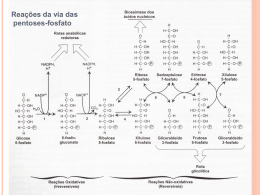
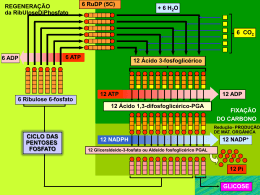
A trajetória comum de um cientista comum Walter Colli Departamento de Bioquímica, Instituto de Química, Universidade de São Paulo Pediram-me para inaugurar essa nova seção do Congresso Anual da Sociedade de Bioquímica e Biologia Molecular (SBBq) que consiste - como assim entendi - em que um cientista sênior, cuja carreira já está consolidada, conte um pouco de sua vida a fim de servir como referência (positiva ou negativa) aos jovens que estão iniciando suas atividades científicas. Não sei porque fui escolhido para ser o primeiro já que há tantos cientistas mais importantes em nossa Sociedade e que poderiam melhor desempenhar essa tarefa. Talvez tenha sido uma homenagem ao fato de eu ter sido Vice-Presidente (1978/1979) e Presidente da Sociedade (1979/1980), bem como o Presidente da Comissão Organizadora do VI Congresso PAABS que ocorreu em São Paulo de 18 a 22 de fevereiro de 1990. A PAABS era a Panamerican Association of Biochemical Societies que mudou de nome para PABMB (Panamerican Association of Biochemistry and Molecular Biology) durante a vigência de meu mandato de Secretário Geral da entidade (1990-1993). Sempre gostei de Genética, mas por problemas familiares prestei vestibular na Faculdade de Medicina da USP. Lá encontrei o Professor Isaias Raw que me convidou a trabalhar com ele logo no primeiro ano. Não era Genética, mas era Bioquímica. Como estudante tive a sorte de cristalizar o citocromo b 5 que era a décima proteína a ser cristalizada e por isso a Nature aceitou a comunicação. Foi meu primeiro trabalho. Formei-me em 1962 e o Professor me convidou para ser Instrutor (Auxiliar de Ensino). Naquele tempo não havia concurso e o Professor era o Senhor de baraço e cutelo. Se ele fosse bom, o Departamento seria bom. Se ele fosse medíocre estabelecia-se uma cadeia de mediocridades. Até agora é assim. Nos concursos viciados contratam-se nulidades que permanecem por 30 anos ou mais, já que as regras aplicadas ao professor universitário são as regras do funcionalismo público. É-se efetivado no serviço público e estabilizado no emprego. Esse é o segredo de polichinelo dos bons Departamentos: contratar os melhores e não os amigos ou os futuros assistentes dos professores mais velhos. No ano de 1963 recebemos para um ano sabático a visita de Maynard Pullman, o descobridor da F1-ATPase. A ligação comum era Efraim Racker na área de fosforilação oxidativa. Com ele e Isaias iniciei meu trabalho de doutoramento (naquele tempo não havia mestrado) estudando síntese proteica em mitocôndria. A ideia era usar sínteses mitocondriais a fim de demonstrar que havia um doador de energia anterior a um composto fosforilado que seria a ligação entre duas proteínas por uma "high energy bond", o composto X~Y. Aquele era o tempo em que havia duas escolas: a de Racker, Green, Lehninger que acreditavam haver um intermediário químico que transferisse energia para ADP e Pi a fim de sintetizar ATP e Mitchell que tinha uma teoria pouco compreendida na época. Nosso trabalho, além de demonstrar que a mitocondria fazia proteínas mostrou também que excesso de oligomicina, que supostamente bloquearia toda a síntese de ATP, permitia ainda um pequeno vazamento suficiente em 5:1 para incorporar leucina radiativa em proteínas mitocondriais. Esse trabalho seria pioneiro se tivesse sido publicado, mas não foi porque Maynard estava inseguro quanto à presença de contaminação bacteriana, que não havia. No entanto, a tese foi mencionada num Annual Review of Biochemistry em artigo de Gottfried Schatz como mais uma demonstração de que mesmo em condições de quase absoluto bloqueio de síntese de ATP havia energia suficiente para conduzir processos biossíntéticos mitocrondriais. Em 1966 defendi meu doutoramento e fui para Nova Iorque fazer pósdoutorado no laboratório de Maynard Pullman para estudar síntese de ácidos graxos em mitocôndria com a mesma finalidade, procurar um intermediário não fosforilado que fosse o doador de energia. É evidente que não o iríamos encontrar, por inexistente, sabemos agora. No entanto, esse estágio rendeu-me duas publicações no JBC que, na época, tinha um índice de impacto (ninguém sabia o que era isso) muito maior do que hoje. Os grandes laboratórios da época só publicavam no JBC. Durante o estágio conheci Michio Oishi, um cientista japonês contratado na mesma instituição logo após seu pós-doutorado. Todos os dias, durante o lanche, discutíamos como isolar e analisar os genes de RNA ribossômico em Bacillus subtilis. Solicitei prorrogação de minha licença à USP e fui para o laboratório dele que ficava um andar abaixo do laboratório do Pullman. Lá isolamos os genes responsáveis pela síntese de rRNA 16S e 23S (5S) e estabelecemos suas relações topológicas no cromossomo. Finalmente eu havia chegado à Genética. Publicamos um trabalho no PNAS e dois no Journal of Molecular Biology. Um deles foi reimpresso em livro editado por Zubay e Marmur. É preciso informar aos jovens que era o ano de 1968 e a aplicação das enzimas de restrição para isolamento de fragmentos e clonagem só apareceu na metade da década de 70. Voltei no Natal de 1969, após 3 anos e 7 meses de experiência pós-doutoral, e procurei meu laboratório no Instituto de Química da USP, pois, o Departamento de Bioquímica havia sido incorporado, como todos os outros, a esse Instituto na Cidade Universitária. No início tive dúvidas de qual linha adotar. Com Bianca Zingales continuei a explorar a região de rDNA de B. subtilis. Seriam os genes (10 ao todo) co-transcritos ou transcritos independentemente? Com Anita Marzzoco fizemos incursão no genoma de Acanthamoeba castellanii. Com Maria Júlia Manso Alves iniciamos um projeto de estudo da membrana do Trypanosoma cruzi. A última linha prevaleceu e estamos nela desde então. Essa história recente eu procurarei resumir na palestra.
Download