Capítulo 2 A Teoria Sintética Introdução Você já deve ter notado que para Darwin estar correto é preciso uma condição básica: tem de haver diferenças entre os organismos de uma determinada espécie, de tal forma, que o ambiente possa selecionar as variações mais bem adaptadas. Essa condição gerava um grande embaraço para Darwin, porque ele não tinha nenhum dado concreto a respeito do que gerava essa variedade de indivíduos. Havia, na época, um total desconhecimento sobre a hereditariedade ( Mendel só publicaria seu trabalho em 1865 que, mesmo assim, ficou esquecido até 1900, porque, aparentemente, ninguém entendeu o que ele quis dizer). Foi apenas no século XX que a genética surgiu como ciência e alguns pesquisadores uniram os conhecimentos dela com a teoria da evolução. A união desses dois ramos da ciência gerou o que chamamos de Teoria Sintética da E v o l u ç ã o ou Neodarwinismo. Um grande feito da teoria sintética foi explicar de onde vêm essas variações entre os indivíduos de uma mesma espécie. As Fontes de Variabilidade Como dito, apenas em 1900 os trabalhos de Mendel foram definitivamente compreendidos. A descoberta de que as características hereditárias estavam “escritas” nos fatores (atuais genes) representaram um grande salto. Logo depois dessa descoberta, vários cientistas verificaram que os genes podem sofrer alterações ao acaso (m u t a ç õ e s gênicas). Por incrível que pareça, a descoberta das mutações fez com que vários cientistas acreditassem que Darwin estava errado. Segundo eles, a seleção natural não seria necessária, visto que novas espécies poderiam surgir a partir de uma simples mutação. Hoje, sabemos que não é assim. Espécies novas não surgem de forma abrupta por causa de uma única mutação. Esse é um processo lento, em que muitas alterações no DNA precisam ocorrer e a seleção natural atuar. Geralmente as alterações nos genes são desvantajosas. Imagine, por exemplo, que cada palavra desse livro seja um gene de um determinado organismo. Agora substitua uma palavra por outra, ao acaso. É claro que é muito difícil que essa substituição faça com que o texto se torne melhor. De qualquer modo, temos vários exemplos de mutações que conferiram vantagens a seus possuidores (leia o quadro abaixo). Contextualizando… Como a maioria das mutações são prejudiciais, praticamente todos os seres vivos possuem um sistema de reparo que as corrige. Além disso, os sistemas enzimáticos responsáveis pela duplicação do DNA são “muito cuidadosos” no seu trabalho, de forma que a quantidade de mutações é pequena. Mas pesquisadores do Laboratório de Mutagênese do Instituto Jacques Monod, da Universidade de Paris, fizeram uma interessante descoberta sobre o tema. Segundo eles, quando bactérias são submetidas a “situações de stresse intenso (como falta de alimento), O SOS (sistema de reparo do DNA) funciona de modo inverso, estimulando a mutação. O SOS aumenta também a variabilidade genética, provocando rearranjos cromossômicos (…). Finalmente, o SOS aumenta a taxa de recombinação, inclusive entre sequências de DNA que não são rigorosamente idênticas. Ao fazer isso, ele facilita a integração eventual de fragmentos de DNA provenientes de uma outra espécie.” Numa situação como a descrita acima, de falta de alimento, alterações no material genético podem ser muito boas, porque, a partir delas, pode surgir, por exemplo, uma nova enzima que permita às bactérias fazer uso de uma nova fonte de alimento que até então elas não conseguiam digerir. Nesse tempo falava-se de uma taxa de mutação mínima, visto que muitas causam problemas, mas esses pesquisadores propõem o termo taxa de mutação ótima, visto que, sem elas, a evolução não ocorre. Prof. Dorival Filho 11 Além das mutações gênicas existem também as a b e r r a ç õ e s ou m u t a ç õ e s cromossômicas. Elas podem ser classificadas em numéricas e estruturais. As numéricas envolvem a alteração na quantidade de cromossomos e as estruturais envolvem a perda ou incorporação de pedaços de cromossomos. As alterações cromossômicas são menos frequentes do que as gênicas, mas, ainda assim, desempenham um importante papel na produção de variabilidade genética. um com uma combinação única de alelos. Mas ainda há uma outra maneira de produzir novas combinações alélicas. Crossing-over ou permutação O crossing-over é uma troca de pedaços entre cromátides de cromossomos homólogos. Com essa troca é possível formar cromossomos, carregando novas combinações de alelos (veja a figura abaixo). Recombinação Gênica Um ser vivo não é formado por poucos genes. Nós, por exemplo, possuímos milhares deles (estima-se que cerca de 40.000). Ora, os genes não trabalham isolados uns dos outros. É a interação de todos eles que fazem um Sem o crossing-over só poderia ser formado dois tipos de gametas: AB e organismo. Portanto, além da criação de ab. Com o crossing-over (representado pelas setas no desenho do meio) um novo gene por mutação, um indivíduo surgem mais duas possibilidades: Ab e aB. Portanto, podemos concluir que as será diferente do outro por causa da mutações produzem novos genes que se combinação de genes que possui. recombinam por meio da segregação A reprodução sexuada por meiose (que independente e do crossing-over gerando produz os gametas) desenvolveu duas uma grande variedade de organismos dentro maneiras de recombinar os genes, formando de uma determinada espécie. Essa variedade sempre organismos diferentes (salvo os é a matéria-prima que o ambiente utiliza para gêmeos univitelinos). selecionar os variantes mais bem adaptados. Segregação independente A maneira como ocorre a segregação dos cromossomos homólogos na anáfase I da meiose é aleatória (veja a figura ao lado). Perceba en- Na situação I o fuso mitótico puxa os cromossomos com os alelos ABC para a esquerda e os alelos abc para a direita formando dois gametas diferentes: ABC e abc. Na situação II os tão que apenas a cromossomos estão dispostos de outra maneira. Assim, pode se formar outros dois gametas: segregação indepen- AbC e aBc. Na situação III os gametas seriam: abC e Abc. dente já produz uma Fazendo alguns cálculos, podemos concluir que o número de combinações possíveis entre os infinidade de indi- cromossomos é de 2 n, onde n é o número de pares de cromossomos. Portanto, um ser víduos diferentes, cada humano pode formar 223 gametas diferentes. Atualize-se Desde a clonagem da ovelha Dolly muitos alunos me perguntam se essa técnica não poderia salvar espécies ameaçadas de extinção. Afinal, à primeira vista, bastaria clonar os últimos sobreviventes. O problema é que a clonagem produz indivíduos geneticamente idênticos e, como você já viu, para que uma espécie evolua é preciso que haja diversidade, variabilidade genética. Sem ela não ocorre a seleção natural, pelo simples fato de que não há o que selecionar. Portanto, mesmo que conseguíssemos fazer 1.000 ou 100.000 clones de um indivíduo representante de uma espécie ameaçada de extinção, essa espécie já teria chegado a seu fim evolutivo. O Teorema de Hardy-Weimberg Prof. Dorival Filho 12 conhecido como terorema de Hardy-Weinberg ou princípio de Hardy-Weinberg. Esse teorema demonstra matematicamente que a proposta do aluno não está correta, desde que se estabeleça algumas condições. Na verdade, o princípio de HardyWeinberg demonstra justamente o contrário. Ele prevê que, se nenhum fator evolutivo estiver atuando na população, a freqüência de alelos permanecerá constante com o passar das gerações (Veja o quadro abaixo). Todas as vezes em que estou explicando a primeira lei de Mendel, algum aluno sempre me faz a mesma pergunta: Se na F2 a proporção de indivíduos com fenótipo dominante é de 75% e a de recessivos é 25%, com o passar das gerações não vai aumentar a frequência do alelo dominante até sumir o recessivo? Foi tentando responder a essa mesma pergunta que o matemático inglês Godfrey H. Hardy (1877-1947) e o médico alemão Wiilhem Weinberg lançaram um teorema que ficaria Fatores Evolutivos Como já dito, o teorema de Hardy-Weinberg diz que, na ausência de fatores evolutivos, a frequência dos alelos em uma determinada população permancerá constante com o passar das gerações. Sem Fatores Evolutivos Freqüências 1 0.8 Freqüência do alelo A Freqüência do alelo a 0.6 0.4 0.2 0 1 2 3 4 5 6 7 8 9 10 Gerações Veja abaixo quais são os fatores evolutivos que alteraram o equilíbrio de Hardy-Weinberg modificando as freqüências gênicas. 1. Mutações As mutações podem alterar a frequência dos alelos. Se a taxa de mutação do alelo A, por exemplo, for diferente do seu alelo a, com o passar das gerações as freqüências se modificarão e o alelo A se tornará cada vez menos frequente. 2. Seleção natural Imagine que um determinado alelo dificulte a sobrevivência de seu possuidor, reduzindo suas chances de chegar a idade adulta e ter filhos. A tendência é que, com o tempo, esse alelo se torne cada vez mais raro, estando a população fora do equilíbrio. Seleção Natural Favorecendo o Alelo a Freqüências 1 0.8 Freqüência do alelo A Freqüência do alelo a 0.6 0.4 0.2 0 1 2 3 4 5 6 7 8 9 10 Gerações Prof. Dorival Filho 13 3. Migração Não pode haver nem imigração nem emigração de indivíduos da população. Isso gera um fluxo gênico que pode ou retirar ou introduzir alelos, alterando suas freqüências. 4. Cruzamentos preferenciais Não pode haver preferência por um determinado fenótipo para cruzamento. Todos os cruzamentos devem ser casuais. Chamamos de Panmixia a qualidade de uma população em que todos os cruzamentos são casuais. 5. Populações pequenas As populações devem ser muito grandes para que não ocorra a chamada deriva genética. A deriva genética dá-se quando, pelo acaso, a freqüência de um alelo aumenta ou diminui drasticamente, podendo até desaparecer. Em uma população pequena isso pode acontecer com muita facilidade. Por exemplo, não é tão difícil você jogar uma moeda para cima e o resultado ser cara cinco vezes seguidas. Agora jogar a moeda 100 vezes e as 100 resultarem em cara é muito mais difícil. Da mesma forma, uma população pequena pode, por coincidência, na passagem de uma geração para outra, formar vários indivíduos homozigotos para o mesmo alelo, por exemplo, AA. Nesse caso, a freqüência do alelo A sobe rapidamente e a do a cai. Numa população grande (assim como no caso das moedas) dificilmente aconteceria isso. Deriva Genética Freqüências 1 0.8 Freqüência do alelo A Freqüência do alelo a 0.6 0.4 0.2 0 1 2 3 4 5 6 7 8 9 10 11 Gerações 6. Efeito do fundador Imagine que uma ilha foi colonizada por poucos representantes de uma determinada espécie de pássaros. Por serem poucos, eles possuem pouca variabilidade genética. Nessa população a deriva genética seria levada ao extremo. Como vimos, vários fatores podem alterar as freqüências gênicas de uma geração para outra. Se você parar para pensar, vai perceber que, na realidade, é muito difícil encontrar uma população em equilíbrio de Hardy-Weinberg, porque sempre, pelo menos, alguns dos fatores descritos acima estarão atuando. Sua importância é chamar a atenção para o fato de que, se as freqüências gênicas não estão constantes, é sinal de que está presente algum fator evolutivo. Portanto, as populações em que esses fatores evolutivos estão ausentes estão em equilíbrio de Hardy-Weinberg. Para que você entenda a equação do equilíbrio de Hardy-Weinberg, imagine uma população em que a frequência do alelo A seja 60% (0,6) e a do seu alelo a, 40% (0,4). As porcentagens significam o seguinte: cada organismo carrega dois alelos. Portanto, em uma população de 50 pessoas, haverá 100 alelos, sendo 60 A e 40 a. Para que nasça uma pessoa com o genótipo AA é necessário que os dois gametas que a formaram transportem o alelo A. Prof. Dorival Filho Como mostrado acima, cada gameta carrega apenas um alelo, visto que é haplóide. Portanto, para sabermos qual é a possibilidade de nascer uma pessoa AA nessa população basta multiplicarmos a frequência dos alelos A. Observe abaixo: Probabilidade de se formar um 0,6 espermatozóide A Probabilidade de se formar um 0,6 óvulo A Probabilidade de nascer um 0,6 x 0,6 = 0,36 indivíduo AA ou 36% 14 0,4 excludentes, como no caso acima, em que, por exemplo, o espermatozóide ou é A ou a, não podendo se dar os dois ao mesmo tempo, devese somar as probalidades. Veja abaixo: 0,4 0,24 + 0,24 = 0,48 ou 48% 0,4 x 0,4 = 0,16 ou 16% Portanto a chance de se formar um heterozigoto nessa população é de 48%. Observe que, ao invés de somar 0,24 + 0,24 você pode simplesmente fazer 2 x 0,24, que o resultado será o mesmo. Agora você já está em condições de entender a expressão do teorema de HardyWeinberg. Considere que: p = freqüência do alelo dominante f(A). q = freqüência do alelo recessivo f(a). Conclusão: Veja agora como calcular probabilidade de nascer um indivíduo aa. Probabilidade de se formar um espermatozóide a Probabilidade de se formar um óvulo a Probabilidade de nascer um indivíduo aa a Agora, muita atenção para a formação do heterozigoto Aa. Ele pode se formar de duas maneiras. Condição 1: Probabilidade de se formar 0,6 um espermatozóide A Probabilidade de se formar 0,4 um óvulo a Probabilidade de nascer 0,6 x 0,4 = 0,24 um indivíduo Aa ou 24% OU Condição 2 Probabilidade de se formar um espermatozóide a Probabilidade de se formar um óvulo A Probabilidade de nascer um indivíduo Aa 0,4 0,6 0,4 x 0,6 = 0,24 ou 24% Freqüência de AA = p x p = p2 Freqüência de Aa = p x q + p x q = 2pq Freqüência de aa = q x q = q2 Como, evidentemente, em uma população qualquer, a freqüência de todos os genótipos possíveis (AA + Aa + aa) é 100% ou 1 a expressão do teorema de Hardy-Weinberg fica: Nesse caso, caímos na regra do “ou” do estudo de probabilidade de genética. Essa regra diz que, quando os eventos são p2 + 2pq + q2 = 1 Exercícios Resolvidos sobre o teorema de Hardy-Weinberg. (PUC-SP) Uma população que está em equilíbrio de Hardy-Weinberg é constituída por 2 mil indivíduos. Sabe-se que 320 deles têm uma certa anomalia, determinada por um gene autossômico recessivo. Entre os indivíduos normais dessa população, qual o número esperado de portadores desse alelo recessivo? Resposta: Se a doença é autossômica recessiva, significa que os 320 indivíduos que a possuem são aa. Numa população de 2 mil indivíduos 320 representam 16% (0,16). Ora, a freqüência do genótipo aa é representada na equação de Hardy-Weinberg por q2. Logo para descobrirmos qual a freqüência do alelo a basta fazermos: q2(faa) = 0.16 q(fa) = 0,4 Portanto, a freqüência do alelo a é de 0,4 ou 40%. Como a soma do a com o A tem que dar 100%, fica fácil descobrirmos a freqüência do alelo A. Basta subtrairmos 1 – 0,4 que acharemos 0,6 (ou 60%). A questão quer saber dentre os indivíduos normais quem é portador do alelo a. Para ser normal e portador do gene recessivo, é preciso ser heterozigoto (Aa). Os heterozigotos são representados por 2pq. Logo basta substituirmos o p e o q na fórmula: 2 . 0,6 . 0,4 = 0,48 (ou 48%). Ou seja, 48% dos dois mil indivíduos da população são heterozigotos. 48% de 2 mil representam 960 indivíduos. Portanto essa é a resposta do problema. 960 indivíduos são normais, mas apresentam o alelo recessivo que determina a anomalia. Prof. Dorival Filho 15 Acaso x seleção natural Para Darwin a seleção natural é criativa. É seu contínuo trabalho de selecionar os variantes mais bem adaptados que vai criando espécies novas. Contudo, muitos cientistas não acreditam que a seleção natural tenha tanto poder assim. Para eles o acaso, por exemplo, pode ser muito mais importante. Para chegar a essa conclusão, eles observaram a imensa variedade genética entre os indivíduos de uma mesma espécie. Ora, se a seleção natural fosse tão presente, não poderia haver tantas formas alélicas para uma mesma característica, afinal, o ambiente selecionaria apenas as melhores. Tentando resolver essa questão um pesquisador japonês chamado Kimura lançou o que ficou conhecido como teoria neutralista. Segundo ela, se encontrarmos dois, três, quatro ou mais alelos ao mesmo tempo numa população, com freqüências constantes, é porque todos eles são equivalentes. N ã o haverá um melhor do que o outro, a ponto de o ambiente poder selecioná-los. Logo, para a seleção natural, eles serão neutros. Durante um bom tempo, houve uma discussão sobre quem estaria com a razão: os que defendiam a seleção natural, ou os defensores do neutralismo. Aparentemente, hoje, há um consenso de que o que vale é o meio termo. Realmente muitas mutações são neutras, não conferem vantagem nem desvantagem. Mas, mutações que provoquem alteração em proteínas essenciais para o bom funcionamento do organismo sofrem a influência da seleção natural. Proteínas, como as histonas, que se ligam ao DNA, são extremamente estáveis do ponto de vista evolutivo. Se compararmos a seqüência de aminoácidos que formam as histonas em peixes, com a de humanos notaremos pouquíssima diferença, apesar de peixes e humanos estarem afastados evolutivamente entre si, por milhares de anos. Já a comparação envolvendo proteínas que não comprometem tão diretamente a saúde mostra que as diferenças são muitas. Portanto, para essas proteínas, várias modificações podem ocorrer ao acaso, porque elas são neutras, não causam nem bem nem mal. Já as histonas, que são importantíssimas, não têm essa mesma “liberdade” para mudar, porque pequenas alterações podem ser eliminadas pela seleção natural. Contextualizando… Você já leu anteriormente uma matéria sobre animais que vivem em cavernas em que lhe foi mostrado como Lamarck e Darwin explicariam a evolução deles. Observe na matéria abaixo explicações sobre a evolução desses organismos troglóbios (que vivem em cavernas) baseadas no neutralismo. Adaptação da matéria “Guano de Morcegos, fonte de vida nas cavernas” publicada pela revista “Ciência Hoje”, no 146. (…) Saber como certas características dos troglóbios evoluíram torna o estudo da vida em cavernas mais interessante. Em geral, esses animais apresentam várias especializações relacionadas ao ambiente cavernícola. (…) Para descrever a primeira hipótese (de como essa evolução ocorre) é fundamental esclarecer antes o que significa o termo “mutação neutra”. (…) Uma mutação neutra é aquela que não influencia nenhum aspecto importante da sobrevivência e/ou da reprodução de um organismo – a “neutralidade”, porém depende do ambiente em que esse organismo vive. Essa hipótese assume que a regressão de certas estruturas, observada em alguns organismos que vivem em cavernas é resultado do acúmulo de mutações neutras (que aparecem casualmente em indivíduos de uma população) durante várias gerações. Sem função nesses ambientes, tais estruturas (olhos, por exemplo) seriam gradativamente reduzidas com o passar das gerações, pois mutações casuais nesse sentido não afetariam a sobrevivência ou reprodução do indivíduo. (…) A hipótese de acúmulo de mutações neutras, portanto, propõe que a regressão de estruturas em muitos troglóbios pode ter ocorrido por causa da ausência de pressões seletivas (como a luz) que eliminariam indivíduos mutantes no meio externo (…). Prof. Dorival Filho 16 O Que a Evolução NÃO é Como todos os conceitos importantes, a evolução gera controvérsia; ela tem sido usada como fundamento intelectual para pontos de vista fiiosóficos, éticos ou sociais e religiosos. Em seu sentido mais amplo, a evolução é meramente mudança e, desse modo, é uma idéia de ampla penetração; notese que galáxias, linguagens e sistemas políticos também evoluem. Evolução biológica é a mudança que ocorre nos organismos em um período de vida maior que o de um único indivíduo. As mudanças que ocorrem no indivíduo durante sua vida não são consideradas evolutivas; Organismos individuais não evoluem. As mudanças que são consideradas evolutivas são aquelas herdáveis via material genético, de uma geração para a outra. Os diversos mecanismos de evolução incluem a seleção natural, que é responsável pelas adaptações dos organismos a diferentes ambientes. Nem a seleção natural, nem qualquer dos outros mecanismos são medidas prévias para alcançar algum fim; a seleção natural, por exemplo, é somente a sobrevivência ou reprodução superior de algumas variantes genéticas, em comparação com outras, sob quaisquer condições ambientais que estejam prevalecendo NO MOMENTO. Desse modo, a seleção natural não pode equipar uma espécie para encarar novas situações futuras nem tem propósito ou direção - nem mesmo o da sobrevivência da espécie. A seleção natural é tão mecânica quanto a gravidade, ela não é moral nem imoral. Portanto, para as pessoas que acreditam que tudo existe para um determinado propósito, a evolução torna-se de difícil compreensão. Além disso, essas pessoas têm dado significados à evolução que nem Darwin nem os biólogos modernos aceitam. Alguns igualaram a evolução a "progresso" das formas de vida "inferiores", em direção às "superiores". A própria palavra progresso implica direção e, como já vimos, a evolução não tem proposito ou direção. Há ainda a suposição de que o que é “natural" é "bom". Dessa forma a lei da seleção natural e o "progresso", este erroneamente atribuído à evolução, foram pregados pelos Darwinistas sociais como um princípio ético que Darwin negava. Esses darwinistas sociais pretenderam, com isso, justificar a competição econômica desenfreada, o capitalismo e o imperialismo, fundados na suposição de que o que é natural é bom. Texto adaptado do /ivro “Biologia Evolutiva" do autor Douglas Futuyma Seleção Natural em Crise? Nesse capítulo você estudou que existem outros fatores que participam do processo evolutivo além da seleção natural proposta por Darwin. Atualmente, existem cientistas que defendem que a seleção natural desempenha um papel fundamental na evolução, tendo definido inclusive o comportamento humano. Na ala oposta, estão aqueles que acreditam que os fatores evolutivos até agora descrito são insuficientes para explicar alguns aspectos morfológicos e fisiológicos. Existem alguns processos fisiológicos nos seres vivos que precisam de um grande número de componentes para que funcione. Detalhe, se for retirado um deles, nada mais funciona. Existem sistemas que só funcionam se todas as peças estiverem presentes no Fonte: A Caixa Preta de Darwin. Prof. Dorival Filho 17 lugar e no momento certo, eles são chamados de sistemas de complexidade irredutível. O pesquisador Michael Behe, defensor dessa idéia, dá alguns interessantes exemplos de sistemas de complexidade irredutível. Um deles é a ratoeira (veja o desenho na página anterior). Observe que basta uma das peças que compõem a ratoeira estar ausente para que ela não funcione mais. Mas, o que isso tem a ver com a evolução? Nos seres vivos existem muitos sistemas de complexidade irredutível e os evolucionistas não têm conseguido explicar satisfatoriamente como eles evoluíram. O problema é que Darwin defendeu que um sistema simples pode evoluir para um mais complexo por meio de uma série de etapas intermediárias, mas, desde que cada etapa seja útil ao seu possuidor. Acontece que em sistemas de complexidade irredutível, as etapas intermediárias não trazem nenhuma vantagem seletiva, porque, lembre-se, ou todas as peças estão juntas ao mesmo tempo ou nada funciona. Um bom exemplo biológico disso é o flagelo bacteriano. Observe no desenho quantas proteínas são necessárias para seu funcionamento. Se uma célula tivesse apenas uma delas, ou mesmo algumas, aparentemente, isso não traria nenhuma vantagem para a célula. Se não traz vantagem como é que a seleção natural favoreceu seu desenvolvimento de forma a posteriormente ter surgido o flagelo Fonte: Biochesmistry inteiro? Essa é uma das questões mais sérias que os evolucionistas enfrentam hoje. • • • • • • Supra-Sumo A única fonte de novos genes é a mutação. A segregação independente e o crossing-over “apenas” criam novas combinações de genes. As mutações são aleatórias. Portanto, não surgem mutações para atender a uma determinada necessidade. As mutações, a segregação independente e o crossing-over, criam uma grande quantidade de organismos geneticamente diferentes dentro de uma espécie. O ambiente, então, tem a matériaprima necessária para fazer a seleção natural. Caso nenhum fator evolutivo (mutações, seleção natural, migração…) esteja atuando, as freqüências de genótipos e alelos permanecerão constantes, com o passar das gerações (Teorema de Hardy-Weinberg). O teorema de Hardy-Weinberg pode ser expressado por: p2 + 2pq +q2. Algumas mutações são neutras, ou seja, não produzem um efeito suficientemente grande para serem “percebidas” pelo ambiente e sofrerem o efeito da seleção natural. Prof. Dorival Filho 18
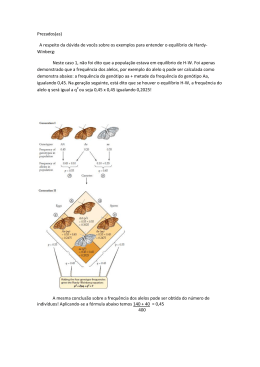
Baixar