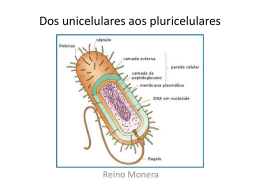
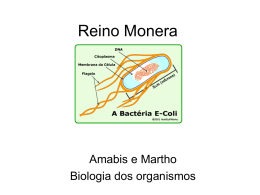
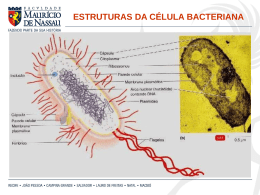
Departamento de Microbiologia Instituto de Ciências Biológicas Universidade Federal de Minas Gerais http://www.icb.ufmg.br/mic Citologia e fisiologia bacteriana Introdução As bactérias possuem várias formas e tamanhos. Elas podem variar de menos de 0,1 µm a até 1,0 mm, ficando o tamanho médio da maioria em torno de 1,0 µm, sendo apenas vistas ao microscópio óptico. O menor grupo de bactérias acreditava-se, até bem pouco tempo, ser as dos gêneros Chlamydia e Rickettsia, com 0,2 µm e 0, 3 µm de comprimento, respectivamente, mas recentemente foram descobertas bactérias, denominadas nanobactérias, que podem chegar a até 0,05 µm de comprimento. Bactérias gigantes também já foram descritas como as da espécie Thiomargarita namibiensis que chegam a medir 0,8 mm de comprimento. Quanto à forma, as bactérias podem ser classificadas basicamente em cocos esféricos, em bacilos (forma de bastão) e em espiral. Essas formas podem ainda apresentar variações. Os cocos, por exemplo, podem também se apresentar ovais (cocoíde), alongados (cocobacilos) ou achatados em uma das extremidades. Os bacilos também podem ter variações em suas extremidades, sendo arredondadas, afiladas (fusobactéria) ou quadradas (retangulares), e em forma de meia lua. As bactérias espirais podem ser curvas com extremidade arredondada, parecendo vírgulas (vibriões), afiladas, ou serem espiraladas, lembrando um saca-rolha (espirilos) ou com o corpo flexível (espiroquetas). Essas células também podem se agrupar formando uma variedade de arranjos. Os cocos podem aparecer isolados, aos pares (diplococos), em cadeia (estreptococos), em cachos (estafilococos), em grupo de quatro células formando uma tétrade ou tetranca, ou em grupo de oito células formando um cubo (sarcina). Os bastonetes também podem se apresentar isolados ou agrupados, podendo ser encontrados em dupla (diplobacilos), formando uma cadeia (estreptobacilos), em paliçada, lembrando uma parede de tijolos, em forma de letra chinesa, ou formando agrupamentos miceliais, lembrando fungos filamentosos (Quadro 1). A forma das células bacterianas é determinada por fatores genéticos e pela presença da parede celular. Remodelagens nos peptideoglicanos que compõem a parede celular podem alterar a forma da célula, indicando que a localização e a síntese desses polissacarídeos influenciam na morfologia bacteriana geral. A genética tem comprovado que um agrupamento de genes, como mreB, mreC, mreD e rodA, codificam fatores que estão envolvidos na síntese de pertideoglicanos e na sua remodelagem. Recentemente, também foi descoberta a existência de citoesqueleto em bactérias, e sua participação como sendo um dos elementos responsáveis pela forma da célula bacteriana. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 1 Quadro 1: Formas básicas e arranjos celulares em bactérias. Formas bacterianas Arranjos celulares Representação esquemábásicas tica das formas Isolados Cocos Espiral Esféricos Chlamydia trachomatis Cocóides Enterococcus Cocobacilos Mycoplasma, Brucella Diplococos Neisseria gonorrhoeae Estreptococos Streptococcus hemolyti- Estafilococos Staphylococcus aureus Tétrades Micrococcus Sarcinas Sarcha ventriculi Isolados Bacilos ou bastonetes Exemplos Extremidades arredondadas Salmonella, E. coil, Pseudomonos Retangulares Bacillus anthracis Fusobactéria Fusobacterium Meia lua Selenomonas Diplobacilos Diplobacillus variabilis Estreptobacilos Bacillus megaterium Em paliçada Corynebacterium Em forma de letra chinesa Corynebacterium diphteriae Micelial Streptomyces Vibriões Vibrio cholerae Espirilos Campylobacter Spirillum Espiroquetas Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic Treponema Leptospira, burgdorferi pylori, pallidum, Borrelia 2 Ultraestrutura bacteriana A célula bacteriana possui várias estruturas, algumas delas sendo essenciais, estando sempre presentes nas células, e outras fazendo parte apenas de determinadas espécies ou grupos de bactérias (Figura 1). Figura 1: Representação esquemática das estruturas de uma célula bacteriana típica. Neste corte longitudinal é possível a visualização das estruturas internas e externas. Um modelo de estudo é a E. coli. 1. Cápsula e glicocálix Nem todas as bactérias possuem essa estrutura, sendo divididas em capsuladas e acapsuladas. Ela é a parte mais externa da célula e é um determinante de virulência de muitas espécies patogênicas, como as dos gêneros Streptococcus, Staphylococcus e Bacillus e as da família Enterobacteriaceae. Algumas bactérias de importância industrial também a possuem, como as Leuconostoc e as Xanthomona, produtoras das gomas dextrana e xantana, respectivamente. A cápsula é uma camada gelatinosa que envolve toda a bactéria e esta fortemente associada à superfície celular, de forma generalizada ela é chamada de glicocálix. É composta por polissacarídeos sintetizados por enzimas presentes na superfície celular, exceto em Bacillus anthracis e Bacillus liceniformis, que é composta de ácido poli-D-glutanato. O glicocálix pode apresentar variações na espessura e na flexibilidade. Se ele é mais condensado e rígido, o glicocálix é chamado de cápsula, se é mais frouxo e não inclui partículas, chama-se camada limosa, ou mucosa ou de slime. O glicocálix não é uma estrutura essencial para a célula bacteriana mais pode desempenhar importantes papéis. Tem função na proteção contra dissecação do meio e pode ser um reservatório de água e nutrientes. A cápsula também atua na aderência da bactéria na superfície do hospedeiro e de substratos, devido a receptores específicos que servem com sítios de ligação com outras superfícies. Uma causa dessa adesão é a formação de biofilmes que são responsáveis por problemas industriais e hospitalares. Na indústria, a formação de biofilmes em tubulações pode perfurá-los causando vazamento de materiais, como óleo. Muitas infecções hospitalares podem ter relação com a formação de biofilmes em cateteres. Mas esses polímeros extracelulares também podem ter fins econômicos na indústria alimentícia e de cosméticos, por exemplo, como emulsificantes e espessantes, além de outras utilizações. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 3 O glicocálix desempenha ainda outras funções. As bactérias encapsuladas patogênicas têm maior capacidade invasiva e são dificilmente reconhecidas e destruídas por fagócitos, além de inibir o sistema de complemento. Os microrganismos formadores de biofilme têm maior resistência a biocidas. 2. Parede célula A parede celular é a estrutura comum a todas as bactérias, com exceção de uma única espécie, Mycoplasma, que são circundadas apenas pela membrana celular. Como já dito, ela atua conferindo rigidez e mantendo a forma celular. Também tem a função de suportar a pressão de turgor sofrida pela célula bacteriana, devido às altas concentrações de solutos dissolvidos em seu interior, e é importante na divisão celular, dando origem ao septo que separa as duas células. A parede celular de procariotos é de difícil visualização ao microscópio óptico, mas pode ser observada ao microscópio eletrônico. A estrutura, os componentes e as funções da parede celular distinguem as bactérias em gram-positivas e gram-negativas e em álcool-ácido resistentes (B.A.A.R), estas última sendo representadas pelo gênero Mycobacteruim. Este grupo de bactérias tem a particularidade de possuir em sua parede cerídeos e ácidos micólicos que lhes confere resistência a corantes ácidos, não sendo facilmente corado pela técnica de Gram como nas demais bactérias. O domínio Archaea também não apresenta uma parede típica de bactérias, tendo somente um tipo de ácido em sua estrutura ou apenas proteínas. O constituinte básico da parede celular das bactérias Gram positivas e negativas são os peptideoglicanos, também chamados de mureína ou mucopeptídio, que são polímeros complexos. As Gram positivas apresentam uma espessa camada dessas macromoléculas (90% da parede), além de ácidos teicóicos e lipoteicóico, enquanto as Gram negativas apresentam uma fina camada de peptideoglicanos (10% da parede), um espaço periplasmático e uma membrana externa, incluindo lipopolissacarídeos (LPS) (Figura 2). Figura 2: Representação esquemática da parede celular de bactérias Gram positivas e Gram negativas. (a) Parede de Gram positivas, mostrando uma espessa camada de peptideioglicanos, englobando ácidos teicóicos e ácidos lipoprotéicos, que se estendem para a membrana celular. (b) Parede de Gram negativa, formada por uma membrana externa, que inclui porinas e lipopolissacarídeos, ligada a uma fina camada de peptideioglicano por lipoproteínas. A camada de peptideoglicano fica dentro do espaço periplasmático, localizado entre as membranas. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic O peptideioglicano é uma estrutura macromolecular covalente formada por rígidas camadas de glicano formando uma trama em meio a flexíveis pontes de peptídeos. Ele é constituído por mais duas partes: um arcabouço, composto por N-acetilglicosamina (GlcNAc) e ácido N-acetilmurâmico (MurNAc) alternados; e um conjunto de cadeias laterais de tetrapeptídios ligados ao MurNAc (Figura 3). Os componentes e a modelagem dos peptideioglicanos estão bem elucidados, mas a sua organização ainda não é bem compreendida. Acreditava-se que eles eram arranjados eram em paralelo à membrana plasmática, mas estudos recentes têm demonstrado que esse arranjo é em posição perpendicular, mudando uma visão tradicional sobre a parede celular de bactérias. 4 Na parede de bactérias Gram positivas o ácido tecóico pode compreender até 50% da massa seca da parede. Ele é um polímero formado por resíduos de glicerol ou ribitol ligados por fosofodiéster, sendo divididos em ácido teicóico e lipoteicóico, estes estão intimamente ligados à porção lipídica da membrana celular. Figura 3: Representação esquemática da estrutura química do peptideoglicano. As repetições constituídas por N-acetilglicosamina (GlcNAc) e ácido N-acetilmurâmico (MurNAc) formam os vertentes de glicano que são ligados por pontes peptídicas cruzadas. Essas ligações são dependentes do ácido diaminopimélico (DAP). Setas vermelhas indicam reações de síntese e as amarelas, clivagem. (a), atividade transglicosilase; (b), atividade transpeptidase, com perda do terminal D-alanina e um pentapeptídeo; (c), atividade transglicosilase lítica; (d), atividade endopeptidase; (e), atividade carboxipeptidase; (f), atividade amidase; (g), atividade N-acetilglicosaminidase. A parede celular das bactérias Gram-negativa é mais complexa. A membrana externa é formada por uma dupla camada lipídica, a sua parte interna é composta basicamente de fosfolipídeos e é hidrofóbica, e a parte voltada para o exterior por lipopolissacarídeos e proteínas e é hidrofílica. O LPS, também chamado de endotoxina, é constituído por um lipídio complexo, o lipídio A, ligado ao antígeno O. As principais proteínas da membrana externa são as porinas que formam poros permitindo a passagem passiva de solutos. O espaço periplasmático, localizado entre a membrana externa e citoplasmática, contém a camada de peptideoglicano, enzimas hidrolíticas, para quebra de macromoléculas, enzimas capazes de inativar drogas e proteínas transportadoras de solutos. As Tabelas 1 e 2 mostram as principais funções dos constituintes das paredes celulares de bactérias Gram positivas e Gram negativas. Tabela 1: Constituintes da parede celular de bactérias Gram positivas e suas principais funções Constituinte Peptideoglicano Ácido teicóico e lipoteicóico Principais funções Possibilita o suporte rígido e a forma celular; protege contra pressão osmótica; é o sítio de ação das penicilinas e das cefalosporinas; é degradada com lisozima Principal antígeno de superfície; promove a aderência do microrganismo às mucosas; facilita a ligação e a regulação da entrada e saída de cátions na célula; regula a atividade das autolisinas na divisão celular; constitui sítios de recepção de bacteriófagos Tabela 2: Constituintes da parede celular de bactérias Gram negativas e suas principais funções Constituinte Peptideoglicano Principais funções Mesma função que na Gram-positiva Espaço periplasmático Contém enzimas envolvidas no transporte, na degradação e síntese de substâncias. As enzimas que inativam drogas podem tornar a bactéria resistente a elas Estabilidade e transporte; fornece uma barreira contra a entrada de moléculas grandes ou hidrofóbicas, como antibióticos, lisozina, detergentes, metais pesados, sais de bile, enzimas digestivas e alguns corantes; fator importante na inibição de fagócitos e do complemento durante a invasão de um hospedeiro Endotoxina promovendo respostas fisiológicas em hospedeiros; potente estimulador da resposta imunológica Membrana externa LPS 3. Membrana citoplasmática A membrana celular bacteriana tem uma estrutura em bicamada lipídica semelhante às deCitologia e fisiologia bacteriana - www.icb.ufmg.br/mic 5 mais membranas celulares, sendo constituída por 70% de proteínas e 30% de fosfolipídios. Diferindo da membrana dos eucariotos pela ausência de esteróides, com exceção dos micoplasmas que incorporam colesterol quando crescem em meio que o contém. Os ácidos graxos dos lipídeos são responsáveis pela hidrofobicidade da parte interna da membrana, enquanto a parte hidrofílica fica voltada para o meio externo aquoso. Interações hidrofóbicas, pontes de hidrogênio e cátions, como Mg++ e Ca++, mantêm a integridade dessa estrutura. A membrana plasmática forma uma barreira, separando o meio interno do externo e a ela são atribuídas muitas das funções encontradas na membrana eucariótica. Suas principais funções é a permeabilidade seletiva e o transporte de solutos, o transporte de elétrons e fosforilação oxidativa em espécies aeróbias, excreção das exoenzimas hidrolíticas, localização das enzimas e moléculas transportadoras que atuam na biossíntese do DNA e a localização dos receptores e outras proteínas do sistema quimiotático dos outros sistemas de transdução sensorial. 4. Citoplasma O citoplasma apresenta duas áreas distintas quando visualizado ao microscópio eletrônico: uma matriz amorfa que contém ribossomos, grânulos nutrientes, vacúolos gasosos, metabólitos e íons; e uma região mais interna contendo o nucleóide. a) Ribossomos: Assim como os de eucariotos, é o sítio de síntese protéica, diferindo apenas em tamanho e em composição química. Os ribossomos bacterianos apresentam tamanhos diferentes. b) Grânulos: O citoplasma contém vários tipos diferentes de grânulos que funcionam como sítios de armazenamento para nutrientes e se coram caracteristicamente com determinados corantes. c) Vacúolo gasoso: Encontrados em procariotos que vivem flutuando em lagos ou mares. Sua parede celular é composta apenas de unidades repetidas de proteínas, formando uma estrutura rígida que permite apenas a entrada de gases. d) Mesossomo: É uma invaginação enovelada da membrana citoplasmática que atua como um apoio para ligar e, posteriormente, separar os cromossomos das células filhas durante a divisão celular. 5. Nucleóide As bactérias não possuem membrana nuclear, ficando o material genético empacotado e compactado em uma região chamada de nucleóide. A única exceção são as planctomicetes, um grupo divergente de bactérias aquáticas, com nucleóide delimitado por uma carioteca. Geralmente, é encontrado nas bactérias um único cromossomo circular contínuo com peso molecular aproximado de 2 a 3 x 109d. Entretanto, pode-se encontrar bactérias com mais de uma cromossomo, como são nas espécies Vibrio cholerae e Brucella melitensis. A quantidade de cromossomos na célula depende da velocidade de crescimento, bactérias que crescem rápido necessitam de mais nucleóide que as de crescimento lento. Nos procariotos algumas espécies têm o cromossomo linear e não circular, como em Burrelia burgdorferi, que parece apresentar repetições formando grampos nas extremidades do DNA, e Streptomyces coelicolor, que apresenta proteínas covalentemente ligadas a essas extremidades. O DNA bacteriano é de cadeia dupla e por ter somente uma cópia de cada gene, todas as bactérias são geneticamente haplóides. A replicação é semiconservativa, os genes estão organizados em operons, não apresentam íntrons, tem alto conteúdo G + C e possuem baixa frequência de sequências repetidas. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 6 6. Plasmídeo Os plasmídeos são pequenos fragmentos de DNA de fita dupla, a maioria é circular, são extracromosomais, replicam-se independentemente do cromossomo e podem existir em número variável na célula. São mais comuns em bactérias Gram negativas. Embora não seja essencial à sobrevivência da célula bacteriana, conferi-lhe vantagens seletivas. A maioria do DNA plasmidial se encontra na forma superenovelada. A replicação do plasmídeo é diferente nas bactérias, enquanto nas Gram negativas acontece de modo similar à replicação em cromossomos, nas Gram positivas, ocorre por círculo rolante, similar ao fago Øx174. Os plasmídeos lineares se replicam com a participação de uma proteína ligada à extremidade 5’ das duas fitas, atuando como iniciador da síntese de DNA. Todos eles carregam consigo os genes para a sua própria replicação. O principal mecanismo de transferência de plasmídeos de uma célula bacteriana a outra é a conjugação. Aqueles que comandam sua própria transferência são chamados de plasmídeos conjugativos. O plasmídeo mais estudado é o de resistência (Fator R), que confere resistência a drogas e a vários inibidores de crescimento. Os genes carregados pelo plasmídeo R codificam proteínas que inativam antibióticos ou interferem na sua captação pela célula. Muitos fatores de virulência são codificados por genes plasmidiais. Alguns outros tipos de plasmídeos são os responsáveis pelo fator de fertilidade bacteriano (Fator F), o plasmídeo COL (Fatos colicolgênico), os responsáveis pela fixação de nitrogênio no solo, os que degradam metais pesados, os codificadores de toxinas, entre outros. 7. Endósporos Nem todas as bactérias formam endósporos, ficando mais restrito a alguns gêneros de Gram positivas, como Bacillus (aeróbio obrigatório), Clostridium (anaeróbio obrigatório), Thermoactinomyces, Sporolactobacillus, Sporosarcina, Sporotomaculum, Sporomusa e Sporohalobacter. Essas estruturas são formadas dentro das células por um processo chamado esporulação, que é um tipo de diferenciação que ocorre em resposta a situações desfavoráveis no meio ambiente, de depleção de alguns nutrientes (com falta de carbono, nitrogênio ou fósforo). Várias mudanças estruturais ocorrem na célula vegetativa até ela se transformar em esporo, envolvendo a produção de enzimas e metabólitos e desaparecimento de vários componentes celulares. Essa diferenciação promove a ativação de muitos genes e modificação da transcrição de RNA polimerase. Estruturalmente, os esporos são muito complexos, possuindo várias camadas. A parte mais externa é o exósporo, um envoltório fino e delgado formado por lipoproteínas; abaixo uma camada de queratina, a capa. Mais internamente se encontra o córtex, constituído de peptideoglicanos frouxamente associados. Por baixo dessas camadas se encontra o protoplasma, que é o citoplasma compactado, e um núcleo, circundados por uma membrana (Figura 4). O endósporo é altamente resistente à dissecação, ao calor e aos agentes químicos. Quando encontram condições favoráveis, germinam produzindo uma célula vegetativa. Um endósporo pode permanecer dormente por muitos tempos (um esporo de Clostridium aceticum foi encontrado após 34 anos em um frasco em um depósito da Universidade da Califórnia; em fragmentos de ruínas de sítio arqueológico romano no Reino Unido, foram encontrados esporos de Thermoactinomyces datados de 2.000 anos). Quando a posição intracelular, o endósporo pode ser central, subterminal ou terminal. Em actinobactérias, os endósporos são exteriorizados, podendo ser únicos, conidiósporos; enfileirados ou esporangiósporos; dentro de um sáculo (Figura 5). Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 7 Figura 4: Representação esquemática das estruturas internas de um endósporo bacteriano. Figura 5: Localização do endósporo na célula bacteriana. 8. Fímbrias e Pili Alguns autores fazem distinção entre fímbrias e pili, mas as considerando como estruturas similares aos flagelos, constituídas de proteínas (pilina), porém sem está relacionadas à motilidade celular. Enquanto as fímbrias (ou pili comuns) são consideradas mais curtas e em maior número, de 100-200 por célula, os pili (ou pili sexuais) são mais longos e presente em menor número, de 1-4 por célula (Figura 6a). Os pili comuns podem apresentar adesinas nas suas pontas e por isso tem a capacidade de fixação, estando relacionados à capacidade de adesão de certas bactérias simbióticas e patogênicas a superfícies, e também à formação de películas ou biofilmes. Os pili sexuais estão envolvidos no processo de conjugação, fixando a bactéria doadora à receptora (Figura 6b). a b Figura 6: Representação esquemática dos apêndices superficiais das bactérias. (a) Visualização de um bastonete (podendo ser E. coli) mostrando vários pili comuns ao longo da superfície celular, um pili sexual, mais longo e flexível, com partículas de fagos aderidas, e um flagelo, mais comprido e mais espesso. (b) Processo de conjugação entre duas células bacterianas, através da presença de um pili sexual. É comum ver vírus específicos se aderirem ao pili. 9. Flagelo Os flagelos são apêndices longos e finos com cerca de 20-25 nm de diâmetro, em forma de chicote, que movem as bactérias na direção de nutrientes e outros atrativos ou as deslocam em direção contrária, no caso de algum fator hostil. São formados por estruturas protéicas em arranjo helicoidal, chamadas flagelinas, que se movem através de um potencial de membrana gerado pela quebra de ATPs. Apresentam-se em número variado nas diversas bactérias, sendo comuns em bastonetes e em bactérias curvas (Vibriões, E. coli, Salmonella, Bacillis, Micrococcus, entre outros), mas também podem estar presentes em alguns cocos. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 8 O comprimento do flagelo é geralmente maior do que o da célula. Sua forma e tamanho são determinados pela flagelina, sua base apresenta uma região mais larga, chamada de gancho, conectando o filamento a região do motor flagelar. A estrutura do flagelo é semi-rígida, quase não sofrendo flexão, deslocando-se rapidamente. O movimento flagelar é rotatório, semelhante a uma hélice, sendo transmitida pelo motor. Os flagelos bacterianos formam arranjos diferentes, sendo encontrado quatro tipos de disposição flagelar na célula: monotríquio (único flagelo polar), lofotríquio (tufo de flagelos nas extremidades), anfitríquio (um flagelo em cada extremidade) e peritríquio (flagelo presentes em toda superfície celular) (Figura 7). Metabolismo bacteriano Os microrganismos têm a capacidade de realizar reações químicas e de organizar as moléculas em estruturas específicas, para que ocorra a replicação celular. O metabolismo bacteriano reúne todas as reações bioquímicas que estes microrganismos podem realizar para a manutenção das atividades vitais da célula. Essas reações podem ser catabólicas, quando há liberação de energia, ou anabólicas, quando ocorre o consumo de energia. Figura 7: Disposição dos flagelos na célula bacteriana. As reações anabólicas envolvem a síntese de compostos orgânicos estruturais, como as proteínas, e funcionais, como os hormônios. Elas são importantes para o crescimento, a construção e o reparo de estruturas celulares. As rea- ções anabólicas envolvem a síntese de compostos orgânicos estruturais, como as proteínas, e funcionais, como os hormônios. Elas são importantes para o crescimento, a construção e o reparo de estruturas celulares. As reações catabólicas envolvem a degradação de compostos orgânicos em moléculas mais simples, fornecendo energia para processos vitais, tais como motilidade, transporte e síntese de moléculas complexas. Quando as formas de obtenção de energia, as bactérias podem manter o metabolismo autotrófico ou heterótrofo. As autotróficas são capazes de utilizar CO2 como principal fonte de carbono, obter energia pela síntese de ATP pela oxidação de compostos inorgânicos (quimiossintetizantes) ou captação de energia luminosa (fotossintetizantes). As heterotróficas ou organotróficas fazem a síntese de ATP a partir da energia liberada pela oxidação de compostos orgânicos, transformando-os em moléculas mais simples e utilizadas como fonte de carbono. O modo como as bactérias podem transformar a energia define seu processo metabólico. As que oxidam compostos químicos empregam os processos de respiração aeróbia, anaeróbia ou fermentação, enquanto as que utilizam luz como fonte de energia, fazem fotossíntese. Na respiração aeróbia ocorre a oxidação do piruvato, na glicólise, a dióxido de carbono e água. Todos os organismos aeróbios têm ampla diversidade e estão presentes em todas as partes do globo. Os microrganismos anaeróbios utilizam outros aceptores finais de elétrons que não sejam o O2, como o sulfato em bactérias do gênero Desulfovibrio. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 9 A fermentação é o processo de obtenção de energia no qual a molécula orgânica a ser metabolizada não é totalmente oxidada, não liberando toda a sua energia. Os substratos mais comuns no processo de fermentação são os açúcares e os aminoácidos, e os produtos obtidos dependem do substrato inicial. Algumas bactérias fermentadoras têm importância econômica, como as espécies Streptococcus thermophilus e Lactobacillus bulgaricus, que são produtoras de iogurte. Todos os organismos fermentadores têm nichos restritos a ambientes anaeróbios, onde a fonte de carbono é aproveitável. A capacidade de crescer em presença ou ausência de oxigênio divide as bactérias em: aeróbias estritas, que crescem onde há disponibilidade de oxigênio, como as do gênero Pseudomonas; microaerófilas, essas requerem uma quantidade reduzida de oxigênio, porque este composto é tóxico a elas em altas concentrações, e altas concentrações de CO2, como são as do gênero Campylobacter; anaeróbias facultativas, que utilizam oxigênio no seu metabolismo, mas podem crescer em sua ausência, como todos os membros da família Enterobacteriaceae; anaeróbias aerotolerantes, podendo suporta a presença do oxigênio, mas sem utilizá-lo no seu metabolismo, exemplo é a espécie Lactobacillus acidophillus; e anaeróbias estritas, que não crescem na presença de oxigênio. Muitas das espécies anaeróbias estritas vivem no solo ou em micro-ambientes em organismos animais que tenham se tornado anaeróbicos, como ferimentos profundos ou a junção das gengivas com os dentes, alguns exemplos são as espécies Clostridium tetani (causadora do tétano), Clostridium botulinum (causadora do botulismo), Porphiromonas gengivallis e Prevotella intermédia (associadas a doenças periodontais). Crescimento bacteriano O crescimento bacteriano deve-se a um aumento do número de células em uma população. As bactérias se dividem por fissão binária, processo no qual uma célula parental se divide para formar duas novas células idênticas àqueles que lhes deu origem. Dessa forma, diz-se que as bactérias possuem crescimento exponencial, ou seja, para qualquer quantidade de células crescendo exponencialmente, quanto maior a quantidade existente dessas células, mais rápido crescerá a população bacteriana. Assim, a taxa de crescimento corresponde à variação do número de células ou da massa celular por unidade de tempo. O crescimento exponencial se apresenta inicialmente com uma taxa lenta de divisão celular, acelerando posteriormente, havendo um número explosivo de células nos estágios finais. O ciclo de crescimento de uma população de bactérias pode ser observado em uma cultura em batelada (crescimento microbiano em um recipiente fechado), o que permitiu a construção gráfica de uma curva de crescimento típica, dividida em quatros fases: fase lag, fase exponencial ou log, fase estacionária e fase de morte ou de declínio. O Gráfico 1 representa a curva de crescimento de uma cultura de bactérias, crescendo em um recipiente fechado. Gráfico 1: Curva de crescimento de uma cultura bacteriana. Neste gráfico é possível distinguir as quatro fases de crescimento. A fase lag, caracterizada por um período de crescimento pouco significativo; a fase log, de crescimento exponencial; a fase estacionária, quando o número de células que se dividem é proporcional àquele de células que morrem; e a fase de declínio, no qual o número de células morrendo é maior do que aquele que se dividem. Lembra-se que a plotação de gráficos de crescimento refletem os eventos que ocorrem em uma população bacteriana e não em uma única célula. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 10 a) Fase lag: Quando uma população microbiana é inoculada em um meio de cultura, não ocorre crescimento imediato. Essa fase é caracterizada por um período de crescimento inexpressivo, podendo variar de curto a longo e até mesmo inexistir. Um inóculo provindo de uma cultura mais velha, em fase estacionária, terá uma fase lag, porque as células que se encontram em falta de determinados nutrientes, requererão de um tempo para sintetizá-los novamente. Outra situação que ocorre essa fase é quando na cultura há células danificadas, necessitando elas de um tempo para sofrerem reparos. b) Fase log: Nesta fase se observa o crescimento exponencial. As células que passam por essa fase geralmente estão em condições saudáveis. A taxa de crescimento exponencial é variável, sofrendo influências do ambiente, como temperatura e composição do meio, e das características genéticas do próprio organismo. Geralmente, ao final da fase log, as bactérias passam a apresentar fenótipos novos, decorrentes do processo de comunicação denominado “quorum sensing”. c) Fase estacionária: Em uma cultura em batelada, esta fase é caracterizada por uma limitação no crescimento da população, decorrente a uma carência de nutrientes, devido ao seu consumo, e/ou a presença de algum produto de excreção que atinge uma concentração inibitória e promove a interrupção do crescimento exponencial. Nesta fase não é observado aumento ou diminuição no número de células, uma vez que está ocorrendo crescimento lento e morte celular ao mesmo tempo e em mesmas proporções, equilibrando o número de células, o que é chamado de crescimento críptico. d) Fase de declínio: Em uma cultura inalterada, após atingir a fase estacionária, as células podem continuar vivas ou morrer por lise celular, entrando assim em uma fase de declínio. Essa fase também pode ser exponencial ou acontecer em uma taxa inferior à taxa de crescimento exponencial, o que é mais comum. Necessidades nutricionais e físicoquímicas para o crescimento As bactérias estão presentes em todos os ambientes, tendo que crescer e sobreviver neles, para isso, possuem uma grande variedade de exigências nutricionais. Nem todos os nutrientes são exigidos nas mesmas quantidades pela célula, os macronutrientes (carbono e nitrogênio, principalmente) são necessários em grande quantidade, os micronutrientes, em menor quantidade, ou apenas traços. Os fatores de crescimento são compostos orgânicos, como as vitaminas, aminoácidos, purinas e pirimidinas, necessários em pequenas quantidades por alguns tipos de células. Não somente as condições nutricionais devem estar adequadas para o crescimento de uma população bacteriana, mas também as condições físico-químicas, que variam de uma espécie para outra, tanto quanto as necessidades nutricionais. Em relação às condições físico-químicas como temperatura e pH, as bactérias apresentam comportamento extremamente variado, podendo ser psicrofílicas, com ótimo em temperaturas baixas (Polaromonas vacuolata – 4ºC), mesofílicas, com ótimo em temperaturas medianas (Escherichia coli – 39ºC), termofílicas, com ótimo em temperaturas altas (Bacillus stearothermophilus – 60ºC), ou hipertermófilas, com ótimo em temperaturas muito elevadas (Thermococcus celer – 88ºC; Pyrolobus fumarii – 106ºC); acidófilas obrigatórias, com ótimo em pH baixo (Thiobacillus e vários gêneros de Archaea), neutófilas, com ótimo em torno do pH 7, ou alcalofílicas, com ótimo em pH alto (bactérias aeróbias marinhas e muitos Bacillus). A água também é um fator que afeta o crescimento microbiano. A sua disponibilidade, expressa geralmente em atividade de água, depende do conteúdo aquoso do ambiente e da concentração de solutos, uma vez que a associação de solutos às moléculas de água, torna-as indisponível ao microrganismo. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic 11 Literatura recomendada CABEEN, Matthew T.; JACOBS-WAGNER, Christine. Bacterial cell shape. Nature reviews/microbiology. Vol. 3. 601-610. August 2005. JAWETZ; MELNICK; ADELBERG. Microbiologia médica. 24 ed. Rio de Janeiro. Editora McGraw-Hill Interamericana do Brasil Ltda. 2009. LEVINSON, Warren; JAWETZ, Ernest. Microbiologia médica e imunologia. 7 ed. Editora Artmed. Porto Alegre. 2005. MADIGAN, Michael T.; MARTNKO, John M.; PARKER, Jack. Microbiologia de Brock. 12 ed. Editora: Artmed. São Paulo. 2010. MURRAY, Patrick R; ROSENTHAL, Ken S; PFALLER, Michael A. Microbiologia médica. 6. ed. Rio de Janeiro. Edotira Elsevier. 2008. TORTORA, Gerard J.; FUNKE, Berdell R.; CASE, Christine L. Microbiologia. 8 ed. Editora Artmed. Porto Alegre. 2009. TRABULSI, Luiz Rachid; ALTERTHUM, Flávio. Microbiologia. 5 ed. São Paulo. Editora: Atheneu. 2008. Citologia e fisiologia bacteriana - www.icb.ufmg.br/mic
Download