Marco Fábio Maia Corrêa
ICTIOFAUNA DEMERSAL DA BAÍA DE GUARAQUEÇABA
(PARANÁ, BRASIL). COMPOSIÇÃO, ESTRUTURA, DISTRIBUIÇÃO
ESPACIAL, VARIABILIDADE TEMPORAL E IMPORTÂNCIA
COMO RECURSO.
Monografia apresentada ao Curso de PósGraduação em Ciências Biológicas – Zoologia,
Setor de Ciências Biológicas da Universidade
Federal do Paraná, como requisito parcial para
obtenção do título de Doutor em Ciências
Biológicas – Zoologia
Orientador : Henry Louis Spach, Dr.
Curitiba
2001
"As verdades que digo não são
para aqueles que não as
compreendem mas para proteger
aqueles que as desconhecem”
Anônimo .
Este trabalho é dedicado à
André Valério Corrêa
(5/05/1971- 07/07/1995).
ii
iii
AGRADECIMENTOS
A Coordenadoria de Aperfeiçoamento de Pesquisa de Nível Superior –
CAPES/PICD, pela bolsa concedida.
Ao Centro de Estudos do Mar da Universidade Federal do Paraná (CEM), pelo
apoio logístico para o meu desenvolvimento acadêmico e profissional.
Aos colegas Dra. Theresinha Monteiro Absher e Hedda Kolm e aos Biólogos Mário
Barletta, Olga Kimiko Maehama, Jynessa Rillo Dutka Gianelli, Giselda Gomes Bonatti
pelo apoio nas coletas de campo.
Ao amigo e "braço direito" Carlos Serafim ("Seu Carlito"), pelos inúmeros
ensinamentos sobre navegação e pesca, pela paciência, incontáveis madrugadas, apoio,
coragem e determinação para que fosse possível atingir os "nossos objetivos".
Á minha mãe Renée Maia Corrêa por seu exemplo de coragem, alegria de viver,
determinação e por compreender e aceitar que amor é abnegação e sacrifício.
Ao Orientador Henry Louis Spach, pela confiança, aconselhamento e paciência
durante a realização deste trabalho.
A funcionários e amigos do CEM Vilma Sueli de Andrade (administradora), Salete
do Rocio Figueiredo Schmidt (secretária da direção), Elair Siuch do Nascimento (secretária
geral) pelo apoio, eficiência e compreensão nos momentos de ausência.
iv
SUMÁRIO
AGRADECIMENTOS
iv
SUMÁRIO
v
LISTA DE FIGURAS
vi
RESUMO
ix
ABSTRACT
x
1. INTRODUÇÃO
1
2. ÁREA DE ESTUDO
7
3. MATERIAL E MÉTODOS
13
4. RESULTADOS
4.1. Parâmetros físico-químicos e meteorológicos
20
4.2. Ictiofauna
4.2.1. Composição, variabilidade temporal e distribuição espacial
25
4.2.2. Diversidade, eqüitatividade e riqueza
36
4.2.3. Estrutura
4.2.3.1 Comprimento e peso
45
4.2.3.2 Estádio de maturação gonadal
63
4.2.4. Comportamento frente a salinidade
82
4.3. Importância como recurso
4.3.1. Das unidade amostrais
84
4.3.2. Das atividades e equipamentos de pesca
85
4.3.3. Dos locais e petrechos preferenciais de pesca
87
4.34. Da ictiofauna
89
5. DISCUSSÃO
5.1. Parâmetros físico-químicos e meteorológicos
91
5.2. Composição, variabilidade temporal e distribuição espacial
93
5.3. Diversidade, eqüitatividade e riqueza
98
5.4. Estrutura
Comprimento e peso
102
Estádio de maturação gonadal
104
Comportamento frente a salinidade
106
5.5. Importância como recurso
119
6. CONCLUSÕES
113
7. REFERÊNCIAS BIBLIOGRÁFICAS
116
8. APÊNDICES
8.1 Tabelas
132
8.2. Fichas de entrevista
160
v
LISTA DE FIGURAS
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
Mapa do litoral do Estado do Paraná, mostrando a posição geográfica dos pontos
amostrais na Baía de Guaraqueçaba.
Fotografia aérea da Baía de Guaraqueçaba, mostrando a localização dos pontos
amostrais (escala 1:50000- ITCF, julho de 1980)
Especificações da rede de arrasto e das portas utilizadas.
Diagrama comparativo dos valores de pluviosidade (em mm), registrados no momento
e nas ultimas 48 horas de amostragem. Fonte: dados de campo e IAPAR.
Diagramas comparativos das médias de temperatura do ar: a) nos meses e b) pontos de
amostragem, observadas na Baía de Guaraqueçaba.
Valores de profundidade (metros) dos arrastos nos: a) pontos e meses de amostragem e
b) pontos amostrais (resultado da comparação pela análise de variância).
Diagramas comparativos das variações observadas nos parâmetros físico-químicos da
água, de superfície e fundo, da Baía de Guaraqueçaba, nos diferentes meses do anos de
1986 e 1987 e pontos amostrais.
Diagramas comparativos das capturas realizadas entre agosto de 1986 e julho de 1987,
na Baía de Guaraqueçaba (Pr, BR), mostrando as freqüências absolutas e pesos totais
nos meses e épocas do ano(a e c) e anuais por pontos amostrais (b e d).
Resultado da análise de variância de Kruskal-Wallis originada da comparação: a) do
número de exemplares capturados (freqüência absoluta) e b) do peso total das capturas
(em gramas), nas diferentes épocas do ano, na Baía de Guaraqueçaba, Pr, BR.
Resultado da análise de variância de Kruskal-Wallis: a) do número de exemplares
capturados (freqüência absoluta) e b) do peso total das capturas (em gramas), nos
diferentes pontos amostrais, na Baía de Guaraqueçaba, Pr, BR.
Distribuição das freqüências absoluta de captura das famílias de peixes nos diferentes
pontos amostrais da Baía de Guaraqueçaba, Pr, Br.
Distribuição das freqüências absolutas de captura das espécies de peixes nos diferentes
pontos amostrais da Baía de Guaraqueçaba.
Diagrama da distribuição dos pontos amostrais (objetos) e das freqüências das espécies
abundantes (descritores) no plano bidimensional dos componentes principais.
a) Dendrograma dos pontos amostrais, resultante da união "UPGA", sobre uma matriz
de similaridade Bray-Curtis, originada de uma matriz de abundância transformada (??)
e b) ordenação dos pontos amostrais no plano bidimensional pelo método "MDS" com
os grupos do dendrograma sobrepostos nos níveis de 80 (linha contínua) e 82 % (linha
tracejada) de similaridade (stress=0,00).
Diagrama da distribuição das freqüências das espécies abundantes (objetos) e dos
meses (descritores), sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição do peso de captura das espécies abundantes (objetos) e dos
meses (descritores), sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição dos pontos amostrais (objetos) e dos pesos de captura das
espécies abundantes (descritores), sobrepostos no plano bidimensional dos
componentes principais.
Distribuição dos valores dos índices de diversidade (N1) e eqüitatividade de Hill (E5),
respectivamente nos meses (a e b) e pontos amostrais (c e d) obtidos na Baía de
Guaraqueçaba, Pr.
Distribuição dos valores dos índices de diversidade (N1) e eqüitatividade (E5) de Hill
(como descritores) e meses amostrais (como objetos), sobrepostos no plano
bidimensional dos componentes principais.
Distribuição dos valores dos índices de diversidade (N1) e eqüitatividade de Hill (E5),
juntamente com os parâmetros meteorológicos e hidrológicos (como descritores) e dos
meses e pontos amostrais (como objetos abreviados respectivamente por três letras e
algarismos arábicos nos círculos), no plano bidimensional dos componentes principais.
Onde O2 = oxigênio dissolvido, pH = potencial hidrogeniônico, T = temperatura,
Prec./48h = precipitação nas 48 horas anteriores às coletas, Prec./Col.= precipitação no
momento da coleta,, Prof. = profundidade, Sal. = salinidade, S = superfície e F = fundo.
Curvas de rarefação para os diferentes pontos amostrais da Baía de Guaraqueçaba, Pr,
Br. Onde E (Sn) é o número esperado de espécies.
vi
8
14
14
21
21
21
23
26
26
28
28
29
31
31
33
33
35
36
39
41
44
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
Diagramas comparativos: a) do número de exemplares de todas as espécies em
freqüência relativa (%) de captura por intervalos de classe de CT (cm), b) do número
de espécies por intervalos de CT médio observado e CT máximo da literatura; c) da
relação entre o CT médio (+ s) obtido e o CT máximo da literatura e d) número de
exemplares em freqüência relativa (%) de captura por intervalos de classe de peso do
corpo (g).
Comparações do: a) CT (cm) médios e b) pesos médios das diferentes espécies
capturadas na Baía de Giuaraqueçaba. Onde e=erro padrão das estimativas nos limites
de (0,05 e 0,10% de significância).
Resultado das comparações dos: a) CT médios (cm) e b) pesos médios (g) pelos
pontos e períodos do ano amostrados. Onde e=erro padrão das estimativas aos níveis de
0,05 e 0,10 significância.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Cathorops spixii (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Citharichtys arenaceus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Etropus crossotus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Genidens genidens (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Isopisthus parvipinnis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Netuma barba (descritores) nos meses e pontos de amostragem (objetos), sobrepostos
no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Sciadeichtys luniscutis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Micropogonias furnieri (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Sphoeroides testudineus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e
pontos encontram-se sobrepostos e indicados por uma seta.
Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de
Stellifer rastrifer (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
Histogramas comparativos do número de exemplares de todas as espécies (%)
capturados na Baía de Guaraqueçaba por: a) sexo e b) estádios de maturação gonadal.
Histogramas do número de exemplares imaturos (freqüência absoluta) nos meses (a) e
pontos amostrais (b), e dos exemplares que efetivamente participaram do processo
reprodutivo nos meses (c) e pontos amostrais (d).
a) Histograma das freqüências de distribuição do número de espécies capturadas por
diferentes estádios de maturação gonadal e b) diagrama do número de exemplares
capturados nos diferentes estádios de maturação gonadal por espécie.
Diagrama da atividade reprodutiva mensal das espécies mais freqüentes. Onde ausente
= 0; Incipiente <=50; Moderada >50 e <=200; Intensa >200 e <=500 e Muito Intensa >
500 exemplares.
Diagrama da atividade reprodutiva espacial das espécies mais freqüentes. Onde ausente
= 0; Incipiente <=50; Moderada >50 e <=200; Intensa >200 e <=500 e Muito Intensa >
500 exemplares.
vii
45
47
49
50
51
53
54
55
56
58
59
60
61
63
64
66
69
69
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
Diagrama da distribuição das freqüências por estádios de maturação gonadal de
Cathorops spixii (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e
pontos encontram-se sobrepostos e indicados por uma seta.
Diagrama da distribuição das freqüências por estádios de maturação gonadal de
Genidens genidens (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e
pontos encontram-se sobrepostos e indicados por uma seta.
Diagrama da distribuição das freqüências por estádios de maturação gonadal de
Isopisthus parvipinnis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e
pontos encontram-se sobrepostos e indicados por uma seta.
Diagrama da distribuição das freqüências por estádios de maturação gonadal de
Sciadeichtys luniscutis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e
pontos encontram-se sobrepostos e indicados por uma seta.
Diagrama da distribuição das freqüências por estádios de maturação gonadal de
Stellifer rastrifer (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e
pontos encontram-se sobrepostos e indicados por uma seta.
Diagrama da distribuição das freqüências por estádios de maturação gonadal de
Sphoeroides testudineus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e
pontos encontram-se sobrepostos e indicados por uma seta.
Distribuição das freqüências relativas acumuladas de exemplares adultos por pontos
médios de intervalos de classe de CT(cm) nas amostras de a) Cathorops spixii, b)
Genidens genidens e c) Isopisthus parvipinnis obtidas na Baía de Guaraqueçaba.
Fonte:dados de campo.
Distribuição das freqüências relativas acumuladas de exemplares adultos por pontos
médios de intervalos de classe de CT(cm) nas amostras de a) Sciadeichthys luniscutis,
b) Sphoeroides testudineus e c) Stellifer rastrifer obtidas na Baía de Guaraqueçaba.
Fonte:dados de campo.
Diagrama comparativo das distribuições das freqüências de ocorrência de adultos e
juvenis das diferentes espécies, em função da salinidade da água de profundidade em
que foram capturados. Onde "s" = desvio e "e"= erro padrões das estimativas.
Diagrama comparativo das freqüências de declarantes por ocupação principal e
secundária na localidade de Guaraqueçaba. Onde: Fun/Pub= funcionário público,
Pres./Ser. = prestação de serviços, Turis. = turismo, Bar/merc.= bar/mercearia, Cano =
canoeiro, Apos. = aposentado e Agri.= agricultura. Fonte:dados de campo.
Diagrama comparativo das porcentagens de declarantes e a utilização do produto
pesqueiro na localidade de Guaraqueçaba. Onde: Sub./Com. = subsistência e comércio.
Fonte: dados de campo.
Relação entre o número de pescadores e: a) a posse de embarcações na localidade de
Guaraqueçaba e b) comparação entre as médias do número de embarcações movi das a
remo e a motor para os pescadores da localidade de Guaraqueçaba. Fonte: dados de
campo.
Diagrama comparativo: a) das médias dos comprimentos das embarcações movidas a
remo e a motor na localidade de Guaraqueçaba. Fonte: dados de campo.
Diagrama comparativo entre o número médio de petrechos de pesca por pescador para
a localidade de Guaraqueçaba. Fonte: dados de campo.
Diagrama mostrando as distribuições dos pescadores, da localidade de Guaraqueçaba,
segundo o número de horas gastas nos locais preferencias de pesca. Fonte: dados de
campo.
Comparação entre o número médio de horas gastas, pelos pescadores de Guaraqueçaba,
nas fainas de pesca nos diferentes locais segundo a ordem de preferência.
Diagrama das distribuições dos pescadores, da localidade de Guaraqueçaba, segundo
os petrechos de pesca e locais onde são utilizados. Fonte: dados de campo.
Plotagem simultânea e ordenação, no espaço dos componentes principais (ACP), dos
descritores (abundância declarada dos recursos na localidade de Guaraqueçaba)
segundo as observações (espécies ictiícas). Fonte: dados de campo.
viii
70
72
74
75
77
78
80
81
82
84
85
85
86
86
87
88
88
90
RESUMO
A composição, estrutura, distribuição espacial, variabilidade temporal da ictiofauna
demersal da Baía de Guaraqueçaba foi estudada, através de amostragens quinzenais, entre
agosto de 1986 e 1987. As amostras foram obtidas por arrastos com portas, com duração
padronizada de 15 minutos, em 4 pontos denominados: 1) furo de Guaraqueçaba, na
porção mais externa e interligada ao sistema estuarino da Baía de Paranaguá, 2) a sudeste
da Ilha do Sambaqui, na porção mediana direita da Baía e entre os morros da Vila e do
Franco, 3) ao norte da Ilha do Sambaqui, na porção mais interna da Baía e no delta
formado pelos Rios Morato, Moratinho e Ipanema e 4) a oeste da Ilha do Sambaqui, na
porção mediana esquerda da Baía e à frente da Serra do Tromomô. Como complemento
foram levantados aspectos sócio-econômicos, da pesca através de 46 entrevistas realizadas
com pescadores de Guaraqueçaba, em setembro e outubro de 1996 e abril de 2000.A
ictiofauna demersal estava composta por 19 famílias, 42 gêneros e 52 espécies de
Actinopterygii. A ictiofauna esteve principalmente representada por espécies comuns nos
estuários tropicais e subtropicais. A maior abundância foi observada no outono e na porção
mais externa; o maior peso de captura foi obtido na primavera e na porção mais externa.
As famílias Ariidae, Sciaenidae, Paralichthyidae e Tetraodontidae dominaram tanto em
freqüência quanto em peso. As espécies Cathorops spixii, Stellifer rastrifer, Genidens
genidens e Sciadeichthys luniscutis foram numericamente dominantes e as três primeiras,
mais Sphoeroides testudineus, apresentaram maior peso de captura. A composição
ictiofaunística dos pontos amostrais foram muito similares, contudo as diferenças nas
abundâncias foram capazes de caracterizá-los como distintos. A região mais interna da
Baía distinguiu-se das demais pela dominância numérica de G. genidens. A diversidade e
eqüitatividade, nos pontos e épocas do ano, foram principalmente afetadas pela redução na
abundância de C. spixii e S. rastrifer, e apresentaram os maiores valores associados: a) às
maiores salinidades (superficial e de profundidade), pH (superficial e de profundidade),
oxigênio dissolvido (de profundidade) e transparência e b) às menores precipitações,
profundidades e teores de oxigênio dissolvido (superficial). Desta forma, entre outros, os
parâmetros meteorológicos (pluviosidade) e hidrológicos (salinidade e profundidade)
estariam desempenhando o principal papel na alteração da composição, diversidade,
eqüitatividade, variabilidade temporal e distribuição espacial da ictiofauna. As áreas
analisadas apresentaram valores de riqueza muito similares. As populações naturais de 38
espécies (73,07%) estavam constituídas por indivíduos com CT médios 50% inferiores ao
CT máximo relatado na literatura sendo em sua maioria composta por juvenis.. As médias
de CT e de peso foram inferiores a 15 cm e 50g em todos os pontos e épocas amostradas.
Ainda, foram observados padrões de estratificação espaço-temporal no CT e pesos médios,
que caracterizaram movimentos freqüentes entre as áreas da Baía e explicaram e
corroboraram o uso (reprodução e recrutamento) e ocupação pela ictiofauna. Foram
também constatadas atividades reprodutivas intensas para C. spixii, S. rastrifer, S.
testudineus e Isopisthus parvipinnis; moderadas para G. genidens e S. luniscutis que
apresentaram preferências temporais e espaciais para reprodução e recrutamento. De modo
geral a reprodução ocorreu entre a primavera e início de verão e o pico do recrutamento no
outono. A comunidade de pescadores de Guaraqueçaba é dependente do peixe como
principal fonte de alimento e a pesca está limitada por motivos operacionais às regiões
próximas das comunidades. As espécies demersais declaradas pelos pescadores como
freqüentemente capturadas coincidiram com aquelas mais abundantes nos arrastos e as
mesmas podem ser consideradas como importante fonte de recursos.
ix
ABSTRACT
The present study investigated the fish species assemblages found in the Guaraqueçaba
Bay. Moreover, this study describes the seasonal changes of the fish species composition
in relation to abundance in number and weight. Fishes were collected at regular intervals
(every 15 days) between August 1986 and July 1987. Samples were taken at 4 stations (1:
Furo de Guaraqueqaba, 2: south-eastern of Ilha do Sambaqui, 3: northem of Ilha do
Sambaqui, 4: western of Ilha do Sambaqui), locaied in Guaraqueqaba Bay, with an otter
trawl. In addition, 46 interviews, in September and October/ 1996 and April/2000, were
done to characterise some aspects of the socio-economy of the fisheries in the region. The
demersal fish species was composed of 19 families, 42 genera and 52 Actinopterygii
species. The fish fauna were represented by many species common to other estuaries in
tropical and subtropical regions located along the Brazilian coast. The greatest capture in
number (15641 fishes) was observed in autumn at the entrance of the bay (station 1).
Greatest capture in weight (236,093.60g) was observed during spring also at the station l.
The most important families in abundance (number and weight) were: Ariidae, Sciaenidae,
Paralichthyidae and Tetraodontidae. Cathorops spixii, Stellifer rastrifer, Genidens
genidens and Sciadeichthys luniscutis were the most important species in number of
individuals, and C. spixii, S.rastrifer, G. genidens and Sphoeroides testudineus showed the
greatest capture in weight. The species composition of each station was very similar,
however, the abundance within the stations were significantly different. G.genidens
showed more individuals in the inner part of the bay. The Hill’s index (Nl) and equitativity
(E5), at each station and during each season, were principally influenced by the reduction
in number of individuals of C. spixii and S. rastrifer during the rainy season (salinity).
Therefore, salinity and rainfall were the variables that controlled the fish species
composition in the estuary. Species richness was also similar between stations. From 52
species captured in the estuary 73.1% showed more juvenile stage with a total length (TL)
(average 50%) smaller than maximum TL found in the bibliography. The average length
and weight for all stations and seasons were less than 15 cm and 50g respectively.
Moreover, movements in the fish assemblage were observed (principally for C. spixii,S.
rastrifer, S. testudineus, Isopistus parvipinis, G. genidens and S luniscutis), between the
station within the bay. These fish movements within the bay were correlated to
reproduction and recruitment. Reproduction time principally occurs between spring and
early summer, and recruitment occurs in autumn. Some socio-economics aspects about the
artisanal fisheries in the Guaraqueqaba Bay are also discussed.
x
1
1. INTRODUÇÃO
As águas mesohalinas protegidas, dos ecossistemas estuarinos e da plataforma
continental marinha, interconectam e interagem através dos canais estuarinos. Através
deles ocorre um intercâmbio ativo de organismos (Weinstem 1988), água (Drunkers
1988 e Seaberg 1988), nutrientes (Peterson et al. 1988), matéria orgânica (Hopkinson
1988) e sedimentos (Nitrouer & DeMaster 1986), entre as marismas e o oceano
adjacente. Isto envolve processos complexos de transporte e mistura (Jansson 1988,
Madden et al. 1988, Kjerfve 1986, Kjerfve & Magill 1989), movimentos migratórios
(Yánez & Nugent 1977, Yãnez-Arancibia et al. 1985, Weinstem 1988), variações de
diversidade e abundância biótica (Livingston 1976, Alvarez-Guillen et al. 1985, Vieira
& Musick 1993); mudanças ontogenéticas nos ciclos de vida (Yãnez-Arancibia et al.
1985 e 1988) além de alterações da dinâmica trófica, produtividade, distribuição
espaço-temporal dos organismos e nos gradientes físico-químicos (Darnell & Soniat
1979, Stevenson 1988). A amplitude destes processos apresenta relação com a
geomorfologia e a hidrodinâmica dos estuários (Odum et al. 1979). Portanto, o estudo
dessa regiões implica na observação das inter-relações entre rio, estuário e oceano.
Ao longo dos últimos 20 anos foi corroborado o conceito de que os estuários
constituem habitats essenciais para
inúmeras espécies de peixes comercialmente
importantes (Saenger & McIvor 1975). Muitas das etapas reprodutivas, incluindo
dispersão de ovos e larvas, distribuição de juvenis e migrações (pouco compreendidas),
estão sincronizadas com o funcionamento dos ambientes estuarinos (Robins 1971,
Bochlert & Mundy 1988, Shaw et al. 1988, Weinstem 1988). Destacam-se nestes
ambientes as florestas de manguezais entre-marés que, embora contenham poucos
habitats diretamente utilizáveis pelas espécies comercialmente importantes (com notável
2
exceção para as ostras), podem fornecer às águas adjacentes nutrientes adicionais (Lao
1983). O aumento do “input” de nutrientes resulta em acréscimo na produtividade
pesqueira (Saenger & McIvor 1975, FAO/UNEP 1980, Gomez 1980, Baglo 1983).
Análises amostrais quantitativas têm demonstrado que maior número de indivíduos
e maior diversidade específica, ocorrem nas águas estuarinas rasas, entre os 30 cm e 1,5
m de profundidade (Hamilton & Snedacker 1984). Reforçam este conceito os
levantamentos ictiofaunísticos marinhos e/ou estuarinos que foram efetuados nos
Estados Unidos, África, Japão e ainda países da Europa, América Central (México) e
América do Sul (Venezuela, Colômbia, Peru e Argentina) (Schultz 1949, Cervigón
1966, Cervigón & Lima 1972, Fischer 1978, UNESCO 1979 e 1980, Castro-Aguirre &
Márquez-Espinoza 1981, Cervigón 1992).
No Brasil, a alta diversidade dos peixes dos ambientes estuarinos pode ser inferida
através das procedências das espécies relacionadas em inúmeros levantamentos e
inventários ictiofaunísticos, entre os quais: os de Fowler (1941), Ribeiro (1907, 1911 e
1915), Carvalho (1941, 1950, 1951), Travassos & Paiva (1957), Pinto (1960), Carvalho
et al. (1968), Vazzoler (1970), Santos (1970), Faleiro (1971), Travassos & Rego-Barros
(1971), D’Incao (1973), Jardim (1973), Roux (1973), Castello (1974), Travassos &
Lima (1975), Vazzoler (1975), Oliveira (1976), Figueiredo (1977), Figueiredo &
Menezes (1978 e 1980), Lema et al. (1979 e 1980), Menezes & Figueiredo (1980 e
1985), Costa (1980), Lucena & Lucena (1981a,b ), Vazzoler et al. (1982), Chao et al.
(1982, 1985), Nonato et al. (1983).
Andreata et. al. (1992) coletaram em 4 áreas da Laguna de Jacarépaguá, 11 famílias
e 15 espécies de peixes. Poeciliidae e Jenynsiidae foram dominantes e estavam
representadas respectivamente pelas espécies Poecilia vivipara, Phalloptychus
januarius, Jenynsia lineata e Xenomelaniris brasiliensis. Consideraram diversidade,
3
riqueza e eqüitatividade baixas. Ainda Vieira & Musick (1994) sumariaram, com base
na literatura e no tipo de aparelho utilizado nas amostragens, a composição
ictiofaunística (principalmente de Actinopterygii) dos estuários tropicais e temperadoquentes do oeste do Atlântico (entre as latitudes de 37o N e 35o S). Constataram que no
norte e nordeste do Brasil (entre as latitudes 8o N e 11o S) foram registradas 94
famílias, 244 gêneros e 427 espécies. Nesta região foram particularmente abundantes as
famílias Ariidae, Sciaenidae, Mugilidae, Haemulidae e Gerreidae. Consideraram a
diversidade baixa para a região de Cabo Frio (23oS) onde foram registradas 37 espécies,
sendo mais abundantes as famílias Atherinidae, Cichlidae, Clupeidae, Gerridae e
Poecilidae. Entre São Paulo (24o S) e Rio Grande do Sul (32o S) registraram 70 famílias,
145 gêneros e 211 espécies, com dominância de Sciaenidae, Ariidae, Mugilidae,
Clupeidae e Atherinidae na maioria dos estuários.
No litoral do Estado do Paraná a literatura registrou 92 famílias, 191 gêneros e 313
espécies, dos quais 80, 179 e 289, respectivamente corresponderam exclusivamente aos
Actinopterygii (Couto & Corrêa, 1992, Castello et al. 1994, Godefroid, 1997 e Chaves
& Corrêa, 1997). Apresentaram maior número de espécies, entre os Chondrichthyes a
família Carcharhinidae (7) e entre os Actinopterygii, a família Myctophidae (37) na
região oceânica (ou plataforma) e Sciaenidae (22) e Carangidae (17) nas regiões
costeiras e oceânicas. Dentre as espécies das regiões costeiras e estuarinas,
aproximadamente 59,74% foram consideradas demersais, 40,26% pelágicas, 88,9%
exclusivamente costeiras, 10,2% costeiro-estuarinas e 0,9% exclusivamente marinhas.
Aproximadamente 54,22%, por seu tamanho e/ou comestibilidade, apresentaram
importância comercial e 45,78% poderiam ser utilizadas como subprodutos (Castello et
al. 1994).
4
Para o sistema estuarino da Baía de Paranaguá foram relatadas 28 e 173 espécies,
respectivamente, de Chondrichthyes e Actinopterygii. Entre os últimos foram
particularmente abundantes as famílias Mugilidae, Ariidae, Sciaenidae, Gerreidae,
Atherinidae, Clupeidae, Carangidae, Serranidae e Tetraodontidae (Castro & Busetti
1985, Corrêa et al. 1986, 1987 e 1988, Corrêa 1987 e 1988, Cunha 1988, Barletta &
Corrêa 1989a,b , Barletta et al. 1989, Maehama & Corrêa 1987, Corrêa & Maehama
1988, Souza 1988, Silva 1989, Silva & Pose 1990, Couto & Corrêa 1992, Barletta et al.
1990, Abilhôa et al. 1997, Lemos et al. 1997, Pinheiro et al. 1997, Pinto et al. 1997,
Godefroid et al. 1997a,b , Bazaluk 1996 e Bazaluk et al. 1997). Aproximadamente 66
espécies eram explotadas por 70,59 % dos pescadores e todas apresentam seu ciclo de
vida, ou parte dele, essencialmente associado com as águas costeiras e estuarinas
(Corrêa et al. 1997).
No sistema em pauta, a composição específica apresentou maior similaridade com a
fauna de águas quentes tropicais ou Caraíbicas (Corrêa 1987 e Godefroid 1997). Por se
tratar de uma área intermediária entre os rios e o oceano, a região está diretamente
relacionada com as migrações dos peixes (Zijlstra 1986 e Castelo et al 1994). Em
hipótese, temperatura e salinidade da água (associada à pluviosidade e às marés) seriam
os principais fatores determinantes da distribuição (Corrêa et al., 1987 e 1995).
A pesca no sistema estuarino da Baía de Paranaguá é caracterizada como artesanal
e de subsistência, com a comercialização dos excedentes de produção (Kraemer 1982,
Corrêa 1987 e 1997, SPVS 1992). Os últimos são comercializados nos mercados
regionais, com alto grau de intermediação (Kraemer, 1982). Acima de 70% da
população das comunidades tradicionais tem na pesca sua ocupação principal e/ou
secundária, com tendência geral ao abandono das outras atividades (SPVS 1992 e
Corrêa et al. 1997).
5
Com base, nas espécies de peixes exploradas, tipos de petrechos e atividades de
pesca, foram distinguidos dois segmentos produtivos: de pesca estuarina (representado
pelas populações tradicionais da região interna da Baía) e de pesca marinha/costeira
(representado pelas comunidades das praias adjacentes e das barras de acesso à
Baía)(SPVS, 1992 e Corrêa et al. 1997).
Os dados de controle e desembarque pesqueiros encontram-se, em sua maioria,
indisponíveis (Kraemer 1982) defasados ou incipientes (Paiva 1996). Segundo Corrêa
(1987), do total de peixes comercializado no mercado do peixe de Paranaguá em 1982,
90,61% procediam do sistema estuarino da Baía de Paranaguá e o restante, das praias e
Estados limítrofes. Do total procedente do sistema, 41,2% tinham origem da região da
Baía das Laranjeiras e 38,97% da Baía de Paranaguá propriamente dita. As praias
adjacentes contribuíram com 0,68% da produção total. A estação mais produtiva estava
compreendida entre o início do verão e o final do inverno.
Simultaneamente às pescarias de rotina, são praticadas em determinadas regiões e
estações do ano a pesca da manjuba (20 e 31 espécies, respectivamente, estuarinas e
marinhas-migrantes), da tainha (Mugil liza) e do cação (várias famílias ) (Barletta &
Corrêa 1989a,b , Corrêa 1994, Spach et al. 1995a , Bazaluk et. al. 1997).
As informações sobre a pesca artesanal do litoral do Paraná apontam para o
esgotamento dos estoques o que, por sua vez, sugere o agravamento do quadro sócio
econômico das comunidades pesqueiras tradicionais (IPARDES 1989a,b , Cunha et al.
1989, SPVS 1992, Corrêa et al. 1997, Karam & Toledo 1997).
Na região de Guaraqueçaba, até então, os estudos relacionados à ictiofauna
abrangeram aspectos, taxonômicos (Ottmann et al. 1992 e
Pinheiro et al. 1994),
parasitológicos (Silveira et al. 1997), biológicos (Gomes-Bonatti 1988, Dutka-Gianelli
& Corrêa 1988), da composição específica e estrutura da pesca do iriko (Ottmann et al.
6
1992, Spach et al. 1995a,b , Bazaluk 1996 e Bazaluk et al. 1997 e Melo et al. 1997), da
composição ictiofaunística (Corrêa et al. 1987 e 1995) e da estrutura social,
relacionados à produção pesqueira (Cunha et al. 1989, Rougeulle 1992, Karam &
Toledo 1997).
De modo geral, os trabalhos mostraram que, no sistema estuarino-baía de
Paranaguá, os conhecimentos sobre a composição e a diversidade ictiíca evoluíram
rapidamente. Contudo, a estrutura e os padrões de distribuição temporal e espacial,
principalmente nas gamboas, nas Baía de Antonina e Guaraqueçaba, e nas enseadas do
Benito e do Itaqui ainda são praticamente desconhecidos (Couto & Corrêa 1982, Corrêa
1994).
O presente trabalho tem por objetivos principais:
a) analisar, por estações do ano e pontos amostrais, a composição específica,
distribuição espacial, variabilidade temporal e estrutura (em peso, comprimento e
proporção sexual) da ictiofauna demersal da Baía de Guaraqueçaba,
b) caracterizar a ictiofauna levantada em relação ao seu comportamento:
- frente a salinidade, em dulcícolas, estuarinos, estuarino-marinhos e
marinhos,
- migratório em residentes e visitantes ocasionais ou temporários,
c) caracterizar meses e estações do ano e a região levantada, em relação a
utilização para a reprodução e/ou crescimento da ictiofauna.
d) caracterizar a ictiofauna observada quanto a sua importância econômica e
social.
7
2. ÁREA DE ESTUDO
O sistema estuarino da Baía de Paranaguá, com uma área líquida de 601 km2 e
o
o
o
o
coordenadas geográficas gerais 25 15’a 25 30’ S e. 48 02’ a 48 45’W, projeta-se
continente adentro na porção norte da planície litorânea do Estado do Paraná (Fig.1).
Comunica-se com o oceano através de canais, separados pela Ilha do Mel, denominados de
Sueste ou Norte (ao norte) e da Galheta (ao sul). Internamente compartimenta-se em
diversos segmentos menores distribuídos ao longo de três eixos principais: a) Baía de
Paranaguá propriamente dita, com orientação leste-oeste à partir da Ilha do Mel, extensão e
largura máximas de 56 por 7 km e cujo extremo oeste é denominado de Baía de Antonina,
b) Baía das Laranjeiras com orientação norte-sul a partir da Ilha das Cobras, extensão e
largura máximas de 30 por 13 km e c) Baía do Pinheiros, de orientação sudeste-noroeste,
extensão e largura máximas de 15 por 3km, que separa a Ilha das Peças da Ilha do Superagüi
e comunica-se com a Baía das Laranjeiras por um canal estreito situado ao norte da Ilha das
Peças e com o Mar de Ararapira através do canal artificial denominado Varadouro (Yoneda
& Absher 1992, IPARDES 1995 e Soares & Barcelos 1995).
O clima da área é do tipo Cfa de Köeppen – pluvial temperado, sempre úmido, com
o
chuvas em todos os meses do ano e temperatura média do mês mais quente acima de 22 C
o
(Bigarella 1978). O mês mais quente é o de janeiro com temperatura média de 25 C e o
o
mais frio o de julho com temperatura média de 17 C (Maack 1981). Conforme IPARDES
(1990), as chuvas são dos tipos ciclônico, orográfico e de convecção. Os meses de maior
pluviosidade são os de verão; fevereiro e julho apresentam respectivamente, o maior (média
304 mm) e o menor (média 61 mm) índice acumulado. Os índices hídricos, segundo a
classificação de Thornthwaite, são sempre superiores a 60, caracterizando tipos úmido (60
a 100) e superúmido (>100) (IAPAR, 1978). Ocorrem em média 20 dias claros ao ano
8
3
4
2
1
1
Paraná
N
Figura 1. Mapa do litoral do Estado do Paraná, mostrando a posição geográfica dos pontos
amostrais na Baía de Guaraqueçaba.
9
(Maack 1981). Predominam na área ventos entre E e SE (41%) com velocidades entre 4 e 6
m/s (INPH 1988).
O mapeamento geológico de superfície da planície costeira vem sendo detalhado e
revisado desde o trabalho pioneiro de Bigarella (1946). Cartas geológicas para a região
foram publicadas por Bigarella & Doubek (1963), Rivereau et al. (1969), Martin et al.
(1988) e Angulo (1992). O último mapeamento geológico (Angulo, 1992) foi feito na escala
1:50000, dividindo a planície costeira nas seguintes unidades sedimentares: 1) Continental,
subdividida em Formação Alexandra, leques aluviais, talus, colúvios e sedimentos fluviais;
2) Costeira (terraços marinhos), subdividida em dunas, depressões intercordões e praias; 3)
Estuarina, subdividida em planícies de maré, fundos rasos, deltas de maré e sedimentos
paleo-estuarinos. Os sedimentos costeiros pertencem a dois tipos de sistema principais: o
de planície costeira com cordões litorâneos e o estuarino. Ambos são representados por
ambientes antigos formados durante períodos em que o mar tinha níveis relativos mais altos
que o atual e por ambientes atuais, tais como praias, planícies de maré, deltas de maré e
dunas frontais.
Dentre os estudos de processos hidrodinâmicos e sedimentares destacam-se
levantamentos de distribuição de salinidade, temperatura e marés (IPqM, 1969), sistemas de
drenagem e sedimentologia das Baías (Bigarella et al. 1970, Bigarella et al. 1978, Soares
& Barcelos 1995), comportamento físico-químico, aporte de água doce e tempo de
residência (Knoppers et al. 1987), causas das marés meteorológicas na região (Marone &
Camargo 1994), quantificação da clorofila, parâmetros químicos e material particulado em
suspensão (Brandini et al. 1985 e 1988) e sobre a dinâmica estuarina (Marone et al. 1995).
De acordo com Knoppers et al. (1987) e Machado et al. (no prelo), o sistema
estuarino da Baía de Paranaguá exibiu um padrão de estratificação salina somente em
situações de fortes marés vazantes, que ocorreram principalmente no período chuvoso
10
(verão). Nos demais períodos, mostrou variação espacial, com águas bem misturadas no
setor externo e progressiva estratificação em direção ao interior da baía. Neste contexto,
uma coluna d’água praticamente homogênea, em todo o sistema, tem sido constatada após
fortes ventos. Estas informações demonstram que a hidrodinâmica da baía é fortemente
influenciada por fatores climáticos (regime de chuvas e ventos), que regulam a intensidade
das marés (Machado et al. no prelo).
Brandini & Thamm (1994) e Machado et al. (no prelo) observaram que a temperatura
apresentava um padrão de distribuição espacial semelhante ao da salinidade. Porém as
variações verticais e horizontais eram de pequena magnitude. Verificaram que a
transparência e o pH exibiam uma relação direta com a salinidade, com valores mais
elevados ocorrendo no período seco de inverno. Em diversas ocasiões, foram registrados
valores de pH mais elevados nas regiões mediana e interna da baía do que na área de maior
influência marinha, sendo o evento atribuído às atividades fotossintéticas (Brandini &
Thamm 1994, Machado et al. no prelo). O oxigênio dissolvido na água de superfície não
apresentou padrão definido, porém o de fundo exibe um gradiente decrescente em direção às
regiões internas da baía. O menor valor (64 % saturação) foi registrado para a água de
fundo da seção mais interna da baía. Este comportamento foi atribuído à variabilidade
espacial nos processos de produção e respiração na coluna d'água e, provavelmente, nos
sedimentos superficiais (Machado et al. no prelo). Relativamente ao material particulado
em suspensão (seston) Machado et al. (no prelo) registrou uma variabilidade acentuada,
com valores variando entre 8,1 e 91,5 mg?dm-3 no setor mediano da baía. Contudo as
variações espaciais e temporais desviaram do padrão esperado, de incremento com a
diminuição da salinidade, possivelmente em conseqüência da complexa hidrodinâmica do
sistema e de processos de ressuspensão dos sedimentos superficiais e penetração da água de
fundo de maior salinidade durante a maré enchente. Em relação à composição do material
11
orgânico particulado, Brandini (1985) verificou que este é constituído basicamente de
detritos vegetais provenientes dos manguezais que circundam a baía e, em menor extensão,
de células fitoplanctônicas. Machado et al. (no prelo) verificaram um padrão de variação
sazonal na composição do material orgânico particulado, em termos da razão carbono
orgânico particulado/clorofila-a: os valores mais elevados foram observados no outono e
no inverno e os menores na primavera e, principalmente no verão. Segundo os autores, este
padrão é conseqüência da variação temporal na magnitude relativa das fontes de matéria
orgânica particulada para o sistema, com maior contribuição do material fitoplanctônico
durante o verão.
As marés e sistemas de correntes ainda são pouco conhecidos. A maré em Paranaguá
foi definida como uma semidiurna pura, com aparecimento de uma maré secundária de três
horas de semiperíodo quando o ciclo lunar se afastando das sizígias se aproxima das
quadraturas. A amplitude foi citada como inferior a 2 m (regime de micromarés) e a
velocidade média da enchente, estimada para o Canal Norte (ou Sueste), em 1,6 nós (0,82
m/s), elevando-se um pouco mais nos períodos da baixa-mar (Bigarella 1978).
Segundo Knoppers & Opitz (1984) e Knoppers et al. (1987) o sistema pode ser
classificado como parcialmente misturado, com tendências à estratificação em períodos de
chuva intensa. No inverno a coluna d’água apresentou-se quase homogênea e a correnteza
máxima atingiu 2,2 nós (1,13 m/s) antes da preamar.
A Baía de Guaraqueçaba, com uma área submersa aproximada de 32 km2 e coordenadas
geográficas gerais 25o 17’ 00” S e 48o 20’ 38”W, está situada no extremo norte da Baía das
Laranjeiras. É delimitada ao sul pela Ilha Pavoça e ao norte pelo delta estuarino, com várias
ilhas rasas planas, formado pelos Rios Morato, do Moratinho e Ipanema. Apresenta
profundidades predominantemente inferiores a 10 m e, nas áreas mais interiorizadas com
características francamente estuarinas, inferiores a 5 m (IPARDES 1989a e 1995). As
12
margens estão circundadas por manguezais e planícies de restinga (IPARDES 1989a e
1995).
A entrada de sedimentos foi estudada por Soares & Barcelos (1995) e ocorre nas
porções mais internas onde deságuam os rios da bacia hidrográfica das Laranjeiras. O
material deste setor deposita-se preferencialmente num baixio ao sul das Ilhas Pavoça e do
Rabelo, é bastante heterogêneo e resulta em alta diversidade textural fortemente influenciada
pela presença de finos. O grau de seleção é muito baixo, indicando pouco transporte desde a
sua remoção da área fonte até o estuário. Já a Baía de Guaraqueçaba possui características
sedimentológicas distintas por receber diretamente os sedimentos da Serra do Mar. Nesta
área a maior parte dos sedimentos, transportados por tração, é retida pelos manguezais que
atuam como verdadeiros filtros e são responsáveis, por exportação, pela elevada
percentagem de matéria orgânica.
Wisniewski et al. 1997 caracterizaram o clima da região de Guaraqueçaba (com base
no período de 1878 a 1995), como do tipo Cfa, com temperatura média anual de 20,9 oC. A
média das máximas é de 26,3 e das mínimas 17,2oC. O mês mais quente é o de fevereiro
com temperatura máxima absoluta de 40,2oC e o mais frio o de junho com temperatura
mínima absoluta de 0,4oC. A média anual de umidade relativa é 85,4% e a de precipitação
2364,80 mm, com máxima precipitação em janeiro (média 659 mm) e mínima em julho
(média 8,1 mm). A média anual de dias com chuva foi de 207, com a média do número
máximo de 252 e mínimo de 127 dias.
13
3. MATERIAL E MÉTODOS
Durante 12 meses (agosto de 1986 e julho de 1987), foi realizado um total de 96
arrastos com duração padronizada de 15 minutos. Os pontos fixos de coleta (Fig. 2)
compreenderam as seguintes áreas: 1) no furo de Guaraqueçaba, ponto mais externo e
local de entrada das águas salinas provenientes do sistema, 2) na porção interna mediana
esquerda, a sudeste da Ilha do Sambaqui e entre os Morros da Vila e do Franco; 3) no
delta formado pelos Rios do Morato e de Guaraqueçaba, ao norte da Ilha do Sambaqui
e, 4) a na porção interna mediana direita, a oeste da Ilha do Sambaqui, à frente da Serra
do Tromomô.
O aparelho utilizado foi a rede de arrasto com portas com malha de 3 cm (entre nós
consecutivos) e 8 x 10 m, respectivamente de boca e comprimento. Foram utilizadas
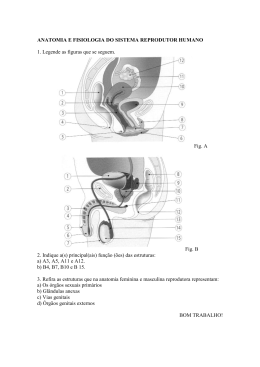
portas com 15 kg e 0,42 x 0,74 m respectivamente de altura e comprimento (Fig.3).
Em todos os pontos amostrais, foram registrados os seguintes parâmetros físicoquímicos, em superfície e fundo, a partir de amostras de água obtidas com a garrafa de
Van Dorn: salinidade (através de refratômetro ATAGO escala 0/40‰), potencial
hidrogeniônico (através de pHmetro portátil digital), temperatura (termômetro químico
escala 1/40°C), oxigênio dissolvido (por titulação, segundo Winkler modificado),
transparência da água (disco de secchi), profundidade (ecobatímetro digital) e tipo de
sedimento (com pegador Van Veen e análise visual).
Os dados de pluviosidade (mm), foram coligidos na estação meteorológica de
Guaraqueçaba (25o 13’S e 48o 48’W, 60 m a.n.m) e gentilmente cedidos pelo serviço de
meteorologia do IAPAR (Instituto Agronômico do Paraná).
Todos os peixes capturados foram congelados e transportados ao laboratório para
identificação ao nível genérico e específico. Foram quantificados, mensurados
14
3
Ilha
do
Sambaqui
2
4
? = 25o17’52,01’’ S
UTM 7.199.390,83 N
Guaraqueçaba
Ilha
do
Pavoçá
1
N
3
? = 48o 19’ 47,72’’ W
UTM 768.846,99 E
Figura 2. Fotografia aérea da Baía de Guaraqueçaba, mostrando a localização dos pontos amostrais (escala
1:50000- ITCF, julho de 1980)
0,42 m
0,74 m
8
m
15 kg
10 m
Figura 3. Especificações da rede de arrasto e das portas utilizadas.
15
(comprimento total ou CT- distância que vai do focinho até a extremidade da nadadeira
caudal, e padrão ou CP- distância que vai da ponta do focinho até o pedúnculo caudal,
obtidos com auxílio de ictiômetro) e pesados (peso total com auxílio de balança com
capacidade para até 5 kg). Foram ainda dissecados para observação do sexo e o grau de
maturação gonadal (segundo escala de Nikolski 1963). Para a família Ariidae, que
apresenta incubação oral de ovos e larvas, foi adicionado o estádio incubando ovos e
larva. Alguns exemplares foram fixados em formol a 10%, preservados em álcool 70 %
e depositados na coleção ictiológica do Centro de Estudos do Mar em Pontal do Sul
(PR).
Para a análise da diversidade (riqueza e eqüitatividade) e eqüitatividade foram
utilizados os índices N1 e E5 de Hill (1973) que são considerados como menos
ambíguos, mais informativos e fáceis de interpretar (Ludwig & Reynolds 1988). Ainda,
o índice de Hill independe do número total de indivíduos na amostra e representa o
número efetivo de espécies na amostra e E5 tende a zero quando alguma das espécies
torna-se mais e mais dominante na comunidade (Ludwig & Reynolds, 1998). Peet
(1974) inclusive sugere o uso deste índice por ser o mais sensível às alterações
quantitativas das espécies raras na composição de uma comunidade. Os índice de
diversidade de Hill é :
N1 = e (H’)
onde H’ é o índice de Shannon , que é estimado em uma amostra por:
Onde ni é o número de indivíduos pertencente a iésima das S espécies na amostra e n é o
número total de indivíduos na amostra.
16
O índice de eqüitatividade E5 é conhecido como a relação de Hill modificada e
representado por:
onde ? é o índice de diversidade de Simpson que é estimado por:
cujos S, n e ni já foram referidos no índice de Shannon, que também está incluído na
fórmula de E5 como H’.
Também foi utilizado o método de rarefação que permite a comparação do números
de espécies entre comunidades pela eliminação do erro provocado pelo tamanho
amostral (Ludwig & Reynolds,1988).
Segundo Schluter & Ricklefs (1993) as comunidades são usualmente definidas por
associações espaciais, funcionais ou taxonômicas ou por interações dinâmicas dentro da
rede trófica. Aqui o termo foi empregado excluindo-se o aspecto trófico.
Para a comparação da similariedade entre os pontos amostrais foi utilizado o
coeficiente Bray-Curtis, a partir da matriz de abundância por espécies (? ?)
transformada, submetido ao agrupamento hierárquico com ligação média ("group
average linking") ou "hierarchical cluster" e à ordenação através da "Non Metric Multi
Dimensional Scaling" (NMDS) (Baroche & Saporta 1982, Legendre & Legendre 1983,
Ludwig & Reynols 1988 e Clarke & Warwick 1994).
Para determinação dos padrões de variabilidade temporal e de distribuição espacialna composição, abundância e diversidade- da ictiofauna e suas relações com os
parâmetros ambientais, os dados foram centrados e reduzidos e submetidos à análise dos
componentes principais -ACP (Baroche & Saporta 1982, Legendre & Legendre, 1983,
17
Ludwig & Reynolds 1988, Rohlf 1989, Johnson & Wichern 1992). A escolha da ACP
baseou-se na sua efetividade para análise de interações complexas. Tradicionalmente (e
foi o caso) esta matriz consiste da variância - covariância ou correlação. Na ACP, os
valores (denominados Eigen) são extraídos em ordem decrescente de magnitude de
modo que os eixos correspondentes ao PCA (componentes) representam a soma
sucessiva da, maior para a menor, variação na matriz (Baroche & Saporta 1982,
Legendre & Legendre, 1983, Ludwig & Reynolds 1988, Rohlf 1989, Johnson &
Wichern 1992, Manly, 1994, Underwood, 1998).
Os resultados, para determinação da estrutura (em peso, comprimento e maturação)
foram analisados através dos programas estatísticos NTSYS-pc (1992) e STATISTICA
(Statsoft Corp. 1997).
Para a avaliação de diferenças significativas na comparação dos parâmetros físicoquímicos (temperatura da água, profundidade, oxigênio dissolvido, salinidade e
transparência) e da densidade ictíica no espaço (pontos amostrais) e tempo
(épocas/meses) foi utilizada a análise uni/multivariada (ANOVA/MANOVA) e sempre
que a distribuição não foi normal (determinada pela prova de Kolmogorov-Smirnov),
foram aplicadas as transformações pertinentes (log ou log (x+1)). O teste a "posteriori"
foi selecionado com base no coeficiente de variação e número de tratamentos (Sokal &
Rohlf 1981, Clarke & Warwick 1994, Sampaio 1998, Underwood 1998). Para algumas
variáveis, nas quais não foi possível atender as suposições relativas à normalidade e
homocedasticidade (teste de Cochran), foi utilizada a prova não paramétrica de KruskalWallis (Siegel 1981, Clarke & Warwick 1994, Sampaio, 1998, Underwood 1998,
Barletta, 1999).
18
Para caracterizar a ictiofauna capturada e o seu comportamento frente aos
parâmetros observados, foram utilizadas as subdivisões propostas por Cariiker (in
Lauff, 1967) e Yánez & Nugent (1977) que propuseram as seguintes categorias:
a) estenohalinos dulcícolas: ovos, larvas, juvenis e adultos somente nos rios e
restritos a salinidades de 0‰.
b) oligohalinos visitantes ocasionais: reprodução e juvenis somente nos rios,
adultos nos rios e nos estuários, no último nunca presentes em salinidades superiores a
5‰.
c) eurihalinos verdadeiramente estuarinos-: ovos, larvas, juvenis e adultos somente
no estuário (nunca em salinidades inferiores a 5‰ e superiores a 25‰).
d) eurihalinos-marinhos :
?? com desenvolvimento ( ovos, larvas e juvenis) em salinidades entre 5 e 25‰,
isto é nos estuários, contudo adultos somente em salinidades superiores a 25‰ ,
?? ovos, larvas, juvenis e adultos marinhos, próximos dos estuários (ciclo de vida
costeiro e em salinidades superiores a 25‰), e com os adultos e juvenis entrando
ocasionalmente no estuário (até salinidades de 5‰).
f) estenohalinos marinhos: com reprodução e desenvolvimento oceânicos
(salinidades superiores a 30‰),somente adultos visitam ocasionalmente as regiões
costeiras (até 25‰).
A análise do grau de importância dos peixes para a economia de subsistência
regional foi estabelecido com base nas informações disponíveis na literatura e em 46
entrevistas (quando ocorreu a homogeneização da variância), realizadas, entre setembro
e outubro de 1996, com pescadores das regiões próximas dos pontos amostrados.
19
As entrevistas foram realizadas com o pescador cabeça da família, todos residentes
em moradias diferentes, e incluíram questões que abrangiam a caracterização: 1) das
unidades amostrais
(idade, estado civil, número da pessoas que coabitam com o
entrevistado, local de residência, ocupação principal - quando praticada acima de 30
horas/semana- e secundária - quando abaixo desse valor), 2) das atividades de pesca
(tipo, locais onde é executada, especificações da embarcação utilizada, número de
horas, tipos de petrechos e suas especificações) e 3) da abundância dos recursos ictiícos
explotados (freqüentes, pouco freqüentes ou ausentes), com base nos nomes vulgares
regionais de 53 espécies selecionadas, independentemente do petrecho utilizado na
captura e a partir do levantamento de Corrêa (1987), por sua importância na economia
regional (Anexo 2).
20
4. RESULTADOS
4.1. PARÂMETROS FÍSICO-QUÍMICOS E METEOROLÓGICOS
Na figura 4 e na tabela 1 são listados os dados mensais de pluviosidade no período
estudado. As
primeiras amostragens, dos meses de novembro, dezembro, janeiro e
fevereiro foram realizadas 48 horas após fortes precipitações (tab.1 e fig. 4). Durante as
coletas, as maiores precipitações ocorreram na segunda amostragem dos meses de
novembro (16 mm) e dezembro (27,6 mm) e na primeira do mês de janeiro (16,7 mm)
(tab.1 e fig. 4).
Na tabela 2 e na figura 5a encontram-se representadas as médias mensais de
temperatura do ar. A comparação das variações das médias mensais caracterizou duas
tendências: a primeira definindo um período no qual a elevação de temperatura ocorreu
com oscilações de pequena amplitude e, a segunda caracterizando um período de queda de
temperatura com oscilações de maior amplitude.
As médias (desvios e erros padrões das estimativas) anuais de temperatura do ar
concernentes aos diferentes pontos amostrais constam da tabela 3 e figura 5b. As
diferenças entre as médias não foram significativas ao nível de 95% de segurança.
Contudo, foi observado um ligeiro incremento nas médias de temperatura do ponto
amostral 1 para o 3 e redução na média de temperatura do ponto 4 (tab. 3 e fig.5b).
As profundidades, nos meses e pontos amostrais, oscilaram de acordo com a altura das
marés e variaram entre 1,52 (ponto 4, agosto/1986) e 7,92 m (ponto 1, janeiro e fevereiro/
1987)(tab. 4 e fig. 6a). Foram significativas (ao nível de segurança de 95%) as diferenças
entre as médias de profundidade dos pontos amostrais 1 a 4, respectivamente: 6,07 (±0,97),
2,75(±0,55), 3,11 (±0,60) e 5,92 (±1,36) m (tab. 5, fig. 6b). Portanto, os pontos 1 e 4
21
80
48h ant. coleta
coleta
70
60
Precipitação (mm)
50
40
30
20
1986
JUL2
JUL1
JUN2
MAI2
JUN1
MAI1
ABR2
ABR1
MAR2
MAR1
FEV2
FEV1
JAN2
JAN1
DEZ2
DEZ1
NOV2
OUT2
NOV1
OUT1
SET2
SET1
AGO2
0
AGO1
10
1987
Figura.4. Diagrama comparativo dos valores de pluviosidade (em mm), registrados no momento e nas
ultimas 48 horas de amostragem. Fonte: dados de campo e IAPAR.
b 30
a 36
±1,96*e
±1,00*e
Média
28
o
Temperatura do ar ( C)
o
Temperatura do ar ( C)
32
±1,96*e
±1,00*e
Média
28
24
26
24
22
16
20
AGO_1
AGO_2
SET_1
SET_2
OUT_1
OUT_2
NOV_1
NOV_2
DEZ_1
DEZ_2
JAN_1
JAN_2
FEV_1
FEV_2
MAR_1
MAR_2
ABR_1
ABR_2
MAI_1
MAI_2
JUN_1
JUN_2
JUL_1
JUL_2
20
1986
1
2
3
4
Pontos amostrais
1987
MESES
Figura. 5. Diagramas comparativos das médias de temperatura do ar: a) nos meses e b) pontos de
amostragem, observadas na Baía de Guaraqueçaba.
ANOVA F(3,91)=86,91 p=0,000
a 10
b
±s
±e
Média
7,5
6,5
Profundidade (m)
Profundidade (m)
8
8,5
Ponto 1
Ponto 2
Ponto 3
Ponto 4
6
4
6,07
5,93
5,5
4,5
3,5
3,11
2
2,75
0
AGO1
AGO2
SET1
SET2
OUT1
OUT2
NOV1
NOV2
DEZ1
DEZ2
JAN1
JAN2
FEV1
FEV2
MAR1
MAR2
ABR1
ABR2
MAI1
MAI2
JUN1
JUN2
JUL1
JUL2
2,5
1,5
1
2
3
4
Pontos amostrais
Figura. 6. Valores de profundidade (metros) dos arrastos nos: a) pontos e meses de amostragem e b) pontos
amostrais (resultado da comparação pela análise de variância).
22
apresentaram uma profundidade média significativamente maior que os pontos 2 e 3,
respectivamente 6,00 (±1,17) e 2,93 (±0,60) m (tab. 6).
Para a temperatura da água não foi observada forte estratificação vertical em nenhum
dos meses do ano e pontos amostrais (tab. 7 e 8 e fig. 7a e b). A análise de variância,
resultante das comparações das médias de temperatura da água entre os meses, pontos
amostrais, superfície/fundo, e da interação entre estas fontes de variação, é mostrada nas
tabelas 9 e 10. As diferenças entre
as médias de temperatura dos meses foram
significativas ao nível de 95% de segurança. As temperaturas dos meses de inverno foram
significativamente inferiores às do final da primavera, início de outono e verão (tab. 11).
Nas tabelas 12 e 13 as médias de salinidade da água, de superfície e fundo, nos meses
e pontos amostrais. A ANOVA da comparação das médias por mês, ponto amostral,
coluna d’água e da interação entre estas fontes de variação é mostrada na tabela 14. Foram
significativas as diferenças entre as médias dos meses, pontos amostrais, da coluna d’água
e da interação dos pontos amostrais/coluna d’água. As menores médias de salinidade
ocorreram nos meses de verão e outono, e as maiores nos meses de final de primavera,
inverno e final de outono, respectivamente épocas de maior e menor pluviosidade (tab.15,
fig. 7c). As menores médias de salinidade, independentemente dos meses e pontos
amostrais, foram observadas para a água de superfície (tab. 16).
Também foi observada a estratificação vertical e horizontal de salinidade entre e nos
pontos amostrais (tab.17 e 18). Houve uma redução significativa da média do ponto
amostral 1/superfície e fundo (furo de Guaraqueçaba) para o 3/superfície (no delta formado
pela embocadura dos rios Morato, Moratinho e Ipanema), sendo que o ponto 4/superfície e
fundo apresentou valores intermediários entre o 2/superfície e fundo e o 3/superfície (tab.
17, 18 e fig.7d).
23
TEMPERATURA DA ÁGUA
a
TEMPERATURA DA ÁGUA
b
ANOVA- Mês F(11,166)=68,69; p<0,00- Coluna d'água F(1,166)=1,02; p<0,31
ANOVA- Ponto F (3,182)=0,18; p<0,91- Coluna d'água F(1,182)=0,20; p<0,65
Pto x Coluna F(3,182)=0,01; p<0,20
Mês x Coluna F(11,166)=0,08; p<0,99
±s Superfície
±e Superfície
Média Superfície
32
34
±s Fundo
±e Fundo
Média Fundo
(ºC)
(ºC)
34
Temperatura da água
Temperatura da água
30
28
26
24
22
±s Superfície
±e Superfície
Média Superfície
32
±s Fundo
±e Fundo
Média Fundo
30
28
26
24
22
20
JUL
JUN
MAI
ABR
MAR
FEV
JAN
DEZ
NOV
OUT
AGO
SET
20
18
SALINIDADE
c
18
1
±s Superfície
±e Superfície
Média Superfície
Pto x Coluna F(3,183)=3,94; p<0,01
30
±s Fundo
±e Fundo
Média Fundo
22
18
14
10
JUL
JUN
MAI
ABR
MAR
FEV
JAN
DEZ
NOV
OUT
SET
AGO
OXIGÊNIO DISSOLVIDO
e
22
18
14
6
1
O2 (% saturação)
±s Fundo
±e Fundo
Média Fundo
±s Superfície
±e Superfície
Média Superfície
140
120
100
80
120
100
80
0
JUL
JUN
MAI
ABR
MAR
FEV
JAN
DEZ
NOV
OUT
20
SET
40
20
AGO
60
1
2
3
4
TRANSPARÊNCIA DA ÁGUA
h
ANOVA -P o n t o sF(3,86)=1,80; p<0,1526
ANOVA Mês
F(11,78)=6,07; p<0,00
1,8
1,8
±s
±e
Média
1,6
±s
±e
Média
1,6
1,4
Transparência (m)
1,4
1,2
1,0
0,8
0,6
1,2
1,0
0,8
0,6
0,4
0,4
Meses
JUL
JUN
MAI
ABR
MAR
FEV
JAN
DEZ
NOV
OUT
SET
0,2
AGO
0,2
±s Fundo
±e Fundo
Média Fundo
140
40
TRANSPARÊNCIA DA ÁGUA
±s Superfície
±e Superfície
Média Superfície
160
60
g
4
Pto x Coluna F(3,163)=0,20, p<0,89
180
160
3
ANOVA- Ponto F(3,3)=2,24; p<0,26 - Coluna d'água F(1,3)=16,14; p<0,03
F(11,147)=0,91; p<0,53
180
0
2
OXIGÊNIO DISSOLVIDO
f
ANOVA- Mês F(11,147)=7,94; p<0,00 - Coluna d'água F(1,147)=2,80; p<0,10
Mês x Prof.
Transparência (m)
±s Fundo
±e Fundo
Média Fundo
10
6
O2 (% saturação)
±s Superfície
±e Superfície
Média Superfície
26
Salinidade (‰)
Salinidade (‰)
26
4
ANOVA- Ponto F(3,183)=13,37; p<0,00- Coluna d'águaF(1,183)=21,14; p<0,00
Mês x Coluna F(11,167)=0,44; p<0,94
30
3
SALINIDADE
d
ANOVA - Mês F(11,167)=12,64; p<0,00- Coluna d'água F(1,167)=27,16; p<0,00
2
0,0
1
2
3
4
Pontos Amostrais
Figura 7. Diagramas comparativos das variações observadas nos parâmetros físico-químicos da água, de
superfície e fundo, da Baía de Guaraqueçaba, nos diferentes meses do anos de 1986 e 1987 e pontos
amostrais.
24
Na tabela 19 são listadas as médias do teor de oxigênio dissolvido na água (de
superfície e de profundidade), expressas em porcentagem de saturação. Na tabela 20 a
ANOVA da comparação entre as médias de oxigênio dissolvido dos meses e da coluna
d’água. As diferenças entre as médias dos meses foram significativas ao nível de segurança
de 95%. De modo geral apresentou tendência a elevação significativa dos meses de
inverno (junho, julho e agosto) para os de primavera (setembro, outubro, novembro e
dezembro)(tab.21 e fig.7e).
Na tabela 22 as médias de teor de oxigênio dissolvido na água de superfície e fundo
por pontos amostrais. Como constatado para a salinidade, as diferenças entre os teores
médios de oxigênio, para a água de superfície e fundo, foram significativas ao nível de
95% de segurança (tab. 23). Foi constatada estratificação vertical, com maiores valores
para as águas superficiais dos pontos amostrais e notável redução na água de superfície do
ponto 2(tab. 24, 25 e fig. 7f).
As médias mensais de transparência da água situaram-se entre 0,67 (janeiro/1987) e
1,32 m (agosto/1986) (tab.26 e fig.7g). As diferenças entre as médias mensais foram
significativas ao nível de 95% de segurança (tab.27). A tendência foi similar àquela
observada para a salinidade e oxigênio dissolvido, isto é, foram significativamente maiores
nos períodos de menor precipitação e, ainda, com redução (embora não significativa) do
ponto amostral mais externo para os mais internos (2 e 3) (tab. 28, 29, 30 e fig. 7g). Como
observado para a salinidade, o ponto 4 apresentou a média da transparência da água
intermediária às dos pontos 1 e 2 (tab. 29, fig. 7h).
25
4.2. ICTIOFAUNA
4.2.1. Composição, variabilidade temporal e distribuição espacial.
Nas tabelas 31 e 32 encontra-se a composição familiar e específica com as
respectivas freqüências absolutas (em número de exemplares e porcentagem) e pesos totais
(em g e porcentagem) de captura, por pontos amostrais. Foi examinado um total de 33868
peixes, com o peso total de 772 777,5 g. Foram capturadas 19 famílias, 43 gêneros e 52
espécies de Actinopterygii.
Na figura 8 (a, b, c, d) os números de exemplares capturados (em freqüência
absoluta) e os pesos totais de captura (g), por épocas do ano e por pontos amostrais.
Na tabela 33 e 34 o resultado da prova de Kruskal-Wallis ANOVA, da comparação
das freqüências de captura por épocas do ano e pontos amostrais. As épocas e pontos
diferiram significativamente entre si quanto às freqüências absolutas de captura.
Comparativamente, um número maior de exemplares foi capturado no outono (15 641,
fig.8a) e no ponto amostral 1 (12 670, fig.8b), correspondendo a 46,18 e 37,41 % do total
obtido, respectivamente, nas estações do ano e pontos amostrais. Uma menor freqüência de
captura foi obtida no inverno (1269 ou 3,75%), e no ponto amostral 3 (4 834 ou
14,27%)(fig.8a e b, 9 e 10a).
Na tabela 33 e 34 o resultado, da prova de Kruskal-Wallis ANOVA, da comparação
das freqüências absolutas e pesos totais de captura por épocas do ano e pontos amostrais.
As diferenças observadas entre os pesos de captura por épocas e pontos foram
significativas ao nível de 95% de segurança. Destacaram-se os altos valores de peso de
captura na primavera (236 093,60 g), no ponto 1 (343 318,00 g) e baixos valores das
capturas de inverno (36 280,70 g) e no ponto 3 (110 515,00 g) (fig.8c e d, 9 e 10b).
As principais famílias capturadas, expressas em ordem decrescente de freqüência
absoluta (número de exemplares) e relativa (%) de captura, foram: Ariidae, Sciaenidae,
26
b
16000
n=33868
14000
12000
12000
10000
Número de Exemplares
14000
Primavera
Verão
8000
6000
4000
2000
10000
8000
JUL
MAI
JUN
ABR
FEV
MAR
JAN
DEZ
NOV
OUT
7953
6000
4834
4000
d
1
2
3
4
360000
Primavera
n=33868
343318,00
300000
Peso Total da Captura (g)
300000
Outono
240000
Verão
180000
120000
60000
240000
180000
179587,06
139356,45
120000
110516,00
60000
Inverno
1
AGO
JUL
JUN
MAI
ABR
FEV
MAR
JAN
DEZ
NOV
SET
0
OUT
Peso Total da Captura (g)
8411
0
360000
0
12670
2000
Inverno
SET
0
c
16000
n=33868
AGO
Número de Exemplares
(Freqüência absoluta)
Outono
(freqüência absoluta)
a
2
3
4
Ponto amostral
Mês/Épocas do ano
Figura 8. Diagramas comparativos das capturas realizadas entre agosto de 1986 e julho de 1987, na Baía de
Guaraqueçaba (Pr, BR), mostrando as freqüências absolutas e pesos totais nos meses e épocas do ano(a e c)
e anuais por pontos amostrais (b e d).
Kruskal-Wallis ANOVA H (3,96)=32,05; p=0,00
a
Kruskal-Wallis ANOVA H ( 3, 96) = 34,84; p =0,00
b
1000
±1.96*e
±1.00*e
Média
600
400
200
Peso Total da Captura (g)
(freqüência absoluta)
Número de Exemplares
800
±1.96*e.
±1.00*e
16000
Média
12000
8000
4000
0
0
Primavera
Verão
Outono
Épocas do Ano
Inverno
Primavera
Verão
Outono
Inverno
Épocas do Ano
Figura 9. Resultado da análise de variância de Kruskal-Wallis originada da comparação: a) do número de
exemplares capturados (freqüência absoluta) e b) do peso total das capturas (em gramas), nas diferentes
épocas do ano, na Baía de Guaraqueçaba, Pr, BR.
27
Paralichthyidae e Tetraodontidae com, respectivamente 24 940 (73,64%), 6 692 (19,76%),
842 (2,49%) e 560 (1,65%) (tab.31 e fig. 11). De modo geral as duas primeiras famílias
foram mais abundantes no ponto amostral 1 e 4, diminuindo consideravelmente no sentido
dos pontos mais internos (2 e 3). Paralichthyidae foi mais abundante no ponto 1,
decrescendo em número no sentido do ponto 4, e Tetraodontidae predominou nos pontos 2
e 4 (tab. 31 e fig.11).
As espécies mais abundantes, também expressas em ordem decrescente de
freqüência absoluta
(número de exemplares) e relativa (%) de ocorrência, foram:
Cathorops spixii, Stellifer rastrifer, Genidens genidens e Sciadeichthys luniscutis
respectivamente com 18 839 (55,62%), 5 073 (14,98%), 3 980 (11,75%) e 1 440 (4,25%).
As demais espécies foram ocasionais e ocorreram em freqüências absolutas e relativas
inferiores, respectivamente, a 700 exemplares ou 2% do total capturado (tab.31 e fig. 12).
Os maiores pesos (em grama e porcentagem) de captura foram obtidos para as
famílias Ariidae, Sciaenidae, Tetraodontidae e Paralichthyidae, com respectivamente 590
905,20 (76,46%); 95 180,50 (12,32%); 45 983,20 (5,95%) e 11 757,90 (1,52%) g (tab.32).
Para as espécies, foram observados em: C. spixii, G. genidens, S. rastrifer e S. testudineus
com respectivamente 449 273,20 (58,14%), 77 558,80 (10,04%), 58 925,10 (7,63%) e 45
781,00 (5,95%) g (tab.32).
Na figura 13 encontra-se a plotagem simultânea das espécies, com freqüências de
captura próximas ou superiores a 1% da captura total (tab.31), como descritores, e dos
pontos amostrais como objetos, nos planos fatoriais da análise dos componentes principais
(ACP). A análise restringiu-se aos 2 primeiros componentes
que explicaram
respectivamente 51,69 e 33,05% da variabilidade total dos dados. Os dois primeiros
componentes associados explicaram 84,74% da variância (tab.35).
28
Kruskal-Wallis ANOVA H(3,96)=8,59 ; p =0,03.
1600
±1.96*e
Peso Total da Captura (g)
1000
800
600
400
±1.00*e
20000
Média
1200
(freqúência absoluta)
±1.96*e
±1.00*e
1400
Número de Exemplares
Kruskal-Wallis ANOVA H (3,96) = 11,33 ; p =0,01
24000
Média
16000
12000
8000
4000
200
0
0
1
2
3
1
4
2
3
4
Ponto Amostral
Ponto amostral
Figura 10. Resultado da análise de variância de Kruskal-Wallis: a) do número de exemplares capturados
(freqüência absoluta) e b) do peso total das capturas (em gramas), nos diferentes pontos amostrais, na Baía de
Guaraqueçaba, Pr, BR.
PONTOS
1
2
3
4
Famílias
Ariidae
Sciaenidae
0
2000
4000
6000
8000
10000
Famílias
Freqüência absoluta
Achiridae
Carangidae
Cynoglossidae
Clupeidae
Diodontidae
Engraulidae
Ephippidae
Gerreidae
Gobiidae
Paralichthyidae
Pomadasyidae
Serranidae
Stromateidae
Tetraodontidae
Triglidae
Trichiuridae
PONTO
1
2
3
4
0
100
200
300
400
500
Freqüência absoluta
Figura 11. Distribuição das freqüências absoluta de captura das famílias de peixes nos diferentes pontos
amostrais da Baía de Guaraqueçaba, Pr, Br.
29
Família Sciaenidae
Família Ariidae
B. ronchus
PONTOS
1
2
3
4
B. bagre
C. spixii
PONTOS
1
2
3
4
C. gracillicirrhus
C. acoupa
C. leiarchus
Espécies
Espécies
C. microlepidotus
G. genidens
C. virescens
I. parvipinnis
M. americanus
M. ancylodon
N. barba
M. furnieri
P. brasiliensis
S. luniscutis
S. brasiliens
S. rastrifer
0
2000
4000
6000
8000
10000
0
Freqüência absoluta
400
800
1200
1600
2000
Freqüência absoluta
Espécies
Outras Famílias
A. lineatus
A. januaria
C. latus
C. edentulus
C. faber
C. spinosus
C. crysurus
C. arenaceus
C. spilopterus
D. rhombeus
D. radiale
E. argenteus
E. crossotus
E. gula
E. melanopterus
G. luteus
G. oceanicus
H. clupeola
L. grossidens
L. laevigatus
O. saliens
O. saurus
P. brasiliensis
P. harroweri
P. paru
P. punctatus
P. tropicus
S. foetens
S. plagusia
S. testudineus
S. vomer
T. lepturus
T. microphthalmus
T. paulistanus
Pontos
1
2
3
4
0
50
100
150
200
250
300
Figura 12. Distribuição das freqüências absolutas de captura das espécies de peixes nos diferentes pontos
amostrais da Baía de Guaraqueçaba.
30
Contribuíram para a formação do componente 1, coordenada positiva, a freqüência
de ocorrência das espécies Citharichthys arenaceus, Sciadeichthys luniscutis, Symphurus
plagusia, Stellifer rastrifer e Cathorops spixii. Para a formação da coordenada negativa,
contribuíram as freqüências de ocorrência das espécies Genidens genidens e
Micropogonias furnieri (fig. 13). Para a formação do componente 2, coordenada positiva,
contribuíram principalmente Sphoeroides testudineus, Netuma barba e, em menor grau,
Micropogonias furnieri. Na coordenada negativa contribuíram principalmente Etropus
crossotus e Isopisthus parvipinnis (fig.13).
Apresentaram correlação positiva com o componente 1, respectivamente nas
coordenadas positiva e negativa, os pontos amostrais 1-2 e 3-4. Por sua vez, apresentaram
correlação positiva com o componente 2, respectivamente com os eixos positivos e
negativos, os pontos amostrais 2 e 1 (fig.13).
Estes resultados mostraram que as amostras do ponto 1 estavam compostas
principalmente pelas espécies C. spixii, S. rastrifer, E. crossotus e I. parvipinnis, com
redução numérica no sentido dos pontos amostrais mais internos (2, 3 e 4). Ainda C. spixii
e S. rastrifer ocorreram em altas freqüências nos pontos 1, 2 e 4, sofrendo redução
significativa apenas no ponto amostral 3 mais interno (fig.13).
No ponto amostral 2 predominaram as espécies, por ordem de abundância, S.
testudineus, N. barba, C. arenaceus, S. luniscutis e S. plagusia.
O ponto 3 foi distinto dos demais pela dominância numérica de G. genidens,
enquanto a composição ictiofaunística dos pontos 1, 2 e 4 foram muito similares. Contudo,
no último ponto ocorreu uma redução numérica significativa para a maioria das espécies
presentes no ponto 1, 2 e 3. É importante destacar que M. furnieri apresentou freqüências
de ocorrência aproximadamente iguais tanto no ponto amostral 2 como no 3 (fig.13 e tab.
31).
31
3
Ponto 2
Sphoeroides testudineus
Componente 2
2
Netuma barba
Citharichthys
arenaceus
Sciadeichthys
luniscutis
Symphurus
plagusia
Micropogonias
furnieri
1
0
Ponto
4
Genidens
genidens
Ponto
3
Stellifer
rastrifer
Cathorops
spixii
-1
Etropus
crossotus
-2
-3,1
-2,1
-1,1
-0,1
0,9
Isopisthus
Ponto
parvipinnis
1
1,9
2,9
Componente 1
Figura 13. Diagrama da distribuição dos pontos amostrais (objetos) e das freqüências das espécies
abundantes (descritores) no plano bidimensional dos componentes principais.
Bray-Curtis (similaridade %)
70
80
12
4
3
90
Stress=0,00
100
Ponto 4
Ponto 2
Ponto 1
Ponto 3
Figura 14. a) Dendrograma dos pontos amostrais, resultante da união "UPGA", sobre uma matriz de
similaridade Bray-Curtis, originada de uma matriz de abundância transformada (?? ) e b) ordenação dos
pontos amostrais no plano bidimensional pelo método "MDS" com os grupos do dendrograma sobrepostos
nos níveis de 80 (linha contínua) e 82 % (linha tracejada) de similaridade (stress=0,00).
32
O dendrograma dos pontos amostrais, obtido à partir de uma matriz de similaridade
Bray-Curtis, com base na abundância transformada(DRQ) de todas as espécies,corroborou
estes resultados (fig.14a). Para melhor visualização, na figura 14b os pontos amostrais
foram ordenados no plano bidimensional da "MDS" com os grupos do dendrograma
sobrepostos nos níveis de similaridade de 80 e 82%.
Na figura 15 encontra-se, nos planos fatoriais da análise dos componentes
principais (ACP), a plotagem simultânea das freqüências numéricas das espécies mais
abundantes (capturas próximas ou superiores a 1% da ocorrência total) como objetos e os
meses de amostragem como descritores.
A análise restringiu-se aos dois primeiros
componentes que explicaram isolada e respectivamente 75,63 e 10,90% e associados
86,53% da variância total dos dados (tab.36). Contribuíram para a formação da coordenada
positiva do componente 1 a maioria dos meses amostrados e da negativa apenas o mês de
julho. Contribuíram para a formação da coordenada positiva do componente 2 os meses de
julho, maio, junho, fevereiro e março, e da coordenada negativa os meses de setembro e
agosto (fig.15). Associaram-se positivamente à maioria dos meses amostrados, e portanto
ao componente 1, as espécies C. spixii, S. rastrifer e G. genidens e negativamente I.
parvipinnis, E. crossotus, S. luniscutis, C. arenaceus, N. barba, M. furnieri e S.
testudineus. As últimas apresentaram correlação positiva com o mês de julho(fig.15).
Estes resultados mostraram a dominância numérica de C. spixii, S. rastrifer e G.
genidens na maioria dos meses amostrados, com a redução do primeiro apenas nos meses
de agosto, setembro e julho (fig.15). Nos últimos meses passaram a dominar S. rastrifer
(agosto), G. genidens (setembro) e E. crossotus, S. luniscutis, C. arenaceus, N. barba, M.
furnieri e S. testudineus (julho).
Na figura 16 os resultados da plotagem simultânea dos pesos totais de captura das
espécies cujas freqüências e/ou pesos foram superiores a 1% (tab. 31 e 32), como objetos, e
DRQ = dupla raíz quadrada
33
Componente 2
1.1
0.1
-0.9
1
23
4 julho
5
6
Isopisthus
parvipinnis
maio
junho
fevereiro
março
abril
outubro
novembro
janeiro
dezembro
Cathorops
spixii
Inverno
Outono
Verão
Primavera
Stellifer
rastrifer
agosto
-1.9
Inverno
1-Etropus crossotus
2-Sciadeichthys luniscutis
3-Citharichthys arenaceus
4-Netuma barba
5-Micropogonias furnieri
6-Sphoeroides testudineus
Genidens
genidens
setembro
-2.9
-2
0
4
6
Componente 1
8
10
Figura 15. Diagrama da distribuição das freqüências das espécies abundantes (objetos) e dos meses
(descritores), sobrepostos no plano bidimensional dos componentes principais.
1.9
56
0.9
4 3
Cathorops
spixii
2
Maio Junho
Março Fevereiro
Janeiro Abril
Outubro Novembro
Componente 2
1
Inverno
Outono
Verão
Primavera
-0.1 Etropus
crossotus
Dezembro
Sphoeroides
-1.1 testudineus
Agosto
Inverno
-2.1
Setembro
Julho
Genidens
genidens
-3.1
-2
0
2
1-Stellifer rastrifer
2-Sciadeichthyes luniscutis
3-Isopisthus parvipinnis
4-Citharichthys arenaceus
5-Micropogonias furnieri
6-Netuma barba
4
6
8
10
Componente 1
Figura 16. Diagrama da distribuição do peso de captura das espécies abundantes (objetos) e dos meses
(descritores), sobrepostos no plano bidimensional dos componentes principais.
34
os meses de amostragem como descritores, nos planos fatoriais da ACP. A análise
restringiu-se aos dois primeiros componentes que explicaram respectivamente 78,72 e
14,35% da variância total dos dados. Os dois primeiros componentes associados
explicaram 93,07% da variabilidade (tab.37). Os meses de maio, junho, março, fevereiro,
janeiro, abril, outubro, novembro, dezembro e agosto contribuíram, em maior grau, para a
formação da coordenada positiva do componente 1 (fig.16). Setembro e julho
contribuíram, também em maior grau,
para a formação da coordenada negativa do
componente 2. Correlacionaram-se positivamente ao componente 1 o peso de captura de
C. spixii e negativamente o peso obtido para as demais espécies com a exceção de G.
genidens. O peso de captura da última apresentou correlação positiva com os meses de
setembro e julho (fig.16).
Os resultados mostraram a dominância em peso de C. spixii na maioria dos meses
amostrados com redução apenas nos meses de setembro (quando G. genidens passou a
dominar) e julho (com a codominância de G. genidens e das demais espécies).
Na figura 17 os resultados da ordenação, no plano bidimensional da ACP, dos
pesos de captura obtidos nos diferentes pontos amostrais como descritores e das espécies
(com freqüências e/ou pesos superiores a 1%) como objetos. A análise restringiu-se aos
dois primeiros componentes que explicaram isolada e respectivamente 74,56 e 22,82% da
variabilidade total dos dados. Associados explicaram 97,38% da variância (tab. 38).
Contribuíram positivamente para formação do eixo 1 os ponto 1, 2 e 4. Para a
formação do eixo 2 contribuiu positivamente o ponto 3 (fig.17). Correlacionaram-se
positiva e negativamente ao eixo 1 respectivamente os pesos de captura de C. spixii e das
demais espécies (exceto de G. genidens). Ao eixo 2 correlacionou-se positivamente o peso
de captura de G. genidens e negativamente os pesos das demais espécies (incluindo o de
C. spixii). Estes resultados demonstraram o predomínio em peso de C. spixii nas capturas
35
dos pontos 1, 2 e 4 e de G. genidens nas capturas do ponto 3. As demais espécies
apresentaram baixos pesos de captura em todos os pontos amostrais (fig.17).
4
3.4
Genidens Ponto 3
genidens
Componente 2
2.4
1.4
1-Stellifer rastrifer
2-Sphoeroides testudineus
3-Sciadeichthys luniscutis
4-Netuma barba
5-Citharichtys arenaceus
6-Etropus crossotus
7-Isopisthus parvipinnis
8-Micropogonias furnieri
9-Chaetodipterus faber
0.4
9
1
7 8
-0.6 6 5 4 3
-1
Ponto 2 Cathorops
Ponto 1
spixii
Ponto 4
2
1
3
5
7
Componente 1
Figura 17. Diagrama da distribuição dos pontos amostrais (objetos) e dos pesos de captura das espécies
abundantes (descritores), sobrepostos no plano bidimensional dos componentes principais
36
4.2.2. Diversidade, equitatividade e riqueza
Nas tabelas 39 e 40 encontram-se discriminados os valores dos índices de diversidade
(N1) e equitatividade (E5) obtidos, respectivamente, para os meses e pontos amostrais.
Na figura 18a é mostrada a distribuição dos valores de N1 nos meses de amostragem.
O número de espécies abundantes foi maior nas amostras dos meses de agosto (9,54) e julho
(8,25), e o menor dos meses de maio (2,97) e junho (3,08). Entre os demais destacaram-se
setembro, dezembro e março quando o número de espécies situou-se próximo de 5. De modo
geral a tendência foi o decréscimo do número de espécies abundantes entre agosto de 1986 e
junho de 1987, com novo aumento no mês de julho do último ano.
Índice de Diversidade de Hill (N1)
a
Eqüitatividade de Hill (E5)
b
jul
jul
jun
jun
mai
mai
abr
abr
mar
mar
fev
fev
jan
jan
dez
dez
nov
nov
out
out
set
set
ago
ago
0
2
4
6
8
10
Índice de Diversidade de Hill (N1)
c
0,3
0,4
Ponto 4
Ponto 3
Ponto 3
Ponto 2
Ponto 2
Ponto 1
Ponto 1
2
4
6
8
10
0,6
0,7
Eqüitatividade de Hill (E5)
d
Ponto 4
0
0,5
0,3
0,4
0,5
0,6
0,7
Figura 18. Distribuição dos valores dos índices de diversidade (N1) e eqüitatividade de Hill (E5),
respectivamente nos meses (a e b) e pontos amostrais (c e d) obtidos na Baía de Guaraqueçaba, Pr.
37
Com relação a equitatividade, o valor de E5 é inversamente proporcional à dominância
numérica de uma das espécies. De modo geral a tendência foi o decréscimo do valor de E5, e
portanto do aumento da dominância em número por uma das espécies, entre os meses de
agosto de 1986 a maio de 1987, com notável exceção para os meses de dezembro e janeiro.
Em junho e julho o valor voltou a aumentar(fig.18b).
Estes resultados refletiram a dominância de C. spixii, portanto a menor diversidade e
equitatividade, nos meses de maio e junho (fig.15 e 18a,b). Do mesmo modo refletiram a
redução numérica desta espécie, com o aumento dos índices de diversidade e equitatividade
nos meses janeiro, dezembro, agosto, setembro e julho, quando também passaram a
predominar S. rastrifer, G. genidens, I. parvipinnis, E. crossotus, S. luniscutis, C. arenaceus,
N. barba, M. furnieri e S. testudineus (fig. 15 e 18a,b).
Considerando-se apenas os pontos amostrais, os valores dos índices de diversidade e
equitatividade aumentaram do ponto amostral 1(N1 = 3,80 e E5 = 0,40), na porção mais
externa da Baía de Guaraqueçaba,
para o 2 (N1 = 5,91 e E5= 0,48) e mantendo-se
aproximadamente constante no ponto 3 mais interno (N1 = 5,70 e E5 = 0,46) (fig.18c e 18d).
O ponto 4, na região mediana oposta ao ponto 2, apresentou o valor de N1 (3,40) inferior ao
do ponto amostral 1 (N1=3,80) e de E5 (0,43) intermediário aos dos pontos 1 (E5=0,40) e 3
(E5=0,46). Estes resultados mostraram tendência similar àquela observada para a salinidade e
transparência da água (fig. 7d e h, 18c e d e tab. 40).
Os resultados também refletiram a distribuição espacial observada na figura 13, onde
pode ser observado o domínio numérico da composição ictiofaunística no ponto 1 por quatro
espécies (E. crossotus, I. parvipinnis, C. spixii e S. rastrifer). A diversidade e equitatividade
aumentaram no ponto 2, onde passaram a ocorrer, além de C. spixii e S. rastrifer, também S.
plagusia, S. luniscutis, C. arenaceus, N. barba e S. testudineus. As duas últimas ocorreram
38
em freqüências similares, e por isso foram detectadas como únicas pelos índices utilizados. No
ponto 3, a diversidade e equitatividade foi similar ao do ponto 2. Contudo, apenas no ponto 3
ocorreu a dominância numérica de G. genidens. O ponto 4 mostrou composição ictiofaunística
similar aos pontos 1 e 3, diferindo dos últimos apenas na redução numérica de exemplares da
maioria das espécies (fig. 13 e 14, tab. 31).
Na tabela 41 encontram-se os valores dos índices de diversidade (N1) e equitatividade
(E5) de Hill, obtidos da relação entre os meses e pontos amostrais.
A figura 19 representa a plotagem simultânea dos meses de amostragem (como
objetos) e os valores dos índices de diversidade (N1) e equitatividade (E5) dos pontos
amostrais (como descritores) nos planos fatoriais resultantes da ACP. A análise restringiu-se
aos dois primeiros componentes que explicaram, respectivamente 34,24 e 31,35 % da
variância total dos dados. Juntos, os dois primeiros componentes, explicaram 65,59 % da
variabilidade (tab. 42). Contribuíram em maior grau para a formação do componente 1,
coordenada positiva, os valores de diversidade e equitatividade dos pontos amostrais 4, 2 e 1 e,
na coordenada negativa, os do ponto 3. Para a formação do componente 2 contribuíram,
também em maior grau, os valores de diversidade dos pontos amostrais 1 e 4 na coordenada
positiva, e negativamente, os obtidos para o ponto amostral 2 (fig.19).
Correlacionaram-se positivamente com a diversidade e eqüitatividade dos pontos 1 e 4,
os meses de agosto (N1/1=5,13 e E5/1=0,59; N1/4=6,57 e E5/4=0,56), julho (N1/1=6,36 e
E5/1=0,53; N1/4=2,67 e E5/4=0,76) e, em menor grau, dezembro (N1/1=3,03 e E5/1=0,50;
N1/4=4,32 e E5/4=0,67) (tab.41). O mês de julho apresentou correlação negativa apenas com
a eqüitatividade e diversidade do ponto 2. Correlacionaram-se positivamente com o último, e
negativamente com os pontos 1 e 4, os meses de junho (N1/2=5,80 e E5/2=0,72) e novembro
(N1/2=7,36 e E5/2=0,93). O mês de setembro apresentou correlação, positiva com os pontos 1
39
3.2
2.2
1.2
0.2
-0.8
Janeiro
E5-Pto3
N1-Pto3 Abril
Julho E5-Pto4
E5-Pto1
N1-Pto1
N1-Pto4
Agosto
Dezembro
Fevereiro
Outubro
Maio
Março
Setembro
E5-Pto2
N1-Pto2
Novembro
-1.8
Junho
-2.8
-2.4
-0.4
1.6
3.6
5.6
Figura 19. Distribuição dos valores dos índices de diversidade (N1) e eqüitatividade (E5) de Hill (como
descritores) e meses amostrais (como objetos), sobrepostos no plano bidimensional dos componentes principais.
(N1=4,29 e E5=0,53), 4 (N1=4,50 e E5=0,56) e principalmente 2 (N1=9,34 e E5=0,82) e,
negativa com o ponto 3(N1=3,17 e E5=0,54) (fig.19 e tab.41).
Os índices analisados correlacionaram-se positivamente com ponto 3, e negativamente
com os pontos 1, 4 e 2, nos meses de janeiro (N1/3=5,89 e E5/3=0,74) e abril de 1987
(N1/3=6,09 e E5/3=0,53). Os meses de outubro, fevereiro, março e maio, apresentaram fraca
correlação negativa com relação aos valores de diversidade de todos os pontos amostrais
(fig.19, tab. 41).
É importante destacar que os valores de diversidade e equitatividade dos pontos 1 e 4
apresentaram correlações positivas e negativas similares ao longo de todo o período analisado,
enquanto que o ponto amostral 2 mostrou-se inversamente correlato com os primeiros apenas
nos meses de novembro e junho (fig.19).
40
Estes resultados quando comparados com os observados nas figuras 12, 13 e 15,
mostraram que as espécies C. spixii e S. rastrifer foram responsáveis pela reunião dos índices
de diversidade e eqüitatividade dos pontos 1, 2 e 4, uma vez que predominaram (em
freqüência) nos meses de dezembro, agosto e setembro (com a maior eqüitatividade para o
último mês no ponto 2). No mês de julho, como já discutido anteriormente, a dominância
numérica destas espécies diminuiu enquanto E. crossotus, I. parvipinnis e G. genidens também
passaram a dominar nos pontos 1, 4 e 3, elevando os valores dos índices de diversidade e
eqüitatividade destes pontos amostrais (fig. 12, 13 e 15). Em outras palavras os resultados
mostraram que a composição ictiofaunística, foi semelhante nos pontos e meses amostrados.
Contudo, a diversidade e eqüitatividade nos pontos amostrais foram principalmente afetadas
pelas variações na abundância de C. spixii e S. rastrifer.
Na figura 20 foram plotados simultaneamente os valores dos parâmetros
meteorológicos (precipitação), hidrológicos (temperatura, salinidade, oxigênio dissolvido, pH,
profundidade dos arrastos, e transparência) e dos índices de diversidade (N1) e eqüitatividade
(E5) como descritores e os meses de amostragem (como objetos) nos planos fatoriais da ACP.
Os dois primeiros componenentes explicaram respectivamente 29,70 e 18,54% da variância
total dos dados (tab.43).
Contribuíram, em maior grau, para a formação do componente 1 (coordenada positiva),
a transparência da água, a salinidade da água (superficial e de profundidade) e, em menor
grau, os índices de diversidade (N1) e de
equitatividade (E5). Os demais parâmetros
meteorológicos (precipitação) e hidrológicos (temperatura, oxigênio dissolvido, pH e
profundidade) também contribuíram para a formação do componente 1, porém na coordenada
negativa.
41
4,1
Set
pH/S
pH/P
Nov
Componente 2
2,1
Out
Jan
0,1
-1,9
O2/P
T/S
T/F
Prec./48h
Jan
Fev
Fev
VERÃO
Jan
Fev
Dez
Out
Mar
Nov
N1
E5
Abr
Set
Set
Mar
Abr
Dez
O2/S Mai
Set
Out
Abr
Dez Fev
Out
Mar
Nov Mai
Jan
Ago
Sal./P
Sal./S
Ago
Ago
Jun
Transp.
Mai
Abr
Jul
Jul
Prof.
Jul
Jun
Jun
Mai
Jul
Ago
Jun
-3,9
-4,1
-2,1
-0,1
1,9
3,9
5,9
Componente 1
Figura 20. Distribuição dos valores dos índices de diversidade (N1) e eqüitatividade de Hill (E5), juntamente
com os parâmetros meteorológicos e hidrológicos (como descritores) e dos meses e pontos amostrais (como
objetos abreviados respectivamente por três letras e algarismos arábicos nos círculos), no plano bidimensional
dos componentes principais. Onde O2 = oxigênio dissolvido, pH = potencial hidrogeniônico, T = temperatura,
Prec./48h = precipitação nas 48 horas anteriores às coletas, Prec./Col.= precipitação no momento da coleta,,
Prof. = profundidade, Sal. = salinidade, S = superfície e F = fundo.
Para a formação do componente 2, coordenada positiva, contribuíram principalmente
os índices de diversidade (N1) e de equitatividade (E5) e, em menor grau, os parâmetros
hidrológicos: pH (superficial e de profundidade) e a salinidade (superficial e de profundidade).
Na coordenada negativa, contribuíram a profundidade dos arrastos e o oxigênio dissolvido da
água superficial.
Correlacionaram-se positivamente:
a) à transparência da água principalmente os meses de inverno (junho, julho e agosto)
e primavera (setembro) nos pontos amostrais 1 e 2.
b) à salinidade (superficial e de profundidade) os meses de inverno (agosto) e
primavera (setembro) nos pontos amostrais 1, 2 e 4.
42
c) aos índices de diversidade e equitatividade os meses de outono (março) e
primavera (setembro, outubro e novembro) apenas no ponto amostral 2,
d) ao pH (superficial e de profundidade) os meses de primavera (outubro e novembro)
e de verão (fevereiro) principalmente no ponto 2,
e) ao oxigênio dissolvido os meses de primavera (outubro) e outono (março) no ponto
3 e verão (dezembro) no ponto 2,
f) à temperatura (superficial e de profundidade), precipitação nas últimas 48 horas e
no momento da coleta os meses de verão (janeiro e fevereiro) nos quatro pontos
amostrais,
g) ao oxigênio dissolvido (na água superficial) os meses de verão (dezembro e
janeiro) e outono (maio) no ponto 4,
h) à profundidade os meses de inverno (junho e julho) e de outono (maio) nos pontos
3 e 4.
A distribuição observada, quando comparada com as obtidas nas figuras 13, 15 e 19,
mostrou que:
a) nos meses de agosto e setembro (fig. 19 e 20) as altas diversidades dos pontos 1 e, em
menor grau,
4 (fig. 19), estavam associadas às espécies S. rastrifer, C. spixii, I.
parvipinnis, E. crossotus e G. genidens (fig. 13) e a alta equitatividade destes meses no
ponto 2 (fig. 19), às espécies S. testudineus, N. barba, C. arenaceus, S. luniscutis, S.
plagusia, S. rastrifer, C. spixii e M. furnieri (fig. 13). Coincidentemente, nos meses em
questão, foram observados os maiores valores de salinidade na água de superfície e de
fundo (agosto nos pontos 1 e 2 e setembro nos ponto 2 e 4) e os menores para a temperatura
da água (superficial e de profundidade), pluviosidade e os teores de oxigênio dissolvido na
água de superfície (fig.20),
43
b) a alta diversidade no ponto 1 e altas eqüitatividades nos pontos 1, 3 e 4 estavam associadas
às espécies anteriormente citadas (item "a", fig. 13) e aos meses de julho, janeiro e, em
menor intensidade, dezembro e abril (fig. 19), quando foram observadas as maiores
profundidades (julho nos pontos 3 e 4), transparências da água (julho no ponto 1),
precipitações e teores de oxigênio dissolvido na superfície (janeiro e dezembro nos pontos
1, 3 e 4) e menores valores de salinidade e pH superficial e de profundidade (julho nos
pontos 3 e 4, janeiro e dezembro no ponto 4)(fig. 20),
c) a maior diversidade do ponto 2 (fig.19) estava associada às espécies S. testudineus, N.
barba, C. arenaceus, S. luniscutis, S. plagusia, S. rastrifer, C. spixii e M. furnieri (fig. 13)
e aos meses de novembro e, em menor intensidade,
setembro e junho (fig.19),
coincidentemente quando foram observados os maiores valores anuais de diversidade,
eqüitatividade e salinidade da água superficial (mês de setembro), pH (mês de novembro),
transparências da água (junho) e de menor profundidade (setembro) (fig.20),
Estes resultados mostraram que os maiores valores de diversidade e eqüitatividade
coincidiram com os meses e pontos amostrais que apresentaram: a) os maiores valores de
salinidade, pH e oxigênio dissolvido na água de profundidade e b) as menores profundidades e
teores de oxigênio dissolvido na água superficial. As maiores temperaturas, precipitações e
teores de oxigênio dissolvido na água de profundidade foram observadas nos meses de verão e
em todos os pontos amostrais, simultaneamente aos menores valores de salinidade (superficial
e de profundidade) e transparência da água.
Na tabela 44 são mostrados os resultados obtidos do cálculo da rarefação e na figura
21 encontram-se representadas as respectivas curvas da estimativa, para cada ponto amostral,
do número esperado de espécies com base em amostras de tamanho crescente e na soma das
probabilidades que cada espécie tem de ser incluída na amostra. Foi utilizado o n=2300 como
44
padrão, que correspondeu ao menor tamanho amostral obtido para os 4 pontos amostrais
analisados. Com base nesse valor, os pontos amostrais puderam ser ordenados em termos das
suas riquezas de espécies. O ponto 3 apresentou a maior riqueza, com um número estimado de
34 espécies, seguido pelos pontos 2, 1 e 4 com 32, 31 e 27 espécies respectivamente.
55
50
45
40
40
42
35
n
E (S )
41
36
30
34
32
31
28
25
20
15
Ponto
Ponto
Ponto
Ponto
10
5
1
2
3
4
0
0
2000
4000
6000
8000
10000
12000
14000
Tamanho da amostra
Figura 21. Curvas de rarefação para os diferentes pontos amostrais da Baía de Guaraqueçaba, Pr, Br. Onde E
(Sn ) é o número esperado de espécies.
45
4.2.3. Estrutura.
4.2.3.1. Comprimento e peso.
Na figura 22(a,b) e tabelas 45,46, 47 foram relacionadas as freqüências absolutas
(em número de exemplares), acumuladas, relativas(%) e relativas acumuladas (%) de
captura por intervalos de classe de CT (cm) e de peso (g). Foram mensurados 18993
peixes, dos quais 18060 (95,09%) exemplares apresentaram CT inferior a 20 cm.
Na figura 22 (c, d) encontram-se relacionadas as freqüências em número de
espécies por intervalos de classes de CT (cm) e as relações entre o CT observado e o CT
máximo, obtido da revisão de literatura por Corrêa (1987) e listados na tabela 45.
14
a
b
100
n=33857
n=18993
12
(500]
(450; 500 ]
(400; 450 ]
(350; 400 ]
Intervalos de Classe
(300; 350 ]
0
(250; 300 ]
0
20
(200; 250 ]
2
(150; 200 ]
4
40
(50; 100 ]
6
60
(0; 50]
8
(100; 150 ]
Número de exemplares
Freqüência relativa (%)
10
( 0; 2 ]
( 4; 6 ]
( 8; 10 ]
(12; 14 ]
(16; 18 ]
(20; 22 ]
(24; 26 ]
(28; 30 ]
(32; 34 ]
(36; 38 ]
(40; 42 ]
(44; 46 ]
(48; 50 ]
(52; 54 ]
(56; 58 ]
(60; 62 ]
(64; 66 ]
(68; 70 ]
(72; 74 ]
(76; 78 ]
(80; 82 ]
Número de exemplares
Freqüência relativa (%)
80
Intervalos de Classe
CT (cm)
Pêso do Corpo (g)
0,00
24
d
CET_EDE
ETR_CRO
GOB_OCE
HAR_CLU
ANC_PAR
SPH_TES
CIT_ARE
PAR_TROP
SYM_PLA
LYC_GRO
CIT_SPI
STE_BRA
ISO_PAR
GEN_GEN
CAT_SPI
PAR_BRAS
STE_RAS
DIP_RAD
MAC_ANC
MEN_AME
SYN_FOE
EUC_MEL
ACH_LIN
POM_COR
PEL_HAR
OLI_SAU
CTE_GRA
PEP_PAR
BAI_RON
TRIN_PAU
BAG_BAG
EUC_ARG
TRIN_MIC
GEN_LUT
EUC_GUL
OLI_SAL
TRIC_LEP
DIA_RHO
CYN_LEI
CYN_VIR
CHI_SPI
MIC_FUR
PRI_PUN
CHL_CRY
CYN_ACO
NET_BAR
CYN_MIC
CHAE_FAB
SCI_LUN
SEL_VOM
LAG_LAE
CAR_LAT
CT Médio
22
CT Máximo
20
(freqüência absoluta)
18
Número de Espécies
16
14
12
10
8
6
4
0,25
0,50
0,75
1,00
> 30
(28;30]
(26;28]
(24;26]
(22;24]
(20;22]
(18;20]
(16;18]
(14;16]
(12;14]
(8;10]
(10;12]
(6;8]
(4;6]
0
<= 2
2
(2;4]
Espécies
c
Intervalos de Classe (cm)
CT Médio+Desvio Padrão/CT Máximo
Figura 22. Diagramas comparativos: a) do número de exemplares de todas as espécies em
freqüência relativa (%) de captura por intervalos de classe de CT (cm), b) do número de
espécies por intervalos de CT médio observado e CT máximo da literatura; c) da relação
entre o CT médio (+ s) obtido e o CT máximo da literatura e d) número de exemplares em
freqüência relativa (%) de captura por intervalos de classe de peso do corpo (g).
46
Na figura 23 (a e b) são apresentadas as espécies e respectivas freqüências de
exemplares mensurados, desvios (s) e erros padrões (e) das estimativas das médias de CT
(cm), CP(cm) e peso (g).
Entre as 52 espécies capturadas, 12 espécies (23,08%) apresentaram CT médio
entre 5 e 10 cm, 35 (67,31%) entre 10 e 20 cm e 5 (9,61%) superaram este intervalo
(tab.45,
fig.22d e 23a). Com relação aos CT máximos relatados na literatura
(Corrêa,1987), 10 espécies (19,23%) não ultrapassam os 20 cm, 20 (38,46%) atingem
entre 20-30 cm, e 22 (42,31%) podem ultrapassar os 30 cm (fig. 22d, c e tab. 46). Estes
resultados mostraram que as curvas de distribuição do número de espécies por intervalos
de classe de CT médio e CT máximo desviaram respectivamente para a esquerda e para a
direita (fig.22d). Portanto as populações de 38 espécies (73,07%) estavam constituídas
principalmente por indivíduos com CT médios 50% inferiores ao CT máximo. Ainda, a
relação entre o CT médio (mais o desvio padrão) das capturas e o CT máximo mostrou que
2 espécies (Cetengraulis edentulus e Etropus crossotus) ou 3,84% atingiram o CT máximo,
6 (Gobionellus oceanicus, Harengula clupeola, Anchoa parva, Sphoeroides testudineus,
Citharichthys arenaceus e Paralichthys tropicus) ou 11,54% atingiram acima de 75% do
CT máximo, 16 (Symphurus plagusia, Lycengraulis grossidens, Citharichtys spilopterus,
Stellifer brasiliensis, Isopisthus parvipinnis, Genidens genidens, Cathorops spixii,
Paralonchurus brasiliensis, Stellifer rastrifer, Diplectrum radiale, Macrodon ancylodon,
Menticirrhus americanus, Synodus foetens e Pomadasys corvinaeformis) ou 30,77%
acima de 50% do CT Máximo e as demais (ou 53,84%) apresentaram um CT médio
somado ao desvio padrão inferior a 50% do CT máximo (fig.22c).
Foram pesados 33857 peixes dos quais 31200 (92,15%) apresentaram peso menor
ou igual a 50 g, 2102 (6,21%) entre 50-100 g, 376 (1,11%) entre 100-150 g e 179 (0,53%)
acima de 200 g (tabela. 45, 47 e fig. 22b, 23b).
47
a
40
CT(cm)
30
±1,96*e
±1,00*e
Média
n=33869
20
0
ACH_LIN
ANC_PAR
BAG_BAG
BAI_RON
CAR_LAT
CAT_SPI
CET_EDE
CHAE_FAB
CHL_CRY
CHI_SPI
CIT_ARE
CIT_SPI
CTE_GRA
CYN_ACO
CYN_LEI
CYN_MIC
CYN_VIR
DIA_RHO
DIP_RAD
EUC_ARG
EUC_GUL
EUC_MEL
ETR_CRO
GEN_GEN
GEN_LUT
GOB_OCE
HAR_CLU
ISO_PAR
LAG_LAE
LYC_GRO
MAC_ANC
MEN_AME
MIC_FUR
NET_BAR
OLI_SAL
OLI_SAU
PAR_BRAS
PAR_TROP
PEL_HAR
PEP_PAR
POM_COR
PRI_PUN
SCI_LUN
SEL_VOM
SPH_TES
STE_BRA
STE_RAS
SYM_PLA
SYN_FOE
TRIC_LEP
TRIN_MIC
TRIN_PAU
10
Espécies
b
600
500
±1,96*e
±1,00*e
Média
n=33869
400
PÊSO (g)
300
200
100
ACH_LIN
ANC_PAR
BAG_BAG
BAI_RON
CAR_LAT
CAT_SPI
CET_EDE
CHAE_FAB
CHL_CRY
CHI_SPI
CIT_ARE
CIT_SPI
CTE_GRA
CYN_ACO
CYN_LEI
CYN_MIC
CYN_VIR
DIA_RHO
DIP_RAD
EUC_ARG
EUC_GUL
EUC_MEL
ETR_CRO
GEN_GEN
GEN_LUT
GOB_OCE
HAR_CLU
ISO_PAR
LAG_LAE
LYC_GRO
MAC_ANC
MEN_AME
MIC_FUR
NET_BAR
OLI_SAL
OLI_SAU
PAR_BRAS
PAR_TROP
PEL_HAR
PEP_PAR
POM_COR
PRI_PUN
SCI_LUN
SEL_VOM
SPH_TES
STE_BRA
STE_RAS
SYM_PLA
SYN_FOE
TRIC_LEP
TRIN_MIC
TRIN_PAU
0
Espécies
ACHI_LIN = Achirus lineatus
ANC_PAR = Anchoa parva
BAG_BAG= Bagre bagre
BAI_RON=Bairdiella ronchus
CAR_LAT=Caranx latus
CAT_SPI=Cathorops spixii
CHAE_FAB=Chaetodipterus faber
CHL_CRY=Chloroscombrus crysurus
CHI_SPI=Chilomycterus spinosus
CET_EDE=Cetengraulis edentulus
CIT_ARE=Citharichtys arenaceus
CIT_SPIL=Citharichthys spilopterus
CTE_GRA=Ctenosciaena gracillicirrhus
CYN_ACO=Cynoscion acoupa
CYN_LEI=Cynoscion leiarchus
CYN_MIC=Cynoscion microlepidotus
CYN_VIR=Cynoscion virescens
DIA_RHO=Diapterus rhombeus
DIP_RAD=Diplectrum radiale
EUC_ARG=Eucinostomus argenteus
EUC_GUL=E. gula
EUC_MEL=E. melanopterus
ETR_CRO=Etropus crossotus
GEN_GEN=Genidens genidens
GEN_LUT=Genyatremus luteus
GOB_OCE=Gobionellus ocenanicus
HAR_CLU=Harengula clupeola
ISO_PAR=Isopisthus parvipinnis
LAG_LAE=Lagocephalus laevigatus
LYC_GRO=Lycengraulis grossidens
MAC_ANCY=Macrodon ancylodon
MEN_AME=Menticirrhus americanus
MIC_FUR=micropogonias furnieri
NET_BAR=Netuma barba
OLI_SAL=Oligoplithes saliens
OLI_SAL=Oligoplithes saurus
PAR_BRAS=Paralonchurus brasiliensis
PAR_TRO=Paralichthys tropicus
PEL_HAR=Pellona harrowerii
PEP_PAR=Peprilus paru
POM_COR=Pomadasys corvinaeformis
PRI_PUN=Prionotus punctatus
SCI_LUN=Sciadeichthys luniscutis
SEL_VOM=Selene vomer
SPH_TES=Sphoeroides testudineus
STE_BRA=Stellifer brasiliensis
STE_RAS=Stellifer rastrifer
SYM_PLA=Symphurus plagusia
SYN_FOE=Synodus foetens
TRIC_LEP=Trichiurus lepturus
TRIN_MIC=Trinectes microphtalmus
TRIN_PAU=Trinectes paulistanus
Figura 23. Comparações do: a) CT (cm) médios e b) pesos médios das diferentes espécies
capturadas na Baía de Giuaraqueçaba. Onde e=erro padrão das estimativas nos limites de (0,05 e
0,10% de significância).
48
O peso médio situou-se abaixo de 100 g em 50 espécies (96,15%) e ultrapassou
este valor em 2 (3,85%) (Menticirrhus americanus e Paralichthys tropicus) (fig. 23b).
Na tabela 48 as médias, desvios e erros padrões das estimativas de CT , CP e peso
por pontos e épocas do ano amostrados de todas as espécies agrupadas (fig.24a). De modo
geral, para todos os pontos amostrais, as médias de CT dos meses de primavera (setembro,
outubro e novembro) sofreram reduções em comparação às demais épocas do ano, com
exceção do CT médio de inverno (junho, julho e agosto) no ponto amostral 3 (tabela.48 e
fig.24a). Nos pontos amostrais 1 e 4 ocorreu a redução gradativa das médias do CT dos
meses de primavera (setembro, outubro e novembro) para os de inverno (junho, julho e
agosto) (tab. 48 e fig.24a). Os pontos 2 e 3 mostraram tendências diferenciadas dos demais,
com baixos CT médios respectivamente nos meses de outono (março, abril e maio) e de
verão (dezembro, janeiro e fevereiro) (fig.24a). Destcaram-se as médias de CT inferiores a
15 cm em todos os ponto e épocas amostradas.
Na figura 24b o resultado da comparação das médias dos pesos das espécies
agrupadas por pontos amostrais associados às épocas do ano. Como esperado, a tendência
dos pesos médios foi similar àquela observada para o CT (fig. 24a). As médias de peso dos
pontos amostrais nas épocas do ano situaram-se abaixo de 50 g.
Nas figuras 25 a 34 foram plotados simultaneamente, para as 10 espécies mais
abundantes, o número de exemplares por intervalos de classe de CT (cm) e de peso (g)
como descritores e os meses e pontos de amostragem como objetos nos planos fatoriais da
ACP. Em todas as espécies a análise restringiu-se aos dois primeiros componentes que
explicaram para:
1) Cathorops spixii respectivamente 39,17 e 18,86 % da variância total dos dados. Os dois
primeiros componentes associados explicaram 58,03 % da variabilidade (tab.49).
Contribuíram positivamente, em maior grau, para formação do eixo 1 os intervalos de
49
15,0
Ponto 1
a
13,5
12,0
10,5
Ponto 2
13,5
12,0
10,5
9,0
15,0
Ponto 3
CT (cm)
9,0
15,0
13,5
12,0
10,5
Ponto 4
9,0
15,0
±1,96*e
±1,00*e
Média
13,5
12,0
10,5
9,0
PRIMAVERA
VERÃO
OUTONO
INVERNO
PRIMAVERA
VERÃO
OUTONO
INVERNO
65
Ponto 1
b
50
35
20
Ponto 2
50
35
20
5
65
Ponto 3
PÊSO (g)
5
65
50
35
20
Ponto 4
5
65
50
35
20
5
Figura 24. Comparações dos: a) CT médios (cm) e b) pesos médios (g) de todas as
espécies agrupadas pelos pontos e períodos do ano amostrados. Onde e=erro padrão das
estimativas aos níveis de 0,05 e 0,10 significância.
50
4.9
Abr
CT<=10
P<=15
Abr
Mar
2.9
Componente 2
Fev
Jun
P(15;20]
Pontos
amostrais
1
2
3
4
Abr
Fev
0.9
CT(10;15]
Out
Set
P>25
Jan Mai
Mar
Jan
Dez
Jun
Fev Jan Fev
Mar
Dez Jan
Jan
CT(15;20]
Set
Out
MaiP(20;25]
-1.1
Nov
demais
CT(20;25]
pontos
Dez CT>25
e meses
Mai
Nov
Dez
-3.1
-1.6
0.4
2.4
4.4
6.4
Componente 1
Figura 25. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Cathorops spixii (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
CT entre (10;15], (15;20] e superior a 20 cm e de peso superior a 30 g (fig. 25). Para a
formação do eixo 2 contribuíram positivamente os intervalos de CT e peso
respectivamente inferiores a 10 cm e 10 g (fig. 25). Correlacionaram-se positivamente
ao eixo 1 o mês de novembro (ponto 1) e, em menor grau, dezembro, outubro e março
(todos no ponto 1). Ao eixo 2 correlacionaram-se positivamente o mês de abril (ponto
2 e, em menor grau, 4) (fig.25). Os meses de março (ponto 2) e abril (ponto1)
apresentaram correlação positiva tanto com o eixo 1 quanto com o 2, enquanto que os
meses de dezembro e outubro (ambos no ponto 1) apresentaram correlação negativa
com o último eixo. Estes resultados mostraram que as freqüências de exemplares nos
intervalos de classes de CT e de peso respectivamente: a) iguais ou inferiores a 10 cm e
10 g aumentaram do mês de novembro (ponto1), dezembro (ponto1), janeiro (ponto 4),
fevereiro (pontos 1 e 4), março (ponto 2) até atingirem o valor máximo em abril
(pontos 1 e, principalmente, 2 e 4), b) (10;15], (15;20] e superiores a 20 cm e a 30 g
51
encontravam-se altas em outubro, novembro e dezembro (no ponto1) e diminuíram nos
meses sucessivos em todos os pontos amostrais (fig. 25). O intervalo de peso entre
(10;20] g começou a aumentar à partir de fevereiro (ponto1 e 4) e foi mais abundante
em março (ponto2) e abril (ponto1).
2) Citharichthys arenaceus respectivamente 72,09 e 16,33% da variância total dos dados,
que associados explicaram 88,42% da variabilidade (tab.49). Contribuíram para
formação do eixo 1 e 2, nas coordenadas positivas, os intervalos de CT entre (5;10] e
de peso igual ou inferior a 5 g (fig. 26). Nas coordenadas positivas e negativas,
respectivamente para o eixo 1 e 2, os CT superiores a 10 cm e os pesos entre (5;15],
7
Componente 2
P<=5
5
Mar
3
CT(5;10]
1
-1
Abr
Abr
Abr
Mai
Set
Fev
Jun
P(5;15]
P(15;25]
CT>=10
demais
pontos
e meses
Jul
P>25
-3
-0.9
Pontos
amostrais
1
2
3
4
1.1
3.1
5.1
Componente 1
Mai
7.1
9.1
Figura 26. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Citharichtys arenaceus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
(15;25] e superiores a 25 g (fig. 26). Para a formação do eixo 2, coordenada positiva,
contribuíram principalmente os intervalos de CT entre (5;10] cm e de peso inferiores a
5 g (fig.26). Apresentaram correlação positiva com o eixo 1 os meses de julho (ponto
52
1) e julho (ponto 2). Correlacionaram-se positivamente com o eixo 2 o mês de março
(ponto 2) e, em menor grau, os meses de abril e fevereiro (ponto 2)(fig.26). O mês de
março (ponto 2) apresentou correlação positiva com ambos os eixos e os de maio
(ponto 2) e julho (ponto 1) correlacionaram-se positivamente com o eixo 1 e
negativamente com relação ao eixo 2 (fig.26). Estes resultados evidenciaram
estratificação temporal em relação ao CT e peso, uma vez que as freqüências de
exemplares nos maiores intervalos de CT e peso mostraram-se baixas nos meses de
fevereiro, março e abril (ponto 2) e aumentaram dos meses de maio/junho (ponto 2)
para julho (no ponto 1). Ao contrário, as freqüências dos exemplares do menor
intervalo de classe de CT e de peso foram maiores nos meses de fevereiro, março e
abril (exclusivamente no ponto 2) e diminuíram nos meses de maio/junho (ponto 2) e
julho (ponto1) (fig. 26). Ainda, o primeiro período pode corresponder a época de
recrutamento da espécie em função da maior abundância dos menores exemplares.
3) Etropus crossotus respectivamente 65,21 e 25,02% da variância total dos dados.
Associados os componentes explicaram 90,22% da variação (tab.49). Contribuíram
positivame nte para a formação do componente 1 os intervalos de CT (8;10] e
superiores a 10 cm e de peso (7;10], (10;13] e superiores a 13 g (fig. 27). Para a
formação do componente 2 coordenada negativa contribuíram os intervalos de CT
iguais e inferiores a 6 e entre (6;7] cm e de peso iguais e inferiores a 7 g (fig. 27).
Correlacionou-se positivamente com o componente 1 o mês de julho (ponto 1).
Apresentaram correlação positiva com o componente 2 os meses de janeiro, fevereiro e
novembro (unicamente no ponto 3) (fig.27). Deste modo, os maiores exemplares
ocorreram principalmente no primeiro mês e ponto e os menores nos últimos meses no
ponto amostral 3. Nos meses de maio (ponto 3) e de junho (ponto 2) as freqüências de
exemplares nos intervalos menores e intermediários de CT e de peso foram baixas e
53
similares. Estes resultados mostraram que no período compreendido entre novembro e
fevereiro ocorreu o recrutamento da espécie na área, tendo como apogeu o mês de
janeiro (fig. 27).
3.4
demais
pontos
e meses
Componente 2
1.4
-0.6
Jul
P>13
P(10;13]
CT>10
Set
Ago
Dez
CT(8;10]
P(7;10]
Mar
Abr
Jun
Mai
-2.6
P<=7
Nov
-4.6
Pontos
amostrais
1
2
3
4
Fev
CT<=6 CT(6;8]
Jan
-6.6
-1
3
7
11
15
Componente 1
Figura 27. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Etropus crossotus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
4) Genidens genidens isoladamente 64,06 e 21,21% da variabilidade. Associados
passaram a explicar 85,27% da variação total dos dados (tab. 49). Contribuíram
positivamente para a formação do componente 1 os intervalos de CT (10;15] e (15;20]
cm e de peso (10;20], (20;30] g (fig. 28). Para a formação do componente 2 coordenada
positiva contribuíram os intervalos de CT (20;25] e superiores a 25 cm e de peso
superiores a 30 g (fig. 28). Na coordenada negativa, ainda do componente 2,
contribuíram os intervalos de CT inferiores e iguais a 10 cm e de peso inferiores e
iguais a 10 g (fig. 28). Correlacionou-se positivamente com o componente 1 o mês de
54
novembro (pontos 3 e, em menor grau, 1). Com os componentes 1 e 2 correlacionou-se
positivamente o mês de dezembro (ponto) e negativamente, para o último, os meses de
6
Dez
4
CT(20;25]
P>30
demais
pontos
e meses
Pontos
amostrais
1
2
3
4
CT>25
Componente 2
2
Fev
Dez
Jan
0
-2
Jun
Mar
Abr
Mar
Ago
Abr
Out
Abr
Nov
CT(15;20]
Nov
P(20;30]
CT(10;15]
P(10;20]
-4
Set
Abr
CT<10 P<10
-6
-2
1
4
7
10
13
Componente 1
Figura 28. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Genidens genidens (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
setembro (ponto 3), abril (pontos 2 e, em menor grau, 4) e de março (ponto 3) (fig. 28).
Estes resultados evidenciaram: a) a existência de estratificação na variabilidade
temporal e espacial com relação ao comprimento e peso e b) que o auge do
recrutamento da espécie na área ocorreu no mês de abril no ponto 2. É importante
destacar a ocorrência de freqüências similares de exemplares com intervalos de CT e
de peso respectivamente inferiores e iguais a 10 cm e 10g e de (10;15] cm e (10;20] g
presentes no mês de setembro no ponto 3. Esta sobreposição temporal de intervalos de
CT e peso pode ser explicada pela justaposição dos exemplares do recrutamento do ano
analisado àqueles do ano anterior.
55
5) Isopisthus parvipinnis, respectivamente 57,39 e 24,24% da variabilidade. Associados
explicaram 81,63% da variação total dos dados (tab.49).Os intervalos de CT (10;15]
cm e de peso (10;15] e (15;20] g contribuíram positivamente para a formação do
componente 1 e os intervalos de CT (15;20] e superiores a 20 cm positivamente para
formação do componente 2 (fig.29). Os intervalos de CT inferiores ou iguais a 10 cm e
de peso inferiores ou iguais a 10 g contribuíram para a formação da coordenada
negativa do último componente (fig.29).
7
CT>20
CT(15;20]
Mar
5
P>20
Componente 2
Out
1
Set
Nov
3
Jan
Nov
Dez
Abr
Pontos
amostrais
1
2
3
4
Set
Jul
P(15;20]
CT(10;15]
P(10;15]
Ago
Mai
-1
-3 demais
pontos
e meses
CT<=10
P<=10
Abr
-5
-1
1
3
5
7
9
11
Componente 1
Figura 29. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Isopisthus parvipinnis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
O intervalo de peso superior a 20 g contribuiu para a formação das coordenadas
positivas de ambos os componentes e o de (10;15] g para a formação da positiva do
componente 1 e negativa do 2 (fig.29).
Correlacionaram-se positivamente ao
componente 1, isto é às classes de CT e de peso intermediárias, os meses de agosto
(ponto 1) e de setembro (ponto 4). O mês de março e abril (ponto 1) apresentaram
correlação positiva respectivamente às maiores e às menores classes de CT e de peso
56
(fig. 29). Os resultados mostraram que nos meses de agosto (ponto 1) e de setembro
(ponto 4) a população de I. parvipinnis estava constituída tanto por exemplares de
classes intermediárias quanto inferiores de CT e de peso, principalmente pelas
primeiras.Em março (ponto 1) os exemplares das maiores classes de CT e de peso
passaram a predominar. Posteriormente em abril, neste mesmo ponto, suas freqüências
foram ultrapassadas pelas das classes menores e intermediárias de CT e de peso, que
correspondeu ao mês de maior recrutamento (fig. 29).
6) Micropogonias furnieri isoladamente 68,25 e 15,52 % da variabilidade. Associados
explicaram 83,77% da variância total dos dados (tab. 49). Contribuiu para formação do
eixo 1, coordenada positiva, o intervalo de peso (25;30] g. Contribuíram para a
formação do eixo 2: 1) na coordenada positiva os intervalos de CT (15;30] cm e de
peso superior a 35 g e 2) na coordenada negativa os intervalos de CT inferior ou igual
a 15 cm e de peso inferior ou igual a 15 e entre (15;25] g (fig. 30). Correlacionou-se
positivamente com o componente 1 e 2 o mês de dezembro (ponto 2 e 3).
4.9
Pontos
amostrais
1
2
3
4
CT(15;30]
2.9
Componente 2
P>35
0.9
Jun
Jan
Mai
Fev
Mar
Out
Nov
Nov
Ago
Set
Set
CT(30;45]
Fev
Mar
-1.1
Dez
P(25;30]
Jan
Nov
Jan
Fev
Abr
p(15;25]
CT<=15
P<=15
Mar
demais
pontos e
meses
Out
Jan
-3.1
-2
Dez
0
2
4
6
8
10
Componente 1
Figura 30. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Micropogonias furnieri (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
57
Correlacionaram-se positivamente com o componente 1 e negativamente com o 2, por
ordem de contribuição, os meses de janeiro e outubro (ponto 3), de março e abril
(ponto 2) e novembro (ponto 3) (fig. 30). Estes resultados mostraram que os maiores
intervalos de classe de CT e de peso foram abundantes no mês de dezembro (nos
pontos 2 e, predominantemente, 3) e os menores em outubro e, em maior grau, janeiro
(ponto 3). Deste modo os recrutas foram inicialmente capturados em outubro, com o
pico de captura em janeiro. As freqüências de exemplares nas maiores e menores
classes de CT e de peso igualaram-se nos meses de novembro (ponto 3) e de dezembro
(em maior grau no ponto 2 e menor no 3), com maior intensidade no último mês. A
partir de dezembro as freqüências de exemplares das maiores e menores classes de CT
e de peso diminuíram. As classes de menor intervalo ressurgiram, em baixas
freqüências, nos meses de março e abril no ponto 2 (fig.30).
7) Netuma barba respectivamente 38,06 e 28,59 % da variação. Associados exprimiram
66,65 % da variabilidade total dos dados (tab.49). Contribuíram para a formação da
coordenada negativa do eixo 1 os intervalos de classe de CT (13;30] cm e de peso
(20;30] e superior a 30 g (fig.31). Participaram na formação da coordenada positiva do
eixo 2 os intervalos de classe de CT e de peso inferiores ou iguais a 15 cm e 10 g.
Correlacionaram-se positivamente, em maior grau, com o componente 1 o mês de
dezembro (ponto1) e com o componente 2 o mês de abril (pontos 2 e 4) e, em menor
grau, para o primeiro os meses de março e maio (ponto 2) e para o segundo novembro
(ponto 4) (fig.31). Estes resultados evidenciaram que as maiores classes de CT e de
peso aumentaram do mês de setembro (em todos os pontos amostrais) para o de
dezembro (ponto 1). Em dezembro (ponto 4) as freqüências das maiores classes de CT
e de peso diminuíram e das menores aumentaram, igualando-se. Em março (ponto 2)
passaram a ser capturados somente exemplares da menores classes, que atingiram as
58
maiores freqüências no mês de abril (pontos 2 e 4) e diminuíram em maio (ponto 2)
(fig.31). Ainda mostraram que o recrutamento da espécie na área pode ter iniciado-se
em dezembro no ponto 1 mais externo e atingido o máximo em abril nos pontos
intermediários 2 e 4.
7.2
5.2
Componente 2
Abr
Abr
Pontos
amostrais
1
2
3
4
CT<=15
P<=10
3.2
Mar
Mai
P(10;20]
1.2
P(20;30]
Dez
Nov
CT(13;30]
P>30
Mar
-0.8
-10
-8
-6
Dez
-4
Dez
Dez Set Abr
Set
Jan
CT>45 CT(30;45]
Out
Jan Fev
-2
0
2
Componente 1
Figura 31. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Netuma barba (descritores) nos meses e pontos de amostragem (objetos), sobrepostos
no plano bidimensional dos componentes principais. Demais meses e pontos encontramse sobrepostos e indicados por uma seta.
8) Sciadeichtys luniscutis isoladamente 38,71 e 33,79 % da variabilidade. Associados
representaram 72,51% da variância total dos dados (tab.49). Contribuíram para a
formação da coordenada positiva dos eixos 1 e 2 os intervalos de CT inferior ou igual
a 10 e entre (10;20] cm e de peso inferior ou igual a 10 e entre (10;20] g (fig.32).
Contribuíram negativamente para formação de ambos os eixos os CT e pesos
superiores a 30 cm e 30 g. Correlacionaram-se positivamente, em maior grau, aos dois
primeiros componentes o mês de abril (ponto 2) e junho (ponto 1) e negativamente o
meses de novembro e outubro (ponto 4). Em outubro e novembro (no ponto 4)
prevaleceram os exemplares dos maiores intervalos de CT e de peso. Em dezembro
59
(ponto1) as freqüências dos últimos diminuiu e as dos menores intervalos de CT e de
peso aumentaram, igualando-se (fig. 32). Em março (ponto 2 e 3) apareceram, em
baixas freqüências, exemplares com CT e peso inferiores ou iguais a 10 cm e 10g, que
atingiram o máximo em abril (ponto2), juntamente com os exemplares das classes de
CT e de peso (10;20] cm e g. Nos meses posteriores, isto é maio e junho (ponto 2),
apenas os exemplares do último intervalo diminuíram, contudo aumentaram no último
mês apenas no ponto amostral 1 (fig.32). Como para as espécies anteriores, estes
resultados parecem estar intimamente relacionados com a atividade reprodutiva. A
presença dos maiores exemplares nos meses de outubro e novembro (no ponto 4),
evidenciaram que a reprodução pode estar sendo realizada neste período e ponto
amostral. Entre os meses de dezembro (ponto 1) e abril (ponto 2) ocorreu o
recrutamento resultante que, provavelmente em junho, abandonou o ponto 2 e dirigiuse para o 1.
8.8
Nov
Pontos
amostrais
1
2
3
4
Componente 2
6.8
4.8
P>30CT(20;30]
CT>30
Dez
2.8
Out
Jun
P(20;30]
Mai
0.8
demais
pontos
e meses
-1.2
-7
-4
Jun
Mai
Mar Abr
Fev
Fev
Mar
AbrJul
Jan
-1
2
Abr
CT(10;20]
P(10;20]
P<=10
CT<=10
Mar
5
8
Componente 1
Figura 32. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Sciadeichtys luniscutis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais.
60
9) Sphoeroides testudineus isoladamente 59,69 e 14,48 % e associados 74,17 % da
variação total dos dados(tab.49). Contribuíram em maior grau para a formação do eixo
1 os intervalos de classe de CT (5;10] e (10;15] cm e de peso (20;30], (30;40] e
superior a 40 g (fig.33). Para a formação do eixo 2, coordenada positiva contribuiu o
intervalo de CT inferior ou igual a 5 cm. O intervalo de peso inferior ou igual a 20 g
contribuiu positivamente para a formação de ambos os eixos. Correlacionaram-se
positivamente ao componente 1 os meses de outubro e março (ponto2) e dezembro
(ponto4) (fig.33).
8.5
Jun
CT<=5
Componente 2
6.5
4.5
P<=20
Pontos
amostrais
1
2
3
4
2.5
Nov
Dez
Ago
Set
0.5
Dez
Fev
-1.5
-1.2
0.8
Jul
Jan
Jan
P(30;40]
CT(10;15]
CT(5;10]
P(20;30]
P>40
CT(10;15]
2.8
4.8
Componente 1
Mar
Out
6.8
8.8
Figura 33. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso
de Sphoeroides testudineus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
Ao componente dois correlacionou-se positivamente, em maior grau, apenas o mês de
junho (ponto 3). As altas freqüências de exemplares nos maiores intervalos de classe
de CT e de peso nos meses de outubro e março (ponto2) e dezembro (ponto 4) podem
estar relacionadas, como suposto para as espécies anteriores, com a atividade
61
reprodutiva (fig.33). Contudo os prováveis exemplares originados do recrutamento (CT
e peso inferiores ou iguais a 5 cm e 20 g) só foram constatados no mês de junho e no
ponto amostral 3 (fig.33).
10) Stellifer rastrifer separadamente 60,06 e 17,87 %
e associados
77,93 % da
variabilidade (tab. 49). Contribuíram para a formação do eixo 1, na coordenada
positiva, o intervalo de CT de entre (10;15] cm e de peso entre (20;30] g (fig.34).
Contribuíram na formação do eixo 2, na coordenada positiva, os intervalos de CT e de
peso superiores a 15 cm e 30 g e na coordenada negativa os intervalos de CT inferior
ou igual a 10 cm e de peso inferior ou igual a 10 e entre (10;20] g (fig. 34).
6.7
Pontos
amostrais
1
2
3
4
CT>15
Out
4.7
Componente 2
P>30
2.7
0.7
-1.3
-3.3
Jan
Out
Dez
Dez
Mai
Fev
Mar
Jul
Out
Mai Dez
Out Fev
Set
Abr
Mar
Abr
demais
pontos e
meses
Abr
-1.7
0.3
2.3
P(20;30]
Jan
CT(110;15]
Nov
Set
P<=10
CT<=10
P(10;20]
4.3
6.3
8.3
Componente 1
Figura 34. Diagrama da distribuição das freqüências por intervalos de classe de CT e peso de Stellifer
rastrifer (descritores) nos meses e pontos de amostragem (objetos), sobrepostos no plano bidimensional dos
componentes principais.
Correlacionaram-se positivamente com o componente 1, em maior grau, o mês de
janeiro (ponto 2) e, em menor grau, novembro (ponto 1) e setembro (ponto3). Com
o componente 2 correlacionou-se positivamente o mês de outubro (ponto 4) e
62
negativamente, em maior grau, abril (ponto 1) e, em menor grau, março (ponto 2) e
setembro (ponto4). Estes resultados mostraram que em outubro (ponto 4) a
população estava composta principalmente por exemplares dos maiores intervalos
de classe de CT e de peso. Em novembro, no ponto 1, a maior freqüência aumentou
para o intervalo de CT entre (10;15] cm e de peso entre (20; 30] g. Em janeiro as
freqüências dos maiores e dos menores intervalos de CT e de peso voltaram
respectivamente a aumentar e diminuir. Os últimos passaram a dominar em abril no
ponto 1. Como já observado anteriormente estes resultados podem estar
relacionados com a atividade reprodutiva da espécie na área.
63
4.2.3.2. Estádio de Maturação Gonadal.
Do total de 25244 peixes examinados, independente da espécie, mês e pontos
amostrais, 17065 (67,60%) dos exemplares correspondiam ao sexo indeterminado, 3505
(13,88%) a machos e 4674 (18,52%) a fêmeas (fig. 35a e tab.50). Com relação ao estádio
de maturação, dos 25171 exemplares analisados, a maioria ou 18399 (73,09%) eram
imaturos, 1947 (7,73%) encontravam-se em descanso reprodutivo. Os restantes
encontravam-se nos estádios em maturação (1056 ou 4,20%), maduro (608 ou 2,42%),
reprodução (763 ou 3,03%) e desovados ou esvaziados (2366 ou 9,41%) e incubando ovos
(32 ou 0,13%)(fig.35b). Com base nestes resultados as populações naturais de peixes da
Baía de Guaraqueçaba estavam constituídas principalmente por exemplares juvenis e
ainda, a região também foi utilizada como local de reprodução.
b
100
100
n=25139
80
60
60
REPRODUÇÃO
MADURO
0
MATURAÇÃO
20
DESCANSO
Macho
0
Fêmea
20
40
IMATURO
40
Número de exemplares
Freqüência relativa (%)
80
Indeterminado
Freqüência relativa (%)
Número de exemplares
n=25244
DESOVADO
a
SEXO
Estádio de Maturação Gonadal
Figura 35. Histogramas comparativos do número de exemplares de todas as espécies (%)
capturados na Baía de Guaraqueçaba por: a) sexo e b) estádios de maturação gonadal.
A distribuição das freqüências dos exemplares imaturos, por mês e pontos
amostrais, mostraram que as maiores freqüências ocorreram respectivamente no mês de
abril e nos pontos intermediários 2 e 4 (tab.51 e fig.36a,b).
Considerando apenas os elementos que participavam da atividade reprodutiva (em
maturação até desovados) as distribuições de freqüência das espécies agrupadas por mês
(fig. 36c), evidenciaram que número de exemplares:
64
a) em maturação atingiu o maior número no mês de setembro diminuindo
gradativamente até o mês de novembro. A partir do último manteve-se
aproximadamente constante e em baixos valores até o mês de abril,
b) maduros apresentaram baixas freqüências em setembro, aumentando em
outubro e diminuindo em novembro. Reapareceram em freqüências incipientes
entre janeiro e abril,
c) em
reprodução
aumentou
entre
setembro
e
outubro
diminuindo
progressivamente até janeiro,
d) desovados ou esvaziados cresceu exponencialmente entre outubro e novembro
decrescendo gradativamente até abril,
b
8000
n=16507
7000
n=16507
40%
35%
30%
5000
25%
4000
20%
3000
15%
2000
10%
JUL
MAI
JUN
ABR
FEV
MAR
JAN
DEZ
NOV
0%
SET
0
OUT
5%
AGO
1000
(freqüência absoluta)
6000
6000
(freqüência absoluta)
Número de Exemplares Imaturos
7000
Número de Exemplares Imaturos
a
30%
5000
25%
4000
20%
3000
15%
2000
10%
1000
5%
0
0%
1
Meses
d
Maturação
Maduro
Reprodução
Desovado
700
Número de Exemplares
(freqüência absoluta)
600
500
3
900
15%
800
12,5%
700
10%
400
7,5%
300
5%
200
4
Maturação
Maduro
Reprodução
Desovado
JUL
JUN
MAI
ABR
MAR
FEV
JAN
DEZ
NOV
OUT
SET
AGO
Meses
15%
500
10%
400
7,5%
300
5%
200
2,5%
100
0%
17,5%
12,5%
600
2,5%
100
0
2
Pontos Amostrais
800
Número de Exemplares
(freqúência absoluta)
c
35%
0
0%
1
2
3
4
Pontos Amostrais
Figura 36. Histogramas do número de exemplares imaturos (freqüência absoluta) nos
meses (a) e pontos amostrais (b), e dos exemplares que efetivamente participaram do
processo reprodutivo nos meses (c) e pontos amostrais (d).
65
As distribuições de freqüência dos exemplares por estádios, que efetivamente
caracterizavam o processo reprodutivo, nos diferentes pontos amostrais (fig. 36d)
mostraram que o:
a) em maturação predominou nos pontos 1 e 3,
b) maduro e em reprodução apresentaram tendências similares, sendo mais
freqüentes no ponto 1, diminuindo no sentido dos pontos 2 e 3. O primeiro
ocorreu no ponto 4 em freqüência intermediária às dos pontos 1 e 2, e o último
ocorreu em freqüência similar à do ponto 2,
c) desovado ou esvaziado foi mais freqüente nos pontos 1 e 4, diminuindo em
proporções similares nos pontos 2 e 3.
Estes resultados evidenciaram a existência de preferências temporais e espaciais na
reprodução e recrutamento. A primeira parece iniciar-se no mês de setembro com o pico
em outubro. Ainda, corroboraram esta assertiva a ocorrência de exemplares desovados ou
esvaziados presentes no mês de novembro. Cabe salientar que o número de exemplares em
reprodução foi significativamente inferior ao de desovados. Este fato ocorreu em função da
dificuldade na captura dos exemplares exatamente no estádio em reprodução, uma vez que
a transição do último para o desovado ou esvaziado ocorre de modo imediato.
Na figura 37 os resultados da comparação do número de espécies por estádios de
maturação gonadal. A tendência geral foi a redução em número dos estádios iniciais para
os mais avançados. Dentre as 52 espécies coletadas 51 (98,08%) foram capturadas no
estádio juvenil, 34 (65,38%) virgem/descanso, 29(55,77%) em maturação, 28(53,84%)
maduro, 17(32,70%) em reprodução, 21 (40,38%) desovado ou esvaziado e 4 (7,69%)
incubando ovos e larvas(fig. 37a). Esta distribuição mostrou que todas as espécies, com
exceção de Cynoscion virescens, utilizaram a área nos estádios iniciais de seu ciclo de vida
(fig.37b). Principalmente as espécies C. spixii, G. genidens e Stellifer rastrifer
66
a
100
90
n=52
51
Imaturo
Descanso
Em maturação
Maduro
Reprodução
Desovado
Incubando
Número de Espécies (%)
80
70
60
34
29
50
28
40
21
30
17
20
10
4
0
b
<=50
(200;300]
Número de Exemplares
(50;100]
(300;400]
(100;200]
>400
Desovado
Reprodução
Maduro
Maturação
Virg/Desc
Imaturo
ACH_LIN
ANC_PAR
BAG_BAG
BAI_RON
CAR_LAT
CAT_SPI
CET_EDE
CHAE_FAB
CHL_CRY
CHI_SPI
CIT_ARE
CIT_SPI
CTE_GRA
CYN_ACO
CYN_LEI
CYN_MIC
CYN_VIR
DIA_ROM
DIP_RAD
EUC_ARG
EUC_GUL
EUC_MEL
ETR_CRO
GEN_GEN
GEN_LUT
GOB_OCE
HAR_CLU
ISO_PAR
LAG_LAE
LYC_GRO
MAC_ANC
MEN_AME
MIC_FUR
NET_BAR
OLI_SAL
OLI_SAU
PAR_BRAS
PAR_TROP
PEL_HAR
PEP_PAR
POM_COR
PRI_PUN
SCI_LUN
SEL_VOM
SPH_TES
STE_BRA
STE_RAS
SYM_PLA
SYN_FOE
TRIC_LEP
TRIN_MIC
TRIN_PAU
Estádio de Maturação Gonadal
Incubando
ESPÉCIE
ACHI_LIN = Achirus lineatus
ANC_PAR = Anchoa parva
BAG_BAG= Bagre bagre
BAI_RON=Bairdiella ronchus
CAR_LAT=Caranx latus
CAT_SPI=Cathorops spixii
CHAE_FAB=Chaetodipterus faber
CHL_CRY=Chloroscombrus crysurus
CHI_SPI=Chilomycterus spinosus
CIT_ARE=Citharichtys arenaceus
CIT_SPIL=Citharichthys spilopterus
CTE_GRA=Ctenosciaena racillicirrhus
CYN_ACO=Cynoscion acoupa
CYN_LEI=Cynoscion leiarchus
CYN_MIC=Cynoscion microlepidotus
CYN_VIR=Cynoscion virescens
DIA_RHO=Diapterus rhombeus
DIP_RAD=Diplectrum radiale
EUC_ARG=Eucinostomus argenteus
EUC_GUL=E. gula
EUC_MEL=E. melanopterus
ETR_CRO=Etropus crossotus
GEN_GEN=Genidens genidens
GEN_LUT=Genyatremus luteus
GOB_OCE=Gobionellus ocenanicus
HAR_CLU=Harengula clupeola
ISO_PAR=Isopisthus parvipinnis
LAG_LAE=Lagocephalus laevigatus
LYC_GRO=Lycengraulis grossidens
MAC_ANCY=Macrodon ancylodon
MEN_AME=Menticirrhus americanus
MIC_FUR=micropogonias furnieri
NET_BAR=Netuma barba
OLI_SAL=Oligoplithes saliens
OLI_SAL=Oligoplithes saurus
PAR_BRAS=Paralonchurus brasiliensis
PAR_TRO=Paralichthys tropicus
PEL_HAR=Pellona harrowerii
PEP_PAR=Peprilus paru
POM_COR=Pomadasys corvinaeformis
PRI_PUN=Prionotus punctatus
SCI_LUN=Sciadeichthys luniscutis
SEL_VOM=Selene vomer
SPH_TES=Sphoeroides testudineus
STE_BRA=Stellifer brasiliensis
STE_RAS=Stellifer rastrifer
SYM_PLA=Symphurus plagusia
SYN_FOE=Synodus foetens
TRIC_LEP=Trichiurus lepturus
TRIN_MIC=Trinectes microphtalmus
TRIN_PAU=Trinectes paulistanus
Figura 37. a) Histograma das freqüências de distribuição do número de espécies
capturadas por diferentes estádios de maturação gonadal e b) diagrama do número de
exemplares capturados nos diferentes estádios de maturação gonadal por espécie.
67
apresentaram números de exemplares imaturos superiores a 2000, Sciadeichthys luniscutis
entre 1006 e 1509, Micropogonias furnieri e Netuma barba entre 503 e 1006 (fig. 37b).
Além disto as últimas também foram capturadas em todos os estádios de maturação
gonadal.
Isto permite concluir que estas espécies utilizaram a área como local de
recrutamento, crescimento e reprodução. Ainda, os machos C. spixii, G. genidens,
Sciadeichtys luniscutis e N. barba utilizaram a área para a incubação oral de ovos e larvas
(fig.37b).
Já as reduções, no número de exemplares e de espécies, observadas nos demais
estádios possivelmente estejam relacionadas ao fato de que a Baía de Guaraqueçaba pode
representar o limite do "continuum" reprodutivo e de distribuição das populações destas
espécies, cujo centro pode estar situado nas áreas adjacentes (fig.37b).
A intensidade reprodutiva (freqüência absoluta do número exemplares adultos e que
efetivamente participaram do processo reprodutivo) das 10 espécies mais abundantes, nos
meses e pontos amostrais, é mostrada respectivamente nas figuras 38 e 39. Foi observada
atividade reprodutiva: a) incipiente (n<=50 exemplares) para Citharichthys arenaceus,
Micropogonias furnieri, Etropus crossotus e Netuma barba, b) moderada (n acima de 50 e
menor ou igual a 200 exemplares) para Sciadeichthys luniscutis e Genidens genidens; c)
intensa (n acima de 200 e menor ou igual a 500 exemplares) para Isopisthus parvipinnis e
Sphoeroides testudineus e d) muito intensa (n>500 exemplares) para Cathorops spixii e
Stellifer rastrifer. De modo geral a atividade reprodutiva foi muito intensa no período
compreendido entre a primavera e final do verão. No outono foi intensa apenas Stellifer
rastrifer e moderada para Isopisthus parvipinnis e Cathorops spixii. Stellifer rastrifer
apresentou os maiores períodos de atividade reprodutiva intensa e muito intensa (entre
setembro e abril) seguida por C. spixii (entre agosto e novembro) e por Sphoeroides
testudineus (em setembro, outubro e em dezembro).
68
A atividade reprodutiva foi incipiente em todos os pontos amostrais para as
espécies Etropus crossotus, Citharichthys arenaceus, Micropogonias furnieri e Netuma
barba. Foi incipiente nos pontos 1,2,3 e moderada no 4 para Sciadeichthys luniscutis.
Mostrou-se moderada em todos os pontos amostrais para Isopisthus parvipinnis, nos
pontos 1, 3 e 4 para Sphoeroides testudineus e somente no ponto 4 e 1 respectivamente
para Sciadeichthys luniscutis e Genidens genidens. A atividade foi intensa no ponto 2 para
Sphoeroides testudineus, no ponto 3 para Genidens genidens e em todos para Stellifer
rastrifer. Cathorops spixii apresentou atividade reprodutiva intensa nos pontos 2 e 3 e
muito intensa nos pontos 1 e 4.
Nas figuras 40 a 45 foram plotados simultaneamente para as espécies com atividade
reprodutiva moderada, intensa e muito intensa, o número de exemplares por estádio de
maturação gonadal como descritores e os meses e pontos de amostragem como objetos nos
planos fatoriais da ACP. Em todas as espécies a análise restringiu-se aos dois primeiros
componentes que explicaram para:
1. Cathorops spixii respectivamente 39,39 e 17,89 % da variância total dos dados.
Os dois primeiros componentes associados explicaram 57,28% da variabilidade
(tab. 52). Para a formação do componente 1 contribuíram em maior grau, na
coordenada negativa, os estádios em maturação, maduro, reprodução e, em
menor grau, o estádio desovado ou esgotado (fig. 40). Na coordenada positiva
contribuíram, porém em menor grau, os estádios imaturo e incubando ovos
(fig.40). Para a formação do componente 2 contribuiu na coordenada positiva o
estádio imaturo e, na coordenada negativa, os estádios desovado e incubando
ovos.
69
JUL
JUN
MAI
ABR
MAR
Meses
FEV
JAN
DEZ
NOV
OUT
SET
CAT_SPI
SPH_TES
STE_RAS
ISO_PAR
GEN_GEN
SCI_LUN
NET_BAR
ETR_CRO
MIC_FUR
CIT_ARE
AGO
0,796
Ausente
1,591
Incipiente
2,386
Moderada
3,182
Intensa
3,978
Muito Intensa
Espécies
Figura 38. Diagrama da atividade reprodutiva mensal das espécies mais freqüentes. Onde
ausente = 0; Incipiente <=50; Moderada >50 e <=200; Intensa >200 e <=500 e Muito
Intensa > 500 exemplares.
4
2
CAT_SPI
STE_RAS
GEN_GEN
SPH_TES
ISO_PAR
SCI_LUN
NET_BAR
MIC_FUR
CIT_ARE
1
ETR_CRO
Pontos Amostrais
3
Ausente
0,5
2Incipiente
Moderada
3,5
5Intensa
Muito Intensa
6,5
Espécies
Figura 39. Diagrama da atividade reprodutiva espacial das espécies mais freqüentes.
Onde ausente = 0; Incipiente <=50; Moderada >50 e <=200; Intensa >200 e <=500 e
Muito Intensa > 500 exemplares.
70
Apresentaram correlação fortemente negativa com o componente 1 e fracamente
positiva com o componente 2, em ordem decrescente de importância para o
componente 1, os meses de outubro e setembro no ponto 1, outubro e agosto no
ponto 4 e outubro nos pontos 3 e 2. O mês de novembro no ponto 1 apresentou
correlação fortemente negativa tanto com o componente 1 quanto com o 2 (fig.
40).
Correlacionaram-se com os componentes 1 e 2, respectivamente nas
coordenadas positiva e negativa, os meses de abril nos pontos 2 e 4 e dezembro
e fevereiro no ponto 1. Estes resultados mostraram a dinâmica temporal e
espacial do processo reprodutivo. As freqüências de exemplares em maturação
aumentaram do mês de agosto (no ponto 4) para o mês de setembro (no ponto1),
atingindo seu ápice em outubro (no ponto 1) (fig.40).
4
Componente 2
Im a t u r o
Abr
Abr
2
Maturação
Out
Maduro
Reprodução
0
Set Ago
O uOt u t
O u t
Jan
Fev
Jan
Jan
Nov Dez
Pontos
amostrais
-2
Fev
1
2
3
4
Nov
Desovado
-4
-8.3
-6.3
-4.3
-2.3
-0.3
Componente 1
D e z
Incubando
1.7
Figura 40. Diagrama da distribuição das freqüências por estádios de maturação gonadal
de Cathorops spixii (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
71
A reprodução ocorreu em processo crescente nos mesmos meses e pontos, com o
ápice também no mês de outubro (no ponto 1), e decrescente até novembro (no
ponto 1). Neste último mês e ponto amostral o número de exemplares desovados
ou esgotados atingiu o seu ápice quando iniciou-se simultaneamente o processo
de incubação oral dos ovos. Em dezembro (no ponto1) a maioria dos exemplares
encontrava-se incubando ovos, processo que estendeu-se até o mês de fevereiro
(também no ponto1). O último mês e ponto correspondeu à incubação dos ovos
dos exemplares que reproduziram-se mais tardiamente. Finalmente, o
recrutamento, iniciado no mês de dezembro (no ponto1), atingiu o seu pico
máximo em abril (nos pontos 4 e 2) (fig.40). É importante destacar que o
processo reprodutivo ocorreu de modo mais intenso no ponto amostral 1 e o
recrutamento nos pontos amostrais 2 e 4. Isto evidenciou que a espécie ocupou
áreas diferenciadas nas diferentes fases do ciclo reprodutivo. Ainda, estes
resultados refletiram fielmente e corroboraram a hipótese formulada
anteriormente de que distribuição das classes de tamanho e peso observadas na
figura 25 estavam relacionadas principalmente a atividade reprodutiva. Além
disso, evidenciaram que os picos de freqüência de exemplares imaturos,
maduros, em reprodução e desovados mostrados nas figuras 36a, b e c refletem
principalmente, por sua abundância, a dinâmica reprodutiva de Cathorops spixii.
2) Genidens genidens respectivamente 60,82 e 16,71% da variabilidade total dos
dados. Os dois primeiros componentes explicaram juntos 77,53% da variância
(tab.52). Contribuíram positivamente para a formação do componente 1 todos
os estádios de maturação gonadal, com o destaque para o estádio desovado ou
esgotado (fig.41). Para a formação do componente 2 contribuíram
72
positivamente, em ordem decrescente de importância, incubação dos ovos,
reprodução e maduro. Contribuíram negativamente, em ordem decrescente de
importância para a formação do mesmo eixo, os estádios imaturo, em
maturação e de descanso reprodutivo (fig.41). Os resultados mostraram que a
maturação das gônadas inicia-se no mês de setembro no ponto 3, atingindo seu
ápice m novembro também no ponto 3. Neste último mês, mas no ponto 1,
ocorreram simultaneamente exemplares desovados, maduros e em reprodução.
Isto permite concluir que o processo reprodutivo ocorreu de modo muito rápido.
A reprodução e incubação dos ovos atingiram seus ápices em dezembro (no
ponto 3) e diminuiram posteriormente em janeiro (no ponto3). Em abril (nos
pontos 3, 2 e 4) aumentou o número de exemplares imaturos. Contudo, os
últimos ocorreram em maiores freqüências no mês de setembro no ponto 3.
4.1
Incubando
Dez
Reprodução
Componente 2
2.1
Jan
Maduro
Nov
0.1
-1.9
Desova
Dez
AbrOut
Abr
Abr Abr
Nov
Descanso
Pontos
amostrais
?1
?2
?3
?4
Maturação
-3.9
Set
Imaturo
-5.9
-1
1
3
5
Componente 1
7
9
11
Figura 41. Diagrama da distribuição das freqüências por estádios de maturação gonadal
de Genidens genidens (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
73
Possivelmente estes exemplares correspondiam aos recrutas do ano anterior. A
distribuição observada espacial e temporal da atividade reprodutiva coincidiu e
explicou a distribuição temporal e espacial dos intervalos de comprimento total e
peso observadas na figura 28. Isto é o aumento no número de exemplares das
maiores classes de CT e de peso nos meses de novembro e dezembro nos pontos
3 e das menores nos meses de setembro (no ponto 3) e abril (nos pontos 2 e 4).
Corroboraram ainda as observações de que as distribuições espaciais e temporais
refletem a atividade reprodutiva.
3) Isopisthus parvipinnis respectivamente 33,77 e 20,70 % da variância total dos
dados (tab.52). Juntos os dois primeiros componentes explicaram 54,47% da
variabilidade (tab.52). Contribuíram para a formação do componente 1 e 2,
simultânea e positivamente, por ordem decrescente de contribuição, os estádios
maduro, desovado (ou esgotados) e em reprodução (fig. 42). Contribuíram
positivamente para o componente 1 e negativamente para o 2, por ordem
decrescente de contribuição, os estádios descanso, maturação e imaturo.
Correlacionaram-se positivamente ao componente 1 os meses de março
(ponto1), setembro (ponto 3 e 4), maio (ponto2) e, em menor grau, abril (ponto
1), novembro (ponto3) e dezembro (ponto2) (fig. 42). Os resultados mostraram
que a maturação das gônadas inicia-se me setembro (nos pontos 3 e 4) e a
reprodução estendeu-se e atingiu o ápice em março (no ponto 1) (fig.42). Os
exemplares desovados (ou esgotados) ocorreram em baixas freqüência à partir
de novembro (ponto3) e dezembro (ponto2), atingindo máximas freqüências
também em março (ponto1) (fig.42).
74
5.5
Reprodução
Componente 2
3.5
Mar
Pontos
amostrais
?1
?2
?3
?4
Maduro
Nov Desovado
Dez
Mai
Nov
Fev
Nov Imaturo
Ago
Abr
Set
1.5
-0.5
Set
-2.5
Descanso
Maturação
-4.5
-0.7
0.3
1.3
2.3
3.3
Componente 1
Set
4.3
5.3
Figura 42. Diagrama da distribuição das freqüências por estádios de maturação gonadal
de Isopisthus parvipinnis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
Finalmente em abril (ponto1) foram capturados os exemplares imaturos. Como
para as espécies anteriores, os resultados das distribuições das freqüências por
intervalos de classe de CT e de peso mostradas na figura 29 explicaram e
refletiram a estratificação dos exemplares em acordo com a atividade
reprodutiva. Isto é a agregação dos maiores exemplares em determinados
pontos amostrais para a realização da reprodução, surgindo gradativamente, nos
meses posteriores, exemplares da menor classe de CT que correspondiam aos
juvenis originados da conclusão do ciclo reprodutivo.
75
4) Sciadeichthys luniscutis respectivamente 35,56 e 18,97 % da variância total dos
dados (tab.52). Juntos os dois primeiros componentes explicaram 54,53% da
variabilidade (tab.52). Contribuíram negativamente para a formação do
componente 1, por ordem decrescente de contribuição, os estádios reprodução,
incubação dos ovos, maduro e desovado (fig.43). Para o componente 2
contribuiu positivamente o estádio imaturo e negativamente o estádio descanso
e em maturação (fig. 43).
Correlacionaram-se em ordem decrescente ao
componente 1 os meses de novembro (ponto4), setembro (ponto4) e dezembro
(ponto 3) (fig.43). Correlacionaram-se positivamente ao componente 2, por
ordem decrescente, os meses de abril (ponto 3), junho (ponto 3), março
(ponto3), maio (ponto3), março (ponto 4) e maio (ponto1) (fig.43).
Correlacionaram-se negativamente ao mesmo componente, em maior grau, os
meses de outubro (ponto 4) e dezembro (ponto 1) (fig.43).
3.6
Nov
Componente 2
1.6
Maduro
Incubando
Set
Dez
Reprodução
Mai
Dez
Abr
Fev
Mar
-0.4
Pontos
amostrais
?1
?2
?3
?4
-2.4
Desovado
Dez
Descanso
Out
-8
Out
Nov
Jan
Mar
Maturação
-4.4
-10
Imaturo
Abr
Jun
Mar
Mai
Mar
Mai
Jun
-6
-4
-2
0
2
Componente 1
Figura 43. Diagrama da distribuição das freqüências por estádios de maturação gonadal
de Sciadeichtys luniscutis (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
76
Os resultados demonstraram que a atividade reprodutiva da espécie iniciou-se
em outubro no ponto 4 (maturação, reprodução e desova) e atingiu seu ápice
em novembro também no ponto 4 (reprodução e incubação dos ovos).
Finalmente em março (no ponto3) o número de recrutas começou a aumentar
até atingir o valor máximo em abril no ponto 3, decrescendo gradativamente
nos meses posteriores (maio e junho nos pontos 3 e 4). A distribuição dos
intervalos de CT e peso podem ser observadas na figura 32 e refletiram a
atividade reprodutiva (fig.43). Ainda demonstraram que os picos de freqüência
de exemplares imaturos, maduros, em reprodução e desovados mostrados nas
figuras 36a, b e c, também refletiram a dinâmica reprodutiva de Sciadeichthys
luniscutis.
5) Sphoeroides testudineus respectivamente 39,88 e 25,93 % da variância total
dos dados (tab.52). Juntos os dois primeiros componentes explicaram 65,81%
da variabilidade (tab. 52). Contribuíram positivamente para a formação dos
dois primeiros componentes os estádios desovado, maturação, reprodução e
maduro e, e negativamente apenas para o componente dois, o desovado e o
descanso
reprodutivo
(fig.44).
Correlacionaram-se
positivamente
aos
componente 1 os meses de outubro (ponto2), dezembro (ponto 4), março
(ponto2) e, em menor grau, setembro (ponto 3), dezembro (ponto2) e julho
(pontos 2 e 4) (fig.44). A distribuição observada mostrou que a atividade
reprodutiva iniciou-se em setembro (nos pontos 3 e 4) e culminou em outubro
(apenas no ponto 2). Em dezembro (no ponto 4) o número de exemplares
desovados (ou esgotados) atingiu o valor máximo (fig.44). Os exemplares
imaturos foram observados em baixas freqüências no mês de junho, julho e
77
março. Isto permite concluir que a espécie não recruta na área examinada. Do
mesmo modo que o descrito para as espécies anteriores as distribuições
temporal e espacial das classes de CT e de peso observadas na figura 33
representaram a estrutura da espécie durante o ciclo reprodutivo.
6
Maduro
Reprodução
Out
4
Set
Componente 2
2
Set
Maturação
FevJanSet Dez
Jun Imaturo
Mai
Abr
Jul
Jan
0
-2
Dez
Desovado
Pontos
amostrais
1
2
3
4
-4
Descanso
Mar
3
5
-6
-1
1
7
Componente 1
Figura 44. Diagrama da distribuição das freqüências por estádios de maturação gonadal
de Sphoeroides testudineus (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
6) Stellifer rastrifer respectivamente 39,28 e 24,47 % da variância total dos dados
(Tab.52). Associados os dois primeiros componentes explicaram 63,75 % da
variabilidade (Tab. 52). Contribuiu positivamente para formação do
componente 1 o estádio imaturo e negativamente todos os demais estádios (fig.
45). Para a formação do componente 2 contribuíram positivamente os estádios
imaturo, desovado (ou esgotado) e reprodução (fig.45). Correlacionaram-se
positivamente ao componente 1, em ordem decrescente de importância, os
meses de abril, janeiro, dezembro e fevereiro ( todos no ponto 4) e
78
negativamente os meses de janeiro (ponto2), novembro (ponto 1) e setembro
(ponto 3) (fig.45).
5.3
Pontos
amostrais
?1
?2
?3
?4
4.3
Componente 2
3.3
Abr
Imaturo
Jan
2.3
Desovado
Jan
Nov
1.2
Reprodução
Descanso
Maduro
Set
Maturação
0.3
Jan
Fev
Fev
Out
Set
Abr
Fev
Dez
Mar
Mai
-0.7
-5.3
-3.3
-1.3
0.7
Componente 1
2.7
4.7
Figura 45. Diagrama da distribuição das freqüências por estádios de maturação gonadal
de Stellifer rastrifer (descritores) nos meses e pontos de amostragem (objetos),
sobrepostos no plano bidimensional dos componentes principais. Demais meses e pontos
encontram-se sobrepostos e indicados por uma seta.
A distribuição observada mostrou que a espécie iniciou a atividade reprodutiva em
setembro (principalmente no ponto 3), com as gônadas tornando-se maduras nos
meses subseqüentes até a reprodução, que foi observada com maior intensidade no
mês de novembro (no ponto 1). Neste mesmo mês surgiram os exemplares
desovados (ou esgotados) que aumentaram em número até o mês de janeiro (no
ponto 2). Em dezembro, janeiro e fevereiro (todos no ponto 4) começou a aumentar
o número de exemplares juvenis que atingiu o valor máximo em abril (no ponto 4).
Como já observado para as espécies anteriores os resultados da distribuição
espacial e temporal das classes de CT e de peso observados na figura 34
demonstraram como estava estruturada a população no ciclo reprodutivo (fig.45).
79
Nas figuras 46 e 47 as curvas de ajustamento (juntamente com os tamanhos da
amostra e o coeficiente de correlação), das estimativas do comprimento total da primeira
maturação, obtidas com base nas freqüências de adultos das populações naturais das
espécies de peixes (com reprodução moderada à muito intensa), presentes na Baía de
Guaraqueçaba. Foram estimados os CT médios de: a) 11,70 cm para Cathorops spixii, b)
13,99 cm para Genidens genidens, c) 14,67 cm para Isopisthus parvipinnis, d) 31 cm para
Sciadeichthys luniscutis, e) 15,99 cm para Sphoeroides testudineus e f) 10,80 cm para
Stellifer rastrifer.
80
Cathorops spixii
a
1,0
Freqüência relativa de adultos
0,9
y= 1 - e
0,8
-6,27
-11
.x
8,06
n=2460
0,7
r=0,9830
0,6
CT 50% =11,70
0,5
0,4
0,3
0,2
0,1
0,0
0
2
4
6
8
10
12
14
16
18
20
22
24
26
28
Comprimento Total (cm)
Genidens genidens
b
1,0
Freqüência relativa de adultos
0,9
0,8
y=1-e
0,7
n = 293
-2,59
-8
.X
5,61
r = 0,9729
0,6
CT50% = 13,99
0,5
0,4
0,3
0,2
0,1
0,0
0
5
10
15
20
25
30
35
Comprimento Total (cm)
Isopisthus parvipinnis
c
1,0
-6,845
y= 1 - e
.X
5,15
n=99
0,8
Freqüência relativa de adultos
-07
r = 0,871369
CT
50%
= 14,67
0,6
0,4
0,2
0,0
0
2
4
6
8
10
12
14
16
18
20
22
24
Comprimento Total (cm)
Figura 46. Distribuição das freqüências relativas acumuladas de exemplares adultos por
pontos médios de intervalos de classe de CT(cm) nas amostras de a) Cathorops spixii, b)
Genidens genidens e c) Isopisthus parvipinnis obtidas na Baía de Guaraqueçaba.
Fonte:dados de campo.
81
Sciadeichthys luniscutis
a
Freqüência relativa de adultos
1,0
-1,692
0,9
y=1-e
0,8
n=76
r = 0,90
0,7
CT 50% = 31,00
-5
.X
3,093
0,6
0,5
0,4
0,3
0,2
0,1
0,0
0
10
20
30
40
50
60
70
80
Comprimento Total (cm)
Sphoeroides testudineus
b
1,0
0,9
y=1-e
Freqüência relativa de adultos
0,8
-7,43
-12
.X
9,11
n=397
r = 0,97
0,7
CT 50% = 15,99
0,6
0,5
0,4
0,3
0,2
0,1
0,0
0
2
4
6
8
10
12
14
16
18
20
22
24
Comprimento Total (cm)
Stellifer rastrifer
c
1,0
y=1-e
0,9
-7
X
5,43
n=1158
0,8
Freqüência relativa de adultos
-6,67
r=0,9116
CT 50%=10,80
0,7
0,6
0,5
0,4
0,3
0,2
0,1
0,0
0
4
8
12
16
20
24
28
Comprimento Total (cm)
Figura 47. Distribuição das freqüências relativas acumuladas de exemplares adultos por
pontos médios de intervalos de classe de CT(cm) nas amostras de a) Sciadeichthys
luniscutis, b) Sphoeroides testudineus e c) Stellifer rastrifer obtidas na Baía de
Guaraqueçaba. Fonte:dados de campo.
82
4.2.4. Comportamento frente a salinidade
Na figura 48 são mostradas as distribuições de freqüências de adultos e juvenis em
relação às salinidades em que foram capturados.
25
25
20
20
JUVENIS
30
Salinidade fundo (‰) JUVENIS
30
15
15
10
5
5
0
30
0
30
25
25
20
20
ADULTOS
ADULTOS
10
15
15
10
10
Média±s
Média±e
Média
5
5
0
0
CYN_MIC CYN_VIR DIA_ROM DIP_RAD EUC_ARGEUC_GUL EUC_MELETR_CROGEN_GENGEN_LUTGOB_OCEH AR_CLU ISO_PAR LAG_LAELYC_GRO
30
25
25
20
20
JUVENIS
30
15
15
10
10
5
5
0
30
0
30
25
25
20
20
ADULTOS
ADULTOS
JUVENIS
ACH_LIN ANC_PAR BAG_BAG BAI_RON CAR_LAT CAT_SPI CET_EDE CHA_FAB CHL_CRY CHI_SPI CIT_ARE CIT_SPI CTE_GRA CYN_ACO CYN_LEI
15
10
5
15
10
5
0
0
MAC_ANC MEN_AME MIC_FUR NET_BAR
OLI_SAL
OLI_SAU PAR_BRAS PAR_TROP PEL_HAR
ACHI_LIN = Achirus lineatus
ANC_PAR = Anchoa parva
BAG_BAG= Bagre bagre
BAI_RON=Bairdiella ronchus
CAR_LAT=Caranx latus
CAT_SPI=Cathorops spixii
CHA_FAB=Chaetodipterus faber
CHL_CRY=Chloroscombrus crysurus
CHI_SPI=Chilomycterus spinosus
CET_EDE=Cetengraulis edentulus
CIT_ARE=Citharichtys arenaceus
CIT_SPIL=Citharichthys spilopterus
CTE_GRA=Ctenosciaena gracillicirrhus
CYN_ACO=Cynoscion acoupa
CYN_LEI=Cynoscion leiarchus
CYN_MIC=Cynoscion microlepidotus
CYN_VIR=Cynoscion virescens
DIA_RHO=Diapterus rhombeus
PEP_PAR
POM_COR
PRI_PUN SCI_LUN SEL_VOM SPH_TES STE_BRA STE_RAS SYM_PLA SYN_FOE TRIC_LEP TRIN_MIC TRIN_PAU
DIP_RAD=Diplectrum radiale
EUC_ARG=Eucinostomus argenteus
EUC_GUL=E. gula
EUC_MEL=E. melanopterus
ETR_CRO=Etropus crossotus
GEN_GEN=Genidens genidens
GEN_LUT=Genyatremus luteus
GOB_OCE=Gobionellus ocenanicus
HAR_CLU=Harengula clupeola
ISO_PAR=Isopisthus parvipinnis
LAG_LAE=Lagocephalus laevigatus
LYC_GRO=Lycengraulis grossidens
MAC_ANCY=Macrodon ancylodon
MEN_AME=Menticirrhus americanus
MIC_FUR=micropogonias furnieri
NET_BAR=Netuma barba
OLI_SAL=Oligoplites saliens
OLI_SAL=Oligoplites saurus
PAR_BRAS=Paralonchurus brasiliensis
PAR_TRO=Paralichthys tropicus
PEL_HAR=Pellona harrowerii
PEP_PAR=Peprilus paru
POM_COR=Pomadasys corvinaeformis
PRI_PUN=Prionotus punctatus
SCI_LUN=Sciadeichthys luniscutis
SEL_VOM=Selene vomer
SPH_TES=Sphoeroides testudineus
STE_BRA=Stellifer brasiliensis
STE_RAS=Stellifer rastrifer
SYM_PLA=Symphurus plagusia
SYN_FOE=Synodus foetens
TRIC_LEP=Trichiurus lepturus
TRIN_MIC=Trinectes microphtalmus
TRIN_PAU=Trinectes paulistanus
Figura 48. Diagrama comparativo das distribuições das freqüências de ocorrência de adultos e juvenis das
diferentes espécies, em função da salinidade da água de profundidade em que foram capturados. Onde "s" =
desvio e "e"= erro padrões das estimativas.
83
Com base nestes resultados as espécies foram agrupadas e foi possível observar dois
grupos principais com várias subdivisões.
1) Espécies que ocorreram nas capturas realizadas em salinidades situadas entre 10 e
25‰:
a) simultaneamente como adultos e juvenis Achirus lineatus, Bairdiella ronchus,
Cathorops spixii, Cetengraulis edentulus, Citharichthys arenaceus, Genidens
genidens,
Gobionellus
oceanicus,
Harengula
clupeola,
Lycengraulis
grossidens, Menticirrhus americanus, Micropogonias furnieri, Netuma barba,
Paralonchurus brasiliensis, Sciadeichthys luniscutis, Sphoeroides testudineus,
Stellifer rastrifer e Symphurus plagusia.
b) somente como juvenis Anchoa parva, Bagre bagre, Chaetodipterus faber,
Chloroscombrus crysurus, Ctenosciaena gracillicirrhus, Cynoscion acoupa, C.
leiarchus, C. microlepidotus, Diapterus rhombeus, Eucinostomus argenteus, E.
gula, E. melanopterus, Geniatremus luteus, Oligoplites saurus , O. salines,
Paralichthys tropicus, Peprilus paru, Prionotus punctatus, Synodus foetens,
Trichiurus lepturus, Trinectes microphtalmus e T. paulistanus.
2) Espécies que ocorreram em salinidades superiores a 25 ‰:
a) simultaneamente como adultos e juvenis C. spilopterus, Etropus crossotus,
Isopisthus parvipinnis, Macrodon ancylodon e Stellifer brasiliensis.
b) somente como juvenis Caranx latus, Chilomycterus
spinosus, Cynoscion
virescens, Diplectrum radiale, Lagocephalus laevigatus, Pellona harrowerii,
Pomadasys corvinaeformis e Selene vomer
84
4.3. IMPORTÂNCIA COMO RECURSO
4.3.1. Das unidades amostrais
Foram entrevistados 46 pescadores com idade média de 45,23 (± 2,24) anos, dos
quais 65,22% eram casados (fig.49a).
Coabitavam com os entrevistados, além da
companheira, em média 3,36 (± 0,33) pessoas, das quais 1,15 (±0,26) também pescava.
Acima de 70% dos entrevistados tem na pesca sua ocupação principal e secundária
(fig.49 e tab.53). Foram constatados aposentados pela pesca (16,13%), mas que
continuavam pescando e as atividades agrícolas, comerciais (bar/mercearia e turismoaluguel de embarcação e guia de pesca), funcionalismo público e de artesanato (canoeiro),
embora presentes mostraram-se irrelevantes (fig. 49 e tab.53).
Acima de 86,95% dos entrevistados declararam realizar a pesca para a subsistência
(consumo próprio ou da família), com a comercialização dos excedentes da produção e
8,69% dos entrevistados, afirmaram realizar a pesca com fins exclusivos de comércio
(fig.50). Foi observado que os entrevistados consomem peixe, em média,
4,54 (± 0,32)
dias na semana.
n=46
Principal
Fun/Pub
Secundária
Pres/Ser
Turis.
Bar/merc
Cano.
Apos.
Agri.
Pesca
0
10
20
30
40
50
60
70
80
90
100
Número de declarantes (%)
Figura 49. Diagrama comparativo das freqüências de declarantes por ocupação principal
e secundária na localidade de Guaraqueçaba. Onde: Fun/Pub= funcionário público,
Pres./Ser. = prestação de serviços, Turis. = turismo, Bar/merc.= bar/mercearia, Cano =
canoeiro, Apos. = aposentado e Agri.= agricultura. Fonte:dados de campo.
85
Utilização do Produto da Pesca
n=46
86,96
Sub./Com.
Comércio
8,69
4,34
Subsistência
0
10
20
30
40
50
60
70
80
90
100
Número de declarantes (%)
Figura 50. Diagrama comparativo das porcentagens de declarantes e a utilização do
produto pesqueiro na localidade de Guaraqueçaba. Onde: Sub./Com. = subsistência e
comércio. Fonte: dados de campo.
4.3.2. Das atividades e equipamentos de pesca.
A percentagem de entrevistados que declararam possuir sua própria embarcação
situou-se acima de 85% (fig. 51a) e, em média, a relação remo/motor por pescador foi de
aproximadamente 1/1 (fig. 51b).
a
n=46
b
t (45)=0,3301 p<0,7429
1,5
Número de Embarcações (freq.absol.)
Embarcação Própria
n=46
89,13
Sim
Não 10,87
0
10
20
30
40
50
60
Número de declarantes
70
80
90
100
±1.96*e
±1.00*e
Média
1,0
0,70
0,60
0,5
0,0
REMO
MOTOR
Figura 51. a) Relação entre o número de pescadores e a posse de embarcações na
localidade de Guaraqueçaba e b) comparação entre as médias do número de embarcações
movidas a remo e a motor para os pescadores da localidade de Guaraqueçaba.Fonte:
dados de campo.
Todas a embarcações eram confeccionadas em madeira e foram constatadas
diferenças significativas entre os comprimentos das embarcações movidas a remo e a
86
motor. As movidas a remo mediam em média 4,68 (±0,633) m e as movidas a motor 7,33
(±0,81)m (fig. 52), as últimas com uma potência média de 11,52 (±1,39) HP.
F(1,56)=43,74 p=0
8,5
±1.96*e
±1.00*e
Comprimento médio (m)
Média
7,33
7,5
6,5
5,5
4,68
4,5
3,5
REMO
MOTOR
Figura 52. Diagrama: a) comparativo das médias dos comprimento das embarcações
movidas a remo e a motor e b) da média da potência dos motores para a localidade de
Guaraqueçaba. Fonte: dados de campo
Na figura 53 encontram-se representadas as comparações das médias dos números
de tarrafas, jerivaus, redes de caceio, fundeio, lanceio, e espinhel por pescador, na
localidade analisada. Em média, os pescadores possuem 0,11 (± 0,32) tarrafas, 1,19 (±
0,86) jerivaus, 1,52 (± 1,60) redes de caceio, 0,93 (± 2,53) de fundeio e 0,09 (± 0,28) de
lanceio. Foram constatados 5 pescadores utilizando o espinhel, 4 praticando o lanceio, 2
utilizando o cerco fixo, e nenhum utilizando o arrasto com portas.
6
n=46
±s
±e
Média
2
Espinhel
Lanceio
Fundeio
Caceio
-2
Jerivau
0
Tarrafa
Freqüência absoluta
4
Figura 53. Diagrama comparativo entre o número médio de petrechos de pesca por
pescador para a localidade de Guaraqueçaba. Fonte: dados de campo.
87
4.3.3. Dos locais e petrechos preferenciais de pesca.
Na tabela 54 encontram-se indicados os locais preferenciais de pesca declarados
por localidades. Os declarantes afirmaram escolher como primeira opção de local de pesca
as regiões situadas próximas à sua própria localidade, restando a segunda e terceira opções
para os locais mais distantes (tab. 54 e fig.54).
Guapicú
B.Pinheiros
B.Guaraqueçaba
Preferência
1 a.
2 a.
I.Peças
3 a.
n=46
B.Tibicanga
I.Rasa
Cananéia
0
40 80 120 160 200 240
Superagüi
Número de Horas
por Mês
I.Cobras
Varadouro
Benito
Itaqui
Tagaçaba
Figura 54. Diagrama mostrando as distribuições dos pescadores, da localidade de
Guaraqueçaba, segundo o número de horas gastos nos locais preferencias de pesca.
Fonte: dados de campo.
Com relação ao número médio de horas mensais utilizados nas fainas de pesca, os
entrevistados gastaram 132,61 (±62,28) pescando nos locais declarados como de primeira
preferência. Nos locais de segunda e terceira opções gastaram, respectivamente, 41,10
(±27,31) e 34,33 (±22,78) horas mensais (fig.55).
88
F(2,60)=17,82 p<0,000001
180
±1.96*e
±1.00*e
Média
160
Número de horas/mês
140
132,61
120
100
80
60
41,10
40
34,33
20
0
1
2
3
Locais/ordem de preferência
Figura 55. Comparação entre o número médio de horas gastos, pelos pescadores de
Guaraqueçaba, nas fainas de pesca nos diferentes locais segundo a ordem de
preferência.
Na figura 58 encontram-se relacionados os aparelhos de pesca e os locais de
utilização.
Guapicú
Tarrafa
B.Pinheiros
Jerivau
B.Guaraqueçaba
Caceio
I.Peças
Fundeio
Lanceio
Espinhel
B.Tibicanga
I.Rasa
n=46
Cananéia
0
Número de
5 10 15 20 25 30 35 40 Pescadores
Superagüi
I.Cobras
Varadouro
Benito
Itaqui
Tagaçaba
Figura 56. Diagrama das distribuições dos pescadores, da localidade de Guaraqueçaba,
segundo os petrechos de pesca e locais onde são utilizados. Fonte: dados de campo.
Foram declarados como petrechos preferencialmente utilizados na própria Baía de
Guaraqueçaba, relacionados por ordem decrescente de importância, o jerivau, o caceio, o
fundeio e a tarrafa. Como já citado anteriormente, o jerivau é utilizado na captura de
89
camarões e o caceio e o lanceio na captura de peixes demersais e pelágicos. Portanto, a
maioria dos pescadores procura capturar o camarão.
4.3.4. Da ictiofauna.
Na figura 57 encontram-se representados os resultados da comparação dos graus de
abundância declaradas pelos entrevistados (ausente, pouco freqüente e freqüente), como
descritores nos planos fatoriais da ACP, para as 53 espécies de peixes (como observações)
freqüentemente comercializadas no Mercado Municipal de Paranaguá. Foram escolhidos
apenas os dois primeiros componentes que juntos explicam 99,99 % da variação total nos
dados (tab. 55).
Foram declaradas pelos pescadores, por ordem decrescente de freqüência,
Cathorops spixii (Ariidae), Harengula clupeola (Clupeidae), Genidens genidens (Ariidae) ,
Bairdiella rochus (Sciaenidae), Mugil curema (Mugilidae), Sardinella brasiliensis
(Clupeidae), Sciadeichthyes luniscutis (Ariidae), Netuma barba (Ariidae), Eucinostomus
spp (Gerreidae), Opisthonema oglinum (Clupeidae), Oligoplites spp (Carangidae) e Mugil
gaimardianus (Mugilidae) (Fig. 57). Stellifer rastrifer (Sciaenidae), a segunda espécie
mais freqüente nos arrastos (tab.31 e Fig. 12), não foi incluída nos questionários por
raramente ser comercializada.
90
4,2
freqüentes
1
componente 2
2,2
2
46
51 52 53
45
47
44
14
43 42 48 50 25-E
4 5
26-T
27-P
49
13
28-L
40 39 41
29-C
7 12 15
30-E
38
31-S
8 11
37
32-C
17 29 33 34
33-B
36
34-E
9 16
28
18
35-P
35
10
36-E
19 27
37-E
32
20
38-E
26
39-N
21 22
40-S
31
41-C
42-C
24
43-C
23
30
44-T
45-R
25
46-S
3
0,2
-1,8
-3,8
6
NG RAU LIDA E
RIC HIURUS LEPTU RUS
AR ALICH TH Y S S PP
O BO TES SU RINA MENS IS
Y NO SC IO N JAM AISC ENC IS
PINEPHELUS NIVEA TUS
1-CATHO RO PS S PIXII
2-HARENGU LA CLU PEO LA
3-GENID ENS G ENIDENS
4-BAIRD IELLA RO NCH US
5-MUG IL CUR EMA
6-SARDINELLA BRA S ILIENSIS
7-SCIADEICH TH YS LU NISC U TIS
8-OLIG O PLITES S PP
9-MUG IL PLA TANUS
10-PO GO NIAS C RO MIS
11-MUG IL G AIMAR DIANU S
12-NETU MA BARBA
13-EUC INOS TO MUS SPP
14-OPIS THO NEMA O G LINUM
15- MENTIC IRRHU S AMEIC A NU S
16-CY NO SC IO N LEIAR CHU S
17-GEO PH AG US BRAS IIENSIS
18-CY NO SC IO N AC OU PA
19-MAC RO DO N A NCY LO DO N
20-MUG IL LIZA
21-MIC R OPO GO N IA S F URNIERI
22-TR AC HIN O TUS C A RO LINUS
23GENYATR EMU S LUTEUS
24-CE NTRO PO MU S UNDEC IMALIS
-3,4
ausentes
C O MBER J APO NIC US
A RA NX LA TUS
AG R E BA G RE
PINEPHELUS G U AZA
O LY DACTY LUS VIRG INIC U S
US ELACH II
PINEPHELUS MIC ROL EPIS
UG ERRES BRASIL IA NUS
EBRIS MIC R OPS
pouco
freqüentes
-1,4
0,6
PHY R AENA PIC UD ILLA
O NO DO N NO BILIS
H IR O CEN TRO DO N BLEEKER IA NUS
H LO RO S CO MBR US CR YS URU S
RA C HINO TUS F ALC A TUS
ACH YCENTRO N C ANA DUS
C O MBER O MO RU S BR ASILIENS IS
47-ANIS OTR EMU S SUR INA M ENSIS
48-ARCH O SA RG US PRO BATO C EPHALU S
49-CY NO SC IO N MIC RO LEPIDO TUS
50-CA RA NX H IPPO S
51-LU TJ ANUS AN ALIS
52-PAR ALO NC HURU S BR ASILIENS IS
53-PEPR ILU S PA RU
2,6
4,6
componente 1
Figura 57. Plotagem simultânea e ordenação, no espaço dos componentes principais
(ACP), dos descritores (abundância declarada dos recursos na localidade de
Guaraqueçaba) segundo as observações (espécies ictiícas). Fonte: dados de campo.
91
5. DISCUSSÃO
5.1. PARÂMETROS FÍSICO-QUÍMICOS E METEOROLÓGICOS.
A maior pluviosidade nos meses compreendidos entre o final da primavera e verão
coincidiram com o padrão citado por Knoppers et al. (1987), para o sistema estuarino da
Baía de Paranaguá.
O incremento, não significativo, das temperaturas do ar e da água entre os pontos 1 e 3
e redução no 4(fig.5b e 6b) pode ser atribuída a maior profundidade dos pontos 1 e 4.
Também ao fato de que as amostragens no ponto 1 e 4 foram sempre realizadas em
períodos de menor temperatura, ou seja início da manhã e as do ponto 4 no final da tarde.
A estratificação vertical da salinidade na coluna d'água em todos os meses do ano,
com maior intensidade nos de maior pluviosidade (tab.4 e 7 e fig.3 e 6c), assim como as
menores médias de salinidade para a água de superfície (fig. 6c), coincidiram com o
comportamento típico das águas estuarinas. Nestas a água salgada, de maior densidade,
procedente do oceano adjacente, penetra próxima do fundo enquanto que a doce, de menor
densidade e procedente dos rios e das chuvas, escoa próxima da superfície (McLusky
1974). A redução significativa da média do ponto amostral 1 (furo de Guaraqueçaba) para
o 3 (no delta formado pela embocadura dos rios Morato, Moratinho e Ipanema) e elevação
no ponto 4 (tab.8 e fig.6d), o último com salinidade média intermediária às observadas nos
pontos 2 e 3 (tab.8 e fig. 6d), estão associados à diluição gradual da água salina (nos
pontos intermediários 2 e 4), procedente do ponto amostral 1, pelo escoamento da água
dos rios situados próximos ao ponto amostral 3.
As maiores médias de percentagem de saturação de oxigênio foram observadas
próximas do fundo na maioria dos meses de amostragem, com uma notável exceção para o
mês de abril (fig. 6e). Estes resultados provavelmente estão relacionados à lâmina de água
salgada, com maior teor de oxigênio, procedente do oceano adjacente.
92
Com relação à inversão observada em abril e à forte redução na água superficial do ponto
2, provavelmente estão relacionadas, como já constatado por Machado et al. e Brandini,
aos processos de produção e respiração nos sedimentos superficiais e na coluna d'água
(mais acentuados nos meses de maior insolação e pluviosidade) e, principalmente com
relação ao ponto 2, ao aporte de matéria orgânica proveniente da cidade de Guaraqueçaba
(Machado? comunicação pessoal).
Os períodos de alta pluviosidade e temperatura, associados aos meses de verão,
destacaram-se dentre os parâmetros analisados na região. O primeiro mostrou ser o
principal responsável pela diminuição da transparência e salinidade da água. Estas
alterações foram observadas com maior intensidade nos pontos mais internos (2, 3 e 4) e
podem estar relacionadas principalmente com o aumento da descarga de água doce
proveniente dos rios e intensificadas pela posição e configuração geográfica dos pontos
amostrais analisados na Baía de Guaraqueçaba. Estes resultados coincidiram aos
observados por Knoppers et. al (1987) e Machado et. al (no prelo) que afirmaram ser a
hidrodinâmica do sistema da Baía de Paranaguá fortemente influenciada pelos fatores
climáticos (regime de chuvas e ventos), os quais regulam a intensidade das marés.
Coincidiram ainda, com observações dos autores citados,
a distribuição espacial de
temperatura da água, apresentando um padrão semelhante ao da salinidade, e a
transparência, exibindo relação direta com a salinidade e com os maiores valores
ocorrendo no período seco de inverno.
?
E.C. Machado – CEM/UFPR.
93
5.2. COMPOSIÇÃO, VARIABILIDADE TEMPORAL E DISTRIBUIÇÃO
ESPACIAL.
Das sete famílias (Sciaenidae, Engraulidae, Gerreidae, Ariidae, Sparidae, Clupeidae e
Haemulidae), relatadas por Vieira & Musick (1994) como principais constituintes (acima
de 75% de ocorrência nas amostras) da rede de arrasto na assembléia de águas profundas
dos estuários do oeste do Atlântico, duas (Ariidae e
Sciaenidae), também foram
abundantes nos arrastos da Baía de Guaraqueçaba (tab. 55). As demais, embora capturadas,
ocorreram em freqüências inferiores a 0,24% da captura total e não foram capturados
exemplares da família Sparidae. Para a região temperado-quente do sudeste do Atlântico,
na costa do Brasil entre São Paulo (24o S) e Rio Grande do Sul (32o S) foram muito
abundantes os juvenis e subadultos de arídeos e scienídeos, coincidentemente aos
resultados obtidos para a Baía de Guaraqueçaba (Vieira & Musick 1994). Contudo, em
Guaraqueçaba, também foram muito abundantes os juvenis e adultos dos Paralichthyidae e
Tetraodontidae (tab.55). Similar ao constatado por Chaves (1995) e Chaves & Vendel
(1997), na região interna da Baía de Guaratuba e situada no litoral do Paraná, onde os
Tetraodontiformes e Pleuronectiformes
predominaram, juntamente com Sciaenidae,
Gerreidae, Ariidae e Haemulidae (incluída em Pomadasyidae por alguns autores), em
diversidade e abundância. Como observado por Saila (1983), Vieira & Musick (1993) e
Nelson (1976 e 1994) estas famílias apresentam ampla distribuição geográfica, refletindo a
ampla faixa de tolerância e de adaptações destes organismos.
As espécies capturadas na Baía de Guaraqueçaba, como já constatado para a as
famílias, também têm ampla distribuição geográfica (dos Estados Unidos até o norte da
Argentina), e todas já foram relatadas como associadas aos estuários e águas costeiras rasas
(<10 m) (Figueiredo 1981, Corrêa 1987, Haimovici et al. 1994).
94
Segundo Kennish (1990) a ictiofauna dos estuários é tipificada pela dominância
numérica de umas poucas espécies. Como esperado este fato também foi observado no
estuário da Baía de Guaraqueçaba. De modo geral a composição específica analisada
coincidiu com a descrita por Vieira & Musick (1994), que citaram como comuns dentre as
espécies de sciaenídeos juvenis e "subadultos" de Stellifer rastrifer em amostragens de
arrasto de fundo realizadas nos estuários da região entre São Paulo e Rio Grande do Sul.
Coincidiram ainda as observações de que entre os arídeos foram numericamente
dominantes Cathorops (Arius) spixii e Sciadeichthyes (A.) luniscutis. Contudo os
resultados diferiram daqueles autores pela superação numérica de Genidens genidens sob
Netuma barba. A discordância observada pode ser atribuída às diferenças latitudinais entre
os estuários, que estão submetidos a diferentes processos e características físico-químicas,
tais como temperatura da água, salinidade, interações intra e interespecíficas, relações
tróficas e ciclos reprodutivos (Kennish, 1990; Pereira, 1994; Agostinho et al., 1997).
Na tabela 56 comparou-se as composições ictiofaunísticas (famílias e espécies mais
abundantes), obtidas por vários autores nos diferentes habitats do sistema estuarino da Baía
de Paranaguá. Desconsiderando-se o método de captura, as famílias registradas como
muito abundantes, relacionadas por ordem do maior para o menor número de citações,
foram: Gerreidae, Sciaenidae, Engraulidae, Clupeidae, Ariidae, Carangidae, Pomadasyidae
e Paralichthyidae. Os Mugilidae, Poeciliidae, Atherinidae, Triglidae e Serranidae foram
abundantes em estudos isolados. Entre todas as famílias constatadas Mugilidae,
Atherinidae e Poecilidae não foram capturadas na Baía de Guaraqueçaba. As primeiras,
grupos dominantes nas águas rasas das zonas temperadas (Haimovici et al., 1994; Vieira &
Musick 1994), são comuns no sistema estuarino da Baía de Paranaguá e principalmente
capturadas com petrechos de pesca para espécies pelágicas, como redes de emalhe e de
cerco (Corrêa, 1987) e na zona de arrebentação (Chao et al. 1982, Maehama & Corrêa
95
1987, Cunha 1988; Monteiro Neto et al. 1990, Figueiredo et al. 1995, Godefroid &
Hofstaetter 1997a, Maciel 1995, Araújo et al. 1997). Contudo, os pescadores de
Guaraqueçaba declararam a família Mugilidae como freqüente nas capturas. Isto permite
concluir que a ausência do mugilídeos nos arrastos está principalmente relacionada com
este método de pesca.
Os poecilídeos são peixes comuns em rios e gamboas do litoral, raramente
ultrapassando os 18 cm, sendo a maioria muito menor e com algumas espécies tolerando
baixas salinidades (Barletta et al. 1990; Nelson, 1994; Cervigón 1996). São geralmente
capturados por aparelhos de pesca de baixa seletividade como picarés, puçás e armadilhas
de malhas diminutas (1mm) (Corrêa 1987, Vendel 1997). Kennish (1990) e Vieira &
Musick (1994) observaram que a similaridade ictiofaunística, nos estuários do oeste do
Atlântico, estava relacionada principalmente com o tipo de aparelho de pesca e ainda com
o tipo de habitat amostrado no estuário. Deste modo, a ausência dos
poecilídeos e
aterinídeos nas amostragens pode estar relacionada às preferência por ambientes diversos
dos analisados e/ou do método de pesca utilizado na Baía de Guaraqueçaba.
Embora a Baía de Guaraqueçaba tenha mostrado configurações e composições
ictiofaunísticas muito similares, foram observados padrões definidos de distribuição
temporal e espacial. As espécies C. spixii e S. rastrifer e G. genidens dominaram
numericamente a Baía de Guaraqueçaba, principalmente na área mais externa e na maioria
das épocas do ano (principalmente na épocas de baixas salinidades associadas aos períodos
de maior pluviosidade) , com notável exceção no inverno (julho). Neste período passaram
a dominar Etropus crossotus (na área mais externa), Sciadeichthys luniscutis, Citharichthys
arenaceus, Netuma barba, Micropogonias furnieri e Sphoeroides testudineus e foram
constatadas as maiores salinidades e transparências da água, diretamente influenciadas pela
menor pluviosidade. Estes fatores associados às interações intra e interespecíficas, relações
96
tróficas e ciclos reprodutivos influenciaram o padrão temporal observado (Livingston
apud Pereira 1994, Kennish 1994, Agostinho et. al. 1997, Barletta 1999, Chaves et al.
2000).
A composição principal da ictiofauna entre os pontos analisados apresentou
diferenças capazes de caracterizá-los como ambientes distintos, visto que as espécies
abundantes foram pouco comuns em alguns deles. Neste sentido foi notável a distribuição
do bagre Genidens genidens que ocorreu principalmente na região mais interna da Baía de
Guaraqueçaba (ponto 3) onde foram constatadas a menor salinidade e, como no ponto
intermediário 2, baixa profundidade. Estas observações permitem concluir que Genidens
genidens prefira ambientes com estas características. Segundo Chao et al. apud Haimovich
et al.(1994) e Fernandes-Pinto et al. (1997), esta espécie utiliza ambientes de baixa
salinidade para reprodução e desova. De fato Genidens genidens utilizou a área para
reprodução. Além disso, os pontos amostrais apresentaram diferenças significativas na
profundidade, salinidade e transparência da água. Isto permite concluir que as
dessemelhanças observadas na composição entre os pontos amostrais tenha ocorrido em
função das diferenças na profundidade e nas características físico-químicas da água, que
são fatores importantes nas configurações de distribuição espacial (e também temporal) da
ictiofauna (Davis 1988, Hook 1991, Matthews et al. 1992, Kennish 1994, Agostinho et al.
1997 e Barletta 1999).
No sistema estuarino da Baía de Paranaguá as maiores freqüências de captura para
o arrasto com portas tem ocorrido no verão e/ou outono e eventualmente - na zona de
arrebentação de Pontal do Sul à Praia de Leste e da Enseada do Benito - no inverno e/ou
primavera (Maehama & Corrêa 1987, Fernandes-Pinto et al. 1997) (tab.56).
Neste
contexto a Baía de Guaraqueçaba apresentou as maiores capturas (em número de
exemplares e peso) na primavera, verão e outono que diferiram dos demais estudos do
97
sistema estuarino (limitados ao verão e/ou outono). Levando-se em consideração que as
espécies mais abundantes capturadas em Guaraqueçaba: a) diferiram daquelas obtidas nos
arrastos do sistema estuarino e B) reproduziram-se e recrutaram da primavera ao final do
outono é razoável supor que estes fatos estão associados. Do mesmo modo que as
diferenças observadas em relação zona de arrebentação e à Enseada do Benito, também
podem ser atibuídas às configurações familiar e específica, suas relações com as variáveis
ambientais e ás interações biológicas, ciclos reprodutivos e de utilização dos habitats
amostrados (Kennish 1994, Barletta 1999). Além disso, embora os estuários sejam
caracterizados como áreas fisicamente instáveis os peixes freqüentemente apresentam
padrões sazonais e espaciais mais ou menos previsíveis de abundância e distribuição (Furia
1996). Segundo Kennish (1994), ainda que apresentem poucas espécies dominantes, as
comunidades ictíicas experimentam mudanças temporais na composição e diversidade em
função da mobilidade básica dos peixes e suas migrações sazonais associadas aos ciclos
reprodutivos e alimentares. Isto foi particularmente válido para a Baía de Guaraqueçaba.
Na Baía de Guaraqueçaba observou-se que na primavera a ictiofauna estava
composta por um menor número de indivíduos com peso elevado e no outono por muitos
exemplares de peso muito inferior, que reunidos elevaram o peso total da captura.
Levando-se em consideração que: a) foram detectadas atividades reprodutivas muito
intensas a moderadas, entre a primavera e o verão, para as espécies Cathorops spixii,
Sphoeroides testudineus, Stellifer rastrifer, Isopisthus parvipinnis, Sciadeichthyes
luniscutis e Genidens genidens b) que as maiores freqüências exemplares em reprodução e
imaturos destas espécies foram capturados respectivamente no outono e na primavera, é
possível concluir que os adultos visitaram a Baía de Guaraqueçaba para reproduzirem-se
entre a primavera e verão e que os picos do recrutamento tenham ocorrido no outono.
Similar ao constatado para a maioria das espécies presentes no sistema estuarino da Baía
98
de Paranaguá que reproduzem-se entre a primavera e final do verão (Corrêa 1987;
Yamaguti et al, 1994) e recrutam do último período até o final do outono (Corrêa, 1987;
Yamaguti et al., 1994).
5.3. DIVERSIDADE, EQUITATIVIDADE E RIQUEZA.
Em regiões subtropicais, a utilização do ambiente pelos grupos de espécies tem
relação com a temperatura (MacFarland 1963, Monteiro Neto et al. 1990, Kennish 1994) e
ainda, as semelhanças entre as comunidades de peixes nos estuários em função das
características físico-químicas da água (Lyons, 1996; Taylor et al., 1996, Barletta, 1999).
Estas afirmativas são particularmente válidas na estruturação da ictiofauna da Baía de
Guaraqueçaba, uma vez que os maiores valores de diversidade e eqüitatividade
correlacionaram-se com os meses e pontos amostrais que apresentaram: a) os maiores
valores de salinidade, pH e oxigênio dissolvido na água de profundidade e b) as menores
profundidades e teores de oxigênio dissolvido na água superficial.
As maiores temperaturas, precipitações e teores de oxigênio dissolvido na água de
profundidade foram observadas nos meses de verão em todos os pontos amostrais,
correlacionando-se aos de menores valores de salinidade (superficial e de profundidade) e
transparência da água. Isto estaria definindo os padrões temporais observados para a Baía
de Guaraqueçaba, com a maior freqüência absoluta em número de espécies na primavera e
de diversidade e eqüitatividade (que excluem as espécies raras) no período de inverno,
quando a abundância e biomassa das espécies dominantes diminuiu. A menor diversidade e
eqüitatividade, ao contrário, foram observadas no verão. A pluviosidade por sua forte
correlação com a temperatura e influência sobre a salinidade, transparência, pH e oxigênio
99
dissolvido, possivelmente desempenhou o papel determinante nas variações de diversidade
e eqüitatividade da Baía de Guaraqueçaba.
Ainda que a época de maior freqüência de captura da Baía de Guaraqueçaba tenha
sido similar às constatadas nos arrastos com portas do resto do sistema estuarino da Baía
de Paranaguá, diferiu do último com relação às épocas – verão e/ou outono – de maior
número de espécies. Na Baía de Guaraqueçaba o maior número de espécies ocorreu no
inverno e primavera, o que coincidiu com o observado por Fernandes-Pinto et al. (1997)
para a Enseada do Benito (tab. 56). A diferença entre épocas pode estar relacionada ao fato
de que a maioria dos estudos analisados foi realizado em áreas geográficas distantes da
Baía de Guaraqueçaba. Ainda todas estão próximas aos canais de acesso ao sistema
estuarino, e sob maior influência do ambiente nerítico adjacente (Bigarella 1978, Lana
1986, Brandini et al. 1988 e Disaró 1995), enquanto que a Enseada do Benito, por sua
proximidade espacial, é muito similar à Baía de Guaraqueçaba. De fato, nos ambientes
costeiros os maiores valores na abundância e no número de espécies de peixes, nos arrastos
de fundo, foram obtidos no verão e os menores no inverno (Chao et al.1985, Paiva-Filho &
Toscano 1987, Giannini & Paiva-Filho 1990), que estão relacionados aos reflexos da
influência da ACAS sobre a plataforma continental (Rocha 1990, Rossi-Wongtschowski &
Paes 1993, Natali Neto 1994).
Considerando que os menores valores de diversidade ocorreram entre o final do
outono (maio) e o início do inverno (junho) e de eqüitatividade no primeiro, quando a
abundância de Stellifer rastrifer, Genidens genidens, Isopisthus parvipinnis Etropus
crossotus,
Sciadeichthys
luniscutis,
Citharichthys
arenaceus,
Netuma
barba,
Micropogonias furnieri e Sphoeroides testudineus diminuiu e C. spixii passou a dominar
na comunidade das áreas mais externa e mediana da Baía de Guaraqueçaba (pontos 1 e 4).
Nestas épocas e locais, foram constatados altos valores de profundidade e transparência da
100
água e baixos valores de salinidade, pH (superficial e de profundidade) e oxigênio
dissolvido (de profundidade). Isto permite concluir que, dentre as outras espécies presentes
na comunidade, C. spixii ocorreu em abundância nestas condições ambientais.
Corroboraram ainda esta afirmação, o fato de que as maiores diversidades e eqüitatividades
anuais foram observadas nos meses agosto, setembro
e julho (inverno/ início da
primavera) nos pontos situados na região mediana (2 e 4) da Baía de Guaraqueçaba,
quando foram constatadas as maiores salinidades (superficial e de profundidade), altos
valores de transparência e baixos valores de temperatura da água, pluviosidade,
profundidade e oxigênio dissolvido na água de superfície. Nestes meses as freqüências de
ocorrência de Stellifer rastrifer, Genidens genidens, Isopisthus parvipinnis, Etropus
crossotus,
Sciadeichthys
luniscutis,
Citharichthys
arenaceus,
Netuma
barba,
Micropogonias furnieri e Sphoeroides testudineus aumentaram e a de C. spixii dimi nuiu
significativamente. Isto evidenciou que as primeiras, por tolerarem estas condições
ambientais,
passaram
a
ocupar
e
dominar
numéricamente
as
áreas,
antes
predominantemente ocupadas pela população de C. spixii , que deslocou-se para áreas
externas da Baía de Guaraqueçaba.
A área mais interna (ponto 3) apresentou altos valores de diversidade na metade do
outono (abril) e de eqüitatividade no verão (janeiro) que, diminuíram simultaneamente nos
meses de inverno (agosto, setembro) e primavera (novembro), quando a abundância de C.
spixii e das outras espécies diminuiu e de G. genidens aumentou. Isto permite concluir que
a abundância das primeiras aumenta temporariamente e a de Genidens genides, por sua
abundância constante, como já discutido anteriormente, parece preferir as condições desta
área. Além disto na metade do outono, as variáveis ambientais não apresentaram
correlação nítida com as freqüências de nenhuma das espécies analisadas. Apresentou
valores intermediários de salinidade, temperatura, profundidade, oxigênio dissolvido, pH e
101
pluviosidade. Provavelmente, esta estabilidade foi a principal responsável pelo aumento no
outono da diversidade na área mais interna da Baía de Guaraqueçaba. Com base nestas
considerações é razoável afirmar que, dentre outros, os parâmetros meteorológicos e
hidrológicos estariam desempenhando um papel importante na alteração da composição,
diversidade, eqüitabilidade e distribuição espacial e variabilidade temporal da ictiofauna da
Baía de Guaraqueçaba.
Alterações sazonais nas taxas de capturas de comunidades ictíicas tropicais e
subtropicais foram constatadas em vários estuários e atribuídas principalmente aos padrões
de reprodução e aumento do recrutamento (Batista e Rêgo 1996, Laroche et al. 1997,
Abilhôa 1998, Pinheiro 1999 e Barletta 1999). Barletta (1999) observou que as assembléias
de peixes no estuário Caeté (Amazônia) apresentaram padrões temporais (épocas de seca e
chuva) de deslocamento espacial, que interferiram nas taxas de captura das comunidades
ictíicas. O mesmo autor constatou que, durante as estações chuvosas, as espécies
parcialmente dependentes do estuário (sensu Barletta 1999) moviam-se para as regiões
inferiores do estuário e propôs ainda que as mudanças sazonais das assembléias de peixes
estuarinos podem ser determinadas pela combinação de flutuações temporais das
abundâncias das espécies ictíicas induzidas pelas chuvas, reprodução e recrutamento, e
pelo recrutamento das espécies marinhas e dulciaqüícolas. Também foram observadas
atividades reprodutivas para as espécies mais abundantes na Baía de Guraqueçaba.
Portanto, concordam com estas proposições as variações observadas nas flutuações das
abundâncias e das distribuições espaço-temporais das espécies ictíicas e as suas relações
com as variáveis ambientais da Baia de Guaraqueçaba.
Segundo Ludwig & Reynolds (1988) o número de espécies de uma área depende da
abundância relativa, da homogeneidade, tamanho da amostra e do tempo gasto na procura.
Ainda, em situações em que o tamanho das amostras não são iguais o método conhecido
102
como rarefação pode ser utilizado para permitir comparações do número de espécies entre
comunidades. Não pode ser esquecido ainda a seletividade amostral do aparelho de pesca
utilizado (Vieira & Musick, 1994). Na tabela 39 é apresentada a comparação da estimativa
do número de espécies com base no número de exemplares capturados, pelos diferentes
autores que utilizaram o arrasto com portas no sistema estuarino da Baía de Paranaguá,
sobrepostos nas curvas de rarefação obtidas para a Baía de Guaraqueçaba. Relativamente à
última, a riqueza de espécies foi superior a estimada para a embocadura do Rio do Maciel,
inferior a dos ambientes de infralitoral próximos da Ilha do Mel e similar, também na
composição, à da Enseada do Benito. Estas diferenças entre as áreas analisadas podem ser
atribuídas ao reflexo do balanço entre dois conjuntos de forças antagônicas. De um lado os
processos abióticos locais e as interações entre espécies que tendem a reduzir a
diversidade. De outro a imigração de outras comunidades para dentro da localidade que
tendem a aumentá-la (Ricklefs & Schluter, 1993).
5.4. ESTRUTURA.
Comprimento e Peso.
As populações de 38 espécies (73,07% do total capturado) estavam constituídas
principalmente por indivíduos com CT médios 50% inferiores ao CT máximo registrado
na literatura. Isto permite concluir que as populações naturais da maioria das espécies
estavam compostas principalmente por indivíduos juvenis. Como já observado e discutido
para a composição da ictiofauna, a variação temporal na estrutura também refletiu a
utilização dos estuários, como já constatado por Reis (1986), Corrêa (1987), Paiva Filho &
Toscano (1987), Corrêa (1991), Chaves (1994), Haimovici et al. (1994), Yamaguti et al.
(1994), Kennish (1994), Corrêa et al. (1995); Maciel (1995), Fernandes-Pinto et al.(1997),
Abilhôa (1998), Pinheiro (1999), Barletta(1999) e Chaves et al. (2000). Na sua maioria as
103
formas mais abundantes são juvenis que utilizam o ambiente estuarino como área de
alimentação e crescimento (Saila 1983, Kennish 1994, Wooton 1998, Chaves et al. 2000).
Foram observadas estratificações temporais e espaciais de CT e peso médio da
ictiofauna e as reduções detectadas nos CT médios retrataram os pontos e épocas de maior
recrutamento das espécies, quando ocorreu o aumento significativo no número de
indivíduos das classes de menor tamanho. Fato já observado por Yamaguti et al. (1994),
Kennish (1994), Fernandes-Pinto et al.(1997), Abilhôa (1998) e Barletta (1999).
A tendência geral observada foi a de redução do CT do inverno para o outono. Fato
também constatado por Abilhôa (1998) em um banco areno-lodoso na Ilha do Mel.
Contudo divergiram deste local as maiores médias de CT e peso que ocorreram na Baía de
Guaraqueçaba na primavera e no banco areno-lodoso no inverno. O mesmo autor propôs,
com base na composição da ictiofauna e nos estádios avançados de maturação gonadal, que
o inverno parecia ser a época de entrada ou saída dos adultos do estuário, provavelmente
relacionadas a alguma atividade reprodutiva. Apontou Sphoeroides testudineus e Genidens
genidens como as espécies de maior tamanho no inverno. Estas espécies também foram
abundantes em Guaraqueçaba, contudo apresentaram maiores médias de CT e de peso no
período de primavera. Diferente do banco areno-lodoso da Ilha do Mel, ambiente situado
próximo do ambiente nerítico, a Baía de Guaraqueçaba corresponde a um dos locais mais
internos do sistema estuarino da Baía de Paranaguá. Estas observações, associadas ao fato
de que na Baía de Guaraqueçaba as maiores freqüências de ocorrência e de atividade
reprodutiva de Sphoeroides testudineus e Genidens genidens ocorreram nas épocas
adjacentes ao do inverno, de que estas espécies reproduzem-se nos estuários e canais
estuarinos (Haimovici et al. 1994) permite concluir que estas espécies deslocam-se para o
interior do sistema estuarino da Baía de Paranaguá para a realização de atividades
reprodutivas.
104
Estádio de maturação gonadal.
As espécies Cathorops spixii, Citharichthys arenaceus, Etropus crossotus,
Genidens genidens, Isopisthus parvipinnis, Micropogonias furnieri, Netuma barba,
Sciadeichthys luniscutis, Sphoeroides testudineus e Stellifer rastrifer são consideradas
como estuarino dependentes para reprodução, recrutamento e/ou alimentação(Reis 1986,
Corrêa 1987, Paiva Filho & Toscano 1987, Corrêa 1991, Chaves 1994, Chaves & Vendel
1997, Haimovici et al. 1994; Castello et al. 1994, Corrêa et al. 1995; Maciel 1995;
Fernandes-Pinto 1997; Abilhôa 1998; Pinheiro 1999, Barletta 1999, Chaves & Bouchereau
2000). De fato todas apresentaram, em maior ou menor grau, atividades reprodutivas e o
recrutamento na Baía de Guaraqueçaba. Ainda foram observados padrões de estratificação
espaço-temporal e em relação ao tamanho, peso e estádio de maturação que caracterizaram
movimentos freqüentes entre as áreas da Baía. Estes movimentos particulares explicam e
corroboram o uso (estrutura CT, peso e e suas relações com a reprodução) e ocupação
espaço-temporal (composição e suas relações com as variáveis ambientais) gerais da
ictiofauna. Fato já constatado por Kennish (1994) que atribuiu as migrações sazonais dos
peixes estuarinos aos ciclos reprodutivos e alimentares e é um indicativo do modo de
utilização do estuário por muitas espécies como áreas de crescimento, reprodução e
alimentação (Reis 1986; Corrêa 1987; Paiva Filho & Toscano 1987; Corrêa 1991; Chaves
1994; Haimovici et al. 1994, Castello et al. 1994; Corrêa et al. 1995; Maciel 1995;
Fernandes-Pinto et al. 1997; Abilhôa 1998; Pinheiro 1999 e Barletta 1999). Ainda por
Barletta (1999) que também contatou movimentos freqüentes e atribuiu o padrão de
distribuição espacial e temporal, não excluindo a integração da reprodução e recrutamento
seqüencial das espécies através do ano, às variáveis ambientais (salinidade associada à
pluviosidade). Isto foi particularmente válido para Guaraqueçaba visto que, de modo geral
a reprodução das 6 espécies que apresentaram atividades de intensa à muito intensa, iniciou
105
no final do inverno, atingiu o máximo na primavera e estendeu-se até o final do outono
(épocas de aumento da temperatura da água e de redução da salinidade – definida pela
estação chuvosa), nas áreas externa (ponto 1 – C. spixii, I. parvipinnis, S. rastrifer),
mediana rasa (ponto 2 – S. testudineus), mediana profunda (ponto 4 – C. spixii e S.
luniscutis) e interna ( ponto 3 – Genidens genidens). De modo geral o número de
exemplares em reprodução foi significativamente inferior ao de desovados, provavelmente
em função do lapso de tempo entre as capturas, uma vez que a transição do estádio em
reprodução para o desovado ou esvaziado ocorre de modo imediato.
O período reprodutivo, constatado entre a primavera/verão, coincidiu com o
relatado na literatura específica (tab58). Contudo, C. spixii e S. luniscutis apresentaram a
defasagem de um mês em relação às outras regiões, e a incubação dos ovos por C. spixii
também prolongou-se pelo mesmo período. Fato possivelmente relacionado às condições
particulares das variáveis ambientais da Baía de Guaraqueçaba e ainda às necessidades
individuais das espécies, já que a desova, eclosão dos ovos e desenvolvimento das larvas
devem ocorrer em épocas com os recursos apropriados (Wootton 1998, MacConnell 1999).
Os tamanhos da primeira maturação, das espécies com atividades reprodutivas
intensas e muito intensas, também difeririam em relação ao constatado para outras regiões
(tab. 58). Possivelmente as diferenças observadas estão relacionadas ao fato de que os
estudos experimentais e observacionais mostraram que a idade e/ou comprimento da
maturidade é uma resposta adaptativa à seleção e varia (em função inversa à taxa de
mortalidade e crescimento) em resposta às alterações ambientais (Wooton 1998,
McConnell 1999).
Já as reduções, no número de exemplares e de espécies, observadas nos demais
estádios de maturação possivelmente estejam relacionadas ao fato de que a Baía de
Guaraqueçaba pode representar o limite do "continuum" reprodutivo e de distribuição das
106
populações destas espécies, cujo centro pode estar situado nas áreas, dos rios ou do sistema
estuarino da Baía de Paranaguá, adjacentes.
Comportamento frente a salinidade.
Muitas classificações das associações de peixes estuarinos são baseadas na sua
tolerância à salinidade e seus padrões de migrações nos estuários (Blaber et al. 1989,
Bulger et al. 1995). Quantificam e qualificam a importância dos estuários como áreas de
reprodução, crescimento e proteção para as espécies ictíicas que os utilizam (Barletta
1999). As 17 espécies capturadas como adultos e juvenis em salinidades situadas entre 5 e
25‰ são comuns nos estuários e freqüentemente capturadas em todos os estádios de
desenvolvimento (Corrêa 1987, Haimovici et al. 1994, , Corrêa 1991, Chaves 1994,
Chaves & Vendel 1997, Haimovici et al. 1994; Castello et al. 1994, Corrêa et al. 1995;
Maciel 1995; Fernandes-Pinto 1997; Abilhôa 1998; Pinheiro 1999, Barletta 1999). Todas
podem consideradas como eurihalinas verdadeiramente estuarinas. Particularmente C.
spixii, C. arenaceus, G. genidens, M. furnieri, N. barba, S. luniscutis, S. testudineus e S.
rastrifer que apresentaram atividades reprodutivas na Baía de Guaraqueçaba. Contudo, M.
furnieri é citada como marinha que desova nas águas costeiras e utiliza os estuários nas
formas larval e juvenil (Yamaguti et al. 1994). Considerando-se que os adultos desta
espécie ocorreram em baixas freqüências em Guaraqueçaba e que a atividade reprodutiva
foi incipiente é possível concluir que de fato esta espécie prefira locais mais próximos da
região costeira para reproduzir-se. Com base nestas considerações é razoável afirmar que
aquelas espécies, com exceção M. furnieri, enquadrem-se na categoria de eurihalinas
verdadeiramente estuarinas (sensu Cariiker in Lauff 1967 e Yánez & Nugent 1977). Por
sua vez M. furnieri pode ser incluída como eurihalina marinha, utilizando-se dos estuários
como local de refúgio e crescimento. Esta proposições coincidiram com Barletta (1999),
107
que incluiu C. spixii e S. rastrifer (com base nos estádios de desenvolvimento capturados e
nos períodos de chuva e seca amostrados no estuário Caeté) entre as espécies
verdadeiramente estuarinas (sensu MacHugh 1967) ou estuarino residentes (sensu Parker
1974) e M. furnieri como espécie marinha estuarino dependente (sensu Chao et al. 1985).
Ainda, considerando-se as 17 espécies de Guaraqueçaba incluídas como eurihalinas
verdadeiramente estuarinas, Barletta (1999) considerou G. oceanicus e G luteus como
verdadeiramente estuarinas e S. testudineus, capturado em baixas freqüências e apenas nos
estádios de pós-larvas e juvenis, como parcialmente dependente do estuário. G. oceanicus
e G luteus ocorreram em baixas freqüências e não apresentaram atividades reprodutivas na
Baía de Guaraqueçaba. Isto permite concluir que, embora possam ser enquadradas como
eurihalinas verdadeiramente estuarinas, preferem outras regiões do sistema estuarino para a
reprodução, recrutamento e crescimento. Com relação a S. testudineus, ao contrário, foi
abundante, apresentou atividade reprodutiva intensa entre setembro e outubro e, face as
baixas freqüências de juvenis, parece não recrutar na Baía. Considerando-se que Barletta
(1999) e Barletta & Bergan (1999) capturaram pós-larvas e juvenis nas áreas rasas dos
manguezais é provável que a espécie recrute em áreas semelhantes do sistema estuarino da
Baía de Paranaguá. Além disso, tanto os adultos quanto os juvenis tem sido relatados como
comuns no sistema estuarino da Baía de Paranaguá (Corrêa 1987, Corrêa 1991, Corrêa et
al. 1995, Fernandes-Pinto 1997; Abilhôa 1998; Pinheiro 1999). Isto permite concluir que a
espécie possa ser incluída como eurihalina verdadeiramente estuarina. Wootton (1999)
reconheceu três tipos de habitat que são ocupados pelos peixes para reprodução,
alimentação e refúgio em períodos de condições bióticas ou abióticas desfavoráveis. Estes
habitats podem estar próximos ou distantes. Levando-se em consideração todo o sistema
estuarino, possivelmente este comporte os três tipos de habitat para a espécie, enquanto
que a amplitude da área estudada por Barletta (1999) comporte dois. Ainda o próprio autor
108
reconheceu que as discordâncias entre as dependências das espécies de peixes nas áreas
estuarinas não residem somente no fato das relações entre o ciclo de vida das espécies e
das condições necessárias durante as fases de seu ciclo de vida. O problema também é
identificar os limites entre o estuário e as áreas costeiras nas regiões estuarinas. Comentou
ainda que no norte da América do Sul as águas costeiras são influenciadas pelo vasto
aporte de água doce proveniente de grandes (Orinoco e Amazonas) e de muitos outros
pequenos rios, tal que as condições estuarinas passam a prevalecer ao longo da costa.
Então é aceitável, como citou o autor, afirmar que certas espécies são dependentes das
condições estuarinas mesmo que capturadas na costa em profundidade de até 25m. Isto
pode ser particularmente válido para o sistema estuarino onde a região mais próxima da
costa sofre tanto a influência direta do oceano adjacente quanto da intrincada drenagem
dos inúmeros rios, podendo apresentar em determinados períodos e locais condições
marinhas e/ou estuarinas (Brandini 1985, Brandini & Thamm 1994, Disaró 1995).
O segundo subgrupo de 22 espécies capturadas em salinidades situadas entre 10 e
25‰, estava composto unicamente por estádios juvenis. Considerando-se que os adultos
das espécies demersais têm, em sua maioria, preferência por águas costeiras de maior
salinidades, visitando ocasionalmente os estuários para se alimentar (Corrêa 1987, Corrêa
1991, Haimovici e al. 1994, Vieira & Musick 1994, Paiva Filho & Toscano 1987, Abilhôa
1998, Pinheiro 199), é possível incluí-las entre os eurihalinos marinhos, com juvenis e
adultos marinhos, próximos dos estuários (ciclo de vida costeiro e em salinidades
superiores a 25‰), e com juvenis e adultos entrando ocasionalmente no estuário (até
salinidades de salinidades de 5‰). De fato Barletta (1999) também incluiu, considerandose as espécies comuns aos dois estuários, C. acoupa, C. faber, O. saliens, O. saurus, S.
vomer, C. leiarchus e C. microlepidotus como espécies marinhas estuarino dependentes
(espécies marinhas que desovam na região costeira e utilizam o ambiente estuarino como
109
área de crescimento durante os primeiro anos de seu ciclo de vida). Dentre as espécies
capturadas em salinidades superiores a 25‰, simultaneamente como adultos e juvenis,
unicamente I. parvipinnis apresentou atividade reprodutiva moderada a intensa. Esta
espécie foi relatada como abundante nos estuários, apresentando movimentos sazonais
relacionados a reprodução, recrutamento, crescimento e alimentação e os padrões descritos
são similares aos constatados na Baia de Guaraqueçaba (Giannini & Paiva Filho 1994,
Chaves et al. 1997). Na Baia de Santos a espécie foi abundante entre março e agosto
(outono e inverno) em águas mais profundas, de baixas temperaturas e altas salinidades e
concentrações de oxigênio dissolvido, onde os jovens do ano foram principalmente
capturados (Giannini & Paiva Filho 1994). Ainda a espécie reproduz-se no manguezal ou
no mar aberto e foi considerada como espécie predadora oportunista (Chaves et al. 1997).
Estas considerações permitem incluí-la como eurihalina marinha (ciclo de vida costeiro e
em salinidades superiores a 25‰), e com os adultos e juvenis entrando ocasionalmente no
estuhrio (ate salinidades de 5‰). Ainda, Giannini (1989) constatou que S. brasiliensis
ocorreu associada, e portanto nas mesmas condições, a I. parvipinnis. Foi relatada ainda
como abundante e freqüentemente capturada associada ao camarão sete-barbas, isto é em
regiões costeiras entre 4 e 14m (Coelho et al. 1987). Não foram constatadas atividades
reprodutivas para S. brasiliensis em Guaraqueçaba. Com base nestas observações S.
brasiliensis também pode ser considerada como eurihalina marinha.
Finalmente as espécies demersais capturadas unicamente como juvenis e em
salinidades superiores a 25‰ são todas freqüentemente capturadas como adultos na região
costeira e marinha, com juvenis ocasionalmente entrando nos estuários (Corrêa 1987,
Gianninni 1989, Haimovici et al 1994, Castello et al. 1994, Corrêa et al. 1995, Abilhôa
1998, Pinheiro 1999). Por apresentarem estas características podem ser consideradas como
eurihalinas com desenvolvimento em salinidades entre 5 e 25 ‰ (estuários) e adultos
110
somente em salinidades superiores a 25 ‰.
5.5. Importância como Recurso
A constatação de que 73,91% da comunidade de Guaraqueçaba tem na pesca sua
atividade principal coincidiu com os resultados obtidos por CORRÊA et al. (1997) que
registraram, na maioria das comunidades litorâneas amostradas (Eufrasina, Almeida,
Guaraqueçaba, Tibicanga e Superagüi), acima de 70% de entrevistados dependendo da
pesca para sobreviver. Corroboraram ainda os padrões relatados por Kraemer (1982) e
Rougeulle (1993), de que a pesca no complexo é de subsistência com a comercialização
dos excedentes de produção. Ainda, pela baixa presença de outros elementos da economia
local, a tendência geral das outras localidades do sistema estuarino da Baía de Paranaguá
do abandono progressivo de outras atividades para dedicação exclusiva à pesca (SPVS
1992).O fato dos pescadores de Guaraqueçaba consumirem peixes entre 4 e 5 dias por
semana permite concluir que a comunidade é dependente do peixe como principal fonte
protéica. Este resultado coincidiu com o observado por Corrêa et al. (1997), que
constataram entre 2,94 a 5,61 dias/semana de consumo de peixes nas demais comunidades
do sistema estuarino da Baía de Paranaguá,. Os mesmos autores observaram que o grau de
dependência do peixe era diretamente proporcional ao grau de isolamento geográfico da
comunidade em relação aos grandes centros urbanos, que possibilitam uma maior
diversificação dos tipos de alimento. Isto parece particularmente válido para Guaraqueçaba
visto as dificuldades de deslocamento terrestre, pouco tempo disponível para atividades
que não a pesca e baixa potência das embarcações que dificultam o acesso aos maiores
centro urbanos (Karam & Toledo 1997).
A pesca parece estar limitada, por motivos operacionais (principalmente barcos a
remo e a motor com até 11 HP e de pequeno tamanho), às regiões próximas das
111
comunidades e coincidiram com Kraemer (1982), Corrêa (1987 e 1997) e SPVS (1992)
que já haviam definido a pesca das comunidades do complexo estuarino-baía de Paranaguá
como tipicamente artesanal, realizada com barcos de baixa autonomia de curso e, por isso,
limitada às regiões estuarinas protegidas.
Os petrechos de pesca utilizados coincidiram com aqueles observados por Corrêa et
al. (1997). Os mesmos autores constataram que os tipos preferidos de petrecho estão
relacionados com o custo, a eficiência na captura e o tipo de recurso procurado. O jerivau é
utilizado na captura de camarões enquanto que as redes de caceio e de fundeio são
utilizadas na captura de peixes demersais e pelágicos. Constataram ainda que as diferenças
entre os comprimentos, alturas e tamanhos das malhas das redes de caceio, fundeio e
lanceio estavam principalmente relacionadas com as limitações operacionais impostas
pelas embarcações e ambientes em que são utilizadas. As redes utilizadas na pesca
costeira, fora do estuário, foram significativamente maiores que as utilizadas pela pesca
estuarina, fato que já havia sido constatado pela SPVS (1992). Quanto ao arrasto com
portas seu uso só é permitido, pela legislação, nas águas costeiras adjacentes ao complexo,
raramente visitadas pelos pescadores entrevistados.
As diferenças observadas entre as médias do número de horas mês gastos por
pescador foram significativamente maiores para as localidades próximas de suas
comunidades. Isto corroborou a afirmação proferida anteriormente de que o tipo de
embarcação, visto que a maioria dos declarantes possuíam embarcações movidas a remo
e/ou a motor de baixa potência, inviabiliza ou dificulta a escolha de locais mais distantes
para a pesca. Os últimos foram sempre declarados como segunda e terceira opções, isto é,
quando inexistem possibilidades, por escassez de recursos pesqueiros, nas regiões
próximas de sua comunidade. Evidentemente, a escolha de áreas mais distantes implica um
aumento considerável do esforço pesqueiro, raramente suportado pela pesca tipicamente
112
artesanal.
Este fato já havia sido observado por CORRÊA et al. (1997), onde os
pescadores optam primeiro pelos locais mais próximos de suas localidades (onde gastam o
maior número de horas), restando a segunda e terceira opções para os locais mais distantes.
As espécies declaradas pelos pescadores como freqüentes foram também as mais
abundantes nos arrastos (Cathorops spixii, Genidens genidens, Sciadeichthyes luniscutis e
Netuma barba). Contudo, divergiram da abundância declarada para Harengula clupeola
Bairdiella ronchus, Mugil curema, Sardinella brasiliensis, Oligoplites saurus, O. saliens,
Mugil platanus, Pogonias cromis, Mugil gaimardianus, Eucinostomus melanopterus e E.
gula e Opisthonema oglinum, que foram capturadas em baixas freqüências ou
encontravam-se completamente ausentes nos arrastos em todos os pontos amostrais).
Porém coincidiram com Corrêa (1987) que referiram-se a estas espécies como abundantes
no complexo-estuarino baía de Paranaguá. Estas diferenças podem ser atribuídas
principalmente à maior diversificação de petrechos utilizados e de áreas visitadas pelos
pescadores.
Considerando-se que 86,95% dos entrevistados pesca para a subsistência (consumo
próprio e da família), que o número médio de dias que os pescadores de Guaraqueçaba
consomem peixe (de 4 a 5 dias na semana), que o número médio de horas mensais gastas
pescando é 132,61 e ainda as abundâncias regionais dos peixes declarados, é razoável
concluir que as seguintes famílias e espécies podem ser consideradas como importante
fonte de recursos:
1) Ariidae, com as espécies Cathorops spixii, Genidens genidens, Sciadeichthyes
luniscutis e Netuma barba,
2) Clupeidae, com Harengula clupeola, Sardinella brasiliensis e Opisthonema
oglinum,
113
3) Sciaenidae, com Bairdiella ronchus, Menticirrhus americanus e Cynoscion
leiarchus,
4) Mugilidae, com Mugil curema, M. gaimardianus e M. liza.
6. CONCLUSÕES
1. A composição ictiofaunistica demersal da Baia de Guaraqueçaba:
?? não diferiu, no nível familiar, da constatada para outros estuários e baias da costa
do Brasil e estava representada principalmente por Ariidae, Sciaenidae,
Tetraodontidae e Paralichthyidae,
?? no nível especifico, foram particularmente abundantes Cathorops spixii, Stellifer
rastrifer,
Genidens
genidens,
Isopisthus
purvipinnis,
Etropus
crossotus,
Sciadeichthys luniscutis, Citharichthys arenaceus, Netuma barba, Micropogonias
furnieri e Sphoeroides testudineus, espécies consideradas como tipicamente
estuarinas e costeiras,
?? diferiu quantita e qualitativamente, nos níveis familiar e específico, dos demais
”habitats” do sistema estuarino da Baia de Paranaguá,
?? apresentou distribuição espacial similar, contudo as diferenças nas abundâncias
especificas permitiram caracterizar as áreas analisadas como distintas,
?? apresentou padrões de variabilidade temporal associados aos parâmetros
meteoro1ógicos (pluviosidade) e hidrológicos (salinidade, transparência e
profundidade), que possivelmente estariam desempenhando o principal papel na
alteraq5o da diversidade, eqiiitatividade e distribuição espacial da ictiofauna.
2. A riqueza ictiofaunística foi menor as das regiões próximas ao ambiente nerítico
adjacente e similar as regiões mais internas do sistema estuarino da Baia de
Paranaguá (como a Enseada do Benito).
114
3. Foram observados, para as espécies mais abundantes, padrões de estratificação
espaço-temporal em relação ao tamanho, peso e estádio de maturação gonadal que
caracterizaram movimentos freqüentes nas áreas da Baia e o uso e a ocupação para
reprodução e recrutamento.
4. A maioria das populações naturais de 38 espécies (73,07%) utilizou a Baia de
Guaraqueçaba como local de desenvolvimento.
5. Utilizaram a Baía de Guaraqueçaba Cathorops spixii, Stellifer ratrifer, Isopisthus
parvipinnis, Genidens genidens, Scideichthys luniscutis e Sphoeroides testudineus
para a reprodução e, com exceção da última, recrutamento. As três primeiras
utilizaram principalmente a área mais externa (ponto 1) e a primeira, juntamente
com S. luniscutis, também a mediana profunda (ponto 4). A mediana rasa (ponto 2)
foi utilizada principalmente por S. testudineus e a mais interna (ponto 3) por G.
genidens.
6. A Baia de Guaraqueçaba possivelmente representa o limite do ”continuum”
reprodutivo e de distribuição de Netuma barba, Etropus crossotus, Micropogonias
furnieri e Citharichthys arenaceus cujo centro pode estar situado nas áreas, dos rios
ou do sistema estuarino da Baia de Paranaguá, adjacentes.
7. O período reprodutivo das espécies mais abundantes estendeu-se do final do
inverno ao final do outono (com o pico na primavera) e o de recrutamento do início
do verão ao final do outono (com o pico no ultimo).
8. A maioria (42,31%) das espécies corresponderiam às eurihalinas marinhas, sete
(17,31%) espécies (entre elas as mais abundantes C. spixii, C. arenaceus, G.
Genidens, N. barba, S. lunicutis, S. testudineus e S. rastrifer) às eurihalinas
verdadeiramente estuarinas e 8 (15,40%) às eurihalinas com desenvolvimento nos
estuários (sensu Cariiker in Lauff 1967 e Yánez & Nugent 1977).
115
9. A comunidade de pescadores de Guaraqueçaba é dependente do peixe como
principal fonte de recurso financeiro e alimentar, e a pesca está limitada, por
motivos operacionais, às regiões próximas da comunidade.
10. As espécies demersais declaradas pelos pescadores como freqüentemente
capturadas coincidiram com aquelas mais abundantes nos arrastos e as mesmas,
associadas às pelágicas, podem ser consideradas como importante fonte de
recursos.
116
7. REFERÊNCIAS BIBLIOGRÁFICAS
Abilhôa, V. 1998. Composição e estrutura da ictiofauna em um banco areno-lodoso na Ilha
do Mel, Paraná, Brasil. Dissertação de Mestrado. Pós-Graduação em Zoologia. Setor
de Ciências Biológicas da Universidade Federal do Paraná. 98p.
Abilhôa, V.; Corrêa, M.F.M.; Pinheiro, P. & Lemos, P.H.B. 1997. Composição e estrutura
da ictiofauna em um banco areno-lodoso na Ilha do Mel, Paraná, Brasil. Resumos do
XII Enc. Bras. de Ictiol. FAPESP/IOUSP. 24 a 28 de fevereiro, p. 152.
Agostinho, A.A.; BINI, L.M. & GOMES, L.C. 1997. Ecologia de comunidades de peixes
de área de influência do reservatório de Segredo. In: Agostinho, A.A. & GOMES, L.C.
Reservatório de Segredo; bases ecológicas para o manejo. Maringá: EDUEM. p. 96111.
Alvarez -Guillen, H.; Yãnez-Arancibia, A. & Lara-Dominguez, A.L. 1985. Ecologia de la
Boca del Carmen, Laguna de Térmitos: El habitat y esctructura de las comunidades de
peces. An. Inst. Cienc. del Mar y Limnol. Univ. Aut. del México 12 (1):107-144.
Angulo, R.J. 1992. Geologia da planície costeira do Estado do Paraná. Tese de
Doutorado. USP. 334p
Araújo, F.G.; Cruz Filho, A.G. da; SILVA, M. de A.; Camargo, Filho, P.R. de O.; Jesus,
E.P. de; Simoni, M.R.F.; Duarte, S. & Moffati, M.S. 1997. Estrutura da comunidade de
peixes jovens da margem continental da Baía de Sepetiba, RJ. Resumos do XII
Encontro Brasileiro de Ictiologia. São Paulo. P.182.
Baglo, M. 1983. Les indices de la construcion du barrage de la lagune de cotonore sur les
activités piscicoles dans le systema lagunaire du Sud-Est Beninois. Bul. de Lison du
Comité Interaficon d’Estudes Hydrauliques, v.52. p.47-55.
Barletta,M. & Corrêa,M.F.M. 1989a.Chondrofauna do complexo estuarino da Baía de
Paranaguá e adjacências, PR, BR. Levantamento e produtividade pesqueira. Res. da IV
Reunião do Grupo de Trab. sobre Pesc. e Pesq. de Tubarões e Raias no Brasil.
UFR, Pernambuco, Tamandaré. p.02.
Barletta,M. & Corrêa,M.F.M. 1989b. Condrofauna da Baía de Paranaguá e adjacência, PR.
Levantamento e Produtividade pesqueira. Res. do XVI Congresso Brasileiro de
Zoologia, Paraíba. p.20.
Barletta,M.;Corrêa,M.F.M. & Sunyé,P.S. 1989. First record of occurrence of a female of
Mobula rochebrunei (Vaill.,1879) in western Atlantic Ocean, Paraná State, Brazil.
Resumos do IV Reunião do Grupo de Trabalho sobre pesca e pesquisa de tubarões
e raias no Brasil.
Barletta, M.;Sunyé, P.S.;Dutka-Gianelli, J.A.R. & Abilhôa, V. 1990. Variação nictimeral e
estacional da Gamboa Perequê (Pontal do Sul, Paraná,Brasil). Relatório final.
Conselho de Ciência e Tecnologia do Paraná, Centro de Biologia Marinha,UFPr.
Inédito.
117
Barletta, M. 1999. Seasonal changes of density biomass and species composition of fishes
in different habitats of the Caeté estuary (North Brazilian coast-east Amazon).ZMT
Bremem Contribution 7. 115p.
Baroche, J.M. & Saporta, G. 1982. Análise de dados. Zahar. Rio de Janeiro. 116 p.
Batista, V.S. & Rêgo, F.N. 1996. Análise de associações de peixes, em igarapés do estuário
do Rio Tibiri, Maranhão. Rev. Bras. Biol. 56:p.163-176.
Bazaluk, P. 1996. Ictiofauna de região de manguezal no verão de 1992-1993 na região
da APA de Guaraqueçaba, no município de Guaraqueçaba, Paraná, Brasil.
Monografia de conclusão de Curso de Bacharelado em Ciências Biológicas. PUCPR.
Bazaluk, P.; Godefroid, R.S.; Hofstaetter, M. & Spach, H.L. 1997. Fauna acompanhante da
pesca da manjuba na Baía de Paranaguá, Paraná. Resumos do XII Enc. Bras. de Ictiol.
FAPESP/IOUSP. 24 a 28 de fevereiro, p. 323.
Beverton, R.J.H. & Holt, S.J. 1957. On the dynamics of exploited fish populations. U.K.
Min. Agr. And Fish., Fish. Invest. 2(19):533p.
Bigarella, J.J. (ed.). 1978. A Serra do Mar e a porção oriental do Estado do Paraná:
um problema de segurança ambiental e nacional (contribuição à geografia,
geologia e ecologia regional). Secretaria de Estado do Planejamento/Associação de
Defesa e Educação Ambiental. 248p.
Bigarella, J.J. & Doubeck, R. 1963. Folha geológica de Paranaguá (Estado do Paraná).
Curitiba, Univ. Paraná. Escala 1:50:000.
Bigarella, J.J.; Duarte, M.G. & Becker, R.D. 1970. Structural characteristics of the dune,
foredune, interdune, beach, beach-dune ridge and sand deposits. Bol. Par. Geoc.,
28/29:9-72.
Bochlert, G.W. & Mundy, B.C. 1988. Roles of behavioral and physical factor in larval and
juvenile fish recruitment to estuarine nursery areas. Am. Fish. Soc. Symp. 3:51-67.
Brandini, F. P. 1985. Ecological studies in the Bay of Paranaguá. I. Horizontal distribution
and seasonal dynamics of phytoplankton. Bolm Inst. Oceanogr. S. Paulo 33(2):139147.
Brandini, F.P.; Thamm, C.A. & Ventura, I. 1988. Ecological studies in the Bay of
Paranaguá. III. Seasonal and spatial variations of nutrients and chlorophyll-a. Nerítica
3(1):1-30.
Brandini, F.P. & Thamm, C.A. 1994. Variações diárias e sazonais do fitoplâncton e
parâmetros ambientais na Baía de Paranaguá. Nerítica, 8 (1/2):55-72.
Carvalho, J. de P. 1941. Nota preliminar sobre a fauna ictiológica do litoral sul do Estado
de São Paulo. Bol. Ind. Anim. 4(3):27-81.
Carvalho, J. de P. 1950. Engraulídeos brasileiros do gênero Anchoa. Bol. Inst. Oceanog.
1(2):43-69.
118
Carvalho, J. de P. 1951. Engraulídeos brasileiros do gênero Anchoviella. Bol. Inst.
Oceanogr. 2(1):41-66.
Carvalho, J. de P.; Tommasi, L.R. & Novelli, M.D. 1968. Lista dos linguados do Brasil.
Contr. Inst. Oceanogr. de São Paulo. Oceanogr. Biol. 14:1-26.
Castello, H.P. 1974. Southern distribution limits for Myliobatis goodei
Chondrichthyes, Myliobatidae. Arq. Ciên. Mar. 14 (1):62-63.
(Garman).
Castello, J.P.; Yamaguti,N.; Corrêa, M.F.M. & Ledo, B.S. de. 1994. Oceanografia
Biológica: Nécton. In: Diagnóstico Ambiental Oceânico e Costeiro das Regiões Sul e
Sudeste do Brasil. Vol. 5. PETROBRÁS. 472 p.
Castro-Aguirre, J.L. & Márquez-Espinoza, A. 1981. Contribuicion ao conocimiento de la
ictiofauna de la Isla de Lobos y zona adyacentes. Vera Cruz, México. Sér. Cient. 22:185.
Castro,E.A. & Busetti,E.T. 1985. Helmintofauna de Ariidae na Baía de Paranaguá,Paraná.
Resumos. do XII Congresso Brasileiro de Zoologia, Campinas. p.220.
Cervigón, F. 1966. Los peces mariños de Venezuela. Caracas. Fund. la Salle Cienc. Nat.
Est. Mar. Margarita . Mon. 11-12. 961p.
Cervigón, F. & Lima, H.H. 1972. Las espécies del género Cynoscion (Pisces-Scaienidae)
de la costa Atlántica de América del Sur, desde Colombia hasta Argentina. Contr.
Cient. Univ. Orient. 3:1-46.
Cervigón, F. 1992. Guia de campo de las especies comerciales marinas y de aguas
salobres de la costa septentrional del Sur América. Fichas FAO de identificación de
especies para los fines de la pesca. FAO. Rome. 513p.
Chao, L.N.; Pereira,L.E.; Vieira, J.P.; Benvenuti, M. & Cunha, L.P.R. 1982. Relação
preliminar dos peixes estuarinos e marinhos da Lagoa dos Patos e região costeira
adjacente, Rio Grande do Sul. Atlântica, V.5, n.1, p. 67-75.
Chaves, P. de T. da C. 1994. A incubação de ovos e larvas em Genidens genidens
(Valenciennes) (Siluriformes, Ariidae) da Baía de Guaratuba, Paraná, Brasil. Revta
bras. Zool. 11(4):p.641-648.
Chaves, P. de T. da C. 1995. Atividade reprodutiva de Bairdiella ronchus (Cuvier)
(Pisces, Sciaenidae) na Baía de Guaratuba, Paraná, Brasil. Revta. bras. Zoo.,32(4):
p.759-766.
Chaves, P. de T. da C. & Bouchereau, J.L. 2000. Use of mangrove habitat for reproductive
activity by the fish assemblage in the Guaratuba Bay, Brazil. Oceanologica Acta
vol.23, n.3. p.273-280.
Chaves, P. de T. da C. & Vendel, A L. 1997. Indicadores reprodutivos das espécies de
Citharichtys Bleeker (Teleostei, Pleuronectiformes) na Baía de Guaratuba, Paraná,
Brasil. Revta. bras. Zool, 14(1):73-79.
119
Chaves, P. de T. da C. & Corrêa, M.F.M. 1997. Composição ictiofaunística da área de
manguezal da Baía de Guaratuba, Estado do Paraná, Brasil (25o 52’S; 48o 39’ W). Rev.
Bras. Zool. 15:p.195-202.
Chaves, P.T.; Bouchereau, J.L. & Vendel, A.L. 2000. The Guaratuba Bay, Brazil (25o
52'S; 48o 39'W), in the life cycle of coastal fish species. International Conference
Sustainability of Estuaries and Mangroves Challenges and Prospects. In: Biodiversity
in Estuaries. Universidade Federal Rural de Pernambuco and International Society for
Mangrove Ecosystems. CD-Rom. 8p.
Clark, K.R. & Warwick, R.M. 1994. Change in marine communities: an approach to
statistical analysis and interpretation. Natural Environment Research Council, UK.
144 p.
Chao, L.N.; Pereira, L.E; Vieira, J.P., Benvenutti, M.A. e Cunha, L.P.R.. 1982. Relação
preliminar dos peixes estuarinos e marinhos da Lagoa dos Patos e região costeira
adjacente, Rio Grande do Sul, Brasil. Atlântica, 5:67-75.
Chao, L.N.; Pereira, L.E. ; Vieira, J.P. 1985. Estuarine Fish Community of the Patos
Lagoon, Brazil: a baseline study. In:Yánez-Arancibia, A. (ed). Fish Community
Ecology in Estuaries and Coastal Lagoons: towards an ecosystem integration.
UNAM:429-450.
Corrêa, M.F.M. 1987. Ictiofauna da Baía de Paranaguá e adjacências. Levantamentos e
Produtividade. Dissertação de Mestrado. Departamento de Zoologia. Universidade
Federal do Paraná. 465p.
Corrêa, M.F.M. 1988. Ictiofauna da Baía de Paranaguá e adjacências (litoral do Estado do
Paraná-Brasil). Res. do XV Congresso Brasileiro de Zoologia, Curitiba. p.344.
Corrêa, M.F.M. 1994. Nécton. Estado do Paraná. In: Petrobrás. Diagnóstico ambiental
oceânico e costeiro das regiões sudeste e sul do Brasil. Vol. IX. p. 33-59.
Corrêa, M.F.M.; Absher, T.M.; Barletta, M.; Dutka-Gianelli, J.A.R.; Bonatti, G.M.G.
1987. Produtividade pesqueira para a região da APA de Guaraqueçaba, PR, BR.
IPARDES/IBAMA. Relatório final. 15p. Inédito.
Corrêa, M.F.M.; Absher, T.M.; Barletta, M.; Dutka-Gianelli, J.A.R.; Bonatti, G.M.G.
1995. Fauna Aquática. In: IPARDES, Diagnóstico Ambiental da APA de
Guaraqueçaba..p.73-83.
Corrêa, M.F.M.; Cerdeiras; P.C.R. & Pieckzarka,J.C. 1988. Levantamento ictiológico do
Rio Guanandi (25o 30'25''S e 45o 45'50''W), Sub-bacia do Rio Nhundiaquara
(Morretes,PR,Brasil). Nerítica, Pontal do Sul, 3(1):37-60.
Corrêa, M.F.M.; Cordeiro, A.A.M. & Justi, I. 1986. Catálogo dos peixes marinhos da
coleção de zoologia da divisão de Geologia e Zoologia da Prefeitura Municipal de
Curitiba. Nerítica, Pontal do Sul 1(1):1-83p.
120
Corrêa, M.F.M. & Maehama, O.K. 1988. Composição e variação temporal da ictiofauna
para as regiões entre as Ilhas do Mel e da Cotinga e Foz do Rio do Maciel (Baía de
Paranaguá, Paraná, Brasil). Resumos do XV Congresso Brasileiro de Zoologia,
Curitiba. p.345.
Corrêa, M.F.M.; Pinheiro, P.C.; Lemos, P.H.B. & Pinto, E.F. 1997. Diagnóstico da pesca
da APA de Guaraqueçaba. In: Zoneamento Econômico-Ecológico da APA de
Guaraqueçaba. Relatório final. IPARDES/IBAMA/FUNPAR. 79p. No prelo.
Costa, F. 1980 Complexo Estuarino Mundaú-Manguaba. Documentário da Ictiofauna. In:
Projeto Levantamento Ecológico Cultural-PLEC. Segunda Etapa. 195p.
Couto, E.C.C. & Corrêa, M.F.M. 1992.Revisão e discussão de trabalhos sobre as espécies
íctíicas da costa paranaense. Resumos. UFRJ.
Cunha, L.H.O.; Rougelle, M.D.; VonBehr, M.F. & Diegues, A.C.S. 1989. Comunidades
litorâneas e unidades de proteção ambiental: convivência e conflitos. O caso de
Guaraqueçaba (Baía de Paranaguá, BR). Série Estudos de Casos 2. Prog. de Pesq. e
Cons. de Áreas Úmidas no Brasil, USP. São Paulo.
Cunha,L.P.R. 1988. Aspectos da ictiofauna das zonas de arrebentação de praias do litoral
do Paraná e do Rio Grande do Sul. Res. do XV Congresso Brasileiro de Zoologia,
Curitiba. p.346.
Davis, T.L.O. 1988. Temporal chances in the fish fauna entering a tidal swamp system in
tropical Australia. Environ. Biol. Fishes, vol. 21., n3., p.161-171.
Darnell, R.M. & Soniat, T.M. 1979. The estuary/continental shelf as an interactive
system. In: Livingston, R.J. (ed.) Ecological processes in Coastal and Marine
Systems. Plenum Press Publ. Corp. N.Y. 548p.
Disaró, S.T. 1995. Associações de foraminíferos da Baía das Laranjeiras, complexo
estuarino Baía de Paranaguá, Paraná, Brasil. Dissertação de Mestrado. Pós-Graduação
em Zoologia do Setor de Ciências Biológicas da Universidade Federal do Paraná. 76p.
Drunkers, J. 1988. Inshore/offshore water exchange in shallow coastal systems. In:
Jansson, B.O. (ed.). Coastal-Offshore Ecosystem interactions. Lectures Notes on
Coastal and Estuarine Studies 22. Springer Verlag, Berlin. 368p.
D’Incao, F. 1973. Sobre ocorrência de Gobionellus oceanicus (Pallas, 1770) no sul do
Brasil (Pisces, Gobiidae). Comun. Mus. Ci. PUCRGS (2):1-20.
FAO/UNEP 1980. The present state of mangrove ecosystems in Southeast Asia and the
impact of pollution: Singapore. Zoology Department of University of Singapore. Rept.
S.C.S./80/WP. 94.
Dutka-Gianelli,J.A.R. & Corrêa,M.F.M. 1988. Dinâmica de crescimento e reprodução de
Cynoscion leiarchus (Pisces-Sciaenidae) para a região de Guaraqueçaba, Paraná, Brasil.
Res. do XV Congresso Brasileiro de Zoologia, Curitiba. p.328.
121
Faleiro, L.C.C. 1971. Primeira ocorrência de Ranzania truncata ( Retzius, 1758) na costa
do Rio Grande do Sul, Br. (Tetraodontiformes, Tetraodontoidei, Milidae). Comun.
Mus. Cien. PUCRS. Zool. 1-12.
Fernandes-Pinto, E.; Corrêa, M.F.M.; Pinheiro, P.C. & Lemos, P.H.B. 1997. Composição,
estrutura e distribuição temporal da ictiofauna da região da Enseada do Benito,
Guaraqueçaba, Paraná. Resumos do XII Enc. Bras. de Ictiol. FAPESP/IOUSP. 24 a 28
de fevereiro, p. 153.
Figueiredo,J.L. 1977. Manual de peixes marinhos do sudeste do Brasil. I. Introdução.
Cação,raias e quimeras. Museu de Zoologia da Universidade de São Paulo. 104p.
Figueiredo, J.L. 1981.Estudo das distribuições endêmicas de peixes da Província
Zoogeográfica Marinha. Argentina . Tese de doutorado, Universidade de São Paulo.
Instituto de Biociências. 121p.
Figueiredo, J.L. & Menezes, N.A. 1978. Manual de peixes marinhos do sudeste do
Brasil. II. Teleostei (1). Museu de Zoologia. Universidade de São Paulo. 110p.
Figueiredo, J.L. & Menezes, N.A. 1980. Manual de peixes marinhos do sudeste do
Brasil. III. Teleostei (2). Museu de Zoologia. Universidade de São Paulo. 90 p.
Figueiredo, G.M.; Garcia, A.M.; Resende, S.M. & Vieira, J.P. 1995. Variação nictimeral
de abundância e tamanho da assembléia de peixes do estuário da Lagoa dos Patos, RS.
Resumos do XI Encontro Brasileiro de Ictiologia. Campinas (SP). p.7.
Fischer, W. (ed.) 1978. FAO species identification sheets for fishery purposes. Western
Central Atlantic. (Fishing Area 31). Rome, FAO, 1978. p.v.
Fowler, H.W. 1941. A list of the fishes known from the coast of Brazil. Arq. Zool.
3(4):115-184.
Furia, R.R. 1996. Composição das capturas de peixes na Enseada das Palmas-Ilha
Anchieta, Ubatuba-SP. Variabilidade Espaço-Temporal. Dissertação de Mestrado.
Instituto Oceanográfico. Universidade de São Paulo.
Giannini, R. & Paiva Filho, A.M. 1990. Os Sciaenidae (Teleostei : Perciformes) da Baía de
Santos(SP), Brasil. Bolm. Inst. Oceanogr., São Paulo, 38(1):69-86.
Gomez ,E.D. 1980. In: FAO/UNEP 1980. The present state of mangrove ecosystems in
Southeast Asia and the impact of pollution: Singapore. Zoology Department of
University of Singapore. Rept. S.C.S./80/WP. 94.
Gomes-Bonatti,G.M. 1988. Reprodução em Anchoa spinifer (Teleostei- Engraulidae) para
a Baía de Guaraqueçaba (Baía de Paranaguá, Paraná, Brasil). Res. do XV Congresso
Brasileiro de Zoologia, Curitiba. p. 276.
Godefroid, R.S.; Hofstaetter,M. & Spach, H.L.1997a . Levantamento icitiofaunístico da
zona de arrebentação da praia de Pontal do Sul, Paraná. Resumos do XII Enc. Bras. de
Ictiol. FAPESP/IOUSP. 24 a 28 de fevereiro, p. 186.
122
Godefroid, R.S.; Hofstaetter,M. & Spach,H.L. 1997b . Estrutura temporal da ictiofauna da
Praia de Pontal do Sul, Paraná. Resumos do XII Enc. Bras. de Ictiol. FAPESP/IOUSP.
24 a 28 de fevereiro, p. 187.
Haedrich, R.L. 1983. Estuarine fishes. In: Ketchum, B.H., ed. Estuaries and Enclosed
Seas. Elsevier Scientific. 183p.
Hamilton,L.S. & Snedacker, S.C. (ed.)1984.
management. UNESCO, Paris. 12. 123p.
Handbook
for
mangrove
area
Hays, W. L. (1988). Statistics (4th ed.). New York: CBS College Publishing.
Haimovici, M. Vooren, C.M. & Catello, J.P. 1994. Nécton do Rio Grande do Sul. In:
Castello, J.P. (Coord.) Diagnóstico ambiental oceânico e costeiro das regiões Sul e
Sudeste do Brasil; Oceanografia Biológica, Nécton. Petrobrás. Rio Grande, v.5., p.81129.
Hill, M.O. 1973. Diversity and evenness: A unifying notation and its consequences.
Ecology 54: p. 427-432.
Hook, J.H. 1991. Seasonal variation in relative abundance and species diversity or fishes in
South Bay. Contributions in Marine Science, v.32, p.127-141.
Hopkinson, C.S. 1988. Patterns of organic carbon exchange between coastal ecosystems;
the mass balance approach in salt marsh ecosystem. In: Jansson,B.O. (ed.) CoastalOffshore Ecosystems interactions. Lectures Notes on Coastal and Estuarine Studies
22. Springer Verlag, Berlin. 368p.
Instituto Agronômico do Paraná (IAPAR). 1978. Cartas climáticas básicas do Estado do
Paraná. Londrina.
Instituto Nacional de Pesquisas Hidroviárias (INPH). 1988. Relatório de apresentação
das medições meteorológicas observadas em Pontal do Sul, Paranaguá (1982 a
1986). Inédito.
Instituto Paranaense de Desenvolvimento Econômico e Social (IPARDES). 1989a.
Zoneamento do litoral paranaense. Convênio SEPL/IPARDES,Curitiba, PR. 175p.
Instituto Paranaense de Desenvolvimento Econômico e Social (IPARDES). 1989b. APA de
Guaraqueçaba. Caracterização sócio-econômica dos pescadores artesanais e
pequenos produtores rurais. IPARDES, Curitiba, PR. 87p.
Instituto Paranaense de Desenvolvimento Econômico e Sócia.l (IPARDES). 1990. MacroZoneamento da Área de Proteção Ambiental de Guaraqueçaba. Volume I.
IPARDES. 254p.
Instituto Paranaense de Desenvolvimento Econômico e Social (IPARDES). 1995.
Diagnóstico Ambiental da APA de Guaraqueçaba. IPARDES. 166 p.
123
Instituto de Pesquisas da Marinha (IPqM). 1969. Estudo das correntes na Baía de
Paranaguá. Arquivos do Porto de Paranaguá. Inédito, p.v.
Jansson, B.O. (ed.) 1988. Coastal-Offshore Ecosystem interactions. Lectures Notes on
Coastal and Esturine Studies 22. Springer Verlag, Berlin. 368p.
Jardim, L.F.A. 1973. Sciaenideos marinhos da costa do Brasil sul. (AcanthopterygiiPerciformes-Scaienidae). Com. Mus. Ci. PUCRGS. Zoo. 3:1-64.
Johnson, G.D. & Wichern, D.W. 1992. Applied multivariate statistical analysis. Pent.
Hall. 642 p.
Karam, K.F. & Toledo,V. 1997. Caracterização sócio-econômica e cultural da população
tradicional da APA de Guaraqueçaba. In: Zoneamento Ecológico Econômico da APA
de Guaraqueçaba. Relatório final. Convênio IPARDES/IBAMA. Inédito. 103p.
Kennish, M. J. 1990. Ecology of Estuaries. CRC Press, Inc. 391p.
Kjerfve, B. 1986. Comparative oceanography of coastal lagoons. In: Wolfe, D.A. (Ed.)
Estuarine variability. Ac. Press Inc. 510p.
Kjerfve, B. & Magill, K.E. 1989. Geographic and hydrodynamic characteristics of shallow
coastal lagoons. Mar. Geol. 88:187-199.
Knoppers, B.A. & Opitz, S. 1984. An annual cycle of particulate organic matter in
mangrove waters. Laranjeira Bay, Southern Brazil. Arq. Biol. Tecnol. 27(1):79-93.
Knoppers, B.A.; Brandini, F.P. & Thamm, C.A. 1987. Ecological studies in the Bay of
Paranaguá II. Some physical and chemical characteristics. Nerítica 2(1):1-36.
Kraemer,M.C. de 1982. Malhas da pobreza: exploração do trabalho de pescadores
artesanais na Baía de Paranaguá. Instituto Hist., Geog. e Etnográfico Paranaense.
Estante Paranista, 22. 168p.
Lana, P. da C. 1986. Macrofauna bêntica de fundos sublitorais não consolidados da Baía
de Paranaguá (Paraná). Nerítica, Pontal do Sul, v.1., n.3., p:79-89.
Lao, P.N. 1983. Mangrove resource management. Fish. Div. M.A.V., S.U.V.A. Tech.
Rept. 5. 23p.
Laroche, J. & Baran, E.; Rasoanandrasana, N.B. 1997. Temporal patterns in a fish
assemblage of a semarid mangrove zone in Madagscar. J. Fish. Biol. 51:p.3-20.
Lauff,G.H. 1967. Estuaries. American Assoc. for Advancement on Science. Publ. 83.
101p.
Legendre, L. & Legendre, P. 1983. Numerical Ecology. Elsevier Scientific Publ.
Company.-New York. 217 p.
124
Lema, T. de ; Lucena, C.A.S. de; Saenger, S. & Oliveira, M.F.T. 1979. Primeiro
levantamento dos tetraodontiformes do sul do brasil, Uruguai e Argentina (Teleostei,
Acanthopterygii). Comu. Mus. Ci. PUCRGS. 20: 1-84.
Lema, T. de; Lucena, C.A.S. de & Lucena, Z.M.D. 1980 Novas adendas à ictiofauna
marinha do extremo sul do Brasil. Iheringia. Sér. Zool. 56:103-120.
Lemos, P.H.B.; Hofstaetter, M. & Godefroid, R.S. 1997. Variação sazonal e composição
alimentar da ictiofauna na zona de arrebentação de Pontal do Sul, Paraná, Brasil.
Resumos do XII Enc. Bras. de Ictiol. FAPESP/IOUSP. 24 a 28 de fevereiro, p. 156.
Livingston, R.J. 1976. Diurnal and seasonal fluctuations of organisms in North Florida
estuary. Estuar. Processes. Acad. Press. 326p.
Lucena, C.A.S. & Lucena, Z.M.S. 1981a. Catálogo dos peixes marinhos do Mus. de
Ciências da Pont. Univ. Católica do Rio Grande do Sul. Elasmobrachiomorphii.
Teleostomi (1a. Parte). Porto Alegre. Com. Mus. Cienc. PUCRGS 21:1-66.
Lucena, C.A.S & Lucena, Z.M.S. 1981b.Catálogo de peixes marinhos do Mus. de
Ciências da Pont. Univ. Cat. do Rio Grande do Sul. Teleostomi (final). Com. Mus.
Ciências PUCRGS, 25:1-80.
Ludwig, J.A. & Reynolds, J.F. 1988. Statistical ecology. A primer methods and
computing. John Wiley & Sons. 337p.
Lyons, J. 1996. Patterns in the species composition of fish assemblages among Wisconsins
streams. Environ. Biol. Fishes, v.4, p.329-341.
Maack, R. 1981.Geografia física do Estado do Paraná. Curitiba. BDT/UFPR/IBPT.
350p.
Machado, E. da C.; Daniel, C.B.; Brandini, N. & Queiroz, R. L.V. de. 1997. Temporal and
spatial dynamics of nutrients and particulate suspended matter Paranaguá Bay, Pr,
Brazil. Nerítica. No prelo.
MacConnell, R.H. 1999. Estudos ecológicos de comunidades de peixes tropicais. Edusp.
534p.
MacFarland, W.N. 1963. Seasonal change in the number and the biomass of fishes from
the surf at Mustang Island, Texas. Publ. Inst. Mar. Sci. Univ. Tex. v.9, p.91-105.
Maciel, N.A.L. 1995. Estudos sobre a composição distribuição, abundância e diversidade
da ictiofauna de três enseadas na região litorânea de Ubatuba, Estado de São Paulo,
Brasil. São Paulo. Dissertação (Mestrado em Oceanografia Biológica)- Instituto
Oceanográfico, Universidade de São Paulo. 141p.
Madden, C.J.; DAY Jr, J.W. & Randall, J.M. 1988. Freshwater and marine coupling in
estuaries of the Mississippi River deltaic plain. Limnol. Oceanogr. 33(4):982-1004.
125
Maehama,O.K.M. & Corrêa,M.F.M. 1987. Composição ictiofaunística para zona de
arrebentação de Pontal do Sul
à Praia de Leste (Litoral de Paraná, Brasil). Res. do
XIV Congresso Brasileiro de Zoologia, Juiz de Fora. p. 231.
Manly, B.F.J. 1998. Multivariate Statistical Methods. A primer. Chapman & Hall. 215p.
MkLusky, D.S. 1974. Ecology of estuaries. Heinemann Educational Books. London.
144p.
Marterer, B.E.L.A. 1986. Peixes de gênero Mugil da Baía de Guaratuba, Paraná. Resumos
do XVII Congresso Brasileiro de Zoologia, Cuiabá, p.111.
Marone , E. & Camargo, R. 1994. Marés meteorológicas no litoral do Estado do Paraná; o
evento de 18 de agosto de 1993. Nerítica, 8 (1/2):73-85.
Marone, E.; Guimarães, M.R.; Pratta,V.P.; Klingenfus, M.E. & Camargo, R. 1995.
Caracterização física das condições oceanográficas, meteorológicas e costeiras das
zonas estuarinas da Baía de Paranaguá, Brasil. VI. Cong. Latino Americano de
Ciencias del Mar, Mar del Plata, Argentina. Setembro.
Martin, L.; Suguio, K; Flexor, J.M. & Azevedo, A.E.G. 1988. Mapa geológico do
Quaternário costeiro dos Estados do Paraná e Santa Catarina. Série Geol. DNPM.
Brasília, 28. 40p. 2 mapas.
Matthews, W.J.; Hough, D.J. & Robison, H.W. 1992. Similarities in fish distribution and
water quality patterns in streams of Arkansas: congruence of multivariate analyses.
Copeia, n.2., p.296-303.
Melo, P.; Hofstaetter, M.; Godefroid, R.S. & Spach, H.L. 1997. Composição específica e
estrutura em tamanho das capturas de engraulídeos na Baia de Paranaguá, Pr. Resumos
do XII Enc. Bras. de Ictiol. FAPESP/IOUSP. 24 a 28 de fevereiro, p. 332.
Menezes,N.A. & Figueiredo,J.L. 1980. Manual de peixes marinhos do sudeste do
Brasil. IV. Teleostei (3). Museu de Zoologia. Universidade de São Paulo. 96p.
Menezes,N.A. & Figueiredo,J.L. 1985. Manual de peixes marinhos do sudeste do
Brasil. V. Teleostei (4). Museu de Zoologia. Universidade de São Paulo. 105p.
Monteiro Neto, C.; Blacher, C.; Laurent, A.A.S.; Snizek, F.N. Canozzi, M.B. & Tabajara,
L.L.C.A. 1990. Estrutura da comunidade de peixes de águas rasas na região de Laguna,
Santa Catarina, Brasil. Atlântica, Rio Grande, v.12, n.2., p:53-69.
Natali Neto, J.F. 1994. Ictiofauna do ecossistema marinho de Ubatuba (23o 36'-24o22'S;
44o33'-45o08'W), SP-Brasil, entre 50 e 100 metros de profundidade: composição,
distribuição, abundância e diversidade. Dissertação de Mestrado. Universidade de São
Paulo. Instituto Oceanográfico. 2v.
Nelson, J.S. 1976. Fishes of the world. New York: Wiley Interc. Public. 416p.
Nelson, J.S. 1994. Fishes of the world. New York: Wiley Interc. Public. 600p.
126
Nitrouer, C.H.A. & DeMaster, D.J. 1986. Sedimentary processes on Amazon Continental
Shelf. Cont. Shelf Res. 6(1/2):380p.
Nikolski, G.V.1963. The ecology of fishes. Academic Press. 1963. 532p.
Nonato, E.F.; Amaral, A.C.Z. & Figueiredo, J.L. 1983. Contribuição ao conhecimento da
fauna de peixes do litoral norte do Estado de São Paulo. Bol. Inst. Oceanogr. 32(2):
143-152.
NTSYS-pc. 1992. Numerical taxonomy and multivariate analysis system. V. 1.7. New
York. Applied Biostatistics Inc.
Odum, W.E., Fisher, J.S. & Pickral, J.C. 1979. Factors controlling the flux of particulate
organic carbon from the estuarine wetlands. In: Livingston, R.J. (ed.). Ecological
processes in Coastal and Marine Systems. Plenum Press Pub. Corp. 548p.
Oliveira, A.M.E. de 1976. Composição e distribuição da ictiofauna, nas águas estuarinas
do Rio Jaguaribe (Ceará-Brasil). Arq. Cienc. Mar. 16(1):9-18.
Ottmann, V.; Seara, C.A.; Godefroid, R.S. & Hoffstaeter, M. 1992. A pesca da manjuba da
Baía de Guaraqueçaba e Laranjeiras. Resumos da V Semana Nacional de
Oceanografia. FURG, Rio Grande. 18 a 23 de outubro.
Paiva, M.P. 1996. Recursos Pesqueiros. In: Levantamento do estado da arte da pesquisa
dos recursos vivos marinhos do Brasil. Min. do Meio Ambiente, dos Recursos
Hídricos e da Amazônia Legal- MMA/Sec. de Coordenação dos Assuntos do Meio
Ambiente - SMA. Relatório Final do Programa Revizée. No prelo.
Paiva Filho, A.M. & Toscano, A.P. 1987. Estudo comparativo e variação sazonal da
ictiofauna na zona entremarés do Mar Casado –Guarujá e Mar Pequeno – São Vicente,
São Paulo. Bol. Inst. Oceanogr. , São Paulo, v.35, n2., p.153-165.
Pauly, D. 1979. Theory and management of tropical multispecies stocks. Review, with
emphasis on Southeast Asian demersal fisheries. ICLARM Studies & Reviews 1: 35 p.
Pauly, D. 1983. Some simple methods for the assessment of tropical fish stocks. FAO
Fish. Techn. Paper 234.
Pereira, L.E. 1994. Variação diurna e sazonal dos peixes demersais na barra do estuário da
Lagoa dos Patos, RS. Atlântica, Rio Grande, v.16, p.5-21.
Pinheiro, P.C. 1999. Dinâmica das comunidades de peixes em três áreas amostrais da Ilha
do Mel, Baía de Paranaguá, Paraná, Brasil. Dissertação de Mestrado. Setor de
Ciências Biológicas, Universidade Federal do Paraná. 171p.
Pinheiro, P.C.; Corrêa, M.F.M. & Spach, H.L. 1994. Caracteres consistentes para
identificação de larvas e juvenis de Anchoa parva (Osteichthyes-Engraulidae). Arq.
Biol. Tecnol. 37(3):747-759.
Pinheiro, P.C.; Corrêa, M.F.M.; Spach, H.L.; Lemos, P.H.B. & Pinto, E.F. 1997.
Composição, variabilidade temporal e espacial da ictiofauna da Ilha do Mel, Pontal do
127
Sul, Paraná. Resumos do XII Enc. Bras. de Ictiol. FAPESP/IOUSP. 24 a 28 de
fevereiro, p. 154.
Pinto, S.I. 1960. Um novo Gobiidae do Estado da Bahia, Brasil (ActinopterygiiPerciformes). Bol. Mus. Nac. 218:p.1-9.
Peterson, D.H.; Hager, S.W. & Schemel, L.E. 1988. Riverine C, N, Si and P transport to
the coastal ocean: an overview. In: Jansson,B.O. (ed.) Coastal-Offshore Ecossystem
interactions. Lectures Notes on Coastal and Estuarine Studies 22. Springer Verlag,
Berlin. 368p.
Reis, E.G. 1986. Reproduction and feeding habitats of the marine catfish Netuma barba
(Siluriformes, Ariidae) in the estuary of Lagoa do Patos, Brazil. Atlântica, Rio Grande,
v.8. p.35-55.
Rholf, F. J. 1989. Numerical taxonomy and multivariate analysis system. Exeter
Publishing Ltd. p.v.
Ribeiro, A.M. 1907. Fauna Brasiliense. Peixes II (Desmobranchios). Arq. Mus. Nac.
14:131-213.
Ribeiro, A.M. 1911. Fauna Brasiliense. Peixes IV. (Eleutherobranchios, Aspirophorus).
Arq. Mus. Nac. 16:1-504.
Ribeiro, A.M. 1915. Fauna Brasiliense. Peixes V. (Eleutherobranchios, Aspirophorus).
Arq. Mus. Nac. 17:1-679.
Ricklefs, R.R. & Schluter, D. 1993. Species Diversity in Ecological Communities.
Historical and Geographical Perspectives. The University of Chicago Press. 416p..
Rivereau, J.C. ; Fuck, R.A.; Muratori, A. & Trein, E. 1969. Paranaguá, folha geológica.
Curitiba, Comissão da Carta Geológica do Paraná. Escala 1:70000.
Robins, C.R. 1971. Distributional patterns of fishes from coastal and shelf waters of the
tropical Western Atlantic. In: Symposium on Investigations Resources of the Caribbean
Sea and Adjacent Regions. Papers on Fisheries Resources. Roma, FAO. p.249-255.
Rocha, G.R.A. 1990. Distribuição, abundância e diversidade da ictiofauna na região de
Ubatuba – SP (23o 20'S- 24o 00'S; 44o 30'W- 45o 30'W), Brasil. Dissertação de
Mestrado. Instituto Oceanográfico.Universidade de São Paulo. 2v.
Rossi-Wongtschowski, C.L. & Paes, E.T. 1993. Padrões espaciais da comunidade de
peixes demersais do litoral norte do Estado de São Paulo-Ubatuba, Brasil. In: PiresVanin, A.M.S. (ed.) Estrutura e função do ecossistema de plataforma continental do
Atlântico sul brasileiro. Publção esp. Inst. Oceanogr.São Paulo (10):169-188.
Rougeulle, M.D. 1993. La crise de la peche artisanale: transformation de léspace et
destructuraton de l’activite- Le case de Guaraqueçaba (Paraná, Brésil). These de
Doctorat de L’Universite de Nantes, UFR Geographie. 410p.
128
Roux, C. 1973. Campagne de la Calypso au large des coutes atlantiques de l’Amerique du
Sud (1961-1962). I. Poissons Tèleostéans du Plateau Continetal Brésilien. Ann. Inst.
Oceanogr. 49 (fasc. suppl.):1-207.
Saenger, P. & McIvor, C.C. 1975. Water quality and fish populations in a mangrove
estuary modified by residential and development. In: Walsh, J. ; Snedacker, S. & Tess,
H. (Ed.) Proceedings of the international Symposium and Biology and
Management of Mangroves. Un. of Florida. p. 753-757.
Saila, S.B. 1983. Some aspects of fish production and cropping. In: Cronin, L. E., ed.
Estuarine Systems. Estuarine Research. Academic Press, New York. Vol.1. 473p.
Sampaio, I.B.M. 1998. Estatística aplicada à experimentação animal. Fundação de
Ensino e Pesquisa em Medicina Veterinária e Zootecnia. Belo Horizonte. 221p.
Santos, E. 1970. Nossos Peixes Marinhos (vida e costumes dos peixes do brasil). Rio de
Janeiro. Zool. Brasilica. F. Briget. 256p.
Santos, E.P. 1978. Dinâmica de populações aplicada a pesca e piscicultura. São Paulo.
HUCITEC/EDUSP. 129p.
Schluter, D.& Ricklefs, R.E. 1993. Species diversity. An introduction to the problem. In:
Ricklefs, R. E. & Schluter, D. 1993. Species Diversity in Ecological Communities.
Historical and Geographical Perspectives. The University of Chicago Press. 1-10.
Soares, C. R. & Barcelos, J. H. 1995. Considerações sobre os sedimentos do fundo das
Baías das Laranjeiras e de Guaraqueçaba. Complexo estuarino da Baía de Paranaguá
(Paraná, Brasil). Bol. Par. de Geociências 43. p.41-60.
Schultz, L.P. 1949. A further contribution to the ichthyology of Venezuela. Proc. Unit.
Stat. Nat. Mus. 99 (3235): 1-21.
Seaberg, W.C. 1988. Observations on the inlet flow patterns derived from numerical and
physical modeling studies. Am. Fish. Soc. Symposium. 3:16-25.
Shaw, R.F.; Rogers, B.D.; Cowan Jr.; J.H. & Herke, W.H. 1988. Ocean-estuary coupling
of ichthyoplancton and nekton in the Northern Gulf of Mexico. Am. Fish. Soc.
Symposium 3: 77-89.
Sheridan, P.F. & Livingston, R.J. 1979. Cyclic trophic relationships of fishes in an
unpolluted, river-dominated estuary in north Florida. In: Livingston, R.J., ed.
Ecological Processes in Coastal and Marine Systems. Plennum Press, New York.
143p.
Silva, M.H.C. 1989. Ictiofauna. In: Almeida, M.V.O; Conti, L.M.P.; Couto,E.C.G.;
Freitas,C.A.F.; Lopes,M.J.S. & Silva,M.H.C. Estudo integrado da foz da Gamboa do
Maciel (Paranaguá, Paraná) durante dois ciclos de maré. Dissertação de Pósgraduação "Latu sensu". Centro de Biologia Marinha, Universidade Federal do
Paraná,Pontal do Sul. p.74-117.
129
Silva,M.H.C. & Pose,L.M. 1990. Composição da ictiofauna da Foz da Gamboa Maciel
(Paranaguá,Paraná) durante dois ciclos de maré. Res. do XVII Congresso Brasileiro
de Zoologia, Londrina. p.338.
Silveira, B.; Hoffstaetter,M.; Godefroid,R.S. & Spach, H.L. 1997. Ectoparasitas de larvas
de peixes das Baías das Laranjeiras e de Guaraqueçaba. Resumos do XII Enc. Bras. de
Ictiol. FAPESP/IOUSP. 24 a 28 de fevereiro, p. 247.
Sociedade Paranaense de Vida Selvagem (SPVS). 1992. Plano integrado de conservação
para a região de Guaraqueçaba, Paraná, Brasil. SPVS/Nature Conservancy,
Curitiba. 2v.129p.
Sokal, R. R. & Rohlf, F.J. 1981. Biometry. W. H. Freeman and Company, 859 p.
Souza,J.R.B. 1988. Levantamento preliminar de peixes juvenis na zona de arrebentação da
Praia de Pontal do Sul, Paraná, Brasil. Relatório da disciplina de Chordata.
Departamento de Zoologia. Universidade Federal do Paraná. Inédito.
Spach, H.L.; Godefroid, R.S. & Hofftaetter, M. 1995 a. Pesca da manjuba (Anchoa parva,
A. tricolor e A. lyolepis, Cetengraulis edentulus e Engraulis anchoita) no litoral do
Paraná. IX Congresso Brasileiro de Engenharia de Pesca. São Luiz do Maranhão.
Spach, H.L.; Hofftaetter, M. & Godefroid, R.S. 1995 b. Estrutura etária das capturas de
manjuba (Anchoa parva e A. tricolor) no estuário da Baía de Paranaguá. I. Adequação
dos protocolos de laboratório. IX Congresso Brasileiro de Engenharia de Pesca. São
Luiz do Maranhão.
StatSoft, Inc. 1997. STATISTICA for Windows [Computer program manual]. Tulsa,
OK: StatSoft, Inc., 2300 East 14th Street, Tulsa, OK 74104, email: [email protected],
WEB: http://www.statsoft.com.
Stevenson, J.C. 1988. Comparative ecology of submersed grass beds in freshwater,
estuarine, and marine environments. Limnol. Oceanogr. 33(4):867-893.
Travassos, H. & Lima, H. de H. (1975). Contribuição ao estudo dos Carangídeos do Brasil.
Arq. Cienc. Mar. 15 (1):33-40.
Travassos, H. & Paiva, M.P. 1957. Lista dos Sciaenidae brasileiros contendo chave de
identificação e proposta de nomes vulgares oficiais. Bol. Inst. Oceanogr. S. Paulo. 8(12):139-170.
Travassos, H. & Rego-Barros, R. 1971. Lista das espécies brasileiras da família Sciaenidae
Owen, 1846 (Perciformes, Percoidei). Arq. Cienc. Mar. 11(2):59-71.
Underwood, A.J.1998. Experiments in ecology. Cambridge University Press. 504p.
UNESCO 1979. Committee for the preparation of CLOFETA. Paris. UNESCO Tec. Pap.
Mar. Sci. 29:1-55.
UNESCO 1980. Preliminary list of marine fishes. Paris. Mar.Inf. 31:1-49.
130
Vazzoler,G. 1970. Ictiofauna da Baía de Santos. I. Sciaenidae. Percoidea. Percomorphii.
Bol. Inst. Oceanogr. 18(1):11-26.
Vazzoler, G. 1975.Distribuição da fauna de peixes demersais e ecologia dos Sciaenidae da
plataforma continental brasileira, entre lat. 29?21’S (Torres) e 31?41’S (Chuí). Bol.
Inst. Oceanogr. 24:85-169.
Vazzoler, A.E.A. de M. 1982. Manual de Métodos para estudos biológicos de
populações de peixes. Reprodução e crescimento. Brasília. CNPq. 106p.
Vazzoler, A.E.A. de M. 1996. Biologia da reprodução de peixes teleósteos: teoria e
prática.Maringá.EDUEM/SBI/CNPq/NUPELIA. 169p.
Vazzoler, A.E.A.M.;Zaneti-Prado, E.M.;Kawakami, E. & Yamaguti, N. 1982. Teleósteos
marinhos coletados entre Cabo Frio (23o S) e Torres (29o 21'S) no programa Faunec
(1975). Dusenia 13(3):p. 127-133.
Vendel, A.L. 1997. Dinâmica e estrutura da assembléia de peixes da gamboa do Baguaçú,
Baía de Paranaguá, Paraná. Monografia de Bacharelado, Ciências Biológicas,
Universidade Federal do Paraná. p.v.
Vieira,J.P. & Musick,J.A. 1993. Latitudinal patterns in diversity of fishes in warmtemperate and tropical estuarine waters of the western Atlantic. Atlântica, Rio Grande,
15:115-133.
Vieira,J.P. & Musick, J.A. 1994. Fish faunal composition in warm-temperate and tropical
estuaries of western Atlantic. Atlântica, Rio Grande:16:31-53.
Weinstein, M.P. (ed.) 1988. Larval fish and shellfish transport through inlets. Am. Fish.
Soc. Symp. 3. 166p.
Wisniewski, C.; Rodrigues, A. S. & Miranda, M. 1997. Diagnóstico do clima e
caracterização sócio-econômica da agricultura e dos sistemas agrícolas. In:
Zoneamento econômico-ecológico da APA de Guaraqueçaba. IPARDES/IBAMA.
No prelo. 46p.
Wooton, R.J. (Ed.) 1998. Ecology of teleost fishes. Kluwer Academic Publishers. Fish and
Fisheries Series 24. 386p.
Yamaguti, Y.; Saccardo, S.A.; Alvares, L.M. de C. & Muto, Y. 1994. Nécton dos Estados
do Rio de Janeiro e São Paulo. In:Castello, J.P. (Coord.). Diagnóstico ambiental
oceânico e costeiro das regiões Sul e Sudeste do Brasil; Oceanografia Biológica:
Nécton. Rio Grande, V.5, p.15-32.
Yánez, L.A. & Nugent,R.S. 1977. El papel ecológico de los peces en lagunas costeras. An.
Centro. Mar. Limnol. UNAM 4(1):1-107.
Yãnez-Arancibia, A.; Lara-Dominguez,, A.L. & Alvarez-Guillen, H. 1985. Fish
community ecology and dynamics in estuarine inlets. In: Yañez-Arancibia, A. (ed.)
131
Fish Community Ecology in Estuaries and Coastal Lagoons: towards an ecosystem
interaction. UNAM Press, México. 645p.
Yãnez-Arancibia, A.; Lara-Dominguez, A.L.; Rojas-Galaviz, J.L.; Sanchez-Gil, P.; Day
Jr, J.W. & Madden, C.J. 1988. Seasonal biomass and diversity of estuarine fishes
coupled with tropical habitat heterogeneity (southern Gulf of Mexico). In: Fish in
Estuaries. J. Fish. Biol. 33(A). 254p.
Yoneda, N.T. & Absher, T.M. 1992. Quadro biofísico e áreas ameaçadas. In: Sociedade de
Pesquisa em Vida Selvagem e Educação Ambiental. Plano Integrado de Conservação
para a região de Guaraqueçaba, Paraná, Brasil. Vol. 1, Capítulo IV, p. 13-16.
Zijlstra, J.J. 1986. Coastal offshore relationships. Importance for invertebrates, fish, birds
and marine mammals. In: Coastal offshore ecosystems relationships. UNESCO
Technical Papers in Marine Science, 48.
132
8. APÊNDICES
133
APÊNDICE I
8.1. TABELAS
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
Valores de precipitação (em mm), nas últimas 48 horas anteriores e no momento das
amostragens realizadas na Baia de Guaraqueçaba, Pr, Br. Fonte:IAPAR.
Médias diárias de temperatura do ar nas datas de amostragem na Baia de
Guaraqueçaba. Onde n=número de amostras (pontos amostrais), s=desvio e e= erro
padrão das estimativas.
Médias anuais de temperatura do ar, por pontos amostrais, da Baia de
Guaraqueçaba, onde n=número de amostragens (amostras quinzenais), s= desvio e
e= erro padrão das estimativas.
Profundidade (em metros) nos pontos amostrais da Baia de Guaraqueçaba, Pr, BR.
ANOVA resultante da comparação entre as médias de profundidades (em metros)
dos arrastos nas diferentes locais do ano e pontos amostrais ( a=0,05).
Probabilidades para o teste da diferença mínima significativa (LSD) obtidos da
comparação das médias de profundidade (em metros) dos arrastos por pontos
amostrais (a=0,05).
Médias mensais de temperatura (oC)da água superficial e de profundidade da Baia
de Guaraqueçaba, Pr, Br. Onde n=número de amostras (amostras quinzenais),
s=desvio e e= erro padrão das estimativas.
Médias anuais de temperatura da água (oC), superficial e de profundidade, por
pontos amostrais na Baia de Guaraqueçaba, Pr, Br. Onde n=número de amostras
(amostras quinzenais), s=desvio e e= erro padrão das estimativas.
ANOVA resultante da comparação das médias de temperatura (oC), na coluna
d’água (superfície e fundo) nos meses amostrais (a=0,05). Dados logaritimizados.
ANOVA resultante da comparação entre as médias de temperatura (oC) na coluna
d’água (superfície e fundo) e pontos amostrais (a=0,05). Dados logaritimizados.
Probabilidades para o teste a ”posteriori” de Scheffe da fonte de variação mês nas
médias de temperatura da água ( oC) (a=0,05).
Médias mensais dos valores de salinidade, para a água de superfície e de
profundidade, observados na Baia de Guaraqueçaba. Onde n=número de amostras
(amostras quinzenais), s=desvio e e= erro padrão das estimativas.
Médias anuais dos valores de salinidade das amostragens quinzenais(n),para a água
de superfície e de profundidade, nos diferentes pontos de amostragem da Baia de
Guaraqueçaba. Onde n=número de amostras (amostras quinzenais), s=desvio e e=
erro padrão das estimativas.
ANOVA resultante da comparação entre as médias de salinidade (‰) dos meses,
pontos e coluna d’água (superfície e fundo) amostrados ( a=0,05).
Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para
a fonte de variação mês nas médias de salinidade da água (‰) (a=0,05).
Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para
a fonte de variação coluna d’água nas médias de salinidade da água (‰) (a=0,05).
Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para
a fonte de variação ponto amostral nas médias de salinidade da água (‰) (a=0,0 5).
Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD) da
interação das fontes de variação ponto amostral/posição na coluna d’água nas
médias de salinidade da água (‰) (a=0,05).
Médias mensais dos teores de oxigênio dissolvido (em % de saturação/100),
observados, na água superficial e de profundidade da Baía de Guaraqueçaba (Pr,
Br.). Onde n=número de amostras (amostras quinzenais), s=desvio e e= erro padrão
das estimativas.
ANOVA resultante da comparação das médias de teor de oxigênio dissolvido (em %
de saturação/100) entre os meses e a coluna d’água (superfície e fundo) (a=0,05).
Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para
a fonte de variação mês nas médias de teor de oxigênio dissolvido (% de
saturação/100) (a-0,05).
136
136
137
137
137
137
138
138
138
138
139
139
139
140
140
140
140
140
141
141
141
134
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
Média anual dos teores de oxigênio dissolvido (em porcentagem de saturação/100),
observados na água superficial e de profundidade, nos diferentes pontos amostrais
da Baia de Guaraqueçaba, Pr, Br. Onde n=número de amostras (amostras
quinzenais), s=desvio e e= erro padrão das estimativas. Fonte: dados de campo.
ANOVA resultante da comparação das medias de teor de oxigênio dissolvido (cm
percentagem de saturação/100) na coluna d’água (superfície e fundo) por pontos
amostrais (a=0,05).
Grupos homogêneos resultantes da diferença mínima significativa (LSD) obtidos da
comparação das médias dos teores de 02 dissolvido (% de saturação/100) na água
por pontos amostrais.
Grupos homogêneos resultantes da diferença mínima significativa (LSD) obtida da
comparação das medias da interação entre o teor de 02 dissolvidos (% de
saturação/100) na coluna d’água (superfície e fundo) e os pontos amostrais.
Médias mensais da transparência da água (em metros) das amostragens realizadas na
Baía de Guaraqueçaba, Pr, Br. Onde n=número de amostragens (amostras
quinzenais3, s=desvio e e= erro padrão das estimativas. Fonte: dados de campo.
ANOVA resultante da comparação das medias da transparência d’água (em metros)
nos meses amostrados (a=0,05). Dados originais logaritimizados.
Grupos homogêneos resultantes da diferença mínima significativa (LSD) obtidos da
comparação das medias da transparência da água (em metros) nos meses
amostrados. Dados logaritimizados.
Médias da transparência da água (em metros), por pontos amostrais, observadas na
Baía de Guaraqueçaba, Pr, Br. Onde n=número de amostras (amostras quinzenais),
s= desvio padrão e e= erro padrão das estimativas. Fonte: dados de campo
ANOVA resultante da comparação das medias da transparência d’água (em metros)
nos pontos amostrados (a=0,05). Analise realizada nos dados originais
logaritimizados.
Freqüências absolutas (f.a.) e percentagem (%) das capturas, por família,
gênero/espécie (em ordem alfabética) e pontos amostrais, obtidas na Baia de
Guaraqueçaba. Onde ponto amostral 1 = furo da 13aia de (luaraqueçaba, ponto
amostral 2 = em frente a vila de (Guaraqueçaba, ponto amostral 3 = foz do Rio
Guaraqueçaba c ponto amostral 4= em frente a Serra do Tromomô.
Pesos totais de captura (absolutos e em porcentagem), por família, gênero/espécies
(em ordem alfabética) e pontos amostrais, na Baia de Guaraqueçaba, Pr, Br. Onde
ponto amostral 1 = furo da Baia de Guaraqueçaba, ponto amostral 2 = em frente a
vila de Guaraqueçaba, ponto amostral 3 = foz do Rio Guaraqueçaba e ponto
amostral 4 = em frente a Serra do Tromomô).
Kruskal-Wallis ANOVA por escore, resultante da comparação das freqüências
absolutas (freq.) e pesos de captura (gramas) por épocas do ano.
Kruskal-Wallis ANOVA por escore, resultante da comparação das freqüências
absolutas (freq.) e pesos das capturas (gramas) por ponto amostral.
Variâncias individuais e cumulativas (%) da ACP resultantes da comparação das 10
espécies mais abundantes (como descritores) e pontos amostrais (como objetos) da
Baia de Guaraqueçaba, Pr, Br.
Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação das
10 espécies mais abundantes (como descritores) e meses de amostragem (como
objetos) da Baia de Guaraqueçaba, Pr, Br.
Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação dos
pesos totais de captura das 10 espécies mais abundantes (como objetos) e meses de
amostragem (como descritores) na Baia de Guaraqueçaba, Pr, Br.
Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação dos
pesos totais de captura das 10 espécies mais abundantes (como objetos) e dos pesos
totais de captura por pontos amostrais (como descritores) na Baia de Guaraqueçaba,
Pr, Br.
Valores dos índices de diversidade (Nl) e equitatividade (E5) de Hill calculados para
os meses de amostragem.
Valores dos índices de diversidade (Nl) e equitatividade (E5) de Hill calculados para
os pontos amostrais.
Relação entre os índices de dive rsidade (Nl e N2) e equitatividade (E5) de Hill,
meses e pontos amostrais da Baia de Guaraqueçaba, Pr, Br.
141
142
142
142
142
142
143
143
143
144
145
146
146
146
146
147
147
147
147
147
135
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
Resultado dos 8 componentes principais (ACP), com as respectivas variâncias
individuais e cumulativas (em %), originados da plotagem simultânea dos meses de
amostragem (como objetos) e os índices de diversidade (Nl) equitatividade (E5) nos
diferentes pontos de amostragem (como descritores) da Baia de Guaraqueçaba, Pr,
Br.
Resultado dos 14 componentes principais (ACP), com as respectivas variâncias
individuais e cumulativas (em %), baseado nos parâmetros meteoro1ógicos
(pluviosidade), hidrológicos superficiais e de profundidade e nos índices de
diversidade (Nl) equitatividade (ES) nos diferentes pontos e meses de amostragem
na Baia de Guaraqueçaba, Pr, Br.
Médias do comprimento total (CT em cm), padrão (CP em cm), e peso (g) e
respectivos desvios (s) e erros padr5es (e) das estimativas para as espécies
capturadas na Baia de Guaraqueçaba (PR, BR).
Número esperado de espécies- E (Sn), em amostras de tamanho aleatório n, com
base nos cálculos de probabilidade de cada espécie ser incluída na amostra (Ludwig
& Reynolds, 1988). Foi utilizado como padrão n=2300 que correspondeu ao menor
tamanho amostral obtido.
Número de exemplares capturados em freqüências absolutas (n), absoluta
acumulada, relativas em % e acumuladas em %, por intervalos de classes de CT
(cm).
Número de exemplares capturados em freqüências absolutas (n), relativas em % e
acumuladas em %, por intervalos de classes de peso (g).
Médias de CT (cm), CP (cm) e peso (g) nos pontos amostrais associados às épocas
do ano, dos peixes capturados na Baia de Guaraqueçaba, com os respectivos
tamanhos das amostras (n) e médias, desvios (+s) e erros (e) padrões das estimativas.
Fonte: dados de campo.
Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação dos
intervalos de classe de CT e de peso das 10 espécies mais abundantes (como
descritores) e meses e pontos amostrais (como objetos) na Baia de Guaraqueçaba,
Pr, Br.
Distribuição das freqüências absolutas (freq. absol.) e relativas (%) por sexo e
estádio de maturação gonadal, das capturas realizadas entre agosto de 1986 e julho
de 1987 na Baia de Guaraqueçaba (Pr, Br).
Distribuição das freqüências absolutas (numero de exemplares) das capturas pox
ponto amostral, mês e estádios de maturação gonadal.
Variâncias individuais e cumulativas (em %) da ACP resultantes da plotagem
simultânea da freqüência por estádio de maturação gonadal (como descritores) e
meses e pontos amostrais (como objetos), das espécies com reprodução moderada
intensa e muito intensa na Baia de Guaraqueçaba, Pr, Br.
Comparação entre as atividades principais e secundárias declaradas pelos
entrevistados de Guaraqueçaba. Fonte: dados de campo.
Locais de pesca declarados por ordem de preferência pelos entrevistados em
Guaraqueçaba (n=46). Fonte: dados de campo.
Resultado dos três primeiros componentes principais (ACP) com as respectivas
variâncias individuais e cumulativas. Fonte:dados de campo
Lista comparativa da composição ictiofaunistica para as capturas com arrasto com
portas a nível familiar, gen6ico e especifico (acima de 75% de ocorrência nas
amostras).
Lista comparativa da composição ictiofaunistica para as capturas, no sistema
estuarino da Baia de Paranaguá, a nível familiar, genérico e especifico (acima de
75% de ocorrência nas amostras).
Estimativa do numero de espécies para o arrasto com portas nos ambientes do
sistema estuarino da Baia de Paranaguá.
Comparação dos resultados relacionados à reprodução, relatados na literatura
especifica e os observados para as espécies mais abundantes da Baia de
Guaraqueçaba, Pr, Br.
148
148
149
150
151
151
152
152
153
153
154
154
155
155
156
157
158
159
136
Tabela 1. Valores de precipitação (em mm), nas últimas 48 horas anteriores e no momento das
amostragens realizadas na Baía de Guaraqueçaba, Pr, Br. Fonte:IAPAR.
Precipitação (mm)
Época do ano
Mes/ano
Amostra
48 horas anteriores
Momento da coleta
Agosto/86
1
19,4
3,2
2
0,0
0,0
Primavera Setembro
1
0,0
0,0
2
3,8
1,2
Outubro
1
0,0
0,9
2
8,6
2,0
Novembro
1
72,6
2,4
2
1,4
16,0
Verão
Dezembro
1
26,2
0,2
2
0,0
27,6
Janeiro/87
1
52,4
16,7
2
9,6
0,6
Fevereiro
1
27,0
1,4
2
0,0
0,0
Outono
Março
1
2,8
0,0
2
0,0
0,0
Abril
1
0,2
0,0
2
15,6
8,4
Maio
1
0,5
0,0
2
0,2
0,0
Inverno
Junho
1
0,2
0,2
2
0,0
0,2
Julho
1
2,4
0,0
2
0,2
2,4
Tabela 2. Médias diárias de temperatura do ar nas datas de amostragem na Baía de Guaraqueçaba. Onde
n=número de amostras (pontos amostrais), s=desvio e e= erro padrão das estimativas.
Época do ano
Mes
Amostra
n
Média ( oC)
(±)s
(±)e
Agosto/86
1
4
21,22
0,58
0,29
2
4
19,75
0,96
0,48
Setembro
1
4
23,37
1,49
0,75
Primavera
2
4
25,62
1,11
0,55
Outubro
1
4
25,50
1,91
0,96
2
4
27,00
2,00
1,00
Novembro
1
4
21,50
0,71
0,35
2
4
28,75
1,66
0,83
Dezembro
1
4
28,87
2,59
1,30
Verão
2
4
29,20
1,26
0,63
Janeiro/87
1
4
30,25
1,26
0,63
2
4
30,87
1,31
0,66
Fevereiro
1
4
24,87
1,38
0,69
2
4
32,50
1,68
0,84
Março
1
4
25,00
2,34
1,17
Outono
2
4
29,25
1,55
0,78
Abril
1
4
28,62
2,93
1,46
2
4
23,25
2,18
1,09
Maio
1
4
27,87
2,33
1,16
2
4
21,62
2,50
1,25
Junho
1
4
24,75
3,09
1,55
Inverno
2
4
21,33
1,53
0,89
Julho
1
4
19,95
0,42
0,21
2
4
22,00
2,45
1,22
137
Tabela 3. Médias anuais de temperatura do ar, por pontos amostrais, da Baía de Guaraqueçaba, onde n
=número de amostragens (amostras quinzenais), s= desvio e e= erro padrão das estimativas.
Ponto amostral
n
Média ( oC)
(±)s
(±)e
1
23
24,57391
3,904917
0,81
2
24
25,50000
3,909298
0,80
3
24
26,36667
4,063054
0,83
4
24
25,85417
4,207858
0,86
Tabela 4.Profundidade (em metros) nos pontos amostrais da Baía de Guaraqueçaba, Pr, BR.
Profundidade (metros)
Época do ano
Mês/ano
Amostra
Ponto1
Ponto2
Ponto3
Agosto/86
1
7,68
2,44
1,52
2
4,57
1,83
1,98
Primavera Setembro
1
7,62
3,04
2,44
2
6,09
4,00
3,00
Outubro
1
5,18
2,74
2,44
2
5,18
2,43
3,05
Novembro
1
6,70
3,04
3,66
2
5,79
3,04
3,66
Verão
Dezembro
1
5,79
3,96
2,74
2
5,18
2,13
3,05
Janeiro/87
1
6,09
2,13
3,65
2
7,92
2,43
2,74
Fevereiro
1
7,92
2,44
3,65
2
6,09
3,04
3,65
Outono
Março
1
7,01
3,35
3,65
2
5,18
2,44
3,48
Abril
1
5,48
3,04
3,35
2
5,80
1,83
3,48
Maio
1
5,49
2,74
3,66
2
5,49
2,44
3,05
Inverno
Junho
1
6,09
2,74
3,96
2
3,04
3,04
Julho
1
6,09
2,74
3,04
2
5,18
3,04
2,74
Ponto4
1,52
4,57
7,31
6,09
4,88
3,96
7,31
5,79
5,79
6,09
6,09
4,88
6,40
7,01
7,62
6,00
4,57
6,10
7,32
6,71
7,31
6,09
6,09
6,70
Tabela 5. ANOVA resultante da comparação entre as médias de profundidades (em metros) dos arrastos
nas diferentes épocas do ano e pontos amostrais (? =0,05).
Fontes de variação
gl (fator) QM (fator) gL (erro) QM (erro)
F
p
Época do ano
3
0,10
79
0,06
1,77
0,16
Ponto amostral
3
4,06
79
0,06
72,30
0,00*
Época x Ponto amostral
9
0,04
79
0,06
0,67
0,74
* significante, CV=0,05, Kolmogorov-Smirnov d=0,13 p < 0,10 (não significante)
Tabela 6. Probabilidades para o teste da diferença mínima significativa (LSD) obtidos da comparação das
médias de profundidade (em metros) dos arrastos por pontos amostrais ( ? =0,05).
Pontos amostrais
Média 1
Média 2
Média 3
Média 4
6,07
2,75
3,11
5,92
1 {1}
0,00*
0,00*
0,97*
2 {2}
0,00*
0,64
0,00*
3 {3}
0,00*
0,64*
0,00*
4 {4}
0,97*
0,00*
0,00*
* significante
138
Tabela 7. Médias mensais de temperatura (oC)da água superficial e de profundidade da Baía de
Guaraqueçaba, Pr, Br. Onde n=número de amostras(amostras quinzenais), s=desvi o e e= erro padrão das
estimativas.
Temperatura da água ( oC)
SUPERFÍCIE
FUNDO
Época do ano Mes/ano
Média
n
(±)s
(±)e
Média
n
(±)s
(±)e
Agosto/86 21,56
8
0,94
0,33
21,75
8
0,71
0,25
Primavera
Setembro
23,10
8
1,07
0,38
22,81
8
0,88
0,31
Outubro
25,38
8
1,13
0,40
24,69
8
1,87
0,66
Novembr
23,38
8
3,42
1,21
23,75
8
3,09
1,09
Verão
Dezembro 28,75
8
1,49
0,53
28,71
8
1,31
0,46
Janeiro/87 31,31
8
0,59
0,21
30,75
8
0,46
0,16
Fevereiro
29,63
8
2,88
1,02
29,00
8
2,20
0,78
Outono
Março
28,69
8
2,03
0,72
28,25
8
1,51
0,53
Abril
26,81
8
1,51
0,53
26,38
8
0,74
0,26
Maio
22,44
8
2,29
0,81
23,19
8
2,76
0,98
Inverno
Junho
21,71
7
1,07
0,41
22,00
7
1,53
0,58
Julho
20,50
8
0,71
0,25
20,63
8
0,83
0,30
Tabela 8. Médias anuais de temperatura da água (oC), superficial e de profundidade, por pontos amostrais
na Baía de Guaraqueçaba, Pr, Br. Onde n= número de amostras (amostras quinzenais), s=desvio e e= erro
padrão das estimativas.
Temperatura da água ( oC)
Ponto
SUPERFÍCIE
FUNDO
Amostral Média
n
(±)s
(±)e
Média
n
(±)s
(±)e
1
24,96
23
3,85
0,80
24,80
23
3,65
0,76
2
25,39
24
3,84
0,78
25,13
24
3,67
0,75
3
25,52
24
3,80
0,78
25,49
24
3,63
0,74
4
25,35
24
4,35
0,89
25,33
24
3,64
0,74
Tabela 9. ANOVA resultante da comparação das médias de temperatura (oC), na coluna d’água
(superfície e fundo) nos meses amostrais ( ? =0,05). Dados logaritmizados.
Fontes de variação gl (fonte)
QM(fonte)
gl (erro)
QM(erro)
F
p (nível)
Mês
11
188,09
166
2,74
68,69
0,00*
Coluna d’água
1
2,81
166
2,74
1,02
0,31
Mês x Coluna
11
0,21
166
2,74
0,08
0,99
* significante, CV=0,06, Kolmogorov-Smirnov d=0,09 p < 0,15 (não significante).
Tabela 10. ANOVA resultante da comparação entre as médias de temperatura (oC) na coluna d’água
(superfície e fundo) e pontos amostrais (? =0,05). Dados logaritmizados.
Fontes de variação gl (fonte)
QM(fonte)
gl (erro)
QM(erro)
F
p (nível)
Ponto amostral
Coluna d’água
Ponto x Coluna
3
1
3
2,54
2,76
0,16
182
182
182
13,84
13,84
13,84
CV=0,15 , Kolmogorov-Smirnov d=0,09 p < 0,15 (não significante).
0,18
0,20
0,01
0,91
0,65
0,20
139
Tabela 11. Probabilidades para o teste a “posteriori” de Scheffe da fonte de variação mês nas médias de
temperatura da água ( oC) (? =0,05).
Época
Mês{No. ordem}
Médias
1
2
3
4
5
6
Inverno
JUL {12}
20,56
xxxx
Inverno
AGO {1}
21,66
xxxx
xxxx
Inverno
JUN {11}
21,86
xxxx
xxxx
Outono
MAI {10}
22,81
xxxx
xxxx
xxxx
Primavera
SET {2}
22,97
xxxx
xxxx
xxxx
Primavera
NOV {4}
23,56
xxxx
xxxx
Primavera
OUT {3}
25,03
xxxx
xxxx
Outono
ABR {9}
26,59
xxxx
xxxx
Outono
MAR {8}
28,47
xxxx
xxxx
Verão
DEZ {5}
28,73
xxxx
xxxx
Verão
FEV {7}
29,31
xxxx
xxxx
Verão
JAN {6}
31,03
xxxx
Tabela 12. Médias mensais dos valores de salinidade, para a água de superfície e de profundidade,
observados na Baía de Guaraqueçaba. Onde n=número de amostras (amostras quinzenais), s=desvio e e=
erro padrão das estimativas.
Salinidade (‰)
SUPERFÍCIE
FUNDO
Época do ano Mes/ano
Média
n
(±)s (±)e
Média
n
(±)s (±)e
Agosto/86
23,13
8
2,53 0,90
23,88
8
2,75 0,97
Primavera
Setembro
21,25
8
2,19 0,77
22,63
8
2,39 0,84
Outubro
17,75
8
2,49 0,88
18,38
8
2,88 1,02
Novembro
15,63
8
7,69 2,72
18,38
8
6,09 2,15
Verão
Dezembro
17,13
8
3,27 1,16
17,88
8
2,80 0,99
Janeiro/87
12,50
8
3,46 1,22
14,88
8
3,09 1,09
Fevereiro
17,25
8
1,75 0,62
18,13
8
2,36 0,83
Outono
Março
21,75
8
3,01 1,06
23,38
8
1,60 0,56
Abril
14,75
8
4,23 1,50
18,38
8
4,03 1,43
Maio
15,38
8
3,34 1,18
16,38
8
3,02 1,07
Inverno
Junho
16,75
8
3,37 1,19
17,00
7
2,71 1,02
Julho
18,38
8
4,21 1,49
19,13
8
3,48 1,23
Tabela 13. Médias anuais dos valores de salinidade das amostragens quinzenais(n),para a água de
superfície e de profundidade, nos diferentes pontos de amostragem da Baía de Guaraqueçaba. Onde
n=número de amostras (amostras quinzenais), s=desvio e e= erro padrão das estimativas.
Salinidade (‰)
SUPERFÍCIE
FUNDO
Ponto
amostral
1
2
3
4
Média
n
(±)s
(±)e
Média
n
(±)s
(±)e
20,17
19,33
14,58
16,46
24
24
24
24
3,37
3,60
4,82
4,47
0,69
0,74
0,98
0,91
22,09
20,08
16,29
17,88
23
24
24
24
3,15
3,16
4,16
3,54
0,66
0,64
0,85
0,72
140
Tabela 14. ANOVA resultante da comparação entre as médias de salinidade (0/oo) dos meses, pontos e
coluna d’água (superfície e fundo) amostrados (? =0,05).
Fonte de variação
gl (fonte)
QM (fonte)
gl (erro)
QM (erro)
F
p
Mês
11
128,11
95
8,06
15,90 0,00*
Ponto amostral
3
173,35
95
8,06
21,51 0,00*
Coluna d’água
1
275,06
95
8,06
34,13 0,00*
Mês x pto
33
3,86
95
8,06
0,48
0,10
Mês x Coluna
11
4,44
95
8,06
0,55
0,86
Pto x Coluna
3
53,42
95
8,06
6,63
0,00*
Mês x Pto x Coluna
33
3,30
95
8,06
0,41
0,10
*significante, CV=0,15 , Kolmogorov-Smirnov d=0,05 p < 0,10 (não significante).
Tabela 15. Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para a fonte de
variação mês nas médias de salinidade da água ( 0/oo) (alfa=0,05).
Época do ano
Mês{No. ordem}
Média
1
2
3
4
Verão
JAN {6}
13,94
xxxx
Outono
MAI {10}
16,12
xxxx
Inverno
JUN {11}
17,37
xxxx
xxxx
Outono
ABR {9}
17,69
xxxx
xxxx
Verão
FEV {7}
17,81
xxxx
xxxx
Primavera
NOV {4}
18,12
xxxx
Verão
DEZ {5}
18,25
xxxx
Primavera
OUT {3}
18,50
xxxx
Inverno
JUL {12}
18,94
xxxx
Primavera
SET {2}
22,25
xxxx
Outono
MAR {8}
22,87
xxxx
Inverno
AGO {1}
23,69
xxxx
Tabela 16. Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para a fonte de
variação coluna d’água nas médias de salinidade da água ( 0/oo) (? =0,05).
Coluna d’água
Média
1
2
Superfície
17,59
xxxx
Fundo
20,00
xxxx
Tabela 17. Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para a fonte de
variação ponto amostral nas médias de salinidade da água ( 0/oo) (? =0,05).
Ponto amostral
Média
1
2
3
4
17,08
xxxx
3
17,33
xxxx
2
19,71
xxxx
1
21,06
xxxx
Tabela 18. Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD) da interação
das fontes de variação ponto amostral/posição na coluna d’água nas médias de salinidade da água (0/oo)
(? =0,05).
Ponto x Coluna {No. Ordem}
Média
1
2
3
4
5
3 superfície{5}
14,58
xxxx
4 superfície{7}
16,29
xxxx
4 fundo{8}
17,87
xxxx
xxxx
2 superfície{3}
19,33
xxxx
xxxx
2 fundo{4}
20,08
xxxx
3 fundo{6}
20,08
xxxx
1 superfície{1}
20,17
xxxx
1 fundo{2}
22,09
xxxx
141
Tabela 19. Médias mensais dos teores de oxigênio dissolvido (em porcentagem de saturação/100),
observados, na água superficial e de profundidade da Baía de Guaraqueçaba (Pr., Br.). Onde n=número
de amostras (amostras quinzenais), s=desvio e e= erro padrão das estimativas.
Oxigênio Dissolvido (% de saturação/100)
Superfície
Fundo
Época do ano Mês/ano
Média
n
(±)s
(±)e
Média
n
(±)s
(±)e
Agosto/86
0,45
8
0,18
0,06
,52
8
0,08
0,03
Primavera
Setembro
0,97
8
0,36
0,13
1,10
8
0,13
0,05
Outubro
1,17
8
0,51
0,18
1,27
8
0,32
0,11
Novembro
0,79
3
0,46
0,27
1,03
4
0,18
0,09
Verão
Dezembro
0,92
4
0,42
0,21
0,96
4
0,08
0,04
Janeiro/87
0,70
7
0,37
0,14
0,87
8
0,29
0,10
Fevereiro
0,74
8
0,31
0,11
0,90
8
0,08
0,03
Outono
Março
0,76
8
0,34
0,12
0,82
8
0,09
0,03
Abril
0,89
7
0,38
0,14
0,58
8
0,09
0,03
Maio
0,70
8
0,35
0,12
0,72
8
0,09
0,03
Inverno
Junho
0,60
7
0,25
0,09
0,70
7
0,06
0,02
Julho
0,63
8
0,24
0,08
0,73
8
0,04
0,01
Tabela 20. ANOVA resultante da comparação das médias de teor de oxigênio dissolvido (em % de
saturação/100) entre os meses e a coluna d’água (superfície e fundo) (? =0,05).
Fonte de variação
gl (fonte)
QM(fonte)
gl (erro)
QM(erro)
F
p (nível)
Mês
11
0,57
147
0,07
7,94
0,00*
Coluna d’água
1
0,20
147
0,07
2,80
0,10
Mês x Coluna
11
0,07
147
0,07
0,91
0,53
*significante, CV=0,04, Kolmogorov-Smirnov d=0,06 p < 0,10 (não significante)
Tabela 21. Grupos homogêneos resultantes do teste Mínima Diferença Significante (LSD), para a fonte de
variação mês nas médias de teor de oxigênio dissolvido (% de saturação/100) (? =0,05).
Épocas do ano
Meses{No. ordem}
Média
1
2
3
4
5
6
Inverno
AGO {1}
0,49
xxxx
Inverno
JUN {11}
0,65
xxxx xxxx
Inverno
JUL {12}
0,67
xxxx xxxx xxxx
Outono
MAI {10}
0,71
xxxx xxxx
Outono
ABR {9}
0,73
xxxx xxxx xxxx
Verão
JAN {6}
0,78
xxxx xxxx xxxx
Outono
MAR {8}
0,79
xxxx xxxx xxxx
Verão
FEV {7}
0,82
xxxx xxxx xxxx
Primavera
NOV {4}
0,91
xxxx xxxx xxxx
Verão
DEZ {5}
0,94
xxxx xxxx
Primavera
SET {2}
1,04
xxxx xxxx
Primavera
OUT {3}
1,22
xxxx
Tabela 22. Média anual dos teores de oxigênio dissolvido (em porcentagem de saturação/100),
observados na água superficial e de profundidade, nos diferentes pontos amostrais da Baía de
Guaraqueçaba, Pr, Br. Onde n=número de amostras (amostras quinzenais), s=desvio e e= erro padrão das
estimativas. Fonte: dados de campo.
Oxigênio Dissolvido (% de saturação/100)
Superfície
Fundo
Ponto
Média
n
s
e
Média
n
s
e
Amostral
(±)
(±)
(±)
(±)
1
0,88
21
0,29
0,06
0,80
21
0,25
0,05
2
0,28
21
0,09
0,02
0,85
22
0,29
0,06
3
0,95
20
0,28
0,06
0,86
22
0,26
0,05
4
0,98
22
0,27
0,06
0,84
22
0,26
0,05
142
Tabela 23. ANOVA resultante da comparação das médias de teor de oxigênio dissolvido (em
percentagem de saturação/100) na coluna d’água (superfície e fundo) por pontos amostrais ( ? =0,05).
Fontes de variação
gl (fonte)
QM(fonte)
gl (erro)
QM(erro)
F
p (nível)
Pontos amostrais
3
1,15
163
0,06
17,52
0,00*
Coluna d’água
1
0,18
163
0,06
2,82
0,09
Pontos x Coluna
3
1,22
163
0,06
18,63
0,00*
*significante, CV=0,30, Kolmogorov-Smirnov d=0,06 p < 0,10 (não significante).
Tabela 24. Grupos homeogêneos resultantes da diferença mínima significativa (LSD) obtidos da
comparação das médias dos teores de O2 dissolvido (% de saturação/100) na água por pontos amostrais.
Ponto amostral{No. ordem}
Média
1
2
2 {2}
0,57
xxxx
1 {1}
0,84
xxxx
3 {3}
0,91
xxxx
4 {4}
0,91
xxxx
Tabela 25. Grupos homeogêneos resultantes da diferença mínima significativa (LSD) obtidos da
comparação das médias da interação entre o teor de O2 dissolvido (% de saturação/100) na coluna d’água
(superfície e fundo) e os pontos amostrais.
Ponto amostral{No. ordem}
Média
1
2
3
2 sup {3}
0,28
xxxx
1 fun {2}
0,80
xxxx
4 fun {8}
0,84
xxxx
xxxx
2 fun {4}
0,85
xxxx
xxxx
3 fun {6}
0,87
xxxx
xxxx
1 sup {1}
0,88
xxxx
xxxx
3 sup {5}
0,95
xxxx
xxxx
4 sup {7}
0,98
xxxx
Tabela 26. Médias mensais da transparência da água (em metros) das amostragens realizadas na Baía de
Guaraqueçaba, Pr, Br. Onde n=número de amostragens (amostras quinzenais), s=desvio e e= erro padrão
das estimativas. Fonte: dados de campo.
Transparência da água (metros)
Época do ano
Mês/Ano
Média
n
(±)s
(±)e
Agosto/86
1,32
6
0,29
0,12
Primavera
Setembro
0,84
7
0,24
0,09
Outubro
0,68
7
0,31
0,11
Novembro
0,87
8
0,15
0,05
Verão
Dezembro
0,75
8
0,13
0,04
Janeiro/87
0,67
8
0,14
0,05
Fevereiro
0,92
8
0,14
0,05
Outono
Março
0,91
7
0,08
0,03
Abril
0,86
8
0,15
0,05
Maio
0,78
8
0,17
0,06
Inverno
Junho
1,09
7
0,44
0,17
Julho
1,23
8
0,22
0,07
Tabela 27. ANOVA resultante da comparação das médias da transparência d’água (em metros) nos meses
amostrados (? =0,05). Dados originais logaritmizados.
Fonte de variação
gl (fonte)
QM(fonte)
gl (erro)
QM(erro)
F
p (nível)
Mês
11
0,30
78
0,05
6,07
0,00*
*significante, CV=0,25 , Kolmogorov-Smirnov d=0,12 p < 0,15 (não significante)
143
Tabela 28. Grupos homogêneos resultantes da diferença mínima significativa (LSD) obtidos da
comparação das médias da transparência da água (em metros) nos meses amostrados. Dados
logaritimizados.
Época do ano
Meses/ No. ordem
Média
1
2
3
4
5
Verão
JAN {6}
0,67
xxxx
Primavera
OUT {3}
0,68
xxxx
xxxx
Verão
DEZ {5}
0,75
xxxx
xxxx
xxxx
Outono
MAI {10}
0,78
xxxx
xxxx
xxxx
Primavera
SET {2}
0,84
xxxx
xxxx
xxxx
Outono
ABR {9}
0,86
xxxx
xxxx
xxxx
xxxx
Primavera
NOV {4}
0,86
xxxx
xxxx
xxxx
xxxx
Outono
MAR {8}
0,91
xxxx
xxxx
xxxx
Verão
FEV {7}
0,92
xxxx
xxxx
Inverno
JUN {11}
1,08
xxxx
xxxx
Inverno
JUL {12}
1,23
xxxx
Inverno
AGO {1}
1,32
xxxx
Tabela 29. Médias da transparência da água (em metros), por pontos amostrais, observadas na Baía de
Guaraqueçaba, Pr , Br. Onde n=número de amostras (amostras quinzenais), s= desvio padrão e e= erro
padrão das estimativas. Fonte: dados de campo.
Transparência (m)
Ponto Amostral
Média
n
(±)e
(?)s
1
1,01
23
0,24
0,05
2
0,85
22
0,35
0,07
3
0,84
22
0,26
0,05
4
0,90
23
0,25
0,05
Tabela 30. ANOVA resultante da comparação das médias da transparência d’água (em metros) nos
pontos amostrados (? =0,05). Análise realizada nos dados originais logaritmizados.
Fonte de variação
gl (fonte)
QM(fonte)
gl (erro)
QM(erro)
F
p (nível)
Ponto Amostral
3
0,14
86
0,08
1,80
0,15
CV=0,31 , Kolmogorov-Smirnov d=0,12 p < 0,15 (não significante).
144
Tabela 31. Freqüências absolutas (f.a.) e percentagem (%) das capturas, por família, gênero/espécie (em
ordem alfabética) e pontos amostrais, obtidas na Baía de Guaraqueçaba. Onde ponto amostral 1 = furo da
Baía de Guaraqueçaba, ponto amostral 2 = em frente a vila de Guaraqueçaba, ponto amostral 3 = foz do
Rio Guaraqueçaba e ponto amostral 4= em frente a Serra do Tromomô.
FAMÍLIA
GÊNERO/ESPÉCIE
ACHIRIDAE
Achirus lineatus
Trinectes microphtalmus
Trinectes paulistanus
ARIIDAE
Bagre bagre
Cathorops spixii
Genidens genidens
Netuma barba
Sciadeichthys luniscutis
CARANGIDAE
Caranx latus
Chloroscombrus crysurus
Oligoplites saliens
Oligoplites saurus
Selene vomer
CLUPEIDAE
Harengula clupeola
Pellona harroweri
CYNOGLOSSIDAE
Symphurus plagusia
DIODONTIDAE
Chilomycterus spinosus
ENGRAULIDAE*
Anchoa parva
Cetengraulis edentulus
Lycengraulis grossidens
EPHIPPIDAE
Chaetodipterus faber
GERREIDAE
Diapterus rhombeus
Eucinostomus argenteus
Eucinostomus gula
Eucinostomus melanopterus
GOBIIDAE
Gobionellus oceanicus
HAEMULIDAE**
Genyatremus luteus
Pomadasys corvinaeformis
PARALICHTHYIDAE
Citharichthys arenaceus
Citharichthys spilopterus
Etropus crossotus
Paralichthys tropicus
SCIAENIDAE
Bairdiella ronchus
Ctenosciaena gracillicirrhus
Cynoscion acoupa
Cynoscion leiarchus
Cynoscion microlepidotus
Cynoscion virescens
Isopisthus parvipinnis
Macrodon ancylodon
Menticirrhus americanus
Micropogonias furnieri
Paralonchurus brasiliensis
Stellifer brasiliensis
Stellifer rastrifer
SERRANIDAE
Diplectrum radiale
STROMATEIDAE
Peprilus paru
SYNODONTIDAE
Synodus foetens
TETRAODONTIDAE
Lagocephalus laevigatus
Sphoeroides testudineus
TRICHIURIDAE
Trichiurus lepturus
TRIGLIDAE
Prionotus punctatus
TOTAL
Família
Gên. Esp.
f.a.
%
f.a.
%
126
0,37
57
0,17
53
0,16
16
0,05
24940
73,64
9
0,03
18839
55,62
3980
11,75
672
1,98
1440
4,25
50
0,15
1
0
2
0,01
20
0,06
3
0,01
24
0,07
44
0,13
35
0,10
9
0,03
250
0,74
250
0,74
17
0,05
17
0,05
60
0,18
13
0,04
36
0,11
11
0,03
133
0,39
133
0,39
6
0,02
1
0
1
0
2
0,01
2
0,01
4
0,01
4
0,01
84
0,25
81
0,24
3
0,01
842
2,49
305
0,90
171
0,50
363
1,07
3
0,01
6692
19,76
23
0,07
20
0,06
14
0,04
2
0,01
80
0,24
23
0,07
473
1,40
68
0,20
14
0,04
579
1,71
78
0,23
245
0,72
5073
14,98
3
0,01
3
0,01
1
0,003
1
0
1
0,003
1
0
560
1,65
16
0,05
544
1,61
45
0,13
45
0,13
10
0,03
10
0,03
Total
33868
*Engraulididae por alguns autores
**Pomadasyidae incluída
100
33868
100
1
PONTOS AMOSTRAIS
2
3
f.a.
%
f.a.
%
f.a.
%
20
25
0
0,06
0,07
0
10
14
4
0,03
0,04
0,01
20
10
10
6
8534
359
134
465
0,02
25,2
1,06
0,40
1,37
0
4223
667
254
678
0
12,47
1,97
0,75
2,00
0
1
3
0
15
0
0
0,01
0
0,04
0
0
12
0
4
8
8
0,02
0,02
97
4
f.a.
%
0,06
0,03
0,03
7
4
2
0,02
0,01
0,01
0
629
2528
76
157
0
1,86
7,46
0,22
0,46
3
5453
426
208
140
0,01
16,10
1,26
0,61
0,41
0
0
0,04
0
0,01
0
1
4
3
4
0
0
0,01
0,01
0,01
1
0
1
0
1
0
0
0
0
0
17
0
0,05
0
8
0
0,02
0
2
1
0,01
0
0,29
103
0,30
20
0,06
30
0,09
4
0,01
11
0,03
2
0,01
0
0
0
2
3
0
0,01
0,01
1
16
2
0
0,05
0,01
12
5
0
0,04
0,01
0
0
13
6
0
0,04
0,02
36
0,11
11
0,03
65
0,19
21
0,06
0
0
0
1
0
0
0
0
0
1
0
0
0
0
0
0
1
0
2
1
0
0
0,01
0
0
0
0
0
0
0
0
0
3
0,01
1
0
0
0
0
0
25
0
0,07
0
16
1
0,05
0
16
0
0,05
0
24
2
0,07
0,01
106
25
267
0
0,31
0,07
0,79
0
167
58
18
0
0,49
0,17
0,05
0
28
52
64
3
0,08
0,15
0,19
0,01
4
36
14
0
0,01
0,11
0,04
0
1
2
1
1
40
22
299
48
7
8
75
117
1816
0
0,01
0
0
0,12
0,06
0,88
0,14
0,02
0,02
0,22
0,35
5,36
5
4
11
1
31
1
31
5
6
239
3
89
1392
0,01
0,01
0,03
0
0,09
0
0,09
0,01
0,02
0,71
0,01
0,26
4,11
1
12
1
0
4
0
66
2
1
296
0
14
659
0
0,04
0
0
0,01
0
0,19
0,01
0
0,87
0
0,04
1,95
16
2
1
0
5
0
77
13
0
36
0
25
1206
0,05
0,01
0
0
0,01
0
0,23
0,04
0
0,11
0
0,07
3,56
0
0
1
0
1
0
1
0
0
0
1
0
0
0
0
0
1
0
0
0
0
0
0
0
6
48
0,02
0,14
1
293
0
0,87
5
49
0,01
0,14
4
154
0,01
0,45
24
0,07
7
0,02
1
0
13
0,04
7
0,02
1
0
1
0
1
0
12670
100
8411
100
4834
100
7953
100
145
Tabela 32. Pesos totais de captura (absolutos e em percentagem), por família, gênero/espécies (em ordem alfabética)
e pontos amostrais, na Baía de Guaraqueçaba, Pr, BR. Onde ponto amostral 1 = furo da Baía de Guaraqueçaba, ponto
amostral 2 = em frente a vila de Guaraqueçaba, ponto amostral 3 = foz do Rio Guaraqueçaba e ponto amostral 4 = em
frente a Serra do Tromomô.
Total
FAMÍLIA
GÊNERO/ESPÉCIE
ACHIRIDAE
Achirus lineatus
Trinectes microphtalmus
T. paulistanus
ARIIDAE
Bagre bagre
Cathorops spixii
Genidens genidens
Netuma barba
Sciadeichthys luniscutis
CARANGIDAE
Caranx latus
C. crysurus
Oligoplites saliens
O. saurus
Selene vomer
CLUPEIDAE
Harengula clupeola
Pellona harroweri
CYNOGLOSSIDAE
Symphurus plagusia
DIODONTIDAE
Chilomycterus spinosus
ENGRAULIDAE*
Anchoa parva
Cetengraulis edentulus
Lycengraulis grossidens
EPHIPPIDAE
Chaetodipterus faber
GERREIDAE
Diapterus rhombeus
Eucinostomus argenteus
E. gula
E. melanopterus
GOBIIDAE
Gobionellus oceanicus
HAEMULIDAE**
Genyatremus luteus
P. corvinaeformis
PARALICHTHYIDAE
Citharichthys arenaceus
C. spilopterus
Etropus crossotus
Paralichthys tropicus
SCIAENIDAE
Bairdiella ronchus
C. gracillicirrhus
Cynoscion acoupa
C. leiarchus
C. microlepidotus
C. virescens
Isopisthus parvipinnis
Macrodon ancylodon
M. americanus
Micropogonias furnieri
P. brasiliensis
Stellifer brasiliensis
S. rastrifer
SERRANIDAE
Diplectrum radiale
STROMATEIDAE
Peprilus paru
SYNODONTIDAE
Synodus foetens
TETRAODONTIDAE
Lagocephalus laevigatus
Sphoeroides testudineus
TRICHIURIDAE
Trichiurus lepturus
TRIGLIDAE
Prionotus punctatus
Família
Peso (g)
%
0,44
3428,10
590905,20
621,10
5650,10
0,73
1625,50
0,21
1853,00
0,24
9263,70
1,20
104,20
0,01
120,50
0,01
1403,50
0,18
1669,50
1249,60
509,00
0,22
0,16
0,07
761,00
639,60
0
0,10
0,08
0,00
385,10
449273,80
77558,80
23005,50
40603,50
0,05
58,14
10,04
2,98
5,25
180,00
263109,70
9478,90
7488,00
9301,40
0,02
34,05
1,23
0,97
1,20
0,80
5,80
408,00
54,00
85,50
0,00
0,00
0,06
0,01
0,01
0
1,80
78,00
0
47,5
1175,00
37,50
0,15
0,00
5650,10
312,50
365,00
203,50
%
4
Peso (g)
%
0,04
0,05
0,03
343,50
78,00
254,50
0,04
0,01
0,03
252,50
167,00
51,00
0,03
0,02
0,01
0
60058,80
8091,00
2758,00
7200,60
0,00
7,77
1,05
0,36
0,93
0
21215,60
55890,70
922,00
3293,00
0,00
2,75
7,23
0,12
0,43
205,00
104889,10
4098,20
11837,50
20808,50
0,03
13,57
0,53
1,53
2,69
0,00
0,00
0,01
0,00
0,01
0
0
247,00
0
22,50
0,00
0,00
0,03
0,00
0,00
0
4,00
137,50
54,00
14,50
0,00
0,00
0,02
0,01
0,00
0,80
0
17,50
0
1,00
0,00
0,00
0,00
0,00
0,00
245,00
36,5
0,03
0,00
580,00
0
0,08
0,00
282,50
0
0,04
0,00
67,50
1,00
0,01
0,00
2652,80
46,00
0,04
0,00
0,01
689,70
1029,50
0,26
0,00
0,13
311,10
550,00
0,34
0,00
0,07
1996,50
1625,50
0,73
0,00
0,21
0
0,09
0,00
0,00
300,00
1210,50
342,50
0,04
0,16
0,04
0
70,00
112,50
0,00
0,01
0,01
10,00
504,50
75,00
0,00
0,07
0,01
290,00
144,00
0
0,04
0,02
0,00
0
492,00
155,00
0,00
0,06
0,02
9263,70
1,20
827,50
0,11
1702,60
0,22
5897,50
0,76
836,10
0,11
30,00
22,50
24,70
27,00
0,00
0,00
0,00
0,00
0
0
0
15,00
0,00
0,00
0,00
0,00
0
22,50
0
0
0,00
0,00
0,00
0,00
30,00
0
24,70
12,00
0,00
0,00
0,00
0,00
0
0
0
0
0,00
0,00
0,00
0,00
120,50
0,02
113,00
0,01
7,50
0,00
0
0,00
0
0,00
1362,50
41,00
0,18
0,01
579,00
0
0,07
0,00
143,00
4,00
270,00
0
0,51
0,34
0,57
0,10
1246,60
445,60
3295,30
0
0,16
0,06
0,43
0,00
2425,30
1007,00
283,50
0
269,60
767,0
645,50
759,00
0,03
0,00
0,00
0,03
0,10
0,08
0,10
370,50
37,00
3974,00
2599,10
4425,80
759,00
0,02
0,00
0,00
0,31
0,13
0,04
0,00
32,50
379,50
201,50
0
0,05
0,00
0,00
0,00
0,05
0,03
0,00
673,00
179,80
504,60
50,30
2531,70
1885,00
7578,00
5258,00
2321,80
9654,70
2669,00
2985,50
58925,10
0,08
0,02
0,07
0,01
0,33
0,24
0,59
0,68
0,30
1,25
0,35
0,39
7,63
5,00
7,00
15,10
27,80
1501,50
1835,00
3786,70
4015,00
1089,00
254,00
2496,00
1847,50
19540,40
0,00
0,00
0,00
0,00
0,19
0,24
0,49
0,52
0,14
0,03
0,32
0,24
2,53
62,50
36,80
274,50
22,50
752,50
50,00
743,80
131,00
762,80
4362,50
173,00
717,50
17271,00
0,01
0,00
0,04
0,00
0,10
0,01
0,10
0,02
0,10
0,56
0,02
0,09
2,23
20,00
124,00
150,00
0
136,00
0
1353,50
49,00
470,00
4294,20
0
147,00
7473,10
0,00
0,02
0,02
0,00
0,02
0,00
0,18
0,01
0,06
0,56
0,00
0,02
0,97
549,50
12,00
65,00
0
141,70
0
1694,00
1063,00
0
744,00
0
273,50
14640,60
0,07
0,00
0,01
0,00
0,02
0,00
0,22
0,14
0,00
0,10
0,00
0,04
1,89
129,70
0,02
0
0,00
23,70
0,00
26,00
0,00
80,00
0,01
55,00
0,01
0
0,00
55,00
0,01
0
0,00
0
0,00
47,00
0,01
47,00
0,01
0
0,00
0
0,00
0
0,00
202,20
45781,00
0,03
5,92
62,50
3864,20
0,01
0,50
13,70
24198,90
0,00
3,13
59,00
4209,00
0,01
0,54
67,00
13508,90
0,01
1,75
3467,50
0,45
0,00
0,01
0,00
1620,00
0,21
0,00
0,00
0,00
650,00
0,08
0,00
0,00
0,00
45,00
0,01
0,00
0,00
0,00
1152,50
0,15
0,00
0,00
0,00
100
343318,00
44,43
139356,50
18,03
110516,00
14,30
179587,10
1,52
12,32
129,70
0,02
55,00
0,01
47,00
0,01
45983,20
5,95
3467,50
0,45
47,30
0,01
Total
772777,50
*Engraulididae por alguns autores
**Pomadasyidae incluída
%
PONTOS AMOSTRAIS
2
3
Peso (g)
%
Peso (g)
0,08
0,16
95180,50
1
Peso (g)
76,46
1212,50
11757,90
Gênero/Espécie
Peso (g)
%
47,30
100
772777,50
29,70
5,00
4,00
5,00
23,24
146
Tabela 33. Kruskal-Wallis ANOVA por escore, resultante da comparação das freqüências absolutas
(freq.) e pesos de captura (gramas) por épocas do ano.
Épocas do ano
n válido
Soma dos escores (freq.)*
Soma dos escores (peso)**
Primavera
28
1461,50
1673,00
Verão
20
1201,50
1108,00
Outono
24
1506,00
1377,00
Inverno
24
487,00
498,00
* Significante (? =0,05 , H (3,n=96)=34,84 p =0,00).
**Significante (? =0,05 , H (3,n=96)=32,08 p =0,00).
Tabela 34. Kruskal-Wallis ANOVA por escore, resultante da comparação das freqüências absolutas
(freq.) e pesos das capturas (gramas) por ponto amostral.
Pontos amostrais
n válido
Soma dos escores (freq.)*
Soma dos escores (peso)**
1
24
1457,50
1512,00
2
24
1094,00
1047,00
3
24
904,00
891,00
4
24
1200,50
1206,00
* Significante (? =0,05 , H(3,n=96)=8,59 p =0,03).
** Significante (? =0,05, H ( 3, n= 96) = 11,33 p =0,01).
Tabela 35. Variâncias individuais e cumulativas (%) da ACP resultantes da comparação das 10 espécies
mais abundantes (como descritores) e pontos amostrais (como objetos) da Baía de Guaraqueçaba, Pr, Br.
Número do Componente
Porcentagem da variância
Porcentagem cumulativa
1
51,68745
51,68745
2
33,05197
84,73942
3
15,26058
100,00000
4
0,00000
100,00000
5
0,00000
100,00000
6
0,00000
100,00000
7
0,00000
100,00000
8
0,00000
100,00000
9
0,00000
100,00000
10
0,00000
100,00000
11
0,00000
100,00000
Tabela 36. Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação das 10
espécies mais abundantes (como descritores) e meses de amostragem (como objetos) da Baía de
Guaraqueçaba, Pr, Br.
Número do Componente
Porcentagem da variância
Porcentagem cumulativa
1
75,62797
75,62797
2
10,89944
86,52741
3
7,75632
94,28373
4
4,44723
98,73096
5
0,94423
99,67519
6
0,24848
99,92368
7
0,04961
99,97329
8
0,01954
99,99283
9
0,00716
99,99999
10
0,00001
100,0000
11
0,00000
100,0000
12
0,00000
100,0000
147
Tabela 37. Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação dos pesos
totais de captura das 10 espécies mais abundantes (como objetos) e meses de amostragem (como
descritores) na Baía de Guaraqueçaba, Pr, Br.
Número do Componente
Porcentagem da variância
Porcentagem cumulativa
1
78,71794
78,71794
2
14,35192
93,06987
3
4,08789
97,15775
4
1,88996
99,04772
5
0,60090
99,64862
6
0,21942
99,86803
7
0,10315
99,97118
8
0,02204
99,99322
9
0,00671
99,99993
10
0,00004
99,99997
11
0,00002
99,99999
12
0,00001
100,0000
Tabela 38. Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação dos pesos
totais de captura das 10 espécies mais abundantes (como objetos) e dos pesos totais de captura por pontos
amostrais (como descritores) na Baía de Guaraqueçaba, Pr, Br.
Número do Componente
Porcentagem da variância
Porcentagem cumulativa
1
74,56146
74,56146
2
22,82273
97,38419
3
2,22085
99,60504
4
0,39496
100,00000
Tabela 39. Valores dos índices de diversidade (N1) e equitatividade (E5) de Hill calculados para os meses
de amostragem.
Índices
Mês/Ano
N1
E5
Agosto
9,54
0,66
Setembro
5,49
0,58
Outubro
4,57
1,50
Novembro
3,96
0,45
Dezembro
5,41
0,58
Janeiro
3,79
0,62
Fevereiro
3,30
0,44
Março
5,03
0,42
Abril
4,00
0,46
Maio
2,97
0,38
Junho
3,08
0,46
Julho
8,25
0,53
Tabela 40. Valores dos índices de diversidade (N1) e equitatividade (E5) de Hill calculados para os
pontos amostrais.
Índices
Pontos Amostrais
N1
E5
1
3,80
0,40
2
5,91
0,48
3
5,70
0,46
4
3,40
0,43
148
Tabela 41. Relação entre os índices de diversidade (N1 e N2) e equitatividade (E5) de Hill, meses e
pontos amostrais da Baía de Guaraqueçaba.
Índices/Pontos/Amostrais
N1
Meses/Ano
Agosto/86
Setembro
Outubro
Novembro
Dezembro
Janeiro/87
Fevereiro
Março
Abril
Maio
Junho
Julho
1
5,13
4,29
2,17
2,83
3,03
3,11
1,95
2,77
4,23
1,92
2,04
6,36
2
4,11
9,34
4,99
7,36
4,90
2,95
4,79
5,58
3,50
5,99
5,80
5,38
3
2,81
3,17
4,65
2,80
3,47
5,89
5,79
4,45
6,09
5,99
1,78
3,93
E5
4
6,57
4,50
2,94
2,90
4,32
2,72
2,81
2,25
2,77
1,73
1,12
2,67
1
0,59
0,53
0,42
0,44
0,50
0,66
0,50
0,37
0,57
0,44
0,48
0,53
2
1,15
0,87
0,73
0,93
0,54
0,61
0,57
0,52
0,49
0,51
0,72
0,44
3
0,37
0,54
0,76
0,38
0,51
0,74
0,52
0,64
0,53
0,56
0,39
0,76
4
0,56
0,56
0,48
0,48
0,67
0,53
0,45
0,45
0,49
0,47
0,28
0,76
Tabela 42. Resultado dos 8 componentes principais (ACP), com as respectivas variâncias individuais e
cumulativas (em %), originados da plotagem simultânea dos meses de amostragem (como objetos) e os
índices de diversidade (N1) equitatividade (E5) nos diferentes pontos de amostragem (como descritores)
da Baía de Guaraqueçaba, Pr, Br.
Número do Componente
Porcentagem da variância
Porcentagem cumulativa
1
34,24535
34,24535
2
31,34724
65,59258
3
15,42919
81,02177
4
6,49452
87,51630
5
4,82921
92,34550
6
4,48473
96,83023
7
2,40597
99,23621
8
0,76379
100,0000
Tabela 43. Resultado dos 14 componentes principais (ACP), com as respectivas variâncias individuais e
cumulativas (em %), baseado nos parâmetros meteorológicos (pluviosidade), hidrológicos superficiais e
de profundidade e nos índices de diversidade (N1) equitatividade (E5) nos diferentes pontos e meses de
amostragem na Baía de Guaraqueçaba, Pr, Br.
Número do Componente
Porcentagem da variância
Porcentagem cumulativa
1
29,70576
29,70576
2
18,53822
48,24398
3
14,64172
62,88570
4
9,19591
72,08161
5
7,73966
79,82127
6
5,71112
85,53239
7
5,01158
90,54397
8
3,16274
93,70671
9
2,72518
96,43189
10
1,69810
98,13000
11
1,11925
99,24925
12
0,43556
99,68478
13
0,24462
99,92940
14
0,07060
100,00000
149
Tabela 44. Número esperado de espécies- E (Sn), em amostras de tamanho aleatório n,
com base nos cálculos de probabilidade de cada espécie ser incluída na amostra
(Ludwig & Reynolds, 1988). Foi utilizado como padrão n=2300 que correspondeu ao
menor tamanho amostral obtido.
Pontos amostrais
Pontos amostrais
n
1
2
3
4
n
1
2
3
4
100
11
12
13
9
2200
31
32
34
27
200
15
16
17
13
2300
31
32
34
28
300
18
18
19
15
2400
31
32
400
19
19
21
16
2500
31
33
500
21
21
23
18
2600
32
33
600
22
22
24
19
2700
32
33
700
23
23
25
20
2800
32
34
800
24
24
26
21
2900
32
34
900
25
25
27
21
3000
33
34
1000 26
26
28
22
3100
33
34
1100 26
27
28
23
3200
33
34
1200 27
27
29
23
3300
33
35
28
30
24
3400
33
35
1300 27
1400 28
29
30
24
3500
33
35
1500 28
29
31
25
3600
34
35
1600 29
30
31
25
3700
34
35
1700 29
30
32
25
3800
34
36
30
32
26
3900
34
36
1800 29
1900 30
31
32
26
4000
34
36
2000 30
31
33
27
4100
34
36
2100 30
32
33
27
150
Tabela. 45. Médias dos comprimentos total (CT em cm ), padrão (CP em cm), e peso (g) e respectivos
desvios (s) e erros padrões (e) das estimativas para as espécies capturadas na Baía de Guaraqueçaba (PR,
BR).
FAMÍLIA
GÊNERO/ESPÉCIE
ACHIRIDAE
Achirus lineatus
Trinectes microphtalmus
T. paulistanus
ARIIDAE
Bagre bagre
Cathorops spixii
Genidens genidens
Netuma barba
Sciadeichthys luniscutis
CARANGIDAE
Caranx latus
C. crysurus
Oligoplites saliens
O. saurus
Selene vomer
CLUPEIDAE
Harengula clupeola
Pellona harroweri
CYNOGLOSSIDAE
Symphurus plagusia
DIODONTIDAE
Chilomycterus spinosus
ENGRAULIDAE**
Anchoa parva
Cetengraulis edentulus
Lycengraulis grossidens
EPHIPPIDAE
Chaetodipterus faber
GERREIDAE
Diapterus rhombeus
Eucinostomus argenteus
E. gula
E. melanopterus
GOBIIDAE
Gobionellus oceanicus
HAEMULIDAE***
Genyatremus luteus
P. corvinaeformis
PARALICHTHYIDAE
Citharichthys arenaceus
C. spilopterus
Etropus crossotus
Paralichthys tropicus
SCIAENIDAE
Bairdiella ronchus
C. gracillicirrhus
Cynoscion acoupa
C. leiarchus
C. microlepidotus
C. virescens
Isopisthus parvipinnis
Macrodon ancylodon
M. americanus
Micropogonias furnieri
P. brasiliensis
Stellifer brasiliensis
S. rastrifer
SERRANIDAE
Diplectrum radiale
STROMATEIDAE
Peprilus paru
SYNODONTIDAE
Synodus foetens
TETRAODONTIDAE
Lagocephalus laevigatus
Sphoeroides testudineus
TRICHIURIDAE
Trichiurus lepturus
TRIGLIDAE
Prionotus punctatus
Total
n
57
53
16
CT (cm)
Média (? ) s (? ) e Máx.*
n
CP (cm)
Média (? ) s
(? ) e
n
Média
Peso (g)
(? ) s
(? ) e
57
53
16
29,29
23,58
31,81
29,65
17,18
26,64
3,93
2,36
6,66
9,82 3,23
9,71 2,43
10,94 2,42
0,43
0,33
0,60
25
30
30
56
50
16
7,71
7,65
8,52
2,64
1,95
1,92
0,35
0,27
0,48
9
7767
2440
593
937
17,88
13,64
13,05
19,59
12,74
3,92
4,10
4,73
6,50
6,58
1,31
0,05
0,01
0,27
0,21
50
30
35
100
120
9
7779
2453
606
950
13,47
11,02
10,40
9,91
10,35
3,71
3,50
3,74
5,24
5,17
1,24
9
0,04 18833
0,07 3979
0,21 672
0,17 1439
42,78
23,86
19,49
34,23
28,22
27,98
19,18
29,36
152,70
71,57
9,33
0,14
0,46
5,89
1,89
1
2
20
3
24
6,20
5,10
14,33
12,77
5,61
1,00
2,25
1,55
1,18
0,70
0,50
0,89
0,24
80
30
45
30
50
1
2
20
3
24
4,80
4,15 0,92
11,58 1,87
10,30 1,10
4,43 0,92
0,65
0,42
0,63
0,19
1
2
20
3
24
0,80
2,90
24,00
18,00
3,56
1,56
10,14
7,00
1,75
1,10
2,27
4,04
0,36
35
9
14,09 1,25
5,46 2,26
0,21
7,46
17
16
35
9
11,24 1,05
4,46 1,74
0,18
0,58
35
9
33,57
4,17
8,34
4,97
1,41
1,65
246
13,28 2,46
0,16
23
-
-
-
-
250
22,60
14,20
0,90
17
10,58
3,78
0,92
61
17
8,65
3,17
0,77
17
95,62
72,41
17,56
13
36
11
12,58
15,07
16,57
0,82
2,33
1,80
0,23
0,38
0,54
15
16
27
13
36
11
10,43 1,52
12,37 1,99
13,63 1,39
0,42
0,33
0,42
13
36
11
23,08
33,62
31,14
4,80
10,71
13,19
1,33
1,78
3,98
132
10,84 5,02
0,44
90
133
8,54
3,89
0,34
133
69,65
143,33
12,43
1
1
2
2
12,50
13,00
9,25 0,35
10,55 1,06
0,25
0,75
40
30
25
22,2
1
1
2
2
9,10
9,80
7,30
7,95
0,28
1,06
0,20
0,75
1
1
2
2
30,00
22,50
12,35
13,50
0,21
2,12
0,15
1,50
4
19,27 6,98
3,49
>20
4
13,60 4,62
2,31
4
15,87
30,12
7,93
81
3
8,55
10,33
3,16
2,64
0,35
1,52
30
25
81
3
6,69
8,53
2,56
2,20
0,28
1,27
81
3
16,82
13,67
19,51
14,22
2,17
8,21
265
171
182
3
10,61
10,53
10,14
26,70
2,33
2,81
1,77
12,90
0,14
0,21
0,13
7,45
15
20
11
50
266
171
182
3
8,72
8,71
8,28
21,53
2,00
2,41
1,51
10,39
0,12
0,18
0,11
6,00
305
171
363
3
13,03
15,20
12,19
253,00
9,33
15,04
5,16
303,79
0,53
1,15
0,27
175,40
23
20
13
2
79
23
421
68
14
571
77
179
3748
12,14
8,12
15,43
15,25
14,42
22,14
11,60
20,45
23,27
11,59
15,10
10,09
9,40
3,71
1,82
4,46
3,18
3,74
2,55
2,76
4,78
4,23
2,44
2,31
1,75
2,39
0,77
0,41
1,24
2,25
0,42
0,53
0,13
0,58
1,13
0,10
0,26
0,13
0,04
35
21
100
60
100
95
24
45
50
60
30
18
20
23
20
14
2
80
23
421
68
14
573
77
179
3777
9,69
6,41
12,06
11,30
11,62
17,77
9,84
16,29
19,99
9,21
11,91
7,74
7,26
3,10
1,38
3,64
0,85
3,15
1,95
5,28
3,74
5,21
2,08
1,96
1,36
2,30
0,65
0,31
0,97
0,60
0,35
0,41
0,26
0,45
1,39
0,09
0,22
0,10
0,04
23
20
14
2
80
23
473
68
14
579
78
245
5070
27,70
8,99
36,04
25,15
31,64
81,96
16,02
77,32
165,84
16,67
34,22
12,18
11,62
27,16
4,57
37,07
3,75
23,65
29,53
13,43
49,09
108,28
12,68
18,26
6,91
21,28
5,66
1,02
9,91
2,65
2,65
6,16
0,62
5,95
28,94
0,53
2,07
0,44
0,30
3
14,50 2,97
1,71
30
3
11,70 2,34
1,35
3
43,23
31,86
18,39
1
14,10
-
-
30
1
10,60
-
-
1
55,00
-
-
1
20,00
-
-
36,5
1
17,20
-
-
1
47,00
-
-
0,32
0,08
16
544
12,64
84,16
6,17
34,80
1,54
1,49
16
543
8,67 1,56
15,13 2,19
0,39
0,09
100
20
16
544
45
35,70 18,76 2,80
150
-
-
-
-
45
77,05
49,30
7,35
10
7,22
40
10
5,76
0,98
0,31
10
4,37
2,38
0,75
18993
1,24
0,39
18785
6,99 1,30
12,21 1,85
Máx.*
16000
1470
10000
2500
3000
3500
33857
*Maiores comprimentos totais relatados na literatura específica (Corrêa 1987),**Engraulididae por alguns autores,
***Pomadasyidae incluída.
151
Tabela. 46. Número de exemplares capturados em freqüências absolutas (n), absoluta acumulada,
relativas em % e acumuladas em %, por intervalos de classes de CT (cm).
CT (cm)
n
(0; 2]
(2; 4]
(4; 6]
(6; 8]
(8; 10]
(10; 12]
(12; 14]
(14; 16]
(16; 18]
(18; 20]
(20; 22]
(22; 24]
(24; 26]
(26; 28]
(28; 30]
(30; 32]
(32; 34]
(34; 36]
(36; 38]
(38; 40]
(40; 42]
(42; 44]
(44; 46]
(46; 48]
(48; 50]
(50; 52]
(52; 54]
(54; 56]
(56; 58]
(58; 60]
(60; 62]
(62; 84]
TOTAL
2
79
418
1844
4054
3174
2944
2785
1924
836
428
206
113
51
26
22
27
11
6
4
5
5
1
7
3
3
1
4
4
2
3
1
18993
Freqüência
acumulada
2
81
499
2343
6397
9571
12515
15300
17224
18060
18488
18694
18807
18858
18884
18906
18933
18944
18950
18954
18959
18964
18965
18972
18975
18978
18979
18983
18987
18989
18992
18993
18993
Freqüência relativa
(%)
0,01
0,42
2,20
9,71
21,34
16,71
15,50
14,66
10,13
4,40
2,25
1,08
0,59
0,27
0,14
0,12
0,14
0,06
0,03
0,02
0,03
0,03
0,01
0,04
0,02
0,02
0,01
0,02
0,02
0,01
0,02
0,01
100,00
Freqüência
acumulada (%)
0,01
0,43
2,63
12,34
33,68
50,39
65,89
80,56
90,69
95,09
97,34
98,43
99,02
99,29
99,43
99,54
99,68
99,74
99,77
99,79
99,82
99,85
99,85
99,89
99,91
99,92
99,93
99,95
99,97
99,98
99,99
100,00
100,00
Tabela. 47. Número de exemplares capturados em freqüências absolutas (n), relativas em % e
acumuladas em %, por intervalos de classes de peso (g).
Peso (g)
n
Freqüência
Freqüência
Freqüência
acumulada
relativa (%)
acumulada (%)
(0; 50]
31200
31200
92,15
92,15
(5; 100]
2102
33302
6,21
98,36
(100; 150]
376
33678
1,11
99,47
(150; 200]
84
33762
0,25
99,72
(200; 250]
28
33790
0,08
99,80
(250; 300]
17
33807
0,05
99,85
(300; 350]
15
33822
0,04
99,90
(350; 400]
8
33830
0,02
99,92
(400; 450]
3
33833
0,01
99,93
(450; 500]
6
33839
0,02
99,95
(500; 600]
18
33857
0,05
100,00
TOTAL
33857
33857
100,00
100,00
152
Tabela 48. Médias de CT (cm), CP (cm) e peso (g) nos pontos amostrais associados às épocas do ano, dos
peixes capturados na Baía de Guaraqueçaba, com os respectivos tamanhos das amostras (n) e médias,
desvios (±s) e erros (±e) padrões das estimativas. Fonte: dados de campo
Ponto
Amostral
Época
Primavera
Ponto 1
Verão
Outono
Inverno
Primavera
Ponto 2
Verão
Outono
Inverno
Primavera
Ponto 3
Verão
Outono
Inverno
Ponto 4 Primavera
Verão
Outono
Inverno
Geral
n
2511
1632
1710
485
902
1303
2172
112
2396
746
531
205
1270
1769
1006
243
18993
CT (cm)
Média (±) s
14,12 3,63
13,46 5,42
12,63 4,32
11,79 3,17
13,25 3,73
12,10 4,43
10,55 2,99
12,07 3,29
12,87 4,45
10,17 4,31
11,78 4,02
13,36 4,42
14,13 6,71
11,78 5,13
11,22 4,64
10,11 3,28
12,46 4,63
(±) e
0,07
0,13
0,10
0,14
0,12
0,12
0,06
0,31
0,09
0,16
0,17
0,31
0,19
0,12
0,15
0,21
0,03
n
2505
1619
1678
472
899
1307
2142
96
2393
758
533
204
1268
1770
990
243
18877
CP (cm)
Média (±) s
11,34 2,96
10,56 4,04
10,11 3,53
9,52
2,66
10,50 2,48
9,52
4,57
8,46
2,37
9,74
2,89
10,19 3,48
7,99
3,60
9,59
3,37
10,67 3,50
11,31 5,12
9,31
4,16
8,98
3,36
8,49
6,94
9,93
3,80
(±) e
0,06
0,10
0,09
0,12
0,08
0,13
0,05
0,30
0,07
0,13
0,15
0,24
0,14
0,10
0,11
0,45
0,03
n
3601
2783
5579
707
915
1882
5502
112
3066
943
613
205
1386
2375
3945
243
33857
Peso (g)
Média (±) s
32,50 18,32
30,54 24,64
22,84 16,85
19,63 20,26
35,51 34,52
22,70 39,63
11,15 14,08
25,11 27,24
24,05 32,70
14,69 19,89
25,77 47,41
34,72 41,59
47,74 123,73
22,96 36,08
13,84 21,82
17,69 37,09
22,82 36,30
(±) e
0,07
0,47
0,23
0,76
1,14
0,91
0,19
2,57
0,59
0,65
1,91
2,90
3,32
0,74
0,35
2,38
0,20
Tabela 49. Variâncias individuais e cumulativas (em %) da ACP resultantes da comparação dos intervalos de classe
de CT e de peso das 10 espécies mais abundantes (como descritores) e meses e pontos amostrais (como objetos) na
Baía de Guaraqueçaba, Pr, Br.
Componente Variância Variância
número
(%)
acumulada
(%)
Cathorops spixii
1
37,77
37,77
2
18,00
55,78
3
12,85
68,63
4
10,92
79,55
5
8,46
88,01
6
5,48
93,49
7
3,20
96,70
8
1,974
98,67
9
1,33
100,00
Citharichthys arenaceus
1
72,09
72,09
2
16,33
88,42
3
8,97
97,40
4
2,14
99,53
5
0,31
99,85
6
0,15
100,00
Etropus crossotus
1
65,21
65,21
2
25,02
90,22
3
5,21
95,44
4
2,29
97,73
5
1,33
99,06
6
0,68
99,74
7
0,22
99,96
8
0,03786
100,00
64,06
Genidens genidens
1
64,06102
2
21,20796
85,27
3
10,81925
96,09
4
2,23915
98,33
5
0,72223
99,05
6
0,47920
99,53
7
0,41936
99,95
8
0,03712
99,98
9
0,01470
100,00
Isopisthus parvipinnis
1
57,39247
57,39
2
24,24533
81,64
3
13,27721
94,91
4
3,47718
98,39
5
1,19393
99,59
6
0,36943
99,95
7
0,03783
99,99
8
0,00663
100,00
Espécie
Espécie
Micropogonias furnieri
Netuma barba
Sciadeichthys luniscutis
Sphoeroides testudineus
Stellifer rastrifer
Componente Variância
Variância
número
(%)
acumulada (%)
1
2
3
4
5
6
1
2
3
4
5
6
7
1
2
3
4
5
6
7
8
1
2
3
4
5
6
7
8
1
2
3
4
5
6
7
68,25
15,52
12,27
2,45
1,51
0,00
38,06
28,59
16,42
10,92
5,37
0,62
0,01
38,71
33,79
13,13
9,66
2,50
1,46
0,53
0,26
59,69
14,48
12,13
6,73
3,78
2,16
1,02
0,00
60,06
17,87
11,26
6,73
2,84
0,97
0,26
68,25
83,77
96,05
98,49
99,99
100,00
38,06
66,65
83,07
93,99
99,37
99,99
100,00
38,71
72,51
85,64
95,26
97,75
99,21
99,74
100,00
59,69
74,18
86,30
93,04
96,82
98,98
99,99
100,00
60,06
77,93
89,19
95,92
98,76
99,74
100,00
153
Tabela 50. Distribuição das freqüências absolutas (freq. absol.) e relativas (%) por sexo e estádio de
maturação gonadal, das capturas realizadas entre agosto de 1986 e julho de 1987 na Baía de
Guaraqueçaba (Pr, Br).
Sexo
Freq. absol.
(%)
Estádio de Maturação
Freq. absol.
(%)
Indeterminado
17065
67,60
Imaturo
16507
65,66
Macho
3505
13,88
Descanso
3839
15,27
Fêmea
4674
18,52
Maturação
1056
4,20
Maduro
608
2,42
Reprodução
763
3,04
Desovado
2366
9,41
Total
25244
100,00 Total
25139
100,00
Tabela 51. Distribuição das freqüências absolutas (número de exemplares) das capturas por ponto amostral, mês e
estádios de maturação gonadal.
Ponto
Amostral
1
2
3
4
TOTAL
Mês
Imaturo
Descanso
AGO
SET
OUT
NOV
DEZ
JAN
FEV
MAR
ABR
MAI
JUN
JUL
Total
AGO
SET
OUT
NOV
DEZ
JAN
FEV
MAR
ABR
MAI
JUN
JUL
Total
AGO
SET
OUT
NOV
DEZ
JAN
FEV
MAR
ABR
MAI
JUN
JUL
Total
AGO
SET
OUT
NOV
DEZ
JAN
FEV
MAR
ABR
MAI
JUN
JUL
Total
129
61
116
411
123
337
230
60
694
115
175
300
2751
4
58
72
19
268
315
201
942
3339
519
407
27
6171
59
967
311
306
342
264
161
215
219
81
64
16
3005
52
191
187
97
237
402
346
221
2601
207
32
7
4580
16507
18
70
173
164
454
333
226
288
345
37
12
29
2149
3
8
29
4
59
92
141
104
22
31
19
4
516
9
207
44
61
47
54
16
15
11
7
0
4
475
14
129
78
10
119
145
69
53
44
31
4
3
699
3839
em
maturação
32
42
177
29
10
11
13
32
38
6
3
9
402
6
13
42
1
16
21
29
7
5
18
5
2
165
0
242
31
8
7
8
2
1
5
1
0
11
316
7
68
52
1
6
4
4
20
5
4
0
2
173
1056
Maduro Reprodução Desovado
Esgotado
27
20
2
41
17
0
41
144
52
87
120
526
4
22
71
6
14
51
9
1
45
15
10
23
32
5
11
2
0
3
0
1
0
2
0
9
266
354
793
0
0
0
18
4
5
66
90
31
3
12
0
1
1
90
17
35
118
20
0
88
2
6
59
3
0
14
4
2
14
0
0
0
1
0
0
135
150
419
1
0
0
25
6
54
23
21
13
4
14
41
5
21
246
24
16
8
3
0
23
0
0
30
1
0
2
1
0
20
0
0
1
4
0
11
91
78
449
1
0
0
14
10
3
61
110
22
7
13
182
5
28
103
2
12
176
3
0
103
20
6
78
3
0
1
0
2
16
0
0
21
0
0
0
116
181
705
608
763
2366
Incubando
ovos
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
32
0
0
0
0
0
0
0
0
32
32
Total
228
231
703
1337
684
752
524
428
1125
163
191
349
6715
13
106
330
39
435
598
479
1120
3383
588
431
34
7556
69
1501
443
434
668
374
205
261
238
110
65
46
4414
74
415
510
342
498
741
525
398
2654
260
57
12
6486
25171
154
Tabela 52. Variâncias individuais e cumulativas (em %) da ACP resultantes da plotagem
simultânea da freqüência por estádio de maturação gonadal (como descritores) e meses e pontos
amostrais (como objetos), das espécies com reprodução moderada, intensa e muito intensa na
Baía de Guaraqueçaba, Pr, Br.
Espécie
Cathorops spixii
Genidens genidens
Isopisthus parvipinnis
Sciadeichthys luniscutis
Sphoeroides testudineus
Stellifer rastrifer
Componente No.
1
2
3
4
5
6
1
2
3
4
5
6
7
1
2
3
4
5
6
1
2
3
4
5
6
7
1
2
3
4
5
6
1
2
3
4
5
6
Variância
39.40
17.87
16.30
15.51
8.88
2.03
60.82
16.71
10.95
6.86
2.44
1.42
0.80
33.78
20.69
17.43
14.91
7.68
5.513
35.56
18.97
16.80
13.29
8.56
6.73
0.07
39.88
25.93
17.01
10.86
4.21
2.10
39.29
24.47
16.78
14.87
2.81
1.77
Variância acumulada
39.40
57.28
73.57
89.09
97.97
100.00
60.82
77.53
88.48
95.34
97.78
99.20
100.00
33.78
54.46
71.90
86.81
94.49
100.00
35.56
54.53
71.33
84.62
93.20
99.92
100.00
39.88
65.81
82.82
93.68
97.89
100.00
39.28
63.76
80.54
95.41
98.22
100.00
Tabela.53. Comparação entre as atividades principais e secundárias declaradas pelos
entrevistados das localidades da Vila Guarani e Eufrasina. Fonte: dados de campo.
Local
Guaraqueçaba n=46
Atividade
Principal
Secundária (%)
(%)
Pesca
73,91
60,87
Canoeiro
0,00
2,17
Turismo
4,35
4,35
Agricultura
0,00
10,87
Aposentado
10,87
0,00
Bar/Mercearia
0,00
4,35
Funcionário público
6,52
0,00
Prestação de Serv.
4,35
17,39
155
Tabela 54. Locais de pesca declarados por ordem de preferência pelos entrevistados em
Guaraqueçaba (n=46). Fonte: dados de campo.
Local de Pesca
Distância da
Ordem de Preferência e
Localidade
Num. Pescadores (%)
(km)
1a
2a
3a
Baía de Guaraqueçaba
2
95,65
Ilha das Peças
19
2,17
2,17
Ilha Rasa
11
6,52
Baía dos Pinheiros
22,5
6,52
2,17
Itaqui
16,5
2,17
Baía de Tibicanga
18
2,17
Superagüi
31
2,17
Guapicú
12
2,17
Tagaçaba
15
2,17
Varadouro
31
2,17
Cananéia
68
2,17
Tabela. 55 . Resultado dos três primeiros componentes principais (ACP) com as
respectivas variâncias individuais e cumulativas. Fonte :dados de campo.
Componente
Porcentagem da variância
Porcentagem cumulativa
1
74,45602
74,45602
2
25,53660
99,99262
3
0,00738
100,00000
156
Tab. 56. Lista comparativa da composição ictiofaunística para as capturas com arrasto com portas a nível familiar, genérico e específico (acima de
75% de ocorrência nas amostras).
Região
Autor
Famílias
Gen. Espécies
Argentina ao Rio da Prata a Baía de
Sciaenidae, Engraulidae,
Variável, com limites de distribuição definidos
USA
Chesapeake
Vieira & Musick
Gerreidae, Ariidae, Sparidae,
por fronteiras geográficas e hidrológicas.
1994
Haemulidae
Sudeste e
São Paulo ao Rio
Sul do Brasil Grande do Sul
Paraná
Guaraqueçaba
Vieira & Musick
1994
Juvenis e subadultos de
Sciaenidae
Ariidae
Dominância numérica de juvenis,
subadultos e adultos de Sciaenidae
Ariidae e
Tatraodontidae
Paralonchurus brasiliensis
Micropogonias furnieri
Isopisthus parvipinnis
Stellifer brasiliensis
Stellifer rastrifer
Cathorops (=Arius) spixii
Sciadeichthyes (=Arius) luniscutis
Netuma barba
N. planifrons
Genidens genidens
Todas as acima com exceção de
N. planifrons
porém com dominância numérica de
C. spixii
N.barba
S.rastrifer
Sphoeroides testudineus
157
Tabela.57. Lista comparativa da composição ictiofaunística para as capturas, no sistema estuarino da Baia de Paranaguá, a nível familiar, genérico e específico (acima de 75% de
ocorrência nas amostras).
Habitat
Zona de
Arrebentação
Região no Sistema da
Baía de Paranaguá
Pontal do Sul a
Praia de Leste
Pontal do Sul
Ilha do Mel
Baguaçú
Gamboa
Perequê
Embocadura
de rios
Rio Ribeirão
Rio do Maciel
Rio do Maciel
Enseadas &
Baías
Benito
Ilha do Mel
Aparelho
Famílias
Espécies
>Capturas
>No. de
Espécies
Fator
Autor
Espera, arrasto de
praia, com portas e
Picaré
Carangidae
Engraulidae
Clupeidae
Sciaenidae
Mugilidae
Caranx latus
Cetengraulis edentulus
Harengula clupeola
Isopisthus parvipinnis
Mugil curema e M. gaimardianus
Verão, Outono e
Primavera
Verão e
Outono
(-)
Maehama & Corrêa
1987
Eucinostomus argenteus
Verão e Inverno
H. clupeola
Achoa tricolor
H.clupeola
Picaré
Atherinella brasiliensis
Outono
Trachinotus goodei, T. carolinus e O.saurus
Picaré
A.parva
H. clupeola
Outono
Espera
Jenynsia lineata
E. argenteus
(-)
Espera
Bairdiella ronchus
Outono
(3 malhas)
Oligoplithes saurus
Licengraulis grossidens
Genidens genidens
Arrasto de praia,
A.brasiliensis
Atherinidae
com portas, puçás e
E.gula e argenteus
(-)
Gerreidae
Sphoeroides testudineus
covos
Tetraodontidae
E.argenteus
Arrasto com portas
Gerreidae
Verão e
Pomadasys corvinaeformis
Pomadasyidae
Outono
Cynoscion. leiarchus
Sciaenidae
Prionotus punctatus
Triglidae
Arrasto com portas
Ariidae
Cathorops spixii e Sciadeichtys luniscutis
Paralichthyidae
Citharichthys spilopterus
Inverno e
Sciaenidae
Stellifer stellifer
Primavera
Verão e
Inverno
R.Ictioplâncton e
Picaré
Arrasto com portas
Arrasto com portas
Ilha do Mel
Ilha do Mel
Guaraqueçaba
Arrasto com portas
Arrasto com portas
Gerreidae
Clupeidae
Engraulidae
Clupeidae
Atherinidae
Carangidae
Engraulidae
Clupeidae
Poeciliidae
Gerreidae
Scienidae
Carangidae
Engraulidae
Ariidae
Gerreidae
Sciaenidae
Gerreidae
Paralichthyidae
Serranidae
Ariidae
Gerreidae
Pomadasyidae
Sciaenidae
Ariidae
Paralichthyidae
Sciaenidae
Tetraodontidae
Cathorops spixii
E. melanopterus
S. rastrifer
E. gula
C.spilopterus
Diplectrum radiale
C. spixii
E. argenteus
P. corvinaeformis
Micropogonias furnieri
C.spixii, Netuma barba
C. arenaceus
S.rastrifer
S.testudienus
Verão e
Inverno
Outono
(-)
Outono
(-)
Verão e
Outono
Temperatura da água,
Godefroid 1997
período e altura das
ondas
Temperatura da
água,salinidade e
Pinheiro,1999
profundidade
Precipitação, temperatura
da água e salinidade
Vendel, 1997
Precipitação, temperatura
da água e salinidade
Barletta et. al. 1990
Precipitação, temperatura
da água e salinidade
Fernandes-Pinto
1997
Precipitação,
temperatura da água e
salinidade
(-)
Silva & Pose,1990
Corrêa &
Maehama.1987
Inverno e
Primavera
Precipitação,
temperatura da água e
salinidade
Verão e
Outono
Verão e
Outono
(-)
Corrêa & Maehama
1987
Verão e
Outono
Verão e
Outono
(-)
Abilhôa 1997
Outono
Verão e
Outono
Temperatura da
água,salinidade e
profundidade
Pinheiro, 1999
Outono
(densidade)
Primavera
(peso)
Inverno e
Primavera
Precipitação,
temperatura da água,
salinidade, profundidade
e transparência
Obs: ">"=maior, "fator" =principais parâmetros apontados no aumento das capturas e número de espécies, (-) não apontado ou não declarado.
Fernandes-Pinto
1997
158
Tabela 58. Estimativa do número de espécies para o arrasto com portas nos ambientes do sistema estuarino da Baía de Paranaguá.
Habitat
Região
Autor
Aparelho
No.
Amostras
No.
Exemplares
(-)
Sistema estuarino da
Baía de Paranaguá
Corrêa1987
Variado
Extensiva
1572
Pontal do Sul
Cunha 1988
Picaré
12
329
Pontal do Sul
Godefroid 1997
12
Extensiva
4641*- 649**
14516
~37380
Zona de
Arrebentação
Gamboa
Pontal do Sul a
Praia de Leste
Baguaçú
Perequê
Rio Ribeirão
Rio do Maciel
R.Ictioplâncton
Picaré
Maehama & Corrêa 1987 Espera, arrasto de praia e
com portas
Vendel 1997
Picaré
Barletta et. al. 1990
Espera
Fernandes-Pinto 1997
Espera (3 malhas)
Arrasto de praia
Silva & Pose,1990
Embocadura de
rios
Rio do Maciel
Enseadas &
Baías
Benito
Ilha do Mel
Ilha do Mel
Ilha do Mel
Corrêa & Maehama
1987
Fernandes-Pinto 1997
Abilhôa 1997
Corrêa & Maehama 1987
Pinheiro 1999
Guaraqueçaba
BAÍA DE
Chaves & Corrêa 1998
GUARATUBA
» Estimado com base na rarefação (Ludwig & Reynolds, 1988)
*ovos **larvas (-) não declarado
No.
Famílias
No.
spp
56
142
(-)
9
32
25383
2581
282
Estimat. No.
spp»
14
36
12 74
70
102
18
21
15
36
43
23
34
6
72
5
5
23
5
Arrasto com portas
Redes de espera
5
5
15
68
5
9
Tarrafa
Puçás
5
5
74
8
Arrasto com portas
12
(-)
(-)
34
(-)
Arrasto com portas
Arrasto com portas
Arrasto com portas
Arrasto com portas
5
48
12
12
784
1338
(-)
893
12
25
(-)
13
26
37
31
31
21 a 26
24 a 30
(-)
21 a 27
Picaré
12
4109
15
31
Arrasto com portas
Arrasto com portas, tarrafa
e espera
96
Extensiva
33868
(-)
19
28
52
60
188
6
16
9
1
7
12
10
2
4 a 12
21
159
Tabela 59. Comparação dos resultados relacionados à reprodução, relatados na literatura específica e os observados para as espécies mais abundantes
da Baía de Guaraqueçaba.
Época
Incubação
1a. maturação
Recrutamento
reprodutiva
oral
cm
Espécie
Literatura
Guaraqueçaba
Literatura
Guaraqueçaba
Literatura
Guaraqueçaba
Cathorops spixii
Nov. a Jan.*
Dez. a Jan.** Nov. a Fev.
M=9,6 e
11,70
> Out(1e 4)
> Abril
F= 9,8*
Set(1) a Nov(1)
(2 e 4)
Genidens genidens
Nov. a Jan.*
Nov. a Jan. *
Nov. a Jan.
15,5 *
13,99
> Dez (3)
> Set (3) e
Jan.
e
Fev.**
Nov(1) a Jan(3)
Abril(3e2)
Isopisthus parvipinnis
Primavera*# Múltipla > Set (4)
(×)
(×)
M=10,9 e
14,67
> Abril(1) e
F=11,7*
Mar(1) e Mai(2)
Maio(2)
Sciadeichthys luniscutis
Nov. a Jan.* > Out(4)eNov(4)
(× )
Out. a Dez.
18,00*
31,00
> Abril(3)
Out. a Dez.
Sphoeroides testudineus
(-)
(× )
(×)
(-)
15,99
Não detectado.
> Set(3) e Out(2)
na área
Set a Dez
Stellifer rastrifer
múltipla ** Múltipla > Nov(1)
(×)
(×)
M=9,7 e
10,80
> Abril(4)
Out. a Dez.**
F=9,5*
Set. a Abr.
Castello et al. 1994* e Chaves & Vendel, 1997**, Chaves et al. 1998 #.
Os números entre parênteses correspondem aos pontos amostrais.
(> ) intensa
(×) não aplicável
(-)não relatado nas literaturas consultadas.
Download