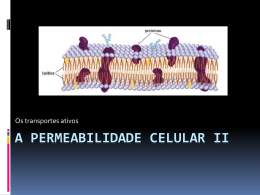
ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br APOSTILA DE FISIOLOGIA 1. INTRODUÇÃO Sabemos que o envoltório celular, a chamada membrana plasmática, possui em sua constituição lipídica proteínas diversas, de variados tipos e tamanhos, o que lhe caracteriza uma imagem mosaica. Mas qual a importância destas proteínas? Principalmente, estes elementos participam de transportes, fluxo de nutrientes e íons do meio intra-celular para o meio extra-celular e vice e versa, função de extrema importância tanto para a sobrevivência celular como também para a manutenção da homeostasia do organismo. É através destes pequenos movimentos de íons e substâncias que se realiza todo o funcionamento do nosso corpo, porém, para que haja estes fluxos, se faz necessária grande quantidade de energia. E de onde vem toda essa energia? Todo o alimento que ingerimos, é convertido através de reações metabólicas em moléculas de energia. Conceitos: • Metabolismo: Anabolismo + Catabolismo • Anabolismo: É a ação de incorporar nutrientes para processar substâncias e extrair substratos necessários para a manutenção das funções corporais. • Catabolismo: Fragmentação enzimática de moléculas grandes, que são reduzidas em moléculas pequenas a fim de serem oxidadas pelas células e posteriormente, eliminadas. • ATP: é a forma que o organismo encontrou para armazenar energia (trifosfato de adenosina). São Três fósforos (P) ligados a uma adenosina. A energia é armazenada entre as ligações de fosfato com a adenosina e pode ser liberada independentemente. O nosso organismo é capaz de gerar energia através de duas vias, via aeróbica (ou oxidativa – com presença de oxigênio) e via anaeróbica (ou glicolítica – sem presença de oxigênio). A via glicolítica é uma seqüência de 10 reações enzimáticas que quebram uma molécula de glicose em 2 ATPs + ácido pirúvico. Com a presença de oxigênio, o ácido pirúvico presente no citoplasma celular (ou piruvato) entra na mitocôndria, onde sofre diversas reações enzimáticas (Ciclo de Krebs), dando origem a 36 moléculas de ATPs. Esta última é a chamada via oxidativa. Sem a presença de oxigênio, o ácido pirúvico em excesso gera o ácido lático (ou lactato), principal causador das dores musculares após as atividades físicas. Portanto, para que seja formada energia em grande quantidade é necessário a presença de oxigênio. Etapas: 1. A glicose é ingerida na alimentação e através da corrente sangüínea é liberada no líquido intersticial. 2. A insulina é um hormônio que estimula os receptores de membrana, facilitando a entrada da glicose na célula através da difusão facilitada. 3. Dentro da célula, a glicose sofre ação de enzimas citoplasmáticas gerando 2 moléculas de ATP + ácido pirúvico 4. O ácido pirúvico é levado através de transportadores até a mitocôndria, onde entra no ciclo de Krebs formando, o ácido oxalacético e 36 moléculas de ATP. 5. O ácido pirúvico não utilizado sobra e, devido a um déficit de oxigênio, gera o ácido lático. Como já mencionado, o ATP é necessário para qualquer função do nosso organismo, seja para a contração muscular, para o impulso nervoso, para a manutenção celular etc. A energia está armazenada entre as ligações da adenosina com os fosfatos. Assim que é quebrada uma destas ligações e é liberada energia, o ATP (trifosfato de adenosina) é convertido em ADP (difosfato de adenosina). Se o ADP for utilizado novamente liberando mais energia, passa a ser chamado de AMP (monofosfato de adenosina). Após a quebra da ligação fosfato/adenosina o fosfato liberado é adicionado a um composto denominado fosfocreatina (creatina + fosfato) formando uma reserva energética. No momento em que for necessária energia rápida sem a presença de oxigênio uma enzima denominada creatina knase extrai o fosfato do composto, adicionando-o a um ADP formando de uma forma bastante rápida o ATP. A este processo denominamos ressíntese de ATP. Profª Cíntia Schneider 1 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br No organismo este processo serve para repor rápidamente os níveis de ATP celular, como no músculo esquelético e cardíaco, permitindo que ele ainda funcione por algum tempo mesmo que não haja oxigênio disponível. Este é um processo que decorre sem a necessidade de oxigênio, porém, este processo não consegue se manter por muito tempo. 2. ORGANISMO EM HOMEOSTASE Homeostasia: Permanente tendência do organismo de manter a constância do meio interno. Relativa independência do organismo em relação às oscilações do ambiente externo. O nosso organismo possui a capacidade de manter-se em equilíbrio, de manter uma constância no meio interno, com valores funcionais, como por exemplo pressão arterial = 120/80 mmHg ou pH do sangue = 7,4. Mas esses valores não são fixos ou seja, existe um valor médio, um intervalo entre o valor máximo e mínimo que mantém sempre em equilíbrio o meio interno, por exemplo, se o pH do sangue estiver 7,43 ou 7,35 não quer dizer que o organismo esteja em desequilíbrio, pois está dentro dos limites, porém se o pH estiver em 8,0 ou 6,5 já é considerado um desequilíbrio orgânico. Outro exemplo, os valores normais para parâmetros sanguíneos de glicose são 75 – 110 mg / 100ml; se o resultado de glicose for de 90mg / 100ml está normal, porém, se o resultado for 120 mg /100ml está fora dos parâmetros de equilíbrio. Dois sistemas atuam na manutenção da homeostase, o sistema nervoso e o sistema endócrino. Em resposta a um estímulo, o sistema nervoso apresenta ação rápida, em curto prazo e com efeito localizado sobre o ponto de desequilíbrio; já o sistema endócrino possui ação mais lenta, a médio e longo prazo, porém de ação duradoura e de efeito amplo sobra o organismo. Os dois sistemas agem de maneira integrada, garantindo a homeostase do organismo, tornando-o capaz de se relacionar com as mudanças do meio externo. Tanto o sistema nervo como o sistema endócrino agem através de sistemas de feedback positivos e negativos. O que é feedback? É um processo que visa restabelecer o equilíbrio do meio interno de modo constante. O aumento ou diminuição de uma função (pressão arterial), provoca uma alteração (física ou química) no organismo, e esta alteração desencadeia uma reação para a correção funcional, garantindo o equilíbrio dinâmico. Através dessas propriedades, o organismo torna-se estável e invariável diante das mudanças exteriores. Feedback Negativo Feedback negativo: é quando a alteração funcional se faz num sentido e a reação para a correção em outro, ou seja, a resposta do sistema de controle é oposta ao estímulo. A maior parte dos sistemas de controle do corpo atua por meio de feedback negativo. Na regulação da concentração de dióxido de carbono, a alta concentração de dióxido de carbono no líquido extracelular aumenta a ventilação pulmonar. Isso, por sua vez, reduz a concentração do dióxido de carbono, visto que, nessa condição, os pulmões eliminam maior quantidade de dióxido de carbono do corpo. Em outras palavras, a alta concentração produz redução da concentração, o que é negativo em relação ao estímulo desencadeante. De modo inverso, caso a concentração de dióxido de carbono caia a valores muito baixos, isso faz com que ocorra aumento por feedback dessa concentração. Essa resposta também é negativa em relação ao estímulo inicial. Profª Cíntia Schneider 2 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br Diversos sistemas contribuem para a regulação da pressão arterial. Um deles, o sistema baroceptor, é exemplo simples e excelente de um mecanismo de controle. Nas paredes da região onde ocorre a bifurcação das artérias carótidas, no pescoço, bem como no arco aórtico, existem muitos receptores neurais, chamados baroceptores, que são estimulados pelo estiramento da parede arterial. Quando a pressão arterial fica elevada, os baroceptores emitem descargas de impulsos para o bulbo, no encéfalo. Nesse local, esses impulsos inibem o centro vasomotor, que, por sua vez, reduz o número de impulsos transmitidos pelo sistema nervoso simpático para o coração e vasos sanguíneos. A falta desses impulsos diminui a atividade bombeadora do coração, além de maior facilidade do fluxo sangüíneo pelos vasos periféricos; esses dois efeitos fazem com que a pressão arterial caia até seus valores normais. De modo inverso, queda da pressão arterial relaxa os receptores de estiramento, permitindo que o centro vasomotor fique mais ativo que o usual, o que provoca a elevação da pressão arterial até seu valor normal. Feedback Positivo O estímulo inicial produz mais estimulação do mesmo tipo, é a retraoalimentação positiva observada em casos nos quais a alteração funcional e a reação se fazem no mesmo sentido, aumentando o desequilíbrio. Ex: # Coagulação do sangue: o rompimento de um vaso sanguíneo dá início à formação do coágulo, que por sua vez ativa diversas enzimas – “fatores de coagulação”- que atuam sobre outras enzimas, ativando o crescimento do coágulo, ativando mais enzimas... Esse processo continua até que a ruptura vascular seja ocluída. # Contrações uterinas: aumento das contrações – cabeça do feto força a passagem pela cérvix – estiramento da cérvix – sinais enviados de volta ao corpo uterino – contrações uterinas mais fortes... Até a expulsão do feto. OBS: O Feedback Positivo causa, por vezes, ciclos viciosos e morte. Ex: # Diminuição do volume sanguíneo (perda de sangue) – diminuição da pressão arterial; diminuição do fluxo sanguíneo p/ o coração – enfraquecimento cardíaco; diminuição do bombeamento cardíaco; diminuição do fluxo sanguíneo coronário (arterial); enfraquecimento cardíaco... Esse ciclo se repete até a morte. Para que haja controle do equilíbrio do meio interno, é necessária uma comunicação contínua entre as células deste organismo, e para que ocorra esta comunicação se faz necessário “portas” de entrada e saída de substâncias, e este fluxo ocorre através das proteínas presentes na membrana plasmática das células. 3. MEMBRANA PLAMÁTICA E TRANSPORTES Estão presentes na membrana plasmáticas dois tipos de proteínas: *proteínas intrínsecas – endoproteína, ectoproteína e transmembrana; *proteínas extrínsecas – interna e externa. Os tipos principais de proteínas participantes de transportes são as do tipo transmembrana, que atuam como um canal de passagem do meio intra-celular para o extra-celular e viceversa. Mas por que são necessários estes canais para a entrada e saída de substâncias? Simplesmente porque há certos tipos de substâncias que não conseguem ultrapassar a membrana celular; substâncias como gases e moléculas pequenas sem cargas elétricas ultrapassam facilmente a camada celular, porém moléculas grandes (como a glicose) ou moléculas com cargas elétricas (Na+; Cl-) não passam pela membrana celular, e nestes casos é necessário um canal específico, que facilite a entrada ou saída destas moléculas. A este fluxo de substância chamamos de transportes de membrana. Existem três tipos de transporte: Difusão simples: transporte de soluto que ocorre a favor do gradiente de concentração de determinada substância, do local de maior concentração para o local de menor concentração, sem que se faça necessária a presença de proteínas carreadoras para facilitar o fluxo desta substância. Profª Cíntia Schneider 3 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br Proteínas carreadoras: proteínas que mudam sua conformação para transportar moléculas muito grandes ou carregadas eletricamente. Ex: As moléculas X passam do compartimento onde estão em maior quantidade (A) para onde então em menor quantidade (B). Transporte Passivo (difusão facilitada): transporte de soluto que ocorre a favor do gradiente de concentração, do local onde a substância está mais concentrada para o local onde está menos concentrada, porém neste caso, com o auxilio de proteínas carreadoras e canais iônicos, pois estas moléculas são muito grandes para ultrapassar a membrana celular, ou possuem cargas elétricas. Transporte Ativo: é o transporte de soluto que ocorre contra o gradiente de concentração, com gasto energético, do local onde está menos concentrado para o local mais concentrado. Osmose: é o transporte de solvente (líquido) do local onde há menos concentração para onde há mais concentração. Profª Cíntia Schneider 4 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br 4. BOMBA DE SÓDIO E POTÁSSIO As células em geral possuem íons dissolvidos em solução sendo então dotadas de cargas elétricas. Os principais grupos portadores de carga elétrica que aparecem no ambiente celular são os íons Na+; K+; Cl-; e A- (ânions orgânicos carregados, como proteínas e aminoácidos). A concentração desses íons é diferente em relação ao ambiente intracelular e o extracelular. Dessa forma, o interior da célula possui uma maior concentração de íons K+ e A- tornando a carga elétrica líquida resultante negativa. O exterior possui uma concentração maior de íons Na+ e Cl- com uma carga elétrica resultante positiva. Essa diferença de polaridade gera uma diferença de potencial elétrico através da membrana celular, chamado de potencial de membrana, sendo definido como a diferença de potencial entre o ambiente intracelular e o extracelular. Devido à diferença de concentração de cada íon entre os ambientes intracelular e extracelular, surge uma força motriz que faz com que esses íons atravessem a membrana celular de forma a equilibrar o gradiente de concentração. Para o potássio, a direção do fluxo de íons é do seu interior para o exterior. Porém, à medida que os íons potássio abandonam o interior da célula, a carga elétrica resultante acaba ficando cada vez mais negativa. Assim, os íons potássio acabam sendo carregados novamente para o interior da célula devido a forças de natureza elétrica (negativo – atrai positivo +). Esse fluxo de íons potássio continua até o momento que as forças de natureza elétrica são compensadas devido à força gerada pelo gradiente de concentração. Para o sódio, o fluxo de íons devido ao gradiente de concentração (potencial químico) possui o sentido do exterior para o interior celular. Nesse caso, como o interior da célula possui carga negativa, a força elétrica acaba carregando os íons sódio para o interior da célula. Assim, os fluxos de íons sódio para o interior da célula são regidos pelos potenciais químicos e elétricos e nenhum fenômeno passivo arrasta os íons para fora da célula. Para os ânions orgânicos, essa transferência através da membrana é dificultada devido aos seus grandes tamanhos moleculares. Os íons cloreto possuem uma contribuição na difusão discreta em relação aos outros grupos já mencionados. Em repouso, os íons potássio e sódio atravessam a membrana através de canais específicos para cada um desses íons, chamados canais de vazamento. Os íons potássio atingem um estado de equilíbrio onde as forças elétricas e químicas se anulam e não ocorre mais fluxo de potássio pela membrana. Já para o sódio, as forças elétricas e químicas se somam e não existe forma de controlar o fluxo. Porém, o número de canais abertos para o transporte de potássio é quase 30 vezes maior que para o sódio, o que não permite que o fluxo de ambos os íons seja tão desequilibrado. Como a célula necessita manter o seu potencial de repouso (negativo no meio intracelular e positivo no meio extracelular), deve existir um mecanismo que transporte os íons potássio e sódio contra os seus gradientes de concentração, fazendo com que a célula mantenha a sua polaridade e seu potencial constante. Esse mecanismo recebe o nome de bomba de sódio-potássio. O funcionamento dessa bomba está baseado em uma proteína (Na+K+ATPase) de elevado peso molecular que atravessa o interior e o exterior da membrana. No lado interno da célula, essa proteína possui sítios de ligação para o sódio, e no exterior, sítios de ligação para o potássio. Assim, a proteína no interior da célula se liga ao sódio carregando-o para o exterior celular. No lado exterior, a molécula sofre uma mudança na sua conformação tornando-se agora capaz de ligar-se com os íons potássio. Para cada três íons sódio transportados para fora, dois íons potássio são transportados para dentro (três íons sódio saem, enquanto 2 íons potássio entram). Isso estabelece a diferença de potencial exigida pela membrana, sendo mais negativo o lado intracelular e mais positivo o meio extracelular. Como a saída de sódio não é acompanhada pela entrada de potássio na mesma proporção, estabelece-se uma diferença de cargas elétricas entre os meios intra e extracelular: há déficit de cargas positivas dentro da célula e as faces da membrana mantêm-se eletricamente carregadas. O potencial eletronegativo criado no interior da célula devido à bomba de sódio e potássio é chamado potencial de repouso da membrana, ficando o exterior da membrana positivo e o interior negativo. Dizemos, então, que a membrana está polarizada. Profª Cíntia Schneider 5 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br Quando estimulada (por um impulso nervoso), uma pequena região da membrana torna-se mais permeável aos íons sódio (abertura dos canais voltagem-dependente de sódio). Como a concentração desse íon é maior fora do que dentro da célula, o sódio atravessa a membrana no sentido do interior da célula. Neste momento, a entrada de grande quantidade sódio é acompanhada pela pequena saída de potássio. Este fluxo de íons inverte o potencial da membrana, ou seja, o meio intra-celular passa a ser positivo e o meio extra-celular torna-se negativo. Esta inversão de potencial é a despolarização, a célula que antes era polarizada (negativamente) despolariza e torna-se positiva internamente. Essa positividade (despolarização) induz o fechamento dos canais voltagem-dependente para o sódio, determinando a parada do fluxo deste íon para o interior da célula. Neste momento, abrem-se canais voltagemdependente de potássio, que, devido ao gradiente de concentração, migra fora da célula. Devido à lentidão da abertura dos canais voltagem-dependente para o potássio, eles só se abrem quando os canais voltagemdependente para o sódio já estão se fechando. Assim, a diminuição do influxo de sódio para o interior da célula e o aumento simultâneo de efluxo de potássio para fora da célula aceleram o processo de repolarização, ou seja, a célula volta a apresentar o meio intracelular negativo e o meio extracelular positivo. Em alguns casos ocorre a lentidão do fechamento ou da abertura dos canais voltagem-dependente. Quando esta demora se dá no fechamento nos canais para o potássio, ocorre a difusão em quantidade excessiva deste íon para fora da célula, criando um déficit de cargas positivas no meio intracelular, ou seja, uma hiperpolarização. Se a demora no fechamento ocorrer nos canais para sódio (chamados canais lentos) e ao mesmo tempo houver demora em abrir os canais voltagem-dependente para o potássio, ocorre o que chamamos de platô, ou seja, uma demora na repolarização. TERMOS: Potencial de repouso (potencial transmembrana) Devido à diferença de concentração entre os meios intra e extracelular forma-se uma ddp (diferença de potencial) entre o interior da célula e o meio extracelular. Este valor é em média = -85 mV e recebe o sinal negativo (-) por convenção (o interior da célula tem grande quantidade de ânions protéicos). Nessa situação a célula é dita polarizada. Essa característica é comum a todas as células do organismo na ausência de estímulos eficazes. Excitabilidade celular É a propriedade que a célula possui de alterar o seu potencial de repouso quando submetida a estímulos eficazes. Despolarização celular: entrada de sódio Quando uma célula recebe um estímulo eficaz ocorre um aumento da permeabilidade do íon sódio (abrem-se os portões dos canais voltagem-dependente para sódio). O Na+ entra na célula a favor do gradiente de concentração levando consigo cargas positivas e gerando uma ligeira despolarização local. Essa despolarização, por sua vez, aumenta ainda mais a permeabilidade ao sódio fazendo com que grandes quantidades deste íon entre na célula. Esse fenômeno é chamado despolarização celular. Potencial de Overshoot: momento durante da despolarização em que o aumento do potencial faz com que ocorra a inativação do fluxo de sódio que cessa a sua entrada na célula. Repolarização Celular: saída de potássio Aproximadamente um milisegundo após a despolarização celular, ocorre um aumento da permeabilidade ao potássio que sai da célula a favor do gradiente de concentração (difusão simples) levando consigo cargas positivas e fazendo com que o potencial caia novamente a valores negativos. Esse processo é denominado repolarização celular. Após esse processo a célula volta a apresentar o seu valor normal de potencial de repouso (-85 mv); porém, as concentrações de Na+ e K+ estão invertidas. A bomba de sódio e potássio repõe as concentrações normais destes íons tornando a célula apta a responder a um novo potencial de ação. Profª Cíntia Schneider 6 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br Hiperpolarização celular: saída excessiva de potássio Em algumas células durante o processo de repolarização celular, a diferença de potencial pode baixar a valores maiores que -85 mv. Esse fenômeno dura apenas milésimos de segundo e imediatamente a célula volta a apresentar o seu potencial de repouso normal. A hiperpolarização ocorre devido a grande permeabilidade da célula aos íons potássio. Potencial de Ação: Os impulsos nervosos são transmitidos através de potencial de ação, que é uma rápida variação do potencial de repouso, ou seja, do potencial de negativo para o potencial de positivo com um rápido retorno para o potencial de repouso negativo, a membrana muda sua polaridade e depois volta ao normal. 5. IMPULSO NERVOSO Os neurônios são as células responsáveis pela recepção e transmissão dos estímulos do meio (interno e externo), possibilitando ao organismo a execução de respostas adequadas para a manutenção da homeostase. Para exercerem tais funções, contam com duas propriedades fundamentais: a irritabilidade (também denominada excitabilidade ou responsividade) e a condutibilidade. Irritabilidade é a capacidade que permite a uma célula responder a estímulos, sejam eles internos ou externos. Portanto, irritabilidade não é uma resposta, mas a propriedade que torna a célula apta a responder. Essa propriedade é inerente aos vários tipos celulares do organismo. No entanto, as respostas emitidas pelos tipos celulares distintos também diferem umas das outras. A resposta emitida pelos neurônios assemelha-se a uma corrente elétrica transmitida ao longo de um fio condutor: uma vez excitados pelos estímulos, os neurônios transmitem essa onda de excitação - chamada de impulso nervoso - por toda a sua extensão em grande velocidade e em um curto espaço de tempo. Esse fenômeno deve-se à propriedade de condutibilidade. Os impulsos nervosos ou potenciais de ação são causados pela despolarização da membrana além de um limiar (nível crítico de despolarização que deve ser alcançado para disparar o potencial de ação). Os potenciais de ação assemelham-se em tamanho e duração e não diminuem à medida em que são conduzidos ao longo do axônio, ou seja, são de tamanho e duração fixos. A aplicação de uma despolarização crescente a um neurônio não tem qualquer efeito até que se cruze o limiar e, então, surja o potencial de ação. Por esta razão, diz-se que os potenciais de ação obedecem à "lei do tudo ou nada". Para transferir informação de um ponto para outro no sistema nervoso, é necessário que o potencial de ação, uma vez gerado, seja conduzido ao longo do axônio. Um potencial de ação iniciado em uma extremidade de um axônio apenas se propaga em uma direção, não retornando pelo caminho já percorrido. Conseqüentemente, os potenciais de ação são unidirecionais - ao que chamamos condução ortodrômica. Uma vez que a membrana axonal é excitável ao longo de toda sua extensão, o potencial de ação se propagará sem decaimento. A velocidade com a qual o potencial de ação se propaga ao longo do axônio depende de quão longe a despolarização é projetada à frente do potencial de ação, o que, por sua vez, depende de certas características físicas do axônio: a velocidade de condução do potencial de ação aumenta com o diâmetro axonal. Axônios com menor diâmetro necessitam de uma maior despolarização para alcançar o limiar do potencial de ação. Nesses de axônios, presença de bainha de mielina acelera a velocidade da condução do impulso nervoso. Nas regiões dos nódulos de Ranvier, a onda de despolarização "salta" diretamente de um nódulo para outro, não acontecendo em toda a extensão da região mielinizada (a mielina é isolante), portanto, os potenciais de ação só ocorrem nos nódulos de Ranvier, onde há os canais voltagem-dependente para sódio. Fala-se em condução saltatória e com isso há um considerável aumento da velocidade do impulso nervoso. Profª Cíntia Schneider 7 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br 6. SINAPSES Os axônios, através de suas terminações, entram em contato com outros neurônios (SNC) e com células efetoras (glândulas, músculo...) (SNP) passando-lhes informações. Estes locais de contato entre estas células são chamados de sinapses. Quando há comunicação entre neurônios, chama-se sinapse interneuronal; quando o contato é entre neurônio e célula efetora, chama-se sinapse neuroefetuadora. As sinapses ainda podem ser divididas entre sinapses químicas e sinapses elétricas. As sinapses elétricas são menos freqüentes, e são exclusivamente interneuronais. Ocorre quando há contato entre as membranas dos dois neurônios, havendo assim uma comunicação direta de íons de uma célula a outra. Este tipo de sinapse não é polarizada, ou seja, ocorre nos dois sentidos. As sinapses químicas são as mais freqüentes, abrangendo todas as sinapses neuroefetuadoras e a maioria das interneuronais. Neste tipo de sinapses, a comunicação se faz através de uma substância química liberada, chamada neurotransmissor. Os principais neurotransmissores são a acetilcolina, o glutamato, o GABA, a dopamina, adrenalina e noradrenalina. Este tipo de sinapse é classificado como polarizada, ou seja, a comunicação se faz em apenas um sentido, da célula pré-sináptica (que possui o neurotransmissor) para a célula pós-sináptica (que possui os receptores para o neurotransmissor). Sinapses Químicas Interneuronais Na grande maioria destas sinapses, o elemento pré-sinaptico é uma terminação axônica, que se comunica com quaquer outra parte de outro neurônio, criando assim, sinapses axodendríticas (axônio – elemento pré-sináptico / dentrito – elemento pós-sináptico); sinapses axossomáticas (axônio – elemento pré-sináptico / pericário – elemtento pós-sináptico); ou sinapses axoaxônicas (axônio – elemento pré-sináptico / axônio – elemento pós-sináptico). Porém, pode ocorrer de o dendrito ou mesmo o pericário (corpo neuronal) ser o elemento pré-sináptico, criando assim, sinpases dendrodendríticas (entre os dendritos das duas células), sinapses dendrossomáticas (entre dendrito e pericário), sinapses somatossomáticas (entre dois pericários), sinapses somatodendríticas (entre pericário e dendrito) e assim por diante. Nas sinapses em que o axônio é o elemento pré-sináptico, o contato se faz de duas maneiras: ou através de uma única ponta dilatada, conhecida como botão sináptico, ou através de pequenas dilatações que ocorrem ao longo de sua arborização, criando varicosidades, chamados botões sinápticos de passagem. Sinapses Químicas Neuroefetuadoras Este contato se faz através entre os axônios dos neurônios periféricos e uma célula não neuronal. Se o contato for com uma célula muscular estriada esquelética, denomina-se junção neuroefetuadora somática, ou placa motora; neste tipo de sinapse o elemento pré-sináptico é uma terminação axônica de um neurônio motor, cujo corpo se localiza no corno anterior da medula espinhal. Se o elemento pós-sináptico for uma célula muscular lisa ou cardíaca, ou se for uma glândula, chama-se junção neuroefetuadora visceral; nestas o elemento pré-sináptico é a terminação nervosa de neurônios do sistema nervoso autônomo simpático e parassimpático, cujos corpos celulares se localizam nos gânglios autonômicos. Profª Cíntia Schneider 8 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br Uma característica das sinapses neuroefetuadoras é a presença de um espaço (virtual) entre o elemento pré e o elemento pós sináptico, chamada fenda sináptica, na qual o neurotransmissor é liberado após o impulso nervoso. Elementos de Uma Sinapse Uma sinapse compreende um elemento pré-sinaptico (que armazena e libera o neurotransmissor), uma fenda sináptica (onde é liberado o neurotransmissor) e um elemento pós-sináptico (que possui os receptores para o nerotransmissor). O elemento pré-sináptico apresenta uma membrana pré-sináptica, sob a qual, se organizam estruturas protéicas, formando uma malha, chamada densidade pré-sináptica. Nesta densidade estão agrupadas e organizadas as vesículas sinápticas, que armazenam os neurotransmissores. A fenda sináptica é o espaço virtual existente entre as duas células da sinapse, no qual será liberado o neurotransmissor. O elemento pós-sináptico é formado pela membrana pós-sináptica, na qual estão inseridos os receptores específicos para os neurotransmissores. A transmissão sináptica ocorre através da ligação do neurotransmissor com o seu receptor específico na membrana pós-sináptica. Transmissão Sináptica Quando o impulso nervoso atinge a membrana pré-sináptica, ocorre uma pequena alteração no potencial de membrana que abre canais voltagem-dependente para o íon cálcio, que entra na célula. O aumento na concentração deste íon dentro célula gera a fusão das vesículas sinápticas com a membrana pré-sináptica, liberando assim, o neurotransmissor armazenado para a fenda sináptica. Após a sua liberação na fenda sináptica, os neurotransmissores se ligam aos seus receptores específicos localizados na membrana pós-sináptica. Dependendo do neurotransmissor liberado e de sua ligação com o receptor específico pode ocorre uma inibição na célula póssináptica ou uma ativação. Por exemplo, a ligação do neurotransmissor GABA com o seu receptor específico, gera a abertura de canais para o íon Cl- na membrana pós-sináptica e a entrada deste íon na célula causa uma hiperpolarização, ou seja, a célula fica mais negativa que o normal; este é um estímulo inibitório. Se houver a ligação da acetilcolina com o seu receptor específico (nicotínico) ocorre a abertura de canais para o íon Na+, e a entrada deste na célula causa uma despolarização, ou seja, um estímulo excitatório. Para que não ocorra inibição ou excitação por tempo prolongado, é necessário que o neurotransmissor que continua na fenda sináptica após a transmissão do impulso nervoso seja rapidamente removido. Esta eliminação pode ocorrer de duas maneiras: por degradação ou por recaptação. Por degradação, ocorre a ação de enzimas, como por exemplo, a acetilcolinesterase, que degrada e elimina a acetilcolina da fenda sináptica. Na recaptação, ocorre uma recaptação do neurotransmissor pela membrana do elemento pré-sináptico, e uma vez dentro da terminação nervosa, o neurotransmissor pode ser reutilizado ou inativado. Esta recaptação pode ser bloqueada por drogas, como a cocaína, por exemplo, causando assim, uma oferta contínua de neurotransmissor na fenda Profª Cíntia Schneider 9 ESCOLA DE MASSOTERAPIA SOGAB Disciplina de Fisiologia Aplicada www.sogab.com.br sináptica, gerando excitação por tempo prolongado; este bloqueio é a causa dos muitos distúrbios psíquicos que afetam os usuários destas drogas. Profª Cíntia Schneider 10
Download