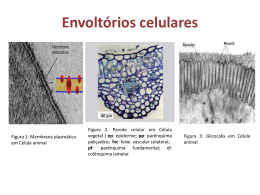
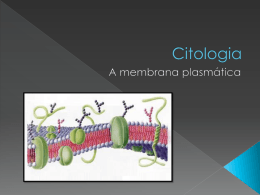
Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 22 ESTRUTURA E FUNÇÕES GERAIS DAS MEMBRANAS BIOLÓGICAS Os metazoários possuem um tegumento de revestimento que separa o meio ambiente externo do interno. O meio interno, por sua vez, é constituído de dois compartimentos: o extracelular e intracelular. Nos animais que possuem um sistema circulatório fechado, o fluido extracelular está distribuído em dois sub-compartimentos: o intersticial e o intravascular (plasma). O espaço intracelular está separado do extracelular através da membrana citoplasmática e ambos os espaços são preenchidos por fluidos cujas composições são bastante distintas. A membrana citoplasmática é uma barreira física, porém permite troca de solvente (água) e de partículas entre os compartimentos extra e intracelular garantindo que ass respectivas composições e osmolaridade sejam precisamente reguladas. Membrana citoplasmática A membrana citoplasmática é constituída de um mosaico de moléculas protéicas incrustadas em uma bicamada de fosfolipídios de consistência fluídica. Esse modelo da membrana citoplasmática é conhecido como mosaico fluido. Os fosfolipídios são moléculas que possuem uma cabeça polar (hidrofílica) e outra apolar (hidrofóbica). Na presença de água as moléculas de fosfolipídios se organizam espontaneamente de modo que os componentes hidrofóbicos voltam-se para dentro da bicamada (cauda) e os hidrofílicos para a água (cabeça). Na bicamada estão espalhados vários tipos de proteínas com as mais variadas propriedades funcionais. Todas as membranas citoplasmáticas compartilham entre si, propriedades fundamentais, mas de acordo com o tipo de célula, possuirá atividades biológicas. Entre essas propriedades podemos destacar: Regulação da composição dos fluidos intracelular e extracelular; Regulação do volume celular; Regulação do metabolismo intracelular determinando a concentração de co-fatores enzimáticos e de substratos; Regulação da atividade metabólica processada por enzimas presentes na membrana; decodificação de sinais químicos e físicos por meio de moléculas receptoras e reguladoras presentes na membrana; Geração e propagação de sinais elétricos; Endocitose e de exocitose. Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 23 PROCESSOS DE TRANSPORTE ATRAVÉS DA MEMBRANA Transporte Passivo ou por Difusão Difusão é o processo de movimento aleatório e espontâneo de partículas suspensas ou dissolvidas em solução cuja dispersão ocorre de uma região de maior concentração para outra de menor concentração, ou seja, a favor do gradiente de concentração (=energia potencial química). Quando o processo ocorre através de uma membrana, a taxa difusional (Js) depende do coeficiente de permeabilidade da membrana à partícula em questão e da área total pela qual ela atravessa. Este fenômeno obedece a lei de Fick sendo matematicamente descrito pela equação : dQs = Ds A dCs dt dx onde: dQs/ dt = taxa de difusão ( Js ) Ds = coeficiente de difusão da subst. s dCs/dx = gradiente de concentração de s entre dois pontos separados por uma distância x A= área total por onde ocorre o transporte Sob o ponto de vista da escala que estamos acostumados a dimensionar as distâncias e o tamanho dos objetos, isto é, em milímetros, centímetros, metros ou quilômetros, o processo de difusão seria um processo muito lento, mas tratando-se das dimensões ao nível celular e intracelular (micrômetros) o processo é bastante eficiente. As partículas podem ser classificadas em função da sua solubilidade e essa propriedade afeta o transporte das partículas através da membrana. Partículas lipossolúveis: atravessam diretamente a bicamada lipídica. Partículas não-lipossolúveis: necessitam de um corredor aquoso como os canais iônicos ou trasportadores especiais. Alguns solutos como as macromoléculas protéicas são completamente impermeáveis. Difusão simples As substâncias lipossolúveis (como o O2, CO2, o álcool, ácido graxo, hormônios esteróides, etc) difundem-se através da bicamada como se o fizessem no fluido circundante a favor do seu gradiente de concentração. O grau de solubilidade lipídica é determinado pelo quociente de partição óleo/água (K). Considerando-se uma membrana permeável a uma partícula s lipossolúvel, rearranjando a lei de Fick, temos: Js = Ds . K . A . dCs dx Ds = coeficiente de difusão K = coeficiente de partição óleo/água A = área da membrana por onde ocorre a difusão dCs = gradiente de concentração de s entre os dois lados dx = espessura da membrana Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 24 O gráfico mostra que a taxa de difusão para as partículas altamente lipossolúveis depende do gradiente de concentração existente através da membrana. Transporte de íons. Os íons só atravessam a membrana através de corredores aquosos formados por canais protéicos denominados canais iônicos. Há basicamente dois tipos de canais: Canais sem comporta: estão sempre abertos, mas são seletivos conforme o raio de hidratação do íon. Reconhecemos assim, canais de sódio, canais de potássio, canais de cálcio, etc. Canais sem comporta: permanentemente abertos Canais com comporta: assumem dois estados: abertos ou fechados. Suas comportas (gates) se abrem ou se fecham mediante agentes externos (neurotransmissores, mudança de potencial da membrana, fosforilação, etc.) Difusão facilitada Algumas moléculas hidrossolúveis como a glicose e aminoácidos não podem atravessar os canais iônicos e usam carreadores protéicos. A partícula a ser transportada se liga a uma proteína da membrana e muda a sua conformação espacial. Essa mudança causa a translocação da partícula de um lado para o outro da membrana. Se o processo for realizado a favor do seu gradiente eletroquímico denominamos essa forma de transporte como difusão facilitada. Como o Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 25 número total de carreadores é limitado, existe uma capacidade máxima de transferência de partículas. Transporte Ativo Até agora descrevemos os mecanismos em que as partículas são transportadas passivamente utilizando apenas a energia livre do próprio sistema. Muitas partículas precisam ser transportadas contra o seu potencial de difusão e, para isso, será necessário consumir energia externa ao sistema como a aquela originada do metabolismo celular. Por isso, esse tipo de transporte é chamado transporte ativo. Há dois tipos de transporte ativo. Transporte ativo primário. O transporte da partícula se realiza com a hidrólise de ATP. Um bom exemplo é enzima ATPase Na/K que hidroliza o ATP e transporta 3Na+ para fora da célula e 2K+ para dentro, ambos contra os respectivos gradientes eletroquímicos. A enzima é conhecida como bomba dependente de Na/K (ou, simplesmente bomba de Na). Transporte ativo secundário (ou acoplado). A partícula é transportada contra o seu gradiente. Nesse caso, ao nível da superfície epitelial do intestino, a glicose é transportada contra o seu gradiente para dentro da célula utilizando a energia livre do gradiente de concentração de outro soluto. O íon Na apresenta um gradiente de concentração de fora para dentro da célula, portanto, dispõe de energia potencial. Muitos solutos são co-transportados contra o seu gradiente usando um carreador. Se o movimento da partícula que pega “carona” e ocorre no mesmo sentido daquele que forneceu a energia é denominado de sinporte e se no sentido contrário, antiporte. Observe que tanto no transporte primário e secundário há consumo de energia; a diferença é está na fonte de energia. Note que na membrana basolateral a glicose é transportada por difusão facilitada, de dentro da célula par o sangue. A figura ilustra ainda o Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 26 transporte ativo primário de Na e K contra os respectivos gradientes de concentração, requerendo ATP. Endocitose e Exocitose O transporte de macromoléculas é realizado através de endocitose (entrada de material para dentro da célula) e exocitose (o contrário). Se houver absorção de liquido apenas, a endocitose é chamada de pinocitose e de sólidos, fagocitose. Potencial Eletroquímico de uma Partícula e Potencial de Difusão A figura abaixo ilustra o processo de difusão quando KCl é acrescentado à água pura no lado 1. Após um certo tempo, os lados 1 e 2 entram em equilíbrio, e passam a apresentar a mesma concentração de K e Cl, não havendo mais fluxo resultante de íons através da membrana. O potencial de difusão de uma partícula s (s) é uma grandeza que expressa o conteúdo de energia livre ou útil que potencialmente poderá ser utilizado para realizar trabalho, isto é, nesse caso, o movimento de partículas. Sob o ponto de vista termodinâmico, quando uma partícula se move de uma região de alto potencial eletroquímico para outra de baixo potencial, a difusão é espontânea e ocorre sem custo adicional de energia. Daí, o processo ser denominado de transporte passivo. Em nosso modelo, no estado de equilíbrio, os dois compartimentos ficaram com as concentrações de soluto e não havia mais movimento resultante de íons. Acontece que as células eucarióticas, ao contrário, apresentam imensas diferenças de concentração de solutos em relação ao meio extracelular e são ativamente mantidas neste estado. Essa diferença é, em parte, devido à impermeabilidade de algumas partículas como proteínas que ficam retidas no interior das células, mas e as várias outras partículas altamente permeáveis, como impedir que a diferença de Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 27 concentração seja dissipada ao longo do tempo? E como explicar a presença de potencial elétrico através da membrana? Existindo uma diferença de potencial eletroquímico entre dois pontos A e B, através da membrana, a partícula s se moverá espontaneamente, a favor desse gradiente. Como o próprio nome diz, o potencial eletroquímico de s leva em conta dois componentes: uma grandeza de natureza química (diferença de concentração) e outra, elétrica (diferença de potencial elétrico): A diferença de potencial elotroquímico de s entre dois pontos A e B ( sA - sB) separados por uma membrana pode ser descrito como : S 1 - s S 2 = RT ln [ s 1 ] + Zs . F. ( 2 - 1 ) ou : [ s 2] = RT ln [ s A ] + Zs . F. ( 2 - 1 ) [ sB] Desta equação podemos deduzir que: Se s > 0 : haverá movimento resultante de s no sentido de 1 2 (não está em equilíbrio); s = 0 : não haverá movimento resultante de s através dos dois compartimentos e o sistema se encontra em estado de equilíbrio dinâmico. s < 0 : haverá movimento resultante de s de 2 1(não está em equilíbrio) Se a partículas s não for um íon, isto é, não está carregada eletricamente, Zs=0 então, s = RT ln [ s 1 ] o potencial de difusão só depende do gradiente químico [ s2] Efeito Donnan No início do século passado, Donnan, um físico-químico observou que quando em dois volumes iguais de água pura são acrescentados KCl em um dos compartimentos, ambos os íons se dissociam e tendem a se difundir passivamente para compartimento adjacente, atingindo em seguida um, estado de equilíbrio ou de eletroneutralidade entre os dois pontos. Continuando com o nosso experimento, se proteínas impermeáveis forem acrescentadas em um dos compartimentos, um novo estado de equilíbrio será atingido, porém, a dinâmica do movimento de íons será diferente. Suponha que sejam adicionadas simultaneamente proteinato de K no lado 1 e de KCl no lado2 de modo que [K]1=[K]2. Ambos os compostos iônicos se dissociarão produzindo íons livres. Apesar do gradiente de concentração do P favorecer o seu transporte, como a membrana lhe é impermeável ela fica retida no lado 1. Em relação ao K não se espera que haja fluxo resultante já que sua concentração é a mesma em ambos os lados. Com relação ao Cl, há uma diferença de concentração e este tenderá a fluir passivamente do lado 2 para o 1, a favor do gradiente eletroquímico. Repare que o movimento de Cl do lado 2 para o 1, tornará o lado 2 positivamente carregado em relação o outro lado, porém sem atingir igualdade de concentração, ou seja [Cl]2[Cl]1, pois o acúmulo de cargas negativas no lado 1 repele a passagem deste ânion. Ao mesmo tempo, este desbalanço de cargas elétricas através da membrana favorecerá um fluxo resultante de K, um cátion, para o lado 1. A distribuição de P continua não afetada: lembre-se ele é impermeável. 28 Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida - Assim, mesmo que haja diferença de concentração de K, Cl e P através da membrana, o sistema se manterá em equilíbrio, sem fluxos resultantes de K e de Cl. Isto mostra que uma condição de equilíbrio não é necessariamente uma condição de igualdade de concentração em ambos os lados da membrana, mas o fato do fluxo resultante das partículas permeáveis ser igual a zero (s = 0). Como resultado desta distribuição, haverá uma diferença de potencial elétrico através da membrana: no lado1 acumulará um excesso de cargas negativas e no lado 2, cargas positivas. A esta distribuição de solutos permeáveis e não permeáveis eletricamente carregadas através de uma membrana semipermeável damos o nome de Equilíbrio de Donnan. Como podemos calcular o valor do potencial de membrana estabelecido nestas condições? Já tínhamos visto que: s = RT ln [ s 1 ] + Zs . F. ( 2 - 1 ) [ s2] Se o sistema está em equilíbrio, então s = 0. Agora podemos calcular a diferença de potencial que existe através da membrana, mesmo havendo diferença de concentração: 2 - 1 = RT ln [ s 2] Zs.F [ s 1] Como ambos os íons estão em equilíbrio, sob esta diferença de potencial elétrico: 2 - 1 = RT ln [K]2 = RT ln [Cl]2 (+1)F [K]1 (-1)F [Cl]1 Equação 1 Ou seja, no estado de equilíbrio de Donnan, obtemos a distribuição recíproca de cátions e de ânions. RT ln [K]2 = RT ln [Cl]2 ou (+1)F [K]1 (-1)F [Cl]1 [K]2 = [Cl]2 [K]1 [Cl]1 Se levarmos em conta a presença de P- - + [P ] + [Cl ]1 = [K ]1 Conhecendo agora algumas propriedades das membranas, vamos analisar o problema, levando-se em consideração uma célula de verdade. Potencial de repouso de uma célula Através da membrana citoplasmática de todas as células do organismo metazoário, existe uma diferença de potencial elétrico (Em). Tal fato pode ser demonstrado, implantando-se um eletrodo de registro no interior da célula e o de referência, do lado de fora. O voltímetro acusará uma diferença de potencial elétrico de -65mV. Isso significa que através da membrana existe uma distribuição desigual de cargas elétricas sendo que a face interna se encontra negativa em relação à externa. Dizemos então que a membrana apresenta uma polaridade e comporta-se 29 Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida como uma bateria (capacitor) que acumula cargas elétricas nas duas faces. Essa diferença de potencial é chamada Potencial de Repouso nos neurônios (e em outras células excitáveis) que não estão em atividade. As membranas dos neurônios e das fibras musculares são capazes de causar fluxos de íons (cargas positivas ou cargas negativas) através da membrana, o que resulta na alteração do Em. A propriedade de uma célula mudar transitoriamente o Em de repouso é chamada excitabilidade e indica a capacidade de gerar bioeletricidade. Além de gerar eletricidade, as células excitáveis são capazes de propagar o impulso elétrico de um ponto a outro da célula, isto é, são dotadas de condutibilidade. Antes de compreendermos como os impulsos elétricos são gerados, precisamos conhecer como o potencial de repouso é gerado e mantido. Potencial de equilíbrio Veja a figura da página seguinte e considere um sistema (A) onde a membrana é permeável apenas ao K+ e impermeável ao Cl-. Além disso, tanto o lado I como II possuem a mesma concentração de KCl. Como não há diferença de concentração, não haverá fluxo resultante de K ( K=0). Se não há fluxo de íons, não haverá nenhuma diferença de potencial elétrico através da membrana (é o que o voltímetro está indicando). Em B, o lado I recebeu uma concentração 10 vezes maior de KCl, ou seja, agora, os K+ poderão se difundir passivamente para o lado II, a favor do seu gradiente de concentração. O movimento de cátions para o lado II tornará esse lado mais positivo em relação ao lado I. Passado algum tempo (C) ainda que o gradiente químico de K continue a impelir os íons K para o lado II, o sistema entra em equilíbrio, isto é, K=0. Por que? A medida que os K se difundem para o lado II, a diferença de potencial elétrico através da membrana vai progressivamente aumentado, até que o gradiente elétrico começa a se opor ao gradiente químico e o sistema entra em equilíbrio. O valor do potencial elétrico através da membrana que se opõe ao gradiente químico do K é chamado então de potencial de equilíbrio do K e pode ser determinado pela equação de Nernst. Em = RT ln [K]in = - 58mV [K]ext a 20oC Por analogia, podemos calcular o potencial de equilíbrio para todos os íons permeáveis de uma célula de verdade, já que as concentrações químicas são conhecidas. Ion K+ Na+ Ca++ Cl Extracelular 5 mM 150 mM 2 mM 150 mM Intracelular 100 mM 15 mM 0,0002 mM 13 mM Razão Extra/Intra 1:20 10:1 .000:1 11,5:1 Eion (37o C) - 80mV + 62mV + 246mV - 65mV Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 30 Pelo que vimos até agora, é fácil imaginar que nem o Na e nem o Ca devem ser os íons responsáveis pelo Em do estado de repouso, já que o potencial de repouso observado é de – 65mV. Então, o Em deve ser causado pelo potencial de equilíbrio do K + já que é o íon mais permeável e não apresenta fluxo resultante. De fato, alterações de concentração externa de K+ causam mudanças significativas no Em. O gráfico ao lado mostra que o aumento de [K]ext reduz progressivamente o Em a zero, ou seja, a membrana vai sofrendo despolarização. No sistema nervoso, qualquer excesso de K é rapidamente seqüestrado pelos astrócitos que garantem uma [K]ext baixa. A equação do potencial de equilíbrio de um íon (equação de Nernst) não leva em consideração a permeabilidade relativa dos íons através da membrana. Então, Goldmann e seus colegas propuseram que apesar da pNa ser muito baixa, haveria uma diferença de permeabilidade entre os íons onde pK > pNa na ordem de 40x. Isso significa que o EM real não poderia ser exatamente o potencial de equilíbrio do K (E K = - 80mV), mas um pouco menor, devido ao escoamento de correntes de Na. Em = RT ln pK[K]ext pK[K]int + + pNa[Na]ext = - 65mV pNa[Na]int E o gradiente de concentração dos íons? Quem origina e o mantém durante o potencial de repouso? Como vimos, o potencial de difusão do K depende da sua elevada permeabilidade à membrana e a existência de um gradiente de concentração (potencial químico). Para manter esse gradiente através de uma membrana permanentemente é necessário consumo de energia externa ao sistema. A ATPase dependente de Na/K resolve o problema transportando 3Na e 2K contra os respectivos gradientes e, assim, garantindo a sua manutenção. Quando a ATPase é bloqueada pela oubaina, os gradientes químicos e elétricos existentes através da membrana celular se dissipam. Apesar de eletrogênica, a contribuição da bomba é insignificante para modificar o Em. Excitabilidade celular: bioleletrogênese e propagação do impulso nervoso Nas células excitáveis, determinados estímulos causam mudanças transitórias no Em a ponto de inverter completamente a polaridade elétrica. Tal evento elétrico é chamado de potencial de ação (PA). O PA começa com uma rápida despolarização seguida de repolarizaçâo. A despolarização rápida é devida à entrada de cargas positivas no interior do neurônio o que vai reduzindo a diferença de potencial, até que ocorra uma completa inversão de polaridade. Neste ponto, a face interna da membrana fica carregada positivamente e a face interna, negativamente. Na repolarizaçâo, o Em volta aos valores do repouso graças à saída de cátions. A saída de cátions é tanta que a membrana chega ficar momentaneamente hiperpolarizada, isto é, torna-se mais negativamente carregada do que no estado de repouso. Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 31 Um neurônio pode ser estimulado com correntes elétricas aplicadas por um eletrodo de estimulação. A figura ao lado mostra o neurônio exibindo vários eventos de PA enquanto está sendo estimulado eletricamente. As alterações do Em durante o PA são causadas pelo fluxo de íons (cargas elétricas) através da membrana. Comportamento dos potenciais de membrana das células excitáveis A estimulação elétrica das células excitáveis causam outros dois tipos de respostas, alem do PA: 1. distúrbios elétricos de baixa voltagem, conhecidos como potenciais eletrotônicos que são reações puramente passivas da membrana à aplicação do estímulo, sem ocorrer alterações de condutância iônica. Propaga-se rápida e passivamente a distâncias curtas. 2. distúrbios conhecidos como resposta ou excitação local que vai além do potencial eletrotônico puro e são causadas por um aumento na condutância da membrana ao Na+ mas a despolarização se mantêm localizada, com distância perde energia potencial. Estes fenômenos ficam restritos, por exemplo, nos dendritos e no soma. O PA é um tipo de distúrbio elétrico de alta voltagem, muito especial e ocorre somente quando a membrana do neurônio for despolarizada até um valor critico. A figura ao lado ilustra um neurônio multipolar recebendo conexões nervosas aferentes. As regiões do corpo celular e dos dendritos são os sítios de recepção dos estímulos nervosos e respondem aos estímulos com potencias elétricos de baixa voltagem (a, b) denominados potenciais pós-sinápticos. Essas respostas elétricas são algebricamente computadas e propagadas eletrotonicamente até a zona de gatilho do PA no cone de implantação do axônio. Somente a partir do cone de implantação, o PA pode ser gerado e propagado ao longo do axônio. 32 Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida MECANISMOS GERADORES E CONDUTORES DO POTENCIAL DE AÇÃO No cone e ao longo de todo o axônio há canais com comporta para o Na e K. Durante o repouso esses canais estão fechados e a membrana está polarizada (cargas negativas na face interna e cargas positivas na face externa). Se o segmento inicial do axônio for despolarizado até o potencial limiar, os + canais de Na com comporta, sensíveis a voltagem, se abrem e o íon difunde-se passivamente para dentro do axônio. A entrada de cargas positivas despolariza a membrana ainda mais, abrindo mais canais de Na+ voltagem dependentes num ciclo de retro-alimentação positiva a ponto de inverter completamente a polaridade da membrana (mais cargas positivas na face interna). A despolarização continua até que o Em alcance valores próximos ao potencial de equilíbrio do Na+ (ENa). O novo Em se opõe ao fluxo passivo de Na e a entrada do cátion diminui. Ao mesmo tempo, os canais de Na voltagem dependentes fecham-se tão rapidamente quanto se abriram, tornando-se temporariamente inativos. Até aqui ficou claro como se dá a fase de despolarização do PA. E como ocorre a repolarizaçâo? Para que o Em volte ao estado de repouso, uma solução seria a de reduzir as cargas positivas da face interna da membrana por meio da saída de K. De fato é isso que acontece. Diferente dos canais de Na voltagem dependentes que se abrem e se fecham rapidamente os canais de K voltagem dependentes se abrem e se fecham lentamente. O K pode agora se difundir passivamente para fora da célula, a favor do seu gradiente elétrico e químico causando a repolarização sem qualquer gasto de energia. Como os canais de K+ fecharem-se lentamente, a face interna da membrana chega a acumular mais cargas negativas do que na condição de repouso, tornando-se momentaneamente hiperpolararizada. Os estudos com os canais unitários de Na e de K deixam claro que durante a despolarização ocorre entrada de corrente (íons) e durante a repolarização, saída ou seja, movimentos de íons através da membrana. É importante salientar que a quantidade de Na e de K que atravessa a membrana durante o PA não causa alteração significativa na concentração dos respectivos íons nos dois lados da membrana. A prova de que a entrada de íon Na é o responsável pelo Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 33 desencadeamento do PA é reforçado pelos seguintes dados: se houver um grande aumento na concentração de Na extracelular, o neurônio se tornará facilmente excitável e se houver redução, muito inexcitável. O modo de ação dos anestésicos locais é um bom exemplo: a lidocaina inibe especificamente os canais de Na voltagem dependentes, impedindo a geração de PA nas células sensoriais causando a analgesia. A TTX é um potente veneno que age do mesmo modo. Na figura da página anterior, o segmento do axônio possui um eletrodo de estimulação (s) e dois eletrodos de registros R1 (mais próximo) e R2 (mais distante). Logo abaixo são mostrados os diferentes pulsos de correntes e os respectivos registros do Em. Repare que o PA só foi gerado apenas quando o estimulo indicado pela seta foi aplicado. Além disso, o PA foi registrado no ponto R1 e R2, ou seja, o evento elétrico se propagou de um lugar para outro e sem alterar a sua amplitude. A intensidade de estimulo capaz de gerar um PA é denominada estímulo limiar e o valor do potencial de membrana é denominado de potencial limiar. Os estímulos que só provocaram alterações de baixa voltagem (potenciais subliminares) são denominados estímulos subliminares. As intensidades acima do estímulo limiar são conhecidas como supralimiares e tem como resposta, aumento na freqüência dos PA. Propriedades do PA Se o axônio fosse bem curto, a condução da informação poderia ser feita por condução eletrotônica, mas como as mensagens neuronais percorrem distâncias longas, é essencial um mecanismo que garanta a total fidelidade da informação, sendo propagada a longas distâncias. Por exemplo, a condução de eletricidade através de cabos elétricos é bem rápida, mas a intensidade do sinal sofre um decaimento exponencial com a distância. Como então contornar a perda de sinal com a distância? A solução foi produzir um evento elétrico auto-regenerativo e auto-propagável como o PA cuja duração e amplitude se propagam imutavelmente ao longo do axônio. Lei do Tudo-ou-Nada: Se um estímulo limiar for aplicado a uma célula excitável, a célula responderá com um PA e nada impedirá que o fenômeno seja adulterado. É por isso que o PA é conhecido como um fenômeno tudo-ou-nada. Isso ocorre por causa das propriedades intrínsecas dos canais voltagem-dependente que se abrem e se fecham sempre de maneira constante. O seu fechamento é “automático”, isto é, não depende de um segundo estimulo para mudar a sua conformação. Assim, quando ocorre uma despolarização da membrana até o limiar, todos os canais se abrem e depois de um tempo, fecham-se automaticamente. Decodificação de intensidade do estímulo. Se o PA é um fenômeno cuja amplitude não varia, como os neurônios decodificam variações de intensidade dos estímulos? A intensidade é codificada em função da freqüência dos PA gerados no axônio (numero de PA / unidade de tempo). A decodificação da variação de intensidade é realizada do estímulo limiar até um máximo. Período refratário: Se um segundo estímulo limiar for aplicado, enquanto o primeiro PA já está em curso, não será possível desencadear outro PA, pois os canais de Na não estarão completamente inativados, isto é, fechados. Este período corresponde ao período refratário absoluto (PRA). A aplicação de um estímulo limiar em uma fase posterior, no curso final da repolarização será possível desencadear um PA, ainda que de menor amplitude. A este período denominamos período refratário Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 34 relativo (PRR). Esta propriedade é também decorrente das propriedades dos canais voltagem dependente e indica que um novo PA só pode ser gerado quando a membrana estiver completamente repolarizada aos níveis de repouso. Curva Intensidade-Duração. Um estímulo forte despolariza rapidamente a membrana até o limiar e precisa de pouco tempo para causar um PA. Já um estímulo mais fraco requer mais tempo para que a mesma quantidade crítica de corrente flua através da membrana para despolarizar o limiar. O gráfico da curva intensidade-duração descreve esta relação. Isto quer dizer que além da intensidade, a duração do estímulo aplicado também é igualmente importante para a manifestação do PA. A intensidade mínima de estímulo capaz de causar um PA é denominada reobase e o tempo necessário para sua aplicação, tempo de utilização. A duração que equivale ao dobro da reobase é denominada de cronaxia. Acomodação: se uma célula excitável for despolarizada muito lentamente até o limiar, não manifestará PA. Isto acontece porque os canais de Na abertos pela despolarização se tornam inativos (se fecham) antes de atingir o potencial limiar, limitando o número de canais críticos necessários para deflagrar o PA. Além disso, como os canais de K voltagem dependentes também se abrem, a membrana se torna refratária. Mecanismo de condução do impulso nervoso ( visite site recomendado nos links ) A figura ilustra a condução do PA em um axônio sem mielina. Repare que o PA está se propagando da esquerda pela direita: assim, primeiro vemos o influxo de Na (despolarização) seguido do efluxo tardio de K (repolarização) Assim que o PA é gerado no cone de implantação (de um neurônio multipolar) ele se propaga ao longo do axônio até o terminal axônico, como se fosse um rastilho de pólvora em combustão. Como o PA se propaga da zona de gatilho até o seu terminal sem depender de um novo estimulo e mantendo a sua amplitude e duração? A membrana citoplasmática é um mal condutor de cargas elétricas, ao contrário dos fluidos extra e intracelulares. Quando uma região da membrana sofre uma despolarização (zona ativa), as correntes iônicas se propagam longitudinal e transversalmente fechando o circuito. Estas correntes eletrotônicas despolarizam a membrana adjacente que se encontra em repouso. Assim, quando ocorre entrada de Na durante o PA, a região adjacente, “empurra” passivamente corrente negativa para a superfície da membrana, despolarizando-a passivamente. Esta resposta local despolariza a membrana até o seu limiar, ou seja, ativam os 35 Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida canais iônicos vizinhos de Na e de K voltagem dependentes, gerando novo PA e assim sucessivamente. Desta maneira o próprio PA serve de estímulo do PA sucessivo, garantindo a sua propagação autoregenerativa, sem qualquer custo adicional de energia metabólica. O PA é conduzido sempre do segmento inicial para o terminal axônico (anterogradamente) e isto não é devido a uma propriedade inerente à membrana ou ao mecanismo de propagação. O sentido da condução não é revertido porque a membrana do axônio onde o PA acabou de ocorrer se torna refratária, i e, os canais de Na se encontram ainda inativos e os de K ainda estão abertos. Portanto, existe um efluxo (saída) de K que hiperpolariza a membrana (tornando-a distante do potencial limiar), barrando qualquer despolarização eletrotônica anterógrada. Ou seja, a existência do período refratário seve como mecanismo de segurança para que o impulso nervoso seja gerado sempre num único sentido (do corpo celular em direção ao terminal sináptico). Velocidade de Condução do PA Tipos de fibras nervosas de um nervo de mamífero Tipo de Fibra Função Diâmetro (m) Velocidade (m/s) Duração do PA (ms) PRA (ms) 0,4 –0,5 0,4-1,0 A B C Propriocepção; Motor somático 10-20 70-120 Tato, pressão 5-12 30-70 Motor 3-6 15-30 Dor, temperatura, tato 2-5 12-30 Neurônio pré-ganglionar autonômico <3 3-15 1,2 1,2 Raízes dorsais Simpático Dor, respostas reflexas Neurônios pós-ganglionares 0,4 – 1,2 0,3 – 1,3 0,5 –2,0 0,7 – 2,3 2 2 2 2 A tabela acima mostra que a velocidade de propagação dos impulsos depende de dois fatores: do calibre e se o axônio é mielinizado ou não. O gráfico informações. ao lado resume estas 1) Diâmetro do axônio. Quanto maior o diâmetro, menor será a resistência ao fluxo de corrente no axoplasma e como conseqüência, maior a veleocidade. 2) Mielinização da fibra. Uma outra maneira de aumentar a velocidade da condução é o de isolar o axônio eletricamente a intervalos fixos. Os gliócitos encapam os axônios com mielina com exceção dos nodos de Ranvier. O PA iniciado no cone é conduzido de nodo a nodo, onde os canais de Na e de K voltagem dependentes estão presentes. O PA no nodo seguinte é causado pela propagação eletrotônica de corrente gerado pelo PA anterior. Assim, ao invés do PA se propagar continuamente como acontece na fibra sem mielina, ocorre aos saltos, gastando menos tempo até os terminais nervosos. O aumento na velocidade de condução nervosa propicia rapidez Curso de Fisiologia 2012 NeuroBiologia Departamento de Fisiologia, IB Unesp-Botucatu Profa. Silvia M. Nishida 36 na transmissão de informações sensoriais, na emissão de comandos motores para os órgãos efetuadores, bem como no processamento de sinais neurais pelo SNC. Potencial de ação de um nervo composto Um nervo periférico é constituído de várias fibras nervosas, tanto motoras como sensitivas (com a exceção de alguns nervos cranianos que possuem ou nervos sensitivos ou motores) e pode conter fibras mielinizadas como não mielinizadas. Tomando-se um nervo composto, é possível registrar o PA composto do nervo, estimulando-o experimentalmente. Na figura abaixo, os registros dos PA do nervo a partir de 3 eletrodos colocados a várias distâncias do eletrodo de estimulação. Note que o PA composto parece ser constituído de vários picos. O que se vê na verdade é a resultante de vários PA unitários das fibras nervosas que compõe o nervo; daí ser denominado de potencial de ação composto. O gráfico ilustra a reconstrução do PA composto mostrando o tamanho relativo dos PA unitários conforme o calibre das fibras e a duração do impulso. Repare que a amplitude e a duração dos PA unitários são diferentes para cada tipo de fibra.
Baixar