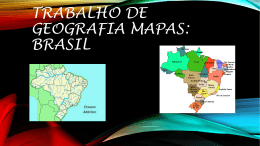
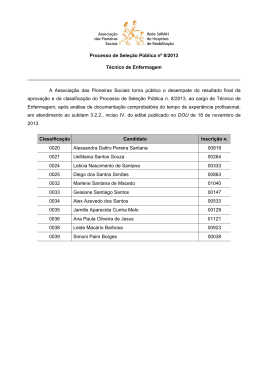
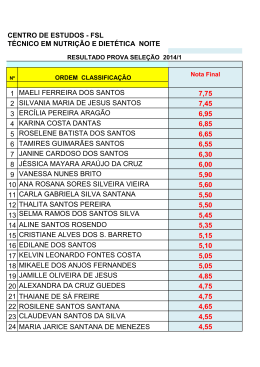
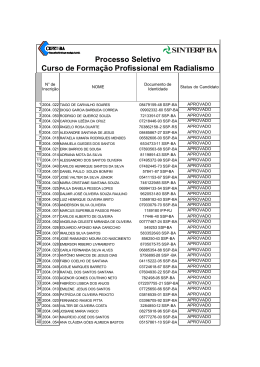
Rev. bras. paleontol. 17(3):363-372, Setembro/Dezembro 2014 © 2014 by the Sociedade Brasileira de Paleontologia doi: 10.4072/rbp.2014.3.07 CARACTERIZAÇÃO PALINOFLORÍSTICA E PALEOAMBIENTAL DA FORMAÇÃO SANTANA (CRETÁCEO INFERIOR), POÇO 4-BO-1-PE, BACIA DO ARARIPE, NORDESTE DO BRASIL HELENA ANTUNES PORTELA, LUZIA ANTONIOLI, RODOLFO DINO Departamento de Estratigrafia e Paleontologia, Faculdade de Geologia, UERJ, Rua São Francisco Xavier, 524 Bloco A, Sala 2030, 20559-013, Rio de Janeiro, RJ, Brasil. [email protected], [email protected], [email protected] MARIA JUDITE GARCIA Laboratório de Paleobotânica e Palinologia, IGc/USP; Rua do Lago, 562, Cidade Universitária, 05508-900, São Paulo, SP, Brasil. [email protected] ABSTRACT – PALYNOFLORAL AND PALEOENVIRONMENTAL CHARACTERIZATION OF THE SANTANA FORMATION (LOWER CRETACEOUS), 4-BO-1-PE WELL, ARARIPE BASIN, NORTHEAST BRAZIL. A paleoenvironmental characterization of the Santana Formation sedimentary rocks (4-BO-1-PE well), Araripe Basin, Late Aptian-Early Albian, was accomplished through qualitative and quantitative analyses of the palynoflora. Palynological and lithological evidence indicate a predominantly continental depositional environment for this section, composed by a fluvial, lacustrine deltaic system in a semi-arid climate. The detection of a single dinoflagellate cyst suggests that small sea ingressions took place during Aptian-Albian times in the region, as also verified in other coeval strata of marginal Brazilian Cretaceous basins. Comparisons with palynofloras from other continents support the hypothesis that this assemblage belongs to the North-Gondwana microfloristic province. Key words: Araripe Basin, Lower Cretaceous, paleoenvironment, paleopalynology. RESUMO – A caracterização do paleoambiente da Formação Santana, bacia do Araripe, de idade Neoaptiana a Eoalbiana, foi realizada através de análise de associações palinoflorísticas recuperadas em testemunhos do poço 4-BO-1-PE. Evidências palinológicas e litológicas indicam um paleoambiente dominantemente continental, composto por um sistema fluvio deltaico lacustre, sob clima quente árido a semiárido. A detecção da presença de um único exemplar de dinoflagelado corrobora a tese do início das pequenas ingressões marinhas que ocorreram durante o Aptiano-Albiano, a exemplo do verificado em outras bacias cretáceas brasileiras de mesma idade. A composição palinoflorística identificada enquadra-se àquelas observadas nas bacias do nordeste brasileiro e insere-se nas características das associações pertencentes à província microflorística do Gondwana-Norte. Palavras-chave: bacia do Araripe, Cretáceo Inferior, paleoambiente, paleopalinologia. INTRODUÇÃO Inferior Pré-Albiana a D. etruscus/Afropollis” de Herngreen et al. (1996). Esta província posiciona-se em uma faixa de aproximadamente 15º de latitude em ambos os lados do paleoequador, e é considerada como representativa de um cinturão climático quente e semiárido. O escopo principal deste trabalho é contribuir na caracterização e determinação das variações do paleoambiente relativo à Formação Santana, bacia do Araripe, bem como na palinoflora contida nos estratos do poço 4-BO-1-PE, perfurado nesta região. Um considerável aumento no conhecimento geológico e paleontológico das sequências aptianas e albianas das bacias mesozoicas brasileiras está sendo alcançado através de recentes revisões estratigráficas e bioestratigráficas realizadas em seções de subsuperfície de poços exploratórios. Entretanto, pesquisas aplicadas vêm demonstrando a necessidade da revisão da classificação litoestratigráfica destas seqüências, bem como da sua palinoflora (Dino et al., 1999; Antonioli, 1998, 2001). A área objeto do presente estudo, pelo seu conteúdo esporopolínico e posição geográfica, insere-se na província “Gondwana Norte” de Brenner (1976), implantada geograficamente na mesma disposição da fitogeoprovíncia III com Dicheiropollis etruscus de Srivastava (1978) e da antiga província WASA (“West African South American”) de Herngreen & Chlonova (1981), atualmente denominada de “província Cretáceo ÁREA DE ESTUDO A bacia sedimentar do Araripe é a mais extensa das bacias interiores mesozoicas do nordeste do Brasil (Figura 1). Localiza-se na província Estrutural Borborema (Brito Neves, 1990), ocupando parte dos estados do Piauí, Pernambuco e Ceará, numa extensão de mais de 8.000 km2. 363 364 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014 CONTEXTO GEOLÓGICO E PALEONTOLÓGICO A bacia sedimentar do Araripe localiza-se entre os meridianos 38º30” e 40º50” de longitude oeste e os paralelos 7º05” e 7º50” de latitude sul (Viana & Neumann, 2002). Esta bacia é estratigraficamente dividida em, da base para o topo: Formação Mauriti; o Grupo Vale do Cariri, constituído pelas formações Brejo Santo, Missão Velha e Abaiara; e o Grupo Araripe, que engloba as formações Rio da Batateira, Santana, Arajara e Exu (Ponte & Appi, 1990). A chapada é constituída por unidades aptianas a cenomanianas, seccionadas por escarpas íngremes que recobrem, em discordância angular, unidades de sequências mais antigas ou repousam diretamente sobre o embasamento cristalino. As sequências sedimentares constituintes da bacia apresentam mergulho em torno de 5ºO (Brito Neves, 1990). A extensão original estimada, para cada uma das diferentes sequências, era bem mais abrangente, considerando os testemunhos remanescentes isolados nas várias pequenas bacias circunvizinhas (Assine, 2007). Durante o Fanerozoico, diversos eventos tectônicos reativaram antigas estruturas do embasamento cristalino, subordinando as bacias interiores do nordeste do Brasil. Dentre elas, a bacia do Araripe é a que apresenta evolução tectono sedimentar mais complexa, constituída por sequências estratigráficas limitadas por discordâncias que representam o registro sedimentar de bacias geneticamente distintas (Assine, 1992). Entre as unidades que constituem a sequência sedimentar do Araripe, a Formação Santana destaca-se no que diz respeito às suas rochas sedimentares e ao seu conteúdo fossilífero. São muito famosas tanto as suas reservas de gipsita quanto sua fauna de peixes, que constitui uma ocorrência ímpar nos registros paleontológicos do Brasil. A Formação Santana é caracterizada por uma grande diversidade litológica verificável ao longo de sua espessura, cujo limite máximo é de aproximadamente 200 m. MATERIAL E MÉTODOS Das cerca de 300 amostras coletadas, foram selecionadas para o presente estudo 53 amostras de calha, coletadas de três em três metros no poço 4-BO-1-PE perfurado na bacia do Araripe (Figura 1). Todas as amostras estão arquivadas e registradas sob os números 2008243 a 2008567 no laboratório de palinomacerais da Faculdade de Geologia (FGEL) da Universidade do Estado do Rio de Janeiro (UERJ). A preparação química das amostras palinológicas selecionadas foi realizada utilizando-se as técnicas de processamento palinológico descritas inicialmente por Uesugui (1979), que são as mesmas utilizadas por Dettmann (1963), Phipps & Playford (1984), Wood et al. (1996) e Antonioli (1998). A amostragem recaiu preferencialmente nas rochas de granulometria fina e coloração mais escura, por derivarem de sedimentos ricos em matéria orgânica e, em consequência, potencialmente portadores de maior volume de palinomorfos. Um mínimo de duas lâminas foi preparado para cada amostra examinada. As lâminas estudadas, contendo os espécimes aqui ilustrados, encontram-se depositadas no laminário palinológico do laboratório de palinomacerais da FGEL-UERJ. Com a utilização do fotomicroscópio Zeiss, e com uma câmera acoplada utilizando o programa Analisys, foram obtidas aproximadamente 200 fotos de palinomorfos. Os palinomorfos selecionados foram marcados através das coordenadas da lâmina England Finder. A análise qualitativa consistiu da individualização dos diferentes morfotipos presentes no material. Tais análises foram efetuadas através da observação das lâminas palinológicas em microscópio óptico. RESULTADOS E DISCUSSÃO Da associação palinológica recuperada foram reconhecidas 76 espécies de palinomorfos, sendo 34 pertencentes a esporos de briófitas e pteridófitas, 38 grãos de pólen de gimnospermas e angiospermas (Apêndice 1); além do reconhecimento de algas dulciaquícolas e da identificação de um exemplar do paleomicroplâncton marinho (dinoflagelado) identificado na porção intermediária da seção estudada. A composição da palinoflora identificada foi subdividida nos seguintes grupos taxonômicos: (i) talófitas (algas, fungos, liquens); (ii) briófitas (esporos de musgos e hepáticas); (iii) pteridófitas (esporos de samambaias, avencas, licófitas e esfenófitas); (iv) elementos gimnospérmicos (grãos de pólen monossacados, bissacados, monocolpados, poliplicados e rimulados); (v) angiospermas (grãos de pólen colpados, monosulcados e porados). As talófitas estão pobremente representadas na palinoflora. Em geral estão presentes na maioria das amostras, contudo em percentuais pouco significativos, sendo o gênero Botryococcus o mais representativo, o qual é usualmente interpretado como um membro das algas Chlorophyceae, de água doce. Entretanto, existe a possibilidade destes palinomorfos terem vivido também em águas salobras costeiras pouco agitadas (Batten & Grenfell, 1996). Em termos de representatividade dentro da seção, de um modo geral, predomina o grupo das gimnospermas seguido pelos esporos triletes (grupo das briófitas e pteridófitas). As angiospermas se apresentam com um menor registro na associação, com valores crescentes em direção ao topo da seção. Através das análises percentuais dos grupos polínicos, foram elaborados os gráficos, onde os esporos triletes estão presentes em praticamente toda a seção estudada, porém em baixa frequência, com valores percentuais de 10% em média (Figura 2). Foram identificados 24 gêneros e 34 espécies, sendo os mais comumente encontrados na associação os gêneros Deltoidospora, Cicatricosisporites e Crybelosporites. Em alguns níveis mais ao topo da seção, é possível perceber um significativo aumento no percentual do gênero Crybelosporites que chega a atingir valores de até 40%, indicando ambiente um pouco mais úmido nesta porção. Os grãos de pólen, afins às gimnospermas dominam praticamente toda a seção, com um percentual de representa- PORTELA ET AL. – CARACTERIZAÇÃO PALINOFLORÍSTICA E PALEOAMBIENTAL DA FORMAÇÃO SANTANA 365 Figura 1. Mapa de localização da bacia do Araripe, localização da área estudada e perfil litológico do poço 4-BO-1-PE (modificado de Assine, 1992). Figure 1. Location map of the Araripe Basin, location of studied area and lithological profile of the 4-BO-1-PE well (modified from Assine, 1992). tividade ao redor de 80%. Observa-se o alto valor percentual de grãos rimulados (Classopollis), na porção inferior da seção, mas que diminui para o topo (Figura 2). Paleoambiente As plantas são consideradas, de modo geral, como organismos bastante sensíveis às alterações dos fatores ecológicos terrestres. Andrews (1961) constatou que, sob condições de mudanças climáticas, as plantas são mais propensas a migrarem para um habitat mais favorável do que se adaptarem às novas condições em seu habitat original. Por conseguinte, mudanças climáticas podem alterar a flora, de modo significativo, num espaço de tempo relativamente curto. As plantas que viveram no Eocretáceo, na sua quase totalidade, são espécies e gêneros já extintos (Lima, 1978). Brenner (1976) e Herngreen & Chlonova (1981), utilizando uma ampla base de dados palinológicos para interpretar paleoclimas no Cretáceo, estabeleceram cinturões microflorísticos com significado climático. A área objeto do presente estudo, pelo seu conteúdo esporo-polínico – predomínio de grãos de pólen afins às Cheirolepidiaceae (Classopolis) que perfaz cerca de 80%; boa representatividade dos grupos Exesipollenites, Araucariacites e Inaperturopollenites e das formas poliplicadas dos gêneros Equisetosporites e Gnetaceaepollenites – e sua ocorrência geográfica (Brasil, Colômbia, Venezuela, Peru, Congo, Gabão, Egito, China e Israel), insere-se na província “Gondwana Norte” de Brenner (1976), que é essencialmente a mesma que a “província Cretáceo Inferior pré-albiana a Dicheiropollis etruscus/Afropollis” de Herngreen et al. (1996). Sedimentos cronocorrelatos aos da Formação Santana, no Brasil, são encontrados na quase totalidade das bacias da margem leste e equatorial brasileira, assim como em algumas bacias interiores, que foram investigadas por Hashimoto et al. (1987), Uesugui (1987), Regali (1989), Dino & Uesugui (1994) e Antonioli (1998). Estudo qualitativo, do material esporo-polínico ora descrito, mostra claramente a predominância de um clima quente em toda seção analisada. Estudo quantitativo mostra a tendência gradual no aumento da aridez e da temperatura em direção ao topo da Formação Santana. Isso se deve ao fato de que as formas indicativas de clima não tropical, como os grãos de pólen bissacados, são extremamente raras na associação. Em contrapartida, há uma abundância de formas típicas de clima quente, como Equisetosporites (Ephedrales), esporos de pteridófitas, como os do gênero Cicatricosisporites (Schizeaceae) e Crybelosporites pannuceus, além do predomínio de Classopollis, que apesar de associado ao grupo das coníferas, é comumente encontrado em condições de clima quente. 366 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014 Figura 2. Correlação entre o perfil litológico e a distribuição dos grupos dos esporos e grãos de pólen ao longo da seção estudada. Figure 2. Lithological profile and distribution of groups of spores and pollen grains. Quanto ao aspecto quantitativo mencionado, baseiase principalmente no aumento percentual do gênero Equisetosporites e das espécies Crybelosporites pannuceus e Afropollis jardinus, verificado nos níveis mais superiores da seção. Lima (1978) registrou na Formação Santana valores de até 55% desse grupo associado à abundância de Afropollis jardinus, que foram usados para indicar aumento da aridez e da temperatura. A caracterização dos grupos mais representativos da seção estudada, apresentada a seguir, tem como produto a identificação de condições paleoambientais e inferências paleoclimáticas para a bacia do Araripe. Esporos Triletes Presentes em quase toda a seção, embora sempre com baixa frequência. Habitam nichos úmidos próximos aos rios e lagos de águas rasas. Produzidos por várias famílias de samambaias, muitos destes esporos lisos não são associados a uma família específica, mas entre elas, certamente estão as famílias Cyatheaceae, Dicksoniaceae, Matoniaceae e Osmundaceae (Van Konijnenburg-Van Cittert, 1978) (Figura 3). Atualmente, a maioria destas famílias ocorre em áreas subtropicais e tropicais úmidas. Alvin (1974) indica que alguns membros da família Matoniaceae (Matonisporites) e Anemiaceae (Cicatricosisporites) são tolerantes a regiões sujeitas a períodos de seca extrema. Convém lembrar que a resistência da Anemiaceae à alternância de períodos úmidos e secos no Cretáceo já havia sido observada por Duarte et al. (2012). Inaperturados Abundantes em quase todo o intervalo estudado. Indicativos de climas secos, habitando regiões tropicais a subtropicais. Segundo Krassilov (1978), um de seus representantes, as araucariaceas, dominam cinturões de baixas latitudes em climas de verão seco com formação de red-beds. As podocarpaceas representadas pelos grãos de pólen Callialasporites (Gamerro, 1965) (Figura 3) são encontradas nestes tipos de clima, podendo também habitar regiões frias e úmidas. Rimulados Os rimulados estão representados, em percentuais relativamente altos dentro da seção, pelo gênero Classopollis (Figura 3). São considerados como tendo sido produzidos por membros das Cheirolepidiaceae bem adaptados a regiões áridas de baixa paleolatitude, climas secos e possivelmente condições de solos salinos próximos à costa. Grupo predominante e bem distribuído dentro da seção, os macrofitofósseis de Cheirolepidiaceae (Brachyphyllum, Frenelopsis, etc.) encontrados nessa unidade possuem características xeromórficas. Existem claras evidências palinológicas e sedimentares de que membros desta família eram halófilos (Upchurch & Doyle, 1981; Vakhrameev, 1970), e sua morfologia é também consistente com adaptação à salinidade. Entretanto, a grande variedade morfológica de membros das Cheirolepidiaceae e ocorrência de grãos de pólen e macrorrestos em depósitos continentais sugere que esta família não estava restrita a habitat costeiro, mas sim adaptada a amplas áreas, variando de costeira a terras altas (Alvin et al., 1978; Batten, 1974; Upchurch & Doyle, 1981). Poliplicados Representados por grãos de pólen da família das Ephedra, do grupo das Gnetales, cujos representantes atuais são os gêneros xeromórficos Ephedra e Welwitschia. Segundo Doyle et al. (1982), as plantas produtoras destes grãos poliplicados adaptam-se bem a regiões arenosas e pantanosas, em ambientes flúvio-deltaicos. Devido ao seu caráter xerofítico, este grupo tem sido usado como uma evidência de aridez na província Gondwana Norte. Estão presentes, com pelo menos, um representante das 15 espécies distribuídas nos gêneros Equisetosporites, Singhia, Gnetaceaepollenites, Steevesipollenites e Eucommiidites em cada nível da seção, com destaque para o gênero Equisetosporites (Figura 4). Bissacados Grupo com uma ocorrência muito baixa na associação, representado pelo gênero Vitreisporites (Figura 3). PORTELA ET AL. – CARACTERIZAÇÃO PALINOFLORÍSTICA E PALEOAMBIENTAL DA FORMAÇÃO SANTANA 367 C A B G D E I K F H L P M Q J O N R S Figura/Figure 3. A, Cyathidites australis Couper, 1953; B, Deltoidospora psilostoma (Kara-Murza) Singh, 1964; C, Leptolepidites verrucatus Couper, 1953; D, Cicatricosisporites avnimelechi Horowitz, 1970; E, Cicatricosisporites brevilaesuratus Couper, 1958; F, Matonisporites silvai Lima, 1979; G, Pilosisporites trichopapillosus (Thiegart) Delcourt & Sprumont, 1955; H, Apiculatisporis sp.1 Dino, 1992; I, Klukisporites pseudoreticulatus Couper, 1958; J, Camarozonosporites rudis (Leschick) Klaus, 1960; K, Crybelosporites pannuceus (Brenner) Srivastava, 1975; L, Araucariacites australis Cookson, 1947; M, Uesuguipollenites callosus Dino, 1994; N, Circulina meyeliana Klaus, 1960; O, Inaperturopollenites simplex Regali, Uesugui & Santos, 1974; P, Callialasporites dampieri (Balme) Dev, 1961; Q, Sergipea variverrucata (Regali, Uesugui & Santos, 1974) Regali, 1987; R, Vitreisporites sp. Dino, 1992; S, Classopollis classoides Pflug, 1953. Escala/Scale bar = 20 μm REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014 368 B E A C D J G H I K F M L N O P Q R S Figura/Figure 4. A, Equisetosporites ambiguus (Hedlund) Singh, 1971; B, Steevesipollenites pentacostatus Regali, 1989; C, Singhia reyrei Lima, 1980; D, Equisetosporites concinnus Singh, 1964; E, Gnetaceaepollenites pentaplicatus Regali, 1989; F, Gnetaceaepollenites jansonii (Pocock) Lima, 1980; G, Gnetaceaepollenites barghoornii Pocock, 1964; H, Bennettitaepollenites sp. 1 Antonioli, 1998; I, Bennettitaepollenites regaliae Dino, 1994; J, Eucommiidites sp. 2 Lima, 1978; K, Brenneripollis reticulatus (Brenner) Júhasz & Góczán, 1985; L, Reyrea polymorphus Herngreen, 1973; M, Retimonocolpites textus (Norris) Singh, 1983; N, Stellatopollis dubius Jardiné & Magloire, 1965, emend. Lima, 1978; O, Afropollis jardinus (Brenner) Doyle, Jardiné & Doerenkamp, 1982; P, Chomotriletes almegrensis Pocock, 1962; Q, Maranhites sp.; R, Botryococcus braunii Kutzing, 1849; S, Subtilisphaera sp. Escala/Scale bar = 20 μm. PORTELA ET AL. – CARACTERIZAÇÃO PALINOFLORÍSTICA E PALEOAMBIENTAL DA FORMAÇÃO SANTANA Angiospermas Grupo bem representado na associação pelo gênero Afropollis (Figura 4). Em termos quantitativos, está mais bem representado nos níveis mais superiores da seção. Habitantes de vários ambientes costeiros, inclusive mangue, com amplo espectro climático (Doyle et al., 1982). Dinoflagelados Grupo representado com apenas um exemplar (Figura 4). A ocorrência de cisto de dinoflagelados já havia sido detectada nos sedimentos pertencentes à Formação Santana, especialmente nos calcários laminados do Membro Crato (Lima, 1978). Cistos de dinoflagelados, descritos por Lima (1971), atribuídos ao gênero Subtilisphaera, fornecem um valioso subsídio para interpretações paleoecológicas dos níveis que os contém, pois sugerem a ocorrência de pequenas ingressões marinhas. Convém lembrar que Arai & Coimbra (1990) identificaram vários níveis com a predominância de cistos de Subtilisphaera dentro da Formação Santana, os quais sugerem a ocorrência de florações fitoplanctônicas (blooms) em ambiente marinho costeiro. CONCLUSÕES A associação palinológica identificada mostra que o início da deposição predominava uma palinoflora essencialmente arbórea, com componentes arbustivos e herbáceos associados. Observa-se, na porção mais superior da seção, aumento dos grãos de pólen inaperturados, representados principalmente pelo gênero Araucariacites. São também comuns, em toda a seção, os grãos rimulados e os esporos triletes, destacando-se formas cicatricosas. A porção intermediária da seção mostra relativo aumento no percentual de formas arbustivas, e elevado acréscimo no número de representantes dos grãos rimulados, assim como maior diversidade de esporos triletes. Os grãos de pólen poliplicados tornam-se mais diferenciados e um pouco mais abundantes. No topo da seção observa-se escassez dos componentes arbóreos, sendo substituídos por vegetação arbustiva, representada pela abundância do gênero Afropollis. Observase agora decréscimo, muito significativo, na distribuição dos grãos de pólen rimulados e significativo aumento na diversidade dos grãos de pólen poliplicados, colpados e esporos perisporados. Em termos quantitativos, dominam os rimulados, com valores ao redor de 60% da associação, seguidos dos inaperturados, que atingem frequência da ordem dos 30%, e dos esporos triletes, com valores da ordem de 10%. Grãos de pólen com afinidades angiospérmicas constituem frações inferiores a 5% do total de espécies que caracterizam as associações. A microflora é muito similar às associações pertencentes à mesma faixa microflorística de outras partes do mundo (Colômbia, Venezuela, Peru, Congo, Gabão, Egito, China e Israel). Insere-se perfeitamente nos cinturões microflorísticos “Gondwana Norte” e “África Ocidental/América do Sul” (WASA) definidos por Brenner (1976) e Herngrenn & Chlonova (1981), respectivamente. 369 Evidências palinológicas e sedimentológicas indicam que a deposição desses sedimentos se deu em ambiente continental, com pequenas ingressões marinhas, marcado pela presença apenas discreta de dinoflagelado (Portela, 2008), o que deve representar um contexto ambiental diferente daquele estudado por Arai & Coimbra (1990). A presença de elementos higrofíticos e xerofíticos identifica uma palinoflora de clima quente, árido a semiárido, com nichos úmidos, em latitude próxima à atual (Antonioli & Dino, 2007). AGRADECIMENTOS A ultima autora agradece ao CNPq pela bolsa de produtividade em pesquisa, processo 306609/2012-6. REFERÊNCIAS Antonioli, L. 1998. Bioestratigrafia, palinologia e paleoambientes da seção Aptiana Eoalbiana da Bacia do Ceará, Nordeste do Brasil. Programa de Pós-Graduação em Análises de Bacias e Faixas Móveis, Universidade do Estado do Rio de Janeiro, Dissertação de Mestrado, 107 p. Antonioli, L. 2001. Estudo palinocronoestratigráfico da Formação Codó, Cretáceo Inferior do Nordeste brasileiro. Programa de Pós-Graduação em Geologia, Universidade Federal do Rio de Janeiro, Tese de Doutorado, 265 p. Antonioli, L. & Dino, R. 2007. Análise palinoestratigráfica da Formação Codó Cretáceo Inferior da Bacia do Parnaíba - NE do Brasil. In: I.S. Carvalho; R.C. Cassab; C. Schwanke; M.A. Carvalho; A.C.S. Fernandes; M.A.C. Rodrigues; M.S.S. de Carvalho; M. Arai & M.E.Q. Oliveira (orgs.) Paleontologia: Cenários de Vida, Interciência, vol. 2, p. 533-546. Alvin, K.L. 1974. Leaf anatomy of Weichselia based on fusainized material. Palaeontology, 17:587-598. Alvin, K.L.; Spicer, R.A. & Watson, J. 1978. A Classopolliscontaining male cone associated with Psedofrenelopsis. Palaeontology, 21:847-856. Andrews, H.N. 1961. Studies in Palaeobotany. New York, John Wiley & Sons, 487 p. Arai, M. & Coimbra, J.C. 1990. Análise paleoecológica do registro das primeiras ingressões marinhas na Formação Santana (Cretáceo Inferior da Chapada do Araripe). In: SIMPÓSIO SOBRE A BACIA DO ARARIPE E BACIAS INTERIORES DO NORDESTE, 1, 1990. Comunicações, Crato, DNPM, p. 225-239. Assine, M.L. 1992. Análise estratigráfica da Bacia do Araripe, Nordeste do Brasil. Revista Brasileira de Geociências, 22:289300. Assine, M.L. 2007. Bacia do Araripe. Boletim de Geociências da Petrobras, 15:371-389. Batten, D.J. 1974. Wealden paleoecology from the distribution of plant fossils. Proceedings of the Geologists’ Association, 85: 433-458. doi:10.1016/S0016-7878(74)80068-4 Batten, D.J. & Grenfell, H.R. 1996. Botryococcus. In: J. Jansonius & D.C. McGregor (eds.) Palynology: principles and applications, AASP Foundation, p. 205-212. Brenner, G.J. 1976. Middle Cretaceous floral provinces and migrations of angiosperms. In: C.B. Beck (ed.) Origin and evolution of angiosperms, Columbia University Press, p. 23-47. 370 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014 Brito Neves, B.B. 1990. Bacia do Araripe no contexto geotectônico regional: In: SIMPÓSIO SOBRE A BACIA DO ARARIPE E BACIAS INTERIORES DO NORDESTE, 1, 1990. Comunicações, Crato, DNPM, p. 21-33. Dettmann, M.E. 1963. Upper Mesozoic microfloras from southeastern Australia. Proceedings of the Royal Society of Victoria, 77:1-148. Dino, R.; Silva, O.B. & Abrahão, D. 1999. Caracterização palinológica e estratigráfica de estratos cretáceos da Formação Alter do Chão, Bacia do Amazonas: In: SIMPÓSIO SOBRE O CRETÁCEO, 5, 1999. Boletim de Resumos, Rio Claro, UNESP, p. 557-565. Dino, R. & Uesugui, N. 1994. Bacias do Recôncavo, Tucano e Jatobá. Boletim de Geociências da Petrobras, 8:163-172. Doyle, J.A.; Jardiné, S. & Doerenkamp, A. 1982. Afropollis, a new genus of early angiosperm pollen, with notes on the Cretaceous palynostratigraphy and paleoenvironments of Northern Gondwana. Bulletin des Centres des Recherches ExplorationProduction Elf-Aquitaine, 6:39-117. Duarte, S.G.; Arai, M. & Wanderley, M.D. 2012. Significado paleoambiental da família Anemiaceae no Cretáceo do Brasil: inferências a partir de dados palinológicos. Revista do Instituto Geológico, 33:1-12. Gamerro, J.C. 1965. Morfología del pollen de Apterocladus lanceolatus Archang. (Coniferae) de la Formación Baqueró, provincia de Santa Cruz. Ameghiniana, 4:133-138. Hashimoto, A.T.; Appi, C.J.; Soldan, A.L. & Cerqueira, J.R. 1987. O neo-Alagoas nas bacias do Ceará, Araripe e Potiguar (Brasil): caracterização estratigráfica e paleoambiental. Revista Brasileira de Geociencias, 17:118-122. Herngreen, G.F.W. & Chlonova, A.F. 1981. Cretaceous microflora provinces. Pollen et Spores, 23:241-555. Herngreen, G.F.W; Kedves, M.; Rovnina, L.V. & Smirnova, S.B. 1996. Cretaceous palynofloral provinces: a review. In: J. Jansonius & D.C McGregor (eds.) Palynology: principles and applications, AASP Foundation, p. 1157-1188. Krassilov, V.A. 1978. Araucariaceae as indicators of climate and paleolatitudes. Review of Palaeobotany and Palynology, 26:113124. doi:10.1016/0034-6667(78)90008-8 Lima, M.R. 1971. Contribuição palinológica à Estratigrafia do Cretáceo do Nordeste do Brasil: análise da perfuração GSB, Serra Branca, PE. Programa de Pós-Graduação em Geociências, Universidade Federal do Rio Grande do Sul, Dissertação de Mestrado, 74 p. Lima, M.R. 1978. Palinologia da Formação Santana (Cretáceo do Nordeste do Brasil). Programa de Pós-Graduação em Geologia Sedimentar, Universidade de São Paulo, Tese de Doutorado, 335 p. Phipps, D. & Playford, G. 1984. Laboratory techniques for extraction of palynomorphs from sediments. Papers of the Department of Geology, 11:1-23. Ponte, F.C. & Appi, C.J. 1990. Proposta de revisão da coluna litoestratigráfica da Bacia do Araripe. In: CONGRESSO BRASILEIRO DE GEOLOGIA, 36, 1990. Anais, Natal, SBG, p. 211-226. Portela, H.A. 2008. Estudo palinológico e palinofaciológico da Formação Santana, Bacia do Araripe, Nordeste do Brasil. Programa de Pós-Graduação em Análises de Bacias e Faixas Móveis, Universidade do Estado do Rio de Janeiro, Dissertação de Mestrado, 108 p. Regali, M.S.P. 1989. Evolução da paleoflora no Cretáceo das margens equatorial e nordeste do Brasil. Revista da Escola de Minas, 42:17-33. Srivastava, S.K. 1978. Cretaceous spore-pollen floras: a global evaluation. In: Biological Memoirs, Lucknow, International Publishers, 130 p. (Palaeopalynology Series 5). Uesugui, N. 1979. Palinologia-técnicas de tratamento de amostras. Boletim Técnico Petrobras, 22:229-240. Uesugui, N. 1987. Posição estratigráfica dos evaporitos da Bacia de Sergipe-Alagoas. Revista Brasileira de Geociências, 17:131134. Upchurch, G.R. & Doyle, J.A. 1981. Paleoecology of the conifers Frenelopsis and Pseudodofrenelopsis (Cheirolepidiaceae) from the Cretaceous Potomac Group of Maryland and Virginia. In: R.C. Romans (ed.) Geobotany II, Springer, p. 167-202. Vakhrameev, V.A. 1970. Range and paleoecology of Mesozoic conifers, the Cheirolepidiaceae. Paleontology Journal, 4:11-25. Van Konijnenburg-Van Cittert, J.H.A. 1978. Osmudaceous spores in situ from the Jurassic of Yorkshire, England. Review of Palaeobotany and Palynology, 26:125-141. doi:10.1016/00346667(78)90009-X Viana, M.S.S. & Neumann, V.H.L. 2002. Membro Crato da Formação Santana, Chapada do Araripe, CE - riquíssimo registro de fauna e flora do Cretáceo. In: C. Schobbenhaus; D.A. Campos; E.T. Queiroz; M. Winge & M.L.C. Berbert-Born (eds.) Sítios Geológicos e Paleontológicos do Brasil, DNPM/CPRM/SIGEP, p. 113-120. Wood, G.D.; Gabriel, A.M. & Lawson, J.C. 1996. Palynological techiniques – processing and microscopy. In: J. Jansonius & D.C. McGregor (eds.) Palynology: principles and applications, AASP Foundation, p. 29-50. Received in January, 2014; accepted in August, 2014. PORTELA ET AL. – CARACTERIZAÇÃO PALINOFLORÍSTICA E PALEOAMBIENTAL DA FORMAÇÃO SANTANA Apêndice 1. Lista de taxon identificados no presente estudo. Appendix 1. List of taxa identified in this study. SPORITES H. Potonié, 1893 Todisporites cinctus Orlowskazwolinska (Malyawkina), 1964 Leiotriletes sp. Cyathidites australis Couper, 1953 Cyathidites crassiangulatus Balme, 1957 Concavisporites sp. Regali, Uesugui & Santos, 1974 Deltoidospora hallii Miner, 1935 Deltoidospora juncta Rouse, 1959 Deltoidospora psilostoma (Kara-Murza) Singh, 1964 Biretisporites potoniaei Delcourt & Sprumont, 1955 Biretisporites deltoidus Dettmann (Rouse), 1963 Undulatisporites undulapolus Brenner, 1963 Leptolepidites verrucatus Couper, 1953 Leptolepidites tumulosus (Döring) Srivastava, 1975 Verrucosisporites sp. 2 Dino, 1992 Apiculatisporis sp. 1 Dino, 1992 Pilosisporites trichopapillosus (Thiegart) Delcourt & Sprumont, 1955 Cicatricosisporites avnimelechi Horowitz, 1970 Cicatricosisporites brevilaesuratus Couper, 1958 Cicatricosisporites nuni Horowitz, 1970 Cicatricosisporites purbekensis Norris, 1969 Cicatricosisporites sp. 1 Dino, 1992 Cicatricosisporites microstriatus Jardiné & Magloire, 1965 Klukisporites foveolatus Pocock, 1964 Klukisporites pseudoreticulatus Couper, 1958 Hamulatisporis albertensis Srivastava, 1972 Cicatricososporites auritus Singh, 1971 Camarozonosporites insignis Norris, 1967 Camarozonosporites rudis (Leschick) Klaus, 1960 Antulsporites sp. cf. A. baculatus Archangelsky & Gamerro, 1966 Matonisporites silvai Lima, 1979 Appendicisporites tricornitatus Weiland & Greifeld, 1953 Rouseisporites reticulatus Pocock, 1962 Densoisporites dettmannae (Srivastava, 1972) Dejax, 1987 Crybelosporites pannuceus (Brenner) Srivastava, 1975 Crybelosporites mamelonatus Lima, 1978 POLLENITES R. Potonié, 1931 Callialasporites dampieri (Balme) Dev, 1961 Callialasporites segmentatus Balme, 1957 Callialasporites lucidus (Pocock) Reyre, 1973 Vitreisporites sp. Dino, 1992 Inaperturopollenites turbatus Balme 1957 (sensu Regali, Uesugui & Santos, 1974) Inaperturopollenites simplex Regali, Uesugui & Santos, 1974 Araucariacites australis Cookson, 1947 Uesuguipollenites callosus Dino, 1994 Sergipea variverrucata (Regali, Uesugui & Santos, 1974b) Regali, 1987 Classopollis classoides Pflug, 1953 Circulina meyeliana Klaus, 1960 Equisetosporites dudarensis (Deák) Lima, 1978 Equisetosporites albertensis Singh, 1964 Equisetosporites ambiguus (Hedlund) Singh, 1971 Equisetosporites concinnus Singh, 1964 Singhia reyrei Lima, 1980 Gnetaceaepollenites jansonii (Pocock) Lima, 1980 Gnetaceaepollenites retangularis Lima, 1980 Gnetaceaepollenites chlatratus Stover, 1964 Gnetaceaepollenites barghoornii Pocock, 1964 Gnetaceaepollenites pentaplicatus Regali, 1989 371 372 REVISTA BRASILEIRA DE PALEONTOLOGIA, 17(3), 2014 Apêndice 1. Continuação. Appendix 1. Continuation. Gnetaceaepollenites oreadis Srivastava, 1968 Steevesipollenites sp. 1 Lima, 1980 Steevesipollenites pentacostatus Regali, 1989 Steevesipollenites grambasti Azema & Boltenhagem, 1974 Steevesipollenites cupuliformis Azema & Boltenhagem, 1974 Eucommiidites sp. 2 Lima, 1978 Cycadopites sp. 4 Antonioli, 1998 Bennettitaepollenites regaliae Dino, 1994 Bennettitaepollenites sp. 1 Antonioli, 1998 Stellatopollis densiornatus (Lima) Lima, 1978 Stellatopollis araripensis (Lima) Lima, 1978 Stellatopollis dubius Lima, 1978 Retimonocolpites textus (Norris) Singh, 1983 Afropollis jardinus (Brenner) Doyle, Jardiné & Doerenkamp, 1982 Brenneripollis reticulatus (Brenner) Júhasz & Góczán, 1985 Reyrea polymorphus Herngreen, 1973 Chomotriletes almegrensis Pocock, 1962
Download