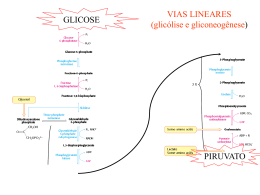
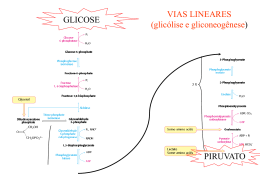
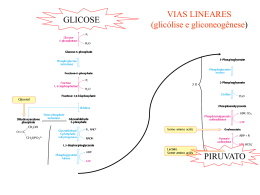
Exemplificando com uma situação extrema... 1960-1970: tratamento da obesidade com jejum prolongado assistido Variações metabólicas durante o jejum plasma 7 Ácidos Graxos - Hidroxybutirato Acetoacetato Glycerol Glicose Metabólitos (mM) 6 5 4 3 2 1 0 0 5 10 15 20 Tempo (dias) 25 30 35 Variações metabólicas durante o jejum 7 500 plasma 6 4 300 3 200 2 1 Glicogênio (mM) Metabólitos (mM) 400 5 Ácidos Graxos - Hidroxybutirato Acetoacetato Glycerol Glicose Glicogênio 100 0 fígado 0 0 5 10 15 20 Tempo (dias) 25 30 35 Variações metabólicas durante o jejum plasma 7 Metabólitos (mM) 6 Ácidos Graxos - Hidroxybutirato Acetoacetato Glycerol Glicose 5 4 3 2 1 0 0 5 10 15 20 Tempo (dias) 25 30 35 Gliconeogênese Síntese nova glicose Em humanos... NUTRIENTE QUANTIDADE (g) Triacilglicerídeos (tecido adiposo) Glicogênio (fígado) Glicogênio (músculo) Glicose (sangue e outros líquidos corporais) Proteína corporal (músculo, principalmente) 9.000 principal reserva 90 250 20 8.000 Por que precisamos sintetizar glicose? Células dependentes de glicose como nutriente glicose CÉLULAS ANAERÓBICAS ADP ATP lactato glicose ADP ATP CO2 hemácias células do cristalino algumas células da retina células da medula renal CÉLULAS COM ISOLAMENTO SELETIVO DA CIRCULAÇÃO SISTÊMICA células do cérebro (BHE) células embrionários (BP) Histórico... Claude Bernard: o fígado é capaz de fornecer glicose à circulação Glicogênio hepático glicose Glicogênio muscular lactato Lactato pode ser convertido em glicogênio no fígado? Journal of Biological Chemistry, 1929 Gerty Theresa Cori Carl Ferdinand Cori (Laureados com Prêmio Nobel em 1947) Lactato desidrogenase (LDH) •reação reversível em condições fisiológicas •enzima tetramérica dois tipos de subunidades: M (músculo esquelético), H (coração) M4 (músculo esquelético) M3H1 M2H2 M1H3 H4 (coração) diferentes valores de Km para os substratos lactato e piruvato glicose hexocinase Go´ = -8.0 kcal/mol ATP ADP glicose-6-fosfato fosfo-hexose isomerase Go´ = -0.6 kcal/mol frutose-6-fosfato fosfofrutoocinase Go´ = -5.3 kcal/mol ATP ADP frutose-1,6-bisfosfato aldolase Go´ = -0.3 kcal/mol triose-fosfato isomerase Go´ = +0.6 kcal/mol gliceraldeído-3-fosfato NAD+ Pi NADH gliceraldeído3P isomerase Go´ = -0.4 kcal/mol 1,3-bisfosfogliceratoato ADP fosfogliceratocinase Go´ = +0.3 kcal/mol ATP 3-fosfogliceratoato fosfoglicerato mutase Go´ = +0.2 kcal/mol Go´ 2-fosfogliceratoato enolase = -0.8 kcal/mol fosfoenolpiruvato piruvatocinase o G ´ = -4.0 kcal/mol ADP ATP piruvato glicose Perfil energético das reações da GLICÓLISE Etapas 1, 3 e 10 são contornadas na via de Gliconeogênese 1 kJ = 0,24 kcal piruvato Reações para conversão de piruvato a fosfoenolpiruvato (PEP) (para reversão da reação da piruvato cinase) NAD+ G3P piruvato NADH G3PDH 1,3BPG PEP piruvato PEPCK oxaloacetato NADH MDH PEP PEPCK PIRUVATO CARBOXILASE oxaloacetato MDH NADH NAD+ malato NAD+ malato G3P lactato NAD+ NAD+ NADH G3PDH piruvato NADH 1,3BPG PEP piruvato PEPCK oxaloacetato NADH MDH PEP PEPCK PIRUVATO CARBOXILASE oxaloacetato MDH NADH NAD+ malato NAD+ malato NAD+ G3P piruvato NADH G3PDH 1,3BPG PEP piruvato PEPCK oxaloacetato NADH MDH PEP PIRUVATO CARBOXILASE PEPCK oxaloacetato MDH aas NADH NAD+ malato NAD+ malato α-cetoglutarato fumarato succinato succinil-CoA F1,6BP glicerol-P DHAP NAD+ G3P piruvato NADH G3PDH 1,3BPG PEP piruvato PEPCK oxaloacetato NADH MDH PEP PEPCK PIRUVATO CARBOXILASE oxaloacetato MDH NADH NAD+ malato NAD+ malato F1,6BP lactato glicerol-P DHAP ácido graxo de cadeia ímpar G3P piruvato G3PDH acil-CoA de cadeia ímpar 1,3BPG PEP piruvato acetil-CoA PEPCK oxaloacetato MDH malato PEP PEPCK PIRUVATO CARBOXILASE oxaloacetato aas propionil-CoA MDH malato α-cetoglutarato fumarato succinato succinil-CoA O consumo de álcool, especialmente por um indivíduo mal alimentado, pode causar hipoglicemia. O álcool ingerido é convertido a acetaldeído no citoplasma do hepatócito, em reação catalisada pela álcool desidrogenase: NAD+ CH3CH2OH NADH CH3COH Utilizando seus conhecimentos sobre a gliconeogênese, tente justificar a hipoglicemia causada pela ingestão de álcool. glicose hexocinase Go´ = -8.0 kcal/mol ATP ADP glicose-6-fosfato fosfo-hexose isomerase Go´ = -0.6 kcal/mol frutose-6-fosfato fosfofrutoocinase Go´ = -5.3 kcal/mol ATP ADP frutose-1,6-bisfosfato aldolase Go´ = -0.3 kcal/mol triose-fosfato isomerase Go´ = +0.6 kcal/mol gliceraldeído-3-fosfato NAD+ Pi NADH gliceraldeído3P isomerase Go´ = -0.4 kcal/mol 1,3-bisfosfogliceratoato ADP fosfogliceratocinase Go´ = +0.3 kcal/mol ATP 3-fosfogliceratoato fosfoglicerato mutase Go´ = +0.2 kcal/mol Go´ 2-fosfogliceratoato enolase = -0.8 kcal/mol fosfoenolpiruvato piruvatocinase o G ´ = -4.0 kcal/mol ADP ATP piruvato Reação para conversão de frutose-1,6-bisfosfato a frutose-6-P (para reversão da reação da fosfofrutocinase) Pi frutose-1,6-bisfosfatase frutose-1,6-bisfosfato frutose-6-P Reação para conversão de glicose-6-P a glicose (para reversão da reação da hexocinase) Pi glicose-6-fosfatase glicose-6-P glicose expressa somente no fígado e no cortex renal F1,6BP F6P glicerol-P glicose G6P F1,6BPase G6Pase lactato DHAP G3P piruvato G3PDH 1,3BPG PEP piruvato PEPCK oxaloacetato MDH malato PEP PIRUVATO CARBOXILASE PEPCK oxaloacetato aas MDH malato α-cetoglutarato fumarato succinato succinil-CoA lactato glicerol aminoácidos Requerimento diário de glicose: 120 g F1,6BP F6P glicerol-P glicose G6P F1,6BPase 40 g 20 g 60 g G6Pase lactato DHAP G3P piruvato G3PDH 1,3BPG PEP piruvato PEPCK oxaloacetato MDH malato PEP PIRUVATO CARBOXILASE PEPCK oxaloacetato aas MDH malato α-cetoglutarato fumarato succinato succinil-CoA Em 1930, Weil-Marlherbe e colaboradores observaram, provocando um certo escândalo no meio científico, que a adição de acetoacetato provocava um aumento na formação de glicose em fatias de rim de rato incubadas na presença de lactato. Por que esses dados pareceram estranhos? T ente explicá-los imaginando de que maneiras uma substância pode estimular uma reação? Agora discuta a sua resposta com base nos resultados encontrados por WeilMarlherbe. De onde vêm os precursores para a síntese de glicose? NEURÔNIO HEMÁCIA CO2 glicose glicose lactato TAG glicerol glicerol + lactato ácidos graxos glicose ADIPÓCITO F1,6BP F6P glicerol-P glicose G6P F1,6BPase G6Pase lactato ácido graxo DHAP ácido graxo G3P piruvato G3PDH acil-CoA 1,3BPG proteínas PEP piruvato acetil-CoA PEPCK oxaloacetato MDH malato PEP aas PIRUVATO CARBOXILASE PEPCK oxaloacetato aminoácidos aas propionil-CoA MDH malato α-cetoglutarato fumarato succinato HEPATÓCITO succinil-CoA CÉLULA MUSCULAR 1980, Van Schaftingen e colaboradores descobriram uma substância capaz de modificar a atividade da fosfofrutocinase isolada de fígado, como mostra a figura abaixo: Esta substância é formada no fígado podendo atingir 20M em ratos bem alimentados e destruída após tratamento com glucagon. Observou-se que esta mesma substância era capaz de inibir a frutose 1,6bisfosfatase com um ki = 0,5M, que é uma concentração próxima daquela necessária para a metade da ativação máxima da fosfofrutocinase. Analise estes dados e procure integrá-los a um esquema metabólico mais geral. Os mesmos autores descobriram, em 1981, uma enzima capaz de sintetizar frutose-2,6-BP a partir de frutose-6P às custas de ATP, à semelhança do que ocorria na reação catalisada pela fosfofrutocinase anteriormente conhecida. Para evitar confusão as duas enzimas foram denominadas fosfofrutocinse-1 (PFK-1), a clássica, e fosfofrutocinase-2 (PFK-2), a que sintetiza frutose-2,6-bisfosfato. Além disso, o mesmo grupo de trabalho em 1982 purificou de fígado de rato, uma enzima capaz de transformar frutose-2,6-BP em frutose-6P. Observaram ainda que a forforilação desta enzima pela proteína cinase AMPc dependente (PKA) provocava um grande aumento em sua atividade. A PFK-2 também é substrato para PKA, sendo o resultado da fosforilação um acentuado decréscimo de sua atividade. Regulação recíproca da GLICÓLISE/GLICONEOGÊNESE por frutose-2,6-bisfosfato Frutose-2,6-bisfosfato sinal / estímulo E Re c AC Gs Gs GTP α GDP α ATP PKA cAMP GTP AMP - + P PFK2 / F2,6Pase Gliconeogênese Fosforilação de proteínas piruvato quinase - P - glicogênio sintase + glicogênio fosforilase P Degradação de Glicogênio P P lipase P + acetil-CoA carboxilase Lipólise A adrenalina age no músculo e no fígado através de sua ligação a receptores adrenérgicos, o que resulta na ativação da PKA. Nos dois tecidos, a PKA catalisa a fosforilação da fosfofrutocinase2/frutose 2,6-bisfosfatase. Entretanto, os efeitos sobre a glicólise em cada um dos tecidos são opostos: no fígado esta via é inibida, enquanto no músculo é ativada. Tente sugerir uma explicação para estes achados. Hue e colaboradores, em 1981 (J. Biol. Chem. 256, 8900-8903), estudaram os efeitos do glucagon no metabolismo de hepatócitos de rato e verificaram alterações dose-dependentes nos níveis de frutose-2,6-bifosfato e na atividade da fosforilase a (enzima que catalisa a degradação do glicogênio), como mostrado na figura abaixo. a) Interprete a figura, ressaltando os efeitos metabólicos do glucagon no fígado em função dos resultados observados. b) Quais seriam os resultados desta experiência se as células usadas fossem células musculares? c) Ao fazer a questão 2a, você concluiu que a produção de glicose poderia ser mantida pelo consumo das proteínas corporais por quantos dias? Reflita sobre o valor encontrado e comente as adaptações que ocorrem ao longo do jejum em relação a esta questão.
Download