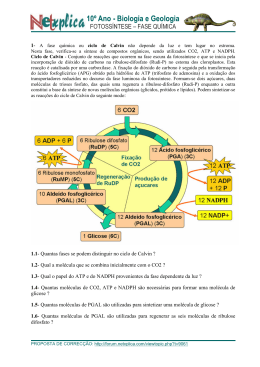
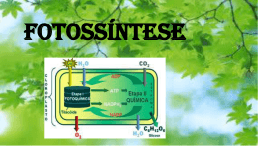
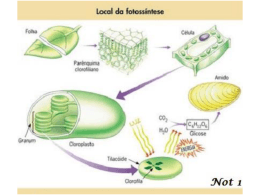
FOTOSSÍNTESE: Sistema fotossintético e fisiologia comparada das plantas de ciclo “C” 3 “C” 4 e CAM. INTRODUÇÃO: A fotossíntese representa o caminho pelo qual praticamente toda a energia penetra na biosfera. Através da fotossíntese as plantas clorofiladas convertem a energia luminosa, proveniente do sol, em energia química. A fotossíntese é uma complexa cadeia de reações, realizada através da energia luminosa por pigmentos específicos (como a clorofila), que resulta na síntese de compostos orgânicos (açúcares) a partir de gás carbônico e água. Todos os demais compostos, essenciais para a estrutura da planta, são produzidos a partir desses compostos orgânicos resultantes da fotossíntese. e d o m r a C o d a i r a M . D . C a t s o C A folha é o órgão sede da fotossíntese, devido sua riqueza em clorofila, sua posição na planta, geralmente exposta à luz, sua forma laminar que permite aproveitamento com eficiência da energia luminosa. O caule de certas plantas e até a raiz de certas espécies de orquídeas, podem realizar fotossíntese, embora a função fotossintética seja preferencialmente desempenhada pelas folhas nos vegetais. Para que as folhas possam sintetizar os alimentos é indispensável, entre outras coisas que suas células verdes disponham de CO2 . O CO2 do ar, penetra na folha através dos estômatos e difunde-se nos espaços intercelulares, dissolvese na água e atravessa o plasmalema das células onde através do citoplasma chegam aos cloroplastos. Nos cloroplastos o CO2 é usado na síntese de açúcares, sendo sua concentração no cloroplasto sempre inferior a do ar. Isto permite um fluxo constante de CO2 do ar para os cloroplastos. A luz pode ser entendida como uma onda de partículas energéticas chamadas fótons. A energia desses fótons é inversamente proporcional ao comprimento de onda dessas prtículas. Assim, a luz azul possui fótons mais energéticos que a luz vermelha, que apresenta comprimento de onda maior. A principal etapa na conversão da energia solar em energia química é a absorção da luz. Um pigmento é qualquer substância que absorve a luz. O padrão de absorção de um pigmento é conhecido como espectro de absorção. Quando um pigmento absorve luz, os elétrons são elevados a um nível mais alto, podendo ocorrer: conversão de energia em forma de calor; transferência da energia sob a forma de energia luminosa (fluorescência ou fosforescência); e captura da energia numa ligação química, como ocorre na fotossíntese. Os pigmentos que participam da fotossíntese incluem as clorofilas, os carotenóides e as ficobilinas. Existem vários tipos diferentes de clorofila que diferem uns dos outros apenas em pequenos detalhes de suas estruturas moleculares. A clorofila a ocorre em todos os eucariontes fotossintéticos e nas algas azuis procarióticas, sendo considerada como essencial para a fotossíntese dos organismos pertencentes a este grupo. A clorofila b é um pigmento acessório e, a exemplo dos outros pigmentos acessórios, amplia o espectro de absorção de luz na fotossíntese. Como a clorofila b absorve luz de comprimento de onda diferentes da clorofila a, aumenta a faixa de luz que pode ser utilizada para a fotossíntese. Os carotenóides são pigmentos lipossolúveis vermelhos, alaranjados ou amarelos, encontrados em todos os cloroplastos e, também, em associação com a clorofila a nas algas azuis procariotas. Os carotenóides que não contêm oxigênio são denominados carotenos, ao passo que os que possuem este elemento são chamados de carotenóis. Na folha verde, a cor dos carotenóides é mascarada pelas clorofilas, que se encontram presentes em quantidades muito mais abundante. Em alguns tecidos, como os de um tomate maduro ou pétalas de uma flor amarela, predominam os carotenóides. Como não são hidrossolúveis, os carotenóides não são encontrados livres no citoplasma, são ligados a proteínas dentro dos plastídios. As ficobilinas são pigmentos acessórios hidrossolúveis, encontrados nas algas azuis, nos cloroplastos das algas vermelhas e em alguns outros grupos de algas eucariotas. e d o m r a C o d a i r a M . D . C a t s o C No cloroplasto, a clorofila e outras moléculas estão empilhadas em estruturas conhecidas como unidades fotossintéticas. Cada unidade contém 250 a 400 moléculas de pigmento, que podem absorver luz e transferi-la para uma molécula reativa. A energia absorvida pela molécula reativa é suficiente para expulsar totalmente um elétron da molécula. A molécula reativa, é portanto, oxidada e perde um elétron, constituindo uma situação por vezes designada como “ter uma lacuna eletrônica”. Para preencher estas lacunas existem dois sistemas fotoquímicos que atuam conjuntamente nas folhas. Assim, os elétrons perdidos por uma molécula reativa são substituídos por outros que provém de uma outra molécula também reativa ou da molécula de água após a fotólise. SISTEMAS DE PIGMENTOS Nas folhas das plantas, existem dois tipos diferentes de sistemas de pigmentos, conhecidos como Sistema de pigmento I e Sistema de pigmento II. O Sistema de Pigmento I contém uma maior proporção de clorofila a em relação à clorofila b que o Sistema de Pigmento II. Em ambos os sistemas encontram-se diferentes tipos de carotenóides. No Sistema de Pigmento I, a molécula reativa é uma forma de clorofila a conhecida como P700, pelo fato de um dos picos de seu espectro de absorção estar situado a 700 nanômetros. Quando P700 é oxidado, descora-se, constituindo o modo pelo qual é detectado. O Sistema de Pigmento II contém também uma molécula de clorofila especializada, capaz de transferir um elétron para uma forma de clorofila a. Seu ponto máximo de reação se situa a 680 nanômetros. Por conseguinte, cada sistema de pigmento faz parte de um sistema fotoquímico diferente. É provável que os sistemas fotoquímicos tenham evoluído separadamente e que o sistema fotoquímico I surgiu primeiro. O sistema fotoquímico I, provavelmente, é capaz de operar isoladamente, porém, de modo geral, atua em conjunto com o sistema fotoquímico II. a t s o C Para que a fotossíntese ocorra, é necessário a presença de um aceptor de hidrogênio, o qual será reduzido. Este aceptor de hidrogênio receberá os prótons hidrogênio provenientes na molécula de água hidrolisada durante as reações no claro. Também provém da água o oxigênio que é librado durante o processo fotossintético e os elétrons que irão substituir os elétrons da clorofila b perdidos quando esta foi excitada pela absorção de luz. . D . C Basicamente a fotossíntese compõem-se de duas etapas: as reações no claro (fosforilação acíclica e cíclica) e reações no escuro (ciclo de Calvin). e d o m r a C o d a i r a M REAÇÕES NO CLARO O processo inicia-se com a absorção de luz pela clorofila a (fotossistema I), que transfere dois elétrons energizados para um aceptor primário, a ferredoxina. Esta transfere esses elétrons ao NADP, que recebe também os + prótons (H ) provenientes da fotólise da água, reduzindo-o a NADPH2. Este será utilizado posteriormente na redução de CO2 a carboidrato, na fase escura da fotossíntese. A absorção de luz pela clorofila b (fotossistema II) resulta na transferência de dois elétrons energizados para a plastoquinona. Estes dois elétrons são substituídos, na clorofila b, pelos dois elétrons provenientes da molécula da água. Da plastoquinona os elétrons são transferidos, através de uma série de aceptores de níveis energéticos cada vez mais baixos, até chegarem na clorofila a, que pode recebê-los por já ter perdido, anteriormente, dois elétrons. A energia liberada nessa transferência de elétrons é usada para sintetizar ATP a partir de ADP e fosfato. Assim a energia luminosa foi convertida em ligações de alta energia do ATP e também em NADPH2, que serão utilizados na fixação do gás carbônico. Os elétrons retirado das clorofilas não retornam aos sistemas fotossintetizantes (FS I e FS II), percorrendo um caminho não cíclico, razão pela qual falamos em fotofosforilação acíclica. Nesse transporte de elétrons associado à liberação de oxigênio formam-se duas moléculas de ATP. Quatro elétrons são liberados na fotólise da água e transportados na cadeia acíclica, e para cada dois elétrons transferidos forma-se uma molécula de ATP. Fosforilação cíclica Constitui uma formação adicional de ATP pelas plantas, para complementar o número de ATPs necessários para a fixação de uma molécula de CO2. A fotofosforilação acíclica fornece NADPH2 , ATP e O2 na proporção de 2:2:1, porém são necessários 3 ATP para fixar 1 CO2 . Assim, o ATP adicional será fornecido pela fosforilação cíclica. . D . C a t s o C Na fosforilação cíclica clorofila a (FS I), absorvendo energia luminosa, tem elétrons excitados que se deslocam para níveis energéticos mais altos, sendo transferido da clorofila para um aceptor de elétrons, a ferredoxina. Desta, ele se desloca para os citocromos, voltando a níveis energéticos mais baixos. Nesta volta, ocorre liberação de energia que é incorporada nas ligações de ATP. Como o elétron sai da clorofila a, voltando a ela, num trajeto cíclico, e como houve produção de ATP por fosforilação, o processo é chamado fosforilação cíclica. Neste processo, que exige a presença de luz e de cloroplastos, forma-se unicamente ATP, não conhecendo-se o número exato de moléculas de ATP que se formam. e d o m r a C o d a i r a M Acredita-se que os mecanismos fotossintéticos mais primitivos atuavam realmente deste modo. Estes mecanismos fotossintéticos não liberavam oxigênio e não se sabe se reduziam o carbono a açúcar. REAÇÕES NO ESCURO Embora chamada desta forma as reações no escuro, acontecem realmente no claro, simultaneamente com as reações ditas no claro. Têm, entretanto, esta denominação por acontecer no estroma dos cloroplastos e não depender de uma fonte luminosa. A energia utilizada nesta fase provém dos ATPs e NADH2 formados durante as reações no claro. As reações no escuro (ciclo de Calvin ou ciclos da ribulose difosfato) consta de uma série de reações nas quais ocorre a carboxilação que consiste na fixação do gás carbônico pela ribulose difosfato (RuDP), formando um composto instável, de 6 carbonos; este sofre hidrólise e se degrada, originando duas moléculas de fosfoglicerato (PGA). Cada molécula de PGA é reduzida a gliceraldeído-fosfato (PGAL), uma triose fosforilada. Para cada molécula de CO2 fixada são produzidas duas trioses. A cada três voltas no ciclo seis trioses são produzidas, destas, 5 participam de uma série de interconversões de açúcares, até regenerar a ribulose difosfato. Apenas 1 sai do ciclo e é utilizada na formação da glicose. Para a produção de uma molécula de glicose são necessárias, portanto, seis voltas no ciclo. Quando se completam a seis voltas no ciclo, então, o primeiro PGAL que sai do ciclo reúne-se ao segundo PGAL para formar hexose-fosfato e, daí, glicose. É importante lembrar que os açúcares não são os únicos compostos orgânicos produzidos pela fotossíntese. O fosfoglicerato (PGA) e outros compostos intermediários do ciclo de Calvin podem ser utilizados na síntese de ácidos orgânicos, aminoácidos e outros produtos. a t s o C As plantas que exibem o PGA como composto resultante do ciclo de Calvin são chamadas de plantas C3 , já que o PGA é um composto de três carbonos. Em outras palavras, plantas C3 são as que fixam o CO2 pelo ciclo de Calvin. . D . C Alguns autores já constataram,entretanto, que algumas espécies de plantas adaptadas a regiões áridas como o milho, a cana-de-açúcar e gramíneas tropicais, são capazes de fixar CO2 em compostos de 4 átomos de carbono ao invés de 3, como no ciclo de Calvin. Nestas plantas aparece ácido málico, ou aspártico, como primeiro produto fotossintético e são chamadas de plantas C4. As plantas C4 fixam o CO2 pelo ácido fosfoenolpiruvato (PEP). e d o m r a C o d a i r a M Há um aspecto anatômico associado a essa via C4 de fixação do CO2; as folhas dessas plantas apresentam uma bainha de células clorofiladas envolvendo o feixe vascular. O CO2 da atmosfera é assimilado nas células do mesófilo que não exibem o ciclo de Calvin, reage com o ácido fosfoenolpiruvato , formando ácido oxalacético, na presença da enzima PEP carboxilase. Posteriormente, esse ácido oxalacético é reduzido a malato ou aspartato, dependendo do tipo de planta, pelo NADPH2 proveniente da fase clara. O malato ou aspartato é levado para as células da bainha vascular onde é descarboxilado, sendo o CO2 utilizado no ciclo de Calvin, enquanto o ácido pirúvico proveniente da descarboxilação, retorna as células do mesófilo, onde reage com o ATP para formar mais moléculas de PEP. Por conseguinte, é apenas nas etapas iniciais da fixação do carbono que a fotossíntese C4 difere da fotossíntese C3. A eficiência das plantas C4, deve-se ao farto dessas plantas absorverem eficientemente o CO2 mesmo quando em baixas concentrações. Isto deve-se ao fato da PEP ter maior afinidade pelo CO2 que a RuDP do ciclo de Calvin. A temperatura ótima para a fotossíntese C4 é muito mais alta que aquela para a fotossíntese C3 e, as plantas C4 florescem mesmo em temperaturas que seriam letais para muitas plantas C3. As plantas C4 estão especialmente bem adaptadas a grandes extremos de intensidade de luz, temperatura e seca. Uma terceira via de fixação do CO2, é a fotossíntese MAC (metabolismo ácido das crassuláceas), que evoluiu independentemente em muitas plantas suculentas, incluindo os cactos. A fotossíntese MAC, representa um sistema especializado de fixação do CO2, destinado a manter um balanço positivo de carbono nos tecidos, ao mesmo tempo que desenvolve-se um eficiente mecanismo de absorção de água. Este mecanismo fotossintético, utiliza também moléculas de 4 carbonos, na fixação do CO2. Nestas plantas, à noite os estômatos se abrem e permite a entrada do CO2 que é assimilado através da reação catalisada pela enzima PEPcarboxilase. O ácido oxalacético produzido é transformado em os ácidos málico e isocítrico, os quais acumulam-se nas folhas, durante a noite. Durante o dia por ação da luz, o malato é descarboxilado sendo que o piruvato formado reage com o ATP e regenera o PEP, enquanto o CO2 liberado no processo é captado pela RuDP do ciclo de Calvin. Este processo é evidentemente vantajoso em condições de alta intensidade luminosa e de falta de água, nas quais vivem a maior parte das plantas suculentas. Estas plantas dependem, em grande parte, do acúmulo durante a noite de carbono para a fotossíntese, pelo fato de seus estômatos estarem fechados durante o dia a fim de retardar a perda d’água. e d o m r a C o d a i r a M . D . C a t s o C FATORES QUE LIMITAM A FOTOSSÍNTESE Vários fatores internos e externos podem influir na taxa de fotossíntese. Como fatores internos podemos citar os pigmentos fotossintetizantes e as enzimas e coenzimas essenciais ao processo. Por exemplo, se a planta for clorótica por deficiência de alguns elementos essenciais como o nitrogênio e o magnésio, não haverá clorofila suficiente para executar a fotossíntese. Os mais importantes fatores que influenciam a fotossíntese são: concentração de CO2, água, luz e temperatura. CO2 Na natureza, o gás carbônico é freqüentemente o fator limitante da fotossíntese. Se a taxa de CO2 no ar estiver baixa, o gás não conseguirá se difundir para os cloroplastos com a mesma velocidade com que ele é consumido. Por outro lado se a concentração de CO2 na atmosfera exceder 1%, passa a ser inibitória, devido a redução do pH pelo ácido carbônico que se forma. A redução do pH promove o fechamento dos estômatos, o que impede a difusão do gás carbônico para os cloroplastos. Neste caso a planta estará carente de CO2, havendo abundância deste gás. LUZ A luz é um fator que limita a fotossíntese à noite e nos momentos que varia sua intensidade. Com grandes intensidades de luz, a taxa fotossintética reduz-se por possíveis danos nos pigmentos fotossintéticos. Em algumas espécies os cloroplástos mudam de posição de acordo com a intensidade luminosa para evitar danos. Durante o dia, quando a fotossíntese está sendo efetuada, haverá uma certa intensidade luminosa, na qual o CO2 consumido na fotossíntese será equivalente ao CO2 liberado na respiração o mesmo acontecendo com o O2 liberado na fotossíntese e consumido na respiração. A essa intensidade luminosa chamamos ponto de compensação fótico. Quando a planta recebe uma intensidade luminosa superior ao ponto de compensação, a taxa fotossintética é maior que a respiratória. Assim, a planta aumenta a produção de matéria orgânica e cresce. O ponto de compensação varia com a espécie: nas plantas de sol, o ponto de compensação é maior que nas plantas de sombra. e d o m r a C o d a i r a M ÁGUA . D . C a t s o C Embora apenas 1% da água absorvida pela planta seja usada na fotossíntese, há evidências que o déficit de água limita a taxa fotossintética. Isso ocorre porque, com a menor hidratação do protoplasma e em particular dos cloroplastos, cai a atividade enzimática, reduzindo os processos metabólicos. Além disso a falta de água promove o fechamento dos estômatos, o que reduz a difusão de CO2. TEMPERATURA Na maioria das espécies tropicais a fotossíntese não ocorre em temperaturas próximas a 5º C. Entretanto, muitas espécies realizam fotossíntese com temperaturas de 0º C ou - 25º C, como alguns líquens e mesmo com - 35º C como algumas coníferas. Para maioria das plantas, entretanto, a temperatura ótima está entre 30º C e 40º C. Temperaturas altas, entre 50 e 60º C, reduzem a taxa fotossintética, provavelmente por determinarem destruição das enzimas e danos no aparelho fotossintetizante. Além disso, a temperatura pode influenciar na abertura e fechamento dos estômatos. QUESTÕES PARA FIXAÇÃO DA APRENDIZAGEM 1. O que entende por fotossíntese ? 2. Quais os pigmentos que participam da fotossíntese ? 3. O que ocorre durante na etapa fotoquímica da fotossíntese ? 4. O que ocorre durante a etapa química da fotossintese ? 5. Quais os fatores limitantes da fotossínte ? Explique cada um deles. BIBLIOGRAFIA INDICADA AO ALUNO a t s o C 1. COUTINHO, L. H. Botânica. v. 2, 7ª ed., São Paulo, Cultrix, 1976. 307 p. il. 2. FERRI, M. G. Fisiologia vegetal 1. 2ª ed. São Paulo: EPU, 1985. 3. MODESTO, Z. M. M. & SIQUEIRA, N. J. B. Botânica. São Paulo, Editora Pedagógica e Universitária - EPU, 1981. . D . C 4. RAVEN, P. H. , EVERT, R. F. & CURTIS, H. Biologia vegetal, 2ª ed., Rio e d o m r a C o d a i r a M de Janeiro, Guanabara Dois, 1978. 724 p. il.
Baixar