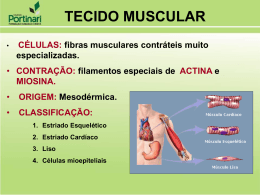
BIOELETRICIDADE Prof. Me Tatiane Quaresma Sendo os seres vivos máquinas elétricas, é natural que seus elementos produzam e usem eletricidade. As células vivas apresentam uma diferença de potencial entre os dois lados da membrana. O interior é sempre negativo, e o exterior, positivo. A origem desses potenciais é distribuição a assimétrica de especialmente de Na+, K+, Cl- e HPO4. = uma íons, O potencial principais: existe sob duas formas a) Potencial de Repouso, ou de estado fixo, mais ou menos em estado estacionário. b) Potencial de Ação, que é uma variação e programação brusca do potencial. ELETRÓDIOS IMPOLARIZÁVEIS Para recolher os potenciais e correntes, é necessário o uso de eletródios que não se polarizem. A polarização dos eletródios é o acúmulo de cargas opostas às que estão sendo medidas e que abaixam o potencial verdadeiro. Para registros superficiais, como do eletrocardiograma, os eletródios são untados com uma pasta eletrolítica, que além de impolarizar, melhora o contato elétrico. De um modo geral, os eletródios são utilizados em pares, sendo um ativo (percebe as diferenças de potencial), e o outro, de referência (sempre em potencial zero). INSTRUMENTAL Milivoltímetros e microamperímetros extensamente usados. são Para se obter um traçado contínuo das variações de voltagem ou corrente, usa-se o osciloscópio. O OSCILOSCÓPIO Consiste em um tubo de raios catódicos (feixe fino de elétrons), e de uma tela que fluoresce com o impacto dos elétrons. O feixe eletrônico tem dois movimentos simples: horizontal, chamado de varredura, e vertical. No eixo do x registram-se movimentos horizontais, no eixo de y, verticais. Para medidas biológicas, é conveniente deixar o feixe “varrendo” no eixo de x, gerando uma fina linha horizontal. Esse eixo vai medir dessa maneira, o Tempo do evento. No eixo de y (vertical), liga-se o potencial: se o potencial se eleva, o feixe sobe, se o potencial se abaixa, o feixe desce. Com a combinação desses movimentos, traçados incrivelmente complexos podem ser registrados com facilidade. Com o uso de grade calibrada, pode-se medir com precisão a magnitude e o tempo de cada evento. POTENCIAL DE REPOUSO, POTENCIAL TRANSMEMBRANA, DE REGIME ESTACIONÁRIO OU DE ESTADO FIXO. Esse potencial tem sua origem em um mecanismo simples, de alternância entre transporte ativo e transporte passivo de pequenos íons. 1ª Fase Os íons Na+ entram passivamente na célula, através do gradiente de concentração. 2ª Fase A célula expulsa esses íons ativamente, ao mesmo tempo que introduz, também ativamente, um íon K+. 3ª Fase Esse íon K+ tem grande mobilidade, e volta passivamente, para o lado externo da membrana, conferindo-lhe carga positiva. Do lado interno, íons fosfato e especialmente proteínas aniônicas fornecem a carga negativa. POTENCIAL DE AÇÃO (PA) REPOLARIZAÇÃO Logo em seguida, fecham-se os canais de Na+, e o íon K+ sai da célula, repolarizando-a. A bomba de sódio se encarrega de expulsar o pequeno excesso de íons Na+ que estava no interior da célula, e tudo volta ao estado inicial. A ELETRICIDADE ANIMAL Contribuição de Galvani e de Volta. A geração de eletricidade por certos peixes já era conhecida quando LUIGI GALVANI descreveu sua célebre observação sobre a contração da pata de rã. Galvani ensinava anatomia em Bolonha (Itália) e conta que certo dia, quando trabalhava com rãs decapitadas e penduradas numa haste de cobre observou que, quando a pata do animal tocava o ferro de um balcão próximo, os músculos se contraíam. Em uma outra experiência, utilizou choque elétrico sobre uma das rãs, e observou a contração muscular. • • • Galvani constatou que com ou sem aplicação de choque elétrico, era possível obter a contração dos músculos das patas posteriores da rã. Dos seus experimentos, concluiu: “o músculo e o nervo constituem uma espécie de condensador de uma própria e peculiar eletricidade que existe em todos os animais vivos”. Galvani acreditava que “ nos músculos se reúne o fluido elétrico, que logo se difunde pelo corpo mediante a rede de nervos, os quais são condutores naturais do fluido elétrico e que se insinuam com suas extremidades dentro dos músculos”. Na época de Galvani, ALEJANDRO VOLTA ensinava física na Universidade de Pavia. Volta, estudando o fenômeno descrito por Galvani, concluiu que os metais podiam produzir eletricidade e, em 1800, construiu o primeiro gerador químico de eletricidade empilhando alternadamente discos de cobre e zinco. Concluiu dizendo que os músculos e o nervos são apenas condutores de eletricidade. Não satisfeito, Galvani relatou os resultados dos novos experimentos nos quais conseguiu obter a contração muscular na pata da rã quando postas em contato com o nervo ciático de outra rã. Hoje se sabe que ambos estavam certos. Em 1887 foi registrado o fenômeno elétrico no coração. SINAIS ELÉTRICOS DO SISTEMA NERVOSO A membrana do neurônio apresenta uma propriedade muito particular que a distingue da maioria das células do organismo. Essa propriedade – excitabilidade - permite que o neurônio produza, conduza e transmita a outros neurônios os sinais elétricos em códigos que constituem a linguagem do sistema nervoso. Canais Iônicos Canais iônicos membrana que passar íons de ou em resposta ou mecânicos. são proteínas integrais de tem a capacidade de deixar modo seletivo, continuamente a estímulos elétricos, químicos Os canais que deixam passar os íons continuamente são chamados canais abertos, e os que só abrem em respostas a estímulos específicos são chamados de canais controlados por comportas. Tanto um como outro pode ser altamente específico: há canais para cátions, como o sódio (Na+), o potássio (K+) e o Cálcio (Ca++), e canais para ânions, como o cloreto (Cl-). Os canais controlados por comportas podem ser abertos por alterações de voltagem que existe naturalmente na membrana entre o interior e o exterior da célula nervosa. Neste caso, são considerados dependentes de voltagem. Outros podem ser abertos por substâncias especificas (ligantes) como neurotransmissores, neuropepitídeos e hormônios, e neste caso são considerados dependentes de ligantes. Finalmente, outros canais são abertos por certos tipos de energia mecânica e radiante que incidem POTENCIAL DE AÇÃO Todo potencial de ação é iniciado com um estimulo de despolarização. Esse estímulo precisa atingir o limiar de excitação do neurônio, quando começará a cadeia de eventos responsáveis pelo potencial. Com os canais de sódio e com o aumento da permeabilidade da membrana ao sódio, seu papel no potencial de repouso irá mudar. Inicia-se, então um grande fluxo de sódio para dentro da célula, causando sua despolarização. Depois que a célula foi despolarizada, os canais de sódio voltagem-dependentes se fecham, dando inicio à fase de repolarização da célula, que começa com a abertura dos canais de potássio. Com o aumento da permeabilidade ao potássio, uma grande quantidade desses íons flui para dentro da célula. Durante esse influxo de potássio, o neurônio não pode se despolarizar de novo, estando em período refratário. Como os canais de potássio permanecem abertos por um período de tempo maior que o necessário para repolarizar a membrana até seu nível de repouso, acaba ocorrendo uma hiperpolarização. Ela fica mais negativa que o normal. Com o tempo, devido a permeabilidade natural ao potássio e a bomba de sódio potássio, a membrana voltará ao potencial de repouso, mas, até lá, ela será mais dificilmente excitada, estando em período refratário relativo. O trabalho desta bomba de sódio e potássio tem um gasto de energia como “ Transporte Ativo “, senão houver o excesso destes elementos no extracelular como no intracelular ou como a combinação de sódio e potássio nas extremidades da bomba com a proporção 3x2 a mesma não ativa. Este processo continua até acabar o excesso (combinação) dos elementos no intra -como no extracelular. Através das vesículas (cápsulas), elas transportam os neurotransmissores que são chamados “ACETILCOLINA ”para os “AXÔNIOS ”. Estes neurotransmissores são fabricados pelos neurônios. Os cálcios que transportam as vesículas até as membranas pré - sináptica. Os cálcios foram absorvidos pelos canais de proteína, e são conhecidos de “ CARREADOR ”. As vesículas ao encostar-se com a membrana pré sináptica se rompem liberando os neurotransmissores na fenda sináptica até chegar na membrana pós - sináptica, está membrana tem canais de encaixe chamado “proteína de canal acetilcolina dependente “, só os neurotransmissores acetilcolina encaixa neste canal de proteína dependente, que na realidade este processo abre o canal para a entrada de sódio, começando o processo idêntico ao do início. DOS NEURÔNIOS ATÉ OS MÚSCULOS DOS NEURÔNIOS ATÉ OS MÚSCULOS Através do axônios onde os canais de proteínas absorvem o cálcio que transporta as vesículas (cápsulas) onde se encontram os neurotransmissores que são levados para a membrana pré sináptica, no contato com a membrana a vesícula se abre liberando os neurotransmissores na fenda sináptica. Os canais das fibras musculares ( proteínas do canal acetilcolina dependente) recebem os neurotransmissores e com isso libera a entrada de sódio nos canais de proteínas, que estimula a membrana da fibra muscular e os túbulos T que por consequência estimula o “retículo sacro plasmático “ que libera para as miofibrilas íons cálcio. Este processo não gasta energia , pois vai do mais concentrado para o menos concentrado. O cálcio aciona a “troponina“ que se encontra no sarcômero, que modifica a “tripomiosina ”que nada mais é uma camada entre a cabeça da miosina e o sitio ativo . Quando a tripomiosina recebe o sinal da troponina, a tripomiosina desloca o filamento fazendo com que a cabeça da miosina entre o contato com o sitio novo. Entre o sitio ativo e a actina contém uma molécula chamada ATP (trifosfato de adenosina) fazendo com que ela flexione agrupando camada por camada entre a cabeça da miosina . Sendo usado 1 fósforo para cada execução desse trabalho, a molécula se torna ADP ( difosfato de adenosina ). Com isso a actina e miosina contrai os sarcômeros, que contrai as miofibrilas, que contrai as fibras musculares e os músculos. Com todo este processo a fibra muscular está contraída, para relaxar é feito um corte no estimulo para o cérebro que corta o P.A que vai dos neurônios até o músculo, fazendo com que a bomba de cálcio do Reticulo Sacro Plasmático bombeie este cálcio que se encontra nas miofibrilas para o retículo sacro plasmático. Quando o processo P.A acontece o núcleo das células produzem neurotransmissores que são levados pelas vesículas. Existem 3 movimentos: Movimentos reflexo: Auto-proteção (P.A não alcança o cérebro). Movimentos automatizados: Movimentos repetitivos, aprendizado (cortex - pouco abaixo do topo do cérebro) Movimentos voluntários: Cérebro que comanda. A função dos túbulos T é enviar os estímulos em todas as direções e estimulá-las ( retículos sacro plasmáticos). O uso das fibras musculares geralmente não é 100% ou todo seu potencial, só é usado a quantidade de fibras necessárias para realizar aquela tarefa exigida. Então quando realizarmos uma tarefa de força ou destreza (colocar uma linha na agulha) que precisamos de precisão, só utilizamos o que precisamos. Sinapse também é conhecida como “PLACA MOTORA”. Glicogênio: Armazenamento de glicose. A dor é uma defesa para o corpo que está acontecendo algo. CONTRAÇÃO DO MÚSCULO ESQUELÉTICO Cerca de 40% do corpo são formados por músculos esqueléticos e quase outros 10% são formados por músculos liso e cardíacos. SARCOLEMA: é a membrana celular da fibra muscular. É formado por uma verdadeira membrana celular, chamada de membrana plasmática. Na extremidade da fibra muscular, as fibras tendinosas se unem, formando feixes, até comporem um tendão muscular que se insere no osso. MIOFIBRILAS: Cada miofibrila, por sua vez, contém, lado a lado cerca de 1.500 filamentos de miosina e 3.000 filamentos de actina. As faixas claras só contém filamentos de actina, e as faixas escuras contêm os filamentos de miosina. E também mostra que as extremidades de actina estão presos ao chamado disco Z. A partir desse disco, os filamentos se estendem, nas duas direções, para se interdigitar com os filamento de miosina. Esses faixas dão ao músculo esquelético e cardíaco sua aparência “Estriada”. A região de uma miofibrila ( ou de toda uma fibra muscular ) situada entre duas linhas Z consecutivas é chamada de sarcômero. Os filamento de actina se sobrepõem totalmente aos outros. SARCOPLASMA: as miofibrilas, no interior da fibra muscular, ficam suspensas em uma matriz, chamada de sarcoplasma, formada pela pelos constituintes usuais. O líquido do sarcoplasma contém grandes quantidade de potássio e de magnésio, de fosfato e de enzimas protéicas. Também está presente número imenso de mitocôndrias que ficam entre e paralelas ãs miofibrilas, situação indicativa da grande necessidade das miofibrilas em contração de quantidade elevada de trifosfato de adenosina (ATP), formado nas mitocôndrias. RETÍCULO SARCOPLASMÁTICO: Também existe no sarcoplasma um imenso retículo endoplasmático, chamado na fibra muscular de “Retículo Sarcoplasmático”. Esse retículo é muito importante para o controle da contração muscular. O MECANISMO GERAL DA CONTRAÇÃO MUSCULAR Um potencial de ação percorre um axônio motor até sua terminações nas fibras musculares. 2. Em cada terminação, há secreção de pequena quantidade de substância neurotransmissoras, chamada Acetilcolina. 3. A acetilcolina atua sobre área localizada da membrana fibro muscular, abrindo númerosos canais protéicos acetilcolina dependentes. 4. A abertura destes canais Acetilcolina-Dependentes permite o influxo de grande quantidade de íons sódio para o interior da membrana da fibra muscular, no ponto da terminação nervosa. Isso produz um potencial de ação na fibra muscular. 5. O potencial de ação se propaga ao longo da membrana da fibra muscular do mesmo modo como o faz nas membranas neurais. 1. 6. O potencial de ação despolariza a membrana da fibra muscular e também penetra profundamente no interior desta fibra, nos túbulos T, que se propagam rapidamente. Ai faz com que o retículo sarcoplasmático libere, para as miofibrilas, grande quantidades de íons Cálcio, que ficam armazenada no seu interior. 7. Os íons cálcio geram forças atrativas entres os filamentos de actina e miosina, fazendo com que deslizem um em direção do outro, o que constitui o processo contrátil. 8. Após o corte de estímulo, é cortado automaticamente o potencial de ação até os músculos por uma fração de segundos, com isso os íons cálcio são bombeados de volta para o retículo sarcoplasmático através da bomba de Cálcio,(aonde existe um transporte ativo, com gasto de energia) até que ocorra um novo potencial de ação muscular; termina a contração muscular. RELAÇÃO ENTRE A VELOCIDADE DE CONTRAÇÃO E A CARGA Um músculo se contrai de uma forma extremamente rápida quando sua contração não sofre oposição de qualquer carga - Quando são aplicadas cargas, a velocidade de contração diminui progressivamente á medida que a carga for aumentando. Quando a carga for aumentada até igualar a força máxima que pode ser gerada pelo músculo, a velocidade de contração é zero, e não ocorre contração. Essa velocidade decrescente da função do aumento da carga é causa pelo fato de que a carga imposta a um músculo em contração é uma força inversa que se opõe a força contratil gerada pela contração do músculo. FIBRAS MUSCULARES RÁPIDAS E LENTAS FIBRAS RÁPIDAS: 1 Fibras muito maiores para uma força maior de contração; 2 Retículo sarcoplasmático extenso, para a liberação rápida de íons cálcio , para desencadear a contração; 3 Grande quantidades de enzimas glicolíticas para a liberação rápida de energia pelo processo glicolítico; 4 Vascularização pouca extensa, pela importância secundária do metabolismo oxidativo 5 Pequeno número de mitocôndrias, igualmente por ser metabolismo oxidativo. FIBRAS LENTAS: 1 Fibras menores; 2 Inervado por fibras nervosas mais finas; 3 Vascularização bem mais extensa, com muitos capilares para fornecimento de quantidades adicionais de oxigênio; 4 Número muito grande de mitocôndrias, permitindo a manutenção de alto nível de metabolismo oxidativo; 5 As fibras contém grande quantidade de mioglobina, proteína contendo ferro, semelhante a hemoglobina das hemácias, a mioglobina acelera de muito o transporte de oxigênio para as mitocôndrias. A mioglobina da aos músculos lentos uma coloração avermelhada, chamados de músculos vermelhos, e os músculos rápidos são chamados de músculos brancos. A UNIDADE MOTORA: Cada motoneurônio que emerge da medula espinhal inerva numerosas fibras musculares: dependendo do tipo de músculos. Todas as fibras musculares inervadas por uma só fibra nervosa motora formam a chamada unidade motora. Os músculos pequenos que reagem rapidamente, cujo controle deve ser bastante preciso, têm unidades motoras com poucas fibras musculares (até apenas duas a três fibras nos músculos laríngeos). Os músculos grandes, que não precisam de um controle muito exato, como por exemplo, o músculo gastrocnêmio, podem Ter unidades motoras com várias centenas de fibras musculares. TÔNUS DO MÚSCULO ESQUELÉTICO: Mesmo quando os músculos estão em repouso, ainda persiste um certo grau de tensão. Isso é chamado de tônus muscular. Impulsos transmitidos do encéfalo para os motoneurônios anteriores correspondentes, em parte, por impulsos que se originam dos fusos musculares localizado nos próprios músculos. FADIGA MUSCULAR: A interrupção do fluxo sanguíneo para um músculo em contração produz fadiga muscular quase total em um minuto ou pouco mais, devido à perda do fornecimento de nutrientes em especial o oxigênio. ATROFIA e HIPERTROFIA MUSCULAR: Deve ser lembrado que um músculo estirado se contrai com mais força que um músculo retraído. Quando um músculo permanece inativo por longos períodos, a velocidade de degradação das proteínas contrateis, bem como a redução de miofibrilas, é maior que a velocidade de respostas. Como resultado ocorre a atrofia muscular. AJUSTE DO COMPRIMENTO MUSCULAR Ocorre um tipo de hipertrofia quando os músculos são estriados além de seu comprimento normal. Isso faz com que sejam adicionados novos sarcômeros nas extremidades das fibras musculares onde elas se fixam as tendões. Inversamente, quando um músculo permanece retraído a comprimento menor que o seu normal por longos períodos, os sarcômeros nas extremidades das fibras desaparecem de modo igualmente rápido. É por esses processos que os músculos são continuamente remodelados para terem o comprimento adequado para uma contração muscular apropriada. A HIPERTROFIA é resultado das fibras musculares isoladas, muito mais acentuada quando o músculo é estirado durante o processo contrátil. CONTRATURA: Ao tentar usar uma musculatura sem aquecimento e tentar fazer um movimento maior que o músculo pode realizar, o mesmo senti que irá romper fibras, e realiza uma contração nesta musculatura indisponibilizando o grupo muscular. CAIMBRA: É quando falta energia para alimentar a bomba de cálcio, tendo esta energia gasto no trabalho excessivo do corpo, MIASTEMIA GRAVIS: Fica impossibilitado de transmitir sinal pela “placa motora “(sinapse) a pessoa possui auto-imunológico, que na realidade produz uma auto-defesa; anticorpos destroem os canais de proteínas. Existem uma enzima acetilcolina esterase que destrói os neurotransmissores que ficam na fenda sinapse, após o corte de P.A no neurônio. E como os canais estão destruídos tendo pouca contração muscular, podendo morrer de parada respiratória pela pouca contração do diafragma. RIGOR MORTIS: É a contração do músculo, fica rígido sem P.A após a morte. Essa contração é causada pela perda total de ATP, que é necessário para as separações das pontes dos filamentos de actina e miosina. Os músculos permanecem neste estado até que as proteínas sejam destruídas, isto pode levar de 15 a 20 horas após a morte. Por hoje é só!
Download