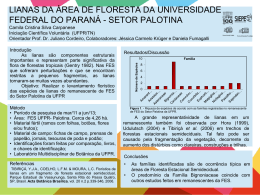
O sentido de enrolamento não influencia o desempenho ecológico de lianas Tiago Jordão Porto; Claudia Pandolfo Paz; Marcel Vaz & Paula Munhoz de Omena Gibson 1996). O fato de não possuírem Introdução A luz é um recurso limitante muitas fibras de sustentação permite para as plantas de subbosque em que algumas espécies de lianas se florestas tropicais (Townsend et al. enrolem no caule de suas hospedeiras 2003), pois somente 4% da radiação (Ingrouille & Eddie 2006), tanto no solar chega ao solo (Chazdon & Fetcher sentido horário como no anti-horário. 1984). A limitação de luz promove uma Apesar de existirem sugestões de que o intensa competição entre as plantas e sentido anti-horário seja mais comum cria uma pressão seletiva que favorece a nas florestas neotropicais (Rundel & evolução que Gibson 1996), estudos empíricos sobre incrementam a obtenção deste recurso. este assunto não foram realizados. Caso Algumas dessas estratégias são (1) o o sentido anti-horário seja realmente o hábito arbóreo, em que plantas crescem mais frequente, é razoável supor que a de modo a sobrepor sua copa em predominância para um sentido confira relação às vizinhas; (2) o epifitismo, em vantagem adaptativa aos indivíduos. A que plantas vantagem poderia ser expressa por meio hospedeiras arbóreas como suporte e de um maior crescimento de indivíduos assim, têm acesso aos estratos próximos com ao dossel e (3) lianas, em que as plantas enrolamento. O crescimento pode ser também utilizam outras plantas como inferido pelo diâmetro do caule dos suporte, mas mantêm o contato com o indivíduos, pois, geralmente, plantas solo (Ribeiro et al. 1999). que vivem mais crescem mais em de herbáceas estratégias utilizam sentido anti-horário de Lianas levam vantagem sobre as diâmetro (Finegan outras plantas na competição por luz, Portanto, quando pois alocam menos recursos em tecidos grupos de lianas com enrolamento para de a direita e para a esquerda é esperado sustentação podendo crescer et al. 1999). comparamos os rapidamente em altura (Rundel & 1 que o grupo com maior crescimento hospedeira. Categorizamos o sentido do tenha, em média, maior diâmetro. enrolamento em horário e anti-horário e O objetivo deste estudo foi obter medimos o diâmetro das lianas, em dados quantitativos sobre o sentido de alturas que variaram de 1,3 m até 2 m enrolamento das lianas em uma floresta em relação ao solo, com paquímetro de de terra firme na Amazônia Central. No precisão de 1 mm. Para testar se há uma caso de haver predominância em algum maior sentido, testaremos se o enrolamento no enrolamento no sentido anti-horário sentido uma utilizamos o teste qui-quadrado (χ2). medida Para testar se existe diferença no indiretamente através do crescimento diâmetro de lianas com sentido horário dos e anti-horário utilizamos o teste de predominante vantagem confere adaptativa indivíduos em termos de frequência crescimento dos indivíduos. Esperamos, Mann-Wittney. caso haja a predominância no sentido Resultados anti-horário, que os de lianas com indivíduos O número de lianas enroladas no enrolados nesse sentido tenham um sentido anti-horário (n = 215) foi 5,5 diâmetro médio maior do que os vezes maior do que o número de lianas indivíduos enrolados no sentido horário. enroladas no sentido horário (n = 39) (χ2 = 121,1; g.l. = 1; p < 0,0001). O diâmetro médio das lianas que enrolam- Métodos no se no sentido horário foi de 10,2 mm fragmento florestal de 100 ha da (dp = 10,4) e das lianas no sentido anti- Fazenda Dimona (02º20’ S - 60º06’ W) horário foi de 13,6 mm (dp = 17,8). ao norte de Manaus, Amazonas, área de Encontramos uma maior frequência de estudo do Projeto Dinâmica Biológica lianas nas menores classes de diâmetro, de Fragmentos Florestais (PDBFF). ou seja, Nossas unidades amostrais foram lianas 79 % tinham menos de 20 mm (Figura avistadas ao longo das trilhas do interior 1). Não encontramos uma relação entre do fragmento. Amostramos apenas as o diâmetro das lianas e o sentido do lianas que tivessem ao menos três voltas enrolamento (U= 3964; p = 0,62; Figura completas 1). Realizamos em o torno estudo da planta 2 Diâmetro (mm) 140 130 120 110 100 90 80 70 60 50 40 30 20 10 0 (0;5] (5;10] (10;15] (15;20] (20;25] (25;30] > 30 Horário Anti-horário Sentido de crescimento Figura 1. Relação entre o diâmetro do caule e o sentido de enrolamento de lianas em um fragmento florestal na Amazônia Central. Os tamanhos dos círculos representam a freqüência das lianas nas diferentes classes de diâmetros. que a chance de termos amostrado uma Discussão Nossos resultados mostraram um grande quantidade de indivíduos de padrão evidente de predominância das apenas algumas espécies que se enrolam lianas com enrolamento no sentido anti- no sentido anti-horário é baixa. horário. Esse resultado poderia ser Ao contrário das nossas explicado se o sentido de enrolamento previsões, o sentido de enrolamento não fosse fixo para cada espécie e se as apresentou relação com o crescimento espécies das mais abundantes fossem lianas, indicando que tal enroladas em um mesmo sentido. característica pode não constituir uma Porém, do vantagem adaptativa. Há uma forte sentido de enrolamento em um mesmo tendência para a busca de explicações indivíduo, indicando que o padrão não é funcionais, fixo, pelo menos para algumas espécies. natural, para os padrões biológicos Ainda, dada a alta riqueza de lianas, 306 encontrados na natureza. Porém, nem espécies distribuídas em 44 famílias todas as características dos organismos (Ribeiro et al. 1999), é razoável supor são adaptativas e podem surgir por observamos inversões originadas por seleção 3 mecanismos independentes de seleção sombreamento a norte do ramo, o que (Futuyma 1996, Williams 1996). Por faria a planta crescer para o sul em exemplo, todas as plantas apresentam busca de luz. Ao cair, a inércia forçaria um movimento rotacional durante o o crescimento que é gerado pela força enrolamento no sentido horário. ramo gravitacional da Terra (Kiss 2009). Um possível mecanismo para oeste criando um Em suma, é provável que o explicativo, mecanismo de enrolamento dependa da também independente de seleção, para o fisiologia do crescimento das lianas padrão de enrolamento das lianas seria (fototropismo baseado na interação entre fototropismo sustentado do ramo jovem) e do efeito e efeito de Coriolis. De acordo com essa de explicação, quando o ápice de um ramo enrolamento jovem se alonga, ele tende a cair em efeito uma direção (Richards 1996). A direção crescimento não é uma característica da queda depende do hemisfério do adaptativa e, portanto, não é suscetível à globo terrestre em que a liana se seleção encontra. Ao sul da linha do Equador, a alongamento não sustentado, sem o qual trajetória do Sol é deslocada para o a liana não poderia enrolar-se. Se o norte da abóbada celeste, fazendo com mecanismo que a queda do ramo seja também para podemos esperar um efeito inverso no o norte. Durante essa queda, a inércia sentido de enrolamento das lianas do causada pelo movimento de rotação da hemisfério norte, que deveriam enrolar- Terra (i.e., efeito de Coriolis) faz com se que o ramo se desloque para oeste, uma horário. e Coriolis. de alongamento Se não o sentido de depender somente do direção do Coriolis, natural, a ao proposto predominantemente contrário for no do válido, sentido vez que a rotação terrestre se dá no sentido oeste-leste. O resultado deste Agradecimentos processo é um crescimento em espiral Agradecemos a Gonçalo Ferraz por nos com início após o encontro da liana com orientar na condução deste trabalho, a a planta hospedeira. Já o enrolamento Paulo Enrique Cardoso pela ajuda nas no sentido horário, que acontece menos análises e ao Glauco e Rafael pela frequentemente no hemisfério sul, pode revisão. ser explicado por um forte 4 Referências de terra-firme na Amazônia Chazdon, R. & N. Fetcher. 1984. Central. Manaus: INPA. Photosynthetic light environments Richards, P.W. 1996. The tropical rain in a lowland tropical forest in forest. Cambridge: Cambridge Costa Rica. Journal of Ecology, University Press. 72:533-564. Finegan, B., M. Camacho & N. Zamora. Rundel, P.W. & A.C. Gibson. 1996. Adaptative strategies of growth 1999. Diameter increment form and physiological ecology in patterns among 106 tree species in neotropical lowland rain forest a logged and silviculturally plants, pp. 33-71. In: Neotropical treated Costa Rican rain forest. biodiversity and conservation Forest Ecology and Management, (A.C. Gibson, ed.). Los Angeles: 121:159-176. The Pulse Copy and Technology Futuyma, D.J. 1986. Evolutionary biology. Sunderland: Sinauer Associates. Ingrouille, M. & B. Eddie. 2006. Plant diversity and evolution. New York: Cambridge University Press. Kiss, J.Z. 2009. Plants circling in outer Center. Townsend, C.R., M. Begon & J.L. Harper. 2003. Essentials of ecology. Oxford: Blackwell Publishing. Williams, G.C. 1996. Adaptation and natural selection: a critique of some current evolutionary space. New Phytologist, 182:555- thought. Princeton: Princeton 557. University Press. Ribeiro, J.E.L.S., M.J.G Hopkins, A. Vicentini, C.A. Sothers, M.A.S Costa, J.M. Brito, M.A.D. Souza, L.H.P. Martins, L.G. Lohmann, P.A.C.L. Assunção, E.C. Pereira, C.F. Silva, M.R. Mesquita & L.C. Procópio. 1999. Flora da Reserva Ducke. Guia de identificação das plantas vasculares de uma floresta 5
Baixar