☰
Explorar
Assinar em
Inscrever-se
Envio
×
Baixar
Sem categoria
ferreira julio
isabela trab de geo mapas
Clique aqui para obter o arquivo
HEC-HMS Exercícios
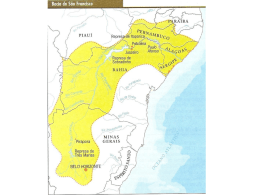
transposição são francisco
hidrosfera - Colégio Energia Barreiros
Bacia do Rio Doce
Biól. Marília Lourenço dos Santos
Salmo 08
Clique aqui para obter o arquivo
O PROBLEMA ENERGÉTICO NO BRASIL