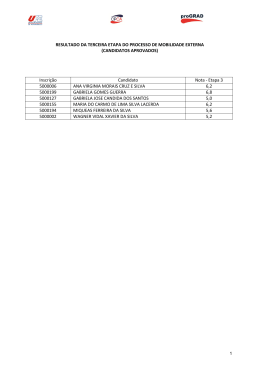
Universidade de Lisboa Faculdade de Medicina de Lisboa Estudo Epidemiológico de Candidíase Invasiva na Unidade de Cuidados Intensivos do Hospital Egas Moniz-Centro Hospitalar Lisboa Ocidental Ana Gomes Paróla Mestrado em Microbiologia Clínica 2008-2010 A impressão desta dissertação foi aprovada pela Comissão Coordenadora do Conselho Científico da Faculdade de Medicina de Lisboa, em reunião de dia 15 de Março de 2011 Universidade de Lisboa Faculdade de Medicina de Lisboa Estudo Epidemiológico de Candidíase Invasiva na Unidade de Cuidados Intensivos do Hospital Egas Moniz-Centro Hospitalar Lisboa Ocidental Ana Gomes Paróla Mestrado em Microbiologia Clínica Dissertação orientada pela Professora Doutora Graciete Freitas Co-orientação pela Professora Doutora Maria Manuel Lopes Todas as afirmações efectuadas no presente documento são da exclusiva responsabilidade do seu autor, não cabendo qualquer responsabilidade à Faculdade de Medicina de Lisboa pelos conteúdos nele apresentados. Agradecimentos Nada que envolva um trabalho desta envergadura pode existir sem esforço, ajuda e compreensão de inúmeras pessoas. Por isso, nesta jornada devo agradecer… À Professora Graciete e à Professora Mané pela sua orientação nestes anos de caos, mesmo que organizado À Helena Brás e à Lavínia pela sua preciosa ajuda no Laboratório de Micologia Ao Dr. Eduardo Monteiro, à Dra. Isabel Rio de Carvalho e à Dra. Teresa Pacheco por terem acreditado neste projecto de tese À Dra. Helena Farinha pelo incentivo na finalização deste projecto Ao Dr. Pedro Santos e Dr. João Furtado pela ajuda preciosa na recolha dos dados populacionais, com sacrifício pessoal para ambos Aos colegas da Farmácia do Hospital Egas Moniz, em especial à Paula Chambel, Luísa Fetal e Ricardo Soares, pelo seu ombro amigo e opiniões francas Ao Nuno, pelas noites mal dormidas, quando já grávida de meses, ficou firme ao meu lado Aos meus pais, pelas múltiplas correcções do texto, quando o cansaço já batia à porta Como diz Jorge Luís Borges no seu Livro de Areia …‖ Ninguém pode ler 2 mil livros. Vivo há quatro séculos e não terei ultrapassado a meia dúzia. Aliás o que importa não é ler, mas reler‖ I Resumo Resumo Desde o início da década de 80 que os fungos têm emergido como agentes patogénicos, especialmente em doentes hospitalizados e imunocomprometidos. A mortalidade associada a este tipo de infecção é elevada, estando relacionada com diferentes tipos de fungos, sobretudo dos géneros Candida, Cryptococcus, e Aspergillus. Em Portugal, os dados epidemiológicos são escassos, pelo que o estudo epidemiológico de candidíase invasiva se reveste de alguma importância. Como tal, propusemo-nos estudar a epidemiologia de Candidíase Invasiva (CI) na Unidade de Cuidados Intensivos do Hospital Egas Moniz, durante o ano de 2009 (Maio a Dezembro), com recolha de dados populacionais e respectiva análise estatística. Realizaram-se testes de susceptibilidade dos isolados frente ao voriconazol, fluconazol, caspofungina, anidulafungina e anfotericina B, através de 3 métodos: microdiluição, Etest® e método difusão em disco. Observou-se uma incidência de 12,8% de CI. A distribuição de espécies foi a seguinte: C. albicans, 59,3%, C. glabrata, 15,5%, C.krusei, 12,4%, C. tropicalis, 9,7% e C. parapsilosis, 3,1%. Resistência ao fluconazol ocorreu numa percentagem minoritária dos isolados, nomeadamente em C. krusei e C. glabrata. Não se observou qualquer isolado resistente ao voriconazol ou à anfotericina B. Relativamente às equinocandinas estudadas, verificou-se CIMs mais elevadas em isolados de C. parapsilosis e C. guilliermondii. Os Etest® e teste de difusão em discos apresentaram uma concordância categórica com o método de referência superior a 93,34% para todos os fármacos testados. Erros Major, Minor e Muito Graves foram observados infrequentemente. Não se observaram diferenças significativas face a dados apresentados na literatura internacional, relativamente à epidemiologia e presença de factores de risco na nossa população. Constatou-se um peso relativo das espécies não-albicans de cerca de 40%, na II Resumo etiologia de CI. Em termos microbiológicos, foram identificadas resistências esperadas para o fluconazol e não susceptibilidade às equinocandinas, com resultados discrepantes entre metodologias empregues. Palavras-chave: Candidíase Invasiva, Cuidados Intensivos, Epidemiologia, Susceptibilidade aos Antifúngicos. III Resumo Abstract Since the early 80's, fungi have emerged as pathogens, especially in hospitalized and immunocompromised patients. The mortality associated with this type of infection is high and is related with different types of fungi, especially Candida, Cryptococcus, and Aspergillus spp.. In Portugal, the epidemiological data are scarce, so the implementation of epidemiological studies in this area is of some importance. As such, we decided to study the epidemiology of Invasive Candidiasis (IC) in the Intensive Care Unit of the Hospital Egas Moniz, during 2009 (May to December), through populational data collection and respective statistic analysis. We proceeded to the testing of antifungal susceptibility to voriconazole, fluconazole, caspofungin, anidulafungin and amphotericin B, using three methods: microdilution, Etest ® method and disk diffusion. We found an incidence of IC of 12.8%. The species distribution was as follows: C. albicans, 59,3%, C. glabrata, 15,5%, C.krusei, 12,4%, C. tropicalis, 9,7% and C. parapsilosis, 3,1%. Resistance to fluconazole occurred in a minority percentage of the isolates, namely in C. krusei and C. glabrata. There were no resistant isolates to voriconazole or amphotericin B. Regarding echinocandins, we observed higher MICs for C. parapsilosis and C. guilliermondii isolates. The Etest® and disk diffusion test showed a high percentage of categorical agreement with the reference method, over 93.34%, for all drugs tested. Major, Minor and Very Major Errors were very infrequently seen. There were no significant differences regarding the epidemiology and the presence of risk factors in our population compared with data presented in international literature. Nonalbicans species corresponded to about 40% of all isolates responsible for IC. In microbiological terms, we identified expected resistance to fluconazole and non-susceptibility to echinocandins, with discrepant results between methodologies used. IV Resumo Key Words: invasive candidiasis, intensive care, epidemiology, antifungal susceptibility. V Índice Geral Índice Geral Agradecimentos ......................................................................................................................... I Resumo ..................................................................................................................................... II Índice ...................................................................................................................................... VI Índice Tabelas e Gráficos ...................................................................................................... IX Abreviaturas ........................................................................................................................... XI I – Introdução Geral ................................................................................................................ 1 1. Infecções em Unidades de Cuidados Intensivos. Infecções fúngicas História e considerações gerais .................................................................................. 1 2. Epidemiologia e etiologia das infecções fúngicas ............................................................... 9 2.1 Infecções por Candida spp. -Conceitos e definições ............................................... 9 2.2 Incidência de infecções fúngicas invasivas ........................................................... 12 2.3 Papel das leveduras como agentes patogénicos em Cuidados Intensivos ............. 13 2.4 Factores de risco da infecção fúngica .................................................................... 18 2.5 Factores de virulência ............................................................................................. 20 2.6 Susceptibilidade in vitro aos antifúngicos ............................................................. 30 2.7 Mecanismos de resistência aos azóis ..................................................................... 35 2.8 Mecanismos de resistência às equinocandinas ...................................................... 40 2.9 Mecanismos de resistências aos polienos .............................................................. 42 3. Tratamento de Infecções Fúngicas Invasivas ................................................................. 46 4. Métodos disponíveis para avaliação de susceptibilidade aos antifúngico .................... 51 II– Estudo Experimental Objectivos do Estudo ............................................................................................................. 53 VI Índice Geral 1. Caracterização da Candidíase Invasiva na UCI do Hospital Egas Moniz .................... 54 1.1 Materiais e métodos ............................................................................................. 54 1.1.1 População em estudo .......................................................................................... 54 1.1.2 Tratamento prévio das amostras ......................................................................... 55 1.1.3 Identificação de fungos ....................................................................................... 55 1.1.4 Tratamento de dados e análise estatística ........................................................... 55 1.2 Resultados ............................................................................................................ 56 1.2.1 Caracterização da população em estudo ............................................................. 56 1.2.2 Distribuição das espécies responsáveis por Candidíase Invasiva........................ 62 1.2.3 Padrão de utilização de terapêutica antifúngica .................................................. 65 1.3 Discussão .............................................................................................................. 67 1.3.1 Incidência de Candidíase Invasiva e caracterização da população em estudo ... 67 1.3.2 Distribuição das espécies de Candida spp........................................................... 78 1.3.3 Padrão de utilização de terapêutica antifúngica .................................................. 83 2. Perfil de susceptibilidade aos antifúngicos....................................................................... 87 2.1 Materiais e métodos ............................................................................................. 87 2.1.1 Agentes antifúngicos .......................................................................................... 87 2.1.2 Preparação do inóculo de leveduras ................................................................... 87 2.1.3 Método de Microdiluição ................................................................................... 87 2.1.4 Método de difusão em discos ............................................................................. 91 2.1.5 Etest® ................................................................................................................. 92 2.1.6 Controlo de Qualidade ......................................................................................... 94 2.1.7 Tratamento de dados e análise estatística ........................................................... 95 2.2 Resultados ............................................................................................................ 97 VII Índice Geral 2.3 Discussão ............................................................................................................ 102 2.3.1 Susceptibilidade aos antifúngicos...................................................................... 102 2.3.2 Concordância entre métodos de avaliação de susceptibilidade aos antifúngicos .................................................................................................................................... 110 3. Conclusões finais ............................................................................................................. 122 Bibliografia Geral ................................................................................................................ 127 Anexos ................................................................................................................................... 151 VIII Índice Geral Índice de Tabelas e Gráficos Tabela 1. Critérios de diagnóstico de infecção fúngica invasiva ........................................... 11 Tabela 2. Características demográficas da população em estudo e estatística de internamento .................................................................................................................................................. 58 Tabela 3. Tipo de cirurgia prévia à admissão ........................................................................ 58 Tabela 4. Tipos de causas de admissão na UCI ..................................................................... 59 Tabela 5. Ponto de Partida de episódios de CI ....................................................................... 60 Tabela 6. Padrão de utilização de antibioterapia (AB) prévia (nºdias) na população estudada .................................................................................................................................................. 60 Tabela 7. Distribuição de leveduras de acordo com origem e espécie ................................... 62 Tabela 8. Tipo de terapêutica instituída ................................................................................. 65 Tabela 9. Padrão de utilização de terapêutica antifúngica prévia (nºdias) na população estudada. ................................................................................................................................... 66 Tabela 10. A análise descritiva da terapêutica prévia com antifúngicos e emergência de isolados resistentes .................................................................................................................. 66 Tabela 11. Critérios de Interpretação para o método de microdiluição ................................. 90 Tabela 12. Critérios de Interpretação para método de difusão em discos .............................. 92 Tabela 13. Critérios de Interpretação de acordo com CLSI ................................................... 94 Tabela 14. Susceptibilidade in vitro de 4 estirpes Candida spp. para controlo de qualidade 98 Tabela 15. Susceptibilidade in vitro de Candida spp. através de 3 métodos: Microdiluição, Etest®, Difusão em Disco ....................................................................................................... 99 Tabela 16. Percentagem de isolados distribuídos pela categoria de susceptibilidade ........... 100 Gráfico 1. Curva de distribuição de idades na população estudada ...................................... 56 IX Índice Geral Gráfico 2. Curva de distribuição de duração de internamento (dias) na população estudada . 56 Gráfico 3. Tipos de admissão na UCI ..................................................................................... 57 Gráfico 4. Causas de admissão na UCI ................................................................................. 57 Gráfico 5. Tipo de co-morbilidades observadas na população em estudo ............................ 59 Gráfico 6. Distribuição das espécies responsáveis por CI ..................................................... 62 X Índice Geral Abreviaturas ALS – Aglutinin Like Sequence AM3 –Antibiotic Medium 3 ATCC – American Type Culture Collection CAD – Coronary Arterial Disease (doença arterial coronária) CI – Candidíase Invasiva CIM - Concentrações Inibitórias Mínimas CIM50 - Concentração Inibitória Mínima, que inibiu 50% dos isolados CIM90 - Concentração Inibitória Mínima, que inibiu 90% dos isolados CLSI - Clinical Laboratory Standards Institute DD – Difusão em Discos DHC – Doença Hepática Crónica DM – Diabetes Mellitus DMSO – DiMethyl SulfOxide DPOC – Doença Pulmonar Obstrutiva Crónica ECMM - European Confederation of Medical Mycology EM – Erro Major EMG – Erro Muito Grave EORTC/MSG - European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group/National Institute of Allergy and Infectious Diseases Mycoses Study Group ET - Etest® EUCAST - European Committee on Antimicrobial Susceptibility Testing HEM – Hospital Egas Moniz XI Índice Geral HTA – HiperTensão Arterial IC – Insuficiência Cardíaca IDSA - Infectious Diseases Society of America IRA – Insuficiência Renal Aguda IRC – Insuficiência Renal Crónica LDL – Low Density Lipoproteins M – erro Minor MD – MicroDiluição MOF – Multiple Organ Failure NA – Não Aplicável NS – Não Susceptível NT/OT – NasoTraqueal/OroTraqueal pós- op. CT – pós-operatório Cirurgia Torácica R - Resistente RPMI 1640 - Roswell Park Memorial Institute 1640 S - Susceptível SAP - Secretory Aspartyl Proteinase S-DD – Susceptível Dose-Dependente TBA – TraqueoBronquite Aguda TP – Tuberculose Pulmonar UCI – Unidade de Cuidados Intensivos VIH – Vírus Imunodeficiência Humana XII História e considerações gerais I –Introdução Geral 1.Infecções em Unidades de Cuidados Intensivos. Infecções fúngicas História e considerações gerais O nascimento da medicina crítica acompanhou a história da medicina e a evolução hospitalar, bem como a introdução de novos conceitos científicos, económicos e funcionais. A existência de hospitais (em latim hospitale ou domus Dei) ascende à Idade Média. A medicina medieval apoiou-se nas obras de grandes sábios da Antiguidade e nos ensinamentos da medicina árabe, observando-se alguns trabalhos contemporâneos das escolas de Salerno e Montpellier. Contudo, a hospitalização era tradicionalmente voluntária, reservada para os mais desfavorecidos da sociedade, muitas vezes com cariz caritativo (140) . A provisão de cuidados de saúde nos hospitais era, até os finais do século XIX, obrigatória somente em situações de epidemias e em casos de doenças infecciosas, como sífilis e lepra (59). A introdução de sistemas/‖seguros‖ de saúde, quer privados quer públicos, e o aumento do número de hospitais, vieram alterar o paradigma da medicina hospitalar, aumentando o número de hospitalizações (59) . O desenho e a funcionalidade dos hospitais tornam-se, assim, relevantes na provisão de cuidados. Florence Nightingale popularizou a existência de enfermarias abertas, tectos altos, janelas em cada lado, com cerca de 30 camas. Este tipo de alas permitia ventilação cruzada e entrada de luz (devido aos tectos altos e janelas em cada lado), de acordo com o conceito de infecção da época (63). Para além disso, este tipo de enfermarias permitia o recurso a uma pequena equipa de enfermagem, fomentando a camaradagem entre doentes. Florence Nightingale reconheceu, -1- História e considerações gerais todavia, a existência de doentes com condições específicas que punham em risco a sua vida e que requeriam cuidados de enfermagem mais específicos. Surge, assim, o primeiro conceito de ―Unidade de Cuidados Intensivos‖ – quartos privados para doentes críticos, com necessidade de atenção redobrada por parte da equipa de enfermagem (147). O conceito de Florence Nightingale depressa se revelou ineficaz em determinadas situações, com limitações no acesso a instalações sanitárias para os doentes, falta de privacidade, pouca flexibilidade na deslocação de camas, conforme o género dos doentes, e impedimento da expansão dos hospitais devido a problemas arquitectónicos, entre outros (63). O início do século XX é relevante na alteração de alguns paradigmas sobre infecção e assepsia(97). A primeira e segunda guerras mundiais revelam-se propulsoras de conhecimento científico, incluindo técnicas cirúrgicas e aplicação dos conhecimentos em anestesia, ambos já em desenvolvimento desde os finais do século XIX (63). O período entre guerras permite o florescimento de alterações de mentalidade. Neste período, observa-se a consolidação dos hospitais como locais de tratamento clínico de excelência e especializado (59,63). É neste âmbito que nascem as primeiras Unidades de Cuidados Intensivos (UCIs). O médico W.E. Dandy abre uma unidade com 3 camas para pós-operatório de doentes neurocirúrgicos. Em 1927, surge a primeira unidade de cuidados de recém-nascidos prematuros, em Chicago (147) . Neste período, observa-se ainda uma tentativa de tornar os hospitais em crescimento mais funcionais, em termos arquitectónicos. Nos anos 30, observa-se uma subdivisão de enfermarias de internamento em secções mais pequenas, com colocação de camas paralelamente à parede e não perpendicularmente, como anteriormente preconizado (63). -2- História e considerações gerais A compartimentalização de enfermarias torna-se generalizada na década de 30-40 e verificase um abandono da teoria miasmática de infecção. A percepção de infecção, inclusivamente de infecção hospitalar (nosocomial), surgiu no séc. XVI (140) . No séc. XVIII, verificou-se a publicação de trabalhos e o desenvolvimento de pesquisa na compreensão da infecção hospitalar. Esta área foi enriquecida com os conhecimentos de Louis Pasteur, William Halsted, Johann Mikulicz-Radecki, entre outros. O avanço dos conhecimentos médicos em bacteriologia e a teoria dos germes enfatizaram a necessidade de criação de técnicas de assepsia, em vez da ventilação de alas, previamente defendida(63, 97). São também estudados e desenhados materiais, incluindo camas e outros, com superfícies mais adequadas aos procedimentos assépticos defendidos (63). A segunda guerra mundial, devastadora, trava o desenvolvimento hospitalar até aqui implementado, mas permite a criação de alas de trauma, especializadas em ressuscitação e cuidados intensivos cirúrgicos (147). No período pós-guerra, o mundo depara-se com necessidades em infra-estruturas e recursos humanos, e a área da saúde não é excepção. A reconstrução é levada a cabo por entidades públicas e/ou privadas. Elevam-se novas preocupações, identificando-se necessidades de alterações arquitectónicas para diminuir o risco de infecções cruzadas nos doentes hospitalizados (63) . Diversos estudos revelam uma percentagem significativa de doentes com infecções adquiridas no hospital, sendo Staphylococcus aureus meticilina resistente, reconhecido como um agente etiológico relevante. Planeia-se, por isso, o desenvolvimento de áreas específicas para tratamento de feridas cirúrgicas, a compartimentalização e rearranjo de enfermarias para aumentar a flexibilidade organizativa de camas e de recursos humanos, a implementação de ventilação mecânica, cuidados ambulatórios, com diminuição de dias de hospitalização, entre outras medidas (63). -3- História e considerações gerais Em 1948-49, uma epidemia de poliomielite, que assola os Estados Unidos e Europa, conduz a necessidades de cuidados aumentadas em doentes com paralisia respiratória. AS UCIs revelam-se de importância extrema na instituição de ventilação manual (através de tubos de traqueostomia) e provisão de cuidados a este tipo de doentes (147). Avanços científicos em ventilação manual e mecânica permitem a organização e implementação de Unidades de Cuidados Intensivos respiratórios e polivalentes. A partir da década de 60-70, observa-se um grande desenvolvimento de Unidades de Cuidados Intensivos (147, 166) . Esta alteração radical na estrutura organizacional dos hospitais continua até a década de 70, quando a situação económica mundial revela a necessidade de poupança (147). Actualmente, a arquitectura hospitalar desenvolve-se a par e passo com a necessidade de controlo de infecções hospitalares graves. Nos dias de hoje, um novo hospital é construído, enquadrando os ensinamentos do passado e incorporando conceitos como controlo de risco de infecção hospitalar (127). A associação entre a construção e o risco de aspergilose invasiva em doentes imunocomprometidos é conhecida há décadas, assim como a associação entre legionelose adquirida no hospital e água potável (127) . A contaminação do ambiente hospitalar tem sido associada à transmissão de Staphylococcus aureus meticilina resistente e enterococos resistentes à vancomicina, entre outros microrganismos (7, 145). A instalação de ventilação com filtros de alta eficiência (HEPA), a existência de quartos com pressão negativa, o correcto acondicionamento de tanques com água potável, a implementação de medidas de esterilização do ambiente (ar e água), a existência de chãos e paredes facilmente laváveis com detergentes e a utilização de materiais com superfícies planas sem poros, bem como o acesso fácil a lavatórios devidamente equipados para lavagem de mãos, aliados à educação dos doentes e profissionais de saúde em medidas de controlo de -4- História e considerações gerais infecção específicas, são directivas que têm vindo a ser aplicadas em muitas instituições hospitalares (63,127). Paralelamente, as UCIs ganharam um papel preponderante na medicina actual, dados os avanços técnico-científicos (farmacoterapia, cirurgias invasivas, técnicas de monitorização não invasivas, avanços laboratoriais, entre outros) registados nas últimas duas décadas, embora o reconhecimento desta especialidade se tenha dado somente nos anos 80. As UCIs também se têm debatido com grandes problemas no controlo de infecções, sobretudo adquiridas no meio hospitalar. As preocupações com bactérias Gram-, como Pseudomonas aeruginosa, e Gram+, como Staphylococcus aureus, existem já há algumas décadas (7, 94). Desenvolvimento mais lento verificou a disciplina de micologia médica. A história da micologia médica começa, de facto, na 1ª metade do século XIX, com a descoberta da primeira micose humana por Gruby e Remak (um caso de tinea favosa). Por outro lado, Gruby e Schöenlein identificam, em 1939, Trichophyton schoenleinii como um agente fúngico patogénico no homem. No fim do século XIX, já Raimond Jacques Sabouraud tinha realizado um estudo extenso sobre fungos patogénicos em infecções fúngicas humanas por dermatófitos, e Robin tinha descoberto Candida albicans (C. albicans) como agente etiológico de mucosite (42,46). Nos Estados Unidos, a descoberta de Blastomyces dermatitidis (B. dermatitidis), em 1894, e Sporothrix schenckii (S. schenkii), em 1896 foram dois marcos históricos da micologia médica. Paralelamente, foram publicados os primeiros dois casos de infecção por Coccidioides immitis (C. immitis), na Califórnia (42) . Caspar Gilchrist e Benjamin R. Schenk seguem as pisadas dos europeus Louis Pasteur e Robert Koch. Os contributos destes autores foram decisivos para a história da infecção por B. dermatitidis, C. immitis e S. schenckii com -5- História e considerações gerais atribuição de nomenclatura, descrição morfológica do fungo e das manifestações da doença (42) . É também neste período que Busse, Buschke e Sanfelice identificam a levedura Cryptococcus neoformans (C. neoformans) (42). No início do séc. XX, observou-se uma grande expansão desta disciplina, com identificação de agentes etiológicos de outras infecções fúngicas como Histoplasma capsulatum, Phialophora verrucosa e Absidia corymbifera (Mucor corymbifera) (42, 45). Os conhecimentos sobre B. dermatitidis, C. immitis e S. schenckii sedimentaram-se. Adicionalmente, foram reportados os primeiros casos de meningite por C. neoformans. Verificou-se ainda distanciação nos estudos levados a cabo pelos europeus, mais centrados nas micoses superficiais, contraposto por um desenvolvimento mais acentuado dos conhecimentos sobre micoses profundas nos Estados Unidos (42, 45). Na década 20, deu-se a primeira tentativa de descoberta e implementação de testes diagnósticos laboratoriais, com o nascimento do primeiro laboratório de micologia médica na Universidade de Columbia, entre outros centros de investigação a nível mundial (42). A década seguinte permitiu a implementação de sistemas de classificação mais adequados, com correcta diferenciação de agentes patogénicos. Recorrem-se a estudos epidemiológicos e ecológicos para correcta caracterização da história natural da doença dos diferentes tipos de infecção fúngica já conhecidos (42, 45). O despoletar da segunda guerra mundial é outro marco histórico para o desenvolvimento da micologia médica. Datam de 1940 e 1946 os primeiros casos conhecidos de CI – endocardite por Candida spp., num doente utilizador de drogas endovenosas, e meningite por Candida spp., num doente com antibioterapia prévia de duração prolongada. São também documentados os primeiros casos de mucormicose rino-cerebral (42,45). -6- História e considerações gerais Os avanços científicos dão-se tanto no conhecimento da epidemiologia das doenças fúngicas como no desenvolvimento de testes laboratoriais. São também desta década os primeiros estudos sobre acitividade antifúngica de algumas substâncias, nomeadamente benzimidazol (primeiro imidazol) e cicloheximida (42). Nas décadas 50-60, a comunidade científica reconheceu, enfim, o papel das infecções fúngicas na sociedade. Procurou-se uma redução drástica da mortalidade associada a este tipo de infecções (entre 50 e 100%). As descobertas de nistatina (primeiro fármaco antifúngico a ser utilizado nos humanos), de anfotericina B e de flucitosina iniciaram o tratamento adequado de infecções fúngicas (42) . É revelada a relação entre agentes fúngicos oportunistas, como Aspergillus spp., Candida spp., Mucor spp. e C. neoformans, e terapêutica antibiótica, imunossupressora ou citotóxica, e condições subjacentes do hospedeiro (entre as quais presença de neoplasias, diabetes e transplantes). Começou, desta forma, a descobrir-se a relação entre infecções fúngicas oportunistas e doentes imunocomprometidos e a tentar desvendar os mecanismos imunológicos implicados (42, 45) . Assuntos como patogénese e interacção fungo-hospedeiro começam a ser estudados com maior atenção. São igualmente realizados os primeiros ensaios clínicos com terapêutica antifúngica, sendo a anfotericina B o agente antifúngico mais promissor até então desenvolvido. Contudo, a comunidade científica debate-se com problemas de formulação adequada, aparecimento de reacções adversas graves e falência terapêutica em situações clínicas severas (42). Os anos 70 permitiram o crescimento exponencial da micologia médica, nas áreas laboratorial, taxonómica, epidemiológica e clínica. O crescimento da incidência e prevalência das infecções fúngicas oportunistas deveu-se, em parte, ao aumento de uso de antibióticos de largo espectro e de terapêuticas imunossupressoras agressivas, ambos responsáveis por um aumento de susceptibilidade do hospedeiro aos agentes etiológicos fúngicos. O -7- História e considerações gerais desenvolvimento de uma nova classe de fármacos, que inclui o cetoconazol, e a utilização de terapêutica combinada anfotericina B + flucitosina, para o tratamento da meningite criptocócica, são contributos relevantes no florescimento da micologia médica como a conhecemos hoje em dia (42, 46). É também na década de 70, mais propriamente em 1975, que a Organização Mundial de Saúde reconhece, na sua 28ª Assembleia Geral, a importância sanitária e as implicações médico-sociais das micoses (46). Os anos 80 e 90 são anos de viragem, com a descoberta do vírus da imunodeficiência humana e o aparecimento de uma nova classe de imunodeprimidos – doentes com SIDA. Esta pandemia deslocou e desdobrou esforços, deixando pouco espaço para o crescimento da micologia médica. Pouco a pouco, retomou-se esta área com o desenvolvimento de novos fármacos da classe dos imidazóis e novas classes de fármacos (42). Ensaios clínicos, estudos multicêntricos de avaliação da terapêutica antifúngica, estudos genéticos, desenvolvimento de testes laboratoriais com base em conhecimentos de imunohistoquímica encontram-se em crescimento, nos dias de hoje (42). -8- Epidemiologia e etiologia das infecções fúngicas 2. Epidemiologia e etiologia das infecções fúngicas 2.1 Infecções por Candida spp. - Conceitos e definições As espécies do género Candida são ubiquitárias e algumas delas constituem a nossa flora microbiológica. Contudo, apenas 10% das espécies descritas se encontram relacionadas com infecção no ser humano (40, 46). As infecções por espécies do género Candida spp. são frequentes na espécie humana, abrangendo desde infecções mucocutâneas até infecções invasivas com envolvimento de qualquer órgão e elevadas taxas de mortalidade. A CI é, de facto, uma infecção relacionada com os avanços técnico-científicos da medicina e das ciências farmacêuticas actuais (40). A realidade é que Candida spp. faz parte da flora endógena normal da pele e dos tractos gastrointestinal e genito-úrinário, estando documentada colonização temporária ou permanente do tracto gastrointestinal em cerca de 40 a 50% dos seres humanos (40, 50) . A colonização é um pré-requisito para o desenvolvimento de candidíase. Alterações na flora endógena das superfícies da pele e mucosas, bem como perda da integridade da barreira do tracto gastrointestinal podem conduzir a um crescimento excessivo de Candida spp. ou permitir a translocação destas leveduras através do intestino. A exposição contínua a outros factores de risco permitirá invasão e disseminação do fungo. A transmissão exógena do fungo também se encontra documentada, correspondendo, no entanto, a uma via de transmissão secundária, de menor relevância. A colonização exógena pode, inclusivamente, coexistir com colonização endógena (40, 50). Não existe, por isso, consenso na definição de CI, excepto para doentes imunocomprometidos. A European Organization for Research and Treatment of Cancer/Invasive Fungal Infections Cooperative Group em conjunto com o National Institute of Allergy and Infectious Diseases Mycoses Study Group (EORTC/MSG) publicaram -9- Epidemiologia e etiologia das infecções fúngicas definições normalizadas de infecção fúngica invasiva, para uso no contexto de realização de estudos clínicos ou epidemiológicos. Estas definições restringiram-se a doentes com cancro ou que sofreram transplante de células hematopoiéticas. Os dois grupos de trabalho consideraram 3 níveis de probabilidade de diagnóstico: infecção provada, provável ou possível, tendo em consideração factores do hospedeiro, sintomatologia clínica e evidência micológica (10, 37, 40). Estas definições apresentam algumas limitações, nomeadamente a dificuldade de transposição para a prática clínica (10, 37, 40). Este problema subsiste na definição de candidemia e candidíase disseminada, vulgarmente denominadas de CI. A candidemia pode ser definida como, pelo menos, uma hemocultura positiva com sinais clínicos temporalmente relacionados. A candidíase disseminada pode definir-se como a existência de um achado histo ou citopatológico com evidência de leveduras de Candida spp. ou de uma cultura positiva obtida de amostras biológicas provenientes de locais anatómicos normalmente estéreis (10). A candidemia é, na prática clínica, interpretada como um sinal evidente de candidíase disseminada. Todavia, existe alguma controvérsia envolvendo a interpretação de uma hemocultura positiva para Candida spp.. De facto, nem todos os doentes com uma hemocultura positiva para Candida spp. apresentam o mesmo risco de desenvolvimento de candidíase disseminada. Diversos factores do hospedeiro podem ser relevantes na interpretação de hemoculturas positivas. Da mesma forma, o isolamento de Candida spp. em amostras de urina, expectoração ou pus de feridas abertas, pode ser de difícil interpretação, quanto à presença de infecção invasiva por Candida spp. (10). Estes factos colocam desafios na correcta interpretação de todos os dados microbiológicos, sendo difícil distinguir colonização por Candida spp. de infecção em doentes críticos internados em UCIs. - 10 - Epidemiologia e etiologia das infecções fúngicas Tabela 1. Critérios de diagnóstico de infecção fúngica invasiva – adaptado de Ascioglu, S., et al.(10) Factores do Hospedeiro Microbiológicos Clínicos Neutropenia <500neutrofilos/mm3 por um período superior a 10 dias; Febre persistente 96h refractária a terapêutica antibiótica adequada; Neutropenia prolongada nos 60 dias antecedentes; Uso recente ( <30 dias) ou recorrente de terapêutica imunossupressora; SIDA; Infecção fúngica provada ou provável num episódio anterior de neutropenia; Sintomas de rejeição aguda de transplante; Uso prolongado de corticosteróides ( 3 semanas) nos 60 dias antecedentes. Resultados positivos em amostras biológicas provenientes de locais normalmente estéreis; 2 Resultados positivos de leveduras na urina na ausência de cateter urinário; Hemocultura positiva para Candida spp.. Infecção do tracto respiratório Major – Infiltrados radiológicos; Minor – Sintomas de tosse, dispneia, hemoptise, dor torácica, efusão pleural; Infecção do Sistema Nervoso Central Major – Evidência radiológica; Minor – Sintomas neurológicos focais, alterações mentais, alterações da bioquímica e da contagem de células do líquido cefalorraquidiano; Infecção fúngica disseminada Lesões da pele papulares ou nodulares sem outra explicação; Achados intraoculares consistentes com diagnóstico de endoftalmite ou corioretinite fúngica; Candidíase crónica disseminada Abcessos no fígado e baço demonstrados por TAC, RM ou ecografia em concomitância com níveis de ALT elevados; Candidemia Os critérios clínicos não são necessários para candidemia ―provável‖, não se encontra definida candidemia ―possível‖. Consequentemente, é necessário compreender a epidemiologia deste tipo de infecção invasiva, conhecer os factores de risco associados e os sintomas clínicos e relacioná-los com a evidência micológica. - 11 - Epidemiologia e etiologia das infecções fúngicas 2.2 Incidência de infecções fúngicas invasivas Desde o início da década de 80 que os fungos têm emergido como agentes patogénicos, especialmente em doentes hospitalizados e imunocomprometidos. Dados recentes, nos Estados Unidos, revelam que a sepsis devida a fungos aumentou 207% entre 1979 e 2000 (86). Dados de um estudo realizado por Sands et al. indicaram que a sepsis foi responsável por 2% de todas as hospitalizações e que 59% dos doentes com sepsis necessitaram de internamento em UCIs (86, 114) . Entre 1979 e 1987, os microrganismos Gram- foram os agentes etiológicos predominantemente causadores de sepsis. Em 2000, 52,1% dos casos de sepsis foram provocados por bactérias Gram+, 37,6% dos casos por bactérias Gram- e 4,6 % dos casos por fungos (86) . A mortalidade associada a este tipo de infecção é elevada e está relacionada com diferentes tipos de fungos, sobretudo leveduras dos géneros Candida e Cryptococcus, e fungos filamentosos do género Aspergillus. Aliás, Candida spp. é a quarta causa de infecção nosocomial da corrente sanguínea (candidemia), responsável por 8 a 10% de todas as infecções da corrente sanguínea adquiridas no hospital (40, 114) . Um estudo europeu EPIC (European Study on the Prevalence of Nosocomial Infections in Critically Ill) reportou que a (40, 52) CI foi responsável por 17% das infecções adquiridas no hospital representou cerca de 10-20% de todas as formas de candidíase . A candidemia (40) . Diversos factores têm contribuído para o incremento de infecções fúngicas invasivas. Os mais importantes incluem: aumento da população imunodeprimida, utilização de antibióticos de largo espectro e de citotóxicos, e aumento do número e técnicas de transplantes (40, 114) . Dados oficiais norte- americanos revelam que a mortalidade atribuível a CI ronda os 15-35% nos adultos, e 10-15% em recém-nascidos e população pediátrica, embora os números sejam muito díspares consoante as populações estudadas e o desenho dos estudos (34, 52, 90). - 12 - Epidemiologia e etiologia das infecções fúngicas Em Portugal, os dados epidemiológicos são escassos. Porém, um estudo epidemiológico num hospital no Porto revelou uma incidência de 2 casos por cada 1000 admissões hospitalares, com mortalidade elevada, de 39,3% (32). 2.3 Papel das leveduras como agentes patogénicos em Cuidados Intensivos Dentro do género Candida, C. albicans é das espécies mais bem estudadas e frequentemente associada a infecção no humano, sendo bem conhecidas as características clínicas das infecções por esta levedura. Conduz frequentemente a infecções mucocutâneas, orofaríngeas ou vulvovaginais. Nas últimas décadas, outras espécies deste género tem vindo a ser estudadas e caracterizadas, devido à crescente incidência de infecções por Candida nãoalbicans (40). A CI encontra-se associada a sepsis severa, choque séptico e falência de múltiplos órgãos (86). Em doentes neutropénicos, a candidemia pode originar complicações graves como candidíase crónica disseminada, de tratamento difícil. Dados estatísticos revelam que doentes com CI podem necessitar de um prolongamento médio de ventilação mecânica invasiva, de 10 dias, e um prolongamento médio de 30 dias de internamento hospitalar (40, 52, 114, 141) . Diversos estudos indicam um risco superior de desenvolvimento de CI em doente com idades extremas, diabéticos, oncológicos (particularmente com neutropenia), do sexo masculino, sob administração de antibióticos de largo espectro, submetidos a procedimentos cirúrgicos extensos e a utilização de dispositivos médicos invasivos como cateter venoso central e cateter urinário (50, 114, 141). Cerca de 90% das infecções a Candida spp. são atribuíveis a C. albicans, C. glabrata, C. parapsilosis, C. tropicalis e C. krusei. A incidência de C. albicans tem vindo a diminuir, correspondendo a cerca de 50-70% consoante região geográfica, com a emergência de - 13 - Epidemiologia e etiologia das infecções fúngicas espécies não-albicans, como C. parapsilosis e C. glabrata. A taxa de incidência de cada espécie é variável consoante a região geográfica (93, 114, 118) . Estudos epidemiológicos em países como Estados Unidos, Reino Unido, França, Alemanha e Noruega, observaram que C. glabrata foi o segundo agente etiológico mais frequente de CI. Pelo contrário, em Espanha, Israel e América Latina, C. parapsilosis foi o segundo agente etiológico mais frequente de CI (57, 84, 85, 111, 114, 118, 153, 164) . Alguns dados apontam para taxas de mortalidade variáveis consoante a espécie de Candida spp. implicada, com taxas de mortalidade superiores em infecções por C. krusei e C. glabrata, e inferiores por C. parapsilosis (40, 114) . Um número alargado de estudos propôs-se identificar factores de risco associados a resultados clínicos negativos, ou mesmo fatais. Factores como idade e gravidade de co-morbilidades relacionaram-se com resultado clínico desfavorável, enquanto factores como ausência ou atraso na administração do tratamento antifúngico, e presença de cateteres foram preditivos independentes de morte por candidemia (40, 114). C. dubliniensis, descrita primeiramente em 1995 por Sullivan et al., apresenta características morfológicas e fisiológicas muito similares a C. albicans, pelo que a sua diferenciação em laboratório tem sido alvo de intensa pesquisa. As técnicas de biologia molecular parecem ser as mais fidedignas na correcta identificação e diferenciação desta espécie (53) . A utilização de meios de cultura específicos, menos onerosos na prática clínica, também têm sido debatidos, para diferenciação das duas espécies. Sugeriu-se inclusivamente que a utilização do meio CHROMagar Candida® poderia diferenciar, com elevadas especificidade e sensibilidade, as colónias (verde escuro C. dubliniensis e verde claro C. albicans) destas duas espécies, podendo esta característica ser utilizada como marcador fenotípico presuntivo de C. dubliniensis (91) . Outros testes fenotípicos têm sido averiguados, com bons resultados e os métodos automatizados ou semi-automatizados também têm sido alvo de escrutínio - 14 - (91, 146) .A Epidemiologia e etiologia das infecções fúngicas evolução destas tecnologias tem permitido um reconhecimento mais adequado desta espécie, com identificação correcta variável entre 60 a 98%, consoante o método utilizado. O VITEK® foi um dos métodos que demonstrou maior fiabilidade (126). Esta espécie encontra-se tradicionalmente associada a candidíase orofaríngea sobretudo em doentes VIH+. Todavia, têm-lhe sido atribuídos diversos casos, raros, de candidemia em doentes oncológicos e num doente pediátrico, e candidíase pulmonar num doente oncológico (68, 114, 146) . Paralelamente, outras espécies como C. guilliermondii, C. famata, C. kefyr, C. lusitaniae, C. norvegensis, C. inconspicua e C. rugosa, apresentaram taxas de isolamento superiores na última década, relacionando-se com formas invasivas de candidíase (40, 114, 116, 153). A distribuição das espécies pelo tipo de infecções que provocam varia consoante diversos factores, incluindo factores do hospedeiro. Diversos autores descreveram um risco aumentado de candidemia por C. glabrata em idades mais avançadas (114, 118) . Outros autores relacionaram a candidemia por C. glabrata com a utilização de antibióticos de largo espectro, utilização profiláctica ou empírica de fluconazol, utilização de cateter venoso central, administração de nutrição parentérica e internamento na UCI. A administração de piperacilina-tazobactam e vancomicina associou-se significativamente com candidemia causada por C. glabrata e C. krusei (114). A relação entre a utilização de terapêutica com fluconazol e emergência de infecção/colonização por C. glabrata tem sido escrutinada. Todavia, alguns estudos têm contestado a relação causal entre estes dois factores, demonstrando que existem outros factores do hospedeiro mais relevantes para o desenvolvimento de infecções por C. glabrata (40) . A observação de existência de taxas mais elevadas de colonização orofaríngea por C. glabrata em determinadas subpopulações, designadamente doentes oncológicos e em idades - 15 - Epidemiologia e etiologia das infecções fúngicas avançadas, permite concluir que a idade avançada e a gravidade de co-morbilidades possuam uma correlação superior com desenvolvimento de candidemia por C. glabrata (85, 114). C. parapsilosis coloniza a pele e apresenta grande facilidade de formação de biofilmes em cateteres ou outros dispositivos invasivos. Encontra-se, por isso, associada a infecções invasivas relacionadas com dispositivos médicos e soluções contaminadas (50, 114, 155) . Encontra-se igualmente associada a infecções invasivas em algumas subpopulações, como crianças e recém-nascidos (114, 155). C. tropicalis apresenta maior relevância em doentes hematoncológicos e doentes neutropénicos. Está documentado que, nestes doentes, a fungemia causada por C. tropicalis é responsável por um internamento mais prolongado em UCIs, quando comparado com doentes com fungemia causada por C. albicans, podendo causar miosite e mialgia severa (40, 114). Por seu turno, C. krusei é relevante em determinadas subpopulações, nomeadamente em doentes oncológicos. Apresenta resistência intrínseca ao fluconazol, pelo que se avaliou a ocorrência da emergência deste agente com a utilização generalizada do fármaco, mas a incidência global da infecção por este agente tem-se mantido estável ao longo dos anos (24%). É um dos agentes frequentes de endoftalmite fúngica e está associada a diarreia nos recém-nascidos (40, 50, 114). O isolamento de espécies mais raras de Candida spp. é cada vez mais frequente (153) . C. guilliermondii causa CI e encontra-se associada ao desenvolvimento de endocardite fúngica em doentes utilizadores de drogas intravenosas. Alguns autores encontraram relação entre o desenvolvimento de infecção invasiva por este agente e realização de cirurgia cardiovascular/abdominal prévia ou utilização de dispositivos médicos invasivos (35, 114) . Por outro lado, C. rugosa tem sido associada a candidemia na presença de cateteres intravenosos ou infecção fúngica invasiva em doentes queimados - 16 - (35, 50, 114) . Ambas as espécies são Epidemiologia e etiologia das infecções fúngicas particularmente comuns na América Latina, com incidências que rondam os 4-5% de todas as candidemias (114). C. lusitaniae, C. norvegensis, C. inconspicua e C. kefyr encontram-se também associadas a infecção disseminada (35). C. norvegensis e C. inconspicua são fenotipicamente semelhantes a C. krusei e existe evidência de serem intrinsecamente resistentes ao fluconazol. Existem casos reportados de infecção causada por C. inconspicua em doentes com VIH e doentes hematoncológicos. C. norvegensis foi isolada em amostras biológicas na Noruega, Japão e Holanda (114) . Outras espécies raras têm sido isoladas no ser humano, designadamente: C. zeylanoides, C. sake, C. valida, C. intermedia, C. humicola, C. pelliculosa, C. viswanathii, C. pulcherrima, C. lipolytica e C. haemulonii (35, 40, 116). Recentemente, a utilização de técnicas de biologia molecular permitiu, ainda, identificar novas espécies responsáveis por infecções no ser humano, fenotipicamente relacionadas com C. glabrata, C. parapsilosis e C. guilliermondii (35, 77, 78, 155) . Desdes os anos 90, que se reconhece C. parapsilosis como um ―complexo de 3 espécies‖: C. parapsilosis, C. orthopsilosis e C. metapsilosis. Apesar desta descoberta, ainda hoje em dia pouco é conhecido sobre as duas últimas espécies desta tríade. Diversos surtos de infecções causadas por C. othopsilosis foram reportados, mas a incidência de infecção em humanos, bem como o perfil de susceptibilidade a antifúngicos encontram-se mal esclarecidos. Estudos retrospectivos e prospectivos têm tentado esclarecer estas questões, identificando C. orthopsilosis e C. metapsilosis em menos de 10% dos isolados previamente classificados como C. parapsilosis. Estes estudos observaram ainda taxas de susceptibilidade elevadas aos azóis, Concentrações Inibitórias Mínimas (CIMs) relacionáveis com as descritas na literatura para C. parapsilosis, para a anfotericina B, assim como CIMs mais baixas relativamente a C. parapsilosis para as equinocandinas (77) . Adicionalmente, foi descoberta uma outra espécie que pode causar - 17 - Epidemiologia e etiologia das infecções fúngicas infecções invasivas, nomeadamente da corrente saguínea, Lodderomyces elongisporus, uma espécie capaz de formar ascosporos, mas fenotipicamente muito similar a C. parapsilosis. Por isso, esta espécie tem sido erroneamente identificada como C. parapsilosis e diversas infecções têm sido erradamente atribuídas a este último agente etiológico (78). De diversos pontos do globo, foram também reportadas infecções pelas espécies C. nivariensis e C. bracarensis, fenotipicamente relacionadas com C. glabrata. Estudos retrospectivos demonstraram a existência de casos raros de infecção causada por estas espécies, atribuídas erradamente a C. glabrata. Entre os casos reportados, encontrou-se resistência aos azóis e equinocandinas, mas os estudos são contraditórios. Lockhart et al. não conseguiram corroborar estes achados, observando, todavia, resistência de C. bracarensis e C. nivarensis à anfotericina B (80). Quanto a C. guilliermondii, encontra-se descrita uma espécie similar, C. fermentati, não existindo, contudo, informação clínica relevante acerca da mesma. Um estudo retrospectivo por Lockhart et al., verificou que uma pequena percentagem de isolados identificados como C. guillermondii correspondiam, na realidade, a isolados de C. fermentati. Quanto ao perfil de susceptibilidade destes isolados não se verificou CIMs elevadas contra equinocandinas, observando-se, contudo, CIMs mais elevadas para o fluconazol (79). 2.4 Factores de risco da infecção fúngica Inúmeros estudos têm tentado, através de análise multivariada, determinar factores de risco associados ao desenvolvimento de CI, de forma a compreender os mecanismos patofisiológicos envolvidos neste tipo de infecção. Entre as variáveis estudadas, têm sido identificados factores extrínsecos e intrínsecos ao hospedeiro, especificamente exposição prévia a antibióticos, tempo de internamento, presença de neutropenia, utilização de - 18 - Epidemiologia e etiologia das infecções fúngicas dispositivos médicos invasivos (cateteres como cateter urinário e cateter venoso central, procedimentos de hemodiálise, entre outros), procedimentos cirúrgicos, como cirurgia gastroabdominal extensa, entre outros (39, 40, 114, 141) . Contudo, a colonização por Candida spp. é um dos principais factores de risco implicado no desenvolvimento de infecção invasiva. Já na década de 1980, Solomkin et al. demonstraram a contaminação de Candida spp. a partir da cavidade abdominal para outros locais do organismo, antecedendo o desenvolvimento de candidemia (40) . Desde então, outros autores evidenciaram que o crescimento fúngico de Candida spp. na cavidade peritoneal é preditivo de infecção fúngica, assim como a presença de leveduras em grandes quantidades nas fezes de doentes oncológicos e recém-nascidos prematuros é um factor de risco para o desenvolvimento subsequente de candidemia. Adicionalmente, outros estudos comprovaram que a colonização de múltiplos locais do organismo por Candida spp. é um factor de risco independente para o desenvolvimento posterior de CI (40, 114, 141) . Alguns autores realizaram análises genotípicas, relacionando geneticamente espécies colonizadoras e responsáveis por infecção invasiva, em doentes neutropénicos e não neutropénicos. Apesar de estudos epidemiológicos demonstrarem que apenas 5 a 15% dos doentes hospitalizados se encontraram colonizados por Candida spp. à entrada, esta proporção facilmente aumenta com o aumento de tempo de internamento e exposição a factores de risco. Dos doentes internados em UCIs, cerca de 86% ficam colonizados por Candida spp., embora somente cerca de 5 a 30% destes doentes desenvolvam CI (40). Do exposto facilmente se compreende a dimensão multifactorial da infecção por Candida spp.. Não obstante, o agente patogénico possui alguns mecanismos que lhe permitem actuar e iludir as defesas do hospedeiro. Assim, a relação agente patogénico–hospedeiro será de extrema importância para a completa compreensão dos mecanismos patofisiológicos da CI. - 19 - Epidemiologia e etiologia das infecções fúngicas 2.5 Factores de virulência Candida spp. possui a capacidade de produzir factores de virulência que lhe permitem colonizar superfícies sintéticas (dispositivos médicos) ou mucocutâneas e, secundariamente, invadir os tecidos do hospedeiro, por disrupção da integridade da membrana celular. Os mecanismos de invasão do hospedeiro têm sido avaliados sobretudo em C. albicans e em populações neutropénicas, como doentes oncológicos sob quimioterapia. O mecanismo primordial para invasão por C. albicans envolve a colonização das superfícies mucocutâneas. Em doentes oncológicos, C. albicans coloniza frequentemente o tracto gastrointestinal, ocorrendo translocação dos fungos, no contexto de neutropenia, induzida por quimioterapia, devida a danos na mucosa intestinal. Os três mecanismos implicados na translocação fúngica são: disrupção do equilíbrio da flora intestinal com crescimento de microrganismos patogénicos; aumento da permeabilidade da barreira intestinal; deficiências no sistema imunitário do hospedeiro (70). O grau de relevância destes mecanismos para a invasão fúngica ainda permanece desconhecido. Porém, estudos em modelos animais evidenciaram a importância de factores como a manutenção do equilíbrio da flora intestinal e da integridade de mucosa em conjunto com a contagem total de neutrófilos na prevenção da invasão fúngica (70) . Estudos sobre outros intervenientes do sistema imunitário do hospedeiro como macrófagos, células dendríticas, monócitos e linfócitos não conseguiram estabelecer, na totalidade, as interacções entre C. albicans e hospedeiro. De facto, alguns estudos enfatizaram o papel da fagocitose na morte de C. albicans. Estudos em modelos animais tentaram ainda revelar as complicadas relações entre o sistema imunitário do hospedeiro, como a libertação de citocinas e actuação dos linfócitos T, e a prevenção da invasão fúngica. Os resultados sugerem que a resposta imunitária Th1, com libertação de interferão gama, IL-23 e IL-12, entre outras citocinas, pode - 20 - Epidemiologia e etiologia das infecções fúngicas estar implicada na prevenção de infecções sistémicas, enquanto uma resposta Th2 poderá a aumentar a susceptibilidade do hospedeiro a infecções invasivas ou sistémicas. A resposta celular inata parece ser muito relevante na prevenção de infecções de mucosas (162). Diversos tipos de alterações morfológicas indutíveis são conhecidos desde longa data no género Candida e em outros fungos, entre as quais o dimorfismo. Entende-se por dimorfismo a variação que consiste na passagem reversível, da fase leveduriforme para a fase filamentosa, dependente das condições de desenvolvimento. A fase filamentosa de C. albicans associa-se, geralmente, à formação do tubo germinativo, considerado a projecção inicial das hifas (69) .O estudo dos factores condicionantes do dimorfismo conduziu a resultados interessantes, revelando a associação da passagem à fase micelar com a temperatura, nutrientes do meio, nomeadamente proteínas, alterações de pH e concentrações de CO2 de 5,5% (69) . Estudos in vitro e em modelos animais revelaram que as formas com hifas ou pseudo hifas são relevantes na invasão do hospedeiro e que podem contribuir eficazmente para um aumento da capacidade de virulência do fungo. As alterações morfológicas de C. albicans encontram-se relacionadas com a expressão do gene UME6, regulador transcripcional da fase filamentosa. Adicionalmente, a pseudohifa parece funcionar como uma forma ―intermediária‖ entre a forma leveduriforme e a fase micelar (21). Dentro do género Candida, C. dubliniensis, espécie filogeneticamente relacionada com C. albicans, possui, também, a capacidade de formar hifas verdadeiras e tubo germinativo (53). Alguns autores formularam a hipótese que a formação de hifas permite, para além de um acréscimo na capacidade de invasão das células hospedeiras, a indução de citotoxicidade e/ou aumento da resistência à fagocitose (48) . Já se encontram descritos outros genes responsáveis - 21 - Epidemiologia e etiologia das infecções fúngicas pela formação de hifas ou pseudohifas em Candida spp. correlacionando-se com a capacidade de virulência das espécies (168). A actividade proteolítica de Candida tem sido implicada na invasão tecidular, encontrando-se já identificadas diversas famílias de genes, nomeadamente a família SAP (Secretory Aspartyl Proteinase), que contém 10 genes e que expressam proteinases. As saps tratam-se de antigénios imunodominantes que conferem virulência a Candida spp., degradando diversas proteínas humanas, como albumina, hemoglobina e imunoglobulina A. Esta actividade proteolítica, que pode ocorrer a diferentes pHs, foi relacionada com a capacidade de invasão tecidular de Candida albicans (14, 48, 69, 75) . Estudos in vitro demonstraram que os genes SAPs1-3 são expressos pelas formas leveduriformes, enquanto os genes SAPs 4-6 são expressos por fungos com hifas (14, 48). Outras espécies não-albicans como C. lusitaniae, C. dubliniensis, C. krusei, C. glabrata, C. tropicalis, C. parapsilosis e C. guilliermondii também expressam proteínases (168) . C. tropicalis possui os genes SAP1-4, enquanto C. dubliniensis apresenta 7 proteinases (com elevada homologia às proteinases de C. albicans) já identificadas (75, 168). Em C. albicans, a perda dos genes SAP2, 4 e 6 conduziu a uma redução importante do crescimento fúngico, adivinhando o papel fulcral da família de genes SAP no crescimento de Candida (48). Por outro lado, alguns autores demonstraram que a parede celular de Candida representa um papel importante na invasão tecidular e na virulência do fungo. Sabe-se que a parede celular de C. albicans não é constante do ponto de vista antigénico (23, 101) . As suas características antigénicas variam de acordo com inúmeras condições, em resposta ao meio onde se encontra. Conhecem-se já diversos antigénios/proteínas, como manano, enolase, proteinases e fosfolipases que podem estar implicados na invasão das células do hospedeiro (23) . A parede celular de C. albicans é composta por polissacáridos, proteínas e lípidos, que formam uma - 22 - Epidemiologia e etiologia das infecções fúngicas rede tridimensional. Esta engloba as glicoproteínas beta-D-glucano (beta-1,6-glucano e beta1,3-glucano), manano e quitina (23) . Os hidratos de carbono correspondem a 80% da composição da parede celular de C. albicans (101). O manano encontra-se correlacionado com invasão em doentes com candidemia (101) . A mananemia (antigenemia) plasmática é transitória e o manano pode formar imunocomplexos com diversos anticorpos em circulação (129). Sabe-se que este componente da parede celular de Candida exibe actividade imunossupressora, apresentando variabilidade (em termos de composição) entre espécies de Candida, estirpes virulentas e avirulentas e entre hifas ou formas leveduriformes. Alguns autores documentaram a relação de fragmentos polissacarídicos do manano e a capacidade de diminuição da resposta celular do hospedeiro, através da realização de estudos in vitro e em animais (101) . Estudos genéticos evidenciaram que a produção de manano é mediada por 3 genes: MNT1, PMT1 e PMT6 (168) . Todavia, encontra-se ainda mal compreendida a função deste componente celular, como factor de virulência de Candida. As proteínas da parede celular de Candida encontram-se ligadas covalentemente à matriz da parede celular e englobam dois tipos: ligadas ao beta-1,6-glucano através do radical glicofosfatidilinositol ou as ligadas directamente ao beta-1,3-glucano (proteínas Pir - proteins with internal repeats) (23) . A expressão destas proteínas pode diferir entre formas leveduriformes, micelares, ou consoante o meio circundante. Entre estas proteínas estão abrangidas adesinas, quitinases (envolvidas na biossíntese da parede celular), glucanosiltransferases e proteinases, entre outras. As funções e capacidade antigénica de diversas proteínas da matriz da parede celular têm sido estudadas e, em alguns casos, as suas actividades ligadas à filamentação e capacidade de invasão tecidular (23, 48, 168). - 23 - Epidemiologia e etiologia das infecções fúngicas Outras proteínas como a hwp-1 e als3 situam-se no tubo germinativo de C. albicans e são sintetizadas exclusivamente durante a produção de hifas (129) . Entre estas, a hwp-1 é uma adesina relevante na CI, tendo sido descrita como factor de virulência (69) . Paralelamente, a als3 pertence à família de genes ALS (Aglutinin Like Sequence), que codifica para glicoproteínas implicadas na adesão da levedura à superfície celular do hospedeiro. São já conhecidos oito genes desta família. A regulação destes genes é modificada por diversas condições, designadamente alterações do meio de crescimento, morfogénese ou fase do crescimento da levedura. A als3 demonstrou estar implicada na adesão endotelial ou epitelial da levedura. Por seu turno, a als1 e als5 ligam-se ao epitélio bucal e fibronectina, respectivamente (69). Outras espécies não-albicans como C. dubliniensis, C. glabrata, C. tropicalis e C. parapsilosis apresentam proteínas da parede celular com homologia com antigénios, como a hwp1, expressada na parede do tubo germinativo de C. albicans (129) . Em C. tropicalis e C. dubliniensis foram também identificadas 3 genes da família ALS, igualmente implicados na adesão às células do hospedeiro. Em C. glabrata, foi identificado o gene EPA-1, também responsável pela produção de uma adesina (168). Os mecanismos de adesão a ligandos do hospedeiro são complexos, mas verificou-se que estão envolvidos diversos ligandos, incluindo componentes da matriz extracelular, fibronectina, entactina, vitronectina, laminina e colagénio, e proteínas plasmáticas como fibrinogénio, plasminogénio e factores do complemento (23). De momento, alguns dos antigénios supracitados têm sido estudados para o desenvolvimento de métodos de diagnóstico imunológicos, mas as suas funções necessitam de esclarecimento futuro. - 24 - Epidemiologia e etiologia das infecções fúngicas Outros antigénios específicos do género Candida têm sido igualmente avaliados, com a finalidade de diagnóstico. Entre estes, um dos mais promissores é a enolase, antigénio com 48 kDa, expresso somente durante a invasão tecidular em doentes com candidemia e inexistente na fase de colonização por Candida spp. (73, 129, 169) . Trata-se de uma enzima glicolítica, presente no citoplasma celular e, em menor quantidade, na camada interior da parede celular (23, 73) de C. albicans e outras espécies do género Candida. papel da enolase como factor de virulência . Alguns autores documentaram o (23,169) . A enolase liga-se ao plasminogénio humano, substância convertida em plasmina pela acção de diversas enzimas, como a urocinase ou activador de plasminogénio. A plasmina apresenta funções relevantes na migração celular, degradação da matriz extracelular ou na fibrinólise. A ligação da enolase fúngica ao plasminogénio, mostrou, em estudos in vitro com C. albicans, encontrar-se associada à activação de plasminogénio, com consequente aumento da permeabilidade da barreira celular (67). Adicionalmente, foram reconhecidas enzimas secretadas com actividade hidrolítica, particularmente as fosfolipases. Estas enzimas correlacionam-se com um aumento de virulência e foram classificadas em diversas classes (A, B, C ou D) de acordo com o éster que clivam (168) . As fosfolipases B possuem grande relevância no género Candida, sendo expressas por outros géneros como C. neoformans e Aspergillus fumigatus (A. fumigatus) (69). Encontram-se identificados 5 genes de fosfolipases classe B, no entanto os genes PLB1 e PLB2 têm sido encontrados em estirpes de indivíduos infectados, correlacionando-se com infecção e invasão sistémica. Porém, o papel destas enzimas não se encontra totalmente determinado (23, 95). - 25 - Epidemiologia e etiologia das infecções fúngicas Para além de enzimas e antigénios de superfície, foram já identificados alterações de metabolismo e produtos secundários do metabolismo de Candida, que podem encontrar-se relacionados com a virulência e a proliferação de espécies deste género (13, 18) . O D-arabinitol é um metabolito que tem sido avaliado para utilização em diagnóstico (169). C. albicans produz quantidades mais elevadas deste metabolito (através da conversão de glucose neste poliol) em doentes com infecção invasiva, relativamente a doentes colonizados ou sem infecção. Contudo, os níveis plasmáticos de D-arabinitol encontram-se mais elevados em doentes com função renal diminuída, pelo que os dados são de difícil interpretação (169). Bernard et al. comprovaram que as espécies C. albicans, C. parapsilosis e C. tropicalis produzem este metabolito in vitro, ao contrário das espécies C. glabrata e C. krusei, sugerindo que pode ter funções fisiológicas importantes, como a manutenção/alteração da osmolaridade celular, permitindo o crescimento em diversos tipos de solutos (18) . Wong et al. evidenciaram a correlação entre os níveis plasmáticos no ser humano (tendo em consideração a excreção urinária do metabolito) e a infecção por C. albicans (169). O switch fenotípico é outro mecanismo que tem sido utilizado para explicar a capacidade de virulência de Candida. As alterações fenotípicas das colónias de C. albicans de lisas e brancas para colónias rugosas e acinzentadas (opacas), observado em 1987 por Soll et al., têm sido alvo de diversos estudos genotípicos (81) . Sabe-se que estas alterações fenotípicas se encontram implicadas no ciclo de vida do fungo e na sua capacidade replicativa. Neste tipo de alteração fenotípica encontra-se envolvida a expressão da proteína wor1. Diversos mecanismos transcripcionais foram implicados na expressão desta proteína (2, 81) . Interessantemente, alguns estudos revelaram que alterações fenotípicas podem correlacionarse com capacidade de infecção invasiva, tratando-se de uma forma de resposta ao stress - 26 - Epidemiologia e etiologia das infecções fúngicas ambiental a que o agente patogénico possa estar sujeito. Diversos autores colocaram a hipótese que o switch para colónias opacas possa beneficiar a capacidade de colonização das mucosas e de infecção de nichos específicos (2, 81) . Alguns estudos identificaram diferenças significativas entre os tipos de colónias (brancas ou opacas), no que concerne à sua relação com o hospedeiro ( 2, 81) . De facto, in vitro, as colónias opacas parecem conseguir fugir à fagocitose, por não secretarem substâncias quimioatractivas dos leucócitos. Por outro lado, as colónias brancas respondem a ferormonas de forma diferente das colónias opacas. Só as colónias opacas possuem a capacidade de reprodução sexuada. Contudo, em resposta à produção de ferormonas, as colónias brancas formam biofilmes, pressupondo-se que este facto pode facilitar o crescimento do fungo, por permitir a aproximação e o reconhecimento de células opacas, capazes de reprodução sexuada. As relações entre as defesas do hospedeiro e as diversas formas fenotípicas de C. albicans, continuam a ser alvo de escrutínio intenso (81). Outros tipos de alterações fenotípicas têm sido descritas noutras espécies de Candida, mas o seu papel como factor de virulência não se encontra totalmente estabelecido (49). A capacidade de ligação a superfícies inertes deve merecer a nossa atenção, com particular detaque na capacidade de formação de biofilmes. A existência de biofilmes é conhecida desde há longa data, mas só recentemente foi aceite, pela comunidade médica, como uma ―entidade‖ a recear. A formação de biofilmes encontra-se melhor estudada no caso de bactérias. O biofilme é definido como uma ―comunidade‖ microbiana (várias microcolónias da mesma espécie ou de diferentes espécies) caracterizada por células ligadas irreversívelmente a um substrato, a uma interface ou umas às outras (23, 36, 156) . Esta ―comunidade‖ encontra-se embebida numa matriz de substâncias extracelulares poliméricas (produzidas pelos microrganismos nela contidos) e apresenta um fenotipo alterado relativamente à taxa de - 27 - Epidemiologia e etiologia das infecções fúngicas crescimento e transcripção genética (23, 36) . As células em biofilmes possuem características diferentes das células em suspensão, no que respeita a comunicação intercelular ou quorum sensing (36, 156) . Os biofilmes são particularmente resistentes em meios onde ocorre rápida passagem de fluidos (logo a tensão de corte é elevada) e as microcolónias que fazem parte do biofilme deformam-se e oscilam à passagem do fluido. No entanto, poderão separar-se do biofilme, em situações de aumento excessivo da tensão de corte, com risco de causarem êmbolos infecciosos (36) . Os atributos do biofilme permitem-lhe também resistir a agentes antimicrobianos (desinfectantes, antibióticos e antissépticos) (36, 142, 156). A relação entre a formação de biofilmes e o aparecimento de infecção tem sido estudada exaustivamente. Evidências epidemiológicas demonstraram que a formação de biofilmes possui um papel no desenvolvimento de quadros infecciosos. Todavia, os mecanismos através dos quais o quadro infeccioso se desenvolve encontram-se mal compreendidos, sendo sugerido por alguns autores o destacamento de microrganismos, ou agregados de microrganismos, dos biofilmes em dispositivos invasivos, resultando em consequentes infecções do tracto urinário ou da corrente sanguínea; a resistência às defesas do sistema imunitário do hospedeiro; provisão de um nicho ecológico para desenvolvimento de microrganismos resistentes a agentes antimicrobianos (36). No género Candida, a formação de biofilmes é relevante em diversas situações clínicas, designadamente endocardite e colonização de dispositivos invasivos como cateter venoso central, cateteres urinários, próteses dentárias, válvulas cardíacas protésicas e outros dispositivos biomédicos (36, 160). A formação de biofilmes tem sido estudada sobretudo em C. albicans. Estudos in vitro demonstraram que as formas leveduriformes aderem primariamente a uma superfície e crescem numa camada basal. Esta camada pode conter algumas hifas ou pseudohifas e cadeias - 28 - Epidemiologia e etiologia das infecções fúngicas tubulares longas de células. À medida que o biofilme amadurece formam-se, na matriz extracelular, hidratos de carbono e proteínas (23, 102) . Entre os hidratos de carbono, o componente mais relevante é o beta-1,3-glucano. A matriz possui ainda outros componentes como fósforo, hexosamina e ácido urónico. Nobile et al. evidenciaram o papel da proteína reguladora zap1 na maturação da matriz extracelular de biofilmes de C. albicans, nomeadamente na biogénese de beta-1,3-glucano, um componente major desta matriz (102) . Outros autores constataram que os biofilmes possuem um aumento de resistência a agentes antimicrobianos que actuam neste componente (102, 156, 160). A maturação do biofilme é acompanhada da produção de álcoois, como produtos secundários, pensando-se que alguns possam estar implicados na sinalização celular e no quorum sensing (23, 102) . A produção de adesinas possui também um papel relevante na formação e maturação de biofilmes de Candida spp.(23, 160) . Fujibayashi et al. identificaram diversas espécies de Candida cuja adesão a superfícies foi comprometida pela actuação de imunoglubulina antiCandida albicans, revelando o papel fundamental da adesão para a colonização e formação de biofilmes por Candida spp. (51). A formação de biofilmes tem sido descrita em diversas espécies de Candida, entre as quais C. albicans, C. lusitaniae, C. krusei, C. parapsilosis, C. glabrata e C. pelliculosa. Hasan et al. utilizaram um modelo murino para compreender a capacidade infecciosa de estirpes com elevada taxa de formação de biofilmes. Neste estudo in vivo, os autores identifcaram as estirpes produtoras de biofilmes e observaram a disseminação de C. albicans por diversos orgãos, nos doentes com este tipo de estirpes. Paralelamente, verificaram existir correlação entre a produção de biofilmes e redução de taxa de crescimento. Estudaram, ainda, a variabilidade na formação/composição de biofilmes entre diversas estirpes de C. albicans, C. lusitaniae, C. krusei, C. parapsilosis, C. glabrata e C. pelliculosa. Os dados obtidos - 29 - Epidemiologia e etiologia das infecções fúngicas permitiram concluir que a formação de biofilme é variável consoante a espécie, e mais relevante nas espécies C. albicans, C. lusitaniae e C. krusei. Constataram, ainda, variabilidade elevada na formação de biofilmes entre estirpes diferentes de C. albicans e C. parapsilosis, explicável, possivelmente, por diferenças na composição de hifas, blastósporos e pseudohifas (60) . Estes resultados são discrepantes de dados apresentados por Tamura et al. (150) . Estes autores identificaram que espécies não-albicans apresentaram biofilmes com propriedades hidrofóbicas e aderência significativamente superiores a biofilmes prouzidos por C. albicans. Todavia, é de ressalvar que a variabilidade na composição de biofilmes e na capacidade de formação de biofilmes poderá estar também relacionada com o local do organismo do hospedeiro onde a espécie de Candida é isolada. Hasan et al. evidenciaram que estirpes de C. glabrata isoladas da urina possuíam elevada capacidade de formação de biofilmes, ao contrário de estirpes de C. glabrata isoladas de hemoculturas, com capacidade mais reduzida de formação de biofilmes (60). 2.6 Susceptibilidade in vitro aos antifúngicos Diversos estudos epidemiológicos, como o programa SENTRY e programa ARTEMIS, conseguiram elucidar a comunidade científica relativamente a perfis de susceptibilidade de Candida spp. contra diversos antifúngicos (93, 116). O fluconazol demonstrou ser activo contra grande parte das espécies de Candida, com taxas de susceptibilidades superiores a 90%. A utilização generalizada de fluconazol na prática clínica conduziu a alguma preocupação, no que respeita à emergência de resistência no género Candida, nomeadamente à emergência de resistência cruzada com outros antifúngicos triazólicos (itraconazol, voriconazol e posaconazol), mas que não tem sido fundamentada por estes programas de vigilância epidemiológica mundiais (93, 116). - 30 - Epidemiologia e etiologia das infecções fúngicas A nível mundial, observou-se um decréscimo de susceptibilidade (menor que 75%) nas espécies C. glabrata, C. guilliermondii, C. rugosa e C. norvegensis (116) . Esta última é fenotipicamente semelhante a C. krusei e existe evidência de ser intrinsecamente resistente ao fluconazol. Dados europeus revelam taxas de resistência de C. glabrata de cerca de 16%, embora os estudos de vigilância global apontem para valores mais elevados (114) . Quanto a C. krusei, apesar da resistência intrínseca ao fluconazol, diversos estudos de avaliação de susceptibilidade revelam diversos isolados com susceptibilidade dose- dependente, apontando (92 ,96, 116) para taxas de resistência (de acordo com as CIMs encontradas) entre 30% a 70% . Apesar de estudos isolados reportarem taxas de resistência significativas em isolados de C. albicans (32, 84) , estudos epidemiológicos à escala mundial não revelam estas tendências (31, 92, 93, 96, 116) . De acordo com o estudo ARTEMIS, a susceptibilidade das diversas espécies de Candida ao voriconazol foi superior quando comparado com a do fluconazol, com excepção de C. tropicalis, sendo de 98% para C. albicans, 82% para C. glabrata, 83% para C. krusei, 96,8% para C. parapsilosis e 91% para C. guilliermondii (116) . Quanto às espécies resistentes ao fluconazol, observou-se uma susceptibilidade superior a 75% nos casos de C. krusei, C. norvegensis e C. dubliniensis. A actividade do voriconazol foi mais reduzida contra espécies resistentes ao fluconazol de C.albicans, C. glabrata, C. tropicalis e C. rugosa (116) . Mallié et al. verificaram, num estudo multicêntrico francês com recurso ao Etest®, que espécies susceptíveis dose-dependentes ao fluconazol apresentaram uma taxa de susceptibilidade de 99% ao voriconazol, na concentração de 2µg/mL, enquanto que espécies resistentes ao fluconazol apresentaram uma taxa de susceptibilidade de cerca de 71%, ao voriconazol (84). Por outro lado, C. rugosa apresentou taxas elevadas de resistência, tanto ao fluconazol como ao voriconazol, podendo atingir os 60% e 38%, respectivamente (115, 116). Estas tendências são - 31 - Epidemiologia e etiologia das infecções fúngicas mais relevantes na América Latina e Ásia, porque na Europa e na América do Norte, a susceptibilidade desta espécie ao voriconazol permanece elevada, mesmo em isolados resistentes aos fluconazol (35, 115) . De qualquer forma, a susceptibilidade reduzida aos triazóis deste agente deve ser tida em consideração, quando é necessário escolher uma terapêutica antifúngica, adoptando-se, preferencialmente, uma política mais conservadora, até execução de teste de susceptibilidade do isolado (115). Por seu turno, a anfotericina B apresenta óptima actividade contra o género Candida sem evolução negativa temporalmente, observando-se, contudo, a emergência de algumas espécies resistentes. Existe ainda alguma dificuldade na avaliação da resistência à anfotericina B pelo método da micro diluição (preconizado pelo CLSI - Clinical Laboratory Standards Institute), com a dificuldade de definição de pontos de corte (26). O Etest® (AB Biodisk) parece conduzir a resultados mais fidedignos e correlacionados com o que ocorre in vivo (108). C. lusitaniae pode desenvolver rapidamente resistência secundária, por switch fenotípico, após exposição ao fármaco, mas CIM≥1µg/mL foi encontrada infrequentemente nesta espécie (35, 114, 148) . Normalmente, as estirpes resistentes à anfotericina B são bastante sensíveis a derivados triazólicos como o fluconazol (35) . Todavia, algumas estirpes resistentes podem apresentar desafios, por baixa susceptibilidade quer à anfotericina B quer a derivados triazólicos (148, 171). Encontra-se já documentada uma percentagem significativa de isolados de C. glabrata e C. krusei com susceptibilidade reduzida à anfotericina B (114) . No estudo SENTRY (1997-2000), Pfaller et al. identificaram 17,4% e 27,8% de isolados resistentes de C. glabrata e C. krusei, respectivamente. Estudos de cinética evidenciaram uma actividade fungicida mais lenta, em relação a estas espécies, quando comparadas com C. albicans. Tal indicia a necessidade de utilização de doses mais elevadas para tratamento de infecções - 32 - Epidemiologia e etiologia das infecções fúngicas causadas por aqueles dois agentes (114, 118) . Encontram-se também reportados casos de resistência primária ou secundária à anfotericina B em estirpes de C. tropicalis (148). A resistência em C. albicans é extremamente rara, embora Nolte et al. tenham descrito uma estirpe de C. albicans deficiente em ergosterol, isolada numa hemocultura de um doente com leucemia. Na África do Sul foram identificadas diversas estirpes de C. albicans resistentes à anfotericina B (16% dos isolados), reconhecendo-se um clade distinto de C. albicans nesta região. Na Europa e na América do Norte, somente uma pequena percentagem de isolados de C. albicans pertence a este clade (114, 148). Alguns autores observaram ainda casos raros de infecção invasiva por C. guilliermondii com falência clínica e resistência comprovada à anfotericina B. Isolados de C. parapsilosis resistentes a anfotericina B foram também detectados, embora raramente (35, 148) . O desenvolvimento de infecção invasiva causada por C. rugosa em doentes sob terapêutica com anfotericina B sugere também reduzida susceptibilidade desta levedura ao fármaco (115) . Quanto a outras espécies raras, alguns isolados de C. kefyr, C. lipolytica, C. orthopsilosis e C. metapsilosis demonstraram CIMs elevadas contra a anfotericina B (35 ,77) . No entanto, este fármaco revelou a melhor actividade, in vitro e in vivo, contra C. pelliculosa, comparativamente a outros agentes antifúngicos (35). C. famata também apresenta alguns desafios para os clínicos, pois estudos de susceptibilidade in vitro revelaram CIMs mais elevadas para equinocandinas, à semelhança de C. guilliermondii e C. parapsilosis, bem como susceptibilidade reduzida ao fluconazol. Encontram-se reportados na literatura internacional casos de falência clínica à anfotericina B lipossómica e falência clínica ao fluconazol. Foi evidenciada resistência in vitro à anfotericina B (35). - 33 - Epidemiologia e etiologia das infecções fúngicas O grupo das equinocandinas apresenta uma óptima actividade fungicida contra todas as espécies de Candida, (CIMs≤2µg/mL em 98,8%) mesmo em espécies resistentes aos triazóis (119) . Moudgal et al. e Vazquez et al. detectaram isolados de C. parapsilosis que exibiam fenótipo de multirresistência às classes de triazóis e equinocandinas (114) . Diversos autores identificaram CIMs ligeiramente mais elevadas (actividades similares mas variações de uma a duas diluições entre os fármacos desta classe) em todas as espécies de Candida para a caspofungina e micafungina, comparativamente à anidulafungina (133) . As espécies C. guilliermondii e C. parapsilosis poderão apresentar CIMs mais elevadas a todos os antifúngicos desta classe (114). Também C. famata e C. lusitaniae apresentaram valores modais de CIMs entre 0,25 e 0,5µg/mL e 0,12 a 0,5µg/mL, respectivamente (119). Alguns estudos reportaram isolados com CIMs muito elevadas, atingindo valores superiores a 8µg/ml (11, 119, 133). Verificaram-se alguns casos de C. glabrata, C. albicans e C. tropicalis com CIMs elevadas às equinocandinas, comparativamente ao esperado globalmente (valor modal). Nas espécies C. glabrata, C. albicans e C. parapsilosis estão ainda descritos diversos casos de resistência clínica à caspofungina, com terapêuticas prolongadas, encontrando-se já documentado um surto de CI por C. parapsilosis resistente à caspofungina, numa unidade de Queimados nos Estados Unidos (11, 114) . A utilização de terapêutica prolongada com caspofungina para tratamento de candidíase orofaríngea por C. albicans em doentes com SIDA tem resultado, igualmente, em casos raros de falência clínica e desenvolvimento de resistência a este antifúngico (11, 114). Hakki et al. descreveram um caso de falência terapêutica à caspofungina num doente oncológico (leucemia mieloblástica aguda) com candidemia por C. krusei e posterior desenvolvimento de endoftalmite (11) . Igualmente, Pelletier et al. descreveram outro caso de falência terapêutica à caspofungina, num doente com CI por C. krusei (133) . No caso da anidulafungina, o estudo multicêntrico publicado por Reboli et al. - 34 - Epidemiologia e etiologia das infecções fúngicas (dados originados em ensaios clínicos) evidenciaram a persistência de CI em oito dos doentes tratados com anidulafungina (134). 2.7 Mecanismos de resistência aos azóis Os fármacos pertencentes a esta classe terapêutica possuem actividade contra o citocromo P450 fúngico, através da inibição da enzima lanosterol 14-α-desmetilase. Esta enzima permite a conversão do lanosterol em ergosterol, e a sua inibição interrompe a síntese membranar na célula fúngica. Estes agentes influem ainda num passo posterior da síntese do ergosterol, através da inibição da ∆20dessaturação (33, 138) . A cascata de biossíntese de ergosterol poderá variar entre espécies de Candida spp.. Na passada década, foram descobertos diversos mecanismos de resistência aos azóis em leveduras do género Candida, com particular enfoque no fluconazol: (1) alteração na enzima lanosterol desmetilase; (2) alterações na esterol ∆5,6desaturase; (3) produção de bombas de efluxo de antifúngicos energia-dependentes (49). (1) A enzima lanosterol 14-α-desmetilase é codificada pelo gene ERG11 e mutações pontuais neste gene estão na origem de alterações da enzima alvo, com decréscimo de afinidade dos derivados azólicos para esta. A expressão excessiva de ERG11 resulta na produção em massa de enzima, criando, por outro lado, necessidade de uma concentração superior de fluconazol a nível intracelular. Obviamente, este evento conduz a uma diminuição da inibição da enzima lanosterol 14-α-desmetilase. Por outro lado, a perda de um alelo, mutações pontuais, recombinação mitótica, entre outros mecanismos, neste gene, podem conduzir ao desenvolvimento de uma estirpe resistente, homozigótica para o gene mutado ( 33, 138, 158). - 35 - Epidemiologia e etiologia das infecções fúngicas (2) A deficiência na enzima ∆5,6desaturase trata-se de outro mecanismo descrito para resistência aos derivados azólicos. A deficiência nesta enzima permite a produção de 14metilfecosterol, com manutenção de viabilidade mesmo com inibição da enzima lanosterol 14-α-desmetilase. Esta enzima é codificada pelo gene ERG3. Em C. glabrata, não se evidenciou resistência em isolados com deficiência em ∆5,6desaturase (49) . Outros autores também não evidenciaram sobre-expressão do gene ERG11 sob pressão farmacológica (161) . Em C. albicans, mutantes homozigóticos para ERG3 evidenciaram resistência ao fluconazol (138) . (3) A produção de bombas de efluxo activas pode ser conseguida através de dois mecanismos: regulação dos genes MDR, com expressão de transportadores de efluxo – bombas MFS (Major Facilitators Superfamily), resultando em CIMs elevadas para o fluconazol; regulação dos genes CDR, resultando em resistência secundária cruzada aos derivados triazólicos (33, 62). O factor de transcrição PDR (pleiotropic drug resistance) parece também ser relevante neste último tipo de mecanismo de resistência, contribuindo para a expressão dos genes CDR (158). Os genes MDR codificam para bombas que actuam por um mecanismo dependente de um potencial de membrana. Os genes CDR codificam para o transporte de ATP. Este mecanismo é bem conhecido em C. glabrata - consegue criar bombas de efluxo dependentes de ATP (ATP-binding cassette efflux pumps), para a expulsão de antifúngicos, através da expressão dos genes CDR1 e CDR2 (33, 62, 158) . Outros genes têm vindo a ser envolvidos neste mecanismo de resistência, como o SNQ2, dependendo da espécie implicada (33, 62, 161) . Como tal, a exposição de C. glabrata a concentrações sub-terapêuticas de fluconazol pode resultar em resistência cruzada entre os diversos derivados triazólicos, conforme notado por diversos autores (33, 62). Em C. glabrata, encontram-se ainda descritos outros mecanismos de resistência - 36 - Epidemiologia e etiologia das infecções fúngicas aos derivados triazólicos, que não parecem ser relevantes noutras espécies de Candida. Investigadores evidenciaram, recentemente, o papel de mutantes petite, que sofrem alterações fenotípicas substanciais, na resistência desta espécie aos derivados triazólicos. Estes mutantes produzem colónias pequenas em meios fermentativos, mas não são capazes de crescer em meios não fermentativos. Estes defeitos devem-se a perdas no ADN mitocondrial ou a mutações no genoma nuclear que afectam a actividade mitocondrial ou actividade respiratória das leveduras. Trata-se de um fenótipo particular, pensando-se que o decréscimo da actividade respiratória possa estar implicada na resistência aos derivados triazólicos. O estudo destes mutantes tem resultado em descobertas inesperadas. Vandeputte et al. evidenciaram a existência de um mutante petite hipersensível ao fluconazol. Verificou-se que esta estirpe sobrexpressava os genes PDR1 e CDR1, implicados na resistência aos azóis, apesar da sua hipersusceptibilidade, conduzindo, por isso, a dados contraditórios (158) . Assim, os mecanismos subjacentes quer à resistência ou à hipersusceptibilidade destes mutantes merecem estudos futuros. Para além disso, Bard et al. reportaram seis isolados de C. glabrata que utilizam o colesterol do hospedeiro em substituição do ergosterol fúngico (12) . Enquanto C. krusei e C. albicans possuem diversos esterois intermediários em comum, C. glabrata só possui lanosterol como único esterol intermediário na biossíntese do ergosterol (71, 104) . Foi possível identificar deficiências nos genes ERG1 e ERG7, responsáveis pela produção de enzimas inactivas e consequente deficiência na cascata de biossíntese do ergosterol (12) . As implicações deste achado não se encontram totalmente estabelecidas. A resistência de C. krusei ao fluconazol aparenta ser mediada pela diminuição da afinidade da enzima alvo, lanosterol 14-α-desmetilase, ao agente antifúngico, de acordo com estudos conduzidos por Orozco et al. (104) . Para além disso, o citocromo P-450 de C. krusei apresenta - 37 - Epidemiologia e etiologia das infecções fúngicas particularidades relativamente ao citocromo P- 450 de C. albicans. Assim, a enzima alvo do fluconazol é diferente em termos qualitativos nas duas espécies. Outros investigadores observaram outros mecanismos de resistência, como diminuição da acumulação de fluconazol na célula fúngica e alterações da ∆5,6dessaturase. Estudos com itraconazol revelaram a existência de estirpes de C. krusei resistentes, por diminuição da acumulação de fármaco na célula fúngica, em oposição a estirpes sensíveis (104) . Não se pode excluir que estirpes de C. krusei com CIMs mais baixas ao fluconazol (dose-dependente, por exemplo) possam utilizar estes mecanismos para diminuirem a susceptibilidade ao fluconazol (96) . De qualquer forma, no género Candida, estes mecanismos podem actuar individual, sequencial ou concertadamente. Outros investigadores têm perseguido hipóteses diferentes, considerando outros possíveis mecanismos de resistência, entre os quais a produção excessiva de saps por C. albicans, conforme documentado num doente com VIH exposto a doses sub-terapêuticas de fluconazol, ou a alterações da expressão de proteínas associadas ao glucano (Glucan Associated Proteins – GAP) como enolase, fosfogliceromutases ou β-(1,3)-glucanases, e da incorporação destes componentes na parede fúngica em estirpes de C. albicans resistentes ao fluconazol (6) . Estes achados sugerem que a exposição de C. albicans ao fluconazol pode alterar a composição e utilização de proteínas a nível da parede fúngica, permitindo o escape desta levedura ao antifúngico e até ao hospedeiro, reflectindo, talvez, alterações metabólicas inespecíficas, independentemente do factor de stress envolvido. Todavia, estes dados carecem de estudos mais elucidativos (6). A interacção de biofilmes com agentes antimicrobianos tem igualmente gerado estudos elucidativos na compreensão de mecanismos de resistência a antifúngicos. - 38 - Epidemiologia e etiologia das infecções fúngicas Os mecanismos de resistência propostos e estudados pela comunidade científica englobam a penetração diminuída do agente antimicrobiano na matriz do biofilme; a alteração da taxa de crescimento de microrganismos formadores do biofilme; o aumento da densidade e/ou envelhecimento celular afectando a resistência a fármacos; ou outras formas de alterações fisiológicas que condicionem o modo de crescimento do biofilme (36). Uma das dificuldades na avaliação de susceptibilidade de biofilmes a antifúngicos é a inexistência de protocolos padronizados (64, 160) . Alguns autores referem, contudo, que fármacos fungicidas (ex: anfotericina B, caspofungina e anidulafungina) possuem maior actividade contra biofilmes, observando-se CIMs mais baixas do que para derivados triazólicos,(64, 156) em que as CIMs contra biofilmes podem ser até 20000x superiores às CIMs observadas em inóculos de células em suspensão (160) . Não obstante, a actividade destes agentes contra biofilmes permanece reduzida. Os fármacos apresentam CIMs consideradas resistentes, e a sua acção de inibição da actividade metabólica dos biofilmes vai decrescendo com a maturação/envelhecimento do biofilme (64, 156). Uma das hipóteses colocadas é a de que ocorre a ligação de antifúngico aos beta-glucanos da matriz extracelular do biofilme. Nett et al. demonstraram, em trabalhos anteriores, a ligação do fluconazol aos beta-glucanos. Desta forma, o biofilme ―sequestra‖ o fármaco (como fluconazol e anfotericina B) evitando a sua actuação sobre as células fúngicas (160). Vediyappan et al. introduziram recentemente novos dados, relativos a mecanismos de resistência de biofilmes a antifúngicos, através de estudos in vitro. Estes autores observaram que a exposição ao fluconazol não condicionou a regulação significativa de genes (160). Nailis et al. estudaram a actividade do fluconazol e anfotericina B sobre biofilmes de C. albicans. Para tal, utilizou concentrações muito elevadas de fluconazol, observando a sua actividade contra biofilmes maduros. Os biofilmes maduros demonstraram tolerância a altas - 39 - Epidemiologia e etiologia das infecções fúngicas concentrações de fluconazol. Pelo contrário, biofilmes jovens demonstraram redução em número de células, quando expostos a doses elevadas de fluconazol. Estes autores estudaram igualmente a expressão de genes, obtendo resultados similares aos de Vediyappan et al., em relação ao fluconazol (99). Muito permanece ainda desconhecido sobre mecanismos de resistência de biofilmes a antifúngicos. Este desconhecimento condiciona também o tratamento de infecções em que a formação de biofilmes possui um papel fulcral. 2.8 Mecanismos de resistência às equinocandinas As equinocandinas possuem como molécula-alvo a β-1,3-glucano sintetase. A inibição desta enzima permite a disrupção da parede celular do fungo, com consequente instabilidade osmótica e lise celular (33) . A β-1,3-glucano sintetase trata-se de um complexo enzimático constituído por, pelo menos, duas subunidades: fksp e rho1p. A primeira é codificada por diversos genes: FKS1, FKS2 e FKS3 (55). Têm sido reportados casos de falência clínica à caspofungina, com identificação de diversos mecanismos de resistência. Encontram-se descritas mutações em diferentes regiões do gene FKS1, na espécie C. albicans. As mutações mais relevantes foram encontradas em regiões delimitadas do gene, nomeadamente nas posições 645 e 641 (11) . Foram também encontradas mutações no gene FKS2. Mutações nestes genes foram igualmente reportadas em estirpes de C. glabrata, C. krusei e C. tropicalis (9, 11, 54). Nas espécies C. guilliermondii e C. parapsilosis, a susceptibilidade reduzida parece estar relacionada com polimorfismos naturais na região fks1p, à semelhança do que ocorre noutras espécies por resistência secundária (98). Estudos de cinética comprovaram que estas mutações conduziram a diminuição da afinidade do complexo enzimático β-1,3-glucano sintetase para a caspofungina - 40 - (55) . Observou-se também Epidemiologia e etiologia das infecções fúngicas que as mutações supracitadas condicionam o aparecimento fenotípico de CIMs mais elevadas, quando comparadas com outras mutações descritas (54) . Por outro lado, as mutações nas posições 641 e 645 conferiram uma capacidade catalítica inferior da β-1,3-glucano sintetase (55) . Interessantemente, os isolados com estas mutações exibiam CIMs elevadas para todos os antifúngicos desta classe, e os estudos de cinética conduzidos evidenciaram resistência cruzada dentro do grupo terapêutico das equinocandinas. Estudos mais aprofundados de Garcia-Effron et al. revelaram diferenças subtis na diminuição de afinidade da subunidade fks1p para os diferentes antifúngicos desta classe, consoante as mutações encontradas (55). Baixench et al. demonstraram, por testes de genotipagem de estirpes de C. albicans, que a utilização prolongada de caspofungina contribuiu para a alteração de marcadores polimórficos (CDC3, EF3 e HIS3) destas. Assim, reportaram a evolução até à homozigotia para o marcador CDC3. A perda de heterozigotia, sob pressão farmacológica, pode ter implicações clínicas relevantes e persistir no tempo. Este tipo de mecanismo também tem sido implicado na resistência de C. albicans ao fluconazol. De qualquer forma, as implicações deste mecanismo na resistência a antifúngicos não se encontram clarificadas (11). Outros autores implicaram a activação de mecanismos compensatórios, como a síntese aumentada de quitina para evitar ou diminuir a disrupção da parede celular, na resistência à caspofungina in vitro (98). Estudos de mecanismos de resistência de biofilmes a esta classe de fármacos têm também sido prolíficos em resultados. Vediyappan et al. verificaram que a exposição de biofilmes de C. albicans à caspofungina conduziu à sobre-expressão, entre outros, dos genes HWP1, ALS3, ECE1, intimamente relacionados com a formação de hifas e biofilmes, sugerindo tratar-se de um mecanismo de defesa contra os efeitos deletérios da caspofungina (160). - 41 - Epidemiologia e etiologia das infecções fúngicas 2.9 Mecanismos de resistência aos polienos Sabe-se que o agente pertencente a esta classe – anfotericina B - actua na membrana celular do fungo através da ligação ao ergosterol, considerando-se diversos mecanismos que permitem a sua acção fungicida. Por um lado, a anfotericina B pode ligar-se a enzimas membranares como a H+ ATPase, com consequente diminuição das reservas energéticas do fungo. Infelizmente, pode também ligar-se à Na+/K+ ATPase de células mamíferas, justificando, em parte, a sua toxicidade no ser humano. Por outro lado, pode conduzir a peroxidação lipídica, com aumento da fragilidade/permeabilidade da membrana celular fúngica, ou a extravasamento de Ca2+. Estes mecanismos podem explicar parcialmente a toxicidade da anfotericina B no ser humano, mas não são suficientes para explicar a gravidade de alguns sintomas reportados (19) . Estudos in vitro permitiram observar a ligação da anfotericina B a receptores das LDL e às próprias LDL, factos comprovados por posteriores estudos em modelos animais. Estes dados foram muito relevantes na construção de formulações de anfotericina B mais inócuas, resultando em diversos tipos de formulações lipídicas (1, 19, 38). Para além disso, diversos autores constataram relações dose-dependentes na toxicidade do composto, concluindo-se que doses elevadas de anfotericina B podem actuar na célula mamífera e danificar o colesterol (1, 19) . Os compostos lipídicos permitem, contudo, utilização de doses mais elevadas e tornam-se mais selectivos para os lípidos fúngicos (1, 38). Toda a investigação nesta área permitiu a criação de diversos modelos explicativos e mais elucidativos do mecanismo de acção da anfotericina B: (A) Interacção com esteróis da membrana; (B) formação de um canal transmembranar (aquoso ou não aquoso, consoante a concentração de antifúngico, e tempo dependente); (C) destruição da integridade da membrana por inserção de monómeros ou outras formas de agregados de anfotericina B na - 42 - Epidemiologia e etiologia das infecções fúngicas bicamada membranar. Todos estes mecanismos resultam num aumento de permeabilidade a catiões monovalentes e extravasamento de catiões através de canais transmembranares. Nem todos estes modelos explicativos foram demonstrados experimentalmente (19). A transferência da substância activa, anfotericina B, das membranas lipídicas para a membrana da célula fúngica também tem sido amplamente estudada. Shimizu et al. identificaram diversos factores implicados neste processo. Constataram que o lipossoma se liga à parede celular da célula fúngica de Saccharomyces cerevisiae de forma temperatura dependente, pois observaram que o fármaco foi transferido para a membrana da célula fúngica e demonstrou actividade antifúngica a 35ºC, mas que a transferência para a membrana da célula fúngica e a actividade fungicida foram reduzidas, a 4ºC. Para além disso, estes autores observaram não ser necessário a ocorrência de disrupção do lipossoma para a transferência de anfotericina B para a célula fúngica. Adicionalmente, o fármaco apresentou uma afinidade superior para o ergosterol em relação ao constituinte lipídico do lipossoma, logo, a composição em lípidos da membrana celular fúngica parece ter relevância na transferência do fármaco do lipossoma (constituinte - colesterol), para a membrana celular fúngica (constituinte- ergosterol). Também a fluidez da membrana lipossómica parece ter relevância na transferência de anfotericina B para a membrana fúngica. Assim, quanto maior a fluidez da membrana lipossómica, maior a libertação da anfotericina B. O lipossoma permite, ainda, a estabilização da substância activa e consequente retenção de actividade biológica. Todos estes factores podem contribuir para a eficácia in vivo (144). Da mesma forma, os mecanismos de resistência do género Candida à anfotericina B carecem de elucidação. Alguns autores observaram resistência à anfotericina B em estirpes deficientes em ergosterol (138) . Liu et al. colocaram a hipótese que tal actividade se possa dever à sobre- expressão dos genes SOD2, YHB1, AOX1, AOX2, CTA1 e GSH1 que codificam para - 43 - Epidemiologia e etiologia das infecções fúngicas proteínas, que respondem ao stress oxidativo causado pela anfotericina B. Estes autores verificaram, ainda, a sub-regulação de genes responsáveis pela biossíntese de ergosterol, como ERG3 e ERG11, inferindo que a célula fúngica poderia utilizar outros esteróis como resposta à pressão farmacológica imposta pela anfotericina B. Também observaram a expressão de genes envolvidos na regulação do metabolismo do ferro e metabolismo dos ácidos gordos como resposta à pressão farmacológica exercida quer pela anfotericina B quer pela caspofungina (76) . As implicações destas observações não se encontram esclarecidas. Sanglard et al. evidenciaram resistência à anfotericina B em mutantes homozigóticos ERG11, com deficiência na enzima lanosterol 14-α-desmetilase. Este mecanismo de resistência permite a evasão do fungo à actividade de outro tipo de antifúngicos, como os derivados triazólicos (138) . A comunidade científica concorda que a resistência a polienos se encontra frequentemente, mas não sistematicamente, associada à resistência a derivados triazólicos, pelo que o mecanismo supracitado, aliado a diminuição/inexistência de ergosterol e substituição por outros produtos intermediários parecem ser de grande relevância na resistência aos polienos (33, 62, 138). Estirpes de C. lusitaniae resistentes à anfotericina B foram estudadas por Young et al., identificando-se dois genes que poderão estar implicados na resistência desta espécie a este polieno: ERG3, que codifica para a enzima ∆5,6desaturase, e ERG6, que codifica para a C24metiltransferase. No entanto, os resultados obtidos são contraditórios (171) . Vandeputte et al. evidenciaram mais uma vez o papel do gene ERG6 na resistência do género Candida à anfotericina B, mais concretamente de C. glabrata. Estes autores observaram a produção de pseudohifas por estirpes de C. glabrata resistentes à anfotericina B, sugerindo maior capacidade de virulência destas estirpes. Evidenciaram que uma única mutação no gene ERG6 foi responsável por uma série de alterações fenotípicas que conduziram a resistência à - 44 - Epidemiologia e etiologia das infecções fúngicas anfotericina B, com aumento da susceptibilidade aos azóis, sugerindo-se que a mutação neste gene diminuiu a actividade de efluxo da parede celular fúngica e aumentou a permeablidade ao azol (159). No que toca a mecanismos de resistência por biofilmes, Vediyappan et al. demonstraram que a exposição de biofilmes de C. albicans à anfotericina B conduziu à sobre-expressão de inúmeros genes, quando comparado com células em suspensão, nomeadamente relacionados com a biossíntese de aminoácidos. Para além disso, verificaram que a anfotericina B se ligou ao biofilme maduro, designadamente aos beta-glucanos(160). Nailis et al. constataram que a anfotericina B revelou actividade contra biofilmes maduros em concentrações na ordem dos 32mg/L, em concordância com dados apresentados em estudos prévios. Contudo, observou-se a sobrevivência/persistência de um pequeno número de células. Estes autores observaram, adicionalmente, diminuição da expressão dos genes CDR1 e MDR1 em biofilmes maduros, relativamente a biofilmes jovens, quando em contacto com anfotericina B (99). - 45 - Tatamento de infecções fúngicas invasivas 3. Tratamento de infecções fúngicas invasivas A terapêutica apropriada em infecções fúngicas invasivas é um factor preditivo de bom prognóstico das mesmas (58) . As evidências reunidas ao longo de diversos anos de estudo do género Candida permitiram, a painéis de peritos, a elaboração de recomendações relativas ao melhor tratamento antifúngico em infecções fúngicas em Cuidados Intensivos (107) . A terapêutica pode ser classificada em 4 géneros: profiláctica, pré-emptiva, empírica ou curativa/documentada. A terapêutica profiláctica cobre todas as situações em que o doente não se encontra infectado. Na terapêutica pré-emptiva, o tratamento é iniciado com base na avaliação de factores de risco do hospedeiro e marcadores de infecção. Como tal, o doente encontra-se em risco elevado de desenvolver uma infecção fúngica e o tratamento antifúngico é iniciado. A terapêutica empírica justifica-se em doentes com sinais clínicos de infecção mas em que o clínico não consegue isolar a fonte de infecção. Na terapêutica curativa/documentada, existe evidência microbiológica aliada a evidência clínica de infecção fúngica invasiva. O microrganismo causador de infecção encontra-se devidamente identificado (58). A terapêutica profiláctica é utilizada em diversos tipos de doentes, nomeadamente doentes neutropénicos com patologias hematoncológicas, após transplante de medula óssea, e receptores de orgãos sólidos, como fígado e pâncreas, evidenciando resultados favoráveis (83, 107, 114) . No contexto de Cuidados Intensivos, alguns estudos apontam para benefícios claros em doentes submetidos a cirurgia abdominal extensa, com consequente prevenção de colonização e invasão intra-abdominal por Candida spp. (107, 163) . No entanto, a comunidade científica permanece dividida, pois os diversos estudos realizados utilizam diferentes desenhos, diferentes populações e diferentes antifúngicos (doses e formulações). Pfaller et al. referem benefícios claros em sub-populações seleccionadas de doentes. Estes autores - 46 - Tatamento de infecções fúngicas invasivas advertem, todavia, para a possibilidade de emergência de estirpes resistentes ao fluconazol e de espécies não-albicans em doentes submetidos a profilaxia. Os resultados na mortalidade dos doentes carecem também de esclarecimentos futuros (58). As terapêuticas empírica e pré-emptiva não possuem dados substanciados por ensaios aleatorizados, pelo que a utilização deste tipo de estratégias permanece controverso (58, 163). A utilização de terapêutica empírica possui, contudo, força de evidência superior em doentes neutropénicos, dado que a utilização de uma formulação lipídica de anfotericina B ou de caspofungina se encontra recomendada pela IDSA (Infectious Diseases Society of America) com força de recomendação A-I (107). Quanto à terapêutica documentada, os fármacos de primeira linha no tratamento de infecções fúngicas invasivas devem ser seleccionados de acordo com a gravidade da situação clínica do doente. Dados retrospectivos apontam para diferenças significativas entre doentes hemodinamicamente estáveis ou instáveis (sepsis, choque séptico, sepsis grave ou síndrome de resposta inflamatória sistémica), pelo que a utilização de antifúngicos de espectro mais alargado no último grupo de doentes pode ser benéfica. Para além disso, é importante focar as diferenças de perfil de farmacocinética/farmacodinâmica dos doentes em Cuidados Intensivos, com variações importantes do volume de distribuição e excreção renal de muitos fármacos (58). Assim sendo, Spellberg et al. propuseram um algoritmo para o tratamento de CI com base no estado hemodinâmico do doente. Em doentes hemodinamicamente estáveis, sem disfunção de orgãos, o fluconazol pode ser considerado como terapêutica de primeira linha. A decisão é baseada no perfil de segurança favorável do fármaco, nos estudos de susceptibilidade mundiais realizados por diversos autores, com demonstração de actividade importante contra Candida spp., e no baixo custo deste fármaco (58). A IDSA também recomenda a utilização de - 47 - Tatamento de infecções fúngicas invasivas fluconazol em doentes não neutropénicos com candidemia (força de recomendação A-I). No entanto, a utilização deste fármaco deve ser preterida se o doente tiver sido exposto recentemente a um azol, se se encontrar previamente colonizado por espécies resistentes ao fluconazol, ou se o doente estiver numa área geográfica em que a probabilidade de infecção por espécies resistentes/com susceptibilidade reduzida aos azóis for elevada (C. krusei/C.glabrata) (58, 90, 107) . Pelo contrário, em doentes hemodinamicamente instáveis, a utilização de equinocandinas parece ser a alternativa mais adequada, conforme Reboli et al. e IDSA (força de recomendação A-III) (58, 107, 134) . A utilização de uma formulação lipídica de anfotericina B pode ser considerada em alternativa (força de recomendação da IDSA A-I). A anfotericina B convencional apresenta problemas de segurança e toxicidade, não sendo recomendada em doentes críticos (90) . A transição de equinocandina para fluconazol é recomendada pela IDSA em candidemias em doentes não neutropénicos, clinicamente estáveis ou com infecção por estirpes que provavelmente apresentam susceptibilidade ao fluconazol (por exemplo C. albicans) (107) . A utilização de voriconazol pode ser considerada em estirpes de C. krusei ou C. glabrata susceptível ao voriconazol (recomendação B-III pela IDSA) (107) . Em infecções por C. parapsilosis, o tratamento inicial com fluconazol é recomendado pela IDSA, com uma força de recomendação B-III (58, 107) . A terapêutica deve ser mantida durante 14 dias, indicando-se também a remoção de dispositivos invasivos como cateteres intravenosos (107). A utilização de fluconazol como terapêutica padrão é co-substanciada por diversos estudos clínicos bem desenhados. Por seu turno, a utilização de equinocandinas em doentes hemodinamicamente instáveis é corroborada pela eficácia e segurança evidenciadas em ensaios clínicos aleatorizados (58, 90, 107) . As equinocandinas apresentam um perfil de segurança favorável, sendo relevante enfatizar, contudo, a probabilidade de interacções - 48 - Tatamento de infecções fúngicas invasivas medicamentosas que requerem ajuste de dose (89, 90). O papel do voriconazol é mais reduzido, pois diversos estudos evidenciaram um perfil de susceptibilidade não previsível em algumas espécies resistentes ao fluconazol. Apresenta, além disso, maior risco de interacções medicamentosas, menor tolerância em relação a outros antifúngicos e menor previsibilidade em termos farmacocinéticos (89, 90) . Pode, contudo, preencher um nicho importante em infecções por C. krusei, C. guilliermondii e C. glabrata susceptível, em doentes que podem fazer o switch de terapêutica endovenosa para terapêutica administrada por via oral (90, 107) .A utilização de anfotericina B deve ser preterida em doentes com infecção por C. lusitaniae, pelas evidências expostas nos capítulos anteriores (107). Em doentes neutropénicos, as recomendações são semelhantes, mas a força de recomendação pela IDSA altera-se em alguns pontos fulcrais. Nesta população, a utilização de fluconazol já apresenta uma força de recomendação bastante menor, ganhando as equinocandinas (caspofungina) e as formulações lipídicas de anfotericina B um papel mais preponderante, mesmo em relação ao voriconazol. Os dados de estudos aleatorizados são reduzidos nesta população e a utilização preferencial de formulações lipídicas de anfotericina B ou de caspofungina na prática clínica não é totalmente co-substanciada pelos diversos estudos. A subida de neutrófilos parece ser um factor crucial no sucesso da terapêutica antifúngica, facilmente perceptível pelos mecanismos de defesa do hospedeiro utilizados para combater infecções fúngicas (107). Em infecções urinárias ou pielonefrites a utilização de fluconazol é preferencial, excepto em caso de leveduras com perfil de susceptibilidade desfavorável. Em situações mais desfavoráveis como candidíase crónica disseminada, osteomielite por Candida spp., candidíase do sistema nervoso central, endocardite e endoftalmite recomenda-se a utilização de antifúngicos com espectro mais alargado e com actividade fungicida, mas o fluconazol em - 49 - Tatamento de infecções fúngicas invasivas altas doses mantém um papel preponderante na terapêutica de manutenção, que é extremamente prolongada (107). - 50 - Métodos disponíveis para avaliação de susceptibilidade a antifúngicos 4. Métodos disponíveis para avaliação de susceptibilidade a antifúngicos Somente em 1992 surge a primeira tentativa de padronização de protocolos para avaliação de susceptibilidade a antifúngicos pelo actual CLSI, baseado num método de macrodiluição em caldo (adaptado posteriormente a microdiluição). O primeiro documento M27-A foi aprovado em 1997, para fungos leveduriformes (108) . Na Europa, o EUCAST (European Committee on Antimicrobial Susceptibility Testing) procedeu ao desenvolvimento de um protocolo padronizado, que se provou ser equivalente ao preconizado pelo CLSI. Este protocolo constitui uma metodologia alternativa, com diferenças subtis em termos metodológicos (108). O método de difusão em discos é simples, desenvolvido para fungos leveduriformes e padronizado pelo CLSI no documento M44-A. No entanto, só possui pontos de corte para o fluconazol e voriconazol (27, 66). Outros métodos têm vindo a ser desenvolvidos e comercializados para utilização na prática diária. Entre estes, um dos mais conhecidos, pela sua facilidade de implementação e boa concordância com o método padronizado de microdiluição, é o Etest®. Trata-se de um método simples, tendo já sido utilizado para determinação de CIMs de fungos leveduriformes e filamentosos (66) . Por seu turno, o Sensititre YeastOne® baseia-se no método de microdiluição, mas utiliza um substracto cromogénico para facilitar a interpretação da CIM. Este método permite a determinação de CIMs para fluconazol, voriconazol, itraconazol, posaconazol, anfotericina B, flucitosina e caspofungina (66, 108) . Outro método disponível é o ATB Fungus 2®. Este método, igualmente baseado no método de microdiluição, permite a determinação de CIMs de fungos leveduriformes para fluconazol, flucitosina, anfotericina B e itraconazol. Pode utilizar a leitura visual ou automatizada (41, 152). Existem ainda métodos totalmente automatizados como o VITEK®, que utiliza espectrofotometria para leitura da CIM. Possui a vantagem relevante de se encontrar acoplado - 51 - Métodos disponíveis para avaliação de susceptibilidade a antifúngicos à identificação da levedura na amostra biológica. Diversos estudos revelaram que o VITEK® apresenta elevada reprodutibilidade e concordância com o método padronizado pelo CLSI (126, 130) . - 52 - Estudo Experimental - Objectivos II Estudo Experimental Objectivos do Estudo 1. Determinar a incidência de CI na UCI do Hospital Egas Moniz (HEM), durante o ano de 2009 (período de Maio a Dezembro); 2. Estudar a distribuição das diferentes espécies de Candida spp. responsáveis por CI; 3. Avaliar o comportamento dos diferentes isolados frente aos antifúngicos mais frequentemente utilizados no tratamento da CI. - 53 - Caracterização da Candidíase Invasiva na UCI do HEM 1. Caracterização da Candidíase Invasiva na UCI do Hospital Egas Moniz 1.1. Materiais e Métodos 1.1.1 População em estudo Neste estudo, a candidemia foi definida como pelo menos uma hemocultura positiva e a candidíase disseminada como uma cultura positiva obtida de amostras biológicas provenientes de locais normalmente estéreis. As amostras de Candida spp. de doentes que preencheram os critérios supracitados foram obtidas na UCI do HEM, e foram colhidas durante o ano de 2009 (período Maio a Dezembro). As amostras biológicas provieram de diferentes locais do organismo, especificamente: sangue, biópsias de tecido, incluindo pele e tecidos moles, e tracto genito-urinário. Foram ainda analisadas amostras provenientes do tracto respiratório, nomeadamente secreções brônquicas, lavados broncoalveolares e expectoração, e amostras obtidas de pontas de cateteres (linhas periféricas ou cateteres centrais). A presença de Candida numa amostra respiratória, mesmo após vários exames repetidos só foi considerada como infecção após eliminação de outra etiologia (tuberculose, pneumonia por vírus ou bactérias e outras etiologias). As amostras foram avaliadas pelo Laboratório de Microbiologia do HEM, e enviadas para o Laboratório de Micologia da Faculdade de Farmácia da Universidade de Lisboa, caso se verificasse a identificação de leveduras do género Candida. Foi ainda preenchida uma ficha clínica do doente com a seguinte informação: idade, etnia, género, data de admissão, condições médicas subjacentes, exposição a procedimentos médicos invasivos, utilização de - 54 - Caracterização da Candidíase Invasiva na UCI do HEM antibioterapia e terapêutica imunossupressora, data de diagnóstico, data de início de terapêutica antifúngica, evolução e resultado – ver Anexo I. 1.1.2. Tratamento prévio das amostras Os isolados foram armazenados a – 20ºC em meio Saboraud líquido com 15% de glicerol, até recuperação por subcultura. Esta foi realizada em meio Sabouraud dextrose ágar suplementado com cloranfenicol, durante 24-48h a 35ºC, para assegurar a pureza e viabilidade dos isolados (26). 1.1.3 Identificação de fungos Todos os isolados de Candida spp. foram identificados no Laboratório de Microbiologia do HEM, tendo as culturas sido enviadas para a Faculdade de Farmácia. 1.1.4 Tratamento de dados e análise estatística Para calcular a taxa de incidência de CI, foi recolhido o número de doentes internados no período em estudo. A taxa de incidência foi calculada em percentagem. A análise estatística foi processada no software EPI Info 2000 3.5 e no software SPSS, PAWS Statistic 18.0., usando como critério de significado estatístico, um nível de significância de 5% (p0,05 e IC de 95%). Foram efectuadas análises exploratórias dos dados mediante o cálculo das frequências absolutas e relativas, medidas de localização e dispersão. A associação estatística entre as variáveis foi determinada através de testes de 2, Teste Exacto de Fisher, Teste t de Student, ANOVA ou Teste Kuskall-Wallis, conforme aplicável. - 55 - Caracterização da Candidíase Invasiva na UCI do HEM 1.2 Resultados Durante o período de estudo foram recebidas, no Laboratório de Micologia da Faculdade de Farmácia da Universidade de Lisboa, 68 amostras. Estas amostras provêem de 46 doentes internados numa Unidade de Cuidados Intensivos, concluindo-se desde já que, de alguns doentes, se isolou diferentes espécies de Candida spp., numa mesma amostra, ou em diferentes amostras e espaços temporais. 1.2.1 Caracterização da população em estudo Quanto aos dados populacionais, poderemos referir que a nossa população apresenta uma média de 71 anos de idade (Gráfico 1) e média de duração de internamento de 29,89 dias (Gráfico 2). Tratou-se maioritariamente de uma amostra caucasiana (97,8%) do sexo masculino (63%), Gráfico 2. Curva de distribuição de duração de internamento (dias) na população estudada Gráfico 1. Curva de distribuição de idades na população estudada Detectaram-se 28 casos de CI em 26 doentes, num universo de 218 doentes internados na UCI durante o período de estudo, correspondendo a uma taxa de incidência de CI de 12,8%. Quanto à incidência de candidemia, foi detectado um caso durante o período de estudo (0,46%). A taxa de letalidade foi de 19,2 % (5 em 26 doentes). A razão de mortalidade - 56 - Caracterização da Candidíase Invasiva na UCI do HEM proporcional foi de 11,1% (5 casos num total de 45 mortes por outras causas, numa população de 218 doentes internados no período de estudo). As características demográficas e clínicas da população em estudo encontram-se espelhadas na Tabela 2. Em relação às causas de internamento, poderá referir-se que o 80% internamento/transferência na/para UCI deveu-se sobretudo a causas médicas 15% 13% 0% (80,2%) - Gráfico 3, com especial Médica relevância para a sepsis (57%), a pneumonia (54%) e insuficiência respiratória Cirurgia urgente Cirurgia coronária Cirurgia electiva Gráfico 3. Tipos de admissão na UCI (61%) - Gráfico 4. 57% 61% 54% 9% 9% 11% 15% 22% Gráfico 4. Causas de admissão na UCI - 57 - 9% 9% 9% Caracterização da Candidíase Invasiva na UCI do HEM Tabela 2. Características demográficas da população em estudo e estatística de internamento Duração Sexo Idade Vent. Cateteres nº dias internamento Masculino Feminino Cateter nº dias Mecânica 27,32 nº dias Traqueostomia Entubação nº dias urinário Média 71,93 30,10 N 29 29 Mediana 74,00 17,00 14,00 11,00 10,00 17,50 Mínimo 40 3 4 2 3 1 Máximo 89 175 175 120 120 65 Intervalo 49 172 171 118 117 64 % Total 63,0% Média 69,47 29,50 N 17 16 Mediana 75,00 15,50 16,00 10,50 16,00 15,00 Mínimo 38 2 3 2 5 3 Máximo 92 145 49 49 49 56 Intervalo 54 143 46 47 44 53 % Total 37% Média 71,02 29,89 N 46 45 Mediana 74,50 16,00 15,00 11,00 14,50 17,00 Mínimo 38 2 3 2 3 1 Máximo 92 175 175 120 120 65 Intervalo 54 173 172 118 117 64 25 20,14 23 62,5% 19 62,2% 18,20 15 15,00 23,90 40 57,1% 21,46 17,35 15 33,3% 42,9% 21,41 32 57,1% 19,53 3 40,6% 37 20 66,6% 13 37,8% 23,20 6 59,4% 14 37,5% 21,37 21,63 9 35 Total % Total 87% 80,4% 69,5% 19,5% 76% Tabela 3. Tipo de cirurgia prévia à admissão Na maioria dos casos, a sepsis e a pneumonia foram causas de admissão catalogadas como médicas, não tendo os doentes sofrido intervenções cirúrgicas. Os doentes cirúrgicos admitidos (n=11) realizaram diversos tipos de - 58 - Tipo cirurgia Frequência Percentagem Abdominal 4 36,4 Neurocirúrgica 4 36,4 Urológica 1 9,1 Torácica 1 9,1 Abdominal/vascular 1 9,1 Total 11 100,0 Caracterização da Candidíase Invasiva na UCI do HEM cirurgia, conforme espelhado na Tabela 3. Contudo, diversos doentes cirúrgicos foram internados na UCIP, devido a sepsis, bacteriemia, traqueobronquite aguda, ou seja, complicações pós-cirúrgicas relevantes - Tabela 4. Tabela 4. Tipos de causas de admissão na UCI Infecção Médica Sepsis Pneumonia Bacteriemia Não (%) 44 0 11 0 11 Sim (%) 59 68 8 11 11 urinária Inf. TBA IRC pós-op pós-op agudizada CT vascular 22 0 33 11 0 22 3 3 Insuf.Resp. IRA 56 22 5 70 Cirúrgica A população estudada apresentou ainda diversas co-morbilidades relevantes (Gráfico 5), designadamente hipertensão arterial (61%), diabetes mellitus (26%), doença pulmonar obstrutiva crónica (39%), insuficiência cardíaca (41%) e doença coronária (28%). Será relevante referir que mais de 10% da população apresentou mais do que 3 co-morbilidades. Esta população apresentou ainda doença auto-imune, em 11% dos casos, e desenvolveu neutropenia, em 8,7% dos casos. Observou-se frequentemente o recurso a terapêutica imunossupressora (33,3%). HTA DM 61% 78% IRC DHC 26,% DPOC 28% 15% Neoplasia TP 41% 39% 22% 9% VIH+ Auto-imune IC CAD 11% 2% 2% Outra Gráfico 5. Tipo de co-morbilidades observadas na população em estudo - 59 - Caracterização da Candidíase Invasiva na UCI do HEM Desta população, somente 26 doentes possuem diagnóstico de candidíase. A presença de Candida numa secreção respiratória só foi considerada infecção após exclusão de outras etiologias. O ponto de partida da infecção, considerando o diagnóstico de candidíase, pode ser observado na Tabela 5. Tabela 5. Ponto de Partida de episódios de CI Candidíase Ponto Origem Amostras (N Partida Episódios) Tracto Cateter Hemocultura Urocultura Outra respiratório Pulmão 19 0 0 1 19 0 Urinário 6 1 0 5 0 2 Hematogéneo 3 0 1 0 1 2 Na Tabela 6 podemos ver o tipo de terapêutica antibiótica utilizada nesta população e o nº dias de terapêutica prévia ao resultado microbiológico (considerada toda a terapêutica realizada no mês anterior). Deve salientar-se a utilização generalizada de antibióticos de largo espectro na nossa população. Para além disso, observou-se a instituição de terapêutica tuberculostática em 2 doentes da população estudada. Tabela 6. Padrão de utilização de antibioterapia (AB) prévia (nºdias) na população estudada. Vancomicina Carbapenem Quinolonas Penicilinas Piperacilina Aminoglicosideos Macrólidos Polimixinas Cefalosporinas Clindamicina Metronidazol Linezolide N(Doentes) 37 13 19 20 14 27 11 5 9 4 4 Média 8,32 9,54 7,74 6,60 11,50 10,33 7,73 25,80 5,56 10,25 7,50 Mediana 7,00 6,00 7,00 6,00 8,50 8,00 8,00 23,00 3,00 9,00 7,50 Moda 3 1 2 2 5 1 8 9 2 4 2 Mínimo 1 1 1 1 3 1 2 9 2 4 2 Máximo 25 41 22 19 32 38 13 53 14 19 13 Nota: nos doentes que não fizeram qualquer terapêutica antifúngica, foi considerada o nº total de dias de utilização de AB. - 60 - Caracterização da Candidíase Invasiva na UCI do HEM Considerando especificamente a subpopulação com diagnóstico de candidíase, verificou-se a prevalência de co-morbilidades como hipertensão arterial (69,2%), doença pulmonar obstrutiva crónica (46,2%), insuficiência cardíaca (42,3%) e diabetes mellitus (30,8%). A subpopulação com diagnóstico de candidíase apresentou uma média de idade de 72,3 anos, similar à média da população geral estudada, uma mediana de 74,5 anos e uma moda de 77 anos. O diagnóstico de candidíase foi mais prevalente no sexo masculino. Relativamente à utilização de procedimentos invasivos, a utilização de cateter venoso central foi observada em 100% da população com diagnóstico de candidíase. A ventilação mecânica foi utilizada em 88,5% (n=23) desta população, enquanto a entubação nasogástrica foi utilizada em 80,8% (n=21), predominantemente mais do que 10 dias. A utilização de cateter urinário foi bastante prevalente nesta subpopulação (n= 19/73,1%), com um padrão de utilização frequentemente superior a 10 dias. Dos doentes traqueostomizados (n=9), 6 (66,7%) desenvolveram candidíase. É ainda de salientar a elevada prevalência de candidíase em doentes neoplásicos (n=6 de um total de 10). Após análise estatística da população não se observou qualquer associação estatisticamente significativa entre co-morbilidades identificadas, factores de risco identificados na literatura internacional e desenvolvimento de CI, comparando os dados de doentes sem diagnóstico de candidíase e com diagnóstico de candidíase. Nesta subpopulação, a utilização de antibioterapia foi relevante, mas sem diferenças significativas (em termos de distribuição) da população que não desenvolveu candidíase. Alguns dados são, todavia, pertinentes referir. A utilização de carbapenemes foi relativamente frequente (n=22), sendo que 57,7% utilizou este grupo farmacoterapêutico por um período superior a 5 dias, previamente ao diagnóstico de CI. Dos 26 doentes com diagnóstico de candidíase, 12 utilizaram previamente piperacilina+tazobactan, o que correspondeu a 60% de todos os doentes que utilizaram este antibiótico (Tabela 6). O número médio de dias de - 61 - Caracterização da Candidíase Invasiva na UCI do HEM utilização de piperacilina+tazobactan foi de 7,3 dias, nesta subpopulação. Pelo contrário, a utilização de cefalosporinas foi mais prevalente nos doentes que não desenvolveram CI (66,6%), encontrando-se descrita somente em 3 doentes com diagnóstico de CI, e por um período de utilização inferior a cinco dias, previamente ao diagnóstico de candidíase. 1.2.2 Distribuição das espécies A distribuição de espécies, de acordo com as amostras recebidas (n=68), foi a seguinte: C. albicans, 72,1% (n= 49), C. glabrata, 7,4% (n=5), , C.krusei, 7,4% (n=5), C. parapsilosis, 4,4% (n=3), C. tropicalis, 4,4% (n=3), C. lusitaniae, 2,9% (n=2) e C. guilliermondii, 1,4% (n=1) – Tabela 7. Tabela 7. Distribuição de leveduras de acordo com origem, espécie e número de doentes em que foram isoladas Espécie N Doentes Origem Hemocultura Urocultura Cateter Secreções brônquicas Outros C. albicans 1 14 4 27 3 36 C. glabrata 0 1 1 2 1 5 C.krusei 0 0 1 4 0 4 C. lusitaniae 0 0 0 2 0 2 C.parapsilosis 0 0 1 2 0 2 C. tropicalis 0 1 0 1 1 3 C. guilliermondii Total (%) 0 0 1 0 0 1 1 (1,5) 16 (23,5) 8 (11,8) 38 (55,9) 5 (7,3) NA Considerando os episódios de CI, a distribuição das espécies encontra-se descrita no Gráfico 6, demonstrando a importância de espécies não-albicans. 9,70% 3,10% C. albicans 12,40% C. glabrata 59,30% C. krusei C. tropicalis 15,50% C. parapsilosis Gráfico 6. Distribuição das espécies responsáveis por CI - 62 - Caracterização da Candidíase Invasiva na UCI do HEM É interessante verificar que 6 episódios (21,4%) de CI se deveram a infecções mistas isto é, por mais do que uma espécie de Candida. As espécies não-albicans foram relevantes neste tipo de infecções, sobretudo C. krusei e C. glabrata. Observaram-se 2 infecções mistas de C. albicans + C.krusei, 1 de C. albicans + C. parapsilosis, 1 de C. glabrata + C. tropicalis, 1 de C. krusei + C. glabrata e 1 de C. albicans + C. glabrata. Na população estudada, o isolamento de determinadas espécies aparentou estar associado a utilização de procedimentos invasivos, presença de co-morbilidades ou utilização de determinada antibioterapia prévia, embora não se tenha encontrado associação estatisticamente significativa entre estes factores e o desenvolvimento de CI. Na amostra estudada, o isolamento de C. albicans foi relevante nos doentes com ventilação mecânica, já que 84,2% (n=32) da população com crescimento de C. albicans utilizou ventilação mecânica invasiva. A entubação nasogástrica demonstrou igualmente relevância em doentes com isolamento de C. albicans, dado que 86,1% (n=31) dos doentes a utilizou. Adicionalmente, nos doentes com IRC e CAD, o isolamento de C. albicans foi relevante, isolando-se em amostras de 42,9% e 53,8% destas subpopulações, respectivamente. Na subpopulação com CAD (n=13), somente 2 doentes desenvolveram candidíase por C. albicans, com ponto de partida pulmonar. Não se observou crescimento da espécie C. lusitaniae em doentes com ventilação mecânica ou entubação nasogástrica, apesar da origem das amostras biológicas (secreções respiratórias), pensando-se que a presença destes dois factores não tenha sido relevante para a colonização/infecção por C. lusitaniae. - 63 - Caracterização da Candidíase Invasiva na UCI do HEM Os 4 doentes com culturas positivas para C. krusei utilizaram carbapenemes (meropenem), com uma média de 5,5 dias de utilização. Estes doentes apresentaram uma mediana de 43 dias de utilização de cateter central, contrariando a mediana de dias de utilização deste dispositivo na população estudada, que foi inferior. Contudo, C. krusei foi mais frequentemente isolada em amostras respiratórias. No caso de C. glabrata, 3 dos cinco doentes com amostras positivas para esta levedura possuíam CAD como co-morbilidade. Todos desenvolveram candidíase, com diversos pontos de partida. Considerando variáveis como sexo e idade, verificou-se a relação não surpreendente entre aparecimento de Candida spp. em urocultura e o sexo (p0,05), com 65% de uroculturas positivas de Candida spp. pertencentes à população feminina. A utilização de antibióticos de largo espectro foi prevalente nos doentes com isolamento de C. albicans, nomeadamente de vancomicina ou linezolide (n=20) e de carbapenemes (n=28). Observou-se uma diferença no padrão de utilização de quinolonas, quando comparado o isolamento de C. albicans vs. espécies não-albicans. Em 80,5% (n=29) dos doentes, em que se verificou isolamento de C. albicans, não foram administradas quinolonas (Tabela 7). Constatou-se que Candida não-albicans foi isolada frequentemente em doentes com administração prévia deste grupo terapêutico. C. lusitaniae desenvolveu-se em dois doentes com IRC, num universo de 7 doentes com IRC. Estes doentes também utilizaram quinolonas como antibioterapia prévia, entre outros fármacos, correspondendo a 15,4% dos doentes que utilizaram este tipo de antibioterapia (Tabela 6). - 64 - Caracterização da Candidíase Invasiva na UCI do HEM 1.2.3 Padrão de utilização de terapêutica antifúngica Verificou-se a instituição de terapêutica antifúngica em grande parte da população estudada (n=32). Pelo contrário, em 14 doentes não foi utilizada qualquer terapêutica antifúngica, não tendo os resultados microbiológicos sido valorizados clinicamente. Relativamente ao padrão de utilização de antifúngicos, o fluconazol foi o antifúngico seleccionado mais frequentemente, tendo sido utilizado em 63% (n=24) dos cursos de antifúngicos (ntotal=38) instituídos nesta população, seguido da caspofungina (n=8/21%), voriconazol (n=3/8%) e, por último, anfotericina B lipossómica (n=3/8%). Alguns doentes realizaram (n=5) mais do que um curso de terapêutica antifúngica na UCI. A anidulafungina foi utilizada num doente integrado num ensaio clínico, tendo sido feito o switch para caspofungina após 4 dias de terapêutica. A média de nº de dias de terapêutica foi de 13,9 dias, com uma mediana de 16 dias e uma moda de 16 dias. Poderemos classificar os cursos de antifúngico instituídos (n=38) de acordo com os seguintes tipos: empírica, profiláctica ou dirigida (curativa/documentada) - Tabela 8. Tabela 8. Tipo de terapêutica instituída Tipo antifúngico Tipo Terapêutica Empírico Profilaxia Dirigida Fluconazol 9 2 13 Voriconazol 2 0 1 Equinocandinas 5 0 3 Polienos 2 0 1 Na tabela 9 encontra-se resumido o padrão de utilização de terapêutica antifúngica prévia (nº dias) ao resultado microbiológico positivo para Candida. - 65 - Caracterização da Candidíase Invasiva na UCI do HEM Tabela 9. Padrão de utilização de terapêutica antifúngica prévia (nºdias) na população estudada. FLUCONAZOL CASPOFUNGINA VORICONAZOL ANFOTERICINA B N (doentes) 6 1 0 2 Média 10 16 16 Mediana 8,5 16 16 Moda 9 Intervalo 17 Mínimo 3 Máximo 21 No universo de doentes (n=8) que realizaram terapêutica antifúngica prévia ao isolamento de Candida, quer por instituição de diversos cursos de antifúngicos ou exposição a antifúngicos no mês anterior na UCI ou noutro serviço clínico, verificou-se a emergência de espécies nãoalbicans, algumas com padrões de resistência/susceptibilidade interessantes. A análise descritiva da terapêutica prévia com antifúngicos e emergência de isolados com resistência/susceptibilidade reduzida encontra-se espelhada na Tabela 10. Tabela 10. Análise descritiva da terapêutica prévia com antifúngicos e emergência de isolados resistentes. Isolado N Origem Amostra Terapêutica prévia (n) Padrão de resistência C. lusitaniae 1 Secreções respiratórias Fluconazol (1) - C. glabrata 2 C. parapsilosis 2(a) C. guilliermondii 1 Cateter C. krusei 2(a) Secreções respiratórias Secreções respiratórias (1) Fluconazol (2) Urocultura (1) Secreções respiratórias (1) 1 –S-DD Fluconazol 1 –S –Fluconazol Caspofungina (1) 2 –NS Anidulafunginab Fluconazol NS- Equinocandinasc Fluconazol (1) 1-R- Fluconazol Anfotericina B Lipossómica (1) 1-S-DD- Fluconazol Cateter (1) (a) amostras provenientes do mesmo doente em espaços temporais diferentes; (b) categorizados como NS pelo método Etest® (c) categorizado como NS pelo método Etest®, CIM=2µg/mL pelo método de microdiluição; - 66 - Caracterização da Candidíase Invasiva na UCI do HEM 1.3 Discussão Como sabemos, as infecções por espécies do género Candida spp. são frequentes na espécie humana, porém a colonização é um pré-requisito para o desenvolvimento de candidíase (40). Não existe, como tal, consenso na definição de CI, excepto para doentes imunocomprometidos, conforme referido anteriormente. Consideram-se 3 níveis de probabilidade de diagnóstico: infecção provada, provável ou possível, levando em linha de conta os factores do hospedeiro, sintomatologia clínica e evidência micológica (10, 37) - Tabela 1. Estas definições apresentam algumas limitações, com dificuldade de transposição para a prática clínica. Existe, pois, controvérsia envolvendo a interpretação de colheitas positivas para Candida spp. quer em hemoculturas, quer em amostras de urina, expectoração ou pus de feridas abertas (10) . Estes factos colocaram-nos desafios na correcta interpretação de todos os dados microbiológicos, sendo difícil distinguir colonização por Candida spp. de infecção em doentes críticos internados em UCIs e, por conseguinte, avaliar dentro da nossa população quais os doentes com diagnóstico de CI. Foi por isso necessário recorrer quer aos dados microbiológicos e ficha de recolha de dados (Anexo I) utilizada neste estudo, quer aos diagnósticos disponíveis no processo clínico electrónico do doente e perfil farmacoterapêutico dos doentes. 1.3.1 Incidência de CI e caracterização da população em estudo No período em estudo, observámos uma incidência de CI de 12,8%. Quanto à incidência de candidemia, foi bastante mais baixa, de 0,46%. A taxa de letalidade foi de 19,2% (5 em 26 doentes). A razão de mortalidade proporcional foi de 11,1% (5 casos num total de 45 mortes por outras causas numa população de 218 doentes). - 67 - Caracterização da Candidíase Invasiva na UCI do HEM Dados de 790 UCIs dos Estados Unidos revelaram que Candida spp. foi responsável por cerca de 5 a 10% de infecções da corrente sanguínea. O estudo europeu EPIC (European Study on the Prevalence of Nosocomial Infections in Critically Ill) reportou que a CI foi responsável por 17% das infecções adquiridas no hospital (40, 52) . A candidemia representou cerca de 10-20% de todas as formas de candidíase (40). O estudo SENTRY realizado entre 1997 e 1999, em que se avaliaram as infecções da corrente sanguínea nos Estados Unidos, Canadá, América Latina e Europa verificou que a candidemia constituiu 3% de todas as infecções sanguíneas adquiridas na comunidade e 9% das nosocomiais ou associadas a cuidados de saúde (52) . Em UCIs observam-se incidências geralmente superiores às observadas numa enfermaria, rondado a candidemia os 10% de incidência. Uma revisão francesa revelou uma incidência média de 6,1/1000 admissões em medicina intensiva (39). Outros autores referem que a incidência geral de CI se situa à volta dos 2% em doentes críticos, existindo, contudo, subpopulações de alto risco em que a incidência pode chegar aos 35%. Nos Estados Unidos, dados concluem que Candida causa cerca de 10% das sepsis em Unidades de Cuidados Intensivos (34). Estudos observacionais revelaram ainda que as formas mais frequentes de candidíase são candidemia, infecções relacionadas com cateter, infecções intra-abdominais e infecções do tracto urinário (90). Em Portugal, o estudo epidemiológico num hospital no Porto (não só em Cuidados Intensivos) revelou uma incidência de 2 casos por cada 1000 admissões hospitalares, com mortalidades elevadas, de 39,3% (32) . Dados oficiais norte-americanos revelam que a mortalidade atribuível a CI ronda os 15-35% nos adultos. Contudo, os estudos - 68 - Caracterização da Candidíase Invasiva na UCI do HEM epidemiológicos são heterogéneos na avaliação da mortalidade a Candida spp., revelando dados díspares, entre 5 a 71% de mortalidade (34, 52, 90, 114, 141). A comparação dos nossos dados com os de estudos internacionais supracitados poderá não ser adequada, por diferenças no tamanho e heterogeneidade das amostras, desenho dos estudos e tipo de análise estatística conduzida. Alguns estudos na literatura internacional apresentam taxas de incidência considerando o n.º doentes/dia (16, 31), dados que não nos foi possível obter. A taxa de letalidade observada neste estudo encontra-se aquém da avaliação da mortalidade descrita no estudo epidemiológico português. Há que referir que, nesse estudo, a mortalidade foi calculada para todos os episódios de fungemia observados, quer por Candida spp. ou por outros géneros, considerando-se uma amostragem superior através da avaliação de todas as hemoculturas positivas para fungos, provenientes de todos os serviços clínicos daquele hospital (32). Todavia, a percentagem de CI, no nosso estudo, encontra-se de acordo com dados provenientes de alguns estudos conduzidos em UCIs, como é o caso do estudo EPCAN, levado a cabo em 73 UCIs espanholas, em que se observou uma incidência de 6,3% de CI (52). Pelo contrário, a incidência de candidemia neste estudo é mais baixa do que seria de esperar, de acordo com o estudo EPCAN - 3,9% de candidemia (52), tendo-se observado um único caso no período de estudo. De facto, dos 3 doentes com diagnóstico de CI disseminada, somente um doente apresentou uma hemocultura positiva. Os outros doentes apresentaram infecções intra-abdominais graves, por mais de uma espécie de Candida. Assim, tal como observado em estudos epidemiológicos internacionais, verificou-se neste estudo que as formas de candidíase com ponto de partida do tracto urinário ou intra-abdominal foram relevantes na nossa população (Tabela 5). Tal poderá relacionar-se com os motivos de internamento em UCI. Como esperado, a sepsis e pneumonia foram causas de admissão médica muito relevantes (Gráfico 4/Tabela 4). No entanto, o número de doentes cirúrgicos admitidos, com realização - 69 - Caracterização da Candidíase Invasiva na UCI do HEM de diversos tipos de cirurgias, não é de menosprezar (n=11). Entre estas, as cirurgias abdominal e neurocirúrgica foram bastante prevalentes (Tabela 3). No que toca às infecções do tracto urinário, deverá referir-se que cerca de 69,5% (n=33) da população estudada utilizou cateter urinário, com uma média de 21 dias de utilização (Tabela 2), dos quais 6 doentes tiveram diagnóstico de candidíase de ponto de partida urinário. Porém, não se encontrou nenhuma associação estatística entre a utilização de cateter urinário e/ou o número de dias de utilização e desenvolvimento de CI. A colonização do tracto urinário de doentes em UCIs por Candida encontra-se bem descrita(20, 34, 39, 40, 114, 141) , tendo estado patente, na nossa amostra, em16 doentes com uroculturas positivas para Candida spp., com uma valorização clínica de 37,5% (n=6) das uroculturas (Tabela 5). Tal prática encontra-se adequada, pois o significado clínico de candidúria, sobretudo em doentes cateterizados é controverso (20, 22) . De qualquer forma, a candidúria encontra-se associada a mortalidade superior, sobretudo em doentes com co-morbilidades, muito embora a presença de candidemia concomitante seja relativamente infrequente (20, 22) . Há que considerar que a formação de biofilmes por Candida é relevante em diversas situações clínicas, incluindo colonização de dispositivos invasivos como cateteres urinários, e evidências epidemiológicas sugerem que estas entidades possam representar um papel no desenvolvimento de quadros infecciosos (20, 22, 36, 160) . Aliás, diversos estudos têm associado o aparecimento de candidúria com determinados factores de risco como sexo feminino, instrumentação do tracto urinário, diabetes mellitus, entre outros (20) . Não é surpreendente verificar que a candidúria também se relacionou na nossa amostra com o sexo feminino (p0,05), com 62,5% de uroculturas positivas de Candida spp. pertencentes à população feminina. Tal associação não ficou demonstrada para a subpopulação com diabetes mellitus. - 70 - Caracterização da Candidíase Invasiva na UCI do HEM Considerando doentes cirúrgicos, diversos estudos indicam um risco superior de desenvolvimento de CI em determinadas subpopulações de doentes submetidas a procedimentos cirúrgicos extensos e/ou a instrumentação com dispositivos médicos invasivos (40, 90, 114, 157) . Em estudos de Blumberg et al. e Wey et al. a cirurgia abdominal prévia demonstrou um risco relativo 7.3 superior para desenvolvimento de CI. Os autores identificaram ainda a utilização de cateter central, insuficiência renal aguda, administração de nutrição parentérica, utilização de múltiplos antibióticos e isolamento de Candida spp. em diversas amostras biológicas, como factores de risco independentes de desenvolvimento de CI (141). A CI de ponto de partida pulmonar foi também muito frequente nesta população, se não a mais frequente. O isolamento de Candida spp. a partir de amostras respiratórias, nomeadamente lavados broncoalveoloares ou secreções brônquicas, é julgado de forma diversificada de acordo com diversos autores e de acordo com o tipo de doentes considerados. Estudos mais antigos em doentes neutropénicos demonstraram que o isolamento de Candida spp. a partir de amostras respiratórias poderia representar um verdadeira infecção, em parte devido ao papel defensivo dos glóbulos brancos polimorfonucleares, deplectados neste tipo de população. A presença de febre persistente e resistente a antibióticos, bem como presença de infiltrados pulmonares, em conjunto com isolamento de Candida spp. em amostras broncoscópicas podem representar condições que põem em risco a vida (135) . Em doentes não neutropénicos, a avaliação deste tipo de amostras e a sua valorização clínica é muito mais difícil, podendo significar colonização e não infecção. Alguns autores argumentam que um lavado broncoalveolar pode ser considerado uma amostra proveniente de um local normalmente estéril e que a pneumonia por Candida é uma entidade a recear (39, 157). Por estes factos, a avaliação clínica destas amostras descartou infecção em cerca de 50% dos doentes - 71 - Caracterização da Candidíase Invasiva na UCI do HEM com amostras respiratórias positivas e a presença de Candida numa secreção respiratória só foi considerada infecção após exclusão de outras etiologias. Importa ainda referir o tipo de doente admitido na UCI durante o período em estudo. A população estudada apresentou como co-morbilidades relevantes hipertensão arterial, diabetes mellitus, doença pulmonar obstrutiva crónica, insuficiência cardíaca, doença coronária, entre outras. Esta população apresentou menos frequentemente doença auto-imune e neutropenia (Gráfico 5). A percentagem de doentes com história de neoplasia ou neoplasia activa foi de 21,7%, não se observando a existência de doenças hematoncológicas na nossa população. Contudo, deve realçar-se a elevada prevalência de candidíase em doentes neoplásicos (n=6, de um total de 10). A utilização de terapêutica imunossupressora, nomeadamente corticoterapia em altas doses, foi relativamente frequente (33,3%). Se pensarmos que múltiplos estudos indicam um risco superior de desenvolvimento de CI e/ou candidemia em doentes com idades extremas (prematuros e idade superior a 70 anos), diabéticos, oncológicos (particularmente com neutropenia), sob administração de antibióticos de largo espectro, doentes do sexo masculino, sob utilização de terapêutica imunossupressora e sob ventilação mecânica invasiva, (31, 32, 40, 85, 114, 141) facilmente observamos a prevalência de alguns destes factores na população analisada. A estadia prolongada em UCI também tem sido associada, como factor de risco independente, ao desenvolvimento de CI (40, 90, 141) . Não podemos esquecer que a análise e interpretação de parâmetros, considerados como factores de risco na literatura internacional, deve ser cuidadosa, pois os resultados obtidos podem depender da heterogeneidade da população em estudo, desenho de estudo e tipo de análise estatística desenvolvida. No nosso estudo, avaliámos a relação entre o desenvolvimento de CI e alguns destes factores, com o objectivo de observar as tendências na nossa população. - 72 - Caracterização da Candidíase Invasiva na UCI do HEM A nossa população apresentou uma média de idades de 71 anos, com um claro desvio da curva de distribuição normalizada de Gauss à direita (Gráfico 1), tratou-se maioritariamente de uma amostra do sexo masculino (63%) e com uma média de duração de internamento de 29,89 dias (Tabela 2). Observou-se um claro desvio da curva de distribuição dos dias de internamento à esquerda (Gráfico 2). Apesar da média de dias de internamento ser 29,89 dias, a mediana foi relativamente mais baixa, e 52% da população não esteve internada mais do que 16 dias. Análises conduzidas em UCIs revelam dados contraditórios relativamente à média de dias após admissão hospitalar em que se observou a ocorrência de casos de CI (31, 32, 90) . Esta avaliação não foi realizada no nosso estudo, tratando-se de uma limitação do mesmo. Para além disso, o isolamento de C. albicans foi relevante em algumas subpopulações com co-morbilidades, como IRC e CAD. O isolamento de C. glabrata também demonstrou importância na subpopulação com CAD. Relativamente à insuficiência renal, esta comorbilidade encontra-se descrita como factor minor predisponente ao desenvolvimento de CI (39) . Quanto à doença arterial coronária, a relação entre as duas variáveis poderá ser bastante mais complexa e envolver mecanismos imunofisiológicos. Investigadores descreveram o potencial aterogénico de alguns agentes patogénicos, incluindo fungos, com implicações quer na iniciação quer na progressão da doença aterogénica (4) . Esta hipótese é corroborada por achados micológicos em placas ateroescleróticas. Jegier et al. identificaram ADN de C. albicans em amostras de tecido aórtico, em doentes com CAD e estenose aórtica. C. albicans relacionou-se com a expressão de moléculas de adesão com actividade pro-inflamatória. O estudo de Jegier et al. foi conduzido em doentes sem história de candidíase ou outro tipo de infecções nos últimos 3 meses, ou sujeitos a qualquer tipo de terapêutica imunossupressora (65) . Contudo, existem estudos em modelos animais que contradizem estes resultados. Assim, argumenta-se que o ADN de diversos agentes patogénicos se pode alojar em tecidos - 73 - Caracterização da Candidíase Invasiva na UCI do HEM saudáveis, sem implicações clínicas ou consequências pro-inflamatórias (65, 167) . De qualquer forma, um aumento de predisposição para aparecimento de Candida spp. em doentes com doença arterial coronária parece improvável. Deverá ser salientado que co-morbilidades cardíacas são, todavia, muito frequentes em doentes em ambiente de Cuidados Intensivos (110). A utilização de ventilação invasiva foi bastante prevalente na nossa população, com utilização em 80,4% dos casos (Tabela 2). Este facto poderá estar ligado ao tipo de causas de internamento mais frequentemente observadas na UCI: sepsis, pneumonia e insuficiência respiratória agudizada. O isolamento de C. albicans foi relevante nos doentes com ventilação mecânica. A associação descrita é fundamentada por diversas fontes bibliográficas (34, 40, 114) . A ventilação mecânica pode funcionar como uma porta de entrada para a colonização e posterior infecção por Candida albicans (114). Não se observou aumento de prevalência de isolamento de C. albicans em doentes com utilização prolongada de ventilação mecânica. É de salientar que não foram avaliados o número de dias de utilização de procedimentos invasivos até ao isolamento de espécies de Candida, pelo que qualquer associação estatística observada teria algumas limitações de interpretação. A entubação nasogástrica também demonstrou relevância em doentes com isolamento de C. albicans. Neste caso, a explicação mais plausível para esta tendência seria que a utilização de sonda nasogástrica permite a colonização da mucosa do tracto gastrointestinal e, em última análise, promove a translocação da levedura através da mucosa gastrointestinal. Diversos factores de risco independentes referidos na literatura internacional poderão estar a causar algum viés nesta tendência, nomeadamente a administração de protectores da mucosa gástrica e a utilização de nutrição parentérica (40, 114) . A utilização de nutrição parentérica e protectores da mucosa gástrica ocorrem, - 74 - Caracterização da Candidíase Invasiva na UCI do HEM frequentemente, em simultâneo com a utilização de entubação nasogástrica. Estes dados não foram recolhidos, não sendo possível avaliar o seu impacto na nossa população. Pelo contrário, os dois doentes com isolamento de C. lusitaniae não apresentaram CI, encontrando-se colonizados a nível do tracto respiratório, sem utilização de ventilação invasiva ou entubação nasogástrica. Tal facto indicia que a colonização do tracto respiratório por este agente poderá ocorrer, independentemente da utilização de determinado tipo de dispositivos invasivos. A epidemiologia de infecções fúngicas por C. lusitaniae encontra-se ainda hoje mal compreendida, sendo difícil a associação deste tipo de infecção a factores de risco independentes. O estudo europeu conduzido pela European Confederation of Medical Mycology (ECMM) só reportou dois casos de candidemia por C. lusitaniae em cerca de 2000 casos, durante um período de 28 meses (154) . Os resultados do estudo ARTEMIS apresentam uma maior percentagem de isolamento deste agente, considerando um muito maior número de amostras biológicas (116). Esta espécie tem sido isolada em amostras humanas de sangue, urina e orofarínge, entre outras. A colonização endógena por Candida spp. destas zonas anatómicas é, de resto, conhecida, fundamentando, em parte, as nossas conclusões (40, 50) . Sanchez et al. reportaram a aquisição exógena (nosocomial) de C. lusitaniae na orofarínge e em amostras de fezes em doentes internados, sem desenvolvimento de infecção por este agente. Estes investigadores não identificaram factores de risco independentes quer para a colonização quer para a infecção por este agente, mas evidenciaram o papel da aquisição exógena nosocomial por esta espécie, numa população imunodeprimida (137). Também a tendência de isolamento por C. lusitaniae em doentes com IRC é de difícil explicação, embora Parkins et al. tenham associado a pré-existência de falência renal com o risco aumentado de mortalidade devido a candidemia (110) . Outros autores referem que a hemodiálise aumenta o risco de colonização por Candida, sendo o índice de risco de 18,1 (39). - 75 - Caracterização da Candidíase Invasiva na UCI do HEM A colonização de linhas vasculares por parte deste agente também se encontra documentada (150) . De facto, um dos doentes em questão sofreu um episódio de agudização da sua insuficiência renal, com necessidade de suporte dialítico contínuo, mas estes dados não se encontram descritos para todos os doentes com IRC da nossa amostra. Como tal, não nos é possível retirar qualquer conclusão destes dados, podendo tratar-se de uma coincidência o isolamento desta espécie em doentes com esta co-morbilidade. Poderia ter sido relevante a avaliação de outros factores descritos na literatura internacional como factores de risco de desenvolvimento de CI, tais como utilização de protectores da mucosa gástrica e nutrição parentérica, técnicas dialíticas, bem como o número de antibióticos utilizados em simultâneo. Estes factos são obviamente limitações do estudo por nós desenvolvido. Para avaliação da importância relativa de factores de risco preditores de infecção fúngica, seria necessário uma estratégia de investigação diferente. Poderemos salientar que 58% (n=27) dos doentes realizaram antibioterapia de largo espectro com um carbapenem + vancomicina ou linezolide. Metade dos doentes tratados com estes antibióticos desenvolveu candidíase. Também a piperacilina+tazobactan foi utilizada frequentemente. Importa, contudo, relevar que a administração de piperacilina+tazobactam e vancomicina têm sido associadas significativamente ao desenvolvimento de candidemia causada por C. glabrata e C. krusei (114) . Na nossa população, não seria possível realizar este tipo de análise, pois só foi verificado um caso de candidemia por C. albicans. Um estudo por Giuliano et al. refere uma correlação entre a utilização de piperacilina e o aumento de leveduras nas fezes e consequente diminuição de bactérias anaeróbias, com identificação dos agentes C. glabrata e C. tropicalis nas fezes (56). Por outro lado, Pultz et al. avaliaram o impacto da utilização de antibioterapia inibidora de anaeróbios obrigatórios, - 76 - Caracterização da Candidíase Invasiva na UCI do HEM como piperacilina+tazobactan, ceftriaxona, metronidazol e clindamicina, na colonização do tracto gastrointestinal por C. glabrata, observando aumento da colonização com a utilização destes fármacos, ao contrário de antibióticos como a levofloxacina e o cefepime (131). Na nossa população verificou-se uma tendência para o isolamento de espécies não-albicans em doentes que utilizaram previamente quinolonas. Paralelamente, a utilização de cefalosporinas foi mais prevalente nos doentes que não desenvolveram CI. Os dados apresentados são corroborados por diversos estudos. Análises recentes do impacto da terapêutica com cefalosporinas de largo espectro na flora intestinal de indivíduos, identificaram alterações não significativas de C. albicans na flora microbiológica (106) . Alvarez et al. avaliaram, através de uma meta-análise, o risco de sobre-infecção por agentes patogénicos, incluindo Candida spp., após exposição a diversos antibióticos de largo espectro. Entre as limitações desta meta-análise, enumeram-se a não avaliação do impacto da ceftriaxona e da ceftazidima (duas cefalosporinas de 3ª geração) e a dificuldade de realização de análise estatística. Esta meta-análise revelou um percentagem relevante de doentes sob terapêutica com ciprofloxacina (25%) ou cefotaxima (8,9%), que desenvolveu sobre-infecção por Candida spp., sem especificação do tipo de espécies mais frequentemente isoladas. Os carbapenemes, cefepime e piperacilina+tazobactan foram responsáveis por uma muito menor percentagem de casos de sobre-infecção (na ordem dos 1,5 a 2,7%). De qualquer forma, a utilização de antibióticos associou-se com maior frequência a sobre-infecções causadas por bactérias do género Enterococcus (3). Um estudo eslovaco retrospectivo evidenciou o papel da terapêutica com quinolonas no desenvolvimento de sobre-infecção por fungos em doentes oncológicos, independentemente das espécies isoladas. Neste estudo, as espécies não-albicans foram responsáveis por 26,8% de todas as fungemias registadas e C. albicans por 43,9% (72). - 77 - Caracterização da Candidíase Invasiva na UCI do HEM 1.3.2 Distribuição das espécies de Candida spp. Outros dados epidemiológicos relevantes caracterizados no nosso estudo incluem a distribuição das espécies isoladas, locais de isolamento e susceptibilidade a antifúngicos. Estudos internacionais revelam que cerca de 90% das infecções são atribuíveis a C. albicans, C. glabrata, C. parapsilosis, C. tropicalis e C. krusei. A incidência de C. albicans tem vindo a diminuir, correspondendo a cerca de 50% a 70% consoante região geográfica, com a emergência de espécies não-albicans, como C. parapsilosis e C. glabrata, com taxas de incidência e importância variáveis consoante a região geográfica (85, 93, 114, 118, 154, 164) . Estudos em Cuidados Intensivos revelaram percentagens mais baixas de incidência de C. albicans (16, 52) . Igualmente, diversos autores que avaliaram a distribuição de espécies isoladas em hemoculturas (candidemia), verificaram baixas taxas de incidência de C. albicans, com emergência de C. glabrata, C. parapsilosis e C. tropicalis (57, 93, 111, 154). Esperar-se-ia, por isso, conformidade com os dados de vigilância epidemiológicos obtidos em estudos internacionais, nomeadamente o Programa ARTEMIS e Estudo SENTRY, entre outros (93, 116, 118). Seria de prever uma percentagem menos significativa de CI causada por C. albicans, com crescimento da importância das espécies não-albicans, como C. glabrata, C. parapsilosis, C. tropicalis e C. krusei. Seria também esperado um aumento da identificação de espécies mais raras como C. guilliermondii, C. kefyr, C. rugosa, C. lusitaniae e C. famata (57, 111, 116, 164) . Apesar de alguns casos raros de candidemia por C. dubliniensis reportados na literatura internacional (68,114,148) , não se previa relevância desta espécie na nossa população. Os resultados obtidos (Gráfico 6) demonstraram que C. albicans correspondeu a 59,3% das espécies de Candida responsáveis por CI, em concordância com dados epidemiológicos internacionais e também nacionais (111, 118, 153) . No estudo ARTEMIS, C. albicans correspondeu a 65,6% de todos os isolados de Candida, (116) o que se encontra de acordo com - 78 - Caracterização da Candidíase Invasiva na UCI do HEM a taxa de isolamento observada no nosso estudo para esta levedura, de 72,1% (Tabela 7). Todavia, os dados do programa ARTEMIS incluem isolados de diversas amostras biológicas, mas os isolados considerados como colonização não foram incluídos na avaliação. Uma taxa de isolamento de C. albicans relativamente similar à nossa foi também observada por Mallié et al. num estudo francês multicêntrico, em que se avaliaram inúmeros tipos de amostras biológicas, pesem embora diferenças significativas na distribuição de isolamentos de espécies não-albicans, relativamente ao nosso estudo (84). Nos doentes com diagnóstico de CI, as espécies não-albicans corresponderam a 40,7% dos isolados (Gráfico 6), com particular relevância para C. glabrata, C. krusei e C. tropicalis. Não obstante, C. albicans manteve-se relevante como agente patogénico na nossa UCI. O isolamento mais frequente de C. albicans em amostras respiratórias era expectável, correspondendo a 71% de todos isolamentos em amostras respiratórias (Tabela 7). Estes dados encontram-se em concordância com os do estudo ARTEMIS (116) . Este facto pode explicar a relevância de C. albicans na nossa amostra, dado que as amostras provenientes do tracto respiratório corresponderam a 55,9% de todas as amostras recolhidas. A frequência de C. albicans em uroculturas (87,5%) não era, todavia, esperada, contrariando estudos epidemiológicos conduzidos em doentes com candidúria, que revelam taxas de isolamento entre os 20 a 70% (20) . Poderá salientar-se que C. glabrata e C. tropicalis foram também isoladas em uroculturas na nossa população, dando origem a CI de ponto de partida urinário, em concordância com diversos estudos epidemiológicos que colocam estes agentes infecciosos como causadores relevantes de candidúria, a seguir a C. albicans (20, 22). Diversos autores relacionaram a candidemia por C. glabrata com a utilização de antibióticos de largo espectro, utilização profiláctica ou empírica de fluconazol, utilização de cateter venoso central, administração de nutrição parentérica e o internamento na UCI. Outros - 79 - Caracterização da Candidíase Invasiva na UCI do HEM descreveram um risco aumentado de candidemia por C. glabrata em idades mais avançadas (40, 49, 85, 114, 118) . No estudo epidemiológico europeu conduzido pela ECMM, a incidência de CI por C. glabrata também demonstrou ser elevada em doentes cirúrgicos, embora sem se observar uma associação estatisticamente significativa (153). Seria, por isso, de esperar que C. glabrata fosse importante num ambiente de Cuidados Intensivos. De facto, a nossa população apresentou alguns dos factores supracitados. O estudo ARTEMIS revelou a importância de C. glabrata em hemoculturas e uroculturas (116) , mas C. glabrata foi isolada de forma heterogénea em amostras biológicas da nossa população, não se observando prevalência de isolamento em uroculturas e qualquer isolamento em hemocultura. Apesar destes dados, observámos uma taxa de incidência de 15,5% na subpopulação com CI, em concordância com estudos epidemiológicos internacionais (entre 10 a 25%) (84, 93, 116, 164). Paralelamente, C. parapsilosis poderia ser, também, relevante num ambiente de Cuidados Intensivos, pela facilidade de formação de biofilmes em cateteres ou outros dispositivos invasivos (22, 40) . Os dados epidemiológicos internacionais referem taxas de incidência que variam, em média, entre 4 e 15%, em diversos tipos de populações (84, 93, 116, 164). Existem, todavia, dados discordantes obtidos por González et al. que referem uma taxa de incidência muito superior, na ordem dos 37,9%, em hemoculturas (57) . Também Costa-de- Oliveira et al. revelam uma taxa de isolamento elevada em hemoculturas, de 25,6% (32). Deve frisar-se que o isolamento de C. parapsilosis no sangue tem demonstrado grande relevância na população pediátrica, onde chega a atingir taxas de incidência que podem rondar os 50% (111, 118) . Dados do estudo SENTRY reportaram que C. parapsilosis só causou infecção em 12% dos doentes com mais de 65 anos, demonstrando maior relevância na população pediátrica (118) . No nosso estudo, observou-se isolamento de espécies não-albicans, de forma heterogénea, em cateter venoso central ou linhas periféricas, na mesma proporção que C. - 80 - Caracterização da Candidíase Invasiva na UCI do HEM albicans. A taxa de isolamento de C. parapsilosis foi bastante baixa, quando comparada com alguns dos dados supracitados, mas este agente demonstrou uma taxa de incidência de 3,1% na subpopulação com CI, similar à observada noutros estudos epidemiológicos internacionais (84, 93, 116, 164) . A média de idades da nossa população também pode explicar, em parte, o reduzido papel de C. parapsilosis, observado no nosso estudo. Deve notar-se, todavia, a baixa detecção de Candida em hemoculturas, no nosso estudo. Como sabemos, o diagnóstico convencional de candidíase inclui a demonstração da invasão tecidular pelo fungo e/ou o seu isolamento em localizações anatómicas habitualmente estéreis. Infelizmente, a cultura dos fungos pode ser morosa e a microscopia directa exige experiência do operador, sendo importante encontrar meios de diagnóstico mais eficazes e precoces (128, 170) . A CI pode ocorrer com hemoculturas negativas ou que positivam tardiamente, com consequências nefastas para o doente, conforme comprovado em estudos mais antigos de Berenguer et al. e estudos mais recentes de Yera et al.. Estes investigadores demonstraram que a utilização simultânea de técnicas de centrifugação das hemoculturas ou de testes serológicos, aumenta a sensibilidade diagnóstica em doentes com hemoculturas negativas (17, 170) . Também Pfaller e Diekema reportaram diferenças nas taxas de solamento de C. glabrata consoante os sistemas ou meios de cultura utilizados para isolamento de microrganismos a partir de hemoculturas (114) . Por isso, existe já alguma variedade de métodos serológicos para Candida spp.. A detecção de ADN é uma técnica promissora, mas a metodologia ainda não se encontra padronizada. Existe risco de contaminação, devido à complexidade de execução e sensibilidade da técnica, bem como dificuldade de distinção entre colonização e infecção. Diversos autores avaliaram a utilização de técnicas combinadas, identificando que a combinação de técnicas de detecção de ADN com técnicas imunológicas conduziram a um aumento da sensibilidade e da especificidade diagnósticas. Analogamente, a detecção - 81 - Caracterização da Candidíase Invasiva na UCI do HEM combinada de anticorpos e/ou antigénios, produziu resultados muito promissores com aumento da sensibilidade e especificidade das técnicas (128, 129) . As dificuldades diagnósticas identificadas em diversos estudos encontram-se espelhadas na baixa percentagem de isolamento de Candida em hemoculturas, apesar do diagnóstico de CI em 26 doentes. Não deixa de ser interessante observar o isolamento de C. guilliermondii e C. lusitaniae, espécies consideradas raras mundialmente, numa amostra de pequenas dimensões. Todavia, nenhum destes doentes desenvolveu CI. É igualmente interessante observar uma diminuta quantidade de isolados de C. tropicalis (Tabela 7). O estudo ARTEMIS refere que C. tropicalis foi isolada frequentemente em UCIs, enfermarias médicas e serviços hematoncológicos, facto não observado no nosso estudo (116) . De facto, C. tropicalis apresenta maior relevância em doentes hematoncológicos e doentes neutropénicos (40, 114). Está documentado que, nestes doentes, a fungemia causada por C. tropicalis é responsável por um internamento mais prolongado em UCIs, quando comparado com doentes com fungemia causada por C. albicans (114) . Na nossa população não se observou a presença de doença hematoncológica concomitante. Este facto poderá ter contribuído para o isolamento modesto desta espécie. C. tropicalis não foi inclusivamente isolado em nenhum doente neutropénico da nossa população. Todavia, esta espécie foi responsável por cerca de 10% dos episódios de CI, em conformidade com dados epidemiológicos mundiais, que referem taxas de incidência na ordem dos 5% a 10% (84, 93, 116, 164). Quanto a C. krusei, o programa ARTEMIS verificou uma taxa de isolamento mais baixa, 22,5%, relativamente à observada na nossa população (116). A incidência global da infecção por este agente tem-se mantido estável ao longo dos anos (2-4%), de acordo com inúmeros estudos em populações heterogéneas (84, 85, 93, 114, 116, 164) , apesar da resistência intrínseca ao - 82 - Caracterização da Candidíase Invasiva na UCI do HEM fluconazol. Tal como C. tropicalis, C. krusei apresenta maior relevância em doentes hematoncológicos e receptores de transplantes de medula óssea (114) . Embora a nossa população não tenha revelado este tipo de co-morbilidades, C. krusei foi responsável por 12,4% dos episódios de CI. Adicionalmente, observou-se isolamento de C. krusei num doente que realizou fluconazol, como terapêutica empírica prévia, na UCI. Este doente teve dois episódios de CI por dois agentes patogénicos, C. krusei e C. albicans, após esta terapêutica, tendo sido necessário a utilização de anfotericina B lipossómica e, posteriormente, de caspofungina. Estes dados e a importância relativa de C. krusei em infecções mistas contribuíram, com certeza, para a percentagem mais elevada de CI por C. krusei constatada no nosso estudo. Este tipo de infecções têm sido descritas por diversos autores, designadamente por Tortorano et al., que descreveram 51 casos de candidemia por duas espécies de Candida, predominantemente por C. albicans + C.glabrata, no estudo europeu conduzido pela ECMM. Estes autores descreveram ainda mortalidade elevada devido a este tipo de infecções (153) . As infecções mistas foram, por isso, relevantes na nossa amostra e poderão ter estado implicada na taxa de letalidade e razão de mortalidade proporcional observadas. 1.3.3 Padrão de utilização de terapêutica antifúngica Hoje em dia, sabe-se que a terapêutica antifúngica adequada a utilizar no tratamento de infecções fúngicas invasivas é um factor preditivo de bom prognóstico (58) . A terapêutica profiláctica pode ser utilizada no contexto de Cuidados Intensivos, em doentes seleccionados, designadamente submetidos a cirurgia abdominal extensa, embora a comunidade científica permaneça dividida quanto a esta questão (58, 107, 163) . Uma meta-análise, conduzida por van Till et al. evidenciou a heterogeneidade dos estudos publicados neste campo, assim como o debate na escolha do tipo de terapêutica profiláctica mais adequada. A sua investigação - 83 - Caracterização da Candidíase Invasiva na UCI do HEM revelou redução da mortalidade e morbilidade relacionadas com infecções fúngicas com a utilização de terapêutica profiláctica, contudo, a efectividade da terapêutica profiláctica, nomeadamente a descontaminação do tracto gastro-intestinal, foi inversamente proporcional ao número de doentes cirúrgicos incluídos (157) . A utilização de terapêuticas empírica e pré- emptiva permanecem controversas, embora existam nichos de utilização do primeiro tipo de terapêutica, em doentes neutropénicos (58, 107). Os conceitos, apesar de diferentes, são de difícil distinção na prática diária. Tal facto conduziu à classificação do tipo de terapêutica utilizada, recorrendo somente ao conceito de terapêutica empírica, conforme especificado pelas justificações clínicas introduzidas na prescrição informatizada dos doentes ou preenchimento do Anexo I. Quanto à terapêutica documentada, o algoritmo proposto por Spellberg indica que a escolha do antifúngico deve basear-se no estado hemodinâmico do doente. Em doentes hemodinamicamente estáveis, sem disfunção de orgãos, o fluconazol pode ser considerado como terapêutica de primeira linha. No entanto, a utilização deste fármaco deve ser preterida caso haja exposição recente a um azol, se verifique colonização prévia por espécies resistentes e/ou caso a epidemiologia geográfica indique elevadas taxas de infecções por leveduras resistentes ao fluconazol. Pelo contrário, em doentes hemodinamicamente instáveis, a utilização de equinocandinas parece ser a alternativa mais adequada (58, 107) . Em doentes neutropénicos, as equinocandinas, designadamente a caspofungina e as formulações lipídicas de anfotericina B ganham um papel de destaque, embora a utilização de fluconazol seja plausível (61, 107). Como tal, o padrão de utilização de antifúngicos na nossa população (ver Tabela 8) encontrase de acordo com o descrito nas recomendações internacionais por painéis de peritos, sendo o fluconazol o antifúngico seleccionado mais frequentemente, seguido da caspofungina, e por - 84 - Caracterização da Candidíase Invasiva na UCI do HEM último o voriconazol e anfotericina B lipossómica. Adicionalmente, dos oito cursos de antifúngico descritos com caspofungina, seis foram utilizados em doentes em sepsis, ou seja doentes hemodinamicamente instáveis. A caspofungina foi ainda utilizada mais frequentemente como terapêutica empírica e o fluconazol como terapêutica dirigida. A utilização de terapêutica empírica foi muito frequente (48,6% dos casos de terapêutica instituída), facto corroborado pelo número de doentes com instituição de terapêutica sem diagnóstico confirmado de CI (18,8% dos doentes). Todavia, esta terapêutica foi instituída num número reduzido de doentes neoplásicos (n=3 de um universo de 10 doentes). Por outro lado, o diagnóstico de candidíase comprovou-se em 10 dos 18 dos doentes (55,5%) que iniciaram terapêutica empírica. Se pensarmos que um atraso na terapêutica, por dados microbiológicos indisponíveis, por exemplo, pode resultar num aumento de mortalidade, conforme observado em alguns estudos epidemiológicos (110, 172) , facilmente se explica esta atitude terapêutica. A elevada percentagem de terapêutica empírica instituída no nosso estudo, com confirmação posterior de diagnóstico de candidíase leva-nos, por isso, a inferir que a terapêutica antifúngica foi instituída atempadamente. O tempo médio desde os resultados microbiológicos e início de terapêutica dirigida/documentada não foi, todavia, determinado. A terapêutica profiláctica foi instituída em reduzida percentagem com fluconazol, nomeadamente em 5% dos doentes, tendo sido seleccionada num doente neutropénico e noutro doente para descontaminação intestinal, embora não estivesse descrita história prévia de cirurgia. Nenhum dos dois doentes desenvolveu estirpes resistentes nem espécies nãoalbicans sob terapêutica com fluconazol, não alimentando as preocupações da comunidade científica sobre a possibilidade de emergência de estirpes resistentes ao fluconazol e de espécies não-albicans, em doentes submetidos a profilaxia (114, 157). - 85 - Caracterização da Candidíase Invasiva na UCI do HEM O padrão de utilização de antifúngicos descrito na nossa população difere de alguns estudos epidemiológicos anteriores, nomeadamente na América do Sul (Brasil), salientando-se, contudo, o período deste estudo (2003 a 2004) (31) . O tempo médio de dias de terapêutica descrito neste estudo, 14 dias, é similar ao observado no nosso estudo, 13,9 dias. Relativamente ao estudo desenvolvido por Costa-de-Oliveira et al., é necessário salientar o tipo de amostra estudada, que difere substancialmente da nossa população. Nesse estudo, a utilização de terapêutica profiláctica foi muito mais frequente, tendo-se observado sobretudo em doentes neoplásicos. A utilização de fluconazol foi prevalente, mas a caspofungina foi utilizada em muito menor percentagem. Será necessário realçar que esse estudo abrange todos os serviços clínicos de um hospital no Porto, pelo que não é exequível uma comparação com a nossa população, exclusivamente de UCIs (32). Zilberberg et al. descreveram um padrão de utilização de antifúngicos numa UCI, semelhante ao constatado no nosso estudo, enfatizando-se, todavia, a percentagem acrescida de caspofungina administrada no estudo supracitado, quando comparado com o observado no nosso estudo. Estes autores verificaram que a escolha do antifúngico foi maioritariamente apropriada, mas que existiram problemas a nível das doses de triazóis seleccionadas e de atraso de mais de 24h na instituição de terapêutica antifúngica (172). Também Massanet et al. avaliaram recentemente a compliance da prática clínica com as recomendações internacionais através da análise dos padrões de utilização de antifúngicos em 9 UCIs. Os resultados foram similares aos descritos por Zilberberg, identificando-se a não descalação da terapêutica e a utilização de fluconazol em doentes expostos previamente a azóis, como potenciais problemas (87). Estes autores denotaram uma utilização mais acentuada de equinocandinas, nomeadamente caspofungina, relativamente ao observado no nosso estudo. - 86 - Perfil de susceptibilidade aos antifúngicos 2. Perfil de susceptibilidade aos antifúngicos 2.1 Materiais e Métodos 2.1.1 Agentes antifúngicos Os antifúngicos foram obtidos através das indústrias farmacêuticas produtoras dos mesmos: os pós de fluconazol, voriconazol e anidulafungina dos Laboratórios Pfizer, o pó de caspofungina dos Laboratórios Merk & Co.. A anfotericina B foi obtida através da compra de Fungizone® - preparação de anfoterina sódica deoxicolato para administração intravenosa. 2.1.2 Preparação do inóculo de leveduras O inóculo foi preparado, a partir 5 colónias com ≥1 mm de diâmetro. Seguidamente, foram suspensas em 5mL de solução salina. Procedeu-se à homogeneização durante 15s e ajustou-se espectrofotometricamente ao padrão de 0,5 McFarland (correspondente a 0,125 de densidade óptica), num comprimento de onda de 530 nm. Os inóculos foram utilizados até duas horas após a sua preparação, tendo sido conservados a 4ºC. 2.1.3 Método Microdiluição Meios de cultura O meio RPMI 1640 é o recomendado neste tipo de técnica, tendo sido produzido de acordo com a constituição descrita no documento M27–S3 do CLSI, com glutamina, fenol e sem bicarbonato de sódio. O meio é tamponado com ácido N-morfolino propanosulfónico (MOPS), devendo assegurar-se um pH entre 6,9 e 7,1 até à concentração final de 0,165mol/L, à temperatura ambiente (29) . No caso dos testes de susceptibilidade à anfotericina B, recorremos ao meio AM3, também de acordo com as recomendações do CLSI - 87 - (26, 29, 82, 136) . Perfil de susceptibilidade aos antifúngicos Ambos os meios, produzidos no Laboratório de Micologia da Faculdade de Farmácia, foram suplementados com glucose até uma concentração final de 20g/L, permitindo melhor visibilidade aquando da leitura das microplacas (26, 29). Preparação das soluções-mãe de antifúngicos A preparação das soluções-mãe e intermediárias de antifúngicos considerou a actividade dos antifúngicos em pó ou em solução cedidos pelos laboratórios produtores, através da seguinte fórmula: Peso (mg)= Volume desejado (mL)x Concentração desejada (µg/mL) Actividade antifúngico (µg/mL) O fluconazol e a caspofungina foram diluídos em água destilada estéril, enquanto as soluções de anfotericina B, voriconazol e anidulafungina foram preparadas com DMSO (26, 29) . A preparação da solução-mãe de anfotericina B teve um passo adicional: ressupensão do pó da ampola de Fungizone® em água e diluição posterior a uma concentração de 1,6mg/mL (149). As soluções-mãe de fluconazol e caspofungina estavam 10x concentradas, relativamente às concentrações requeridas para a técnica de microdiluição. Pelo contrário, as substâncias com baixa solubilidade em água (anfotericina B, voriconazol e anidulafungina), requereram concentrações superiores, nomeadamente 100x, relativamente às concentrações requeridas nos ensaios, de forma a evitar possíveis precipitações em diluições subsequentes. As soluções-mãe foram conservadas a -60ºC, por um período de seis meses, em frascos esterilizados de polietileno ou polipropileno. Não foi realizada filtração, pois seria necessário assegurar que não ocorreria adsorção dos antifúngicos ao filtro utilizado (26, 29). As diluições subsequentes procederam-se em meio RPMI 1640 para todos os antifúngicos, excepto anfotericina B, cuja solução foi diluída em meio AM3. Para os antifúngicos - 88 - Perfil de susceptibilidade aos antifúngicos fluconazol e caspofungina procedeu-se a uma diluição de 1:5. Para os antifúngicos voriconazol, anfotericina B e anidulafungina procedeu-se a 2 diluições consecutivas - 1:10 e 1:5. Preparação das microplacas (26, 29) As microplacas possuem 96 poços de fundo em U. Com uma micropipeta, procedeu-se à distribuição de 100µL de meio RPMI ou AM3 nas cúpulas 2 a 12. Posteriormente, adicionouse à cúpula 1 200µL de solução de antifúngico preparada anteriormente. Fizeram-se diluições 1:2 até à cúpula 10, rejeitando-se 100µL. O poço 1 continha a concentração mais elevada de antifúngico e o poço 10 a mais baixa. Os poços 11 e 12 serviram de controlo de crescimento das leveduras e de esterilidade do meio, respectivamente. Inoculação das microplacas (26, 29) Primeiro, a suspensão do inóculo foi diluída 1:50, seguida de uma diluição de 1:20 em meio RPMI 1640, o que resultou numa concentração de 1x103 a 5x103 UFC/mL. De seguida, procedeu-se à inoculação das microplacas com 100 µL de inóculo. Tal resultou numa diluição de 1:2 na concentração do inóculo, entre 0,5x103 a 2,5x103 UFC/mL. Para avaliação da anfotericina B utilizou-se o meio AM3. Leitura das microplacas (26, 29) A incubação realizou-se a 35ºC durante 24h a 48h dependendo do antifúngico testado. As microplacas foram lidas com a ajuda de um espelho de leitura. O crescimento em cada poço foi comparado com o poço de controlo de crescimento (poço 11), definindo-se uma pontuação numérica, de acordo com a seguinte escala: 0 opticamente limpa; 1 ligeiramente turva; 2 grande decréscimo na turvação (aproximadamente 50% de turvação); 3 ligeira redução na turvação; 4 ausência de redução na turvação. Para a anfotericina B, a CIM é definida como a mais baixa concentração em que há ausência de crescimento (0), o que corresponde a uma - 89 - Perfil de susceptibilidade aos antifúngicos inibição de 100%. A leitura foi realizada às 24h. No caso do azóis, a determinação das CIMs pode ser menos nítida e está definida como a mais baixa concentração de antifúngico que origina um score 2 ou seja grande decréscimo na turvação (aproximadamente 50% de inibição). A utilização desta escala permite a distinção entre isolados resistentes e de susceptibilidade intermédia. A leitura do teste de susceptibilidade ao fluconazol e voriconazol foi realizada às 48h. Quanto às equinocandinas, a leitura foi realizada às 24h. A CIM encontra-se definida como a mais baixa concentração de antifúngico que origina um score 2. Critérios de Interpretação Para classificação dos isolados, quanto à susceptibilidade in vitro aos antifúngicos testados, recorremos aos pontos de corte estabelecidos pelo CLSI no documento M27-A3 e descritos abaixo na Tabela 11. Tabela 11. Critérios de Interpretação de acordo com CLSI para o método de microdiluição (26, 29) Intervalos das CIMs (µg/mL) Susceptível dose dependente Resistente Antifúngico Susceptível Fluconazol ≤8 16-32 ≥64 Voriconazol ≤1 2 ≥4 Caspofungina ≤2 >2 Anidulafungina ≤2 >2 Não susceptível Para a anfotericina B não se encontram definidos quaisquer valores críticos, sugerindo-se contudo a utilização do ponto de corte de 1µg/mL. Assim, uma CIM<1µg/mL sugere um isolado potencialmente sensível, enquanto uma CIM1µg/mL sugere um isolado resistente (26, 29) . - 90 - Perfil de susceptibilidade aos antifúngicos 2.1.4 Método de difusão em discos Meios de cultura O meio Mueller-Hinton é o recomendado neste tipo de técnica, de acordo com o documento M44-A do CLSI, tendo sido suplementado com glucose até uma concentração final de 20g/L e um pH entre 7,2 e 7,4. O meio foi ainda suplementado com azul de metileno a uma concentração de 0,5µg/mL. Esta operação permite melhor visibilidade aquando da leitura dos diâmetros. As placas de ágar utilizadas possuíam um diâmetro de 90 mm e uma profundidade de 4 mm (27, 28). O meio foi preparado no Laboratório de Micologia da Faculdade de Farmácia. Discos de antifúngicos Foram obtidos discos impregnados com anfotericina B (100µg), fluconazol (25µg) e voriconazol (1µg) (Bio-Rad Laboratories, Amadora, Portugal) e foram acondicionados no frigorífico à temperatura ≤8ºC, de acordo com o laboratório fabricante, até ao prazo de validade das embalagens. Inoculação das placas de ágar Para inoculação da placa, procedeu-se de acordo com o preconizado para o método Etest® (Folheto Informativo Etest®, AB BIODISK, Solna, Suécia) que consiste na impregnação de uma zaragatoa no inóculo de leveduras previamente preparado. A zaragatoa deve estar humedecida, sem excesso de líquido. A placa de ágar Mueller-Hinton foi inoculada recorrendo à aplicação da zaragatoa por toda a superfície da placa, com formação de estrias. O processo foi repetido mais duas vezes rodando a placa de ágar cerca de 60º, de forma a assegurar uma distribuição uniforme do inóculo. Por último, foi aplicada a zaragatoa nos bordos da placa de ágar, descrevendo uma circunferência. De seguida, foram aplicados os discos impregnados com os agentes antifúngicos, com auxílio de uma pinça, não se - 91 - Perfil de susceptibilidade aos antifúngicos excedendo a aplicação de cinco discos por placa de 90 mm. Procedeu-se à incubação das placas a 35-37ºC, durante um período de 24h. Leitura dos diâmetros A leitura dos diâmetros realizou-se de acordo com o documento M44–S2, considerando-se 80% de inibição de crescimento para aferição do diâmetro de inibição para os fármacos fluconazol e voriconazol ( 27, 28). Não existem pontos de corte definidos para a anfotericina B, pelo que se utilizaram diversas referências bibliográficas, para definição dos critérios de interpretação (100, 139). Critérios de interpretação Os critérios de intrepretação encontram-se mencionados na Tabela 12. Tabela 12. Critérios de Interpretação de acordo com CLSI para método de difusão em discos, Negri et al. e Saubolle et al. (27, 28, 100, 139) Zona de diâmetro (mm) Antifúngico Susceptível Susceptível dose dependente Resistente Anfotericina B >10 - ≤10 Fluconazol ≥19 15-18 ≤14 Voriconazol ≥17 14-16 ≤13 2.1.5 Etest® Meios de cultura O meio RPMI 1640 é um dos recomendados neste tipo de técnica, tendo sido produzido de acordo com a constituição descrita no documento M27–S3 do CLSI. No caso do teste de susceptibilidade à anfotericina B, recorremos ao meio AM3. Ambos os meios foram - 92 - Perfil de susceptibilidade aos antifúngicos produzidos no Laboratório de Micologia da Faculdade de Farmácia e suplementados com glucose até uma concentração final de 20g/L e 15% de ágar. Acondicionamento das tiras teste O acondicionamento das tiras teste realizou-se de acordo com as recomendações do fabricante (Folheto Informativo Etest®, AB BIODISK, Solna, Suécia), a -20ºC, até ao prazo de validade das embalagens. Inoculação das placas de ágar Para inoculação da placa, procedeu-se de de acordo com o preconizado para o método Etest®, (Folheto Informativo Etest®, AB BIODISK, Solna, Suécia) e descrito anteriormente para o método de difusão em discos. Procedeu-se à aplicação das tiras Etest® na superfície da placa, com auxílio de uma pinça. Procedeu-se à incubação das placas a 35ºC, durante um período de 24-48h. Leitura das elipses de inibição A leitura das elipses de inibição realizou-se de acordo com as instruções do fabricante (Folheto Informativo Etest®, AB BIODISK, Solna, Suécia), considerando-se a CIM como a concentração obtida quando a elipse de inibição e crescimento intercepta a tira de Etest®. Para a anfotericina B, a CIM foi lida no ponto de uma inibição de 100% de crescimento. Para os azóis, caspofungina e anidulafungina a CIM foi lida no primeiro ponto de inibição significativa/decréscimo importante no crescimento, considerando 80% de inibição de crescimento. A leitura das CIMs realizou-se às 24h ou 48h, consoante as espécies de Candida. - 93 - Perfil de susceptibilidade aos antifúngicos Critérios de Interpretação Os critérios de interpretação são idênticos aos utilizados para o método de microdiluição tabela 13. Tabela 13. Critérios de Interpretação de acordo com CLSI (26,29) Intervalos das CIMs (µg/mL) Susceptível dose dependente Resistente Antifúngico Susceptível Fluconazol ≤8 16-32 ≥64 Voriconazol ≤1 2 ≥4 Caspofungina ≤2 >2 Anidulafungina ≤2 >2 Não susceptível Para a anfotericina B não se encontram definidos quaisquer valores críticos, sugerindo-se, contudo, a utilização do ponto de corte 1µg/mL. Assim, uma CIM<1µg/mL sugere um isolado potencialmente sensível, enquanto uma CIM1µg/mL sugere um isolado resistente (26, 29). 2.1.6 Controlo de Qualidade Selecção das estirpes de referência As estirpes de referência foram seleccionadas de acordo com as recomendações dos documentos M27–S3, M44-A e recomendações do fabricante (Folheto Informativo Etest®, AB BIODISK, Solna, Suécia), considerando os antifúngicos testados. Assim, o controlo de qualidade foi realizado com as estirpes Candida parapsilosis ATCC 22019, Candida krusei 6258, e recorreu-se à estirpe de referência Candida albicans ATCC 90028 (26, 27). Para alguns fármacos, não existem intervalos de valores definidos para avaliação de estirpes de referência. O controlo com a estirpe de referência C. albicans ATCC 90028 para a anfotericina B, pelo método de microdiluição, foi realizado às 24h. Contudo, o intervalo de CIMs para esta estirpe só se encontra documentado para um período de incubação de 48h - 94 - (29) . Utilizou-se também a Perfil de susceptibilidade aos antifúngicos estirpe de referência C. lusitaniae ATCC 200950. A utilização deta estirpe de referência é sugerida pelo CLSI, quando se utiliza o meio AM3, para avaliação da reprodutibilidade e adequabilidade do meio (26). Acondicionamento das estirpes de referência As estirpes de referência foram subcultivadas em meio Sabouraud dextrose ágar suplementado com cloranfenicol, a 35ºC durante 24-48h. Para um armazenamento por um período de tempo reduzido, as estirpes foram mantidas entre 2 a 8ºC. Para um armazenamento prolongado, recorreu-se à congelação das estirpes a -70ºC em meio Saboraud líquido suplementado com 15% de glicerol. Antes do armazenamento, comprovou-se a viabilidade e pureza destas após incubação de 24h (26). Teste das estirpes de referência e de controlo de qualidade Recorreu-se aos procedimentos atrás descritos (microdiluição, teste de difusão em discos e Etest®) após subcultura a partir das estirpes armazenadas (incubação 24-48h), com a finalidade de avaliar a manutenção das suas CIMs. 2.1.7 Tratamento de dados e análise estatística Para tratamento dos dados microbiológicos, analisou-se a CIM50, CIM90, intervalo de CIMs e percentagem de isolados resistentes (57, 164) . Avaliou-se a distribuição de espécies pelas diferentes amostras biológicas e a concordância de resultados entre os métodos utilizados. Para esta última análise, recorreu-se aos valores interpretativos descritos pelo CLSI, determinando-se o acordo categórico (%) entre método de difusão em disco/ método Etest® e os resultados obtidos pelo método de microdiluição (padrão) (100) . Considerou-se ainda a percentagem de concordância dos resultados obtidos pelos diferentes métodos (Etest® e método de microdiluição) em duas diluições. Definiram-se erros major (EM) como isolados - 95 - Perfil de susceptibilidade aos antifúngicos considerados resistentes pelo método difusão em disco/Etest® e considerados susceptíveis pelo método de microdiluição. Erros muito graves (EMG) definiram-se como isolados considerados sensíveis pelo teste de difusão em disco/ Etest® e resistentes pelo método de microdiluição. Erros minor (M) ocorreram quando o resultado de um dos testes foi sensível ou resistente, e do outro teste foi susceptível dose dependente (100, 130). - 96 - Perfil de susceptibilidade aos antifúngicos 2.2 Resultados Das 68 amostras recebidas foram analisadas 60. Tal deveu-se ao facto de algumas amostras terem sido incorrectamente identificadas/enviadas ou por não ter sido possível, pela deterioração da amostra, realizar qualquer tipo de determinação de susceptibilidade. Relativamente à susceptibilidade aos antifúngicos dos isolados estudados, resistência ou susceptibilidade dose-dependente foi observada raramente. A resistência ao fluconazol ocorreu numa percentagem minoritária dos isolados, nomeadamente em isolados de C. krusei e C. glabrata. Todavia, a percentagem de isolados resistentes diferiu consoante o teste de susceptibilidade utilizado (Tabelas 15 e 16). Não se observou qualquer isolado resistente ao voriconazol, incluindo isolados resistentes ao fluconazol, ou à anfotericina B. O mesmo não poderá ser dito relativamente às equinocandinas estudadas, observando-se CIMs elevadas em isolados de C. parapsilosis e C. guilliermondii. Não deixa, contudo, de ser pertinente referir a ―resistência‖ cruzada (não susceptibilidade) observada no isolado de C. guilliermondii, de acordo com o método Etest®. Os resultados relativos aos isolados de C. parapsilosis diferiram consoante o teste de susceptibilidade utilizado, evidenciando-se, não obstante, CIMs elevadas para ambas as equinocandinas. No caso da anfotericina B e do voriconazol, 90% das espécies de Candida apresentaram CIMs ≤0,25µg/mL, pelo método preconizado pelo CLSI. 90% das espécies de Candida foram inibidas por concentrações de anidulafungina ≤0,063µg/mL. Pelo contrário, a caspofungina apresentou uma CIM90 ≤0,5µg/mL. Considerando cada uma das espécies, C. albicans revelou CIMs90 ≤0,5µg/mL, ≤0,125µg/mL, ≤0,25µg/mL, ≤0,25µg/mL e ≤0,063µg/mL para fluconazol, voriconazol, anfotericina B, caspofungina e anidulafungina, respectivamente. As espécies C. krusei e C. glabrata apresentaram prevalentemente CIMs mais elevadas ou idênticas a 0,25µg/mL para o - 97 - Perfil de susceptibilidade aos antifúngicos voriconazol, se considerarmos o método de microdiluição. Pelo método Etest®, foram observadas CIMs mais elevadas no caso de C. glabrata, com CIM ≥0,75µg/mL em 3 dos 4 isolados. Estes isolados apresentaram, paralelamente, resistência ou susceptibilidade dosedependente ao fluconazol. Através deste método, C. krusei revelou CIM=0,5µg/mL contra o voriconazol, em 2 dos 3 isolados testados. A caspofungina evidenciou CIMs mais elevadas que a anidulafungina, especialmente considerando as espécies não-albicans C. glabrata e C. krusei, não excedendo, todavia, CIMs de 0,5µg/mL para ambas as espécies. A anfotericina B revelou CIMs≤0,125µg/mL em espécies não-albicans, como C. tropicalis, C. parapsilosis, C. guilliermondii e C. glabrata. O controlo de qualidade realizado encontra-se resumido na Tabela 14, com cálculo de valores médios. Tabela 14. Susceptibilidade in vitro de 4 estirpes de Candida spp. para controlo de qualidade (28, 29) Estirpes Testadas Anfotericina B Método Microdiluição CIM µg/mL* Método Difusão em Disco Diâmetro em mm* Candida parapsilosis ATCC 22019 0,75(24h) Candida krusei ATCC 6258 0,38 (24h) Candida albicans ATCC 90028 0,25 (24h) Fluconazol 1 16 Voriconazol 0,03 0,25 Caspofungina 0,5 (24h) 0,25 (24h) Anidulafungina 1 (24h) 0,12 (24h) Fluconazol - - 30 Voriconazol - 24 32 * Valores Médios - 98 - 0,5 Candida lusitaniae ATCC 200950 0,75 (24h) Perfil de susceptibilidade aos antifúngicos Tabela 15. Susceptibilidade in vitro de Candida spp. através de 3 métodos: Microdiluição, Etest®, Difusão em Disco MD (µg/mL) Espécie Antifúngico Todas espécies (n=60) Fluconazol Voriconazol Caspofungina Anidulafungina Anfotericina B C. albicans (n=44) Fluconazol Voriconazol Caspofungina Anidulafungina Anfotericina B ET (µg/mL) DD (mm) Intervalo R 50% 90% Intervalo R 50% 90% Intervalo R 0,125-64 0,03-0,5 0,016-4 0,016-2 0,03-0,5 2 0 1 0 0 0,25 0,03 0,125 0,016 0,063 8 0,25 0,5 0,063 0,25 0,25-256 0,012-1 0,023-32 0,002-32 0,002-0,5 4 0 1 4 0 1 0,038 0,125 0,006 0,19 24 0,5 0,5 0,125 0,38 17-43 22-44 0 0 14-24 0 0,125-1 0,03-0,25 0,016-0,5 0,016-0,125 0 0 0 0 0 0 0 0 0 1,5 0,19 0,25 0,012 0,25 0 0 0,002-0,38 0,75 0,032 0,125 0,004 0,19 23-43 29-44 0 0,5 0,125 0,25 0,063 0,25 0,25-2 0,012-0,75 0,023-0,38 0,002-0,064 0,03-0,5 0,125 0,03 0,125 0,016 0,063 4-64 0,063-0,5 0,063-0,5 0,03-0,063 0,063-0,125 1 0 0 0 0 3-64 0,094-1 0,032-0,5 0,006-0,032 0,19-0,38 1 0 0 0 0 17-38 28-43 0 0 15-19 0 32-64 0,125-0,5 0,25-0,5 1 0 0 3 0 0 17-18 22-31 0 0 Anidulafungina 0,016-0,063 0 Anfotericina B C. lusitaniae (n=2) Fluconazol Voriconazol Caspofungina Anidulafungina Anfotericina B C..parapsilosis (n=3) Fluconazol Voriconazol Caspofungina Anidulafungina Anfotericina B C. tropicalis (n=3) Fluconazol Voriconazol Caspofungina Anidulafungina Anfotericina B 0,063-0,25 0 64-256 0,038-1 0,125-0,5 0,0060,125 0,19-0,5 0 14-19 0 0,5-2 0,03-0,03 0,063-0,5 0,063-0,063 0,063-0,125 0 0 0 0 0 1,5-2 0,023-0,023 0,25-0,25 0,016-0,064 0,064-0,125 0 0 0 0 0 27-35 32-39 0 0 17-17 0 0,5-1 0,03-0,03 0,5-4 0,5-1 0,03-0,063 0 0 1 0 0 0,75-2 0,023-0,096 0,38-0,75 3-8 0,125-0,25 0 0 0 3 0 23-35 28-43 0 0 15-21 0 0,125-2 0,03-0,5 0,016-0,125 0,016-0,063 0,03-0,03 0 0 0 0 0 0,75-1 0,032-0,75 0,032-0,38 0,008-0,016 0,064-0,125 0 0 0 0 0 27-31 27-29 0 0 15-16 0 8 0,25 2 2 0,03 0 0 0 0 0 8 0,38 ≥32 ≥32 0,047 0 0 1 1 0 22 22 0 0 18 0 C. glabrata (n=4) Fluconazol Voriconazol Caspofungina Anidulafungina Anfotericina B C. krusei (n=3) Fluconazol Voriconazol Caspofungina 14-24 0 0 C. guilliermondii (n=1) Fluconazol Voriconazol Caspofungina Anidulafungina Anfotericina B - 99 - Perfil de susceptibilidade aos antifúngicos Torna-se ainda pertinente a análise de isolados distribuídos pela categoria de susceptibilidade consoante o teste de susceptibilidade utilizado, bem como a concordância de resultados entre os métodos utilizados (Tabela 16). Tabela 16. Percentagem de Isolados distribuidos pela categoria de susceptibilidade Isolados (%) Discrepância (N) Antifúngico Método S SDD R Fluconazol MD 88,34a 6,67 3,34 ET 90 3,34 DD 93,34 MD Concordância Categórica Concordância (%) 2 diluições (%) EMG EM M 6,67 0 0 2 96,67 66,67 6,67 0 1 0 2 95 NA 98,34a 0 0 ET 98,34a 0 0 0 0 0 100 88,34 DD 100 0 0 0 0 0 100 NA Anfotericina B MD 100 0 0 B ET 100 0 0 0 0 0 100 80 DD 100 0 0 0 0 0 100 NA MD 98,34 - 1,67 ET 98,34 - 1,67 1 1 - 98,34 73,34 DD - - - - - - NA NA MD 100 - 0 ET 93,34 - 6,67 0 4 0 93,34 68,34 DD - - - _ _ _ NA NA Voriconazol Caspofungina Anidulafungina (a) Num isolado de C. albicans não foi possível determinar a CIM para o Fluconazol e Voriconazol de acordo com método de diluíção. Neste mesmo isolado não foi possível determinar a CIM para o Voriconazol pelo método Etest®; Facilmente observamos diferenças relevantes entre os dois métodos Etest® e teste de difusão em discos, quando comparamos os resultados com o método de microdiluição. O Etest® e o teste de difusão em discos apresentaram uma concordância categórica superior a 93,34% para todos os fármacos testados. Ambos os testes apresentaram 100% de concordância categórica relativamente ao método padrão, em relação aos fármacos voriconazol e anfotericina B. - 100 - Perfil de susceptibilidade aos antifúngicos Considerando a concordância em duas diluições, as divergências do Etest® relativamente ao método padronizado pelo CLSI aumentam. Assim, a percentagem de concordância decai para 66,6% no que concerne o fluconazol. A concordância entre os dois métodos é superior a 80% para outros antifúngicos testados, voriconazol e anfotericina B, mas não demonstra resultados similares nas equinocandinas, revelando percentagens de concordância idênticas à observada para o fluconazol. Importa ainda referir o tipo de erros detectados, quando comparados os Etest® e teste de difusão em discos com o método de microdiluição. EMG foram observados infrequentemente (n=2), nomeadamente na avaliação do fluconazol através do teste de difusão em discos e na avaliação da susceptibilidade às equinocandinas pelo Etest®. Os EM e M foram igualmente infrequentes (n=5/n=4), tendo-se observado EM na avaliação da susceptibilidade às equinocandinas pelo Etest® e M na avaliação do fluconazol através dos dois métodos não padronizados pelo CLSI. - 101 - Perfil de susceptibilidade aos antifúngicos 2.3 Discussão 2.3.1 Susceptibilidade aos antifúngicos Diversos estudos epidemiológicos, como o estudo SENTRY e programa ARTEMIS, identificaram a manutenção de taxas de actividades muito elevadas (>90%) do fluconazol contra grande parte das espécies de Candida (93, 116) . Observou-se, porém, um decréscimo de susceptibilidade (75%) nas espécies C. glabrata, C. guilliermondii, C. rugosa e C. norvegensis (116). As taxas de resistência de C. glabrata ao fluconazol diferem de acordo com áreas geográficas, oscilando acima dos 16% (114, 116) . Conforme reportado por diversos autores, C. krusei apresentou, surpreendentemente, taxas de susceptibilidade (isolados S-DD) entre 30% a 70%, sensivelmente (92, 96, 116). C. albicans tem permanecido altamente sensível a este antifúngico (31, 92, 93, 96, 116) . Esperava-se, por isso, que o fluconazol demonstrasse actividade contra grande parte das espécies de Candida, com taxas de susceptibilidades superiores a 90%, excepto em possíveis isolados de C. krusei, C. glabrata, C. guilliermondii, C. rugosa e C. norvegensis. De facto, o fluconazol demonstrou, no nosso estudo, enorme robustez, com uma taxa de resistência global reduzida, de 3,34%, de acordo com o método padronizado pelo CLSI. Os isolados resistentes pertenceram às espécies C. glabrata e C. krusei. É de reforçar, que, tal como identificado por diversos autores (92, 96, 116) , C. krusei apresentou CIMs consideradas dose-dependente, em 66,6% dos casos, de acordo com o método de microdiluição. O método Etest® considerou todos os isolados resistentes e o método de difusão em discos considerou os 3 isolados susceptível dose-dependente ao fluconazol. A resistência de C. krusei ao fluconazol, considerada intrínseca, aparenta ser mediada por diminuição da afinidade da enzima alvo, lanosterol 14-α-desmetilase, ao agente antifúngico. - 102 - Perfil de susceptibilidade aos antifúngicos Encontram-se descritos outros mecanismos de resistência (104). Não se exclui, hoje em dia, que estes mecanismos possam ocorrer em estirpes S-DD (96) . Assim, a validade prática dos testes de susceptibilidade contra o fluconazol, em C. krusei, é reduzida. Numa análise mais detalhada da susceptibilidade a este fármaco, verificamos algumas diferenças em relação a parâmetros como CIM90 e valores modais de CIMs descritos na literatura. Matar et al. descreveram CIM90 para C. albicans na ordem dos 0,25µg/mL contra o fluconazol pelo método de microdiluição, similar ao observado no nosso estudo, de 0,5µg/mL (Tabela 15). Os mesmos autores descreveram CIM90, para o mesmo agente, de 2µg/mL pelo método Etest®. Observámos, neste caso, valores idênticos para o Etest® (1,5µg/mL) (88). O panorama de susceptibilidade ao voriconazol é muito favorável, com taxas de susceptibilidade superiores a 83%, de acordo com o reportado no programa ARTEMIS, se considerarmos C. glabrata e C. krusei, e superior a 90%, sem contar com estas espécies (116). Nas espécies resistentes ao fluconazol, tem-se constatado mundialmente uma susceptibilidade ao voriconazol superior a 75% nos casos de C. krusei, C. norvegensis e C. dubliniensis, mas actividade mais reduzida contra outras espécies como C. albicans, C. glabrata, C. tropicalis e C. rugosa (116) . Mallié et al. reportaram susceptibilidade muito favorável de C. glabrata ao voriconazol, na ordem dos 71% a 99%, conforme se tratavam de isolados resistentes ou S-DD ao fluconazol (84). Previa-se, por isso, um óptimo perfil de susceptibilidade na maior parte das espécies de Candida e não foi encontrado nenhum isolado resistente a este antifúngico, de acordo com os 3 métodos realizados (Tabela 15). Matar et al. e Quindós et al. descreveram CIM90 na ordem dos 0,03µg/mL para Candida albicans, contra o voriconazol, pelo método de microdiluição, um pouco aquém do observado no nosso estudo (0,125µg/mL), mas similar ao observado por - 103 - Perfil de susceptibilidade aos antifúngicos Pfaller et al., em estudos mais antigos (88, 132) . Os mesmos autores (Matar et al.) descreveram CIM90 para o mesmo agente de 0,047µg/mL pelo método Etest® (88) . Pelo contrário, observámos valores significativamente mais altos para o Etest® (0,19µg/mL) na nossa amostra. Ainda pelo método Etest®, observaram-se CIMs mais elevadas no caso de C. glabrata, com CIM ≥0,75µg/mL em 3 dos 4 isolados analisados, de forma idêntica ao constatado por Maillé et al., e similar à CIM90 apresentada por Quindós et al. (1µg/mL), na sua meta-análise (84, 132) . Estes isolados apresentaram paralelamente resistência ou susceptibilidade dose-dependente ao fluconazol. Ao contrário do relatado por Panackal et al. e Maillé et al., não observámos nenhum isolado de C. glabrata com CIM≥2µg/mL (84, 105). A produção de bombas de efluxo activas tem sido implicada como mecanismo de resistência cruzada ao fluconazol e voriconazol. Sabe-se que C. glabrata consegue criar bombas de efluxo dependentes de ATP, para a expulsão de antifúngicos, e que a exposição de C. glabrata a concentrações sub-terapêuticas de fluconazol pode resultar em resistência cruzada entre os diversos derivados triazólicos (33, 62) . Dos cinco doentes com isolamento de C. glabrata, constatou-se, em dois casos, história de terapêutica antifúngica prévia, nomeadamente fluconazol (Tabela 10). As CIMs destes isolados contra o fluconazol foram de 4 e 16µg/mL, susceptível e susceptível dose-dependente, respectivamente. Ambos os doentes tiveram um episódio de CI, com necessidade de nova terapêutica antifúngica. Estes isolados apresentaram CIMs de 0,063 e 0,25µg/mL, contra o voriconazol pelo método de microdiluição, respectivamente. Por seu turno, o Etest® revelou uma CIM mais elevada, de 0,75µg/mL, para o isolado S-DD ao fluconazol. Não se observou, por isso, qualquer tipo de resistência cruzada, sendo significativo, todavia, a detecção de CIMs mais elevadas de C. glabrata para o voriconazol, pelo método Etest®, iguais ou superiores à CIM90 para todas as espécies testadas (Tabela 15). - 104 - Perfil de susceptibilidade aos antifúngicos Para C. krusei, constatou-se um intervalo de CIMs entre 0,125 e 0,5µg/mL, pelo método de microdiluição. Também aqui se verificaram CIMs≥CIM90 observada para todas as espécies, em concordância com dados apresentados por Quindós et al.(132). Devemos, porém, reforçar que nenhum isolado apresentou CIM≥1µg/mL pelo método de microdiluição, e que só um isolado de C. glabrata apresentou CIM=1µg/mL, pelo Etest®. Num isolado de C. albicans não foi possível determinar a CIM pelo método de microdiluição, tanto para o fluconazol como voriconazol, e pelo Etest®, para o voriconazol. Por seu turno, a anfotericina B não tem revelado uma evolução negativa temporalmente, mantendo óptima actividade contra o género Candida. Encontra-se descrito o rápido desenvolvimento de resistência secundária de C. lusitaniae por switch fenotípico, após exposição ao fármaco, embora CIMs≥1µg/mL sejam infrequentes (35, 114, 148) . Estudos epidemiológicos identificaram também uma tendência de redução de susceptibilidade de isolados de C. glabrata e C. krusei à anfotericina B (114, 148). No estudo SENTRY, Pfaller et al. identificaram 17,4% e 27,8% de isolados de C. glabrata e de C. krusei resistentes à anfotericina B, respectivamente (118) . Encontram-se também reportados casos de resistência à anfotericina B em estirpes de C. tropicalis, C. guilliermondii (com falência clínica e resistência comprovada à anfotericina B), C. parapsilosis e alguns isolados de C. kefyr, C. lipolytica, C. orthopsilosis e C. metapsilosis (35, 77, 115, 148) . A resistência em C. albicans é extremamente rara, embora tenha sido identificada com frequência preocupante na África do Sul (16% dos isolados), reconhecendo-se um clade distinto de C. albicans nesta região (114). No nosso estudo, a anfotericina B demonstrou excelente actividade contra todas as espécies de Candida, com uma CIM90≤0,25µg/mL e CIMs≤0,125µg/mL em espécies não-albicans, como C. tropicalis, C. parapsilosis, C. guilliermondii e C. glabrata. Curiosamente, os únicos - 105 - Perfil de susceptibilidade aos antifúngicos isolados com CIMs>0,25µg/mL foram de C. albicans, mas não se observou nenhum isolado com CIM>0,5µg/mL. C. lusitaniae evidenciou CIMs díspares, mas ainda assim ≤0,125 µg/mL. Pelo Etest®, não se observou nenhum isolado com CIM>0,5 µg/mL, a CIM90 para todas as espécies foi de 0,38µg/mL e as espécies C. lusitaniae e C. tropicalis apresentaram CIMs≤ 0,125µg/mL. Estes resultados encontram-se em concordância com os observados por Diekema et al., no seu estudo de avaliação de susceptibilidade de espécies raras de Candida (35) . Não se observou a emergência de C. tropicalis ou C. lusitaniae após terapêutica prévia com anfotericina B. Os valores determinados encontram-se abaixo dos apresentados na metaanálise de Quindós et al., pese embora o facto de estes autores terem compilado resultados, determinados com diversos métodos: Neosensitabs® e Sensistitre® YeastOne microdiluição (CLSI), EUCAST, Etest®, (132) . Dados apresentados por Swinne et al. também referiram CIMs90 superiores para as espécies não-albicans testadas, tendo inclusivamente observado isolados de C. glabrata, C. parapsilosis, C. krusei e C. tropicalis com CIMs≥1µg/mL, (149) o que não foi verificado no nosso estudo. Constatou-se, porém, um episódio de re-infecção por C. krusei, após terapêutica com anfotericina B lipossómica para um episódio anterior de CI causada pelo mesmo agente (Tabela 10). Para os dois isolados em causa, as CIMs contra a anfotericina B foram de 0,125µg/mL e 0,063µg/mL, observando-se uma redução no valor da CIM de C. krusei, no isolado pós-terapêutica com anfotericina B lipossómica. Quanto ao grupo das equinocandinas, apresenta uma óptima actividade fungicida contra todas as espécies de Candida (119). Diversos autores identificaram CIMs ligeiramente mais elevadas em Candida spp. para a caspofungina e micafungina, comparativamente à anidulafungina (133). - 106 - Perfil de susceptibilidade aos antifúngicos As espécies C. guilliermondii, C. parapsilosis, C. famata e C. lusitaniae apresentaram, em estudos epidemiológicos, CIMs mais elevadas, transversalmente a todos os antifúngicos desta classe (119, 133) . Verificaram-se casos de C. glabrata, C. albicans e C. tropicalis com CIMs elevadas às equinocandinas, comparativamente ao esperado globalmente (valor modal). Nas espécies C. glabrata, C. albicans, C. tropicalis, C. parapsilosis e C. krusei estão ainda descritos casos de resistência clínica à caspofungina, com terapêuticas prolongadas (11, 54, 114) . No caso da anidulafungina, Reboli et al. evidenciaram a persistência de CI em oito dos doentes tratados com anidulafungina (134) . No seguimento destes case reports, identificaram- se mutações no genes FKS (FKS1, FKS2 e FKS3) que codificam para a β-1,3-glucano sintetase, um complexo enzimático, e a enzima alvo destes fármacos (33, 54, 55) . Encontram-se descritas mutações em diferentes regiões do gene FKS1, na espécie C. albicans e mutações no gene FKS2, em estirpes de C. glabrata. Este tipo de mecanismo de resistência encontra-se também descrito em estirpes de C. krusei (11, 54) . Nas espécies C. guilliermondii e C. parapsilosis, a susceptibilidade reduzida parece relacionar-se com polimorfismos naturais na região fks1p (98) . Contudo, dado a frequência rara destes acontecimentos, não era de esperar que o mesmo ocorresse neste estudo, esperando-se, obviamente, uma óptima actividade fungicida contra todas as espécies de Candida, mesmo em espécies resistentes aos triazóis. Os nossos resultados diferiram consoante o método utilizado. A percentagem de susceptibilidade observada pelo método de microdiluição foi de 100% e 98,34% para a anidulafungina e caspofungina, respectivamente. Considerando o Etest®, a susceptibilidade foi sensivelmente mais baixa, de 98,34% e 93,34% para a caspofungina e anidulafungina, respectivamente. A caspofungina evidenciou CIMs mais elevadas que a anidulafungina, factos corroborados pela literatura internacional (133) . A CIM90 para todas as espécies foi de 0,5µg/mL para a caspofungina e 0,063µg/mL para a anidulafungina. Olhando para o valor - 107 - Perfil de susceptibilidade aos antifúngicos modal para todas as espécies, a caspofungina apresentou um valor modal de 0,12µg/mL e a anidulafungina de 0,015µg/mL, o que ainda reforça mais as diferenças de CIMs observadas entre as duas equinocandinas. Devemos, contudo, notar que os valores modais para C. albicans e espécies não-albicans são discrepantes para os dois fármacos, sobretudo se considerarmos as espécies C. parapsilosis e C. guilliermondii. Assim, caspofungina e anidulafungina apresentaram valores modais de 0,12µg/mL e 0,015µg/mL, e CIM90 de 0,25µg/mL e 0,063µg/mL, para a espécie C. albicans, respectivamente. Foi notória a discrepância observada para a caspofungina, no nosso estudo, relativamente aos valores apresentados por Pfaller et al. (119). Todavia, Messer et al. reportaram CIMs90 para C. albicans similares às observadas no nosso estudo (92) . Por seu turno, as espécies não-albicans apresentaram valores modais de 0,5µg/mL e 0,063µg/mL, para a caspofungina e anidulafungina, respectivamente, claramente superiores para ambas as equinocandinas. Em estudos mais antigos, Pfaller et al. reportaram C. glabrata, C. tropicalis e C. krusei com valores modais de CIM entre 0,015 e 0,06µg/mL, para este grupo terapêutico (119) . No nosso estudo, a caspofungina apresentou CIMs claramente mais elevadas às reportadas por estes autores para espécies não-albicans. No entanto, o intervalo de CIMs observado, no nosso estudo, para todas as espécies, foi de 0,016-4µg/mL e de 0,016-2µg/mL para caspofungina e anidulafungina, respectivamente, demonstrando ser relativamente similar entre as duas equinocandinas, tal como reportado em estudos prévios de Andes et al. (5). Deve reforçar-se a NS cruzada observada no isolado de C. guilliermondii, pelo Etest®, com discrepância de resultados entre este método e o método padronizado pelo CLSI. De qualquer forma, ambos os fármacos exibiram CIM=2µg/mL, para este isolado, pelo método padronizado. C. guilliermondii é considerada uma espécie rara e um estudo conduzido por - 108 - Perfil de susceptibilidade aos antifúngicos Diekema et al. identificaram isolados de C. guilliermondii com menor susceptibilidade às equinocandinas, com CIMs mais elevadas (2 a 16x) que as observadas para outras espécies de Candida; 5,2% resistentes à anfotericina B, com CIMs entre 2 µg/mL e 64 µg/mL; e uma CIM90=8µg/mL para o fluconazol. Interessantemente, o nosso isolado exibiu uma CIM relativamente elevada para o fluconazol (8µg/mL), em comparação com a CIM90 para todas as espécies, e baixa quer para o voriconazol quer para a anfotericina B, de 0,25µg/mL e 0,03µg/mL, respectivamente, em concordância com o reportado por Diekema et al. (35). Os resultados relativamente aos isolados de C. parapsilosis diferiram consoante o teste de susceptibilidade utilizado. Assim, a percentagem de NS para a caspofungina e anidulafungina foi de 33,3% e 0%, respectivamente, de acordo com o método de microdiluição. As CIMs foram, em ambos os casos, iguais ou superiores a 0,5µg/mL, com um intervalo de 0,5-4µg/mL e 0,5-1µg/mL, respectivamente. Se considerarmos o método Etest®, verificamos diferenças substanciais no padrão de NS de C. parapsilosis: 0% para a caspofungina e 100% para a anidulafungina, com intervalos de CIMs de 0,38 a 0,75µg/mL e 3 a 8µg/mL, para caspofungina e anidulafungina, respectivamente. Estes dados são concordantes com evidências microbiológicas descritas na literatura internacional (119, 133). Um novo artigo científico de Arendrup et al. veio evidenciar o risco de estirpes de Candida poderem conter a(s) mutação(ões) fks, sem a(s) evidenciarem fenotipicamente, com CIMs>2µg/mL, reportando EMG em diversos testes, incluindo no método padronizado pelo CLSI e no Etest® (8). O estudo epidemiológico português de Costa-de-Oliveira et al. evidenciou percentagens de resistência superiores às verificadas no nosso estudo, chegando a atingir os 20% de C. albicans S-DD ou resistente ao fluconazol, e 10% de C. albicans S-DD ou resistente ao voriconazol. Estes investigadores verificaram ainda elevada percentagem de resistência - 109 - Perfil de susceptibilidade aos antifúngicos (100%) de C. parapsilosis à caspofungina. Contrariamente ao nosso estudo, estes autores consideraram isolados com CIMs ≥2µg/mL como resistentes (32) . Ao contrário das nossas constatações, um isolado de C. lusitaniae exibiu resistência à anfotericina B, não sendo evidente se se tratou de resistência primária ou secundária ao fármaco (32). As nossas constatações demonstram, apesar de tudo, óptima actividade dos ―velhos‖ fármacos fluconazol e anfotericina B, salvo determinados nichos, e encontram-se maioritariamente em concordância com observações de investigadores a nível mundial. 2.3.2 Concordância entre métodos de avaliação de susceptibilidade aos antifúngicos Existem diversos métodos disponíveis, incluindo métodos padronizados pelo CLSI ou pelo EUCAST, e métodos automatizados que têm vindo a ser desenvolvidos e comercializados para utilização na prática diária. É verdadeiramente pertinente uma avaliação cuidada dos vários tipos de testes disponíveis e da sua fiabilidade e limitações. Neste estudo permitimo-nos realizar uma análise de concordância de resultados, dado termos realizado diversos tipos de métodos. Uma análise da concordância categórica entre os 3 métodos realizados apurou uma concordância categórica superior a 93,34% entre o Etest® e o método de difusão em discos, relativamente ao método de microdiluição, para todos os fármacos testados (Tabela 16). Estes achados encontram-se de acordo com estudos levados a cabo por Torres et al. e Negri et al.(100, 151). Pfaller et al. referenciaram inclusivamente uma baixa percentagem de EMG, EM e M entre o método de microdiluição e teste de difusão em discos, para o fluconazol. Considerando as diferentes espécies, estes autores verificaram que a concordância entre o método de microdiluição e o teste de difusão em discos era menor para C. glabrata, C. parapsilosis e C. tropicalis, o que não foi inteiramente constatado no nosso - 110 - Perfil de susceptibilidade aos antifúngicos estudo (122), dado que verificámos um EMG e um EM para C. glabrata, mas não se constatou qualquer erro para as espécies C. parapsilosis e C. tropicalis. Observámos, contudo, um erro Minor para C. krusei. Negri et al. referenciaram uma concordância categórica entre o Etest® e o método de difusão em discos relativamente ao método de microdiluição muito similar à observada por nós, para a anfotericina B, mas relativamente menor para o fluconazol. A sua avaliação de EMG, EM e M revelou uma percentagem significativa de M para o fluconazol, com 4% de falsos susceptíveis observados pelo método de difusão em discos, devido a uma pobre diferenciação de isolados S-DD (100) . Estas observações encontram-se em concordância com as nossas constatações. Igualmente, Matar et al. reportaram percentagens significativas de M e de EMG devido a menor correlação (≤83%) entre isolados considerados resistentes pelo método de microdiluição e considerados susceptíveis, susceptíveis dose-dependente ou resistentes pelo método de difusão em discos. Também constataram que isolados considerados S-DD pelo método difusão em discos eram, na realidade, S-DD ou resistentes pelo método de microdiluição. Considerando os diâmetros dos halos, na avaliação de susceptibilidade do fluconazol, verificaram um intervalo entre 0 e 47 mm (88) . No nosso estudo, observámos valores entre 17 e 43 mm para o fluconazol, revelando dificuldade de determinação de espécies consideradas resistentes pelo método de microdiluição (Tabela 15). A utilização de meio Mueller Hinton suplementado com azul de metileno, bem como a utilização de períodos de incubação de 24h, têm sido apontados como passos relevantes na metodologia para uma correcta avaliação dos halos de inibição e diminuição dos chamados fenómenos de trailing, ou arrastamento (113, 121) . Também a correcta utilização de estirpes de referência e o controlo de qualidade adequado são muito relevantes, tal como demonstrado por Barry et al.. A utilização da estirpe Candida krusei 6258 é desaconselhada no controlo de qualidade de testes - 111 - Perfil de susceptibilidade aos antifúngicos de difusão em discos para o fluconazol, pelo risco de não surgir halo de inibição ou zonas de (15) diâmetro ≤18 mm . Não se verificaram EM devido a determinação de falsos resistentes (Tabela 14 e Tabela 16). Todavia, o teste de difusão em discos revelou, no nosso estudo, concordância pobre com o método de microdiluição para as espécies C. glabrata e C. krusei. Independentemente deste facto, este método revelou-se, neste estudo, muito reprodutível na avaliação de isolados susceptíveis, relativamente ao voriconazol e anfotericina B, como observado por Matar et al. e Negri et al. (88, 100). Deve ser realçado que não existem pontos de corte de diâmetros de halos de inibição preconizados para a anfotericina B. Para a definição de susceptibilidade baseámo-nos em dados antigos, propostos por Saubolle e Hoeprich. No seu artigo científico publicado em 1978, propuseram um ponto de corte de susceptibilidade de ≥10 mm para a anfotericina B, mediante a realização da correlação entre as CIMs observadas pelo método de microdiluição e o teste de difusão em discos(139). Este ponto de corte tem sido utilizado por outros autores em estudos de avaliação de susceptibilidade Candida spp. à anfotericina B (100) . Os intervalos para o diâmetro dos halos de inibição, observados por Saubolle e Hoeprich, foram semelhantes aos observados no nosso estudo. Verificámos um intervalo entre 14 e 24 mm e um valor modal de 18 mm. Dados de Saubolle e Hoeprich revelaram um intervalo entre 13 e 18 mm e valor modal de 16 mm. Verificámos o aparecimento de 3 isolados com halos de inibição de 14 mm. Os resultados do método de microdiluição verificaram CIMs de 0,125µg/mL para os 3 isolados. O Etest® determinou CIMs entre os 0,19 e 0,5µg/mL. Em dois dos casos, trataram-se de isolados de C. krusei. Estes dados encontram-se de acordo com um estudo recente de Espinel-Ingroff et al., em que referiram a necessidade de rever os pontos de corte para a anfotericina B, considerando que CIMs≤1µg/mL se correlacionam com diâmetros ≥15 mm e CIMs≥2µg/mL se correlacionam com diâmetros ≤13 mm (44). - 112 - Perfil de susceptibilidade aos antifúngicos Noutro estudo publicado pelo mesmo autor, onde avaliou o método de difusão em discos e outros métodos comerciais, observou-se maior reprodutibilidade com tempos de incubação de 48h, uma boa actuação do meio Mueller-Hinton e correlação entre CIMs e diâmetros testados pelo método de difusão em disco (R=0,681) (43) . Devemos referir que, no nosso estudo, o período de incubação para a anfotericina B foi de 24h. Não obstante, a controvérsia relativamente aos pontos de corte mais adequados na avaliação da susceptibilidade à anfotericina mantém-se. Quanto aos métodos de microdiluição e o Etest®, diversas questões se encontram por resolver, incluindo a avaliação de susceptibilidade à anfotericina B. A utilização do meio AM3 tem gerado algumas críticas devido a risco superior de variabilidade interlotes e diminuição da reprodutibilidade dos métodos. Também os ponto de cortes de CIMs para estes métodos ainda não se encontram definidos para a anfotericina B, apesar de trabalhos de Rex et al., e, mais recentemente, de Park et al., na tentativa de realização de correlação entre dados in vitro e in vivo (109) . De qualquer forma, este último autor identificou o meio AM3 como superior ao clássico meio RPMI, para avaliação das CIMs pelo método de microdiluição. Constatou valores de CIMs mais elevados para o Etest®, com 48% dos isolados com CIMs≥0,38µg/mL (109) . Por seu turno, Lozano-Chiu et al. não verificaram diferenças significativas nas CIMs obtidas pelo método de microdiluição, com o meio AM3, apesar da variação de lotes utilizados, pressupondo que a variabilidade interlote pode ser, de facto, uma falsa questão (82). Sugeriram, contudo, que o ponto de corte, com o meio AM3, para o método de microdiluição deveria ser reduzido para ≤0,5µg/mL, por realizar uma separação mais eficiente entre isolados resistentes e sensíveis (82) . As técnicas de citometria de fluxo reportaram grande fiabilidade do meio AM3 para determinadas espécies de Candida, como C. - 113 - Perfil de susceptibilidade aos antifúngicos albicans e C. parapsilosis, mas menor fiabilidade na detecção de isolados resistentes de C. krusei, C. glabrata, C. lusitaniae e C. tropicalis (24). Para a anfotericina B, o Etest® tem revelado, em alguns estudos, superioridade, relativamente ao método de microdiluição, na separação de isolados susceptíveis de resistentes, conforme constatado por diversos autores (108) . Neste método permanecem algumas questões relativamente ao melhor meio, se RPMI se AM3. Peyron et al. observaram maior facilidade de discriminação de isolados resistentes de C. lusitaniae com o meio RPMI,(112) ao invés de outros estudos que reportaram resultados muito favoráveis com o meio AM3 (47, 74, 165) . Num estudo publicado em 1998, Pfaller et al. não encontraram superioridade na avaliação de isolados resistentes e distinção destes, com a utilização do meio AM3, relativamente ao meio RPMI (120) . Wanger et al. observaram, interessantemente, alguma discrepância na concordância espécie a espécie, com a utilização dos dois meios, face ao método de microdiluição (165). No nosso caso, utilizando o meio AM3 apoiados nos diversos estudos publicados, e observámos uma concordância categórica de 100% e concordância em duas diluições de 80%, com o método padronizado pelo CLSI. Estes factos encontram-se em concordância com os verificados por outros autores, designadamente Negri et al., que observou uma concordância categórica de 97% e concordância em duas diluições de 83%, utilizando, embora, o meio RPMI (100). Tal como notado pelos autores supracitados, o intervalo de CIMs que é permitido determinar no teste de microdiluição e no Etest® é muito diferente, considerando todos os fármacos testados, e, como tal, pode conduzir a alguma discrepância nos resultados das CIMs em isolados susceptíveis. Devemos salientar que, tal como notado por Wanger et al. e Chatuverdi et al., observámos uma concordância em duas diluições inferior a 80% para os isolados de C. krusei, mas não para C. glabrata e C. lusitaniae - 114 - (24, 165) . Não obstante, deverá Perfil de susceptibilidade aos antifúngicos ser enfatizado que num dos isolados de C. krusei, que apresentou inclusivamente um halo de inibição de 14 mm pelo teste de difusão em discos, se verificou um valor de Etest® de 0,5µg/mL, superior ao observado pelo teste de microdiluição. O doente em questão, realizou terapêutica com anfotericina B lipossómica, verificando-se posteriormente um novo episódio de CI por C. krusei, com necessidade de um segundo curso de antifúngico com caspofungina. O segundo isolado apresentou uma CIM, pelo Etest®, de 0,38µg/mL e um halo de inibição de 18 mm. Estudos de Clancy e Nguyen, entre outros, têm sugerido a diminuição do ponto de corte de resistência para ≥0,38µg/mL (109). Estudos de correlação in vivo e in vitro levados a cabo por Park et al. não conseguiram comprovar a utilização deste ponto de corte para o Etest® (109) . Tal como os dados publicados por estes investigadores, muitos dos nossos isolados ―caíram‖ nesta CIM. Assim, a definição de novos pontos de corte é uma questão extremamente pertinente, embora não seja perceptível quais os mais adequados para cada tipo de teste. Por sua vez, uma avaliação da concordância entre os dois testes para os dois triazóis testados revela uma concordância categórica de 96,67% para o fluconazol e 100% para o voriconazol. Quanto à concordância em duas diluições, revelou-se inferior para ambos os fármacos, de 66,67% e 88,34%, respectivamente. Estes resultados encontram-se parcialmente de acordo com os dados observados por Negri et al., que reportaram uma percentagem de concordância em duas diluições de 64% para o fluconazol (100) . Tal como estes investigadores, observámos erros M, mas não EM para o fluconazol (Tabela 16), o que também terá contribuído para a elevada percentagem de concordância categórica. Negri et al. observaram uma percentagem de falsos resistentes para o Etest®, enquanto nós observámos M, com categorização de resistente pelo Etest® e S-DD pelo método de microdiluição. Os erros supracitados - 115 - Perfil de susceptibilidade aos antifúngicos contribuíram para uma quantidade significativa de discrepâncias em duas diluições, tendo conduzido a uma percentagem de concordância em duas diluições inferior a 70%. Dados de Matar et al. também revelaram a prevalência de M quando testaram triazóis. O chamado fenómeno de arrastamento (trailing) poderá estar na base das discrepâncias entre métodos, e foi observado em ambos os fármacos no estudo de Matar et al. (88). No nosso estudo, não evidenciámos discrepâncias tão marcadas para o voriconazol como para o fluconazol, o que se repercutiu na percentagem de concordância em duas diluições (Tabela 16), mas que, ainda assim, provou ser inferior à reportada por Matar et al. (88). Outros autores também comprovaram percentagens de concordância em duas diluições elevadas (Pfaller et al., e Chryssanthou et al.) (25, 123). Um estudo antigo de Sewell et al. referiu diferenças entre as percentagens de concordância em duas diluições para o método de midrodiluição, quando comparado com o método de macrodiluição. O factor contributivo para estas discrepâncias foi o tempo de incubação utilizado no método de microdiluição (24h ou 48h), sobretudo em espécies como C. glabrata e C. tropicalis (143) . O tempo ideal para a leitura das CIMs tem vindo a ser debatida pelo CLSI, verificando-se que as leituras às 24h podem obter CIMs clinicamente mais significativas, devido aos fenómenos de arrastamento (trailing), que podem ocorrer mais frequentemente com tempos de incubação mais alargados. Estima-se que cerca de 5% dos isolados possam demonstrar este tipo de comportamento para o fluconazol. Os pontos de corte para o fluconazol foram definidos com base em estudos que utilizaram a leitura às 48h. Dados de Pfaller, Rex e Revankar, entre outros autores, comprovaram que os mesmos pontos de corte poderiam ser utilizados para a leitura às 24h para o fluconazol (26) . Para o voriconazol, ainda não existem estudos suficientes que comprovem a utilização da leitura às 24h pelo método de microdiluição (26) . Matar et al. demonstraram que a concordância entre o método - 116 - Perfil de susceptibilidade aos antifúngicos microdiluição e Etest®, para os dois triazóis, é superior se a leitura for realizada às 24h. Deve, contudo, ressalvar-se que utilizaram, no seu estudo, meio Mueller Hinton, em vez de RPMI (88) . Outros aspectos relevantes na avaliação da susceptibilidade aos triazóis, designadamente voriconazol, são os pontos de corte que se encontram, neste momento, estabelecidos para os métodos microdiluição e difusão em discos. As categorias criadas basearam-se numa séries de ensaios clínicos e correlações entre os dados in vivo e in vitro, com especial atenção para parâmetros farmacocinéticos e farmacodinâmicos (117, 125). Contudo, existem alguns problemas com estas categorizações, conforme apontado por diversos micologistas. Primeiro de tudo, a utilização da categoria S-DD poderá ser pouco pertinente e poderá convidar o clínico, à semelhança do realizado para o fluconazol, a escalar doses, num fármaco com uma cinética não linear. Por outro lado, um ponto de corte de resistência ≥4µg/mL poderá não ser suficiente para diferenciar espécies resistentes in vivo, já que a escolha dos pontos de corte considerou maioritariamente dados de C. albicans (9) . Em resposta a estas fragilidades, dados de Pfaller et al. revelaram que 99% dos isolados de Candida avaliados foram inibidos por concentrações ≤1µg/mL (92% para C. glabrata e 100% para C. krusei). Por outro lado, isolados de C. glabrata com CIM ≥4µg/mL revelaram, concomitantemente, resistência ao fluconazol. Aliás, observou-se, em estudos in vitro, uma correlação muito significativa entre as CIMs de voriconazol e fluconazol para esta espécie. Pfaller et al. observaram que, cerca de 39% dos isolados resistentes ao fluconazol, apresentaram CIMs≥4µg/mL, 44% apresentaram CIMs≥2µg/mL e 17% CIMs≤1µg/mL. Para além destes dados, os resultados in vivo demonstraram eficácia entre 72 a 92%, excepto para C. glabrata (55%). Da aplicação da regra 90/60, o voriconazol obteve resposta clínica em isolados com CIMs≥4µg/mL, em 60% dos casos. Este conjunto de dados permitiu o estabelecimento de pontos de corte para o - 117 - Perfil de susceptibilidade aos antifúngicos voriconazol, pelo método de microdiluição (117) . Estes dados, compilados com informação de estudos prévios, como do programa ARTEMIS (123) , permitiram ainda a proposta de estabelecimento de ponto de cortes para o voriconazol pelo método de difusão em discos, pois os dois métodos, microdiluição e difusão em discos, demonstraram uma concordância categórica muito elevada para as 3 categorias preconizadas (117, 123) . No nosso estudo, também evidenciámos concordância categórica de 100% entre os dois métodos, distribuição das espécies por CIMs≤1µg/mL e distribuição dos diâmetros para valores >17 mm, aliás ≥22 mm (Tabela 15). As questões levantadas por Arendrup et al., permanecem, contudo, pertinentes e novos desafios poderão ser encontrados na reavaliação de pontos de corte para o voriconazol, pelas metodologias supracitadas (9). Alguns desafios prendem-se ainda com outra classe de fármacos, as equinocandinas. Estes fármacos têm sido alvo de intensa pesquisa na tentativa de permitir uma utilização de metodologias de avaliação de susceptibilidade in vitro com adequada correlação in vivo. Apesar de já existirem dados relativos à avaliação de susceptibilidade destes fármacos pelo método de difusão em disco, não nos foi possível utilizar este método, por indisponibilidade de discos comercializados. Assim, realizámos somente a avaliação de susceptibilidade por 2 métodos – microdiluição e Etest®, tendo obtido resultados interessantes para ambos os fármacos testados. Verificou-se uma concordância categórica entre 93,34 e 98,34% (Tabela 16). A concordância em duas diluições demonstrou ser bastante menor, de 68,34% para a anidulafungina e de 73,34% para a caspofungina (Tabela 16). Os valores de concordância em duas diluições apresentados por Chryssanthou et al. demonstraram, igualmente, percentagens menores às exibidas para o - 118 - Perfil de susceptibilidade aos antifúngicos voriconazol, por exemplo, com 87% de concordância em duas diluições (leitura às 48h), ainda assim superior ao observado no nosso estudo (25). Estes autores constataram CIMs pelo Etest® muito inferiores às observadas pelo método de microdiluição, para a caspofungina. Verificaram ainda a categorização inadequada de um isolado de C. guilliermondii, considerado susceptível pelo Etest® e NS pelo método de microdiluição. Uma das observações mais relevantes foi que se obteve maior correlação entre os métodos com tempos de leitura às 24h para ambos os métodos, quando comparado com a leitura às 48h (25). No nosso estudo, foram observadas CIMs com um valor modal de 0,006µg/mL, pela metodologia Etest® para a homóloga anidulafungina, em comparação com o valor modal de 0,016µg/mL obtido pelo teste de microdiluição. Para a caspofungina, ambos os métodos apresentaram um valor modal de 0,125µg/mL. Observámos erros relevantes nesta classe, pelo método Etest®, para as espécies C. guilliermondii e C. parapsilosis, com 5 EM e 1 EMG. Ao contrário dos autores supracitados, não verificámos CIMs mais baixas para C. guilliermondii pelo Etest®, mas sim num isolado de C. parapsilosis, no que concerne a caspofungina. O método Etest® referenciou como não susceptíveis à anidulafungina todos os isolados de C. parapsilosis, em desacordo com as CIMs obtidas pelo método de microdiluição. Os pontos de corte propostos pelo CLSI continuam a gerar alguma discussão, por se tratar de um assunto tão recente. Novos dados obtidos em modelos animais, sobre diferenças na farmacodinâmica de determinadas espécies, revelaram a necessidade de utilização de doses mais elevadas deste grupo terapêutico para tratamento de C. parapsilosis (5) . Garcia-Effron et al. e Odabasi et al. evidenciaram o potencial papel do soro no aumento significativo das CIMs, implicando, especialmente, a eficácia da anidulafungina e micafungina (55, 103) . Os primeiros autores sugeriram, por isso, um ponto de corte de não susceptível >0,5µg/mL para estes dois fármacos, reforçando que este ponto de corte não iria criar muitos falsos - 119 - Perfil de susceptibilidade aos antifúngicos ―resistentes‖, dado que 99% dos isolados testados tinham CIMs≤0,5µg/mL (55) . Da mesma forma, Arendrup et al. são da opinião que será necessário a utilização de pontos de corte mais baixos para diferenciar adequadamente estirpes susceptíveis de não susceptíveis. Estes autores verificaram a existência de isolados de Candida, com mutações no gene FKS, mas com CIMs≤2µg/mL. Para além disso, avaliaram a concordância dos diversos métodos disponíveis, incluindo o método de microdiluição e Etest®, face à detecção de mutantes fks e a correcta diferenciação de estirpes selvagens. Reportaram uma percentagem elevada de isolados mutantes incorrectamente identificados como isolados susceptíveis, de acordo com os pontos de corte propostos pelo CLSI (8) . Quando utilizados os pontos de corte propostos por estes autores, as estirpes mutantes foram consideradas susceptíveis somente em cerca de 10% dos casos. Os mesmos autores reportaram uma percentagem inferior de EM pelo método de microdiluição, relativamente ao Etest®, mas superior de EMG, para todas as equinocandinas testadas. A avaliação da anidulafungina resultou em menor número de EMG e EM, relativamente às outras equinocandinas testadas, com idêntica actuação de ambos os métodos (8) . Estes dados reforçam a dificuldade, no nosso estudo, de avaliação dos resultados microbiológicos produzidos quer pelo Etest® quer pelo método de microdiluição. Será relevante enfatizar a percentagem superior de mutantes não susceptíveis classificados como susceptíveis pelo método de microdiluição, de acordo com o estudo supracitado. À luz dos pontos de corte sugeridos por Garcia-Effron et al.para a anidulafungina, os nossos isolados de C. parapsilosis e C. guilliermondii seriam considerados não susceptíveis à anidulafungina, quer pelo Etest®, quer pelo método de microdiluição. Todavia, é pertinente realçar que a correlação entre a mutação FKS e a elevação de CIMs pode ser clinicamente relevante, mas desconhece-se se a mesma mutação em isolados com CIMs baixas terá a mesma relevância clínica (124) . Esta questão é deveras pertinente, sobretudo se considerarmos que uma meta- - 120 - Perfil de susceptibilidade aos antifúngicos análise identificou taxas de sucesso terapêutico com caspofungina no tratamento de CI por C. parapsilosis e por C. guilliermondii, superiores a 70%, apesar de todas as limitações inerentes a este tipo de análise e impossibilidade de correlação com dados in vitro (30). Esta área apresenta-se em constante evolução, afigurando-se pertinentes todas as questões anteriormente focadas. Face ao exposto, parece-nos prematuro a utilização de pontos de corte tão abrangentes como 2µg/mL, pelo menos para algumas espécies de Candida. - 121 - Conclusões finais 3. Conclusões Finais Os fungos têm emergido como agentes patogénicos, especialmente em doentes hospitalizados e imunocomprometidos. Candida spp. tem sido considerada a quarta causa de infecções da corrente sanguínea, responsável por 8 a 10% de todas as infecções da corrente sanguínea adquiridas no hospital. Em UCIs observam-se incidências geralmente superiores às observadas numa enfermaria, pois diversos estudos indicam um risco superior de desenvolvimento de CI em doente com idades extremas, oncológicos, sob administração de antibióticos de largo espectro, submetidos a procedimentos cirúrgicos extensos e a utilização de dispositivos médicos invasivos, por exemplo. Este tipo de doentes são internados com frequência em UCIs. Para além disso, dados oficiais norte-americanos revelam que a mortalidade atribuível a CI ronda os 15-35% nos adultos. Logo, existe um esforço continuado para uma melhor caracterização desta problemática devido às elevadas mortalidade e morbilidade associadas a este tipo de infecções. O estudo epidemiológico por nós desenvolvido, com o intuito de podermos contribuir para um melhor conhecimento do que se passa em Portugal, permite-nos realçar alguns pontos: Observou-se uma incidência de CI de 12,8%, com uma taxa de letalidade de 19,2% e razão de mortalidade proporcional de 11,1%. Pelo contrário, a incidência de candidemia, foi baixa, de 0,46%; O isolamento da espécie C. albicans manteve-se muito frequente na nossa população, correspondendo a 87,5% dos agentes isolados em uroculturas e 71% em amostras respiratórias. Esta espécie foi responsável por cerca de 60% de todos os episódios de CI; - 122 - Conclusões finais As CI causadas por espécies não-albicans foram frequentes, correspondendo a cerca de 40% dos casos de CI; As infecções mistas tiveram grande expressão na nossa reduzida amostra, de 21,4%; As formas mais frequentes de candidíase foram infecções de ponto de partida pulmonar, infecções intra-abdominais e infecções do tracto urinário; Não se observaram diferenças significativas face a dados apresentados na literatura internacional, relativamente à epidemiologia e distribuição das espécies responsáveis por CI; Verificou-se relação entre o isolamento de Candida spp. em urocultura e o sexo, com 65% de uroculturas positivas de Candida spp. pertencentes à população feminina; Notou-se uma tendência para o isolamento de espécies não-albicans, em doentes que utilizaram previamente quinolonas; O padrão de terapêutica antifúngica demonstrou concordância com as recomendações de paineis de peritos, com utilização prevalente de fluconazol; Constatou-se a diminuída actividade do fluconazol contra as espécies C. glabrata e C. krusei, mas este fármaco manteve óptima actividade contra o género Candida; Os fármacos voriconazol e anfotericina B revelaram óptima actividade contra Candida spp.. O voriconazol demonstrou óptima actividade contra C. glabrata e C. krusei; Constatou-se não susceptibilidade das espécies C. guillermondii e C. parapsilosis às equinocandinas, com resultados discrepantes entre metodologias empregues; Os Etest® e teste de difusão em discos apresentaram uma concordância categórica com o método de referência superior a 93,34% para todos os fármacos testados. A concordância em duas diluições entre o Etest® e o método de referência revelou-se inferior para todos os fármacos, entre os 66,67% e 88,34%; - 123 - Conclusões finais Erros Major, Minor e Muito Graves foram observados infrequentemente, nomeadamente na avaliação do fluconazol e das equinocandinas. Como tal, poderemos constatar conformidade dos resultados apresentados no nosso estudo com dados epidemiológicos descritos em diversos estudos internacionais. Verificámos ainda a prevalência na nossa população de diversos factores de risco preditivos de CI, descritos por outros autores. Contudo, a interpretação dos dados populacionais e microbiológicos e comparação com dados apresentados por outros autores revelou-se difícil por heterogeneidade de amostras, desenhos de estudo e metodologias empregues. Deveremos realçar o peso da CI na mortalidade geral da população estudada, com uma percentagem de razão de mortalidade proporcional de 11,1%. A nossa população revelou-se idosa e com inúmeras co-morbilidades associadas, factos que terão contribuído para este valor. Paralelamente, as infecções mistas tiveram grande expressão na nossa população, o que também poderá ter contribuído para a taxa de letalidade e razão de mortalidade proporcional observadas neste estudo. O isolamento da espécie C. albicans manteve-se muito frequente na nossa população, demonstrando a sua relevância clínica em UCIs. Por outro lado, as espécies não-albicans também demonstraram uma importância relativa significativa, embora de forma discrepante. C. parapsilosis não demonstrou grande relevância na nossa população, ao contrário de C. glabrata, C. krusei e C. tropicalis. Alguns factos poderão ter contribuído para estas discrepâncias. As quinolonas poderão estar implicadas, pois a utilização deste tipo de antibioterapia poderá contribuir para o isolamento de determinadas espécies de Candida, designadamente C. glabrata, que foi o segundo agente mais relevante de CI na nossa população. Adicionalmente, o número de doentes cirúrgicos foi relativamente elevado, encontrando-se descrito 2 episódios de CI de ponto de partida intra-abdominal num universo - 124 - Conclusões finais de 28. Estes factos também parecem ter contribuído para a importância relativa de espécies não-albicans, nomeadamente C. glabrata. Diversos autores referem que a CI por C. parapsilosis é mais relevante na população pediátrica, logo a sua baixa incidência na nossa amostra poderá dever-se à idade média da população estudada. O padrão de utilização da terapêutica antifúngica demonstrou concordância com as recomendações de paineis de peritos, apesar da grande frequência de instituição de terapêutica empírica. A utilização de fluconazol foi prevalente, sendo interessante constatar a elevada taxa de incidência de CI por C. krusei, observada na nossa população. Em termos microbiológicos, foram identificadas resistências esperadas para o fluconazol, mas em menor percentagem que antecipado por dados na literatura internacional, com actividade reduzida contra as espécies C. glabrata e C. krusei. O fluconazol, voriconazol e anfotericina B revelaram óptima actividade contra Candida spp., ressalvando, contudo, algumas fragilidades das metodologias empregues. O voriconazol demonstrou óptima actividade contra C. glabrata e C. krusei, apesar de se terem observado CIMs mais elevadas, relativamente às outras espécies de Candida. A observação de não susceptibilidade das espécies C. guillermondii e C. parapsilosis às novas equinocandinas encontra-se de acordo com inúmeros estudos internacionais. Nestas duas espécies deve, contudo, realçar-se os resultados de susceptibilidade discrepantes entre metodologias empregues, no estudo das equinocandinas, assuntos que têm apaixonado os microbiologistas a nível mundial. Este assunto permanece em discussão cerrada e os dados apresentados, apesar da pequena amostragem, são relevantes, sugerindo a necessidade de revisão dos pontos de corte propostos para estes fármacos, sobretudo se consideramos as espécies supracitadas. Os antifungigramas possuem cada vez mais provas de resultados fidedignos, com boa correlação in vitro e in vivo, amplamente corroborado pelas concordâncias categóricas observadas entre metodologias. - 125 - Conclusões finais Este estudo revela, contudo, fragilidades em alguns pontos dos métodos utilizados, para diferentes grupos terapêuticos, espelhados na percentagem mais reduzida de concordância, em duas diluições, entre o método de microdiluição e o método Etest®, assim como nos erros detectados nos métodos não padronizados, Etest® e método de difusão em discos. A pequena amostragem estudada demonstrou a importância dos testes de susceptibilidade e da sua introdução na prática clínica. Face ao exposto, considera-se pertinente o aprofundamento do conhecimento da susceptibilidade do género Candida a antifúngicos, em Portugal, com estudos multicêntricos. - 126 - Bibliografia Geral 1. Adler-Moore, J.P., and R.T. Proffitt. 2008. Amphotericin B lipid preparations: what are the differences?. Clin. Microbiol. Infect. 14: 25-36. 2. Alby, K., and R. J. Bennett. 2009. Stress-Induced Phenotypic Switching in Candida albicans. Mol. Biol. Cell. 20: 3178-3191. 3. Álvarez, C., J. M. Ramos, R. San Juan, C. Lumbreras, and J. M. Aguado. 2005. Riesgo de sobreinfección asociado com el uso de antibióticos. ¿Todos los antibiótico son iguales? Rev. Esp. Quimioterap. 18: 39-44. 4. Ameriso, S. F., E. A. Fridman, R. C. Leiguarda, and G.E. Sevlever. 2001. Detection of Helicobacter pylori in Human Carotid Atherosclerotic Plaques. Stroke. 32: 385-391. 5. Andes, D., D. J. Diekema, M. A. Pfaller, J. Bohrmuller, K. Marchillo, and A. Lepak. 2010. In Vivo Comparison of the Pharmacodynamic Targets for Echinocandin Drugs against Candida Species. Antimicrob. Agents Chemother. 54: 2497-2506. 6. Angiolella, L., M. M. Micocci, S. D’Alessio, A. Girolamo, B. Maras, and A. Cassone. 2002. Identification of Major Glucan-Associated Cell Wall Proteins of Candida albicans and Their Role in Fluconazole Resistance. Antimicrob. Agents. Chemother. 46: 1688-1694. 7. Antonelli, M., E. Azoulay, J. Chastre, G. Citerio, G. Conti, D. De Backer, F. Lemaire, H. Gerlach, J. Groeneveld, G. Hedenstierna, D. Macrae, J. Mancebo, S. M. Maggiore, A. Mebazaa, P. Metnitz, J. Pugin, J. Wernerman, and H. Zhang. 2008. Year in review in Intensive Care Medicine, 2007. II. Haemodynamics, pneumonia, infections and sepsis, invasive and non-invasive mechanical ventilation, acute respiratory distress syndrome. Intensive Care Med. 34: 405-422. 8. Arendrup, M. C., G. Garcia-Effron, C. Lass-Flörl, A. G. Lopez, J-L. Rodriguez- Tudela, M. Cuenca-Estrella, and D. S. Perlin. 2010. Echinocandin Susceptibility Testing of - 127 - Candida Species: Comparison of EUCAST EDef 7.1, CLSI M27-A3, Etest, Disk Diffusion, and Agar Dilution Methods with RPMI and IsoSensitest Media. Antimicrob. Agents Chemother. 54: 426-439. 9. Arendrup, M. C., and D. W. Denning. 2007. Does One Voriconazole Breakpoint Suit All Candida Species?. J. Clin. Microbiol. 45: 2093-2094. [Letter] 10. Ascioglu, S., J. H. Rex, B. de Pauw, J. E. Bennett, J. Bille, F. Crokaert, D. W. Denning, J. P. Donnelly, J. E. Edwards, Z. Ejavec, D. Fiere, O. Lortholary, J. Maertens, J.F. Meis, T. F. Patterson, J. Ritter, D. Sellelag, P. M. Shah, D. A. Stevens, and T. J. Walsh. 2002. Defining Opportunistic Invasive Fungal Infections in Immunocompromised Patients with Cancer and Hematopoietic Stem Cell Transplants: An International Consensus. Clin. Infect. Dis. 34: 7-14. 11. Baixench, M-T., N. Aoun, M. Desnos-Ollivier, D. Garcia-Hermoso, S. Bretagne, S. Ramires, C. Piketty, and E. Dannaoui. 2007. Acquired resistance to echinocandins in Candida albicans: case report and review. J. Antimicrob. Chemother. 59: 1076-1083. 12. Bard, M., A.M. Sturm, C. A. Pierson, S. Brown, K. M. Rogers, S. Nabinger, J. Eckstein, R. Barbuch, N. D. Lees, S.A. Howell, and K.C. Hazen. 2005. Sterol uptake in Candida glabrata: rescue of sterol auxotrophic strains. Diagn. Microbiol. Infect. Dis. 52: 285293. [Abstract] 13. Barelle, C. J., C. L. Priest, D. M. MacCallum, N. A. R. Gow, F. C. Odds, and A. J. P. Brown. 2006. Niche-specific regulation of central metabolic pathways in a fungal pathogen. Cell. Microbiol. 8: 961–971. 14. Barelle, C. J., V. M. S. Duncan, A. J. P. Brown, N. A. R. Gow, and F. C. Odds. 2008. Azole antifungals induce up-regulation of SAP4, SAP5 and SAP6 secreted proteinase genes - 128 - in filamentous Candida albicans cells in vitro and in vivo. J. Antimicrob. Chemother. 61: 315322. 15. Barry, A., J. Bille, S. Brown, D. Ellis, J. Meis, M. Pfaller, R. Rennie, M. Rinaldi, T. Rogers, and M. Traczewski. 2003. Quality Control Limits for Fluconazole Disk Susceptibility Tests on Mueller-Hinton Agar with Glucose and Methylene Blue. J. Clin. Microbiol. 41: 3410-3412. 16. Bassetti, M., E. Righi, A. Costa, R. Fasce, M. P. Molinari, R. Rosso, F. B. Pallavicini, and C. Viscoli. 2006. Epidemiological trends in nosocomial candidemia in intensive care. BMC Infect. Dis. 6: 21 17. Berenguer, J., M. Buck, F. Witebsky, F. Stock, P. A. Pizzo and T. J. Walsh. 1993. Lysis—centrifugation blood cultures in the detection of tissue-proven invasive candidiasis disseminated versus single-organ infection. Diagn. Microbiol. Infect. Dis. 17: 103-109. [Abstract] 18. Bernard, E. M. K. J. Christiansen, S-F. Tsang, T. E. Kiehn, and D. Armstrong. 1981. Rate of Arabinitol Production by Pathogenic Yeast Species. J. Clin. Microbiol. 14: 189194. 19. Brajtburg, J., and J. Bolard. 1996. Carrier Effects on Biological Activity of Amphotericin B. Clin. Microbiol. Rev. 9: 512-531. 20. Bukhary, Z. A. 2008. Candiduria: a Review of Clinical Significance and Management. Saudi J. Kidney Dis. Transplant. 19: 350-360. 21. Carlisle, P.L., M. Banerjee, A. Lazzell, C. Monteagudo, J. L. López-Ribot, and D. Kadosh. 2009. Expression levels of a filament-specific transcriptional regulator are sufficient to determine Candida albicans morphology and virulence. Proc. Natl. Acad.Sci. U.S.A. 106: 599-604. - 129 - 22. Cauda, R. 2009. Candidaemia in Patients with Inserted Medical Device. Drugs. 69: s33s38. 23. Chaffin, W. L. 2008. Candida albicans Cell Wall Proteins. Microbiol. Mol. Biol. Rev.72: 495-544. 24. Chaturvedi, V., R. Ramani, and J. H. Rex. 2004. Collaborative Study of Antibiotic Medium 3 and Flow Cytometry for Identification of Amphotericin B-Resistant Candida Isolates. J. Clin. Microbiol. 42: 2252-2254. 25. Chryssanthou, E., and M. Cuenca-Estrella. 2002. Comparison of the Antifungal Susceptibility Testing Subcommittee of the European Committee on Antibiotic Susceptibility Testing Proposed Standard and the Etest with the NCCLS Broth Microdilution Method for Voriconazole and Caspofungin Susceptibility Testing of Yeast Species. J. Clin. Microbiol. 40: 3841-3844. 26. Clinical and Laboratory Standards Institute. 2008. Reference method for broth dilution antifungal susceptibility testing of yeasts. Approved standard: third edition (M27A3). Wayne, PA: Clinical and Laboratory Standards Institute. 27. Clinical and Laboratory Standards Institute. 2004 Methods for antifungal disk diffusion susceptibility testing of yeasts. Approved guideline (M44-A). Wayne, PA: Clinical and Laboratory Standards Institute. 28. Clinical and Laboratory Standards Institute. 2007. Zone diameter interpretive standards, corresponding minimal inhibitory concentration (MIC) interpretive breakpoints, and quality control limits for antifungal disk diffusion susceptibility testing of yeasts. Informational supplement (M44-S2). Wayne, PA: Clinical and Laboratory Standards Institute. - 130 - 29. Clinical and Laboratory Standards Institute. 2008. Reference method for broth dilution antifungal susceptibility testing of yeasts. Informational supplement (M27-S3). Wayne, PA: Clinical and Laboratory Standards Institute. 30. Colombo, A. L., A. L. Ngai, M. Bourque, S. K. Bradshaw, K. M. Strohmaier, A. F. Taylor, R. J. Lupinacci, and N. A. Kartsonis. 2010. Caspofungin Use in Patients with Invasive Candidiasis Caused by Common Non-albicans Candida Species: Review of the Caspofungin Database. Antimicrob. Agents Chemother. 54: 1864-1871. 31. Colombo, A. L., M. Nucci, B. J. Park, S. A. Nouér, B. Arthington-Skaggs, D. A. da Matta, D. Warnock, and J. Morgan for the Brazilian Network Candidemia Study. 2006. Epidemiology of Candidemia in Brazil: a Nationwide Sentinel Surveillance of Candidemia in Eleven Medical Centers. J. Clin. Microbiol. 44: 2816-2823. 32. Costa-de-Oliveira, S., C. Pina-Vaz, D. Mendonça, and A. G. Rodrigues. 2007. A first Portuguese epidemiological survey of fungaemia in a university hospital. Eur. J. Clin. Microbiol. Infect. Dis. doi:10.1007/s10096-007-0448-4. 33. Cuenca-Estrella, M. 2010. Antifúngicos en el tratamiento de las infecciones sistémicas: importancia del mecanismo de acción, espectro de actividad y resistencias. Rev. Esp. Quimioter. 23: 169-176. [Artigo em espanhol] 34. del Palacio, A., J. Villar, and A. Alhambra. 2009. Epidemiología de las candidiasis invasoras en población pediátrica y adulta. Rev. Iberoam. Micol. 26: 2-7. [Artigo em espanhol] 35. Diekema, D. J., S. A. Messer, L. B. Boyken, R. J. Hollis, J. Kroeger, S. Tendolkar, and M. A. Pfaller. 2009. In VitroActivity of Seven Systemically Active Antifungal Agents against a Large Global Collection of Rare Candida Species as Determined by CLSI Broth Microdilution Methods. J. Clin. Microbiol. 47: 3170-3177. - 131 - 36. Donlan, R. M., and J. W. Costerton. 2002. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 15: 167-193. 37. Donnelly, J.P. 2006. Consensus definitions for invasive fungal disease: Strengths, limitations, and revisions. Med. Mycol. 44: s285-s288. 38. Dupont, B. 2002. Overview of the lipid formulations of amphotericin B. J. Antimicrob. Chemother. 49: 31-36. 39. Eggimann, P., and D. Pittet. 2000. Candidoses invasives en réanimation. Schweiz Med. Wochenschr. 130: 1525-1537. [Artigo em francês] 40. Eggimann, P., J. Garbino, and D. Pittet. 2003. Epidemiology of Candida species infections in critically ill non-immunosuppressed patients. Lancet Infect. Dis. 3: 685-702. 41. Eraso, E. M. Ruesga, M. Villar-Vidal, A. J. Carrillo-Muñoz, A. Espinel-Ingroff, and G. Quindós. 2008. Comparative evaluation of ATB Fungus 2 and Sensititre YeastOne panels for testing in vitro Candida antifungal susceptibility. Rev. Iberoam. Micol. 25: 3-6. 42. Espinel-Ingroff, A. 1996. History of Medical Mycology in the United States. Clin. Microbiol. Rev. 9: 235-272. 43. Espinel-Ingroff, A. 2006. Comparison of Three Commercial Assays and a Modified Disk Diffusion Assay with Two Broth Microdilution Reference Assays for Testing Zygomycetes, Aspergillus spp., Candida spp., and Cryptococcus neoformans with Posaconazole and Amphotericin B. J. Clin. Microbiol. 44: 3616-3622. 44. Espinel-Ingroff, A., E. Canton, D. Gibbs, and A. Wang. 2007. Correlation of NeoSensitabs Tablet Diffusion Assay Results on Three Different Agar Media with CLSI Broth Microdilution M27-A2 and Disk Diffusion M44-A Results for Testing Susceptibilities of Candida spp.and Cryptococcus neoformans to Amphotericin B, Caspofungin, Fluconazole, Itraconazole, and Voriconazole. J. Clin. Microbiol. 45: 858-864. - 132 - 45. Esteves, J. A., J. D. Cabrita, and G. N. Nobre. 1990. Ecologia, p.129-147. In J. A. Esteves, J. D. Cabrita, and G. N. Nobre (ed.), Micologia Médica, 2nd ed. Fundação Calouste Gulbenkian, Lisboa. 46. Esteves, J. A., J. D. Cabrita, and G. N. Nobre. 1990. Referência cronológica, p.VIII. In J. A. Esteves, J. D. Cabrita, and G. N. Nobre (ed.), Micologia Médica, 2nd ed. Fundação Calouste Gulbenkian, Lisboa. 47. Favel, A., F. Peyron, M. De Méo, A. Michel-Nguyen, J. Carrière, C. Chastin, and P. Regli. 1999. Amphotericin B susceptibility testing of Candida lusitaniae isolates by flow cytofluorometry: comparison with the Etest and the NCCLS broth macrodilution method. J. Antimicrob. Chemother. 43: 227-232. 48. Felk, A., M. Kretschmar, A. Albrecht, M. Schaller, S. Beinhauer, T. Nichterlein, D. Sanglard, H. C. Korting, W. Schäfer,1 and B. Hube. 2002. Candida albicans Hyphal Formation and the Expression of the Efg1-Regulated Proteinases Sap4 to Sap6 Are Required for the Invasion of Parenchymal Organs. Infect. Immun. 70: 3689-3700. 49. Fidel, Jr., P. L., J. A. Vasquez, and J.D. Sobel. 1999. Candida glabrata: Review of Epidemiology, Pathogenesis, and Clinical Disease with Comparison to C. albicans. Clin. Microbiol. Rev. 12: 80-96. 50. Fridkin, S. K., and W. R. Jarvis. 1996. Epidemiology of Nosocomial Fungal Infections. Clin. Microbiol. Rev. 9: 499-511. 51. Fujibayashi, T., M. Nakamura, A. Tominaga, N. Satoh, T. Kawarai, N. Narisawa, O. Shinozuka, H. Watanabe, T. Yamazaki, and H. Senpuku. 2009. Effects of IgY against Candida albicans and Candida spp. Adherence and Biofilm Formation. Jpn. J. Infect. Dis. 62: 337-342. - 133 - 52. Galván, B. and F. Mariscal. 2006. Epidemiología de la candidemia en UCI. Rev. Iberoam. Micol. 23: 12-15. [Artigo em espanhol] 53. Gamboa, A. R., M. Mendoza, A. Fernández, and E. Díaz. 2006. Detection of Candida dubliniensis in patients with candidiasis in Caracas, Venezuela. Rev. Iberoam. Micol. 23: 8184. 54. Garcia-Effron, G., D. J. Chua, J. R. Tomada, J. DiPersio, D. S. Perlin, M. Ghannoum, and H. Bonilla. 2010. Novel FKS Mutations Associated with Echinocandin Resistance in Candida Species. Antimicrob. Agents Chemother. 54: 2225-2227. 55. Garcia-Effron, G., S. Park, and D. S. Perlin. 2009. Correlating Echinocandin MIC and Kinetic Inhibition of fks1 Mutant Glucan Synthases for Candida albicans: Implications for Interpretive Breakpoints. Antimicrob. Agents Chemother. 53: 112-122. 56. Giuliano, M., M. Barza, N. V. Jacobus, and S. L. Gorbach. 1987. Effect of BroadSpectrum Parenteral Antibiotics on Composition of Intestinal Microflora of Humans. Antimicrob. Agents Chemother. 31: 202-206. 57. González, G. M., M. Elizondo, and J. Ayala. 2008. Trends in Species Distribution and Susceptibility of Bloodstream Isolates of Candida Collected in Monterrey, Mexico, to Seven Antifungal Agents: Results of a 3-Year (2004 to 2007) Surveillance Study. J. Clin. Microbiol. 46: 2902-2905. 58. Guery, B. P., M. Arendrup. G. Auzinger, E. Azoulay, M. B. Sá, E. M. Johnson, E. Müller, C. Putensen, C. Rotstein, G. Sganga, M. Venditti, R. Z. Crespo,and B. J. Kullberg. 2008. Management of invasive candidiasis and candidaemia in adult non neutropenic intensive care unit patients: Part II. Treatment. Intensive Care Med. doi 10.1007/s001134-008-13339-6. - 134 - 59. Hähner-Rombach, S. 2004. Hospitalization: A contentious Issue for Patients and Health Funds in Baden, 1893-1914. Med. Hist. 48: 329-350. 60. Hasan, F., I. Xess, X. Wang, N. Jain, and B. C. Fries. 2009. Biofilm formation in clinical Candida isolates and its association with virulence. Microbes Infect. 11: 753-761. 61. Herbrecht, R., U. Flückiger, B. Gachot, P. Ribaud, A. Thiebaut, and C. Cordonnier. 2007. Treatment of invasive Candida and invasive Aspergillus infections in adult haemotological patients. EJC Supplements. 5: 49-59. 62. Hernáez, M. L., J. Pla, and C. Nombela. 1997. Aspectos moleculares y genéticos de la resistencia a azoles en Candida albicans. Rev. Iberoam. Micol. 14: 150-154. [Artigo em espanhol] 63. Hughes, J. 2000. The "Matchbox on a Muffin": The Design of Hospitals in the Early NHS. Med. Hist. 44: 21-56. 64. Jain, N., R. Kohli, E. Cook, P. Gialanella, T. Chang, and B. C. Fries. 2007. Biofilm Formation by and Antifungal Susceptibility of Candida Isolates from Urine. Appl. Environ. Microbiol. 73: 1697-1703. 65. Jegier, B., R. Jaszewski, P. Kurnatowski, K. Kuba, and M. Lelonek. 2010. Mycotic DNA in Non-Atherosclerotic Aortic Wall of Coronary Patients Is Associated With sICAM-1 Expression. Circ. J. 74: 749-753. 66. Johnson, E. M. 2008. Issues in antifungal susceptibility testing. J. Antimicrob. Chemother. 61: i13-i18. 67. Jong, A.Y., S. H. M. Chen, M. F. Stins, K. S. Kim, T-L. Tuan, and S-H. Huang. 2003. Binding of Candida albicans enolase to plasmin(ogen) results in enhanced invasion of human brain microvascular endothelial cells. J. Med. Microbiol. 52: 615-622. - 135 - 68. Kalkanci, A., N. Kokturk, E. Senol, K. Acar, O. Guzel, B. Sancak, S. Kustimur, and R. Haznedar. 2005. Could Candida dubliniensis be involved in lung fungal balls? Rev. Iberoam. Micol. 22: 157-159. 69. Karkowska-Kuleta, J., M. Rapala-Kozik, and A. Kozik. 2009. Fungi pathogenic to humans: molecular bases of virulence of Candida albicans, Cryptococcus neoformans and Aspergillus fumigatus. Acta Biochim. Pol. 56: 211-224. 70. Koh, A. Y., J. R. Köhler, KK. T. Coggshall, N. Van Rooijen, and G. B. Pier. 2008. Mucosal Damage and Neutropenia are required for Candida albicans Dissemination. PLoS Pathog. 4(2): e35. doi:10.1371/journal.ppat.0040035. 71. Koul, A., J. Vitullo, G. Reyes, and M. Ghannoum. 1999. Effects of voriconazole on Candida glabrata in vitro. J. Antimicrob. Chemother. 44: 109-112. 72. Krcmery, V., E. Oravcova, S. Spanik, M. Mrazova-Studena, J. Trupl, A. Kunova, K. Stopkova-Grey, E. Kukuckova, I. Krupova, A. Demitrovicova and K. Kralovicova. 1998. Nosocomial breakthrough fungaemia during antifungal prophylaxis or empirical antifungal therapy in 41 cancer patients receiving antineoplastic chemotherapy: analysis of aetiology risk factors and outcome. J. Antimicrob. Chemother. 41: 373-380. 73. Laín, A., M. D. Moragues, J. C. G. Ruiz, J. Mendoza, A. Camacho, A. del Palacio, and J. Pontón. 2007. Evaluation of a Novel Enzyme-Linked Immunosorbent Assay To Detect Immunoglobulin G Antibody to Enolase for Serodiagnosis of Invasive Candidiasis. Clin. Vaccine Imunol. 14: 318-319. 74. Law, D., C. B. Moore, and D. W. Denning. 1997. Amphotericin B resistance testing of Candida spp.: a comparison of methods. J. Antimicrob. Chemother. 40: 109-112. - 136 - 75. Linares, C. E. B., E. S. Loreto, C. P. Silveira, P. Pozzatti, J. M. Santurio, and S. H. Alves. 2007. Enzimatic and Hemolytic Activities of Candida dubliniensis Strains. Rev. Inst. Med. Trop. Sao Paulo. 49: 203-206. 76. Liu, T.T., R. E. B. Lee, K. S. Barker, R. E. Lee, L. Wei, R. Homayouni, and P. D. Rogers. 2005. Genome-Wide Expression Profiling of the Response to Azole, Polyene, Echinocandin, and Pyrimidine Antifungal Agents in Candida albicans. Antimicrob. Agents Chemother. 49: 2226-2236. 77. Lockhart, S. R., S. A. Messer, M. A. Pfaller, and D. J. Diekema. 2008. Geographic Distribution and Antifungal Susceptibility of the Newly Described Species Candida orthopsilosis and Candida metapsilosis in Comparison to the Closely Related Species Candida parapsilosis. J. Clin. Microbiol. 46: 2659-2664. 78. Lockhart, S. R., S. A. Messer, M. A. Pfaller, and D. J. Diekema. 2008. Lodderomyces elongisporus Masquerading as Candida parapsilosis as a Cause of Bloodstream Infections. J. Clin. Microbiol. 46: 374-376. 79. Lockhart, S. R., S. A. Messer, M. A. Pfaller, and D. J. Diekema. 2009. Identification and Susceptibility Profile of Candida fermentati from a Worldwide Collection of Candida guilliermondii Clinical Isolates. J. Clin. Microbiol. 47: 242-244. 80. Lockhart, S. R., S. A. Messer, M. Gherna, J. A. Bishop, W. G. Merz, M. A. Pfaller, and D. J. Diekema. 2009. Identification of Candida nivariensis and Candida bracarensis in a Large Global Collection of Candida glabrata Isolates: Comparison to the Literature. J. Clin. Microbiol. 47: 1216-1217. 81. Lohse, M. B., and A. D Johnson. 2009. White–opaque switching in Candida albicans. Curr. Opin. Microbiol. 12: 650–654. - 137 - 82. Lozano-Chiu, M., P. W. Nelson, M. L. Lancaster, M. A. Pfaller, and J. H. Rex. 1997. Lot-to-Lot Variability of Antibiotic Medium 3 Used for Testing Susceptibility of Candida Isolates to Amphotericin B. J. Clin. Microbiol. 35: 270-272. 83. Maertens, J.A., P. Frère, C. Lass-Flörl, W. Heinz, and O. A. Cornely. 2007. Primary antifungal prophylaxis in leukaemia patients. 2007. EJC Supplements. 5: 43-48. 84. Mallié, M. J. M. Bastide, A. Blancard, A. Bonnin, S. Bretagne, M. Cambon, J. Chandenier, V. Chauveau, B. Couprie, A. Datry, M. Feuilhade, R. Grillot, C. Guiguen, V. Lavarde, V. Letscher, M. D. Linas, A. Michel, O. Morin, A. Paugam, M. A. Piens, H. Raberin, E. Tissot, D. Toubas, and A. Wade. 2005. In vitro susceptibility testing of Candida and Aspergillus spp. to voriconazol and other antifungal agentes using Etest®: results of a French multicentre study. Int. J. Antimicrob. Agents. 25: 321-328. 85. Marchetti, O., J. Bille, U Fluckiger, P. Eggimann, C. Ruef, J. Garbino, T. Calandra, M-P. Glauser, M. G. Täuber, and D. Pittet, for the Fungal Infection Network of Switzerland (FUNGINOS). 2004. Epidemiology of Candidemia in Swiss Tertiary Care Hospitals Secular Trends, 1991-2000. Clin. Infect. Dis. 38: 311-320. 86. Martin, G.S., D. M. Mannino, S. Eaton, and M. Moss. 2003. The Epidemiology of Sepsis in the United States from 1979 through 2000. N. Engl. J. Med. 348: 1546-1554. 87. Massanet P, P. Corne, M. Conseil, PF. Perrigault, L. Lachaud, S. Jaber, B. Calvet, O. Jonquet, and J. Reynes. 2010. Antifungal treatments of candidemia in non-neutropenic patients: evaluation of practice in intensive care units. Med. Mal. Infect. 40: 644-649. [Abstract] 88. Matar, M. J., L. Ostrosky-Zeichner, V. L. Paetznick, J. R. Rodriguez, E. Chen, and J. H. Rex. 2003. Correlation between Etest, Disk Diffusion, and Microdilution Methods for - 138 - Antifungal Susceptibility Testing of Fluconazole and Voriconazole. Antimicrob. Agents Chemother. 47: 1647-1651. 89. Mazzei, T. and A. Novelli. 2009. Pharmacological Properties of Antifungal Drugs with a Focus on Anidulafungin. Drugs. 69: s79-s90. 90. Méan, M., O. Marchetti, and T. Calandra. 2008. Bench-to-bedside review: Candida infections in the intensive care unit. Crit. Care. 12: 204. 91. Mesa, L. M., N. Arcaya, O. Cañas, Y. Machado, and B. Calvo. 2004. Evaluación de los caracteres fenotípicos para diferenciar Candida albicans de Candida dubliniensis. Rev. Iberoam. Micol. 21: 135-138. [Artigo em espanhol] 92. Messer, S. A., G. J. Moet, J. T. Kirby, and R. N. Jones. 2009. Activity of Contemporary Antifungal Agents, Including the Novel Echinocandin Anidulafungin, Tested against Candida spp., Cryptococcus spp., and Aspergillus spp.: Report from the SENTRY Antimicrobial Surveillance Program (2006 to 2007). J. Clin. Microbiol. 47: 1942-1946. 93. Messer, S. A., R. N. Jones, and T. R. Fritsche. 2006. International Surveillance of Candida spp. and Aspergillus spp.: Report from the SENTRY Antimicrobial Surveillance Program (2003). J. Clin. Microbiol. 44: 1782-1787. 94. Moellering, R.C., J.R. Graybill, J.E.McGowan, and L.Corey. 2007. Antimicrobial resistance prevention initiative- an update: Proceedings of an expert panel on resistance. Am. J. Infect. Control. 35: s1-s23. 95. Mohan das, V., and M. Ballal. 2008. Proteinase and phospholipase activity as virulence factors in Candida species isolated from blood. Rev. Iberoam. Micol. 25: 208-210. 96. Mokaddas, E. M., N. A. Al-Sweih, and Z. U. Khan. 2007. Species distribution and antifungal susceptibility of Candida bloodstream isolates in Kuwait: a 10-year study. J. Med. Microbiol. 56: 255-259. - 139 - 97. Moulin A. M. 1985. Os frutos da ciência, p. 92-105. In J. Le Goff, and A. Freire (ed.), As Doenças têm história, 1st ed. Terramar Editores, Distribuidores e Livreiros, lda, Lisboa. 98. Munro, C. A. 2010. Fungal echinocandin resistance. Biol. Rep. 2:66 (doi:10.3410/B266) 99. Nailis, H., D. Vandenbosch, D. Deforce, H. J. Nelis, and T. Coenye. 2010. Transcriptional response to fluconazol and amphotericin B in Candida albicans biofilms. Res. Microbiol. 161: 284-292. 100. Negri, M., M. Henriques, T. I. E. Svidzinski, C. R. Paula, and R. Oliveira. 2009. Correlation Between Etest®, Disk Diffusion, and Microdilution Methods for Antifungal Susceptibility Testing of Candida Species From Infection and Colonization. J. Clin. Lab. Anal. 23: 324-330 101. Nelson, R. D., N. Shibata, R. P. Podzorski, and M. J. Herron. 1991. Candida Mannan: Chemistry, Suppression of Cell-Mediated Immunity, and Possible Mechanisms of Action. Clin. Microbiol. Rev. 4: 1-19. 102. Nobile, C. J., J. E. Nett, A. D. Hernday, O. R. Homann, J-S. Deneault, A. Nantel, D. R. Andes, A. D. Johnson, and A. P. Mitchell. 2009. Biofilm Matrix Regulation by Candida albicans Zap1. PLoS Biol. 7(6): e1000133.doi:10.1371/journal.pbio.1000133. 103. Odabasi, Z., V Paetznick, J.H. Rex, and L. Ostrosky-Zeichner. 2007. Effects of Serum on In Vitro Susceptibility Testing of Echinocandins. Antimicrob. Agents Chemother. 51: 4212-4214. 104. Orozco, A. S., L. M. Higginbotham, C. A. Hitchcock, T. Parkinson, D. Falconer, A. S. Ibrahim, M. A. Ghannoum, and S. G. Filler. 1998. Mechanism of Fluconazole Resistance in Candida krusei. Antimicrob. Agents Chemother. 42: 2645-2649. - 140 - 105. Panackal, A.A., J. L. Gribskov, J. F. Staab, K. A. Kirby, M. Rinaldi, and K. A. Marr. 2006. Clinical Sgnificance of Azole Antifungal Drug Cross-Resistance in Candida glabrata. J. Clin. Microbiol. 44: 1740-1743. 106. Panagiotidis, G., T. Bäckström, C. Asker-Hagelberg, A. Jandourek, A. Weintraub, and C. E. Nord. 2010. Effect of Ceftaroline on Normal Human Intestinal Microflora. Antimicrob. Agents Chemother. 54: 1811- 1814. 107. Pappas, P. G., C. A. Kaufman, D. Andes, D. K. Benjamin, Jr., T. F. Calandra, J. E. Edwards, Jr., S. G. Filler, J. F. Fisher, B-J. Kullberg, L. Ostrosky-Zeichner, A. C. Reboli, J. H. Rex, T. J. Walsh, and J. D. Sobel. 2009. Clinical Practice Guidelines for the Management of Candidiasis: 2009 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 48: 503-535. 108. Paredes, C. V. T. 2009. Actualización en pruebas de susceptibilidad antifúngica. Rev. Chil. Infect. 26: 144-150. [Artigo em espanhol] 109. Park, B. J., B. A. Arthington-Skaggs, R. A. Hajjeh, N. Iqbal, M. A. Ciblak, W. LeeYang, M. D. Hairston, M. Phelan, B. D. Plikaytis, A. N. Sofair, L. H. Harrison, S. K. Fridkin,and D. W. Warnock. 2006. Evaluation of Amphotericin B Interpretive Breakpoints for Candida Bloodstream Isolates by Correlation with Therapeutic Outcome. Antimicrob. Agents Chemother. 50: 1287-1292. 110. Parkins, M. D., D. M: Sabuda, S. Elsayed, and K. B. Laupland. 2007. Adequacy of empirical antifungal therapy and effects on outcome among patients with invasive Candida species infections. J. Antimicrob. Chemother. 60: 613-618. 111. Péman, J., E. Cantón, M. Gobernado, and the Spanish ECMM Working Group on Candidaemia. 2005. Epidemiology and antifungal susceptibility of Candida species isolatade - 141 - from blood: results of a 2-year multicentre study in Spain. Eur. J. Clin. Microbiol. Infect. Dis. 24: 23-30. 112. Peyron, F., A. Favel, A. Michel-Nguyen, M. Gilly, P. Regli, and A. Bolström. 2001. Improved Detection of Amphotericin B-Resistant Isolates of Candida lusitaniae by Etest. J. Clin. Microbiol. 39: 339-342. 113. Pfaller, M. A., L. Boyken, S. A. Messer, R. J. Hollis, and D. J. Diekema. 2004. Stability of Mueller-Hinton Agar Supplemented with Glucose and Methylene Blue for Disk Diffusion Testing of Fluconazole and Voriconazole. J. Clin. Microbiol. 42: 1288-1289. 114. Pfaller, M. A., and D. J. Diekema. 2007. Epidemiology of Invasive Candidiasis: a Persistent Public Health Problem. Clin. Microbiol. Rev. 20: 133-163. 115. Pfaller, M. A., D. J. Diekema, A. L. Colombo, C. Kibbler, K. P. Ng, D. L. Gibbs, V. A. Newell, and the Global Antifungal Surveillance Group. 2006. Candida rugosa, an Emerging Fungal Pathogen with Resistance to Azoles: Geographic and Temporal Trends from the ARTEMIS DISK Antifungal Surveillance Program. J. Clin. Microbiol. 44: 35783582. 116. Pfaller, M. A., D. J. Diekema, D. L. Gibbs, V. A. Newell, J. F. Meis, I. M. Gould, W. Fu, A. L. Colombo, E. Rodriguez-Noriega, and the Global Antifungal Surveillance Group. 2007. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2005: an 8.5 Year Analysis of Susceptibilities of Candida species and Other Yeast Species to Fluconazole and Voriconazole Determined by CLSI Standardized Disk Diffusion Testing. J. Clin. Microbiol. 45: 1735-1745. 117. Pfaller, M. A., D. J. Diekema, J. H. Rex, A. Espinel-Ingroff, E. M. Johnson, D. Andes, V. Chaturvedi, M. A Ghannoum, F. C. Odds, M. G. Rinaldi, D. J. Sheehan, P. Troke, T. J. Walsh, and D. W. Warnock. 2006. Correlation of MIC with Outcome for - 142 - Candida Species Tested against Voriconazole: Analysis and Proposal for Interpretive Breakpoints. J. Clin. Microbiol. 44: 819-826. 118. Pfaller, M. A., D. J. Diekema, R. N. Jones, S. A. Messer, R. J. Hollis, and the SENTRY Participants Group. 2002. Trends in Antifungal Susceptibility of Candida spp. Isolated from Pediatric and Adult Patients with Bloodstream Infections: SENTRY Antimicrobial Surveillance program, 1997 to 2000. J. Clin. Microbiol. 40: 852-856. 119. Pfaller, M. A., L. Boyken, R. J. Hollis, J. Kroeger, S. A. Messer, S. Tendolkar, and D. J. Diekema. 2008. In Vitro Susceptibility of Invasive Isolates of Candida spp. to Anidulafungin, Caspofungin, and Micafungin: Six Years of Global Surveillance. J. Clin. Microbiol. 46: 150-156. 120. Pfaller, M. A., S.A. Messer, and A. Bolmström. 1998. Evaluation of etest for determining in vitro susceptibility of yeast isolates to amphotericin B. Diagn. Microbiol. Infect. Dis. 32: 223-227. 121. Pfaller, M.A. 2008. New Developments in the Antifungal Susceptibility Testing of Candida. Current Fungal Infection Reports 2: 125 –133. 122. Pfaller, M.A., K. C. Hazen, S. A. Messer, L. Boyken, S. Tendolkar, R. J. Hollis,and D. J. Diekema. 2004. Comparison of Results of Fluconazole Disk Diffusion Testing for Candida Species with Results from a Central Reference Laboratory in the ARTEMIS Global Antifungal Surveillance Program. J. Clin. Microbiol. 42:3607-3612. 123. Pfaller, M.A., D. J. Diekema, S.A. Messer, L. Boyken, and R. J. Hollis. 2003. Activities of Fluconazol and Voriconazol against 1,586 Recent Clinical Isolates of Candida Species Determined by Broth Microdilution, Disk Diffusion, and Etest Methods: Report from The ARTEMIS Global Antifungal Susceptibility Program 2001. J. Clin. Microbiol. 41: 14401446. - 143 - 124. Pfaller, M.A., L. Boyken, R. J. Hollis, J. Kroeger, S. A. Messer, S. Tendolkar, R. N. Jones, J. Turnidge, and D. J. Diekema. 2010. Wild-Type MIC Distributions and Epidemiological Cutoff Values for the Echinocandins and Candida spp. J. Clin. Microbiol. 48: 52-56. 125. Pfaller. M.A., D. J. Diekema, and D.J. Sheehan. 2006. Interpretive Breakpoints for Fluconazole and Candida Revisited: a Blueprint for the Future of Antifungal Susceptibility Testing. Clin. Microbiol. Rev. 19: 435-447. 126. Pincus, D. H., D. C. Coleman, W. R. Pruitt, A. A. Padhye, I. F. Salkin, M. Geimer, A. Bassel, D. J. Sullivan, M. Clarke, and V. Hearn. 1999. Rapid Identification of Candida dubliniensis with Commercial Yeast Identification Systems. J. Clin. Microbiol. 37: 35333539. 127. Piteira C. 2007. Doenças Infecciosas Transmitidas através do Meio Ambiente., p.8-13. in C. Piteira, A Qualidade do Ar Interior em Instalações Hospitalares, 1st ed. Lidel, edições técnicas, lda, Lisboa. 128. Pontón, J. 2009. Utilidad de los marcadores biológicos en el diagnóstico de la candidiasis invasora. Rev. Iberoam. Micol. 26: 8-14. [Artigo em Espanhol] 129. Pontón, J., and A. del Palacio. 2007. Avances y limitaciones del diagnóstico precoz de las infecciones invasoras causadas por levaduras. Rev. Iberoam. Micol. 24: 181-186. [Artigo em espanhol] 130. Posteraro, B., R. Martucci, M. La Sorda, B. Fiori, D. Sanglard, E. De Carolis, A.R. Florio, G. Fadda, and M. Sanguinetti. 2009. Reliability of the Vitek 2 Yeast Susceptibility Test for Detection of In Vitro Resistance to Fluconazole and Voriconazole in Clinical Isolates of Candida albicans and Candida glabrata. J. Clin. Microbiol. 47: 1927-1930. - 144 - 131. Pultz, N. J., U. Stiefel, M. Ghannoum, M. S. Helfand, and C. J. Donskey. 2005. Effect of Parenteral Antibiotic Administration on Establishment of Intestinal Colonization by Candida glabrata in Adult Mice. Antimicrob. Agents Chemother. 49: 438-440. 132. Quindós, G. A. J. Carrillo-Muñoz, E. Eraso, E. Cantón, and J. Pemán. 2007. Actividad antifúngica in vitro de voriconazol: Nuevos datos después de los primeros años de experiencia clínica. Rev. Iberoam. Micol. 24: 198-209. [Artigo em espanhol] 133. Quindós, G., and E. Eraso. 2008. Actividad antifúngica in vitro de la anidulafungina. Rev. Iberoam. Micol. 25: 83-91. [Artigo em espanhol] 134. Reboli, A. C., C. Rotstein, P. G. Pappas, S. W. Chapman, D. H. Kett, D. Kumar, R. Betts, M. Wible, B. P. Goldstein, J. Sschranz, D. S. Krause, and T. J. Walsh for the Anidulafungin Study Group. 2007. Anidulafungin versus Fluconazol for Invasive Candidiasis. N. Engl. J. Med. 356: 2472-2482. 135. Rello, J., M-E. Esandi, E. Díaz, D. Mariscal, M. Gallego and J. Vallès. 1998. The Role of Candida spp. Isolated From Nonneutropenic Patients Bronchoscopic Samples in Nonneutropenic Patients. Chest. 114: 146-149. 136. Rex, J. H., C. R. Cooper, W. G. Merz, J. N. Galgiani, E. J. Anaissie. 1995. Detection of Amphotericin B-Resistant Candida Isolates in Broth-Based System. Antimicrob. Agents Chemother. 39: 906-909. 137. Sanchez, V., J. A. Vazquez, D. Barth-Jones, L. Dembry, J.D. Sobel, and M. J. Zervos. 1992. Epidemiology of Nosocomial Acquisition of Candida lusitaniae. J. Clin. Microbiol. 30: 3005-3008. 138. Sanglard, D., F. Ischer, T. Parkinson, D. Falconer, and J. Bille. 2003. Candida albicans Mutations in the Ergosterol Biosynthetic Pathway and Resistance to Several Antifungal Agents. Antimicrob. Agents Chemother. 47: 2404-2412. - 145 - 139. Saubolle, M. A., and P. D. Hoeprich. 1978. Disk Agar Diffusion Susceptibility Testing of Yeasts. Antimicrob. Agents Chemother. 14: 517-530. 140. Saunier, A. 1985. A vida quotidiana nos hospitais da Idade Média, p. 205-220. In J. Le Goff, and A. Freire (ed.), As Doenças têm história, 1st ed. Terramar Editores, Distribuidores e Livreiros, lda, Lisboa. 141. Schelenz, S. 2008. Management of candidiasis in the intensive care unit. J. Antimicrob. Chemother. 61: i31-i34. 142. Seneviratne, C. J., L. J. Jin, Y. H. Samaranayake, and L. P. Samaranayake. 2008. Cell Density and Cell Aging as Factors Modulating Antifungal Resistance of Candida albicans Biofilms. Antimicrob. Agents Chemother. 52: 3259-3266. 143. Sewell, D. L., M. A. Pfaller, and A. L. Barry. 1994. Comparison of Broth Macrodilution, Broth Microdilution, and ETest Antifungal Susceptibility Tests for Fluconazole. J. Clin. Microbiol. 32: 2099-2102. 144. Shimizu, Kosuke, M. Osada, K. Takemoto, Y. Yamamoto, T. Asai, and N. Oku. 2010. Temperature-dependent transfer of amphotericin B from liposomal membrane of Ambisome to fungal cell membrane. J. Control. Release. 141: 208-215. 145. Shulman, L., and David Ost. 2005. Managing infection in the critical care unit: how can infection control make the ICU safe? Crit. Care Clin. 21: 111-128. 146. Silva, V., M. Cabrera, M. C. Díaz, C. Abarca, and G. Hermosilla. 2003. Prevalencia de serotipos de Candida albicans en aislamientos de hemocultivo en Chile y primer caso de candidemia por Candida dubliniensis. Rev. Iberoam. Micol. 20: 46-51. [Artigo em espanhol] 147. Society of Critical Care Medicine. History of Critical Care. 2001. Society of Critical Care Medicine. Disponível - 146 - em: http://www.sccm.org/AboutSCCM/History_of_Critical_Care/Pages/default.aspx. Acedido 2 de Novembro 2009. 148. Sterling, T. R., and W. G. Merz. 1998. Resistance to amphotericin B: emerging clinical and microbiological patterns. Drug Resist. Updat. 1: 161-165. 149. Swinne, D., M. Watelle, and N. Nolard. 2005. In vitro activities of voriconazol, fluconazole, itraconazole and amphotericin B against non Candida albicans yeast isolates. Rev. Iberoam. Micol. 22: 24-28. 150. Tamura, N., M. Negri, L. Bonassoli, and T. Svidzinski. 2007. Fatores de virulência de Candida spp isoladas de cateteres venosos e mãos de servidores hospitalares. Rev. Soc. Bras. Med. Trop. 40: 91-93. 151. Torres D., N.A., C. A. Álvarez M. and M. A. Rondón S. 2009. Evaluación mediante tres técnicas de susceptibilidad a fluconazol en especies de Candida aisladas en pacientes com infecciones invasoras. Bogotá – Colombia. Rev. Chil. Infect. 26: 135-143. [Artigo em espanhol] 152. Torres-Rodrıguez, J. M., and E. Alvarado-Ramírez. 2007. In vitro susceptibilities to yeasts using the ATB® FUNGUS 2 method, compared with Sensititre Yeast One® and standard CLSI (NCCLS) M27-A2 methods. J. Anitmicrob. Chemother. 60: 658-661. 153. Tortorano, A. M., A. L. Rigoni, E. Biraghi, A. Prigitano, M. A. Viviani, and the FIMUA-ECMM candidaemia study group. 2003. The European Confederation of Medical Mycology (ECMM) survey of candidaemia in Italy: antifungal susceptibility patterns of 261 non-albicans Candida isolates from blood. J. Antimicrob. Chemother. 52: 679-682. 154. Tortorano, A. M., J. Peman, H. Bernhardt, L. Klingspor, C. C. Kibbler, O. Faure, E. Biraghi, E. Canton, K. Zimmermann, S. Seaton, R. Grillot the ECMM Working Group on Candidaemia. 2004. Epidemiology of Candidaemia in Europe: Results of 28- - 147 - Month European Confederation of Medical Mycology (ECMM) Hospital-Based Surveillance Study. Eur. J. Clin. Microbiol. Infect. Dis. 23: 317-322. 155. Trofa, D., A. Gácser, and J. D. Nosanchuk. 2008. Candida parapsilosis, an Emerging Fungal Pathogen. Clin. Microbiol. Rev. 21: 606-625. 156. Valentín, A., E. Cantón, J. Pemán, and G. Quindós. 2007. Actividad in vitro de la anfotericina B y la anidulafungina sobre biopelículas de Candida albicans y Candida tropicalis. Rev. Iberoam. Micol. 24: 272-277. [Artigo em espanhol] 157. van Till, J. W. O., O. van Ruler, B. Lamme, R. JP Weber, J. B. Reitsma, and M. A. Boermeester. 2007. Single-drug therapy or selective decontamination of the digestive tract as antifungal prophylaxis in critically ill patients: a systematic review. Crit. Care. 11: R126 (doi:10.1186/cc6191). 158. Vandeputte, P. G. Tronchin, F. Rocher, G. Renier, T. Bergès, D. Chabasse, and J-P. Bouchara. 2009. Hypersusceptibility to Azole Antifungals in a Clinical Isolate of Candida glabrata with Reduced Aerobic Growth. Antimicrob. Agents Chemother. 53: 3034-3041. 159. Vandeputte, P. G. Tronchin, T. Bergès, C. Hennequin, D. Chabasse, and J-P. Bouchara. 2007. Reduced Susceptibility to Polyenes Associated with a Missense Mutation in the ERG6 Gene in a Clinical Isolate of Candida glabrata with Pseudohyphal Growth. Antimicrob. Agents Chemother. 51: 982-990. 160. Vediyappan, G., T. Rossignol, and C. d'Enfert. 2010. Interaction of Candida albicans Biofilms with Antifungals: Transcriptional Response and Binding of Antifungals to BetaGlucans. Antimicrob. Agents. Chemother. 54: 2096-2111. 161.Vermitsky, C-P., and T. D. Edlind. 2004. Azole Resistance in Candida glabrata: Coordinate Upregulation of Multidrug Transporters and Evidence for a Pdr1-Like Transcription Factor. Antimicrob. Agents Chemther. 48: 3773-3781. - 148 - 162. Villar, C. C., and A. Dongari-Bagtzoglou. 2009. Immune defence mechanisms and immunoenhancement strategies in oropharyngeal candidiasis. Expert Rev. Mol. Med. 10: e29. doi:10.1017/S1462399408000835. 163. Viscoli, C. 2009. Antifungal Prophylaxis and Pre-Emptive Therapy. Drugs. 69: s75-s78. 164. von Zepelin, M. B., L. Kunz, R. Rüchel, U. Reichard, M. Weig, and U. Groß. 2007. Epidemiology and antifungal susceptibilities of Candida spp. to six antifungal agents: results from a surveillance study on fungaemia in Germany from July 2004 to August 2005. J. Antimicro. Chemother. 60: 424-428. 165. Wanger, A., K. Mills, P. W. Nelson, and J. H. Rex. 1995. Comparison of Etest and National Committee for Clinical Laboratory Standards Broth Macrodilution Method for Antifungal Susceptibility Testing: Enhanced Ability To Detect Amphotericin B-Resistant Candida Isolates. Antimicrob. Agents Chemother. 39: 2520-2522. 166. Weil, M.H. 1973. The Society of Critical Care Medicine, its history and its destiny. Crit. Care Med. 1: 1-4. 167. Wright, S.D., C. Burton, M. Hernandez, H. Hassing, J. Montenegro, S. Mundt, S. Patel, D. J. Card, A. Hermanowski-Vosatka, J. D. Bergstrom,C. P. Sparrow, P. A. Detmers, and Y-S. Chao. 2000. Infectious Agents Are Not Necessary for Murine Atherogenesis. J. Exp. Med. 191: 1437-1491. 168. Yang, Y-L. 2003. Virulence Factors of Candida species. J. Microbiol. Immunol. Infect. 36: 223-228. 169. Yeo, S. F., and B. Wong. 2002. Current Status of Nonculture Methods for Diagnosis of Invasive Fungal Infections. Clin. Microbiol. Rev. 15: 465-484. - 149 - 170. Yera, H., B. Sendid, N. Francois, D. Camus, and D. Poulain. 2001. Contribution of Serological Tests and Blood Culture to the Early Diagnosis of Systemic Candidiasis. Eur. J. Clin. Microbiol. Infect. Dis. 20: 864-870. 171. Young, L.Y., C. M. Hull, and J. Heitman. 2003. Disruption of Ergosterol Biosynthesis Confers resistance to amphotericin B in Candida lusitaniae. Antimicrob. Agents Chemother. 47: 2717-2724. 172. Zilberberg, M. D., M. H. Kollef, H. Arnold, A. Labelle, S. T. Micek, S. Kothari, and A.F. Shorr. 2010. Inappropriate empiric antifungal therapy for candidemia in the ICU and hospital resource utilization: a retrospective cohort study. BMC Infect. Dis. 10: 150. - 150 - Anexo I – Ficha de recolha de dados Isolado de _________________ Local de Isolamento ________________________ Data da Colheita ___/___/___ Sexo F □ M Processo _______________ Data de Nascimento ___/___/___ Data de Admissão Hospitalar ___/___/___ Data de Admissão na UCI ___/___/___ Tipo de Admissão Médica □ Cirurgia Urgente Data de Saída da UCI ___/___/___ Cirurgia (até 30 dias antes da admissão) Abdominal □ □ Vascular □ Coronária □Cirurgia Electiva □ Infecção Urinária Infecção Cirúrgica □ Outra □ IRA □ Pneumonia □ □ Outra □ HTA □ Diabetes Mellitus □IRC □ DHC □ □Traqueobronquite □ DPOC □ Neoplasia □ TP □ VIH+ □ Insuficiência respiratória □ D. Auto-imune □ IC □ CAD □ □ Pós op. vascular imediato □ IRC agudizada Cardíaca □ Neurocirurgia Outra □ Pós op. cardiotorácico imediato □ Exposição a Meios Invasivos Cateteres □ Sim □ Não □ nº___ nº Dias ___ Situação Imunológica Sem Evidência de Imunodeficiência Ventilação Mecânica □ de ___/___/___ a ___/___/___ Cateter Urinário □ de ___/___/___ a ___/___/___ □ Entubação NT/OT □ Traqueostomia □ Neutropenia «500 □ Terapêutica Imunossupressora □ Outra □ Terapêutica Antimicrobiana Mudança Tipo de Data de Data de Antimicrobiano Agente Local de Isolado Infecção Resultado§ de Terapêutica* □ Co-morbilidades Causa de admissão Sepsis /choque séptico / Sepsis severa /MOF □ Bacteriemia □ Torácica □ Coronária Início Fim Antimicrobiano+ 1) 2) 3) 4) 5) * Indicar se Empírica; Dirigida; Sem Indicação Formal; Profilaxia; + Indicar se: Falência; Toxicidade; Dirigida com TSA; § Indicar se Melhorado; Agravado; Não avaliável - 151 -
Baixar