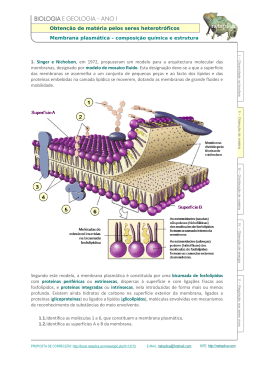
Veterinarian Docs www.veterinariandocs.com.br Fisiologia Fisiologia Básica -Introdução Quatro humores: sangue – quente e úmido fleuma – fria e úmida bile amarela – quente e seca bile negra – fria e seca Historia da fisiologia: -Claudio Galeno: pai da fisiologia experimental -Andreas Vesalius: Livro – a estrutura do corpo humano -Willian Harvey: estudo anatômico sobre o movimento do coração e do sangue em animais. Fisiologia comparada. -Luigi Galvani: poder da eletricidade no movimento muscular (eletrofisiologia). -Alessandro Volta: contração observada em patas seria resultado de eletricidade gerada pelos materiais utilizados para conectar nervos e músculos de rãs nas preparações. -Antonie Lavoisier: verificou que o principio de conservação de energia aplicava-se também aos seres vivos. Metabolismo -Johannes Müller: teoria celular, unidade fundamental dos seres vivos. -Carl Ludwig: invenção do quimógrafo. 1 www.veterinariandocs.com.br -Claude Bernard: introdução ao estudo da medicina experimental, liquido intersticial. -Walter B. Cannon: homeostasia Membrana celular É uma estrutura elástica delgada e deformável. É composta, quase inteiramente, por proteínas e lipídios. Sua composição aproximada é de: 55% proteínas 25% fosfolipídios 13% colesterol 4% outros lipídios 3% carboidratos A estrutura básica é a bicamada lipídica, que é uma fina película de lipídios, com apenas duas moléculas de espessura. Grandes moléculas de proteínas globulares estão inseridas, a intervalos irregulares, nessa película lipídica. Os constituintes mais abundantes das membranas celulares são proteínas e fosfolipídios. A molécula de fosfolipídio consiste de um grupo terminal polar e de duas cadeias não-polares, hidrofóbicas, ac. graxos. A bicamada lipídica básica é composta por moléculas de fosfolipídios. Uma extremidade de cada molécula de fosfolipídio é solúvel em água, isto é, hidrofílica. A outra extremidade só é solúvel em gorduras, isto é, é hidrofóbica. É a extremidade fosfatídica que é hidrofílica, enquanto a extremidade de AG é hidrofóbica. A característica especial da bicamada lipídica é a de que ela é um fluido, e não um sólido. Por conseguinte, partes da membrana podem, literalmente, fluir de um ponto para outro, ao longo da superfície da membrana. As proteínas, ou outras substâncias dissolvidas ou que flutuam na bicamada lipídica, se difundem para todas as áreas da membrana celular. As moléculas ajudam a determinar o grau de permeabilidade da bicamada aos constituintes hidrossolúveis dos líquidos corporais, e também é responsável pela fluidez da membrana. Proteínas da membrana celular: a maioria são glicoproteínas. São divididas em 2 porções: -Integrais: (intrínsecas) que atravessam a membrana, de um lado a outro. -Periféricas: (extrínsecas) que ficam presas a uma das superfícies da membrana, não penetrando no seu interior. 2 www.veterinariandocs.com.br Muitas das proteínas integrais formam canais (ou poros) estruturais pelos quais as moléculas de água e das substâncias hidrossolúveis, especialmente íons, podem se difundir entre os líquidos extracelulares e intracelulares. Outras proteínas integrais atuam como proteínas carreadoras para o transporte de substâncias que, de outro modo, não conseguiriam atravessar a membrana. Por vezes, chegam a transportar substâncias na direção oposta à sua direção natural de difusão, o que é chamado de ‘transporte ativo’, outras ainda atuam como enzimas. As proteínas periféricas atuam quase que inteiramente, como enzimas ou outros tipos de controladores do funcionamento celular. *Quando a composição iônica do meio é alterada, as proteínas periféricas são muitas vezes removidas da membrana. As proteínas integrais da membrana estão embebidas na membrana por intermédio de interações hidrofóbicas com o interior da membrana. Essas interações hidrofóbicas podem ser rompidas por detergentes. Modelo Mosaico-Fluido: muitos dos componentes moleculares de membranas celulares estão livres para se difundirem no plano da membrana. A maioria dos lipídios e proteínas se movem livremente no plano da dupla camada, mas eles saltam de uma monocamada fosfolipídica para outra em freqüências muito baixas. Composição lipídica: -fosfolipídios principais (fosfolipídios que contém colina ,lecitinas e esfingomielinas) são responsáveis pelas propriedades de permeabilidade passiva das membranas; -aminofosfolipídios; -fosfatidilglicerol, fosfatidilinositol e cardiolipina; *O bifosfato de fosfatidilinositol, quando clivado pela fosfatase C, libera Trifosfato de Inositol (IP3) e diacilglicerol. O IP3 é liberado no citossol, onde atua em receptores do retículo endoplasmático causando liberação de Ca2+. Colesterol: é o principal componente da membrana. Funciona como um ‘tampão’ de fluidez. Ele tende a manter a fluidez. Glicolipídios: os domínios de CHO dos glicolipídeos funcionam como receptores de antígenos. Composição protéica: variável com a função da membrana. Atuam como enzimas, proteínas de transporte, receptores para hormônios e neurotransmissores. Glicoproteínas: as cadeiras de CHO das glicoproteínas estão localizadas na superfície externa das membranas plasmáticas. Fibronectina: é uma glicoproteína, que contribui para a adesão das células. *Aquaporinas: canais de água na membrana. 3 www.veterinariandocs.com.br *Soro caseiro: transporte ativo secundário, com a entrada de 1 glicose e 1 Na há entrada de 250 moléculas de água (por isso hidrata). Líquido extracelular Líquido intracelular Na+ 142 mEq/l 10 mEq/l K+ 4 mEq/l 140 mEq/l Ca++ 2,4 mEq/l 0,0001 mEq/l Cl- 103 mEq/l 4 mEq/l Glicose 90 mg/dl 0 a 20 mg/dl Transporte pelas membranas, mas não através delas Endocitose: É o processo que permite ao material penetrar na célula, mas sem passar através da membrana plasmática. -Fagocitose: captação de moléculas -Pinocitose: captação de moléculas solúveis. *Depressões Revestidas: são regiões cobertas por uma proteína (Clatrina). Essas depressões revestidas estão envolvidas na endocitose mediada por receptor. Exocitose: é um processo que se assemelha ao reverso da endocitose. A liberação de neutrotransmissores pelas terminações nervosas pré-sinápticas é feita por exocitose. Fusão das vesículas de membrana: os conteúdos de um tipo de organela podem ser transferidos para outra organela pela fusão das membranas dessas organelas. Em algumas células, os produtos de secreção são transferidos do retículo endoplasmático para o aparelho de Golgi. A liberação de proteínas e neurotransmissores por exocitose requer a fusão de vesículas contendo as moléculas a serem liberadas com a membrana plasmática. Transporte de moléculas através das membranas Difusão: 2 tipos (facilitada e simples) -Difusão simples: o movimento cinético molecular das moléculas, ou dos íons, ocorre através da abertura da membrana, ou através dos espaços intermoleculares, sem necessidade da ligação com proteínas transportadoras na membrana. A velocidade da difusão é determinada pela quantidade de substância disponível, pela velocidade do movimento cinético e pelo nº e tamanho das aberturas na membrana celular. 4 www.veterinariandocs.com.br Ocorre por causa do movimento térmico aleatório, também denominado Movimento Browniano. A difusão resulta na distribuição uniforme de átomos ou moléculas. Difusão facilitada: necessita da interação com uma proteína transportadora das moléculas ou dos íons. A proteínas transportadora facilita a passagem das moléculas, ou dos íons, através da membrana por unir-se com eles quimicamente e lançá-los, através da membrana. *A difusão é rápida para distâncias pequenas. A regra do polegar é que uma molécula típica demora 1 ms para se difundir 1 μm. Entretanto, o tempo necessário para a difusão aumenta com o quadrado da distância na qual ela acontece. Ex.: o aumento da distância de difusão de 10x significa que será necessário tempo 100x mais longo para que ela se complete. Coeficiente de difusão (d): é proporcional à velocidade com que a molécula em difusão pode se mover no meio circundante. Quanto maior for a molécula, e mais viscoso o meio, menor é a D. O D é inversamente proporcional ao peso molecular. D = k.T/п.r.n K = constante de Boltzmann T = temperatura absoluta r = raio da macromolécula n = viscosidade do meio Permeabilidade à moléculas hidrossolúveis: as moléculas hidrossolúveis muito pequenas e sem carga atravessam as membranas celulares muito mais prontamente do que o previsto pela sua solubilidade lipídica. A água penetra nas membranas plasmáticas, cerca de 100x mais rapidamente do que o previsto pelo seu raio molecular. Há 2 razões para a alta e incomum permeabilidade à água: -moléculas pequenas e hidrossolúveis podem passar entre 2 moléculas adjacentes de fosfolipídios. -a existência de proteínas na membrana chamadas aquaporinas, que formam canais que permitem o fluxo elevado de água. *A permeabilidade das membranas para as moléculas hidrossolúveis e sem carga diminui à medida que aumenta o tamanho das moléculas. Os íons, devido à carga, são relativamente insolúveis em solventes lipídicos e as membranas não são muito permeáveis à maioria dos íons. A difusão iônica através da membrana ocorre principalmente através dos canais iônicos protéicos, que atravessam a membrana. 5 www.veterinariandocs.com.br As membranas plasmáticas têm proteínas específicas que permitem a transferência de metabólitos vitais para dentro e fora da célula. Osmose: é o fluxo de água que ocorre através de membrana ‘semipermeável’, do compartimento onde a concentração do soluto é menor para o compartimento onde a concentração é maior. A membrana semipermeável é definida como sendo uma membrana permeável à água, mas impermeável à solutos. A osmose acontece porque a presença de soluto diminui o potencial químico da água. A água tende a fluir de onde seu potencial químico é maior para onde seu potencial químico é menor. A diminuição do potencial químico da água em uma solução também reduz a pressão de vapor, abaixa o ponto de congelamento e aumenta o ponto de ebulição da solução, comparada com a água pura. Movimento efetivo de água através da membrana celular, fazendo com que a célula fique inchada ou murcha, dependendo da direção do movimento efetivo. *pressão de vapor: é a pressão exercida por um vapor quando este está em equilíbrio dinâmico com o líquido que lhe deu origem, ou seja, a quantidade de líquido (solução) que evapora é a mesma que se condensa. Permeabilidade dos canais protéicos: muitos dos canais protéicos são altamente seletivos para o transporte de um ou mais íons ou moléculas específicos. Isso resulta das características do próprio canal, como o diâmetro, o formato e a natureza das cargas elétricas ao longo das superfícies internas. Ex.: canais de sódio. As superfícies internas do canal têm cargas fortemente negativas. Essas potentes cargas negativas atraem os pequenos íons desidratados de sódio (Na) para dentro desses canais, na verdade, conduzindo o sódio para longe de suas moléculas hidratantes de água. Uma vez dentro do canal, os íons sódio se difundem em qualquer direção. Assim, o canal de sódio é especificamente seletivo para a passagem dos íons sódio. Os canais de potássio são menores que os de sódio e não estão carregados negativamente. Portanto, não existe qualquer força atrativa poderosa atraindo os íons para o interior do canal, razão pela qual os íons não são afastados das moléculas de água que irão hidratá-los. *a forma hidratada do potássio é menos que a do sódio. Pois o sódio atrai muito mais moléculas de água que o potássio. Sódio (Na): fora da célula Potássio (K): dentro da célula Comportas dos canais protéicos: controlam a permeabilidade iônica dos canais. A abertura e o fechamento são controlados por 2 mecanismos principais: 6 www.veterinariandocs.com.br -Comportas por voltagem: as comportas se abrem quando há uma perda de carga no interior da célula, e as comportas se abrem para que entre íons. Ex.: sódio, potássio -Comportas químicas: são controladas por ligações de substâncias químicas às proteínas, acarretando alteração conformacional. Ex.: acetilcolina. Difusão facilitada: também denominada Difusão mediada por carreador. A difusão facilitada difere da difusão simples através de um canal aberto da seguinte maneira: apesar de a velocidade de difusão através do canal aberto aumentar proporcionalmente com a concentração da substância a ser difundida, na difusão facilitada a velocidade de difusão aproxima-se do valor máximo, quando a concentração aumenta. *A velocidade de difusão da difusão facilitada é limitada. O motivo por essa velocidade ser limitada é pelo motivo de a proteína carreadora ter em seu interior receptor específico para a substância a ser carregada. A proteínas recebe a substância, há alterações conformacionais, e depois a substância é liberada para o interior da célula. E assim não há como aumentar a velocidade de difusão, se há só um receptor por proteína. Fatores que afetam a velocidade efetiva de difusão 1-Efeito da diferença de concentração sobre a difusão efetiva através da membrana: a velocidade efetiva de difusão para dentro da célula é proporcionalmente à concentração no exterior menos a concentração interna. 2-Efeito do potencial elétrico da membrana sobre a difusão de íons – equação de Nernst: os íons se movimentarão através da membrana mesmo quando não existe qualquer diferença de concentração capaz de causar sua movimentação efetiva. Equação de nernst: a equação é usada para computar a diferença de potencial elétrico necessária para produzir uma força elétrica, que é igual e oposta a força de concentração. FEM = ± 61 log C1 FEM: força eletromotriz C2 C1: concentração 1 C2: concentração 2 Efeito da diferença de pressão através da membrana: quando a pressão é mais alta em um dos lados da membrana que no outro, significa que a soma de todas as 7 www.veterinariandocs.com.br forças das moléculas que golpeiam os canais nesse lado da membrana é maior que o outro lado. 1- Se a diferença de potencial medida através da membrana é igual à diferença de potencial calculada pela equação de Nernst, este está em equilíbrio eletroquímico através da membrana, e não haverá fluxo. 2- Se o potencial elétrico tem o mesmo sinal daquele calculado pela equação de Nernst, mas é maior em módulo do que o valor calculado, a força elétrica é maior que a força de concentração. Portanto, o movimento resultante tende a ocorrer na direção da força elétrica. 3- Quando a diferença de potencial elétrico é do mesmo sinal, mas é numericamente menor que a calculada menor do que o calculado pela equação de Nersnt, a força de concentração é maior que a elétrica. Por isso, o movimento resultante tende a ocorrer na direção da força de concentração. Osmolalidade Serve para enunciar a concentração de uma solução em termos de número de partículas. 1 molécula de glicose com PM= 180g é igual à 1 osmol de glicose Porque a molécula de glicose não se dissocia em meio aquoso. 1 molécula de cloreto de sódio com PM= 58,5g é igual à 2 osmois de NaCl. Porque a molécula de NaCl se dissocia em 2, e o nº de moléculas osmoticamente ativas é 2x maior que a do soluto não dissociado. Transporte ativo: Os sistemas de transporte ativo permitem o transporte de seus substratos contra gradientes de concentração ou de potencial eletroquímico. Como estes processos requerem energia, os processos ativos devem estar ligados ao metabolismo energético de alguma maneira.Os transportadores ativos utilizam ATP. Às vezes é necessária a grande concentração de substância no líquido intracelular, apesar do líquido extracelular conter apenas pequena concentração. -Primário: quando a energia deriva diretamente do fracionamento do ATP -Secundário: quando a energia deriva da que foi armazenada na forma de diferença de concentração iônica entre os 2 lados da membrana, geradas inicialmente pelo transporte ativo primário. Bomba de sódio-potássio: um processo que bombeia Na para fora da célula e K para dentro da célula. Essa bomba é responsável pela manutenção das diferenças de 8 www.veterinariandocs.com.br concentração do sódio e do potássio e também pelo estabelecimento da voltagem negativa dentro das células. -Componentes Físicos da Bomba: a proteína carreadora é um complexo de duas proteínas globulares separadas, uma maior denominada subunidade α e a menor subunidade denominada β. A proteína maior tem 3 funções: -tem 3 locais receptores para a fixação dos íons sódio na porção da proteína que protrude no interior da célula. -tem 2 locais receptores para íons potássio no exterior. -a porção interna dessa proteína, perto dos locais de fixação do sódio, tem atividade ATPase. -Como ocorre: quando 2 íons K fixam-se no exterior da proteína carreadora, 3 íons Na fixam-se na parte interna, a função da ATPase é ativada. A seguir ocorre a clivagem de uma molécula de ATP, que será fracionada à ADP, com liberação de energia contida em uma ligação fosfato de alta energia. E assim ocorre uma transformação conformacional na proteína, fazendo com que os 3 Na saiam e os 2 K entrem. -Importância: consiste em controlar o volume das células. Sem o funcionamento desta, a maioria das células do corpo sofreria tumefação. Como: dentro da célula existe grande nº de proteínas e de outros compostos orgânicos que não conseguem sair da célula. A maioria deles tem carga negativa e, portanto, coleta também, ao seu redor, inúmeros íons positivos. E assim, todas essas substâncias tendem a causar osmose da água para dentro da célula. Se isso não for impedido, a célula incha até explodir. Pelo motivo de saírem 3 Na e entrarem 2 K, há uma perda de íons para fora da célula e assim também mandando água para fora da célula. Se a célula percebe que está ‘inchando’ (entrando água), ativa o mecanismo de bomba de sódio-potássio. *A bomba de sódio-potássio é considerada ‘eletrogênica’, porque define uma diferença de cargas fora e dentro da célula (potencial elétrico). Transporte ativo primário de cálcio: O transportador tem duas formas básicas denominadas E1 e E2. Os 2 sítios de ligação do Ca2+ na conformação E1 tem alta afinidade pelo Ca e são acessíveis pelo lado do citossol. Na conformação E2 os sítios de ligação de Ca tem afinidade muito baixa e são acessíveis na luz do Retículo Sarcoplasmático. Existe 2 bombas de cálcio. Uma que fica na membrana e bombeia o cálcio para fora da célula e outra que fica dentro do citoplasma e bombeia Ca++ para organelas vesiculares internas. A proteínas carreadora funciona como ATPase, clivando o ATP à ADP. Como o Ca++ é um segundo mensageiro importante, o nível citossólico para o Ca++ está sujeito a complexa regulação. Entre as proteínas transportadoras da membrana 9 www.veterinariandocs.com.br que participam na regulação do nível citossólico de Ca++ estão as Ca++ - ATPases localizadas nas membranas plasmáticas. A Ca++ - ATPase da membrana plasmática é regulada pela calmodulina. Na presença de Ca++ em concentração submicromolar, o complexo Ca++ com a calmodulina liga-se a sítio específico da Ca++ - ATPase da membrana plasmática. Esta ligação causa a dissociação do peptídeo auto-inibitório do domínio de ligação do ATP e, assim, ativa a Ca++ - ATPase. A maioria das células armazena Ca++ no retículo endoplasmático ou em outra vesículas de armazenamento intracelular, tal com o retículo sarcoplasmático das células musculares. O Ca++ é concentrado nessa vesículas pelas Ca++ - ATPases. *SERCA: Ca++ - ATPases dos retículos sarcoplasmáticos e endoplasmáticos. Troca de Na+ e Ca++: Certas células eletricamente excitáveis, como as cardíacas, têm mais um mecanismo adicional para controlar o nível intracelular de Ca++. Nas células cardíacas, a diminuição do Ca++ intracelular, que ocorre em cada diástole, é causada pela proteína trocadora de sódio/cálcio e pela Ca++ - ATPase do RS. O trocador de Na/Ca é estimulado em níveis micromolares de Ca++, pela ligação do complexo Ca++calmodulina, a um local específico da proteína trocadora. Co-transporte de Na+, K+ e Cl-: Muitas células, epiteliais e não-epiteliais, contêm uma proteína de membrana plasmática que medeia o transporte simultâneo (co-transporte) de Na+, K+ e Cl- do fluído extracelular para o citossol. O co-tranportador é ativado pelo murchamento da célula, o que leva a um influxo de Na+, K+ e Cl- e estes geram uma força osmótica para restaurar o volume celular. *O transportador Na+, K+ e Cl- é especificamente inibido por drogas como furosemida e bumetanida (diuréticos de alça). Transporte facilitado de glicose: A glicose difunde muito pouco pela membrana plasmática. As membranas plasmáticas de diferentes tipos de células contêm proteínas transportadoras, que medeiam o transporte facilitado de glicose e de monossacarídeos relacionados. Eritrócitos, hepatócitos, adipócitos e células musculares possuem transportadores de glicose. -Em adipócitos e células musculares, a taxa de transporte de glicose através da membrana plasmática é aumentada pela insulina. -Nos eritrócitos e em certos neurônios o transporte de glicose é estimulado por níveis reduzidos de ATP, e por níveis aumentados de ADP e AMP. A anóxia no 10 www.veterinariandocs.com.br músculo cardíaco e o exercício no músculo esquelético estimulam o transporte de glicose. Co-transporte de glicose juntamente com íons sódio: A proteína carreadora tem 2 locais de fixação externamente, um para o sódio outro para a glicose. Além disso a concentração de Na é muito alta fora da célula, o que proporciona a energia para o transporte. O Na e a glicose são transportados juntamente nesse mecanismo. Transporte Facilitado: ocorre via um transportador que não necessita de energia. *não é deprimido pelos inibidores metabólicos. *eles não podem transportar substâncias contra seu gradiente de concentração. Ex.: monossacarídeos Propriedades Coligativas 1-Pressão Osmótica: a pressão do lado A que é suficiente para anular a entrada de água pura é denominada Pressão Osmótica da solução do lado A. uma pressão no sentido inverso ao da osmose ou no mínimo com a mesma intensidade daquele que o solvente faz para atravessar a membrana semipermeável. A essa pressão, capaz de impedir o fenômeno da osmose. П = RT (фic) П: pressão osmótica R: constante de gás ideal T: temperatura absoluta Ф: coeficiente osmótico i: numero de íons formados pela dissociação de uma molécula de soluto. c: concentração molar do soluto Transporte através da membrana mediado por proteína: algumas substâncias entram e saem da célula via proteínas intrínsecas (transportadores) ou canais. Os processos de transporte mediados por proteína são capazes de transportar substâncias através das membranas muito mais rapidamente que a difusão simples. Um transportador liga-se ao soluto a ser transportado em um lado da membrana, e então o transportador sofre mudanças conformacionais que permitem ao soluto ser liberado no outro lado da membrana. Se o transportador não tem ligação com energia metabólica, o soluto transportado irá fluir do lado da membrana onde é mais concentrado para o lado onde é menos concentrado, constituindo o denominado 11 www.veterinariandocs.com.br Transporte Facilitado. Certos transportadores estão acoplados à energia metabólica, e podem usar a energia para transportar o soluto contra seu gradiente de energia, de onde ele é menos concentrado para onde é mais concentrado. Este é chamado Transporte Ativo. As duas classes mais importantes de canais iônicos são os controlados por voltagem e os controlados por ligantes. A probabilidade de um canal controlado por voltagem estar aberto depende do valor da diferença de voltagem transmembranosa e a probabilidade de um canal controlado por ligante estar aberto depende da concentração da substância (ligante), como acetilcolina, que regula o canal. Propriedades do transporte mediado: 1- Uma substância é transportada muito mais rapidamente por transporte mediado do que as outras moléculas que têm o mesmo peso molecular e mesma solubilidade lipídica, mas atravessam a membrana por difusão simples. 2- O transporte mostra a cinética de saturação. Quando a concentração do composto transportado é aumentada, a taxa de transporte primeiramente aumenta, mas há uma concentração acima da qual a taxa de transporte não aumenta mais. Nesse ponto, o sistema de transporte está saturado com o composto transportado. 3- A proteína mediadora apresenta especificidade química: somente as moléculas com a estrutura química requerida são transportadas. A especificidade da maioria dos sistemas de transporte não é absoluta, e em geral é mais ampla do que a especificidade da maioria da enzimas. Todavia, a relação chave-fechadura entre uma enzima e seu substrato também pode ser aplicada às proteínas transportadoras. Na+, K+ - ATPase: Oscila entre 2 estados (E1 e E2). Na conformação E1 os sítios de ligação dos íons estão voltados para o citossol, e têm alta afinidade pelo Na+ e baixa afinidade pelo K+. O Na+ do citossol é liberado para no líquido extracelular. E o K+ é liberado no citossol. Em cada ciclo da bomba, uma molécula de ATP é hidrolisada, três Na+ são ejetados do citossol e dois K+ são captados pelo citossol. Devido aos gradientes iônicos criados pela bomba, o Na+ tende a se difundir passivamente de volta para o citossol e o K+ tende a se difundir para fora da célula. Para o fluido extracelular. Uma vez criado, um gradiente de concentração, representa uma forma de energia potencial química que pode ser utilizada para realizar trabalho. *No intestino delgado, a glicose e a galactose são absorvidas por transporte ativo secundário acoplado ao Na+. A presença de Na+ no lúmen aumenta a absorção de glicose, e vice-versa. A terapia de reidratação oral é freqüentemente empregada nas diarréias graves. Os pacientes ingerem uma solução contendo NaCl, glicose, K+ e 12 www.veterinariandocs.com.br HCO3-. A absorção de Na+ e glicose no intestino delgado promove a absorção osmótica de água, o que facilita a reidratação do paciente. Potenciais de membrana e potenciais de ação: Equação de Goldman-Hodgkin-Kartz: permite o cálculo do potencial de membrana no interior da membrana, quando dois íons positivos univalentes (Na, K) e um íon negativo (Cl) estão envolvidos. FEM(mV)= - 61.log. CNai.PNa + CKi.Pk + CClo.PCl CNao.PNa + Cko.Pk + CCli.PCl Ci: concentração interna do íon Co: concentração externa do íon P: permeabilidade do íon *Se a membrana tem permeabilidade zero para os íons potássio e cloreto, o potencial da membrana passa a ser determinado exclusivamente pelo Na. Ficando assim a Equação de Nernst. *As variações nas permeabilidades do sódio e potássio são as principais responsáveis pela condução dos sinais nos nervos. Equilíbrios iônicos e potenciais de repouso: O citoplasma em geral é eletricamente negativo em relação ao fluido extracelular. Essa diferença de potencial elétrico através da membrana plasmática em uma célula em repouso é denominada potencial de repouso de uma membrana. O potencial de repouso da membrana tem papel central na excitabilidade de células musculares e neurais, e em outras respostas celulares. Equilíbrio iônico: a grandeza que permite comparar as contribuições relativas da concentração iônica e do potencial elétrico ao movimento de um íon é denominada Potencial Eletroquímico de um íon. μA(X+) – μb (X+) = R.T.ln[X+]A + zF (EA + EB) [X+]B Δμ= diferença de potencial eletroquímico R= constante dos gases idéias T= temperatura absoluta ln[X+]A = logaritimo natural da razão das concentrações de X+ [X+]B z= valência (2+ para o Ca, -1 para o Cl) 13 www.veterinariandocs.com.br F= número de Faraday (EA – EB)= diferença de potencial elétrico O 1º termo da equação R.T.ln[X+]A /[X+]B é a tendência de X+ se mover de A para B por causa da diferença de concentração O 2º termo zF (EA + EB) é a tendência de os íons se moverem de A para B por causa da diferença de potencial elétrico. Δμ descreve a diferença de energia potencial que existe entre um mol de íon X do lado A e um mol de íons X do lado B, que resulta tanto da diferença de concentração quanto da diferença de potencial elétrico. *Os íons X tendem a se mover espontaneamente de um maior potencial eletroquímico para um menor. Se Δμ é positivo, os íons tendem a se mover de A para B. Se Δμ é negativo, os íons tendem a se mover do lado B para A. Se μA > μb, os íons tendem a fluir espontaneamente do lado A para o B. Equilíbrio de gibbs-donnan: O citoplasma contém proteínas, polifosfatos, ac. nucléicos e outras substâncias que não podem atravessar a membrana. As propriedades estacionárias dessa mistura de íons permanentes e não permanentes são descritas pelo Equilíbrio de Gibbs-Donnan. Equação de condutância de corda: Especifica que o potencial da membrana é uma média ponderada dos potenciais de equilíbrio de todos os íons para os quais a membrana é permeável. A maneira pela qual a interação de gradientes de íons cria um potencial de repouso na membrana. Considerando a concentração de K+, Na+ e Cl- na membrana plasmática de uma célula: Em= gk Ek + gNa ENa + gCl ECl Σg Σg Σg g= condutâncias da membrana para os íons indicados E= potencial de equilíbrio *Quanto mais permeável a membrana é a um íon, maior é a condutância da membrana para aquele íon. Potencial de repouso: O potencial de repouso da membrana das fibras nervosas de grande diâmetro, quando não estão transmitindo sinais nervosos, é cerca de 90 mV. Isto é, o potencial no interior da fibra é 90 mV mais negativo que o potencial no líquido extracelular. 14 www.veterinariandocs.com.br *Transporte Ativo de Íons Sódio e Potássio, através da membrana: Bomba de Sódio e Potássio. -Todas as membranas celulares apresentam bomba de sódio-potássio. Origem do potencial de repouso normal de membrana: 1-Contribuição do Potencial de Difusão do Potássio: usando a fórmula de Nernst o potencial seria de -94 mV. Portanto, se os íons potássio fossem o único causador do potencial de repouso, esse potencial de repouso no interior da fibra também seria igual à – 94 mV. 2-Contribuição da Difusão de Sódio através da membrana neural: usando a fórmula de Nernst o potencial seria de + 61 mV. Como interagir os 2 potenciais? Usando a equação de Goldman.Sendo que o K é mais permeável que o Na, lógico que o K contribui muito mais para o potencial da membrana. Usando-se a equação de Goldman, obtem-se um potencial interno da membrana de – 86 mV. 3-Contribuição da Bomba de Na+ e K+: O fato de mais íons sódio serem bombeados para o exterior do que mais potássio para o interior produz uma perda contínua de cargas positivas do interior da membrana, isso cria um adicional de negatividade (cerca de – 4 mV) no interior. Portanto o verdadeiro potencial de membrana, com o qual todos esses fatores operam ao mesmo tempo é cerca de – 90 mV. Potencial de ação: é uma rápida alteração no potencial da membrana seguida por um retorno ao potencial de repouso da membrana. -Se propaga com a mesma forma e tamanho ao longo de todo o comprimento de um nervo ou célula muscular. -É a base da capacidade de transmissão de sinais das células nervosas. -Permite que todo o comprimento destas longas células se contraia quase simultaneamente. -As proteínas dos canais para íons dependentes de voltagem na membrana plasmática são as responsáveis pelos potenciais de ação. -Respostas Sublimiares: a Resposta Local Uma alteração do potencial da membrana de -90 mV para –70 mV é uma despolarização porque diminui a diferença de potencial. Uma alteração do potencial da membrana de -90 mV para -110 mV é uma hiperpolarização. 15 www.veterinariandocs.com.br *Diz-se que a mudança de potencial é conduzida com atenuação. (o tamanho da alteração de potencial depende da distância entre o eletrodo e o ponto de injeção da corrente. O tamanho da modificação do potencial diminui exponencialmente com a distância do local de injeção da corrente). Se pulsos de corrente despolarizante progressivamente maiores são aplicados à membrana plasmática, um potencial de membrana limiar pode ser atingido no qual um tipo diferente de resposta, o potencial de ação, ocorre. O potencial de ação difere das respostas locais em dois importantes aspectos: 1- Ele é uma resposta muito maior, na qual a polaridade do potencial de membrana reverte (o interior fica + e o exterior -). 2- O potencial de ação se propaga sem atenuação por todo o comprimento do nervo ou fibra nervosa. *Ao contrário de uma resposta local, seu tamanho não diminui com a distância. -Tudo-ou-nada: quando se aplica um estímulo superior ao estímulo do limiar, o tamanho e a forma do potencial de ação não se alteram, o tamanho do potencial de ação não aumenta com estímulos de maiores intensidades. Um estímulo, ou não deflagra um potencial de ação ou produz um potencial de ação completo. Mecanismos iônicos dos potenciais de ação O potencial de membrana finalmente retorna ao valor de repouso quase tão rapidamente quanto se despolarizou. Após a repolarização, observa-se uma hiperpolarização transiente que é conhecida como ‘pós-potencial hiperpolarizante’. Mecanismos iônicos do potencial de ação em um axônio gigante de lula: O potencial de repouso da membrana (Em) é aproximadamente -70 mV. O potencial de equilíbrio do K+ (Ek) é aproximadamente -100 mV. Uma elevação na gk hiperpolariza e uma diminuição na gk despolariza a membrana. Uma elevação na gNa despolariza e causa reversão da polaridade da membrana. O potencial de ação é causado por elevações sucessivas na condutância da membrana plasmática para os íons de sódio e potássio. A condutância para o Na+, gNa, aumenta muito rapidamente durante a parte inicial do potencial de ação. A condutância ao potássio, gK, aumenta mais lentamente, atinge seu pico por volta do meio da fase de repolarização e depois retorna mais lentamente para os níveis de repouso. *A elevação da condutância a cada um dos íons irá aumentar sua capacidade de trazer o potencial de membrana para o seu potencial de equilíbrio. A rápida elevação na 16 www.veterinariandocs.com.br gNa durante a fase inicial do potencial de ação faz com que o potencial de membrana se mova para o potencial de equilíbrio do Na+. O rápido retorno do potencial de membrana para valores de repouso é causado pela rápida redução de gNa e pela elevação progressiva de gK. Canais para íons e Comportas: Hodgkin e Huxley propuseram que as correntes dos íons Na e K passam por canais distintos, cada um deles com características próprias. Canal para Na+: várias α-hélices que atravessam completamente a membrana e provavelmente circundam o canal para o íon. O canal para Na possui tanto comporta de ativação como a comporta de inativação, que são responsáveis pelas alterações na gNa durante o potencial de ação. Para atravessar a parte mais estreita do canal, conhecida como ‘filtro de seletividade’, um íon deve eliminar a maior parte da água de hidratação substituindo a interação com a água por interação com resíduos de aminoácidos. Comportamento unitário dos canais para íons: Os canais para íons oscilam espontaneamente entre dois estados de condutância, em um estado aberto e outro fechado. A probabilidade de cada canal estar no estado aberto é maior quando a membrana está despolarizada. Potenciais de ação em músculo cardíaco e liso M.Cardíaco: a rápida despolarização inicial e reversão da polaridade são causadas pela entrada rápida de Na no meio intracelular, através de canais para Na que são semelhantes aos canais para Na dos nervos e músculos. Os canais para Na são chamados de canais rápidos. Canais lentos: pertencem a uma classe particular de canais para Ca, chamados de canais para Ca de tipo L. O Ca penetra na célula e é essencial para a concentração celular, pois estimula a liberação de mais Ca do RS. A repolarização da célula ventricular ocorre pelo fechamento dos canais para Ca de tipo L e pela abertura muito mais tardia de canais para K. Acomodação: Quando um nervo ou célula muscular são lentamente despolarizados, o limiar normal pode ser ultrapassado sem que um potencial de ação seja disparado. Velocidade de condução: 17 www.veterinariandocs.com.br A velocidade da condução eletrônica em um nervo ou fibra muscular é determinada pelas propriedades elétricas do citoplasma e da membrana plasmática da célula. Efeito do diâmetro da fibra: maior diâmetro apresenta maior velocidade, por causa da diminuição da resistência. Efeito da mielinização: a mielina consiste da membrana plasmática da células de Schwann, que se enrola ao redor da fibra nervosa, isolando-a. Interrupções na bainha de mielina ocorrem a cada intervalo. Estas regiões são conhecidas como Nós de Ranvier. A mielina aumenta a condução da fibra. A mielina aumenta a velocidade por: -eleva a constante de comprimento do axônio -diminui a capacitância do axônio -restringi a geração de potenciais de ação dos nós de ranvier. *Os canais para Na e para K que participam do potencial de ação são altamente concentrados nos nós de Ranvier. Portanto, o potencial de ação é regenerado somente nos nós de Ranvier. – Condução Saltarória Transmissão sináptica: Sinapse: estrutura especializada que permite comunicação elétrica entre as células. Existem dois tipos: elétrica e química. Em uma sinapse elétrica as duas células estão conectadas por ‘canal juncional’. E em uma sinapse química o neurônio présináptico libera substâncias neurotransmissoras que se liga a receptores protéicos específicos na membrana plasmática das células pós-sinápticas para alterar o potencial de membrana destas. Junção neuromuscular: (placa motoras terminais) São as sinapses entre os axônios de neurônios motores e fibras musculares esqueléticas. Estrutura da junção neuromuscular: Fendas Sinápticas: onde situam-se as terminações do axônio motor, na superfície das células musculares. Dobras Juncionais: membrana plasmática que reveste as células musculares, formando dobras. Nas terminações do axônio há muitas vesículas sinápticas, de superfícies lisas, que contêm acetilcolina. Fenda Juncional: contém material amorfo rico em CHO, separa a terminação do axônio e a célula muscular. 18 www.veterinariandocs.com.br As moléculas do receptor para Acetilcolina estão concentradas nas aberturas das dobras juncionais. As vesículas sinápticas nas terminações neurais e os sítios especializados são chamados de ‘Zonas Ativas’ Acetilcolinesterase: enzima que cliva acetilcolina em acetato e colina, é distribuída na superfície externa da membrana pós-juncional. Sinapse da trasmissão neuromuscular: Potencial de ação no terminal do axônio motor pré-sináptico, aumentando a permeabilidade e influxo de Ca2+ no terminal axônio. Liberação de acetilcolina da vesícula sináptica na fenda sináptica e difusão de acetilcolina para a membrana pósjuncional. Ligação da acetilcolina com receptores específicos na membrana pósjuncional. Aumento da permeabilidade da membrana pós-juncional a Na+ e K+ causa PPM (potencial de placa motora). Despolarização de áreas de membrana muscular adjacente à placa motora e disparo de um potencial de ação. *PPM: é transitório porque a acetilcolina é rapidamente hidrolisada em colina e acetato. Sinapse de acetilcolina: acetilcolina é produzida por condensação de acetil coenzima A e colina. Neurônios motores e seus axônios estão entre as poucas células capazes de sintetizar acetilcolina. Embora acetil-CoA seja produzida por neurônios, a colina não é sintetizada por neurônios motores. A colina é obtida por captação ativa do fluido extracelular. Liberação quântica de transmissor: Acetilcolina não é liberada continuamente por terminais nervosos pré-juncionais, é liberado em pacotes. A quantidade de acetilcolina contida em uma vesícula é chamada de ‘Quantum’ de acetilcolina. *PPMMs: potenciais de placa motora em miniatura – ocorrem mesmo que os neurônios motores não sejam estimulados. Cada PPMM despolariza a membrana pós-juncional em aproximadamente 0,4 mV. Esta despolarização não é suficiente para desencadear um potencial na membrana muscular. Ação da colinesterase e recaptação da colina: A acetilcolinesterase está concentrada na superfície externa da membrana pósjuncional e na lâmina externa. Drogas que inibem esta enzima são chamadas de Anticolinesterase. O neurônio motor não pode sintetizar colina. Portanto, a recaptação das fendas sinápticas fornece a colina necessária para a ressíntese de acetilcolina. Mecanismo iônico do potencial de placa motora: Os canais para cátions que a acetilcolina abre na membrana pós-sináptica diferem dos canais para cátions dependentes de voltagem dos nervos e músculos pelo 19 www.veterinariandocs.com.br fato de operarem independentemente do potencial de membrana. Os canais pósjuncionais são ativados pela ação da acetilcolina e não pelo potencial de membrana. Sinápses elétricas: Canais juncionais: conectam as duas células que fazem uma sinapse elétrica. *Uma mudança do potencial de membrana de uma célula é transmitida para a outra célula por fluxo direto de corrente através dos canais juncionais. Geralmente sinapses elétricas permitem condução em ambas as direções. Neste aspecto diferem das sinapses químicas. Certas sinapses elétricas conduzem com menor resistência em uma direção do que em outra, isso chama-se de ‘Retificação’. Conéxon: arranjo hexagonal – canal juncional Conexina: cada uma das subunidades é uma proteína única. *Os canais podem se fechar em resposta ao aumento da concentração intracelular de Ca2+ ou H+ em uma das células. Sinapses elétricas ocorrem por todo o sistema nervoso central e periférico de vertebrados. São úteis em vias reflexas, nas quais há transmissão rápida entre células. Sinapses químicas: Quando um neurônio faz uma sinapse química com outro, o terminal nervoso pré-sináptico se alarga para formar um ‘botão terminal’. Por causa da estrutura e organização da sinapse química, a condução é unidirecional. A condução unidirecional nas sinapses químicas contribui para a organização do sistema nervoso central dos vertebrados. Nas sinapses químicas, o transmissor liberado pelos neurônios pré-sinápticos altera a condutância da membrana plasmática pós-sináptica a um ou mais íons. *Potenciais de ação NÃO são produzidos na sinapse. Eventualmente, a despolarização ou hiperpolarização é conduzida ao cone de inserção do axônio, região do neurônio onde o axônio se origina. *A região do cone de inserção-segmento inicial tem limiar mais baixo que a membrana plasmática. Relações de entrada e saída: Sinapses podem ser classificadas em um-para-um, umpara-muitos ou muito-para-um com base na relação entre entrada e saída. Um-Para-Um: como na junção neuromuscular, a entrada e a saída são as mesmas. Um-Para-Muitos: um potencial de ação único na célula pré-sináptica provoca vários potenciais de ação na célula pós-sináptica. 20 www.veterinariandocs.com.br Muitos- Para-Um: um potencial de ação em uma célula pré-sináptica, não é suficiente para estimular a célula pós-sináptica a disparar um potencial de ação. Então necessita de vários potenciais de ação de diferentes neurônios em uma única célula, para estimulá-la. Potenciais pós-sinápticos inibitórios e excitatórios: Um neurônio motor espinhal responde de duas maneiras ao potencial de ação dos neurônios que fazem sinapse com ele. A resposta pós-sináptica é uma despolarização transitória ou uma hiperpolarização transitória. PPSE: (potencial pós-sináptico excitatório) = despolarização, levando o neurônio próximo ao limiar. PPSI: (potencial pós-sináptico inibitório) = hiperpolarização, levando o neurônio longe do limiar. Somação das entradas sinápticas: Somação Espacial: ocorre quando duas entradas separadas chegam quase simultaneamente. Os dois potenciais pós-sinápticos são então adicionados. Se: PPSE + PPSE = despolarização PPSE + PPSI = uma cancela a outra Somação Temporal: ocorre quando dois ou mais potenciais de ação em um neurônio pré-sináptico ocorrem em rápida sucessão. Modulação da atividade sináptica: Quando um axônio pré-sináptico é estimulado repetidamente, a resposta póssináptica pode aumentar a cada estimulo. Este fenômeno é chamado ‘Facilitação’. Quando um neurônio pré-sináptico é estimulado tetanicamente (muitos estímulos em freqüência alta) por vários segundos, ocorre uma ‘Potenciação PósTetânica’, que é um aumento da resposta pós-sináptica. Fadiga Sináptica: depressão neuromuscular na junção neuromuscular. Em alguns casos, um decréscimo no conteúdo quântico (a quantidade de transmissor por vesícula sináptica) contribui para a fadiga sináptica. 21 www.veterinariandocs.com.br Mecanismos iônicos de potenciais pós-sinápticos em neurônio motor espinhal: PPSE: é causado por um aumento transitório na condutância da membrana póssináptica ao Na+ e ao K+ em resposta ao neurotransmissor. PPSI: aumento da concentração de Cl- na membrana pós-juncional. Que resulta da liberação de transmissores na sinapse inibitória, permite a entrada de Cl- na célula pós-sináptica, o que a hiperpolariza. Neutrotransmissores Acetilcolina: é o transmissor usado por todos os axônios motores que emergem da espinha dorsal. Aminas Biogênicas Transmissoras: nerepinefrina, epinefrina, dopamina, serotonina e histamina. *Dopamina, norepinefrina e epinefrina são Catecolaminas. Aminoácidos Transmissores – Glicina: o aminoácido mais simples. É um neurotransmissor inibitório liberado por certos interneurônios espinhais. Ácido γ-aminobutítico (GABA): presente somente em certos neurônios no SNC. Funciona como um transmissor inibitório. *Os receptores pós-sinápticos para a glicina e GABA são ambos canais de Cl- ativados por ligantes, que permitem o influxo de Cl- para hiperpolarizar o neurônio póssináptico. Oxido Nítrico – NO: é um transmissor entre neurônios inibitórios dos sistema nervoso entérico. Peptídeos Neuroativos Certas células liberam peptídeos que atuam em baixas concentrações para excitar ou inibir neurônios. Afetam os neurônios em concentrações mais baixas e a ação perdura mais que os neurotransmissores. São sintetizados nos corpos celulares dos neurônios. Neuropeptídeos podem atuar como hormônios, como neurotransmissores ou como neuromoduladores. *Neurotransmissor: muda a condutância da célula-alvo a um ou mais íons. *Neuromodulador: modula a transmissão sináptica. Pode atuar prés-sinapticamente para mudar a quantidade de transmissor liberado em resposta a um potencial de ação. Peptídeos Opióides: 3 classes 22 www.veterinariandocs.com.br -encefalinas -endorfinas -dinorfinas Opióides inibem os neurônios no cérebro envolvidos na percepção da dor. Peptídeos opióides estão entre os analgésicos mais potentes conhecidos. Outros neuromoduladores: ATP e adenosina funcionam como neuromoduladores no SNC, autônomo e periférico. Receptores para Neutransmissores Receptores Ionotrópicos: são canais para íons ativados por ligantes. Receptores Metabotrópicos: não são canais para íons ativados por ligantes, mas participam do 1º passo de uma cascata de sinalização. Receptores inibitórios: Receptores para GABA e Glicina A sinapse inibitória mais comum no SNC usa a glicina ou GABA como transmissor. Glicina: na medula espinhal GABA: no cérebro Receptores para GABA e glicina são canais para Cl- ativados por ligantes que medeiam o influxo de Cl- nos neurônios. A corrente de Cl hiperpolariza e, assim, inibe o neurônio. *Barbitúricos são usados como sedativos e anticonbulsivantes. Se liga a distintos sítios nos receptores para GABA e aumentam a probabilidade de abertura de canais de Cl, em resposta ao GABA. Mecanismo molecular e celular de liberação de neutransmissor: As vesículas sinápticas menores, que contêm as pequenas moléculas clássicas de neurotransmissores, são carregadas com o transmissor nas terminações nervosas. O neurotransmissor clássico é sintetizado nas terminações nervosas e acumulado nas vesículas sinápticas por transporte ativo secundário que catalisa a troca de neurotransmissores por H. As vesículas grandes que liberam neuropeptídeos podem fundir-se com a membrana pré-sináptica em múltiplos locais para liberar seu conteúdo. Devido à entrada de Ca nos terminais nervosos, através de canais para cálcio dependentes de voltagem, a concentração local de Ca nos terminais nervosos eleva-se e desencadeia a fusão de uma pequena fração das vesículas ancoradas. As vesículas fundidas liberam o transmissor na 23 www.veterinariandocs.com.br fenda sináptica. As vesículas vazias, envolvidas por clatrina, são rapidamente recolhidas. *Clatrina: proteína que desempenha um importante papel no processo de formação de vesículas membranares no interior das células eucariontes. Músculo estriado esquelético: As células musculares são altamente especializadas na conversão de energia química em energia mecânica. O músculo esquelético é um músculo estriado que está sob controle voluntário. As estriações das células musculares esqueléticas resultam do arranjo altamente ordenado das moléculas de actina e miosina. A força é gerada pela interação de actina e miosina musculares, um processo que requer elevação transitória do Ca intracelular. Organização do músculo esquelético: Fibras musculares: cada músculo é composto de numerosas células chamadas fibras musculares. Endomísio: envolve cada uma dessas fibras As fibras musculares individuais estão agrupadas em Fascículos. Perimísio: envolve cada fascículo. *Dentro do perimísio estão os vasos sanguíneos e nervos. Finalmente, fascículos são agrupados para formara o músculo Epimísio: envolve o músculo. Cada fibra de músculo esquelético contém feixes de filamentos chamados Miofibrilas, correndo ao longo do eixo da célula. O padrão estriado das células resulta de um padrão repetitivo nas miofibrilas. A miofibrila pode ser subdividida longituninalmente em sarcômeros. O sarcômero é demarcado por duas linhas escuras, chamadas linhas Z, e é unidade contrátil no músculo. Em cada lado da linha Z existe uma linha clara (banda I), que contém filamentos finos, composto de actina. A área entre as duas bandas I, dentro do sarcômero, é a banda A, que contém proteínas Miosina. Uma área clara está presente no centro do sarcômero, e é chamada banda H. Os filamentos finos de actina se estendem da linha Z ao limite da banda H. Cada miofibrila na fibra muscular é envolvida por retículo sarcoplasmático (RS). Túbulos T: invaginações no sarcolema, estendem-se para o interior da fibra muscular até próximo dos limites da banda A. 24 www.veterinariandocs.com.br A porção do RS próxima aos túbulos T é chamada cisterna terminal, e é o local de liberação de Ca. *Os dímeros da proteína tropomiosina se estendem por todo o filamento de actina, cobrindo os sítios de ligações da miosina nas moléculas de actina. Um complexo de troponina constituído de três subunidades (troponina T, troponina I e troponina C). A tropomiosina tem possibilidade de inibir ligações da miosina ao filamento de actina. A troponina T liga-se à troponina, a troponina I facilita a inibição da ligação actina-miosina e a troponina C liga p Ca++. A ligação da troponina C com o Ca++ promove o movimento da tropomiosina sobre o filamento de actina, expondo os sítios de ligações de miosina, e, desse modo, ocorre a interação entre filamentos de actina e miosina e a contração do sarcômero. A miosina é uma proteína grande. A região da cabeça se estende do filamento grosso em direção ao filamento fino de actina, e é a porção de molécula que pode se ligar à actina. A atividade de ATPase da miosina reside na cabeça globular. Controle da atividade do músculo esquelético: Nervos motores e unidades motoras; Especificamente, cada músculo esquelético é invervado por um motoneurônio α. Os nervos motores se ramificam no músculo, cada ramos inervando uma única fibra muscular. Uma unidade motora consiste de um motoneurônio e todas as fibras musculares inervadas por ele. A junção neuromuscular formada por um motoneurônio α é chamada placa terminal. A acetilcolina liberada pelo motoneurônio na junção neuromuscular inicia um potencial de ação na fibra muscular, que rapidamente se propaga ao longo de seu comprimento. Acoplamento excitação-contração: Quando um potencial de ação é transmitido ao longo do sarcolema da fibra muscular e então para os túbulos T, Ca++ é liberado pelas cisternas terminais do RS para o sarcoplasma. Esta liberação de Ca++ do RS eleva a concentração intracelular de Ca++ que, por sua vez, promove a interação actina-miosina e contração. Este aumento no Ca++ inicia uma contração chamada ‘Abalo’. O mecanismo de elevação do Ca++ intracelular envolve uma interação entre proteínas no túbulo T e na cisterna terminal do RS adjacente. O túbulo T é uma invaginação do sarcolema, que se estende para dentro da fibra muscular. Há uma ligação entre os túbulos T e as cisternas terminais, chamados de pés. Como este se liga à droga rianodina, ele é chamado de Receptor Rianodina (RYR). 25 www.veterinariandocs.com.br Na membrana do túbulo T supõe-se que o RYR interaja com uma proteína chamada receptor para Diidropiridina (RDHP). O RDHP é um canal para Ca++ controlado por voltagem. Entretanto o influxo de Ca++ para dentro da célula, através do RDHP não é necessário para liberação do Ca do RS. Em vez disso, a liberação do Ca da cisterna terminal RS resulta da mudança conformacional do RDHP, por meio de uma interação proteína-proteína, abre o RYR liberando Ca++ no mioplasma. O relaxamento do músculo ocorre à medida que o Ca intracelular é seqüestrado de novo pelo RS. A captação do Ca pelo Rs é devida a ação da bomba de Ca ++ (Ca2+ ATPase). Interação actina-miosina: Formação de Ponte O processo de contração é regulado pelos filamentos finos. O Ca++ liberado do RS liga-se a troponina C. Uma vez ligada ao Ca++, a troponina C facilita o movimento da molécula associada de tropomiosina em direção à depressão de actina. Esse movimento da tropomiosina expõe o sítio de ligação da miosina no filamento de actina, permitindo a formação de uma ponte e desse modo a geração de tensão; Ciclo da formação de pontes – encurtamento do sarcômero Uma vez que a miosina e actina estejam ligadas, a mudança da conformação na molécula de miosina, dependente de ATP, provoca movimento dos filamentos de actina em direção ao centro do sarcômero. Isto reduz o comprimento do sarcômero e contrai a fibra muscular. No estado de repouso, a miosina hidrolizou parcialmente o ATP. Quando o Ca++ é liberado da cisterna terminal do RS, ele se liga a tropomiosina C, que por sua vez promove movimento de tropomiosina sobre o filamento de actina. Isso então permite que a cabeça ‘energizada’ da miosina se ligue à actina adjacente. A miosina sofre então uma mudança de conformação, denominada ‘ação de engrenagem’,que puxa o filamento de actina em direção ao centro do sarcômero. A miosina, então, liga um novo ATP e parte da energia é utilizada para recolocar a cabeça na posição inicial e retornar ao estado de repouso. Se o Ca++ intracelular estiver ainda elevado, a miosina produzirá um outro ciclo de formação. O ciclo continuará até a bomba SERCA transportar o Ca++ de volta para o RS. À medida que o nível de Ca++ cai, o íon se dissocia da tropomiosina C, o complexo troponina-tropomiosina se movimenta e bloqueia os sítios de ligação da miosina no filamento de actina. Rigor Mortis: músculo fica rígido. Tipos de músculos esqueléticos: Tipicamente, os músculos de contração lenta são recrutados antes das fibras de contração rápida, devido a maior excitabilidade dos motoneurônios que inervam os 26 www.veterinariandocs.com.br músculos de contração lenta. A alta capacidade oxidativa das fibras musculares de contração lenta suporta a atividade contrátil mantida. As fibras musculares de contração rápida, por outro lado, tendem a ser grandes e tipicamente têm baixa capacidade oxidativa e alta capacidade glicolítica. As unidades motoras de contração rápida estão assim mais bem adaptadas a curtos períodos de atividade quando altos níveis de força são exigidos. As fibras de contração rápida podem ser convertidas em fibras musculares de contração lenta (e vice-versa)m dependendo do padrão de estimulação. A estimulação elétrica crônica do músculo de contração rápida resulta na expressão da miosina de contração lenta e diminui a expressão da miosina de contração rápida, com o aumento da capacidade oxidativa. Essas características se correlacionam com o nº de mitocôndrias presentes na fibra. O mecanismo desta mudança na expressão gênica é desconhecido, mas pode ser secundário a uma elevação da concentração intracelular de Ca++ de repouso. Como as fibras rápidas dependem do metabolismo glicolítico elas entram em fadiga rapidamente. Conseqüentemente, elas são usadas apenas ocasionalmente e por breves períodos de tempo. E as fibras lentas satisfazem as demandas metabólicas com a fosforilação oxidativa. Conseqüentemente, esses músculos entram em fadiga mais lentamente, e por isso, são usados mais para atividades de sustentação (postura). SERCA 1 no músculo de contração rápida. SERCA 2 no músculo de contração lenta. A atividade da SERCA 1 é maior que da SERCA 2. Portanto a recaptação de Ca++ pelo RS ocorre mais rapidamente em músculos rápidos. Modulação da força de contração: Recrutamento: Um meio simples de aumentar a força de contração de um músculo é recrutar mais fibras musculares. As unidades motoras de contração lenta tendem a ser pequenas e são inervadas por um motoneurônio α, que é facilmente excitado. As unidades motoras de contração rápida, tendem a ser grandes e são inervadas por motoneurônios α que são mais difíceis de excitar. As unidades motoras lentas são recrutadas primeiro que as rápidas. A vantagem dessa estratégia de recrutamento é que as primeiras fibras musculares recrutadas são as que têm alta resistência à fadiga. Além disso, o pequeno tamanho das unidades motoras de contração lenta permite um controle fino em baixos níveis de força. Tetania: Um único potencial de ação libera Ca++ suficiente para causar um abalo muscular. Porém, a duração desta contração é muito curta porque o Ca++ é muito rapidamente bombeado e volta para dentro do RS. Se o músculo é estimulado uma segunda vez antes do completo relaxamento, a força de contração aumenta. A freqüência de estímulos necessária para produzir tetania depende do tipo de fibra, lenta ou rápida, da unidade motora. 27 www.veterinariandocs.com.br Tônus do músculo esquelético: Mesmo em repouso os músculos normalmente exibem algum nível de atividade contrátil. Esta firmeza ou tônus é causada por um baixo nível de atividade contrátil em algumas unidades motoras que se deve a arcos reflexos dependentes do fuso muscular. O tônus no músculo esquelético é distinto do ‘tônus’ do músculo liso. Fonte de energia durante a contração: ATP: o músculo tem pequenos estoque de ATP, capaz de suportar algumas contrações somente. Fosfato de Creatina: é utilizada para converter ADP em ATP e assim repor o estoque de ATP. As enzimas CPK catalisa a reação. A fadiga do m. esquelético durante o exercício intenso está associada à depleção do estoque de fosfato de creatina. Carboidratos: as células musculares contêm glicogênio que fornece glicose para a fosforilação oxidativa e glicólise. As células musculares podem obter glicose do sangue, através da insulina. Ácidos Graxos e Triacilgliceróis: importante fonte de energia durante períodos prolongados. Os A.G. são submetidos à β-oxidação dentro da mitocôndria. Fadiga: NÃO é o resultado da depleção dos estoques de energia. Produtos metabólicos parecem ser importantes fatores no inicio da fadiga. Durante exercícios intensos, o acúmulo de fosfato inorgânico e ácido lático no citoplasma causa fadiga muscular. O acúmulo de ácido lático reduz o pH citoplasmático e inibe a alteração actina-miosina. Este declínio no pH reduz a sensibilidade do Ca++ da interação actina-miosina por alteração da ligação do Ca++. O fosfato inorgânico tem sido também implicado como um importante fator no desenvolvimento da fadiga durante o exercício intenso. O músculo esquelético exibe considerável plasticidade fenotípica. O crescimento anormal está associado a hipertrofia celular causada pela adição de mais miofibrilas e mais sarcômeros nas extremidades da célula para igualar o crescimento do esqueleto. Treinamento de força induz a hipertrofia muscular, enquanto o treinamento de resistência aumenta a capacidade oxidativa de todas as unidades motoras envolvidas. Os regimes de treinamento não são capazes de alterar o tipo de fibra ou a expressão das isoformas de miosina. Músculo liso: Também deve ser capaz de realizar contrações econômicas (consumindo o mínimo de ATP). Tipos: 28 www.veterinariandocs.com.br -Unitário: as células são acopladas eletricamente, de modo que a estimulação elétrica de uma é seguida pelo estímulo das células musculares lisas adjacentes. Isto resulta numa onda de contração como a observada na peristalse. -Multiunitário: as células não são acopladas eletricamente, de modo que a estimulação de uma célula não resulta necessariamente na ativação da célula muscular lisa adjacente. -Músculo Liso Fásico: que exibe uma atividade rítmica ou intermitente -Músculo Liso Tônico: que está constantemente ativo. Contatos célula a célula: Existe uma variedade de contatos especializados entre as células do m. liso. Estes contatos permitem a ligação mecânica e a comunicação entre as células. Vários tipos de junções são encontrados no M. liso. A ligação funcional das células é proporcionada pelos canais juncionais (gap junctions). Os canais juncionais formam vias de baixa resistência entre as células. Junções aderentes (placas densas ou placas de fixação) fazem a ligação mecânica entre as células do m. liso. Cavéolas: representam invaginações na membrana do m. liso (análogas Túbulos T). O RS se estende por toda a célula do M. liso, apesar de haver regiões juncionais onde ele toca regiões do sarcolema e/ou cavéolas. Células e membranas: Núcleo único, localizado centralmente. As células do m.liso não possuem túbulos T, as invaginações do sarcolema do m. esquelético que fazem as conecções elétricas para o RS. Entretanto, o sarcolema do m. liso possui fileiras longitudinais de pequenas bolsas, tipo saco, denominadas Cavéolas, que aumentam a superfície-volume das células. De importância, os canais intracelulares para Ca++ no RS do m. liso incluem o Receptor de Rianodina RYR que é similar ao encontrado no RS do m. esquelético e o canal para Ca++ dependente do IP3. Aparelho contrátil: A ausência de estriações no m. liso não implica em ausência de ordem. Os filamentos grossos e finos são organizados em unidades contráteis que são análogas aos sarcômeros. Os filamentos finos do m. liso possuem composição de actina e tropomiosina e estrutura similar ao m. esquelético. 29 www.veterinariandocs.com.br O conteúdo de miosina do m. liso é somente um quarto do encontrado no m. estriado. Estes grupos de filamentos grossos, interdigitados com os filamentos finos, se conectam a corpúsculos ou áreas densas, e representam o equivalente ao sarcômero. Citoesqueleto: O citoesqueleto nas células do m. liso serve de ponto de fixação para os filamentos finos e permite a transmissão da força para a extremidade da célula. O aparatil contrátil do m. liso não é organizado em miofibrilas e não existem linhas Z. Os equivalentes funcionais das linhas Z nas células do m. liso são os corpúsculos densos e áreas densas que formam a banda ao longo do sarcolema. Controle da atividade do m. Liso: A atividade contrátil do m. liso pode ser controlada por inúmeros fatores, incluindo hormônios, nervos autonômicos, atividade de marcapassos e uma variedade de drogas. Os potenciais de ação no m. liso são altamente variáveis e nem sempre necessários para iniciar uma contração. Regulação da contração: Contração do m. liso ocorre em resposta a uma elevação de Ca++. No m. esquelético: a elevação de Ca++ afeta a actina (contração regulada pela actina) No m. liso: a elevação de Ca++ afeta a miosina (contração regulada pela miosina) Especificamente, um aumento na concentração intracelular de Ca++ ativa uma cinase protéica dependente de Ca++-calmodulina, que fosforila a cadeia regulatória da miosina, permitindo que ela interaja com a actina. O ciclo de formação das pontes de miosina no m. liso é similar ao m. estriado, no qual após a fixação ao filamento de actina a ponte sofre uma ação de alavanca tracionando o filamento fino em direção ao centro do filamento grosso, gerando força. O ADP e o Pi são liberados pela cabeça da miosina neste momento, permitindo que o ATP se ligue. O ciclo de formação das pontes continua enquanto a ponte de miosina permanecer fosforilada. A ciclagem de formação das pontes continua com a hidrólise de um ATP por ciclo, até que a concentração de Ca++ caia. Contração fásica x tônica: Durante uma contração fásica, a concentração de Ca++ citoplasmática, a fosforilação das pontes e a força atingem um pico de depois retornam a um valor basal. Durante uma contração tônica, a concentração de Ca++ citoplasmática e a fosforilação das pontes declinam após uma rápida elevação inicial, ma não retornam aos níveis basais. 30 www.veterinariandocs.com.br Quando a concentração de Ca++ citoplasmática cai durante contrações tônicas, a probabilidade de uma ponte ser desfosforilada aumenta. O músculo se relaxará se a concentração de Ca++ cair abaixo do necessário para a ligação à calmodulina e a ativação da cinase da cadeia leve da miosina. Energia e metabolismo: O músculo liso utiliza uma quantidade 300 vezes menor de ATP. Todas as necessidades metabólicas são prontamente atendidas pela fosforilação oxidativa. A fadiga do m. liso não ocorre, a menos que a célula seja privada de suprimento de O2. O RS também regula a concentração de Ca++ do citoplasma. Os canais para o Ca++ no retículo sarcoplasmático se abrem em resposta a sinais químicos em vez de sinais elétricos. Neurotransmissores ou hormônios que atuam através de receptores no sarcolema podem ativar a fosfolipase C, seguida da geração do 2º mensageiro. IP 3. O IP3 ativa o canal para o Ca++ dependente de IP3 no RS. O RS do m. liso também contém canais para Ca dependentes de Ca (RYR). O Ca++ é reacumulado no retículo sarcoplasmático pela SERCA. 31 www.veterinariandocs.com.br Referências Bibliográficas DUKES. Fisiologia dos Animais Domésticos. 12 a ed. Rio de Janeiro: Guanabara Koogan, 2006. BERNE, R.M.; LEVY, M.N. Fisiologia. 5 a ed. Rio de Janeiro: Elsevier, 2004. 32 www.veterinariandocs.com.br
Download