO, M. H. F. Análises fisiológicas e enzimáticas em abacaxi submetido à tecnologia de radiação ionizante. Ciência e Tecnologia de Alimentos, v. 27, p. 602-607, 2007. SILVA, J. P. da. Qualidade de infrutescência de abacaxizeiro „MD-2‟ cultivado sob diferentes relações K/N. 2010. 100f. Dissertação (Mestrado em Agronomia) – UFPB, Paraíba. SILVA, M. V. da; ROSA, C. I. L. F.; BOAS, E. V. de B. V. Conceitos e Métodos de controle do escurecimento enzimático no processamento mínimo de frutas e hortaliças. B.CEPPA, v. 27, p. 83-96, 2009. SILVA, S.; TASSARA, H. Abacaxi. In: SILVA, S.; TASSARA, H. Frutas no Brasil. São Paulo: Nobel, 2001. p.25-27. SILVA, C. C. M.; SILVA, M. V. L. e SANTOS, J. R. Estudo das características físicoquímicas de ésteres etílicos obtidos a partir do óleo de mamona, Ricinus communis L. In: 28ª Reunião Anual da Sociedade Brasileira de Química. Poços de Calda-MG, 2005. SINGLETON, V. L.; ROSSI, J. A. Colorimetry of total phenolics with phosphomolybdicphosphotungstic acid reagents. American Journal of Enology and Viticulture, v.16, p.144-158, 1965. SHALINI, G. R.; SHIVARE, U. S.; BASU, S. Thermal inactivation kinetics of peroxidase in mint leaves. Journal of Food Engineering, v. 85, p. 147–153, 2008. TEISSON, C. Le brunissement interne de l‟ananas. IHistorique. II-Material et méthodes. Fruits, v.34, p. 245-281,1979. 72 TEIXEIRA, G. H. A. et al. Use of modified atmosphere to extend shelf life of fresh-cut carambola (Averrhoa carambola L. cv. Fwang Tung). Postharvest Biology and Technology, v. 44, p. 80–85. 2007. TIWARI, B.K. et al. Inactivation kinetics of pectin methylesterase and cloud retention in sonicated orange juice. Innovative Food Science and Emerging Technologies, v. 10, p. 166–171. 2009. THÉ, P. M. et al. Influência de tratamentos pós-colheita sobre a atividade enzimática de abacaxi cv. Smooth Cayenne. B.CEPPA, v. 24, p. 423-442, 2006. THÉ, P. M. et al. Modificações na atividade enzimática em abacaxi ‘Smooth Cayenne’ em função da temperatura de armazenamento e do estádio de maturação. Ciência e Agrotecnologia, v.25, p.364-370, 2001. THÉ, P. M. P.; NUNES, R. de P.; CARVALHO, V. D. de. Efeitos de tratamentos póscolheita sobre fatores que influenciam na textura de abacaxis cv. Smooth Cayenne. Revista Ceres, v. 56, p. 705-712, 2009. TOMÁS-BARBERÁN, F. A.; ESPÍN, J. C. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. Journal of the Science of Food and Agriculture, v. 81, p. 853-879, 2001. TOIVONEN, P. M. M.; BRUMMELL, D. A. Review. Biochemical bases of appearance and texture changes in fresh-cut fruit and vegetables. Postharvest Biology and Technology, v. 48, p. 1-14, 2007. UENOJO, M.; PASTORE, G. M. Isoloamento e seleção de microrganismos pectinilíticos a partir de resíduos provenientes de agroindústrias para produção de aromas frutais. Revista Ciência e Tecnologia de Alimentos, v. 26, p. 509-513, 2006. URLAUB, R. Enzymes in fruit and vegetable juice extraction. In: WHITEHUSRT, R. J.; LAW, B. A. Enzyms in Food Technology, p.144-183. Sheffield Academic Press (CRC Press), Sheffield. 2002. 73 VÁMOS-VIGYÁZÓ, L. Polyphenol oxidase and peroxidase in fruits and vegetables. CRC – Critical Reviews in Food Science and Nutrition, v.15, p. 49-127, 1981. VASCO, C.; RUALES, J.; KAMAL-ELDIN, A. Total phenolic compounds and antioxidant capacities of major fruits from Ecuador. Food Chemistry, v. 111, p. 816–823, 2008. VANDRESEN, S. Caracterização físico-química e Comportamento reológico de sucos de Cenoura e laranja e suas misturas. 2007. 133f. Dissertação (Mestrado em Engenharia de Alimentos) - Universidade Federal de Santa Catarina, Florianópolis. VEDANA, M. I .S. Efeito do processamento na atividade antioxidante da uva. 2008. 85f. Dissertação (Mestrado em Ciência e Tecnologia de alimentos) - Programa de PósGraduação em Ciência e Tecnologia de Alimentos, Universidade Federal do Paraná, Paraná. VENTURA, J. A. et al. „Vitoria‟: Nova cultivar de abacaxi resistente a fusariose. DCM-INCARPER, Documento 148, Espírito Santo, 2006. <http://www.incaper.es.gov.br/servicos/images/abacaxi_vitoria.pdf>. Disponível Acesso em: em: 20 março 2011. VIDAL, J. R. M. B.; GASPARETO, C. A. Comportamento reológico da polpa de manga (Mangífera indica L. ) – efeito dos sólidos insolúveis. In: Boletim da Sociedade Brasileira de Ciência e Tecnologia de Alimentos, Campinas, v.20, p.172-175, 2000. YILDIRIM, A.; MAVI, A.; KARA, A. A. Determination of antioxidant and antimicrobial activities of Rumex crispus L. extracts. Journal of Agricultural and Food Chemistry, v.49, p. 4083-4089, 2001. WHITAKER, J. R. Principles of enzymology for the food science. New York: Marcel Dekker, 1972, 636 p. 74 WHITAKER, J. R.; LEE, C. Y. Recent advances in chemistry of enzymatic browning. In: WHITAKER, J. R. (Ed.). Enzymatic browning and its prevention. Washington: American Chemical Society, 1995. p. 2-7. WU, X. et al. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. Journal of Agricultural and Food Chemistry, v.52, p.4026-4037, 2005. ZADERNOWSKI, R., M. et al. Composition of phenolic acids in sea buckthorn (Hippophae rhamnoides L.) berries. Journal of the American Oil Chemists‟ Society, v. 82, p. 175-179, 2005 ZAWISTOWSKI, J.; BILIADERIS, C. G.; ESKIN, N. A. M. Poliphenol oxidase. In: ROBINSON, D. S.; ESKIN, N. A. M. (Ed.) Oxidative enzymes in foods. London: Elsevier Science, 1991. p. 218-220. ZHOU, H. et al. Delayed storage and controlled atmosphere storage of nectarines: two strategies to prevent woolliness. Postharvest Biology and Technology, v. 18, p. 133-141, 2000. ZHOU, Y. et al. Enzymes associated with blackheart development in pineapple fruit. Food Chemistry, v. 80, p. 565–572, 2003. ZOCCA, F. et al. Detection of pectinmethylesterase activity in presence of methanol during grape pomace storage. Food Chemistry, v. 102, p. 59–65, 2007. 75 7.0 Anexos Anexo 01. Gráficos obtidos no experimento de otimização da extração enzimática. Figura 01(a). Gráfico de Pareto para extração de PER. Figura 01(b). Gráfico Bidimensional de contorno para extração de PER. 76 Figura 01(c). Gráfico bidimensional de contorno para extração de PFO. Figura 01(d). Gráfico bidimensional de contorno para extração de PME. Figura 01(e). Gráfico de Pareto para extração de PME. 77 Figura 01(f). Gráfico bidimensional de contorno para extração de PG. Pareto Chart of Standardized Effects; Variable: PG 2 factors, 1 Blocks, 11 Runs; MS Pure Error=13,50904 DV: PG (2)NaCl(L) 9,27627 (1)pH(L) 3,756507 NaCl(Q) pH(Q) 1Lby2L -2,29795 -1,63694 ,6088086 p=,05 Standardized Effect Estimate (Absolute Value) Figura 01(g). Gráfico de Pareto para extração de PG. 78 Anexos 02. Gráficos obtidos no experimento de inativação da enzima PER. Pareto Chart of Standardized Effects; Variable: % INATIVAÇÃO 3 factors, 1 Blocks, 18 Runs; MS Pure Error=1,003367 DV: % INATIVAÇÃO (2)TEMPERATURA(L) 89,40051 TEMPERATURA(Q) 51,71374 (1)CONCENTRAÇÃO(L) 26,46374 TEMPO(Q) 13,68367 (3)TEMPO(L) 12,63366 2Lby3L 8,248669 CONCENTRAÇÃO(Q) 6,070128 1Lby2L -2,80603 1Lby3L 2,678964 p=,05 Standardized Effect Estimate (Absolute Value) Figura 02(a). Gráfico de Pareto para a inativação de PER com ácido ascórbico. Pareto Chart of Standardized Effects; Variable: % inativação 3 factors, 1 Blocks, 18 Runs; MS Pure Error=,0972667 DV: % inativação (2)temperatura(L) 266,0781 temperatura(Q) 197,7465 (3)tempo(L) 56,24921 tempo(Q) 47,39895 1Lby3L -46,581 Concentração(Q) -44,4868 (1)Concentração(L) 35,68435 2Lby3L 7,13056 1Lby2L -5,24873 p=,05 Standardized Effect Estimate (Absolute Value) Figura 02(b). Gráfico de Pareto para a inativação de PER com mestabissulfito de sódio. Pareto Chart of Standardized Effects; Variable: % inativação 3 factors, 1 Blocks, 18 Runs; MS Pure Error=1,261391 DV: % inativação (2)Temperatura(L) 59,13516 Temperatura(Q) 40,83479 Tempo(Q) 7,615482 (3)Tempo(L) 6,051306 Concentração(Q) -4,17148 (1)Concentração(L) -4,05439 1Lby2L 3,61288 1Lby3L -2,62331 2Lby3L -1,67107 p=,05 Standardized Effect Estimate (Absolute Value) Figura 02(c). Gráfico de Pareto para a inativação de PER com cloreto de cálcio. 79 % inativação = 23,298352419362 + 7,1730811931683*x + 1,7096088260765*x2 + 24,232291056662*y + 14,564810108048*y2 + 2,92125*0*y 120 100 80 60 40 20 i % inativação = 23,298352419362+24,232291056662*x +14,564810108048*x2 +3,424393188822*y+3,8539101400247*y2+2,92125*x*y+0 % inativação = 23,298352419362 + 7,1730811931683*x + 1,7096088260765*x2 + 24,232291056662*y + 14,564810108048*y 2 + 2,92125*0*y 1,5 1,5 1,0 0,5 0,5 TEMPO min TEMPERATURA °C 1,0 0,0 -0,5 -0,5 -1,0 -1,0 -1,5 -1,5 ii 0,0 -1,0 -0,5 0,0 0,5 1,0 CONCENTRAÇÃO Ac. Ascorb. mg/L 1,5 120 100 80 60 40 20 -1,5 -1,5 iii -1,0 -0,5 0,0 0,5 1,0 1,5 120 100 80 60 40 20 TEMPERATURA °C Figura 02(d). Superfície de resposta para o efeito temperatura e concentração de acido ascórbico (i), gráfico bidimensional de contorno para os efeitos temperatura e concentração de acido ascórbico (ii) e temperatura e tempo (iii) na inativação enzimática. 80 % inativação=30,779266245175+22,455139510485*x+17,340429544132*x² +4,7470423406699*y+4,1564236089085*y²-0;57875*1*x-5,13625*1*y +0,78625*x*y-0,88954677 120 100 80 60 40 i Fitted Surface; Variable: % inativação DV: % inativação 1,5 1,50 1,25 1,0 1,00 0,75 0,5 temperatura: 2 tempo: 3 0,50 0,0 0,25 0,00 -0,25 -0,5 -0,50 -0,75 -1,0 -1,00 -1,25 -1,5 ii i -1,6 -1,2 -0,8 -0,4 0,0 0,4 0,8 1,2 1,6 -1,4 -1,0 -0,6 -0,2 0,2 0,6 1,0 1,4 temperatura: 2 120 100 80 60 40 -1,50 ii -1,5 -1,0 -0,5 0,0 0,5 1,0 1,5 120 100 80 60 40 20 Concentração: 1 i ii Figura 02(e). Superfície de resposta para o efeito tempo e temperatura (i); gráfico ii bidimensional de contorno para os efeitos tempo e temperatura (ii) e temperatura e concentração de metabissulfito de sódio (iii) na inativação enzimática. 81 Fitted Surface; Variable: % inativação DV: % inativação 1,5 1,0 Tempo: 3 0,5 0,0 -0,5 -1,0 -1,5 -1,5 i -1,0 -0,5 0,0 0,5 1,0 1,5 100 80 60 40 20 Temperatura: 2 z=26;195891851905-1;2321829136412*x-1;3172979030932*x^2 +17;971953293654*y+12;895087059813*y^2+1;4346085409253*x*y +4;24394015 ii 80 60 40 20 Figura 02(f). Gráfico bidimensional de contorno quando se aplicou cloreto de cálcio em termos de porcentagem de inativação enzimática para o efeito tempo e temperatura (i) e superfície de resposta para os efeitos temperatura e concentração de cloreto de cálcio (ii). 82 Anexos 03. Gráficos obtidos no experimento de inativação da enzima PFO. Pareto Chart of Standardized Effects; Variable: % inativação 3 factors, 1 Blocks, 18 Runs; MS Pure Error=,6163667 DV: % inativação (2)Temperatura(L) Temperatura(Q) (1)Conc. Ac. Ascorbico(L) (3)Tempo(L) Tempo(Q) 1Lby3L 1Lby2L Conc. Ac. Ascorbico(Q) 2Lby3L 73,62166 30,57979 29,89797 15,3099 12,84958 -5,90389 -4,42679 1,037441 ,7430523 p=,05 Standardized Effect Estimate (Absolute Value) Figura 03(a). Gráfico de Pareto para a inativação de PER com ácido ascórbico. Pareto Chart of Standardized Effects; Variable: % de inativação 3 factors, 1 Blocks, 18 Runs; MS Pure Error=,4479083 DV: % de inativação (2)Temperatura(L) (1)conc. MS(L) Temperatura(Q) (3)Tempo(L) 1Lby2L conc. MS(Q) 2Lby3L Tempo(Q) 1Lby3L 75,48655 45,01172 37,50694 30,20375 -24,2487 9,016229 -7,79423 5,166133 -4,33013 p=,05 Standardized Effect Estimate (Absolute Value) Figura 03(b). Gráfico de Pareto para a inativação de PER com metabissulfito de sódio. Pareto Chart of Standardized Effects; Variable: % de inativaçao 3 factors, 1 Blocks, 18 Runs; MS Pure Error=1,119771 DV: % de inativaçao (2)Temperatura(L) 56,76438 Concentraçao CC(Q) -15,2225 (1)Concentraçao CC(L) 13,89096 (3)Tempo(L) 12,2618 Tempo(Q) -9,86547 Temperatura(Q) 6,205623 1Lby2L -3,28634 1Lby3L 2Lby3L -2,19089 0, p=,05 Standardized Effect Estimate (Absolute Value) Figura 03(c). Gráfico de Pareto para a inativação de PER com cloreto de cálcio. 83 i 80 70 60 50 z=61;465828615814+6;3516403089265*x+3;2524943622243*y +2;7885044115204*y^2-1;22875*0;*x-1;63875*x*y+0; z=61;465828615814+15;640471635941*x+6;702340445388*x^2 +3;2524943622243*y+2;7885044115204*y^2-1;22875*0;*x-1;63875*0;*y +0; 1,5 1,0 Tempo: 3 0,5 0,0 -0,5 110 100 90 80 70 60 -1,0 -1,5 -1,50 -1,25 -1,00 -0,75 -0,50 -0,25 ii 0,00 0,25 0,50 0,75 1,00 1,25 1,50 Temperatura: 2 z=61;465828615814+6;3516403089265*x+3;2524943622243*y +2;7885044115204*y^2-1;22875*0;*x-1;63875*x*y+0; 1,5 Tempo: 3 1,0 0,5 0,0 -0,5 -1,0 -1,5 -1,5 iii -1,0 -0,5 0,0 0,5 1,0 1,5 80 70 60 50 Conc. Ac. Ascorbico: 1 Figura 03(d). Superfície de resposta para o efeito tempo e concentração de ácido ascórbico (i), gráfico bidimensional de contorno para os efeitos tempo e temperatura (ii) e tempo e concentração de ácido ascórbico (iii) na inativação enzimática. 84 z=61;290546100252+13;670637000461*x+7;0578958903423*x^2 +5;4699090306075*y+;97214080635975*y^2-5;7377049180328*0;*x -1;0245901639344*0;*y-1;844262295082*x*y+0; 110 100 90 80 70 60 50 Figura 03(e). Superfície de resposta para o efeito, tempo e temperatura quando se aplicou metabissulfito de sódio na inativação de PFO. Fitted Surface; Variable: % de inativação 3 factors, 1 Blocks, 18 Runs; MS Pure Error=,4479083 DV: % de inativação 1,5 1,0 Tempo: 3 0,5 0,0 -0,5 -1,0 -1,5 -1,5 -1,0 -0,5 0,0 0,5 1,0 1,5 90 80 conc. MS: 1 Figura 03(f). Gráfico bidimensional de contorno para os efeitos tempo e concentração de metabissulfito de sódio na inativação enzimática de PFO. 85 z=71;794411396116+3;977602204174*x-4;5291819016944*x^2 +16;254181972065*y+1;8463710434302*y^2-1;2295081967213*x*y+0; 1,5 Temperatura: 2 1,0 0,5 0,0 -0,5 -1,0 -1,5 -1,5 i -1,0 -0,5 0,0 0,5 1,0 1,5 100 90 80 70 60 50 40 30 Concentraçao CC: 1 Fitted Surface; Variable: % de inativaçao 3 factors, 1 Blocks, 18 Runs; MS Pure Error=1,119771 DV: % de inativaçao 1,5 Tempo: 3 1,0 0,5 0,0 -0,5 -1,0 -1,5 -1,5 ii -1,0 -0,5 0,0 0,5 1,0 1,5 90 80 70 60 Concentraçao CC: 1 Fitted Surface; Variable: % de inativaçao 3 factors, 1 Blocks, 18 Runs; MS Pure Error=1,119771 DV: % de inativaçao iii z=71;794411396116+16;254181972065*x+1;8463710434302*x^2 +3;5111007424365*y-2;9352936654133*y^2-1;2295081967213*0;*x+0; 100 90 80 70 60 50 40 Figura 03(g). Gráfico bidimensional de contorno para o efeito temperatura e concentração de cloreto de cálcio (i), tempo e concentração de cloreto de cálcio (ii), e superfície de resposta para os efeitos tempo e temperatura (iii).
Download