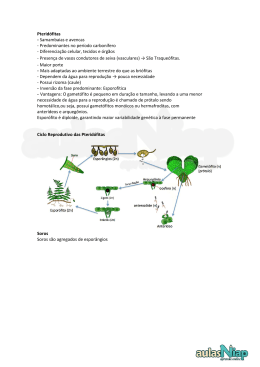
UNIVERSIDADE ESTADUAL DO CEARÁ CENTRO DE CIÊNCIAS DA SAÚDE – CCS CURSO DE CIÊNCIOAS BIOLÓGICAS Bruno Edson Chaves Pteridófitas : as primeiras plantas vasculares Fortaleza-CE Julho, 2006 UNIVERSIDADE ESTADUAL DO CEARÁ - UECE 2 CENTRO DE CIÊNCIAS DA SAÚDE – CCS CURSO DE CIÊNCIOAS BIOLÓGICAS Bruno Edson Chaves Pteridófitas : as primeiras plantas vasculares Trabalho apresentado como requisito parcial da Universidade Estadual do CearáUECE de aprovação na disciplina de Botânica I, ministrada pela professora Jeanne Barros Leal de Pontes Medeiros, no curso de Ciências Biológicas Fortaleza, Ceará SUMÁRIO 3 Introdução......................................................................................................... pág 5 Plantas vasculares sem sementes – As Pteridófitas........................................ pág 6 1- ocorrência e distribuição ........................................................................ pág 6 2- origem do esporófito ............................................................................... pág 9 3- evolução ............................................................................................... pág 13 4- características gerias do grupo ............................................................ pág 16 5- classificação ......................................................................................... pág 18 6- particularidades dos principais filos ...................................................... pág 20 Filo Rhyniophyta ................................................................................. pág 20 Filo Zosterophyllophyta ....................................................................... pág 21 Filo Lycophyta ..................................................................................... pág 22 Filo Trimerophyta ................................................................................ pág 25 Filo Psilophyta...................................................................................... pág 25 Filo Sphenophyta ................................................................................ pág 28 Filo Pterophyta .................................................................................... pág 30 7- Importância econômica ........................................................................ pág 35 8- Importância ecológica .......................................................................... pág 36 Conclusão ...................................................................................................... pág 37 Bibliografia ..................................................................................................... pág 38 4 Introdução Na divisão do reino Plantae entende-se por Pteridophyta, organismos vegetais cuja já existe presença de vasos condutores – do tipo xilema e floema-, para a condução de seiva bruta e elaborada respectivamente, portanto pertencem ao grupo das traqueófitas, sendo as pteridófitas os primeiros organismos a adquirem vasos. O aparecimento dos vasos condutores possibilitou uma maior diversidade de formas, permitindo-as atingir maiores dimensões do que qualquer outra planta terrestre existente até então, transformando-as nas primeiras plantas a abandonar por completo o meio aquático e conquistarem o ambiente terrestre. Estas plantas são encontradas nos mais variados habitats, e como características a maioria possui caule, raízes e folhas porém não há a formação de sementes, muito menos de flores, o que os colocam no grupo das Criptógamas. Caracterizado pela marcada alternância de fases no seu ciclo de vida, sendo a fase duradoura a esporofítica e gametofítica efêmera. As pteridófitas, assim como as demais plantas vasculares, tem uma grande importância e representatividade na flora. Constituem um grupo taxonômico informal, no qual estão reunidos sete filos, são eles: Rhyniophyta, Zosterophyllophyta, Trimerophytophyta, Lycophyta, Psilotophyta, Sphenophyta e Pterophyta. Porém destes sete filos somente os quatro últimos são encontrados atualmente, os outros três estão extintos. Em comum estas plantas tem somente as características gerais do grupo supracitados, a ausência de sementes e tecidos vasculares que transportam água e sais minerais para as plantas. Cada uma tem a sus particularidade e sua forma de vida. No estudo das pteridófitas procura-se entender o seu processo evolutivo das plantas, pois essas, com dito anteriormente, foram de fundamental importância para a ocupação progressiva do ambiente terrestre e o aumento da independência do meio aquático ou seja, pontos chaves para vida vegetal na terra. 5 Plantas vasculares sem sementes – As Pteridófitas 1- Ocorrência e distribuição As pteridófitas são um grupo de plantas bastante heterogêneo, e são encontradas nos mais variados habitats, que vão desde de ambientes de altas latitudes e altitudes ao interior de florestas pluviais tropicais e de vegetação arbustiva subdesértica a rochas costeiras e mangues; embora sejam normalmente achados nas regiões de clima tropical. Gostam de ambientes úmidos e sombrios, em ambientes de florestas vivendo como plantas epífitas ou hemiepífita, podendo ainda ser vistas sobre rochas, ou no solo. Como exemplo de plantas epífitas podem ser citadas as samambaias e as avencas, uma informação interessante refere-se ao que foi descrito na literatura, no qual cerca de 1/3 de todas as espécies de samambaias tropicais cresce sobre troncos e ramos de árvores como epífitas, sendo o dendezeiro uma das hospedeiras preferidas dessas pteridófitas. Apenas um pequeno número de espécies é capaz de crescer fora de um ambiente de floresta úmida, e sabendo que a maioria desse tipo de florestas está na região tropical pode-se deduzir que a maior diversidade de espécies nos trópicos esteja relacionada a condições relativamente estáveis que existem em muitos ecossistemas tropicais, nos quais as espécies tornam-se ajustadas a nichos ecológicos mais estreitos. Porém tais plantas vasculares ainda podem ser encontradas até em desertos, ou em ambientes aquáticos, no que se refere a este último ambiente os poucos representantes são duIcícolas, porém, nenhum marinho. Pelo fato de apresentarem vasos condutores podem apresentar grande diversidade de formas; podendo variar bastante de tamanho desde pequenas pteridófitas aquáticas como a Salvinia, até espécies arborescentes como o samambaiaçu, Cyathea com mais de 5m. As pteridófitas possuem cerca de 9.000 a 12.000 espécies, das quais cerca de 3.250 ocorrem nas Américas. Alguns autores estimam que aproximadamente 35% das espécies existentes na América ocorrem no Brasil, o que dá mais ou menos um número de 1.150 espécies de pteridófitas existentes no Brasil. 6 A região sudeste-sul do Brasil possui em torno de 600 espécies e abriga um dos centros de endemismo e especiação de pteridófitas no Continente Sul-Americano. O que se deve ao fato de parte dessa região apresentar a combinação de clima tropical úmido, montanhas e ecossistemas florestais. De acordo com alguns autores, as montanhas promovem alta riqueza de espécies de pteridófitas, os mesmo ainda afirmam que tal fato ocorre devido à variedade de ambientes criados por diferentes tipos de solos, rochas, elevações, inclinações, exposições à luz e microclimas das regiões montanhosas. Na zona Norte, os estudos realizados na região amazônica verificaram a ocorrência de quase 400 espécies, e é bem provável que possa existir bem mais do que esse número. Na região nordeste do Brasil, de modo geral, as pteridófitas tem uma distribuição relativamente ampla: no Maranhão ocorrem em áreas remanescentes de Floresta Perenifólia Higrófila Costeira citando 15 espécies; no Piauí há a citação de quatro espécies; destacam-se as ocorrências das pteridófitas nesta região, nas diversas zonas fitogeográficas que abrangem desde a Serra do Baturité, no estado do Ceará, até a Chapada Diamantina, no estado da Bahia, além daquelas ocorrentes em áreas de caatinga. Existem ainda estudos sendo realizados sobre a pteridoflora de Pernambuco que constataram a presença de pteridófitas na zona das Caatingas, além de sua presença nos remanescentes de mata atlântica. Em geral, quando falamos exclusivamente de samambaias, (as pteridófitas mais popularmente conhecidas) é certo que sua maior diversidade encontra-se nos trópicos onde cerca de 75% das espécies são encontradas. Em números podemos dizer que nas regiões temperadas, como nos Estados Unidos e Canadá, somente cerca de 380 espécies de samambaias foram descritas, enquanto cerca de 1.000 ocorrem no pequeno país tropical de Costa Rica, na América Central. A partir do dado referente às samambaias e de algumas informações supramencionadas pode tentar fazer uma relação com os outros tipos de pteridófitas na qual 80% das espécies de pteridófitas ocorrem em áreas tropicais, sendo mais comuns em montanhas tropicais e subtropicais úmidas. Esta forte correlação entre pteridófitas e florestas, deve-se ao fato de que as primeiras, no geral, são grandemente dependentes de outras plantas para lhes prover condições de proteção e sobrevivência. Em 7 condições de sombreamento na floresta, uma considerável proporção de esporos de pteridófitas deve ser capaz de germinar. Foi demonstrado, ainda, que em algumas florestas neotropicais as pteridófitas representam cerca de 10% do total do número de espécies de plantas vasculares apresentando-se em maior número de espécies do que qualquer família de angiospermas herbáceas. 8 2 – Origem do esporófito No decorrer do curso da evolução as plantas tiveram a tendência de estabelecer um esporófito livre e independente, quanto à nutrição, e tal esporófito tornase mais evidente pela primeira vez nas Pteridófitas. Essa evolução desta fase do desenvolvimento da planta foi de fundamental importância para a conquista do ambiente terrestre uma vez que possibilitou maior resistência e possibilitou um maior crescimento vegetal. Vários estudiosos de morfologia vegetal tentaram explicar a natureza do caule das primitivas Pteridófitas. Sabe-se que as os pteridófitos já existiam no Siruliano, ou seja, a mais de 380 milhões de anos e como o passar do tempo às plantas foram evoluindo, desde simples Rhyniophytos (que tinha como característica um caule dicotomicamente ramificado) aos Pterphytas (que tinha rizóides, folhas, e raízes). Foram emitidas, então, três teorias gerais sobre a natureza dos primeiros esporófitos, são elas: Que ele consistia nas folhas e o eixo folhoso surgiu mais tarde no decurso da evolução do caule; Que os apêndices do eixo evoluíram de áreas superficiais do eixo; E que o esporófito é inteiramente de natureza axial e que as folhas e ramos resultam de uma modificação de ramos. De acordo com estudo aprofundado de fósseis de alguns Psilófitos, tem-se conhecimento de que os primeiros pteridófitos eram esporófitos independentes, sem raízes nem folhas, constituídas por um eixo de ramificação dicotômica. Como será mostrado mais à frente a Rhynia citado acima, pode ter sido (de acordo com estudos realizados), progenitor dos Psilófitos pois ambas tem ramificações dicotômicas. E foi tomando por base essas comparações evolutivas da natureza caulinar das Pteridófitas mais primitivas que chegou a conclusão de que seria impossível as folhas terem precedido o eixo. Vários estudiosos, então lançaram teorias tomando por base a idéia da natureza axial do caule seguida por sua evolução até as ramificações simpódicas na constituição do eixo. Zimmermann, então reuniu as idéias baseadas nesses estudos e lançou a teoria do teloma – Figura 1a. No qual afirmava que o eixo caulinar primitivo era ereto, ramificado dicotomicamente, sem folhas e de estrutura semelhante aos de 9 Figura 1 – a)esquema mostrando a teoria do teloma; b) formação dos megáfilos; c) formação dos microfilos; d) derivção de uma ramificação dicitômica a simpódica 10 Rhynia. Os telomas os mesomas formam o sistema telómico, onde os telomas férteis originam os esporângios. As folhas foram agrupadas em dois grupos: as macrófilas - Figura 1b - e as microfilas – figura 1c. Estas últimas só apresentavam uma única nervura desde a base até o ápice. As folhas microfilas são sempre dotadas de um limbo com um sistema de nervuras muito ramificado, em que a nervação pode ser aberta ou fechada. Seguiu-se então uma mudança ramificação, de dicotômica a simpódica – Figura 1d - , num eixo ereto e sem folhas. Disto resultou um eixo com uma sucessão de sistemas telômicos. Houve então a necessidade da fusão dos telomas, dispostos lado a lado, trazendo como conseqüência uma folha constituída por pecíolos e limbo, com um sistema de nervuras ramificadas dicotomicamente e abertas. E as micrófilas originaram num eixo reto e áfilo com ramificação dicotómica, onde houve um sobrepujamento como resultado da passagem à ramificação simpódica. Uma outra teoria foi formulada por Bower, esta é chamada de Teoria da enação. Tal teoria afirma que as folhas licopodias são estruturas novas que surgiram em superfícies que não tinham saliência. Admitiu-se, então, a presença de expansões (enações), de natureza mais ou menos semelhantes a espinhos, surgiram na superfície lisa de um eixo se folhas e que, com o tempo, se tornaram achatadas e foliáceas. Após o aparecimento do cordão vascular (nervura) disposto até à extremidade da folha as enações passaram a ter ligação com o cilindro vascular do eixo. Porém as teorias acima mostradas demonstram apenas como evoluiu o sistema caulinar, mas esquece de mencionar como evoluiu o sistema vascular, este um mecanismo de fundamental importância para a sobrevivência das plantas no meio terrestre. Paralelo a evolução do sistema caulinar ocorreu à evolução do sistema vascular. Os tecidos vasculares primários constituíam o cilindro central ou estelo, do caule, da raiz no corpo primário da planta. A evolução dos tecidos vasculares surgiu da necessidade da planta transportar água e nutrientes; com isso, as plantas desenvolveram diversas organizações vasculares. São conhecidos vários tipos de estelo, entre eles o protostelo (cilindro maciço), sifanostelo (cilindro maciço sem a preenchimento e com parênquima) e eustelo (cilindro em fuso). A diversificação dos arranjos vasculares no processo evolutivo implica também, entre outros fatores, na fotossíntese e no transporte de água às folhas. 11 A distribuição relativa dos sistemas vasculares influencia nas principais diferenças nas estruturas da raiz, caule e folha, como por exemplo: protostelo forma micrófilos, sifanostelo forma micrófilos e megáfilos e eustelo forma apenas megáfilos, estes deramm origem as folhas atuais. Estes tecidos vasculares – xilema primário e floema primário -, tiveram que se tornar mais resistentes cada célula dos elementos traqueais, desenvolveu um preenchimento com lignina característico, para poder levar a seiva para o topo da planta, além de dar maior resistência à planta. 12 3- Evolução As pteridófitas constituem um grupo vegetal com características peculiares. De acordo com estudos da paleobotânica as primeiras pteridófitas compreendem vegetais vasculares muito simples que surgiram por volta de 400 milhões de anos durante a era Paleozóica dos períodos Siluriano ao Devoniano (figura 2) . Neste período existia um considerável número de Psilófitos (que como será visto mais na frente, tais organismos apresentavam uma organização bem simples), estes ocupavam uma região de transição entre a água e a terra. Figura 2 – período em que surgiram as primeira pteridófitas Figura 3 – Período carbonífero. Inicia-se o surgimento de grandes florestas pantanosas Mas nesse mesmo período – meados do Devoniano – os três mais importantes exemplares de plantas vasculares resumia-se aos filos Rhyniophyta, Zosterophyllophyta e Thimerophytiphyta. Tais filos apresentavam uma estrutura relativamente pequena e morfologia simples e primitiva. Não apresentavam raízes, na fase esporofítica, constando apenas de ramos com folhas muito pequenas, quando presentes. Ao final desse período estes três filos foram extintos e grupos mais complexos tornaram-se dominantes. Ainda na Era Paleozóica, segue-se o período Carbonífero (Figura 3) que teve como características o surgimento de grandes florestas pantanosas, tropicais e subtropicais, seguido por, no permiano, uma a extinção de algumas espécies de pteridófitas, como exemplo pode citar as Lycophytas arbóreas. Viver em Terra firme inicialmente não foi fácil, para solucionar esse problema de sustentação em terra firme, cada célula desenvolveu um preenchimento com lignina em sua parede celular que lhe possibilitou maior resistência à planta e conseqüentemente proporcionou a forma ereta da planta além de possibilitar um 13 crescimento maior do vegetal – muito embora as primeiras plantas como a Rhynia, possuía aproximadamente meio metro de altura. Diz-se que elas foram pioneiras no que se refere: à presença de vasos condutores de seiva, o xilema e o floema (solucionando, assim, a dificuldade de transporte de água e alimentos através da planta); e na presença de estômatos (porém neste ponto alguns autores discordam e afirmam que as primeiras plantas a apresentarem estômatos foram as briófitas do filo Anthocerophyta), apesar de que estas primeiras plantas eram destituídas de raízes verdadeiras, no lugar apresentava ainda rizóides e seu caule era subterrâneo do tipo rizoma. Era composta por talos cilíndricos, fotossintetizantes e que possuíam cutícula e em cuja extremidade se encontrava o esporófito. A geração gametofítica tornou-se gradualmente menor e protegida e dependente do esporófito, quanto à nutrição. Uma outra diferença que elas adquiriram no decorrer do curso evolutivo é que tais plantas foram os primeiros vegetais com Heterosporia, ou seja, que produz esporos diferentes por meiose, sendo estes denominados Micrósporo e micrósporo. Então, a partir das características acima citadas adicionadas a outras que serão mais na frente discutidas, quando se classifica os primeiros representantes das pteridófitas fala-se que são vegetais criptógamos vasculares e cormófitos. Porém as primeiras plantas vasculares, mesmo tentando se tornar independente da vida aquática, da mesma forma que as briófitas, dependiam da água para a fecundação. Quando se estuda e compara morfologicamente os vegetais atuais e os fósseis das primeiras plantas vasculares (as pteridófitas) e tenta achar explicações para o modo de evolução desses organismos vegetais, verifica-se que esses vegetais não encaixam-se na lacuna entre pteridófitas e plantas inferiores na escala evolutiva a partir daí tenta-se desvendar com teorias a forma de evolução desses vegetais, pode-se destacar,m então, algumas teorias, citadas a baixo. A primeira teoria postula a origem a partir das algas verdes, parenquimatosa com talos complexos, alternância de gerações isomórfica e meristemas apicais. Alguns defensores dessa teoria não relacionam os pteridófitos com qualquer grupo de algas. Outros autores defendem a idéias de que essa evolução deu-se a partir de algas pardas, que crescem próximas ao limite superior das marés e vivem nos níveis mais elevados da zona intercotidal. Todos que acreditam nessa teoria afirmam que a relação 14 filogenética entre briófitas e pteridófitas foi devido a uma evolução paralela. Entretanto não pode negar que muitas características entre esses dois taxa são tão fortes que há razões para acreditas numa relação filogenética. A segunda teoria crer, portanto, em uma relação direta ou indireta das pteridófitas com as briófitas, a partir de duas linhas evolutivas divergentes de plantas terrestres que já possuem arquegônio, embora estes muito primitivo. Porém ouros defensores dessa teoria, seguindo por outra linha de raciocínio, afirmam que os pteridófitos provêm de Anthoceros. Uma terceira teoria ainda afirma que as plantas vasculares evoluíram com as várias contribuições do genoma dos fungos que levou a especialização de várias células; Sendo o corpo vegetal um mosaico onde seriam encontradas várias células de algas e fungos adicionado de várias formas intermediárias. Também não se pode esquecer a teoria que afirma que pode ter havido a possibilidade de um organismo parasita tenha se integrado ao a planta, transferindo seus genes para o hospedeiro, e tenha ajudado a planta a evoluir. 15 4 – Características gerais do grupo As pteridófitas são plantas vasculares ou traqueófitas com vasos condutores do tipo xilema ou lenho e floema ou líber - que solucionou o problema de transporte de água e alimento através da planta -, e foi através desses vasos que a seiva bruta, contendo água e sais minerais, e a seiva elaborada, rica principalmente em açúcares dissolvidos na água, são transportadas. Tal sistema permite grande velocidade no transporte de materiais.Sobre este aspecto ainda é importante citar que elas sintetizam lignina. Tal característica permitiu que estas plantas a atingir maiores dimensões do que qualquer outra planta terrestre existente até então - atualmente são ditas como plantas de porte variado-, transformando-as nas primeiras plantas a abandonar por completo o meio aquático e conquistarem o ambiente terrestre. Apresentam o corpo na forma de cormo, isto é, com raiz (que formam o sistema radicular e é protegido pela coifa), caule e folhas (que formam o sistema caulinar) verdadeiros. Mas não possuem sementes, muito menos de flores e frutos, reproduzindo-se por esporos (que são agrupados em soros) o que os colocam no grupo das Criptógamas. Possuem folhas grandes denominadas frondes que geralmente são divididas em folíolos; as folhas jovens ficam enroladas e são chamadas de báculos. Em geral, a folha é a única parte visível da planta, pois o caule é subterrâneo ou fica rente ao solo, com crescimento horizontal. Este tipo de caule, que lembra uma raiz, é chamado rizoma. Podem apresentar propagação vegetativa, pois o rizoma pode, em determinados pontos, desenvolver folhas e raízes, dando origem a novos indivíduos, e se seguido por possível apodrecimento do rizoma em certos pontos, essas plantas podem tornar-se indivíduos independentes. Algumas, ainda apresentam diferenciação em suas folhas de tal forma que algumas folhas se encarregam das funções vegetativas, estas são denominadas de trofófilo e um segundo tipo se encarrega das funções vegetativas e das funções reprodutivas, estas são chamadas de esporófilo. A esse fenômeno dá-se o nome de heterofilia. Sobre as traqueófitas sem semente ainda podemos afirmar que tem como substância de reserva o Amido, além de alguns lipídios, polissacarídeos e proteínas. Sua parede celular é composta de celulose, hemicelulose, lignina., esta última de 16 fundamental importância na evolução dessas plantas. Além da sua forma de crescimento, como as outras traqueófitas, ser axial. Estas plantas apresentam um ciclo de vida de alternância de gerações (com fase gametofítica e esporofítica) heteromorfas, sendo que a fase esporofítica a fase dominante (a planta propriamente dita) sendo independente do gametófito, este último é a fase transitória e recebe o nome de prótalo, e também é haplóide. O prótalo é uma estrutura geralmente pequena de coloração verde e em forma de lâmina vivendo acima do solo. Em alguns casos pode ser ainda saprófito e ser encontrado dentro do solo, sendo neste caso incolor. De qualquer forma ele tem um período de vida curto não passando de algumas semanas, somente em situações especiais, caso não haja a fecundação o prótalo, é que este pode viver mais tempo podendo viver até alguns anos. Além disso, são oogâmicas como todas as plantas vasculares. Possuem arquegônios contendo a oosfera grande e imóvel. Os gametas masculinos são pequenos anterozóides multiflagelados que nadam ou são conduzidos até a oosfera. Em condições adequadas, as paredes do anterídio se rompem, liberando os anterozóides que nadam até o arquegônio, que ali penetram por um canal, atingindo a oosfera. Ocorre então a fecundação, sempre com a participação da água. O zigoto germina sobre a planta-mãe, dando origem ao esporófito dominante, que possui esporos, com uma parede bem definida (exina) impregnada com esporopolenina, em esporângios; esses esporos, ao germinarem, darão origem a uma nova planta-mãe. O ambiente são os mais variados podendo ser encontradas desde ambientes de altas latitudes e altitudes até o interior de florestas pluviais tropicais e de vegetação arbustiva sub-desértica a rochas costeiras e mangues. Mas a maior parte das pteridófitas habita as regiões tropicais, mas podem ser encontradas em regiões temperadas até mesmo desérticas. 17 5 – Classificação A tabela que segue logo a baixo exemplifica como são classificadas as pteridófitas atualmente. Antes de ler a tabela têm que saber que este grupo de vegetais vascularizados pertence, assim como todas as plantas, ao Domínio Eukaria e do Reino Plantae. O sistema de classificação botânica variou bastante nos últimos anos. Antes existia o filo pteridófitas com suas diversas classes. Mas atualmente o termo Pteridófita, designa apenas um grupo taxonômico não formal, formado por sete filos (demonstrados abaixo) – que eram as antigas clases e com diversos gêneros. DIVISÃO Rhyniophyta CLASSE Rhyniopsida ORDEM Rhyniales FAMÍLIA Rhyniaceae Cooksoniacea Zosterophyllophyt Zosterophyllopsid a a Zosterophyllales Zosterophyllaceae Asteroxylales Asteroxylaceae Protolepidodendral es Protolepidodendrace ae Drepanophycaceae Lycophyta Lycopodiaceae Lepidodendrales Lepidodendraceae Selaginellas Selaginellaceae Isoetales Isoetaceae Pleuromeiales Pleuromeiaceae Trimerophytales Trimerophytaceae Psilotales Psilotacea Lycopsida Trimerophytophyt Trimerophytopsid a a Psilotophyta Lycopodiales Psilotopsida GÊNERO Rhynia Horneophyton Cooksonia Zosterophyllium Crenaticaulius Rebuchia Sawdonia Asteroxylon Protolepidodendro m Leclercquia Baragwanathia Drepanophycus Lycopodium phylloglossum Lycopodites Lepidodendron Lepidoohloios Lepidostrobus Sigillaria Stigmaria Selaginella Selaginellites Isoetes Stylites Isoetites Pleuromeia Trimerophyton Psilophyton Pertica Psilotum Tmesipteris 18 Sphenophyta Hyeniales Hyeniaceae Calamitales Calamitaceae Sphenophyllales Sphenophyllaceae Equisetales Equisetaceae Ophioglossopsida Ophioglossales Ophioglossaceae Marattiopisida Marattiales Marattiaceae Sphenopsida Hyenia Calamites Arthropitys Asterophyllites Sphenophyllum Equisetum Equisetites Cladoxylopsida Coenopteridopsida Schizaeceae Gleicheniaceae Osmundaceae Matoniaceae Pterophyta Filicales Polypodiaceae Filicopsida Cyatheaceae Hymenophyllaceae Marsileales Salviniales Marsileaceae Salviniacecae Ophioglossum Botrychium Marattia Angiopteris Psaronius Schizaea Gleichenia Osmunda Matonia Adiantum Asplenium Blechnum Dryopteris Pellaea Polypodium Pteridium Woodsia Cheilanthes Platycerium Polystichum Cnemidaria Cyathea Trichomanes Marsiela Regnellidium Azolla Salvinia Destes organismos acima apresentado vale ressaltar que alguns estão extintos, como por exemplo os filos Rhyniophyta, Zosterophyllophyta, e Trimerophytophyta. Além das classes Cladoxylopsida e Coenopteridopsida. Além de algumas ordens de Lycophyta. 19 6 – Particularidades do principais filos FILO RHYNIOPHYTA Em solos do final do Silúrico, mais ou menos há 420 milhões de anos atrás, foram encontrados os primeiros fósseis de plantas vasculares completas, estas pertenciam na sua maioria a divisão Rhyniophyta, como exemplo pode-se citar os gêneros: Cooksonia e Rhynia. O primeiro é a mais antiga planta vascular conhecida e habitava superfícies lodosas e, possuía caules aéreos delgados que cresciam para cima cerca de 6.5 cm, também possuía esporângios globosos. A Rhynia, por sua vez, é o membro mais conhecido desse filo, habitava brejos e possuía caules aéreos ramificados dicotomicamente com cerca de vinte centímetros de comprimento revestidos com uma cutícula e possuidores de estômatos, Ambas possuíam talos eretos, sem folhas e apresentando esporângios terminais e dicotomicamente ramificados. Não diferenciavam raízes, caule e folhas, mas possuíam rizomas subterrâneos. Os esporângios destas primeiras plantas terrestres eram inseridos nos ápices dos talos férteis e produziam um único tipo de esporo, por isso é dito como uma planta homosporada. Esta estrutura pode ser melhor visualizada na figura 4 . O gametófito era monóico, cilíndrico, subterrâneos e bifurcados, com números rizóides. Careciam de clorofila e provavelmente mantinham associações com fungos endofíticos Possuía anterozóides multiflagelados e espiralizados. Os anterídios eram hemiesféricos, rodeados por uma capa de células estéreis e o arquegônio era Figura 4 – Reconstrução de esporófito de uma Rhynia, observa-se a presença de talos dicotomicamente ramificados e os esporângios fusiformes na terminação do talo parcialmente fundido no gametófito. Estas primeiras plantas não eram muito altas, faziam pouca sombra, 20 reteriam a umidade e levariam à acumulação de detritos orgânicos entre elas Os primeiros organismos terrestres terão, assim, determinado a formação do humos (um solo muito orgânico). Atualmente agrupa os gêneros Psilotum, Tmesipteris mas a maiorias dos membros deste filo se extinguiu por volta de 380 milhões de anos atrás, entre estes membros extintos pode-se citar Psilophyton (figura 5 ), Rhynia e Asteroxylon Figura 5 – Reconstrução de esporófito de uma Psilophyton, com detalhe para seu esporângios na terminação do talo. FILO ZOSTEROPHYLLOPHYTA Evoluindo das Rhyniophyta no começo do devoniano, as zosterófitas são um grupo de plantas pequenas, que viveu no devoniano entre 408 até 370 milhoes de anos atrás. Este grupo ocupou um lugar chave na evolução de plantas da terra. De acordo com fósseis achados, foi observado que este grupo vegetal possui características típicas de licófitas, por isso acreditam serem ancestrais das licófitas. As Zosterófitas (figura 6 ) não possuem nem folhas e eram dicotomicamente ramificadas, no qual cada haste se dividia em duas hastes de tamanho igual .Não possuíam raízes verdadeiras, seus caules aéreos eram revestidos por uma cutícula, alguns ainda possuíam estômatos nos ramos mais acima, transparecendo que os de baixo ficassem imersos na água, o que demonstrava que o grupo era, possivelmente, aquático. 21 Figura 6 – esta reconstrução de um zosterophyllophyta, que mostra como se organizava os numerosos esporófito e no detalhe um esporângio terminal. Eles produziam ramos laterais que se dividiam em dois eixos, um que crescia pára cima e outro que crescia voltado para baixo, este último funcionava com raízes. Entre outras características desse filo pode ser citado ainda que a fotossíntese era realizada provável-mente toda sobre as hastes da planta e que tinham esporângios globosos formados lateralmente em pedicelos curtos. FILO LYCOPHYTA Os vegetais da divisão Licophyta (do grego lycos - lobo; podos - pé; e phyton – planta) também possui seus precursores no Devoniano e muito provavelmente seus progenitores foram as primeiras Zosterophyllphyta, são representados pelos licopódios (plantas homosporadas) e selaginelas (heterosporadas). Atualmente possuem 10 a 15 gêneros e aproximadamente 1000 espécies conhecidas. Os esporófitos da maioria dos gêneros deste filo são constituídos por um rizoma (caule subterâneo horizontal) ramificado que emite ramos aéreos e raízes verdadeiras. As folhas são vascularizadas e dispõem-se espiraladamente ao redor do caule e são do tipo microfila. O rizoma e as raízes também são vascularizados. Os estudos relacionados a essa divisão mostram que elas possuem pelo menos três ordens extintas. E as outras três conhecidas atualmente constituem inteiramente de ervas; cada ordem inclui uma única família. Embora sejam plantas herbáceas, seus ancestrais fósseis atingiam dimensões arbóreas. Os esporofilos localizam-se em ramos modificados, com entrenós muito curtos, os estróbilos, tais 22 estruturas são onde os esporângios encontram-se reunidos, situados na axila de folhas modificadas com função de proteção. A partir da meiose, nos esporângios, ocorre uma diferenciação formando esporos haplóides que originam os gametófitos bissexuados, que podem ser estruturas verdes, irregularmente lobadas (ex.: Lycopodiella) ou estruturas micorrízicas subterrâneas (ex.: Lycopodium) – figura 7-. As licopodófitas podem ser homosporadas como briófitas ou psilófitas. Entretanto, alguns gêneros podem apresentar esporângios diferenciados originando dois tipos de esporos: os megásporos (esporos femininos) de tamanho maior originados no megasporângio, e o micrósporos (esporos masculinos) de tamanho menor originados no microsporângios. Plantas que possuem esse tipo de diferenciação de esporos e esporângios são denominadas heterosporadas. Figura 7- um exemplar de Lycopodium, Figura 8 – Aspecto de uma selaginella Lycopodium obcurum Os esporos irão germinar e darão origem a gametófitos cilíndricos, haplóides, maciços, sempre dióicos e clorofilados nas espécies hetrosporadas. Estes por sua vez originarão arquegônios ou anterídios. Até que os arquegônios e anterídios de um Lycopodiaceae se desenvolvam completamente, pode ocorrer um período de cerca de 15 anos. E como foi falado anteriormente ainda haverá a necessidade de água para a fecundação, o anterozóide biflagelados nada, então, até o arquegônio, em seguida há sucessivas divisões celulares e ocorre o crescimento do embrião interior do arquegônio, originando o esporófito, que permanece por um tempo preso ao gametófito e depois se torna independente. O ciclo de vida de Lycophyta está representado na figura 10. As duas Ordens mais importantes dessa divisão são: Lycopodiaceae, e Selaginellae (figura 8). 23 A primeira é, como falado anteriormente, homosporada e possui folas sem lígulas. O esporófito é herbáceo podendo ou não ficar restritos a parte terminal (figura 9) dos ramos e constituem estróbilos definidos. O gametófito é totalmente ou parcialmente subterrâneo. E existem três famílias: Protolepidodendracea, Lycopodiaceae e a das Isoetaceae. Figura 9 – Lycopociaceae, observar o esporófito terminal Já a ordem Selaginellae compreende organismos de caule herbácio, sem apresentar qualquer sinal de engrossamento (o que difere das Lycopodiaceae que apresentam engrossamento secundário do caule da raiz). As folhas podem ser iguais ou dimórficas todas apresentando lígula. As raízes se formam a partir da extremidade distal dos ramos (rizóforos) do caule. Os esporófitos são heterospóricos, como dito anteriormente, e situase em estróbilos nas extremidades dos ramos. famílias: E existem Miadesmiacea duas e a Lepidodendrales Figura 10 - ciclo de vida de uma selaginella, representando o ciclo de vida do filo Lycophyta 24 FILO TRIMEROPHYTA Os organismos pertencentes ao filo Trimerophyta (figura 11) são espécies que provavelmente evoluíram diretamente dos Rhyniophyta, sendo um grupo de plantas também devonianas, estando presentes no período de aproximadamente entre 408 à 360 milhões de anos atrás. Possuíam crescimento axial e se diferenciavam dos outros grupos da época por possuir muitas ramificações (diferente dos filos anteriores que posdsuiam ramificação dicotômica), e somente algumas das extremidades possuíam esporângios. Isto permitiu que estas plantas crescessem continuamente e muito maiores do que Rhynia. Trimerophytopsida, assim como as Rhyniopsida, possuíam talos áfilos, rizomas e os esporângios terminais.Mas diferenciavam-se no que se refere ao porte, Trimerophytopsida eram muito maiores atingindo até 4 m de altura, e pelo sistema de ramificação mais complexo, com eixo principal e ramos laterais, e esporângios agrupados em cachos e com mecanismo de ruptura ou deiscência para liberação dos esporos. E justamente por essa grande semelhança com as Rhyniopsida é que por muito tempo ela foi posta como sendo da outra ordem. Estes organismos provavelmente forma os progenitores das plantas modernas com sementes Figura 11 – representante de Trimerophyta FILO PSILOPHYTA Os Psilophyta (do grego psilos = nu; e phyton = planta) compreende a divisão de vegetais vasculares mais simples - que supõem, em geral, seu caráter de primitividade - e que durante muito tempo não foi considerado como um grupo taxonômico coerente pelos paleobotânicos, por que se tratava de uma planta muito diferente de qualquer outra conhecida atual ou fóssil. Porém com a descoberta de restos de plantas idênticas na natureza tornou-se claro a identificação desta planta e as agruparam todas na divisão das Psilophyta. O nome para o grupo foi aplicado originalmente em 1927 para um grupo de plantas fósseis da era Paleozóica dos períodos do médio Siluriano ao Devoniano 25 inferior e médio – com idades superiores a 300 milhões de anos -, incluindo Psilophyton e Rhynia, que mais tarde foi expandida para incluir dois gêneros de plantas atuais com uma estrutura muito similar, tais gêneros são: Psilotum (figura 12)e Tmesipteris (figura 13); mas estes estão hoje em dia classificados separadamente numa divisão usualmente chamada de Psilotophyta. Figura 12 – representante de Psilotum Figura 13 – representante de Tmesipteris Constituem vegetais que têm esporófitos que diferem de todas as outras plantas vasculares, pois são desprovidos de raízes na fase esporofítica (em seu lugar existe um rizóides unicelulares, com micorrizas associadas). O esporófito possui um tamanho relativamente pequeno quando comparado às demais pteridófitas e está, geralmente, diferenciado numa parte subterrânea provida de rizóides e em uma outra área menor, o caule, este é delgado e muito ramificado (ramificações dicotômicas de toda a planta ou de suas partes terminais), cilíndrico, fotossintetizante e com um sistema vascular prostotélico pouco lignificado, ainda pode ser citada a presença de estômatos. Na maioria dos gêneros os ramos aéreos são destituídos de folha, em vez disso apresenta apenas escamas. Mas num pequeno número as folhas são pequenas, simples, sem nervuras e densamente inseridas. Os esporângios (FIGURA 14a) terminais são reunidos em grupos de três formando, assim, os sinângios (figura 14b), estes estão situados em ramos laterais muito curtos. A partir da meiose ocorrida nos esporângios surgem esporos cujo desenvolvimento origina os gametófitos. O gametófito monóico e temporário é muito pequeno e tem um formato cilíndrico. É saprófito e independente do esporófito, apesar de ser totalmente aclorofilado. Encontra-se sempre em simbiose com fungos. O ciclo de vida está representado pela figura 15 26 Figura 14 - Esporângio de Psilophyta, em a, pode ser visto três esporângio que juntos formam os sinângios (b) Alguns gêneros como Psilotum e Tmesipteris podem ser encontrados sempre em simbiose com fungos, tais fungos são zigomicetos endomicorrízicos. O primeiro gênero é característico de regiões tropicais e subtropicais e o segundo nativo da Nova Zelândia e Austrália. No Brasil este filo é representado pelo gênero Psylotum, uma planta herbácea com ramificação dicotômica Que em muito se assemelham ao gênero Rhynia, um gênero que existiu no período Devoniano. Na planta adulta, os eixos produzem esporângios trilobados em ramos laterais muito curtos; o gênero Psylotum é homosporado, os esporos são produzidos no esporângio e após a germinação, os esporos originam o gametófito bixexuado, que é uma estrutura aclorofilada (portanto saprófita) subterrânea, com associação de micorrizas. Os anterozóides de Psilotum são multiflagelados e necessitam de água para nadar até a oosfera; o esporófito originado sexualmente fica, inicialmente, preso ao gametófito, absorvendo seus nutrientes, mas depois solta-se do pé, que permanece no gametófito. O gênero Tmesipteris, cresce como uma epífita sobre samambaias arbóreas e outras plantas e fendas de rochas. Diferentemente do gênero Psilotum que possui apêndices em formas de escamas, as espécies deste gênero possui apêndices em forma de folhas além destes serem maiores que as escamas, mas em geral no que se refere à outros aspectos deste gênero é muito similar ao Psilotum. 27 Figura 15 – ciclo de vida de um Psilotophyta FILO SPHENOPHYTA Este filo é representado atualmente por um único gênero o Equisetum (figura16), cujas espécies são conhecidas por cavalinhas - cujo nome surge devido a semelhança do seu caule com uma cauda de cavalo. Este gênero é o único na família Equisetaceae, que, por sua vez, é a única família da ordem Equisetales e da classe Equisetopsida (também conhecida como Arthrophyta em livros antigos). Análises moleculares recentes o enquadram como sendo um gênero dentro das Pteridophytas, relacionando-as aos Marattiales. Porém estes estudos ainda são muito questionados e até mesmo a sistemática vive mudando constantemente. Este gênero teve sua origem, diferente dos outros, no final do Devoniano e foram importantes membros da flora durante o período Carbonífero. O gênero é comum nas cidades, está presente em quase todos continentes. são plantas perenes mas podem secar no inverno no caso da planta estar em regiões 28 temperadas, mas também pode permanecer sempre verde (para algumas espécies tropicais, e a espécie temperada Equisetum hyemale). A maioria delas são muito pequenas crescendo de 0,2 – 1,5 m de altura, embora possa haver São encontrados em terrenos paludosos e úmidos. Possui tamanho variável. Em geral compreendem vegetais vasculares com folhas micrófilas (diminutas) que se distribuem verticiladamente na região do nó, alguns autores ainda falam que não existem estruturas foliares. Nos entrenós encontram-se sulcos longitudinais, alternando com arestas longitudinais mais ou menos salientes, eles são estriados e, em suas células epidérmicas, eles apresentam sílica. Apresentando raízes verdadeiras (vascularizadas). Os caules são estriados (firmes e reforçadas com depósitos de sílica nas células epidémicas) – figura 17 - demonstrando rugosidade, articulados e ocos; dividen-se em nós curtos e entrenós compridos; aparecem de sistemas subterrâneos que permanecem vivos até mesmo durante as estações desfavoráveis, já que em tais estações a parte aérea morre. Esta mesma estrutura é verde e fotossensível, sendo ela responsável pela maior parte da fotossíntese realizada - a atividade fotossintetizante das folhas é desprezível. Apresentam raízes adventícias que surgem dos nós dos rizomas e ainda caules aéreos que surgem de ramificações desses rizomas subterrâneos. Figura 16 - Equisetum representante do filo Sphenophyta Figura 17 – Caule estriado, mostrando os nós e entrenós Figura 18 – Estróbilos de Sphenophyta 29 No ápice dos ramos férteis encontram-se os densos estróbilos (figura 18), que diferenciam das de Lycopodophyta. Em Sphenophyta os estróbilos podem estar localizados em ramos estrobilíferos aclorofilados (E. arvense) ou nas extremidades de ramos vegetativos (E. hyemale), mas, em geral são formados nas extremidades de diferentes ramos vegetativos. Os esporângios encontram-se reunidos em esporangióforos, estes, por sua vez, estão reunidos nos estróbilos no ápice do caule. São homosporadas, originando-se nos esporângios apenas um tipo de esporo a partir da meiose. Quando os esporos estão maduros os esporângios contraem-se e rompem-se ao longo de sua superfície interna, liberando os esporos. Um fator que ajuda no processo de dispersão dos esporos é a presença de elatérios originados de sua parede celular e que, espiralizam–se com a umidade e pela perda de água distendem-se auxiliando na dispersão dos esporos. Os gametófitos haplóides, originados a partir do desenvolvimento desses esporos, são: muito pequenos (no máximo 1cm, com algumas exceções), membranosos, dióicos, podendo apresentar dimorfismo sexual ou monóicos, neste caso apresentando protoginia, observando-se inicialmente o aparecimento dos arquegônios e, apenas após o desaparecimento destes, o de anterídios. Possuem vida livre e estabelecem-se principalmente sobre a lama que tenha sido recentemente inundada e esteja cheia de nutrientes. Possuem anterídios multiflagelados que nadam até a oosfera (uma característica comum a todos as pteridófitas). Esta Divisão apresenta apenas 1 gênero atual, Equisetum, com espécies ocorrendo tanto em regiões temperadas com tropicais. Porém este filo compreende mais quatro ordens Hyeniales, Pseudoborniales, Sphenophyllales, Calamitales, todas estas somente na forma de fósseis. FILO PTEROPHYTA As Pterophytas (conhecidas também como filicíneas) atuais, é o filo mais conhecido entre todas as Pteridófitas, compreendem um grupo muito diverso de 30 plantas vasculares sem sementes, sendo representado por seis ordens denominadas: Marattiales, Ophioglossales, Psilotales, Osmundales, Polypodiales e Hydropteridales. Quando muitas vezes queremos exemplificar uma pteridófitas, utilizamos como exemplo uma samambaia (um exemplar de pterophyta), este exemplo tem como a finalidade demonstrara quão próximo de nós está os organismos desse filo, que inclusive é denominado coletivamente como pteridófitas ou samambaias (figura 19), de tamanha a sua importância para este grupo. Outras plantas muito importantes desse filo são as avencas e o xaxim. Este filo engloba aproximadamente cerca de 11.000 espécies no mundo e 1300 espécies no Brasil, sendo que a maioria é encontrada em regiões tropicais – onde cerca de 75% das espécies são encontradas- , pois nesse tipo de ambiente existe umidade e sombra (tudo o que elas precisam) tendo como destaque os vegetais epifíticos, aquáticos ou aqueles que possuem caule aéreo. Considerando as critógamas este grupo é o que apresenta maior diversidade de formas decrescimento. O tamanho e habitat são muito variáveis; no que se refere ao tamanho, pode-se encontrar desde espécies trepadeiras até árvores muito grandes. De acordo com estudos de paleobotânicos algumas famílias de samambaias apresentam abundante registro fóssil, sendo alguns desses registros anteriores ao período Carbonífero, o que nos leva a concluir que as samambaias estão entre as plantas atuais com ancestrais mais antigos. De maneira geral, apesar da ausência de crescimento secundário limitar seu crescimento, as Pteridófitas foram um componente importante das formações vegetais do Paleozóico. Este filo compreende criptógamas vasculares com folhas e raízes verdadeiras, portanto vascularizados.Tais folhas são, na maioria dos grupos, macrofilas simples ou composta (pinadas), com folíolos ligados entre si por uma nervura central e sua vascularização deixam lacunas no cilindro vascular, além de que permite que estes grupos apresentem folhas com um tamanho maior que as demais traqueófitas descritas até agora. Neste caso os folíolos ficam ligados entre si por uma nervura central denominada Raque. Dentre as folhas simples, o limbo 31 pode ser pinatisecta (contínuo) ou isolados (pinatifida) A ramificação pinada pode ser simples (pinada) ou se repetir por uma (bipinada), duas (tripinada) ou mais vezes. Mas algumas samambaias são muito pequenas e tem folhas inteiras, como no caso da Lygodium. Na maioria dos grupos, a face inferior das folhas cresce mais que a superior a esse processo dar-se o nome de vernação circinada, resultando na formação do que se denomina báculo. (figura 20) Figura 19 – samambaia representando o filo Pterophyta Figura 20 - Báculo O caule é muito pequeno, tendo de quatro a seis centímetros de diâmetro, pode ser do tipo rizoma (lembrando uma raiz) ou aéreo. O gênero Botrychium é a única samambaia atual conhecia a formar um câmbio vascular. Existem dois tipos de classificação para as samambaias: as eusporangiadas ou leptosporangiadas. No caso da primeira classificação, é produzido um eusporângio, esporângio de origem multicelular, tais células estão localizadas na superfície do tecido a partir do qual o esporângio será produzido As ordens Marattiales, Ophioglossales e Psilotales são eusporangiadas; no segundo caso o esporângio será formado a partir de uma única célula inicial. Os esporângios encontram-se reunidos em soros, esporocarpos, espigas ou sinângios – Neste caso enquadram-se Polypodiales e Hydropteridales. A célula que vai originar este tipo de esporângio vai sofrer sucessivas divisões, formando duas camadas: a mais interna originará uma grande parte do pedicelo ou permanece inativa; a mais externa originará, mediante um padrão de divisões celulares, dará origem a uma esporângio pedicelado, com uma cápsula globosa 32 que possui uma parede com uma camada de células de espessuras. Do lado interno dessa parede existe uma estrutura nutritiva com duas camadas de células de espessura denominada tapete. A massa interna diferencia-se e cada célula por meiose originará quatro esporos. Na ordem Osmundales o esporângio tem origem a partir de uma única célula. Entretanto, mas de uma célula participa do desenvolvimento do esporângio. Assim a ordem Osmundales não caracteriza-se nem com eusporangiada e nem como leptosporangiada. Nos grupos nos quais os esporângios encontram-se reunidos em soros, pode haver dois tipos de classificação diferente. A primeira é feita de acordo com a membrana que protege este soro, quando está na face inferior do folíolo, o soro é protegido por uma membrana chamada indúzio, quando está na borda do folíolo protegido por uma dobra e chamado falso indúzio. Uma outra diferença refere-se a uma estrutura diferenciada, o anel ou ânulo que, através de movimentos higroscópicos advindos da dessecação -, é responsável pelo rompimento do estômio (camada de células com menor resistência), liberando os esporos, que irão germinar, formando um novo gametófifo (prótalo). A maioria das samambaias atuais é homosporada, com exceção dos representantes de duas ordens aquáticas. Uma característica importante de muitas Pteridófitas extintas e atuais é a grande semelhança, em aparência, desenvolvimento e comportamento entre as folhas e sistemas de ramificação. Em algumas espécies não é possível se diferenciar as partes do sistema aéreo, resultado da evolução de apêndices laterais de alguns grupos ancestrais. As Pteridófitas se diferenciam das Gimnospermas por serem na maioria homosporadas e por não apresentarem de maneira geral crescimento secundário. Como esse grupo é muito grande não como falar de particularidades sem mencionar pelo menos três tipos, são eles: Ophioglossales e Marattiales; Fillicales; e as ordens Marsileaales e Salviniales. No primeiro grupo então enquadram-se plantas de regiões temperadas, cujas existe apenas uma única folha, esta consiste em duas partes: uma porção 33 vegetativa, ou lâmina e um segmento fértil. Os gametófitos são estruturas subterrâneas, tuberosas, alongadas, com numerosos rizóides. Além de serem eusporangiadas, o que difere das demais. É no segundo grupo, que estão a maioria das samambaias conhecidas (cerca de 35 família e 320 gêneros). São leptosporângidas e homosporadas. A maioria tem rizomas sifonostélicos. Pode formar raiz verdadeira, mas esta rapidamente murcha e o restante são raízes adventícias.As folhas, ou frondes, são megáfilos (sendo as únicas plantas vasculares sem sementes que possuem megáfilos bem desenvolvidos) e representam a parte mais conspícua do esporófito e são formadas pelas pinas que estão presas ao raque.A folha desenvolve-se por vernação circinada, originando báculos. Os esporângios estão agrupados em soros que muitas vezes estão cobertos pelo indúsio. Os esporos dão origem ao gametófito de vida livre, bissexuado, na forma de protalo. A superfície inferior do gametófito origina o arquegônio e os anterídios, que quando há o amadurecimento dos anterozóides – flagelados -, ocorre a liberação deste para ocorrer a fecundação. A pós a fecundação forma-se o zigoto que por divisões celulares irás originar o esporófito adulto que quando adquire um nível de fotossíntese suficiente para se manter enraizar-se e o gametófito desintegra-se. Os esporos serão produzidos nos soros e após sua liberação ia originar o gametófito. No último grupo enquadram-se as samambaias aquáticas. Elas são heterosporadas . Sua estrutura reprodutiva é denominada de esporocarpo. Possuem gametófitos extremamente especializados. Podem estabelecer relações simbiontes com cianobactérias. 34 2.7- Importância econômica As pteridófitas são normalmente usadas como plantas ornamentais, neste aspecto as samambaias e avencas são muito cultivados pelas sua beleza e a forma de suas folhas. O emaranhado das raízes dos fetos arborescentes são muito utilizados por poderem servir como substrato para o cultivo de orquídeas. Uma outra importância refere-se ao uso de carvão mineral a partir dessas plantas, durante o carbonífero as pteridófitas dominavam grandes áreas, com espécies que chegavam a 30 ou 40metros de altura. Estas, sob condições especiais, transformaram-se em carvão mineral (carvão de pedra), utilizado, atualmente, como combustível. O tronco do samambaiaçu (figura 21) é utilizado para fazer xaxim (figura 22). Sobre o Xaxim ele é visto como um vaso orgânico - é um entrelaçado de raízes adventícias que saem do caule de certas samambaias que retém umidade. E devido a exploração Algumas pteridófitas são intensa vem sendo ameaçado de extinção. figura 21 - Samambaiaçu usadas como vermífugos como exemplo podemos citar o Apidium filix mas, desta planta é extraído do rizoma o vermifugo e utilizado no combate à teníase. Outras espécies foram muito utilizadas para polir potes e panelas, na época colonial e de expansão de fronteiras. Tradicionalmente, ainda, são usadas na culinária oriental, alguns brotos de samambaia têm ação comprovadamente cancerígena. As avencas também podem ser comestíveis e algumas espécies podem ser medicinais. Figura 22 - Xaxim 35 2.8- Importância ecológica As pteridófitas são importantes componentes da flora e são fundamentais para o desenvolvimento e estabelecimento de outros grupos vegetais e animais. Porém, há algumas que podem ser seletivas em relação ao seu habitat, presentes apenas em áreas mais conservadas de mata fechada. Contribuem na manutenção da umidade no interior da floresta, absorvendo água pelas raízes densas e distribuindo-a gradualmente ao solo e ao ar, desenvolvendo a microfauna e microflora do substrato, extremamente necessárias para o equilíbrio ecológico do ambiente. Sendo plantas sensíveis às mínimas variações das condições climáticas, as pteridófitas constituem-se em importante banco de dados das características pretéritas e atuais de uma dada região. Ainda apresentam importância ecológica pois são base da cadeia alimentar. Porém algumas espécies como Salvinia molesta e Pteridium aquilinum, são plantas invasoras e podem infestar enormes áreas de lagos e rios (no caso da primeira) e terrenos queimados (no caso da segunda) e sua presença no ambiente pode funcionar até como bioindicador de problemas ambientais. 36 Conclusão O conhecimento sobre a importância das pteridófitas, além de todos os seus aspectos nos fazem refletir como é importante estudar esses organismos. Pos eles servem de gancho na evolução de plantas aquáticas a terrestres. Além de este grupo é muito pouco estudado e necessita de mais atenção dos pesquisadores já que para poder compreender melhor o processo evolutivo dos vegetais precisa ter um conhecimento mais aprofundado sobre esses organismos 37 Biblografia RODRIGUES, Silvane Tavares, ALMEIDA, Samuel Soares de, ANDRADE, Laíse de Holanda Cavalcante et al. Composição florística e abundância de pteridófitas em três ambientes da bacia do rio Guamá, Belém, Pará, Brasil. Acta Amaz., 2004, vol.34, no.1, p.35-42. ISSN 0044-5967. PIETROBOM, Marcio Roberto e BARROS, Ivã Carneiro Leão. Pteridófitas de um remanescente de Floresta Atlântica em São Vicente Férrer, Pernambuco, Brasil: Pteridaceae. Acta Bot. Bras., out./dez. 2002, vol.16, no.4, p.457-479. ISSN 0102-3306. SANTIAGO, Augusto César Pessôa e BARROS, Ivã Carneiro Leão. Pteridoflora do Refúgio Ecológico Charles Darwin (Igarassu, Pernambuco, Brasil). Acta Bot. Bras., out./dez. 2003, vol.17, no.4, p.597-604. ISSN 0102-3306. XAVIER, Sergio Romero da Silva e BARROS, Ivã Carneiro Leão. Pteridoflora e seus aspectos ecológicos ocorrentes no Parque Ecológico João Vasconcelos Sobrinho, Caruaru, PE, Brasil. Acta Bot. Bras., out./dez. 2005, vol.19, no.4, p.775-781. ISSN 0102-3306. SOUSA, Maria Cristina de, GUILLAUMET, Jean-Louis, AGUIAR, Izonete de Jesus Araújo. Ocorrência e distribuição de pteridófitas na reserva florestal Walter Egler, Amazônia Central, Brasil. Acta Amaz., 2003, vol. 33, n° 4, p. 555-562. FIGUEREDO, Jane B., SALINO, Alexandre. Pteridófitas de quatro Reservas Particulares do Patrimônio Natural do Sul da Região Metropolitana de Belo Horizonte, Minas Gerais, Brasil. Lundiana, 2005, vol. 6, n° 2, pág 83-94. ISSN 1676-6180. RAVEN, P. H.; EVERT, R. F. & EICHORN, S. E. 2001. Biologia Vegetal. 6ª Ed. Editora Guanabara Koogan. 906p. SMITH, G. 1950. Botânica Criptogâmica. 3ª Edição Vol. 2: Briófitas e Pteridófitas. Fundação Calouste Gulbenkian. Lisboa. 386p. FERRI, MÁRIO GUIMARÃES. Botânica. 15ª Edição. Vol. 2: Morfologia Externa das Plantas. Editora Nobel. São Paulo. 1983. 147p. MAUSETH, JAMES D. Botany an Introduction to Plant Biology. 3ª edição Jones and Bartlett publishers. 2003 TRACHEOPHYTA . Disponível em: <http://www.dse.ufpb.br/lafic/Tracheophyta_geral.doc>. Acessado em: 06 jul. 2006 PTERIDÓFITAS. Disponível em: <http://professores.unisanta.br/maramagenta/pteridofitas.asp> . Acessado em 26 Jun. 2006. 38 TECIDOS VEGETAIS: ESTRUTURA E ENQUADRAMENTO EVOLUTIVO. Disponível em: < http://azolla.fc.ul.pt/aulas/BiologiaCelular/docs/Tec_Vegetais.pdf>. Acessado em 06 Jun. 2006 CHLOROBIONTA (GREN PLANTS) – 2. Disponível em: <http://www.palaeos.com/Plants/default.2.htm>. Acessado em 06 Jun. 2006 DEVONIANO VEGETAIS PRIMITIVOS . Disponível em: <http://www.geocities.com/historiadaterra/devegepri.html>. Acessado em 06 Jun. 2006. PTERIDÓFITAS. Disponível em: <http://www.geocities.com/CapeCanaveral/Lab/6969/pterid.htm>. Acessado em 30 Jun 2006. PTERIDOFITA: EVOLUÇÃO PARA PTERIDÓFITAS. Disponível em: <http://www.bio2000.hpg.ig.com.br/pteridofitas.htm>. Acesso em: 30 jun. 2006. A CLASSE POLYPODIOPSIDA (PTEROPHYTA). Disponível em: <http://www.dse.ufpb.br/lafic/Polypodiopsida.doc>. Acessado em: 05 Jul. 2006 39
Download