☰
Explorar
Assinar em
Inscrever-se
Envio
×
Baixar
Sem categoria
FELIPE DE ALMEIDA DIAS
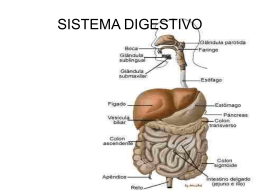
Sistema Digestório
Órgãos associados ao Trato Digestivo
Tipos de Associações entre os Animais
Tema: Tecido epitelial de secreção (glândulas).
Sistema dgestivo
Sistema digestório1
Aves
Clique aqui para visualizar.
ANFÍBIOS